Embed Size (px)
Citation preview

THE JOURNAL 0 1993 by The American Society for Biochemistry and
OF BIOLOGICAL CHEMISTRY Molecular Biology, Inc
Vol ,268, No. 6, Issue of Februery 25. pp. 4399-4406,1993 Printed in U. S. A .
Soluble c-Kit Proteins and Antireceptor Monoclonal Antibodies Confine the Binding Site of the Stem Cell Factor*
(Received for publication, September 11, 1992)
Janna M. Blechman$§, Sima Lev$, Maria Felice BrizzinII , Orit LeitnerS, Luigi Pegoraron, David Givol$, and Yosef Yarden$** From the $Department of Chemical Immunology, The Weizmann Institute of Science, Rehovot 76100, Israel and the flDipurtmento di Scienze Biomediche e Oncobgia Umanu, Universita di Torino, Torino 10126, Italy
The binding of the stem cell factor (SCF) to the c-kit- encoded receptor tyrosine kinase stimulates a variety of biochemical responses that culminate in cellular pro- liferation, migration, or survival. The extracellular domain of p145”” consists of five immunoglobulin-like domains. To confine the ligand binding site to this portion of the receptor we generated a panel of murine monoclonal antibodies (mAbs) to the Kit protein and identified two mAbs that efficiently displaced recep- tor-bound SCF and also inhibited proliferation of SCF- dependent human megakaryocytes. To map the epi- topes of these mAbs we constructed and expressed soluble portions of the extracellular domain of Kit, which included either the two amino-terminal Ig-like domains (denoted Kit 1-2), three Ig-like domains (Kit 1-2-3), or the entire extracellular portion (Kit-X). All three recombinant proteins were recognized by the ligand inhibitory mAbs, suggesting that the SCF bind- ing site resides in the amino-terminal half of the ecto- domain. Consistent with this conclusion, all of the sol- uble proteins inhibited SCF binding to Kit-expressing cells, and they also underwent specific covalent cross- linking to the radiolabeled ligand. However, whereas Kit 1-2-3 and Kit-X displayed comparable ligand affin- ities, deletion of the third Ig-like domain, in Kit 1-2, involved significant reduction in SCF binding. Hence, the binding site of SCF probably includes Ig-like do- mains l and 2, but structural determinants distal to this portion may also participate in ligand recognition.
~ ~~ ~~
Mice with mutations in the Steel (S l ) locus on chromosome 10 exhibit a complex phenotype that includes defects in melanogenesis (white hair coat), gametogenesis (sterility), and hematopoiesis (macrocytic anemia) (Bennett, 1956; McCulloh et al., 1964, 1965; Mintz and Russell, 1979; Silvers, 1979; Russell, 1979). S1 alleles contain deletions in a gene that encodes a transmembrane protein which functions as a pre- cursor for a peptide growth factor (Zsebo et al., 1990b; Huang et al., 1990; Copeland et al., 1990). The soluble growth factor was identified independently on the basis of its ability to
* This work was supported by National Institutes of Health Grant CA 51712, by the Citrina Foundation, an AIRC grant, and Consiglio Nazionale Ricerche Grant 92.02237.PF39. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Recipient of a postdoctoral fellowship from the Israel Cancer Research Fund.
11 Fellow of AIRC. ** Recipient of a research career development award from the Israel
Cancer Research Fund. To whom correspondence should be ad- dressed. Tel.: 972-8-343026; Fax: 972-8-344141.
promote formation of colonies from early hematopoietic pro- genitors (Zsebo et al., 1990a), its differential proliferative effect on normal mast cells, but not mast cells derived from W mutant mice (Nocka et al., 1990), and by its action on MC- 6 mast cells (Williams et al., 1990). This cytokine was respec- tively named stem cell factor (SCF),’ c-kit ligand, and mast cell growth factor. Besides its essential in vivo role in hema- topoiesis, melanogenesis, and gametogenesis, in vitro SCF acts synergistically with other growth factors including eryth- ropoietin, granulocyte-colony stimulating factor (G-CSF), granulocyte-macrophage colony stimulating factor (GM-CSF) and interleukins (ILs) 3 and 6 (reviewed in Besmer, 1991). SCF exists both as a cell surface form and as a soluble form that corresponds to 164 amino-terminal residues of the long protein. In addition, two alternatively spliced SCF mRNAs encode variant transmembrane proteins that differ in the rate of processing into the soluble form (Flanagan et al., 1991).
All of the biological effects of SCF are initiated by its binding to a cell surface receptor that is encoded by the c-kit proto-oncogene (Yarden et al., 1987; Qiu et al., 1988). The mouse kit gene is allelic with the white spotting ( W ) locus on chromosome 5 (Chabot et al., 1988; Geissler et al., 1988). Early observations indicated that W mutants are phenotypically indistinguishable from 5’1 mutant mice and therefore predicted that these two loci interact on the same functional pathway (Bennett, 1956; Russell, 1979). This prediction has been con- firmed recently through the identification of the S1 and W gene products as a ligand (SCF) and its cognate receptor (Kit), respectively. High affinity binding of noncovalently held dimers of SCF to ~ 1 4 5 ~ ’ ~ protein rapidly induces dimeri- zation of the receptor (Blume-Jensen et al., 1991; Lev et al., 1992a). This stimulates the intrinsic cytoplasm-facing cata- lytic domain of Kit that functions as a tyrosine-specific pro- tein kinase. The next step in the signaling pathway includes tyrosine phosphorylation and physical association with a dis- tinct set of proteins that generate intracellular second mes- sengers (Lev et al., 1991; Rottapel et al., 1991; Funasaka et al., 1992). The interactions of the ligand-stimulated Kit with cellular substrates is mediated, at least in part, by the hydro- phillic kinase insert domain that lies within the catalytic core and strongly couples to proteins that contain src homology domains (Lev et al., 1992b; Rottapel et al., 1991).
The Kit protein belongs, both structurally and functionally,
’ The abbreviations used are: SCF, stem cell factor; CSF, colony- stimulating factor; G-CSF, granulocyte CSF; GM-CSF, granulocyte- macrophage CSF; IL, interleukin; PDGF, platelet-derived growth factor; mAb, monoclonal antibody; Kit-X, a recombinant extracellu- lar domain of Kit/SCF receptor; CHO, Chinese hamster ovary; PBS, phosphate-buffered saline; bp, base pair(s); PAGE, polyacrylamide gel electrophoresis; EDAC, l-ethyl-3-(3-dimethylaminopropyl)car- bodiimide; EGF, epidermal growth factor.
