Embed Size (px)
Citation preview

BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 234, 549–553 (1997)ARTICLE NO. RC976678
Soluble Overexpression in Escherichia coli,and Purification and Characterization of Wild-TypeRecombinant Tobacco Acetolactate Synthase
Soo-Ik Chang,*,1 Moon-Kyeong Kang,* Jung-Do Choi,* and Sung Keon Namgoong†*Department of Biochemistry, Chungbuk National University, Cheongju 361-763, Korea; and†Department of Chemistry, Seoul Women’s Univesity, Seoul 139-774, Korea
Received April 14, 1997
characterized with respect to its isozyme pattern andAcetolactate synthase (ALS) is the first common en- subunit structure. In contrast, none of the plant ALS
zyme in the biosynthesis of L-leucine, L-isoleucine, and enzymes has been purified yet since in plants ALS isL-valine. The wild-type ALS gene from Nicotiana taba- present in low abundance and is unstable when iso-cum was cloned into the bacterial expression vector lated from plants. Accordingly, physical and kineticpGEX-2T. The resulting recombinant plasmid pGEX-
properties in addition to the isozyme pattern and sub-ALS2 was used to transform Escherichia coli strainunit structure of ALS from higher plants are not wellXL1-Blue, and the wild-type tobacco ALS (wALS) wascharacterized [15]. As a useful alternative, expressionexpressed in the bacteria as a protein fused with gluta-of plant ALS gene in bacteria could be used to purifythione S-transferase (GST). The fusion product GST-the plant ALS which are in low amounts.wALS was purified in a single step on a glutathione-
Previously, the sulfonylurea-resistant ALS geneSepharose column. The purified GST-wALS was sensi-(SurB) from N. tabacum has been functionally ex-tive to a sulfonylurea herbicide, and was lost itspressed in Escherichia coli [16]. In addition, both thesensitivity to end products, L-valine, L-leucine and L-
isoleucine. These results suggest that the purified re- wild-type and mutant Arabidopsis ALS genes havecombinant tobacco ALS was functionally active, and been functionally expressed in Escherichia coli [17-that the sulfonylureas may not bind to the feedback 18], and Brassica napus ALS has been expressed inregulatory site on the plant ALS. q 1997 Academic Press Salmonella typhimurium [19]. None of these plant
ALS enzymes expressed in the bacteria, however,have not been purified to homogeneity yet. The prob-lems could probably be due to i)lengthy purification
Acetolactate synthase (ALS, EC 4.13.18) is an en- steps with low recovery and ii)inefficient expressionzyme catalyzing the first common step in the biosynthe- in heterogous system.sis of branched chain amino acids in bacteria, yeast, The purpose of this study is to prepare a functionaland higher plants. It is the target of several classes of plant ALS, which is a valuable material for the develop-structurally unrelated herbicides, including the sulfo- ment and rational design for new herbicides. For thisnylureas [1-3], the imidazolinones [4], the triazolopyri- purpose, the wild-type tobacco ALS genes were cloned,midines [5], the pyrimidyl-oxy-benzoate [6-7], the pyri- expressed in E. coli, and purified as a fusion proteinmidyl-thio-benzenes [7], and the 4,6-dimethoxypyri- with glutathione S-transferase (GST), a protein part-midines [8]. It is feedback-inhibited by the end products ner that makes possible one step purification using glu-L-valine, L-leucine, and L-isoleucine in both microor- tathione (GSH) agarose affinity column chromatogra-ganisms and in plants [9-12]. In bacteria [13] and yeast phy. Previously, an amino-terminal fragment of the[14], ALS has been purified to homogeneity and is well sulfonylurea herbicide-resistant tobacco ALS gene was
cloned in the bacterial expression vector pGEX-2T ina similar manner, but the purified fusion product was1 To whom correspondence should be addressed. Fax: 82-431-67-
2306. E-mail: [email protected]. not in a active form [20].Abbreviations used: ALS, acetolactate synthase; GST, glutathione In this study, we report the first purification of the
S-transferase; GSH, glutathione; PCR, polymerase chain reaction; wild-type tobacco ALS enzyme in highly pure and ac-GST-wALS, the wild-type tobacco ALS fused with GST; IPTG, isopro-tive form and its interaction with a sulfonylurea herbi-pyl-b-D-thiogalactoside; EPPS, N-(2-hydroxyethyl)piperazine-N *-3-
propanesulfonic acid. cide. The results suggest that the purified recombinant
0006-291X/97 $25.00Copyright q 1997 by Academic PressAll rights of reproduction in any form reserved.
