Embed Size (px)
Citation preview

This article was downloaded by: [Emory University]On: 19 August 2013, At: 07:36Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH,UK
Tropical ZoologyPublication details, including instructions forauthors and subscription information:http://www.tandfonline.com/loi/ttzo20
Social differentiation anddivision of labour in thequeenless ant Platythyreaschultzei Forel 1910(Hymenoptera Formicidae)Martin H. Villet aa Department of Zoology, University of theWitwatersrand, P.O. Wits, 2050, South AfricaPublished online: 01 Aug 2012.
To cite this article: Martin H. Villet (1991) Social differentiation and divisionof labour in the queenless ant Platythyrea schultzei Forel 1910 (HymenopteraFormicidae), Tropical Zoology, 4:1, 13-29, DOI: 10.1080/03946975.1991.10539472
To link to this article: http://dx.doi.org/10.1080/03946975.1991.10539472
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all theinformation (the “Content”) contained in the publications on our platform.However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness,or suitability for any purpose of the Content. Any opinions and viewsexpressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of theContent should not be relied upon and should be independently verified withprimary sources of information. Taylor and Francis shall not be liable for anylosses, actions, claims, proceedings, demands, costs, expenses, damages,and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of theContent.
This article may be used for research, teaching, and private study purposes.Any substantial or systematic reproduction, redistribution, reselling, loan,

sub-licensing, systematic supply, or distribution in any form to anyone isexpressly forbidden. Terms & Conditions of access and use can be found athttp://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013

Tropical Zoology 4: 13-29, 1991
Social differentiation and division of labour in the queenless ant Platythyrea schultzeiForel1910 (Hymenoptera Formicidae)
MARTIN H. VILLET
Department of Zoology, University of the Witwatersrand, P. 0. Wits 2050, South Africa
Received 15 December 1989, accepted 30 October 1990
Colonies of Platythyrea schultzei Forel 1910 are obligately queenless, containing only 10-40 morphologically and anatomically similar, monomorphic workers. The role of the queen is filled by a single inseminated worker in each colony. The ovaries of this worker are physiologically active, while oogenesis is repressed in its nestmates.
Colony labour is organised into five groups of tasks: care of eggs, care of larvae, care of cocoons, and general colony maintenance, including foraging. The mated worker monopolize the task of caring for the eggs. Organisation of the remaining ants to carry out the other tasks shows a clear pattern of age polyethism, through which all workers except the mated worker pass. There is no indication that division of labour is based on or biased by size differences amongst the workers.
The behavioural marker (carrying eggs) for the mated worker allows its experimental removal from each colony. In each case its role is taken over by a single, young, uninseminated worker that undergoes ovarial development and assumes the same behavioural characteristics and control over its nestmates' oogenesis as the previous mated worker. Thus insemination does not appear to be a prerequisite for the physiological differentiation of an individual to fill the role of reproductive.
KEY WORDS: Ponerinae, queenlessness, polyethism, reproduction, morphology, milking larvae.
Introduction Materials and methods Results
Morphological differentiation Reproductive differentiation Behavioural differentiation
Discussion Acknowledgements References
14 14 16 16 18 18 24 27 27
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013

14 M.H. Villet
INTRODUCTION
Queenlessness is an important phenomenon in ants because it implies the loss of a developmental pathway that once gave rise to the morphologically distinct, reproductive queen caste, a fundamental characteristic of eusocial ant societies. Its advent creates the need for changes in the behavioural roles normally filled by the worker caste, and in particular their role in reproduction. Decoupling of behaviour and morphology in the worker caste has been recorded in many ants (CALABI 1988), but there is evidence that there may be some limit to the ability of one caste to assume the role of another (WILSON 1984). Queenlessness might also affect the division of labour, another characteristic of eusociality.
Queenless societies in which mated workers assume the role of the queen have been reported in a number of species of ponerine ants in the tribes Ectatommini and Ponerini (WARD 1981, PEETERS & CREWE 1985, WILDMAN & CREWE 1989, FuKuMOTO et al. 1989, PEETERS & HIGASHI 1989, WARE et al. 1990), and this form of social organisation appears to be unique to the subfamily. Queenlessness has been reported in the myrmicine ants Pristomyrmex pungens Mayr 186(> (TSUJI 1988) and Megalomyrmex spp. (BRANDAO 1987), but there is evidence that these species have retained a developmental pathway that results in the production of ergatoid queens (BRANDAO 1987, TsUJI 1988). Relatively little is known about the regulation of reproduction in queenless species.