4399

4400 SCF Binding Site of Kit
to a subgroup of receptor tyrosine kinases which includes the receptors for the platelet-derived growth factors (PDGFs), colony-stimulating factor 1, and a few receptors for still unknown ligands. These type 111 receptor tyrosine kinases (Yarden and Ullrich, 1988) are characterized by extracellular ligand binding portions that contain five immunoglobulin (1g)-like domains (Williams and Barclay, 1988; Hunkapiller and Hood, 1989). In addition, the ectodomains of these recep- tors share many common short stretches of amino acids that are distributed throughout the whole length of the extracel- lular portions (Yarden et al., 1987). This raises the question of where along the extracellular structures the differential ligand binding specificities reside. One possibility is that a specific Ig-like domain, or a specific interdomain stretch, functions as a ligand binding site. The Ig homology unit is characterized by a primary sequence of 70-100 residues in length, with an essentially invariant disulfide bridge spanning 40-60 amino acids. Several other relatively conserved residues are involved in establishing a tertiary structure referred to as an antibody fold (Amzel and Poljak, 1979). This structure is shared by all members of the Ig supergene family, which includes molecules with immunological functions, and pro- teins with no known immune function, such as cell adhesion molecules and receptors for lymphokines and growth factors (Williams and Barclay, 1988; Hunkapiller and Hood, 1989). Kit, like other nonimmunological members of the family, contains Ig homology units that correspond to the C2 type or the V type of Ig-like domains.
It is presently unclear how the compact and globular struc- tures of the Ig homology units of receptor tyrosine kinases participate in the establishment of ligand binding. However, soluble recombinant portions of the PDGF receptor (Duan et al., 1991) and Kit/SCF receptor (Lev et al., 1992c) fully retain high affinity ligand binding and also undergo ligand-induced dimerization. These observations indicated that all of the structural determinants of ligand binding are confined to the extracellular portions of these receptors. To confine the ligand binding cleft of Kit further we generated ligand-competitive monoclonal antibodies (mAbs) and recombinant soluble por- tions of the ectodomain. By using these reagents we obtained results that provide strong evidence that the SCF binding site is confined to the amino-terminal portion of Kit.
EXPERIMENTAL PROCEDURES
Materials-Radioactive materials were purchased from Amersham Corp. Protein A coupled to Sepharose was from Pharmacia LKB Biotechnology Inc. or prepared in our laboratory. Recombinant hu- man SCF and transforming growth factor-a were from Amgen (Thou- sand Oaks, CA). Recombinant human IL-3 was a gift from Genetic Institute (Cambridge, MA), and recombinant human GM-CSF was a gift from Sandoz (Basel, Switzerland). Molecular weight standards for gel electrophoresis were obtained from Bio-Rad. Calf serum was from HyClone Laboratories (Logan, UT). Rabbit antibodies to mouse Igs were from Jackson ImmunoResearch. Unless otherwise indicated, all other chemicals were from Sigma. Polyclonal antibodies to human Kit/SCF receptor were raised in rabbits that were injected with a recombinant Kit-X protein. The extracellular domain of the ~185"" receptor was purified from the medium of cells that were transfected with the corresponding portion of the neu cDNA.'
Cell Culture-CHO cells were cultured in F-12 medium containing 10% calf serum and antibodies and were maintained in a humidified incubator with 5% CO, in air at 37 "C. For transfection experiments a CHO subline that lacks the dihydrofolate reductase (dhfr) gene was used (Urlaub and Chasin, 1980). Conditioned media were obtained by culturing the cells without serum. For binding experiments the monolayers were grown on fibronectin (Boehringer Mannheim)- coated dishes (10 pg/ml fibronectin in PBS). M07e cells were rou- tinely grown as a suspension culture in Iscove's modified Dulbecco's
' I. Stancovski, unpublished data.
medium supplemented with 5% fetal calf serum and 5 ng/ml recom- binant human IL-3. Growth factor deprivation was achieved by washing the cells twice in Iscove's modified Dulbecco's medium and culturing them for 24 h in Iscove's modified Dulbecco's medium supplemented with 5% fetal calf serum at a concentration of 2 X lo6/ ml. Cells were preincubated for 50 min with different concentrations of the indicated antibody and seeded at 50,00O/well in 96-well plates with either 10 ng/ml GM-CSF, 10 ng/ml IL-3, or 30 ng/ml SCF. Following a 24-h incubation, [rneth~l-~Hlthyrnidine was added (2 pCi/ well) for the last 4 h. Alcohol/acid-precipitable radioactivity was determined by liquid scintillation counting.
Mice-BALB/c and CD2/Fl mice were obtained from the Experi- mental Animals Center of the Weizmann Institute of Science.
Buffered Solutions-HNTG buffer contained 20 mM HEPES (pH 7.5), 150 mM NaC1, 0.1% (w/v) Triton X-100, and 10% glycerol. Solubilization buffer contained 50 mM HEPES (pH 7.5), 150 mM NaC1, 1% (w/v) Triton X-100, 10% glycerol, 2 mM EDTA and 1 mM phenylmethylsulfonyl fluoride. High wash contained 50 mM HEPES (pH 7.5), 500 mM NaCl, 0.2% (w/v) Triton X-100, 0.1% SDS, and 5 mM EDTA. Medium wash was like high wash except 150 mM NaCl. Low wash contained 10 mM Tris (pH 8.0), 0.1% (w/v) Triton X-100, and 5 mM EDTA. PBS contained 137 mM NaCl, 2.7 mM KC1,7.9 mM NaHP04 and 1 mM KHZPO, (pH 7.2).
Generation of Monoclonal Antibodies to Human Kit/SCF Recep- tor-Kit-X was purified from the medium of transfected CHO cells by using lectin chromatography as we described previously (Lev et al., 1992~). Ten micrograms of the protein was injected intraperito- neally into BALB/c mice (2 months old). Two additional injections were given at intervals of 2 weeks. The antisera were tested 10 days after the third injection by using a direct binding assay to T-18 cells that overexpress c-kit. The spleens of mice that developed strong immune response were selected, and their splenocytes were fused with NSO myeloma cells by using polyethylene glycol (Galfre et al., 1987). Hybridomas were selected for growth in hypoxanthine/aminopterin/ thymidine medium, and their supernatants were screened by using an assay of antibody binding to T-18 cells. Positive wells of hybri- domas were tested further by an immunoprecipitation assay using either radiolabeled Kit-X or T-18 cell extract followed by Western blotting with a rabbit anti-Kit-X antibody. Hybridomas that scored positive were cloned twice by limiting dilut,ion. Large quantities of specific mAbs were produced by preparation of ascites fluid in CD2/ F, mice. mAbs were purified from ascites fluid by affinity chromatog- raphy on a Sepharose-protein A column that was equilibrated with 1.5 M glycine buffer (pH 8.9) containing 3 M NaC1. Elution of the antibodies from the column was done with 0.1 M citrate buffer (pH 5.0 for IgG, and pH 3.0 for other classes).