549
AID BBRC 6678 / 692d$$$101 05-09-97 10:28:44 bbrcg AP: BBRC

Vol. 234, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
tant was applied to a 5-mL GSH-Sepharose 4B column (Pharmacia-tobacco ALS was functionally active, and that the sulfo-LKB, USA) pre-equilibrated with the resuspension buffer and thennylureas may not bind to the feedback regulatory sitewashed with 10 volume the resuspension buffer to remove unboundon the plant ALS. proteins. The GST-wALS fusion was recovered from the column witha buffer of 50 mM Tris-HCl, pH 9.6, 15 mM GSH. The enzyme wasstored in aliquots at0707C. In order to obtain the recombinant wALSMATERIALS AND METHODSseparated from GST, the purified GST-wALS (1 mg) was subjectedto overnight digestion at 47C in the presence of 10 units of thrombin.Materials. Bacto-tryptone, yeast extract and bacto-agar wereAn aliquot was analyzed by SDS-PAGE [25]. Protein quantitationspurchased from Difco Laboratories (Detroit, USA). Restriction andwere determined by the method of Bradford [26].modifying enzymes were from Boehringer Mannheim (Mannheim,
Germany) and Amersham Corporation (Arlington Heights, USA). Assays of ALS. Enzyme activities of the purified GST-wALS orOligonucleotides were obtained from BIONEER (Chungbuk, Korea). wALS were measured according to the method of Westerfeld [27]The plasmid pSB1 containing the wild-type tobacco ALS gene was with a modification. The reaction mixture (200 mL) contained 50 mMobtained from Dr. B. J. Mazur (E. I. Du Pont de Nemours & Co., potassium phosphate, pH 7.0, 20 mM pyruvate, 25 mM thiamine-Wilmington, USA). The expression vector pGEX-2T was obtained pyrophosphate, 50 mM MgCl2, and 5 mM flavine adenine dinucleo-from Dr. Sang-Ki Paik (Chungnam National University, Taejon, Ko- tide in the presence or absence of various concentrations of a sulfonyl-rea). Metsulfuron methyl, a sulfonylurea herbicide, was obtained urea herbicide (metsulfuron methyl) or end products (L-valine,from Dr. Dae-Whang Kim (Korea Research Institute of Chemical L-leucine and L-isoleucine). Assays were initiated by adding the puri-Technology). fied GST-wALS or wALS enzymes and terminated by adding 20 mL
of 6 N H2SO4 after 1 h at 377C. The acidified reaction mixtures wereConstruction of the expression vectors for GST-wALS, pGEX-ALS2.heated for 15 min at 607C after which 0.2 mL of 0.5% w/v creatineThe wild-type Nicotiana tabacum ALS gene was cloned into expres-was added. Then 0.2 mL of 5%-naphthol was added and the solutionssion vector pGEX-2T, a tac promoter-based vector designed to ex-were heated for an additional 15 min at 607C. The absorbances ofpress cloned insert as a fusion with glutathione S-transferase [21].the solutions were determined at 525 nm. Specific activities of ALSThis vector incorporates a linker and thrombin cleavage site betweenwere expressed as mmol acetoin/min/mg protein. The IC50 value forthese two domains to allow cleavage and removal of the fusion part-inhibition by inhibitors is defined as the concentration of metsulfuronner after purification of the expression using glutathione (GSH) aga-methyl which inhibits ALS activity 50% in a 60-min fixed time assayrose column chromatography. First, the vector pWals was con-carried out as described above. The data were fit to the equationstructed by transfering the ALS gene into pBluescript SK(0) by si-
multaneously amplifying the gene from the plasmid pSB1 and% Activity Å 100/(1 / [I]/IC50) [1]introducing both BamHI and BamHI restriction sites at the 5* and 3 *
ends, respectively, of the coding strand by polymerase chain reaction(PCR) with the oligonucleotide primers 1 and 2: where % activity equals the amount of activity in the presence of
various inhibitor concentrations as percent of an untreated control,and [I] equals the inhibitor concentration. IC50 was calculated by1: 5*-CATCTCCGGATCC
BamHIATGTCCACTACCCAAA-3 *
non-linear least-squares and the Simplex method for error minimiza-tion [28].