The Platythyreini occur throughout the tropics. No queen caste has been reported from 25 of the 34 species of Platythyrea Roger 1863, including P. schultzei (BROWN 1975, KuGLER 1976). Because colonies of this species contain fewer than 50 members, they allow observation and dissection of whole colonies rather than samples of colony members. It was thus chosen for a study of the effects of queenlessness on division of labour and reproduction.
MATERIALS AND METHODS
Platythyrea schultzei Forel1910 is a medium-sized (8.5-9.5 mml, black ant with reddish legs. It is distributed through Zaire, Zambia, Zimbabwe, Namibia and South Africa (WHEELER eta!. 1922), and shows little morphological variation in its workers (ARNOLD 1915, BROWN 1975).
Seven colonies of were collected near drainage lines in mixed woodland near Hoedspruit (24° 28' S 30° 43' E), eastern Transvaal. Special care was taken to ensure that the entire colony and its brood were collected, and a flat-bottomed stone was left in the excavated hole for returning foragers to gather beneath; these were collected in the evening and during the morning of the next day. Colonies were housed in the laboratory in plastic freezer boxes 310 X 210 X 70 mm, covered with glass to minimize disturbances, and provided with a modified Lubbock nest covered with red Perspex. Water and live adult and larval dermestids and tenebrionids were provided. Laboratory temperatures were 20-23 °C and 50-80% RH.
Each day newly emerged ants were marked on the thorax with enamel modeling paint, using a different colour for each colony. An identifying number was written in waterproof ink onto this mark with a fine drafting pen. The ants were returned to their nest after their labels had dried, and were quickly accepted by their nestmates. A record was kept of the date on which each ant emerged. All other colony members were marked in the same way when the nests were first installed in the laboratory.
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013

Social differentiation in Platythyrea schultzei 15
Morphological differentiation
To quantify the size distribution of the species, the smallest distance between the eyes (interocular width) and the length of the antenna! scape, excluding the articulation, were measured for each ant. A least-squares linear regression model was fitted to the log-transformed data, which were also checked for normality.
Behavioural differentiation
A catalogue of behaviours was produced from 8 months of casual observation of four laboratory colonies, from the ethograms of two colonies (see below) and from observations of forced nest translocation in three laboratory colonies. The completeness of the behavioural catalogue was assessed from the ethogram (see below) using a log-Poisson distribution (FAGEN & GoLDMAN 1977).
Activity within two nests was recorded on video tape over 6 hr periods using a National NV8050 time lapse video recorder and a National WV 1500E/C television camera fitted with a close focus lens. A built-in clock, displaying date, hour, minute and second, was displayed on the screen and included in the recording. From these recordings the sequences, frequencies and durations of behaviours were quantified by focusing sampling on 34 ants (from a total of 41) for 30 min each (ALTMANN 1974).
Association between particular pairs of behaviours was investigated in two ways using the data derived from the video tape recordings. A behavioural transition probability matrix (HERBERS & CuNNINGHAM 1983) was used to assess the probability of one behaviour giving way to another. A hierarchical cluster analysis was performed on the matrix using an asymmetrical clustering algorithm (HUBERT 1973, CHILKO 1983) that is equivalent to single-linkage clustering when applied to a symmetrical matrix, to reveal how behaviour was organised.
This approach is limited to single-step transitions, and is thus capable of detecting the association of behaviours into tasks, but does not reveal roles, which are groups of functionally similar workers. In order to overcome these limitations it is necessary to measure the association of behaviours over longer time periods. To do this an association matrix was drawn up to show the number of ants which showed two given behaviours in their repertoires (HERBERS & CuNNINGHAM 1983). This approach offers some indication of the fidelity with which tasks are attended, i.e. the roles workers fill.
To investigate the basis of polyethism, each numbered colony member which eclosed in the laboratory was observed once a week for about 10 min, and the behaviours it performed during this period were recorded. Behaviours repeated during an observation period were noted only once for each ant. Ants lived as long as 21 weeks, but because sample sizes beyond 18 weeks were too small to be meaningful, only week 1 to week 18 of each ant's lifespan were used to assess age polyethism. The data were analysed using the approach of SEELY (1982): an ethogram was calculated for each week, and the values for each behaviour standardised across the weeks by dividing by the respective maximum value of each behaviour.