Construction and Expression of Mini-Kit-XProteins-To construct expression vectors that direct the synthesis of portions of the extra- cellular domain of Kit we used the previously described human Kit- X cDNA cloned into the Bluescript (pBS; Stratagene, La Jolla, CA) plasmid (Lev et al., 1992~). The kit 1-2 expression vector, containing the 5' 719 bp of human c-kit cDNA was constructed as follows. An EcoRI-ClaI fragment was cut out from the pBS-kit-X plasmid, and the resulting ends were filled in with Klenow I enzyme and ligated with T4 ligase. This resulted in juxtaposition of two in-frame stop codons 3' next to the c-kit region that encodes Ig-like domains 1 and 2. A BarnHI-XbaI fragment (704 bp) was cut out of the resulting plasmid and inserted into the pLSV-DHFR mammalian expression vector (Lev et al., 1992c) that was precut with XhoI and XbaI downstream of the internal SV40 early promoter. The kit 1-2-3 expression vector, containing 960 bp from the 5' end of human c-kit, was similarly constructed from pBS-kit-X plasmid that was cleaved with BarnHI and AsnI restriction endonucleases to yield a 960-bp fragment. This was ligated into a BamHI-SalI-cut shuttle vector (PATH2). After transformation of bacteria the resulting plasmid was cut with BamHI and ClaI, and the gel-purified fragment was ligated into pBS-kit-X vector from which the c-kit insert was removed by using BarnHI and ClaI. The resulting plasmid contained the 5' 960 bp of c-kit and an in-frame stop codon at its 3' end. A BamHI-XbaI fragment was then extracted from this plasmid and fused with a XhoI- and XbQI-CUt pLSV-DHFR vector. The mammalian expression vectors encoding Kit 1-2 and Kit 1-2-3 were then separately CO- transfected with a plasmid that confers neomycin resistance to dhfr- negative CHO cells (Urlaub and Chasin, 1980). Stably expressing cells were selected by their ability to grow in gentimycin (0.8 mg/ml) containing medium. Single-cell colonies were screened by biosyn- thetic labeling and immunoprecipitation of the corresponding [35s] methionine-labeled Kit protein.

SCF Binding Site of Kit 4401
Radiolabeling of Proteins-Affinity-purified rabbit antibodies to mouse F(ab'):! were labeled with "'1 by using the chloramine-T method (Hunter and Greenwood, 1962). SCF was labeled by using IODO-GEN (Pierce Chemical Co.) as follows. IODO-GEN-coated tubes (2 pg/50 pl of chloroform) were incubated with SCF (8 pg) and Nal2'I (1 mCi) for 15 min a t 22 "C. The reaction mixture was then subjected to gel filtration on a Sephadex G-25 (fine) column (0.6 X 7 cm) that was presaturated with bovine serum albumin.
Biosynthetic Labeling and Immunoprecipitation-Subconfluent monolayers of cells growing in six-well dishes (Costar) were washed with methionine-free Dulbecco's modified Eagle's medium and grown for 16 h in the same medium supplemented with 10% dialyzed calf serum and 50 pCi of [R'S]methionine/ml. The cell supernatants were spun for 10 min a t 4 "C in an Eppendorf centrifuge and subjected to immunoprecipitation. Protein A-conjugated Sepharose beads (3 mg/ sample) were suspended in HNTG buffer and incubated for 30 min a t 22 "C with either a polyclonal rabbit antiserum to Kit-X (10 pl) or with rabbit anti-mouse F(ab')2 (5 pg) . The latter was followed, after a brief wash, by incubation with mAbs (10 pg). The antibody-conju- gated beads were washed three times with HNTG and incubated with the cell supernatants for 2 h a t 4 "C. The immune complexes were washed twice with each of the high, medium, and low salt buffers. Then gel sample buffer was added to the washed immunoprecipitates, and they were heated a t 95 "C (5 min) and resolved by SDS-PAGE and autoradiography. In the case of lZ'I-labeled samples or samples that were not radioactively labeled, the immunoprecipitates were washed three times with HNTG.
Western Blotting-Washed immunoprecipitates were mixed with SDS gel sample buffer and subjected to SDS-PAGE. The gel-sepa- rated proteins were transferred electrophoretically onto nitrocellulose filters. Filters were first saturated for 1 h at 22 "C with blocking solution (10% low fat milk in 20 mM Tris-HC1 (pH 7.6) and 17 mM NaC1). Antisera were then added in the same solution, and the incubation was carried out for 1 h. For detection the filters were washed three times (5 min each wash) with PBS, 0.05% Tween 20 and reacted for 45 min at room temperature with horseradish perox- idase-conjugated protein A. The enzyme was removed by washing as above. The filters were reacted for 1 min with a chemiluminescence reagent (ECL, Amersham) and exposed to an autoradiography film for 1-15 min.
Binding Assays of Radiolabeled Ligands-The binding buffer con- tained Dulbecco's modified Eagle's medium supplemented with 0.1% bovine serum albumin and 20 mM HEPES-buffered at pH 7.5. The assays were carried out in 24-well dishes, and the monolayers were equilibrated in binding buffer 15 min prior to adding the radiolabeled ligand. For the determination of antibody binding the monolayers were incubated at 22 "C for 90 min with hybridoma supernatants or with various concentrations of the purified mAbs. Following a brief wash with PBS, 0.1% bovine serum albumin, 1251-labeled rabbit anti- mouse F(ab')? (10' cpm/well) was added, and the incubation was continued for an additional 90 min. Cell-bound radioactivity was determined by solubilization and harvest in 0.2 M NaOH and 0.1% SDS. For the determination of "'I-SCF binding cell monolayers were incubated a t 4 "C with 12'I-SCF for 2 h in the presence of various concentrations of unlabeled SCF. The amount of nonspecific binding was determined by performing the assay in the presence of a 100-fold excess of unlabeled SCF. At the end of the binding reaction the cells were washed, and their bound radioactivity was determined as de- scribed above. Ligand displacement analyses were performed in a similar way except that "'I-SCF (2 nM) was incubated with cellular monolayers for 4 h a t 4 "C in the presence of increasing concentra- tions of soluble Kit proteins.
Covalent Cross-linking of Radiolabeled SCF-Affinity-purified re- combinant proteins (Kit-X, Kit 1-2-3, Kit 1-2, and Neu-X) were incubated in PBS, 0.1% bovine serum albumin with "'I-SCF (100 ng/ml, lo5 cpm/ng) in the presence of various concentrations of unlabeled SCF or transforming growth factor-n. The total reaction volume was 0.06 ml. After 4 h a t 22 "C the proteins were subjected to covalent cross-linking by adding disuccinimidyl suberate to a 0.5 mM final concentration, and the reaction was allowed to continue for 40 min a t 22 "C. Gel sample buffer was then added, and the reaction was terminated by boiling and SDS-PAGE. For cross-linking of SCF to cultured cells the ligand was incubated with the cells for 2 h a t 4 "C, washed, and the ligand-receptor complexes were covalently cross- linked by adding EDAC (15 mM) in PBS and further incubation for 40 min a t 22 "C. The cells were then lysed in solubilization buffer, and the Kit receptor was immunoprecipitated as described above.
RESULTS
Generation of Ligand-competitive Monoclonal Antibodies to the KitISCF Receptor-In an attempt to determine the con- tribution of specific regions of the extracellular domain of Kit to the binding of SCF, we generated a panel of site-specific mouse mAbs to Kit by immunizing mice with a recombinant whole ectodomain of human Kit (Kit-X; Lev et al., 1992~). The capacity of selected hybridoma clones to recognize spe- cifically human Kit was tested by immunoprecipitation of unlabeled Kit-X and Western blotting with rabbit polyclonal antibodies (Fig. 1). The specificity of this blotting assay was indicated by the lack of signal in immunoprecipitates of the extracellular portion of p185"" (denoted Neu-X), which was used as a control antigen. To confirm the specificity of the mAbs to p145"', they were assayed for binding to c-kit- transfected CHO cells (cell line T-18). All of the antibodies that immunoprecipitated Kit-X also recognized T-18 cells but did not react with the parental untransfected CHO cells (data not shown). The results of the binding of two mAbs, K44 and K45, to c-kit-overexpressing cells are depicted in Fig. 2A.