2: 5*-ATGCGGATCCBamHI
TCAAAGTCAATAGG-3 *
RESULTS
The PCR was carried out as described [22]. Each reaction containedExpression and purification of the wild-type recombi-50 ng of template DNA, 100 pmol of the primers 1 and 2, 200 mM
dNTPs in 50 mM KCl, 10 mM Tris (pH 8.3), and 1.5 mM MgCl2 in nant tobacco ALS. We cloned the wild-type tobacco100 mL. The reactions were overlayed with 100 mL of mineral oil and ALS gene [29] including part of the chloroplast transitsubjected to 30 cycles of the following program: 947C, 1 min 30 sec; peptide into the bacterial expression plasmid pGEX-2T557C, 2 min; 727C, 3 min. PCR products were subcloned by ethanol
as described in Materials and Methods. Induction ofprecipitation, restriction digestion with BamHI, and ligation withthe wild-type tobacco ALS fused with GST, GST-wALS,BamHI-treated pBluescript SK(0). Finally, the ALS gene was ex-
cised from the pWals with BamHI, isolated from 1% agarose gel, and by IPTG was established by analyzing crude bacterialcloned into the BamHI site of pGEX-2T. The resulting expression extracts on SDS-PAGE as shown in Fig. 1, lane 2. Anvector pGEX-ALS2 was used to transform the E. coli strain XL1- 91 kDa band corresponding to the fusion protein com-Blue cells. Plasmid DNA was isolated according to the alkaline lysis
posed of GST (26 kDa) and the wild-type tobacco ALSmethod, and transformation of XL1-Blue cells was achieved using a(65 kDa), wALS, was detected in pGEX-ALS2 afterstandard CaCl2 transformation protocol [23]. DNA sequence analysis
of the PCR product was performed by dideoxynucleotide chain termi- IPTG induction. In order to find optimum conditionsnation method [24]. for the expression of GST-wALS, several parameters