To assess physical polyethism, the distance between the eyes of each ant was measured as described under morphological differentiation. These data were combined with the behavioural data of the age polyethism study, and the head measurements of workers exhibiting a particular behaviour were compared between behaviours in two ways. First an analysis of variance was performed, using a least significant difference (LSD) criterion to test for differences in mean head measurement between pairs of behaviours (HERBERS & CuNNINGHAM 1983). Secondly, although the variation in head measurements in this species is small, size-related differences in behaviour may be expressed through the amount of its life which each worker spends in each role. The entry for each ant was therefore weighted by the number of weeks in which it displayed each given behaviour, and the analysis repeated.
Reproductive differentiation
After the behavioural observations were completed, workers' reproductive organs were dissected in insect Ringer's solution, stained with methylene blue and examined under a dissecting
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013

16 M.H. Villet
microscope for oocytes and corpora lutea in the ovaries. The number of ovarioles, the general development of the oocytes and the appearance of the spermatheca were noted, along with the age of the ant, wherever possible. The spermatheca was squashed in insect Ringer's solution between a microscope slide and coverslip and its contents examined under a Leitz Labrolux 11 phase contrast microscope for sperm.
RESULTS
Colonies were small, containing 9-38 members (Table 1). They were found up trees in dead, dry branches and in shallow, unstructured nests under stones, often occupying deserted termite galleries. Colonies appear to be monodomous, and attacked alien conspecifics when these were placed near their nests in the laboratory. Larvae and cocoons were sorted into piles, but kept in the same area of the nest, in both field and laboratory nests. Many parts of the nests contain the remains of cocoons and prey items, and no detritus was found around any of the nest entrances.
The remains of ants (Camponotus Mayr 1861, Crematogaster Lund 1831, Polyrhachis F. Smith 1858, Plectroctena Smith 1858, Bothroponera Mayr 1862 and Pheidole Westwood 1841 soldiers), termites (Macrotermes Holmgren 1910 soldiers), small beetles and bugs of several families, and grasshopper nymph were found in the nests. Workers foraged alone, both in the field and in the laboratory, searching up tree trunks and on the ground. The Crematogaster and Polyrhachis species recovered from P. schultzei middens were also arboreal foragers (H.G. RoBERTSON pers. comm.).
Males were collected in the field in November, and many eclosed from cocoons in the laboratory between November and February. Adult males lived as long as 4 weeks after eclosing, and tended to leave the nest after 2 weeks. They did not return to the nest after they had left. Males transferred from one laboratory colony to another entered the new nests and were generally quickly accepted by the workers.
Morphological difierentation
Female colony members were all similar in morphology. Only workers were found, and no queen or queen-like individual could be distinguished by the presence of either ocelli of flight sclerites, or the atrophied remnants of these structures. Neither was the gaster of any individual enlarged in any way, as is characteristic of ergatoid ponerine queens (VILLET 1989a).
Table 1. Composition of Platythyrea schultzei colonies used in this study.
Colony Date of Eggs Larvae Cocoons Males Workers collection
1 02.06.1986 9 1 3 0 22 2 03.09.1986 18 0 0 0 38 3 05.11.1986 17 22 10 0 9 4 05.11.1986 34 32 20 2 16 5 05.11.1986 30 34 16 4 30 6 19.11.1986 55 40 22 0 21 7 11.11.1987 29 16 17 0 12
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013
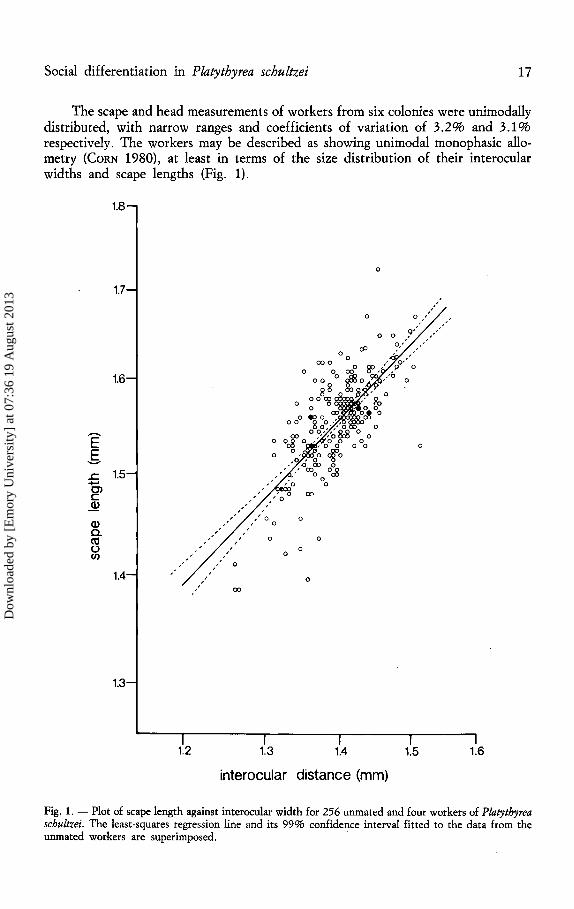
Social differentiation in Platythyrea schultzei 17
The scape and head measurements of workers from six colonies were unimodally distributed, with narrow ranges and coefficients of variation of 3.2% and 3.1% respectively. The workers may be described as showing unimodal monophasic allometry (CoRN 1980), at least in terms of the size distribution of their interocular widths and scape lengths (Fig. 1).