Evidently both antibodies displayed saturable surface binding. To test the possibility that some of the mAbs that we
generated are directed to the ligand recognition site of Kit, the binding of l2'II-SCF to T-18 cells was determined in the presence or absence of each mAb. Fig. 2B depicts the results that were obtained with mAbs K44 and K45. In contrast to the K45 antibody, which did not affect binding of lZ5I-SCF even a t high antibody concentrations, mAb K44 stoichiomet- rically inhibited ligand binding at an approximately 1:l ratio of antibody to SCF. Only one other antibody, K57, similarly affected the binding of SCF (data not shown), implying that both mAbs K44 and K57 are directed to the SCF binding site or to a nearby determinant. To support this possibility further we determined the effect of these mAbs on covalent cross- linking of Iz5I-SCF to ~ 1 4 5 ~ ' ' . In the absence of mAbs two ligand-receptor complexes could be immunoprecipitated from "-18 cells (Fig. 3A). These complexes correspond to receptor monomers and dimers (Lev et al., 1992a). However, incubation of the cells with "'1-SCF in the presence of K44 or K57 mAb completely abolished both cross-linked receptor forms (Fig. 3A). In contrast, mAbs that did not inhibit SCF binding to T-18 cells (e.g. K45 and K49) exerted only limited effects on cross-linking of '"I-SCF to the receptor. Similarly, mAbs K44 and K57 did not immunoprecipitate preformed covalent com- plexes of '"I-SCF with ~ 1 4 5 ~ ' ' (Fig. 3B), although they did recognize the Kit-X protein (Fig. 1B) and the full-length
Kit- X Neu-X
f 116
t 85 I6 Ab: Anti Kit-X (rabbit)
FIG. 1. Selection of hybridomas producing Kit-specific an- tibodies. Supernatants of individual clones of hybridoma cells were incubated with Kit-X protein (approximately 200 ng) or with Neu-X protein (200 ng) as indicated. After gel electrophoresis and protein transfer to a nitrocellulose filter, the blot was probed with a polyclonal rabbit antibody to Kit-X followed by horseradish peroxidase-labeled protein A. Chemiluminescence (ECL kit, Amersham) and autoradi- ography (5-min exposure) were used to obtain the shown autoradi- ogram. IP, immunoprecipitated; IB, immunoblotted.

4402 SCF Binding Site of Kit
[Ab] ; nM
[Ab]; nM
FIG. 2. Antibody binding to Kit-overexpressing cells and competition with SCF. Panel A , confluent monolayers of T-18 cells, a CHO subline that overexpresses the human Kit protein (Lev et al., 1991), were incubated for 90 min a t 22 "C with different concentrations of mAb K44 (closed circles) or K45 (open squares). After a brief washing step the monolayers were further incubated for 90 min a t 22 "C with radiolabeled rabbit Ig specific to mouse IgG. This was followed by extensive washing of the unbound radioactivity and counting the cell-associated radioactivity in a y-counter. Panel B, confluent monolayers of T-18 cells were incubated with lZ5I-SCF (2.5 ng/ml, lo5 cpm/ng) in the presence of different concentrations of mAb K44 (closed circles) or K45 (open squares). Following 4 h of binding at 4 "C the monolayers were washed extensively with ice-cold PBS, and the cell-associated radioactivity was determined in a y- counter. Nonspecific binding of '251-SCF was determined by perform- ing the binding reaction in the presence of a 100-fold excess concen- tration of unlabeled SCF, and it was subtracted from the total amount of bound radioactivity. The results shown are representative of two experiments.
receptor (data not shown) when it was not cross-linked to the ligand. On the basis of these experiments we concluded that mAbs K44 and K57 recognize determinants that are close to the SCF binding site of ~ 1 4 5 ~ " , unlike other antibodies that interact elsewhere on the receptor molecule. In separate ex- periments we addressed the possibility that K44 and K57 are directed against the same epitope. To this end, we radiolabeled K44 and analyzed its displacement from Kit-expressing cells by native SCF or K57 mAb. Interestingly, SCF efficiently displaced surface-bound '251-K44, but K57 was ineffective a t concentrations that inhibited SCF binding to T-18 cells (data not shown). We therefore concluded that the epitopes of mAbs K44 and K57 are nonoverlapping.
The capacity of mAb K44 to inhibit SCF binding to Kit was examined in uitro by using the M07e human megakar- yoblastic cell line. These cells display absolute dependence for survival and proliferation on GM-CSF and IL-3 (Avanzi
I P Ab! Anti-Klt-X ( rabbi t )
FIG. 3. Effect of monoclonal antibodies on cross-linking of ''9-SCF to Kit and immunoprecipitation of the ligand-recep- tor complexes. Monolayers of T-18 cells (approximately lo6 cells) were incubated with 2 nM '*'I-SCF in the presence of the indicated mAbs a t 2 pg/ml (panel A ) or without antibodies (panel B ) . Following 2 h a t 4 "C the cells were washed, and the ligand-receptor complexes were covalently cross-linked by a 40-min incubation a t 22 "C with 15 mM EDAC. The complexes were then immunoprecipitated (ZP) with a polyclonal antibody to Kit-X (panel A ) or with the indicated mAbs (panel B ) and analyzed by electrophoresis in 5% acrylamide gels followed by autoradiography (24 h a t -70 "C with an intensifier screen). The locations of molecular mass marker proteins are indi- cated by bars, and the corresponding masses are given in kDa. Note that both dimers and monomers of Kit were resolved.
TABLE I Effect of anti-Kit mAbs on DNA synthesis of M07e megakaryocyte
cells M07e cells (50,00O/well) were deprived of growth factors 24 h
before starting the experiment. Antibodies were added to the cells 50 min prior to the addition of growth factors. The incorporation of [3H] thymidine into DNA was determined by a 4-h pulse that was given after 20 h of incubation a t 37 "C with antibodies and growth factors. The results given are averages of triplicate determinations in which the variation did not exceed 10%. The experiment was repeated twice.
Treatment ['HIThymidine incorporated
Deprived cells SCF (30 ng/ml) GM-CSF (10 ng/ml) K44
10 5 d m 1
SCF (30 ng/ml) + K44 0.5 pg/ml 5 d m 1
10 d m 1
5 rg/ml 10 ccg/ml
5 d m 1
GM-CSF (10 ng/ml) + K44 (10 pg/ml) K49
SCF (30 ng/ml) + K49 0.5 pg/ml
10 u d m l
cpm 1,463
57,966 59,632
1,069 1,196
3,536 2,622 2,977
68,587
1,552 2,773
50,715 39,383 28.667
et ai., 1990). More recently it has been reported that M07e cells can proliferate also in the presence of SCF (Hendrie et al., 1991). When tested on M07e cells, mAb K44 completely inhibited SCF-induced incorporation of [3H]thymidine into macromolecules (Table I). However, K44 had no effect on the induction of DNA synthesis by GM-CSF (Table I) or IL-3 (data not shown). As expected, mAb K49, which does not affect SCF binding, exerted only a limited inhibitory effect on SCF-stimulated M07e cells.