Expression and purification of GST-wALS. XL1-Blue cells con- were adjusted. We found that the optimum growth tem-taining the pGEX-ALS2 plasmid were grown at 307C in Luria-Ber- perature for the recombinant E. coli was determinedtani (LB) medium in a 2-L round-bottomed flask to an A600 of 0.5- to be 307C, and that the optimum level of IPTG for GST-0.8. The wild-type tobacco ALS fused with GST, GST-wALS, was
wALS induction was 0.3 mM, added when the cultureinduced with 0.1-1.0 mM isopropyl-b-D-thiogalactoside (IPTG). Cellsreached an A600 of 0.7 (data not shown). Western blotwere grown for additional 4h at 307C and harvested by centrifugation
at 5,000g for 10 min. The cell paste was stored at 0707C. analysis of the crude extract indicated that the fusionFor the purification of GST-wALS, the cell paste was thawed and protein reacted with antibodies [20] against GST or
suspended in resuspension buffer (50 mM EPPS, 10% ethylene glycol, amino-terminal fragment of sulfonylurea resistant to-10 mM MgCl2, 1 mM pyruvate, 4 mg/mL Leupeptin, 3 mg/mL Aproti-bacco ALS (data not shown). The fusion protein wasnin, 2 mg/mL Pepstatin A, pH 7.5). The suspension was lysed by
sonication at 47C and centrifuged at 20,000g for 15 min. The superna- soluble and was found in the supernatant of the extract
550
AID BBRC 6678 / 692d$$$101 05-09-97 10:28:44 bbrcg AP: BBRC
Vol. 234, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
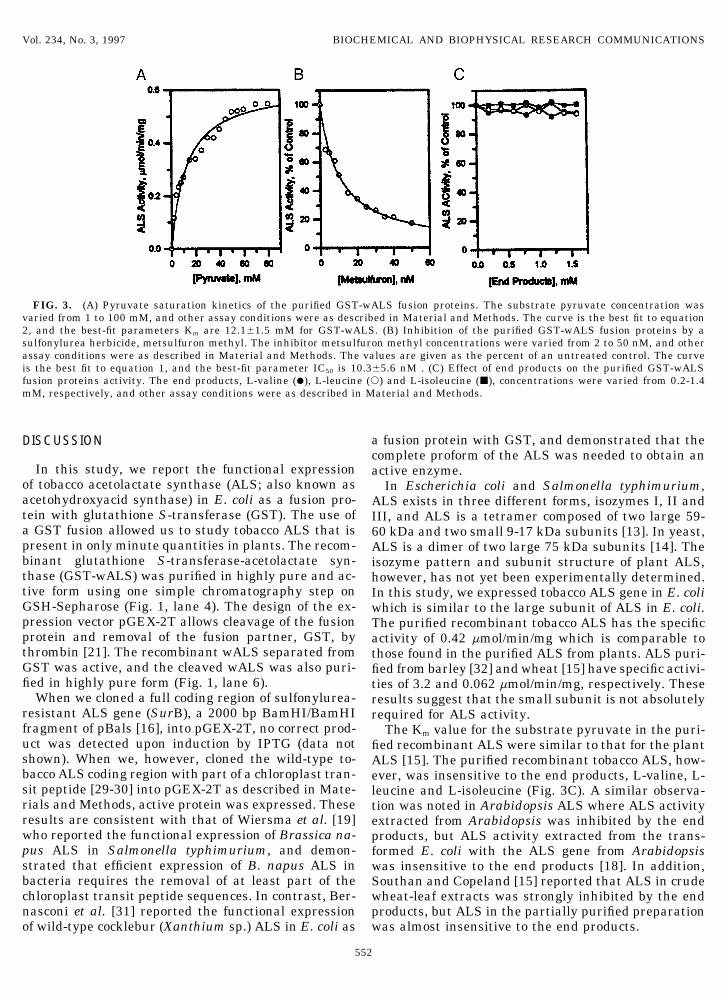
pyruvate saturation kinetics of GST-wALS. Saturationof GST-wALS by pyruvate appeared hyperbolic. Thekinetic data were fit to the equation
v Å Vmax/(1 / Km/[pyruvate]) [2]
by non-liner least-squares and the Simplex methodfor error minimization [28]. In this equation Vmax ismaximum velocity, and Km is the Michaelis constantfor pyruvate. The curve in Fig. 3A has been calcu-lated with equation 2 and the best-fit parametersVmax Å 0.61{0.02 mmol/min/mg protein and Km Å12.1{1.5 mM.