1.8
1.7
1.6
-E E -.c 1.5 +-' C> c Q)
Q) c. «) () fJ)
1.4 ,'
1.3
1.2
00
' 0 "" ,' 0
,' 0 0 0
0
0 0
0
1.3
0
0
1.4
interocular distance (mm)
1.5 1.6
Fig. 1. - Plot of scape length against interoculat width for 256 unmated and four workers of Platythyrea schultzei. The least-squares regression line and its 99% confidence interval fitted to the data from the unmated workers are superimposed.
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013

18 M.H. Villet
Reproductive differentiation
Every worker was found to possess a pair of ovaries composed without exception of four ovarioles each, and a spermatheca. However, only one individual in each colony was found to be in reproductive condition. The ovaries of this ant were well developed, being about 3 mm long, and contained corpora lutea and oocytes in varying stages of development, including two or three virtually mature oocytes 1.4 mm in length. The spermatheca contained live sperm. The head measurements of these mated workers were similar to those of their nestmates (t = - 0.271, P = 0. 787).
The ovaries of the remaining workers contained qualitatively similar densities of tracheoles to mated workers, but no oocytes or evidence of recent laying, since corpora lutea were absent. There was no trace of sperm in the spermathecae, and the ovarioles were about 2 mm long. The age of individuals did not affect any of these findings.
All of the eggs were removed from three nests. In the first six eggs were laid in 120 hr, in the second seven in 120 hr, and the last five in 168 hr. Since there was only one mated worker in each nest, these figures correspond to oviposition rates of 1.2, 1.4 and 0.7 eggs/reproductive ant/day respectively. Eggs were 1.41±0.06 mm long (1.24-1.57, n= 64).
Behavioural diHerentation
The ethograms provided sample coverages (FAGEN & GoLDMAN 1977) of more than 99.5% in all cases (Table 2), indicating that the behaviours seen were adequately sampled. The number of behaviours in each ethogram were within the 95% confidence limits of the estimated repertoire sizes (Table 2). The estimates agreed well with the observed catalogue sizes, except for that of virgin workers, which was understimated by at least three behaviours. Part of this discrepancy is due to the inclusion of specialised behaviours associated with nest moving and foraging. Mating and defensive behaviours are missing from both the ethogram and the catalogue.
The estimates of repertoire size imply that behaviour was adequately sampled in that the probability of seeing a new behaviour using the same observation techniques (P= 0.0001) was negligible. A total of 44 behaviours was recorded for workers, and 20 for the males (Table 2). Even if the various resting behaviours are grouped together, the total is still 42 for the workers. Ten of these behaviours were not observed during the construction of the ethogram, and therefore could not be quantified. No behaviours which could be interpreted as dominance or submissiveness were seen.
The behaviours of carrying (WC) and dragging (WD) adults have been distinguished because workers may carry nestmates in a stereotyped way, by grasping them by their mandibles and holding them over their backs, but when a foreign ant enters the nest, it is dragged out by either its legs or antennae. Nestmates are recruited by carrying only, and this occurs only during nest translocation. Necrophoria was not seen because ants did not die inside the nest, but left it when they became weak, and died outside.
Two unusual behaviours were present which have not been recorded in any ant species before, and therefore need explanation.