Construction and Expression of Two Deletion Mutants of the Extracellular Portion of Kit-In an attempt to localize the epitopes of mAbs K44 and K57 that are apparently located close to the SCF binding cleft, we constructed two deletion mutants of the Kit-X protein. The unique endonuclease re- striction sites EcoRI and AsnI of the human c-Kit cDNA

SCF Binding Site of Kit 4403
(Yarden et al., 1987) were used to introduce stop codons a t these sites, thereby terminating translation a t either the amino- or the carboxyl-terminal side of the third Ig domain of Kit (Fig. 4). The modified cDNAs were ligated separately into an eukaryotic expression vector that contained the dhfr gene as a selectable marker. These plasmids were used to transfect dhfr-deficient CHO cells (Urlaub and Chasin, 1980), and drug-resistant colonies were grown individually. Clones that overexpressed the shorter deletion mutant, containing Ig domains 1 and 2 (denoted Kit 1-2), or the longer protein that contained also domain 3 (Kit 1-2-3), were selected by using an immunoprecipitation assay. A representative immunopre- cipitation analysis of the selected clones is shown in Fig. 5. Monolayers of the selected cell lines and a control untrans- fected cell line were incubated for 16 h with [35S]methionine. The biosynthetically labeled Kit proteins that were secreted into the growth medium were then immunoprecipitated with a polyclonal rabbit antibody that was raised against the whole extracellular domain. As shown in Fig. 5, a 36-kDa protein was detectable in the medium of cells that were transfected with the kit 1-2 plasmid, whereas a 65-kDa protein was secreted by kit 1-2-3-transfected cells. These molecular masses are in agreement with the expected sizes of the recom- binant glycosylated proteins (Fig. 4 and Qiu et al., 1988). In addition to the indicated p36 and p65 proteins, two other bands of 120 and 180 kDa appeared in the immunoprecipi- tates. We attribute these molecules to nonspecific interactions with the polyclonal antiserum because they were precipitated also from untransfected cells (Fig. 5).
Ligand Inhibitory mAbs Recognize the Recombinant Kit Proteins-In the next step we analyzed the ability of Kit- specific mAbs to recognize Kit 1-2-3 and Kit 1-2 proteins by using an immunoprecipitation assay. As shown in Fig. 6, Kit 1-2-3 was recognized by four mAbs, including the ligand- competitive K44 and K57 antibodies. In addition, these two antibodies could immunoprecipitate the shorter mutant (Fig. 6, right panel), implying that their epitopes are confined to Ig
Kit 1-2-3
Kit TK2
FIG. 4. Structural representation of the Kit protein and recombinant forms of its extracellular domain. The various proteins are schematically presented, and the Ig-like domains are shown as loops. The proteins are drawn to scale, and the designations of their functional domains are indicated at the bottom. Also shown are the locations of the unique sites of restriction endonucleases that were used to construct the recombinant proteins. Numbers 1-5 indi- cate the respective Ig-like domains. The locations of all of the cysteine residues of the extracellular domains are indicated by a circle and C. Tryptophan residues that belong to the consensus structure of the Ig homology unit are shown by a circle and W. SP, signal peptide; TK, tyrosine kinase.
Kit-X 4' 4
J I P Ab: Anti Kit-X(rabbit)
FIG. 5. Expression of portions of the extracellular domain of Kit in CHO cells. cDNAs corresponding to the whole extracellular domain of Kit (denoted Kit-X) or to portions of it (denoted Kit 1-2 and Kit 1-2-3; see Fig. 4) were cloned into the pLSV-DHFR eukaryotic expression vector and used to transfect cultured CHO cells. Clones of cells that overexpressed the transfected genes were selected and grown separately. Biosynthetically labeled supernatants from lo6 cells were harvested after a 16-h incubation of the indicated cell lines with ["S]methionine. The Kit proteins were immunoprecipitated from the cleared supernatants by using a rabbit polyclonal antibody to Kit-X and visualized by SDS-PAGE and autoradiography (48 h at -70 "C with an intensifier screen). The locations of the Kit proteins are indicated by arrows. Also shown are the locations of molecular mass marker proteins (in kDa). None refers to control untransfected CHO cells.
Kit 1-2-3
I8 Ab: Anti Kit-X ( rabbi t )
FIG. 6. Recognition of Kit 1-2 and Kit 1-2-3 by different monoclonal antibodies. Supernatants of CHO cell lines that over- express Kit 1-2-3 or Kit 1-2 (as indicated) were harvested and concentrated 20-fold in an ultrafiltration cell (Amicon). For control we used a CHO cell line that secretes the extracellular domain of the Neu protein (labeled Neu-X). Aliquots of 0.5 ml of each supernatant were then subjected to immunoprecipitation (ZP) with the indicated mAbs. The immunocomplexes were resolved by SDS-PAGE (8.5% acrylamide), transferred onto nitrocellulose, and immunoblotted (ZB) with a rabbit polyclonal antibody to Kit-X. Detection was performed by using chemiluminescence and autoradiography (left panel, 1 min; right panel, 15-min exposure of the x-ray film). An arrow marks the location of Kit 1-2 protein, and bars indicate the locations of molec- ular mass marker proteins.
domains 1 and 2 and the intervening sequences. Interestingly, mAbs K27 and K69 did not react with Kit 1-2-3, suggesting that their recognition sites are distal to the first three Ig domains of Kit. Both K45 and K49 mAbs, which do not interfere with ligand binding, recognized Kit 1-2-3 and also Kit 1-2 (Fig. 6 and data not shown). In conclusion, the portion of the Kit protein which is included in the short deletion mutant Kit 1-2 contains the binding sites of mAbs K44 and K57, and by extension also the SCF binding site or part of it.