(2) Interaction of ALS with herbicides. ALS activityof GST-wALS was assayed in the presence of metsulf-uron methyl, a sulfonylurea herbicide. IC50 values weredetermined for GST-wALS. GST-wALS was extremelysensitive to inhibition by metsulfuron methyl. The
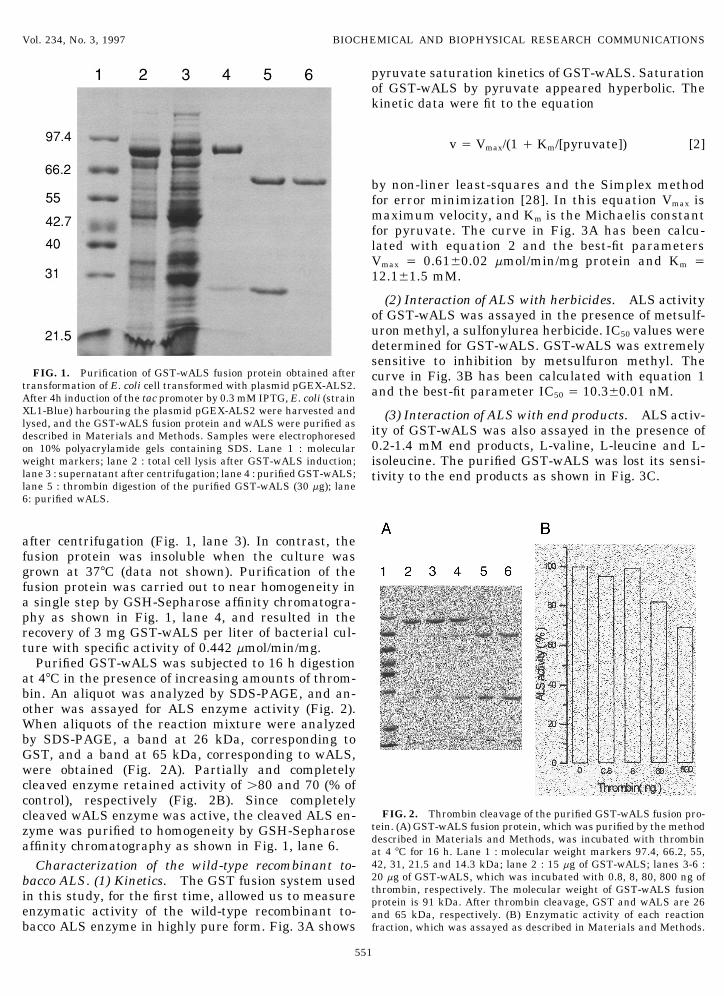
FIG. 1. Purification of GST-wALS fusion protein obtained after curve in Fig. 3B has been calculated with equation 1transformation of E. coli cell transformed with plasmid pGEX-ALS2. and the best-fit parameter IC50 Å 10.3{0.01 nM.After 4h induction of the tac promoter by 0.3 mM IPTG, E. coli (strainXL1-Blue) harbouring the plasmid pGEX-ALS2 were harvested and (3) Interaction of ALS with end products. ALS activ-lysed, and the GST-wALS fusion protein and wALS were purified as ity of GST-wALS was also assayed in the presence ofdescribed in Materials and Methods. Samples were electrophoresed
0.2-1.4 mM end products, L-valine, L-leucine and L-on 10% polyacrylamide gels containing SDS. Lane 1 : molecularweight markers; lane 2 : total cell lysis after GST-wALS induction; isoleucine. The purified GST-wALS was lost its sensi-lane 3 : supernatant after centrifugation; lane 4 : purified GST-wALS; tivity to the end products as shown in Fig. 3C.lane 5 : thrombin digestion of the purified GST-wALS (30 mg); lane6: purified wALS.
after centrifugation (Fig. 1, lane 3). In contrast, thefusion protein was insoluble when the culture wasgrown at 377C (data not shown). Purification of thefusion protein was carried out to near homogeneity ina single step by GSH-Sepharose affinity chromatogra-phy as shown in Fig. 1, lane 4, and resulted in therecovery of 3 mg GST-wALS per liter of bacterial cul-ture with specific activity of 0.442 mmol/min/mg.