Milking larvae (LM): late instar larvae of P. schultzei possess a ventral abdominal
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013
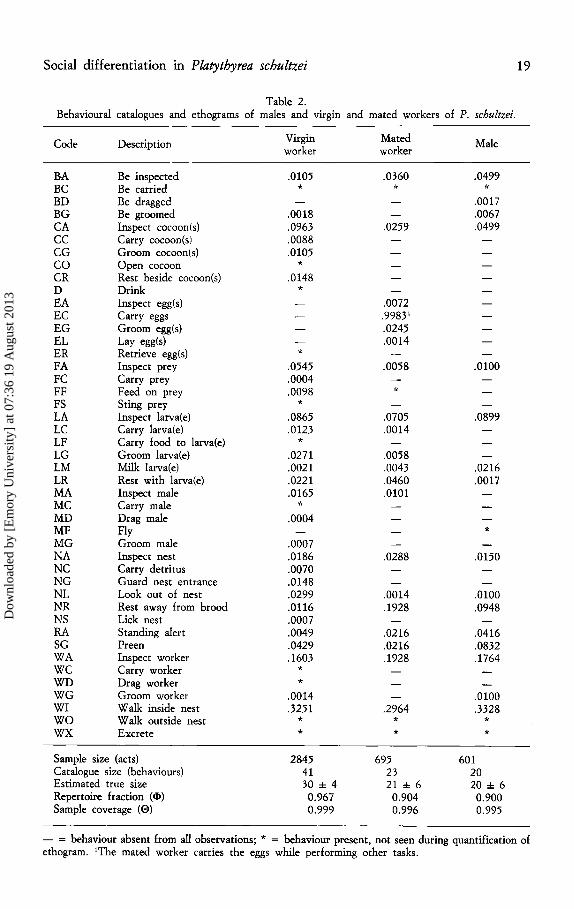
Social differentiation in Platythyrea schultzei 19
Table 2. Behavioural catalogues and ethograms of males and virgin and mated workers of P. schultzei.
Code Description Virgin Mated Male worker worker
BA Be inspected .0105 .0360 .0499 BC Be carried * * * BD Be dragged .0017 BG Be groomed .0018 .0067 CA Inspect cocoon(s) .0963 .0259 .0499 cc Carry cocoon(sJ .0088 CG Groom cocoon(s) .0105 co Open cocoon * CR Rest beside cocoon(s) .0148 D Drink * EA Inspect egg(s) .0072 EC Carry eggs .9983' EG Groom egg(s) .0245 EL Lay egg(s) .0014 ER Retrieve egg(s) * FA Inspect prey .0545 .0058 .olOO FC Carry prey .0004 FF Feed on prey .0098 * FS Sting prey * LA Inspect larva(e) .0865 .0705 .0899 LC Carry larva(e) .0123 .0014 LF Carry food to larva(e) * LG Groom larva(e) .0271 .0058 LM Milk larva(e) .0021 .0043 .0216 LR Rest with larva(e) .0221 .0460 .0017 MA Inspect male .0165 .o101 MC Carry male * MD Drag male .0004 MF Fly * MG Groom male .0007 NA Inspect nest .0186 .0288 .0150 NC Carry detritus .0070 NG Guard nest entrance .0148 NL Look out of nest .0299 .0014 .olOO NR Rest away from brood .0116 .1928 .0948 NS Lick nest .0007 RA Standing alert .0049 .0216 .0416 SG Preen .0429 .0216 .0832 WA Inspect worker .1603 .1928 .1764 we Carry worker * WD Drag worker * WG Groom worker .0014 .0100 WI Walk inside nest .3251 .2964 .3328 wo Walk outside nest * * * wx Excrete * * *
Sample size (acts) 2845 695 601 Catalogue size (behaviours) 41 23 20 Estimated true size 30 ± 4 21 ± 6 20 ± 6 Repertoire fraction ( cJ>) 0.967 0.904 0.900 Sample coverage (0) 0.999 0.996 0.995
- = behaviour absent from all observations; * = behaviour present, not seen during quantification of ethogram. 'The mated worker carries the eggs while performing other tasks.
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013

20 M.H. Villet
gland and tubercle that workers and males were often seen to lick (HANRAHAN et al. 1987, VILLET et al. 1990b). Since no other part of the larva was licked, and because grooming of the rest of the larva is restricted to the female caste, this behaviour was distinguished from normal grooming. Milking is not the same as larval haemolymph feeding (MASUKO 1986, 1989), since the adults do not feed on larval haemolymph (VILLET et al. 1990b).