Inhibition of Binding of Radiolabeled SCF to Wild-type Kit by Soluble Recombinant Proteins-Because of the soluble nature of the recombinant truncation mutants of Kit, simple ligand binding assays were not feasible. In addition, immobi- lization by using antibodies could interfere with the binding assay. We therefore tested the ability of soluble Kit proteins
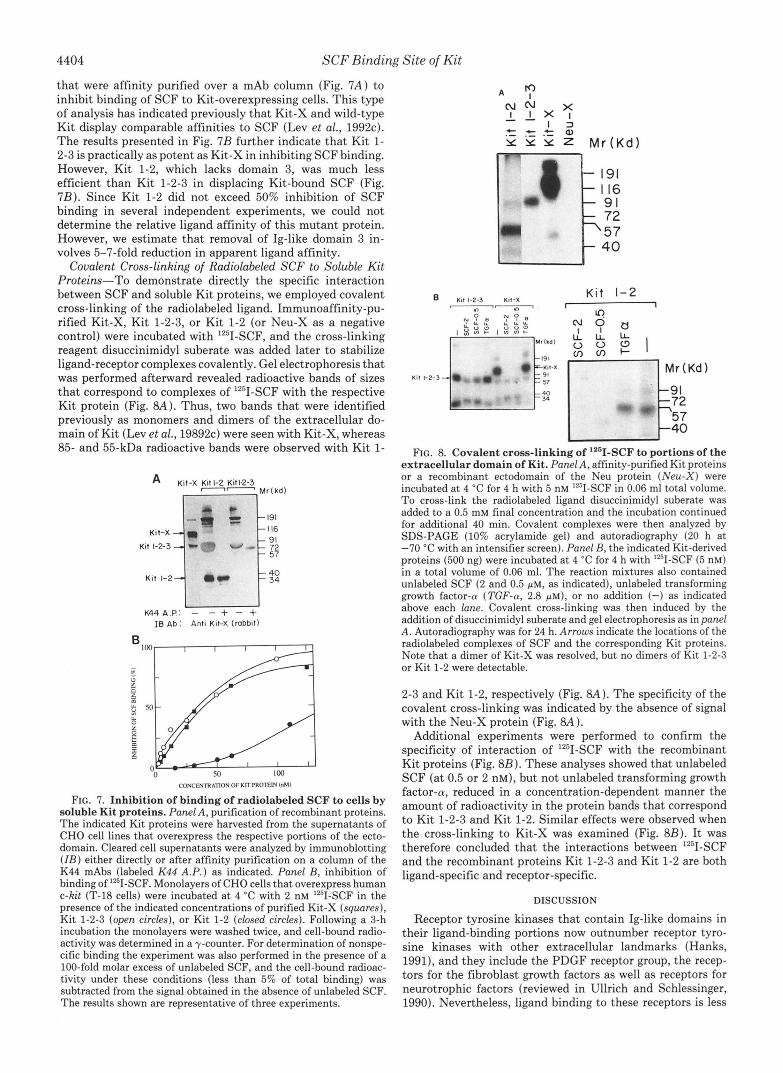
4404 SCF Binding Site of Kit
that were affinity purified over a mAb column (Fig. 7A) to inhibit binding of SCF to Kit-overexpressing cells. This type of analysis has indicated previously that Kit-X and wild-type Kit display comparable affinities to SCF (Lev et al., 1992~). The results presented in Fig. 7B further indicate that Kit 1- 2-3 is practically as potent as Kit-X in inhibiting SCF binding. However, Kit 1-2, which lacks domain 3, was much less efficient than Kit 1-2-3 in displacing Kit-bound SCF (Fig. 7B). Since Kit 1-2 did not exceed 50% inhibition of SCF binding in several independent experiments, we could not determine the relative ligand affinity of this mutant protein. However, we estimate that removal of Ig-like domain 3 in- volves 5-7-fold reduction in apparent ligand affinity.
Covalent Cross-linking of Radiolabeled SCF to Soluble Kit Proteins-To demonstrate directly the specific interaction between SCF and soluble Kit proteins, we employed covalent cross-linking of the radiolabeled ligand. Immunoaffinity-pu- rified Kit-X, Kit 1-2-3, or Kit 1-2 (or Neu-X as a negative control) were incubated with '251-SCF, and the cross-linking reagent disuccinimidyl suberate was added later to stabilize ligand-receptor complexes covalently. Gel electrophoresis that was performed afterward revealed radioactive bands of sizes that correspond to complexes of IZ5I-SCF with the respective Kit protein (Fig. 8A). Thus, two bands that were identified previously as monomers and dimers of the extracellular do- main of Kit (Lev et al., 19892~) were seen with Kit-X, whereas 85- and 55-kDa radioactive bands were observed with Kit 1-
A Kit-X Kit 1-2 Kit1.2-3 -- Mr(kd)
I-
Kit 1-2 i rn- 57 72
K44A.P: - - 4- - -t IB Ab: Anti Kit-X (robbit)
CONCENlRAllON OF KTT P R O E I N ("MI
FIG. 7. Inhibition of binding of radiolabeled SCF to cells b y soluble Kit proteins. Panel A, purification of recombinant proteins. The indicated Kit proteins were harvested from the supernatants of CHO cell lines that overexpress the respective portions of the ecto- domain. Cleared cell supernatants were analyzed by immunoblotting ( I B ) either directly or after affinity purification on a column of the K44 mAbs (labeled K44 A.P.) as indicated. Panel B, inhibition of binding of lZ5I-SCF. Monolayers of CHO cells that overexpress human c-kit (T-18 cells) were incubated at 4 "C with 2 nM "'1-SCF in the presence of the indicated concentrations of purified Kit-X (squares), Kit 1-2-3 (open circles), or Kit 1-2 (closed circles). Following a 3-h incubation the monolayers were washed twice, and cell-bound radio- activity was determined in a y-counter. For determination of nonspe- cific binding the experiment was also performed in the presence of a 100-fold molar excess of unlabeled SCF, and the cell-bound radioac- tivity under these conditions (less than 5% of total binding) was subtracted from the signal obtained in the absence of unlabeled SCF. The results shown are representative of three experiments.
B
Kif 1-2-3
K i t 1-2 - Mr (Kd 1 p; 57
FIG. 8. Covalent cross-linking of "'I-SCF to portions of the extracel lular domain of Kit. Panel A, affinity-purified Kit proteins or a recombinant ectodomain of the Neu protein (Neu-X) were incubated a t 4 "C for 4 h with 5 nM '2'I-SCF in 0.06 ml total volume. T o cross-link the radiolabeled ligand disuccinimidyl suberate was added to a 0.5 mM final concentration and the incubation continued for additional 40 min. Covalent complexes were then analyzed by SDS-PAGE (10% acrylamide gel) and autoradiography (20 h a t -70 "C with an intensifier screen). Panel B, the indicated Kit-derived proteins (500 ng) were incubated at 4 "C for 4 h with "'I-SCF (5 nM) in a total volume of 0.06 ml. The reaction mixtures also contained unlabeled SCF (2 and 0.5 p ~ , as indicated), unlabeled transforming growth factor-cu (TGF-a, 2.8 p ~ ) , or no addition (-) as indicated above each lane. Covalent cross-linking was then induced by the addition of disuccinimidyl suberate and gel electrophoresis as in panel A. Autoradiography was for 24 h. Arrows indicate the locations of the radiolabeled complexes of SCF and the corresponding Kit proteins. Note that a dimer of Kit-X was resolved, but no dimers of Kit 1-2-3 or Kit 1-2 were detectable.
2-3 and Kit 1-2, respectively (Fig. 8A). The specificity of the covalent cross-linking was indicated by the absence of signal with the Neu-X protein (Fig. 8A).