Purified GST-wALS was subjected to 16 h digestionat 47C in the presence of increasing amounts of throm-bin. An aliquot was analyzed by SDS-PAGE, and an-other was assayed for ALS enzyme activity (Fig. 2).When aliquots of the reaction mixture were analyzedby SDS-PAGE, a band at 26 kDa, corresponding toGST, and a band at 65 kDa, corresponding to wALS,were obtained (Fig. 2A). Partially and completelycleaved enzyme retained activity of ú80 and 70 (% ofcontrol), respectively (Fig. 2B). Since completely
FIG. 2. Thrombin cleavage of the purified GST-wALS fusion pro-cleaved wALS enzyme was active, the cleaved ALS en-tein. (A) GST-wALS fusion protein, which was purified by the methodzyme was purified to homogeneity by GSH-Sepharosedescribed in Materials and Methods, was incubated with thrombinaffinity chromatography as shown in Fig. 1, lane 6. at 4 7C for 16 h. Lane 1 : molecular weight markers 97.4, 66.2, 55,42, 31, 21.5 and 14.3 kDa; lane 2 : 15 mg of GST-wALS; lanes 3-6 :Characterization of the wild-type recombinant to-20 mg of GST-wALS, which was incubated with 0.8, 8, 80, 800 ng ofbacco ALS. (1) Kinetics. The GST fusion system usedthrombin, respectively. The molecular weight of GST-wALS fusionin this study, for the first time, allowed us to measure protein is 91 kDa. After thrombin cleavage, GST and wALS are 26
enzymatic activity of the wild-type recombinant to- and 65 kDa, respectively. (B) Enzymatic activity of each reactionfraction, which was assayed as described in Materials and Methods.bacco ALS enzyme in highly pure form. Fig. 3A shows
551
AID BBRC 6678 / 692d$$$101 05-09-97 10:28:44 bbrcg AP: BBRC
Vol. 234, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 3. (A) Pyruvate saturation kinetics of the purified GST-wALS fusion proteins. The substrate pyruvate concentration wasvaried from 1 to 100 mM, and other assay conditions were as described in Material and Methods. The curve is the best fit to equation2, and the best-fit parameters Km are 12.1{1.5 mM for GST-wALS. (B) Inhibition of the purified GST-wALS fusion proteins by asulfonylurea herbicide, metsulfuron methyl. The inhibitor metsulfuron methyl concentrations were varied from 2 to 50 nM, and otherassay conditions were as described in Material and Methods. The values are given as the percent of an untreated control. The curveis the best fit to equation 1, and the best-fit parameter IC50 is 10.3{5.6 nM . (C) Effect of end products on the purified GST-wALSfusion proteins activity. The end products, L-valine (l), L-leucine (s) and L-isoleucine (j), concentrations were varied from 0.2-1.4mM, respectively, and other assay conditions were as described in Material and Methods.
a fusion protein with GST, and demonstrated that theDISCUSSIONcomplete proform of the ALS was needed to obtain an
In this study, we report the functional expression active enzyme.of tobacco acetolactate synthase (ALS; also known as In Escherichia coli and Salmonella typhimurium,acetohydroxyacid synthase) in E. coli as a fusion pro- ALS exists in three different forms, isozymes I, II andtein with glutathione S-transferase (GST). The use of III, and ALS is a tetramer composed of two large 59-a GST fusion allowed us to study tobacco ALS that is 60 kDa and two small 9-17 kDa subunits [13]. In yeast,present in only minute quantities in plants. The recom- ALS is a dimer of two large 75 kDa subunits [14]. Thebinant glutathione S-transferase-acetolactate syn- isozyme pattern and subunit structure of plant ALS,thase (GST-wALS) was purified in highly pure and ac- however, has not yet been experimentally determined.tive form using one simple chromatography step on In this study, we expressed tobacco ALS gene in E. coliGSH-Sepharose (Fig. 1, lane 4). The design of the ex- which is similar to the large subunit of ALS in E. coli.pression vector pGEX-2T allows cleavage of the fusion The purified recombinant tobacco ALS has the specificprotein and removal of the fusion partner, GST, by activity of 0.42 mmol/min/mg which is comparable tothrombin [21]. The recombinant wALS separated from those found in the purified ALS from plants. ALS puri-GST was active, and the cleaved wALS was also puri- fied from barley [32] and wheat [15] have specific activi-fied in highly pure form (Fig. 1, lane 6). ties of 3.2 and 0.062 mmol/min/mg, respectively. These
When we cloned a full coding region of sulfonylurea- results suggest that the small subunit is not absolutelyresistant ALS gene (SurB), a 2000 bp BamHI/BamHI required for ALS activity.fragment of pBals [16], into pGEX-2T, no correct prod- The Km value for the substrate pyruvate in the puri-uct was detected upon induction by IPTG (data not fied recombinant ALS were similar to that for the plantshown). When we, however, cloned the wild-type to- ALS [15]. The purified recombinant tobacco ALS, how-bacco ALS coding region with part of a chloroplast tran- ever, was insensitive to the end products, L-valine, L-sit peptide [29-30] into pGEX-2T as described in Mate- leucine and L-isoleucine (Fig. 3C). A similar observa-rials and Methods, active protein was expressed. These tion was noted in Arabidopsis ALS where ALS activityresults are consistent with that of Wiersma et al. [19] extracted from Arabidopsis was inhibited by the endwho reported the functional expression of Brassica na- products, but ALS activity extracted from the trans-pus ALS in Salmonella typhimurium, and demon- formed E. coli with the ALS gene from Arabidopsisstrated that efficient expression of B. napus ALS in was insensitive to the end products [18]. In addition,bacteria requires the removal of at least part of the Southan and Copeland [15] reported that ALS in crudechloroplast transit peptide sequences. In contrast, Ber- wheat-leaf extracts was strongly inhibited by the endnasconi et al. [31] reported the functional expression products, but ALS in the partially purified preparation
was almost insensitive to the end products.of wild-type cocklebur (Xanthium sp.) ALS in E. coli as
552
AID BBRC 6678 / 692d$$$101 05-09-97 10:28:44 bbrcg AP: BBRC

Vol. 234, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
4. Shaner, D. L., Anderson, P. C., and Stidham, M. A. (1984) PlantThe reasons for the insensitivity of the recombi-Physiol. 76, 545–546.nant plant ALS or the partially purified ALS from
5. Gerwick, B. C., Subramanian, M. V., Loney-Gallant, V., andplants to the end products, the feedback inhibitors,Chandler, D. P. (1990) Pest. Sci. 29, 357–364.are not known. One possibility is that these ALS were
6. Babczinski, P., and Zelinski, T. (1991) Pest. Sci. 31, 305–323.not in the native conformation or were modified dur-7. Choi, J.-D., Moon, H., Chang, S.-I., Chae, J.-K., and Shin, J.-H.ing purification [15]. The other possibility is that a (1993) Korean Biochem. J. 26, 638–643.
second subunit which is responsible for the feedback 8. Shim, H.-O., Kim, D.-W., Chang, S.-I., and Choi, J.-D. (1995) J.sensitivity of ALS does not exist in these ALS prepa- Biochem. Mol. Biol. 28, 471–476.ration. The latter possibility suggests that a second 9. Miflin, B. J., and Cave, P. R. (1972) J. Exp. Bot. 23, 511–516.subunit of ALS exists in plants [18]. However, a sec- 10. Gollop, N., Chipman, D. M., and Barak, Z. (1983) Biochim. Bio-
phys. Acta 748, 34–39.ond subunit has not yet been identified for plant ALS11. Rathinasabapathi, B., Williams, D., and King, J. (1990) Plantas well as yeast ALS.
Sci. 67, 1–6.Mutants that are resistant to branched chain amino12. Mourad, G., Williams, D., and King, J. (1995) Planta 196, 64–acids have been reported, and the resistance has been
68.shown to be a consequence of point mutations in the13. Schloss, J. V., Dyk, D, E., Vasta, J. F., and Kutny, R. M., (1985)ALS gene coding sequence [33-34]. For example, the
Biochemistry 24, 4952–4959.serine to leucine change at position 214 of tobacco ALS
14. Poulsen, C., and Stougaard, P. (1989) Eur. J. Biochem. 185, 433–is responsible for L-valine resistance [34]. In addition, 439.the sensitivity of ALS to the herbicides was not altered 15. Southad, M. D., and Copeland, L. (1996) Physiol. Plant. 98, 824–in L-valine resistant mutants of tobacco or Arabidopsis 832.[33, 35]. Recently, Hervieev and Vaucheret [34] sug- 16. Kim, H.-J., and Chang, S.-I. (1995) J. Biochem. Mol. Biol. 28,
265–270.gested a hypothesis that the herbicide and branched17. Smith, J. K., Schloss, J. V., and Mazur, B. J. (1989) Proc. Natl.chain amino acid binding sites on ALS are not identical.