Additional experiments were performed to confirm the specificity of interaction of '251-SCF with the recombinant Kit proteins (Fig. 8B). These analyses showed that unlabeled SCF (at 0.5 or 2 nM), but not unlabeled transforming growth factor-0, reduced in a concentration-dependent manner the amount of radioactivity in the protein bands that correspond to Kit 1-2-3 and Kit 1-2. Similar effects were observed when the cross-linking to Kit-X was examined (Fig. 8B). It was therefore concluded that the interactions between Iz5I-SCF and the recombinant proteins Kit 1-2-3 and Kit 1-2 are both ligand-specific and receptor-specific.
DISCUSSION
Receptor tyrosine kinases that contain Ig-like domains in their ligand-binding portions now outnumber receptor tyro- sine kinases with other extracellular landmarks (Hanks, 1991), and they include the PDGF receptor group, the recep- tors for the fibroblast growth factors as well as receptors for neurotrophic factors (reviewed in Ullrich and Schlessinger, 1990). Nevertheless, ligand binding to these receptors is less

SCF Binding Site of Kit 4405
understood than in the case of the EGF receptor and the insulin receptors. By using chimeras between chicken and human EGF receptors (Lax et al., 1989) and direct peptide mapping of the binding sites of lZ5I-EGF (Lax et al., 1988; WU et al., 1990) and EGF-competitive mAbs (Wu et al., 1989), it was shown that the receptor’s portion that lies between the two cysteine-rich sequences specifies ligand binding. How- ever, it appears that the amino-terminal portion of the EGF receptor also participates in the formation of the ligand bind- ing cleft. Similarly, the binding of insulin and insulin-like growth factor-1 to their related receptors appears to involve both the amino-terminal portions of the receptors (Wedekind et al., 1989; Kjeldsen et al., 1991) and their cysteine-rich domains (Gustafson and Rutter, 1990; Yip et al., 1991). The latter may function as a core binding cleft whereas ligand specificity is probably determined by adjacent sequences.
Interestingly, studies of ligand interactions with receptor tyrosine kinases that contain Ig-like motifs also indicate that noncontiguous receptor segments may create the ligand bind- ing sites of such receptors. In addition, determinants of spec- ificity to different ligands may not coincide. For example, the amino-terminal three Ig-like domains of the PDGF receptor confer the ability to bind different PDGF isoforms (Heidaran et al., 1990), but specificity of the a receptor to PDGF-AA resides in the second Ig-like domain and it is distinct from the PDGF-BB specificity determinant (Heidaran et al., 1992). Likewise, the carboxyl-terminal half of the third Ig-like do- main of variants of the fibroblast growth factor receptors determines ligand specificity to the keratinocyte growth factor or basic fibroblast growth factor, but binding of acidic fibro- blast growth factor is determined elsewhere in the receptor (Werner et al., 1992; Miki et al., 1992; Yayon et al., 1992).
By using SCF-competitive mAbs we localized the ligand binding site to the two amino-terminal Ig-like domains of Kit (Fig. 3). However, a recombinant protein that contained only the first two Ig-like domains (Kit 1-2) displayed reduced affinity to SCF, as compared with the whole ectodomain of Kit or a soluble protein encompassing the three amino-ter- minal Ig-like domains (Fig. 7B). It is therefore conceivable that part of the high affinity binding site resides in the receptor’s portion that is defined by the amino-terminal two Ig-like domains, but determinants which are included in the third Ig loop are also involved in the formation of the ligand binding cleft. Covalent cross-linking experiments (Fig. 8) also support our conclusion that the major portion of the SCF binding cleft is confined to the first and second Ig-like do- mains of Kit. However, this type of experiment cannot resolve differences in ligand binding affinities.
An independent support to the amino-terminal localization of the SCF binding site was obtained recently in our labora- tory by utilizing chimeric human-mouse Kit protein^.^ Since murine Kit does not bind human SCF, it was possible to localize the binding site of human SCF through replacement of individual Ig-like loops of mouse Kit by the corresponding parts of human Kit. This approach identified the second Ig- like domain of human Kit as the core of the binding site but indicated that the adjacent Ig domains are also involved in ligand recognition. Our preliminary data further suggest that the effect of domain 3 is caused by a decrease in the rate of ligand dissociation. Presumably, SCF binding to the second Ig-like domain of Kit is followed by a conformational change that allows the third domain to fold over the binding cleft and thereby inhibit ligand dissociation. Unfortunately, our soluble Kit proteins do not provide a convenient system in
S. Lev, J. M. Blechman, S-I. Nishikawa, D. Givol, and Y. Yarden, submitted for publication.
which to test this model, and we currently employ chimeric Kit proteins to address the mode of involvement of individual domains.
The possibility that interdomain packing, rather than a contiguous protein sequence, defines the ligand binding site of Kit, is consistent with our observation that the SCF- competitive mAbs (K44 and K57) recognize conformation- dependent epitopes of Kit (data not shown). The amino- terminal location of the binding site is also reminiscent of the finding that domain 2 of the aPDGF receptor confers speci- ficity to PDGF-AA (Heidaran et al., 1992). Interestingly, the first and the second Ig-like domains of another receptor with five such motifs, the intercellular cell adhesion molecule 1, contain the binding site for the integrin LFA-1 and also for a subgroup of human rhinoviruses (Staunton et al., 1990). Pre- sumably, distal localization of the binding sites for soluble ligands ( i e . SCF and PDGF), integrin, or a bulky virus makes these cellular receptors accessible to interactions.
In conclusion, our present study confined the ligand binding site of Kit to its amino-terminal portion and implicated the involvement of noncontiguous protein sequences in SCF rec- ognition. Further dissection of the amino-terminal half of the ectodomain of Kit by internal deletions should make it pos- sible to define more precisely the interaction between SCF and Kit. Alternatively, crystallization of the recombinant Kit proteins, and especially Kit 1-2-3, may provide molecular insights into the binding of SCF to its target cells.
Acknowledgments-We thank Dr. Kris Zsebo (Amgen Center) for recombinant human SCF, Magda David for help in the generation of mAbs, and Miriam Fagan for typing this manuscript.
REFERENCES Amzel, L. M., and Poljak, R. J. (1979) Annu. Reu. Biochem. 48,961-967 Avanzi, G. C., Brizzi, M. F., Giannotti, J., Ciarletta, A., Yang, Y., Pegoraro. L.,
Bennett, D. (1956) J. Morphol. 9 8 , 199-234 Besmer, P. (1991) Curr. Opin. Cell Biol. 3,939-946 Blume-Jensen P., Claesson-Welsh L., Siegbahn, A. Zsebo, K. M., Westermark,
Chabot, B., Stephenson, D. A., Chapman, V. M., Besmer, P., and Bernstein, A. B., and Heldin, C.-H. (1991) EdBO J. 10,412114128
Copeland, N. G., Gilbert, D. J., Cho, B. C., Donovan, P. J., Jenkins, N. A,, (1988) Nature 335,8&89
Coseman, D., Anderson, D., Lyman, S. D., and Williams, D. E. (1990) Cell
Duan, D . 3 R., Pazin, M. J., Fretto, L. J., and Williams, L. T. (1991) J. Biol. Chern. 266,413-418
Funasaka, Y., Boulton, T., Cobb, M., Yarden, Y., Fan, B., L man, S. D., Flanagan, J. G., Chan, D., and Leder, P. (1991) Cell 64,1025-1035
Williams, D. E., Anderson, D. M., Zakut, R., Mishima, Y., an8Halaban. R. (1992) Mol. Biol. Cell 3 , 197-209
Galfre, G., Howe, S. C., Milstein, C., Butcher, G. W., and Howard, J. C. (1987) Nature 266,550-552
Geissler, E. N., Ryan, M. A., and Housman, D. E. (1988) Cell 55,185-192 Gustafson, T. A., and Rutter, W. J. (1990) J. Biol. Chern. 265,18663-18667 Hanks, S. K. (1991) Curr. Opin. S t ru t . Biol. 1,369-383 Heidaran, M. A., Pierce, J. H., Jensen, R. A., Matsui, T., and Aaronson, S. A.