Acad. Sci. USA 86, 4179–4184.We showed that the purified wild-type recombinant18. Singh, B., Szamosi, I., Hand, J. M., and Misra, R. (1992) Planttobacco ALS was potently inhibited by a sulfonylurea
Physiol. 99, 812–816.herbicide but was lost its sensitivity to the feedback19. Wiersma, P. A., Hachey, J. E., Crosby, W. L., and Moloney M. M.inhibitors (Figs. 3B and 3C). These results suggest that (1990) Mol. Gen. Genet. 224, 155–159.
the sulfonylureas may not bind to the feedback regula- 20. Chang, S.-I., Kang, M.-K., Kim, H.-J., Choi, J.-D., and Nam-tory site on the plant ALS, and thus results in this goong, S. K. (1996) J. Biochem. Mol. Biol. 29, 462–467.study are consistent with the hypothesis of Hervieev 21. Smith, D. B., and Johnson, K. S. (1988) Gene 69, 31–40.and Vaucheret [34]. 22. Saiki, R. K., Gelfand, D. H., Stoffel, S., Scharf, S. J., Higuchi, R.,
In conclusion, we demonstrated that expression of Horn, G. T., Mullis, K. B., and Erlich, H. A. (1988) Science 239,487–491.plant ALS gene in E. coli as a fusion protein with GST
23. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecularcan provide a useful alternative to purification of plantCloning: A Laboratory Manual, 2nd ed., Cold Spring HarborALS enzymes which are in low amounts and cumber-Laboratory Press, Cold Spring Harbor, New York.some to purify. This study will allow us to permit detail
24. Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc. Natl.kinetic and structural analyses of substantial quanti- Acad. Sci. USA 74, 5463–5467.ties of the purified ALS. 25. Laemmli, U. K. (1970) Nature 227, 680–685.
26. Bradford, M. M. (1951) Anal. Biochem. 72, 248–254.ACKNOWLEDGMENTS 27. Westerfeld, W. W. (1943) J. Biol. Chem. 161, 495–502.
28. Nelder, J. A., and Mead, R. (1965) Comput. J. 7, 308–313.We thank Dr. Babara J. Mazur (E. I. Du Pont de Nemours &
29. Mazur, B. J., Chui, C. F., and Smith, J. K. (1987) Plant Physiol.Co., USA) for plasmid pSB1, Dr. Sang-Ki Paik (Chungnam National85, 1110–1117.University, Korea) for plasmid pGEX-2T, and Dr. Dae-Whang Kim
30. Lee, K. Y., Townsend, J., Tepperman, J., Black, M., Chui, C. F.,(Korea Research Institute of Chemical Technology) for metsulfuronMazur, B., Dunsmuir, P., and Bedbrook, J. (1988) EMBO J. 7,methyl. This work was supported by the Korea Science and Engi-1241–1248.neering Foundation (92-46-00-01, 95-0402-09-01-3).
31. Bernasconi, P., Woodworth, A. R., Rosen, B. A., SubramanianM. V., and Siehl, D. L. (1995) J. Biol. Chem. 29, 17381–17385.REFERENCES
32. Durner, J., and Boger, P. (1988) Z. Natturforsch. 43c, 850–856.33. Wu , K., Mourad, G., and King, J. (1994) Planta 192, 249–255.1. Chaleff, R. S., and Mauvaris, C. J. (1984) Science 224, 1443–
1445. 34. Hervieu, F., and Vaucheret, H. (1996) Mol. Gen. Genet 251, 220–224.2. LaRossa, R. A., and Schloss, J. V. (1984) J. Biol. Chem. 259,
8753–8757. 35. Relton, J. M., Wallsgrove, R. M., Bourgin J. P., and Bright,S. W. J. (1986) Planta 169, 46–50.3. Ray, T. B. (1984) Plant Physiol. 75, 827–831.
553
AID BBRC 6678 / 692d$$$101 05-09-97 10:28:44 bbrcg AP: BBRC













![Research Article Positive Selection of Squalene Synthase ...downloads.hindawi.com/journals/ijg/2019/5913491.pdf · [24]. SQS1 of Panax ginseng (PgSS1) overexpression resulted in enhanced](https://img.pdfslide.us/doc/110x75/6063d34d2bd3a12ccb72c37a/research-article-positive-selection-of-squalene-synthase-24-sqs1-of-panax.jpg)