Heidaran, M. A., Yu, J.-C., Jensen, R. A., Pierce, J. H., and Aaronson, S. A.
Hendrie, P. 0.. Miyazawa, K., Yang, Y. C., Langefeld, C. D., and Broxmeyer,
Huang, E., Nocka, K., Beier, D. R., Chu, T. Y., Buck, J. Lahm, H. W., Wellner,
Hunkapiller T., and Hood, L. (1989) Adu. Immunol. 44 , l -63 Hunter, W. M., and Greenwood, F. C. (1962) Nature 194,495-496 Kjeldsen, T., Andersen, A. S., Wiberg, F. C., Rasmussen, J. S., Schaffer, L.,
Sci. U. S. A. 88,4404-4408 Balschmidt, P., Moller, K. B., and Moller, N. P. H. (1991) Proc. Natl. Acad.
Lax, I., Burgess, W. N., Bellot, F., Ullrich, A., Schlessinger, J., and Givol, D. (1988) Mol. Cell. Biol. 8, 1831-1834
Lax, I., Bellot, F., Howk, R., Ullrich, A., Givol, D., and Schlessinger, J. (1989) EMBO J. 8,421-427
Lev, S., Givol, D., and Yarden, Y. (1991) EMBO J. 10,647-654 Lev, S., Yarden, Y., and Givol, D. (1992a) J. Biol. Chern. 267,15970-15977 Lev, S., Givol, D., and Yarden, Y. (1992h3 Proc. Natl. Acad. Sci. U. S. A. 8 9 ,
Lev, S., Yarden, Y., and Givol, D. (199%) J. Biol. Chern. 2 6 7 , 10866-10873 McCulloh, E. A., Siminovitch, L., and Till, J. E. (1964) Science 144 , 844-846 McCulloh, E. A., Siminovitch, L., Till, J. E., Russell, E. S., and Bernstein, S.
E. (1965) Blood 26,399-410 Miki, T., Bottaro, D. P., Fleming, T. P., Smith, C. L., Burgess, W. H., Chan,
A. M.-L., and Aaronson, S. A. (1992) Proc. Natl. Acad. Sci. U. S. A. 8 9 , 246- 250
and Clark, S. C. (1990) J. Cell. Physiol. 146,458-464
63, 175-183
(1990) J. Biol. Chern. 265,18741-18744
(1992) J. Biol. Chern. 267,2884-2887
H. E. (1991) Exp. Hernatol. 19, 1031-1037
D., Leder, P., and Besmer, P. (1990) Cell 63,225-23’3
678-682

4406 SCF Binding Site of Kit Mintz, B., and Russell, E. S. (1957) J. Exp. Zool. 134,207-237 Park, L. S., Martin, U., Mochizuki, D. Y., Boswell, H. S., Burgess, G. S., Nocka, K., Buck, J., Levi, E., and Besmer, P. (1990) EMBO J. 9,3287-3294 Qiu, F., Ray, P., Brown, K., Barker, P. F., Jhanwar, S., Ruddle, R. H., and Wu, D., Wang L., Sato, G. H., West, K. A., Harris, W. R., Crabb, J. W., and
Coseman, D., and Steward, D. L. (1990) Cell 63,167-174
Rottapel, R., Reedijk, M., .Williams, D. E., Lyman, S. D., Anderson, D. M., Wu, D., Wang, L., Chi, Y., Sato, G. H., and Sato, J. D. (1990) Proc. Natl. Acad. Sato, J. D. (i989) J. Bioi. Chem. 264,17469-17475
Russell, E. S. (1979) Adu. Genet. 20,357-459 Silvers, W. K. (1979) The Coat Colors of Mice: A Model for Gene Action and Yarden, Y., Kuang, W. J., Yang-Feng, T., Coussens, L., Munemitsu, S., Dull,
Staunton, D. E., Dustin, M. L., Erickson, H. P., and Springer, T. A. (1990) Cell J. 6,3341-3351 T. J., Chen, E., Schlessinger, J., Francke, U., and Ullrich, A. (1987) EMBO
Besmer, P. (1988) EMBO J. 7 , 1003-1011
Pawson, T., and Bernsteln, A. (1991) Mol. Cell Bzol. 11,3043-3091 Sci. U. S. A. 87,3151-3155 Yarden, Y., and Ullrich, A. (1988) Annu. Reu. Bwchem. 57,443-478
Interaction, pp. 206-241, Springer Verlag, New York
61,243-254 Ullrich, A,, and Scblessinger, J. (1990) Cell 6 1 , 203-212 Urlaub, G., and Chasin, L. A. (1980) Proc. Natl. Acad. Sei. U. S. A. 77,4216- Yip, C. C., Grunfeld, C., and Goldfine, I. (1991) Ekehemistry 30,695-701
Yayon, A., Zimmer, Y., Guo-Hong, S., Avivi, A., Yarden, Y., and Givol, D. (1992) EMBO J. 11 , 1885-1890
4220
Brandenburg, D. (1989) BwL Chem. Hoppe-Segler 370,251-258 Wedekind, F., Baer-Pontzen, K., Bala-Mohan, S., Choli, D., Zahn, H., and B., Buchdev, R. K., Yushenkoff, V. N., Birkett, N. C., Williams, L. R.,
Zsebo, K. M., Wypych, J., McNiece, I. K., Lu, H. S., Smith, K. A., Karkare, S.
Werner, S.. Duan, D.3. R.. de Vries. C.. Peters. K. G.. Johnson. D. E.. and K. E. (1990a) Cell 63. 195-201 Satyagul, N. V., Tung, W., Bosselmann, R. A,, Mediaz, E. A., and Langley,
Williams, L. T. (1992) Mol. Cell. Bid. i2 ,82-88 Williams, A. F., and Barclay, A. N. (1988) Anno. Reu. Immunol. 6,381-405
Zsebo, K. M., Williams, D. A., Geissler, E. N., Broudy, V. C., Martin, F. H.,
Williams, D. E., Eisenman, J., Baird, A., Rauch, C., Ness, K. V., March, C. J., (1990b) Cell 6 3 , 213-224 Atkins, H. L., Hsu, R. Y., Birkett, N. C., Okino, K. H., and Murdock, D. C.





