Retrieving egg(s) (ER): as discussed below, one ant in each nest normally monopolised the task of carrying the eggs. If another ant found some eggs lying loose, it would pick them up and then appeared to search for the egg-bearer. When it found her, whether she was carrying other eggs at the time or not, it thrust the retrieved eggs at her mandibles until she took them. The eggs were always returned to the same ant in each nest. This behaviour was therefore distinguished from simply carrying eggs. Workers do carry eggs during nest translocation, after soliciting them from their normal bearer.
Dissection revealed that the ants that monopolised tending the eggs were the only ones in the nest that were inseminated. The age polyethism study showed that these ants filled this role in each nest for over 26 weeks (most workers lived less than 21 weeks in the laboratory). Their ethogram (Table 2) was therefore quantified separately. These ants carried the eggs even while performing other acts, putting them down only to feed and preen. It is then that other workers picked up the eggs, and exhibited the behaviour of retrieving eggs (ER). ER was also seen when the laboratory nests were disturbed and the egg-bearer dropped some eggs.
The egg-carrying worker were removed from three nests and dissected, confirming their status as mated workers. The eggs were left in the nest. Initially several different workers carried the eggs, but after 3 or 4 days one of the more recently eclosed workers (1-3 weeks old) would assume a monopoly over the task. In one case this lasted 15 weeks, until the study ended. Upon dissection several weeks after removing the mated workers, it was found that although the replacements were uninseminated, they had well developed ovaries containing mature oocytes and several corpora lutea. None of their contemporary nestmates showed any ovarial development, although workers only 1-2 days younger or older were also present in the nest. It is emphasized that only a single replacement developed in each colony.
Of the behaviours of the remaining workers, walking within the nest (WI) was the most frequent, followed by inspection of nestmates and brood. There were also common in males, which frequently rested too. Almost half of the acts were related to colony maintenance, a fifth to social interactions, and 28% to brood care (Table 2). The duration of any given behaviour was highly variable, and most coefficients of variation were greater than 80% (Table 3). About 40% of the colony time budget was spent in some form of resting behaviour. The tempo of activity in the nest was moderate, workers performing 256 acts/ant/hour, excluding resting time and acts. Of this active component, 23% went to brood care, 50% to colony maintenance, 20% to personal maintenance, and only 6% to social interactions.
Evidence of division of labour was obtained from the behavioural transition analysis. Apart from the tending of eggs, three further groups of behaviours were defined (Fig. 2): tending larvae, tending cocoons, and general nest-related tasks, including a subgroup relating to foraging and activity about the nest entrance. Preening was associated with all behaviours.
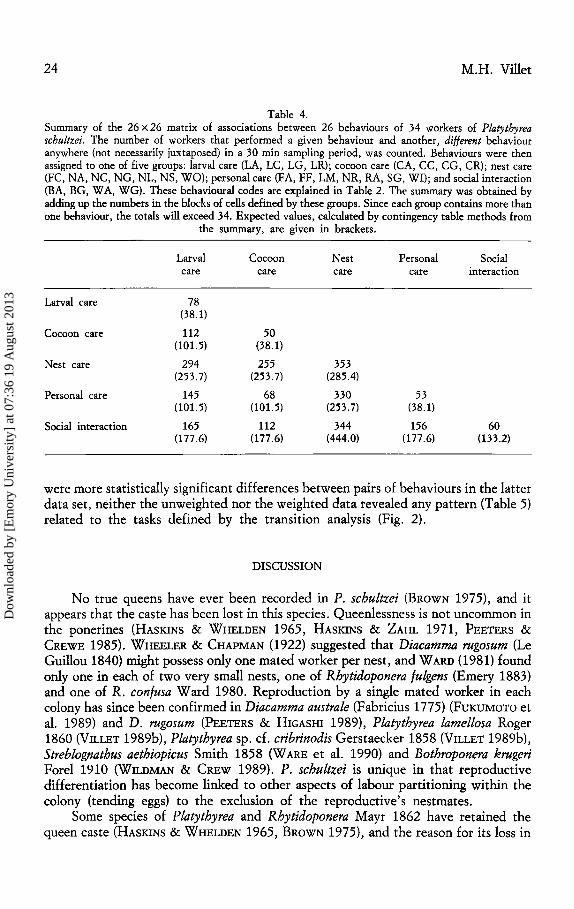
The association matrix (Table 4) indicated that workers tended to concentrate on
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013
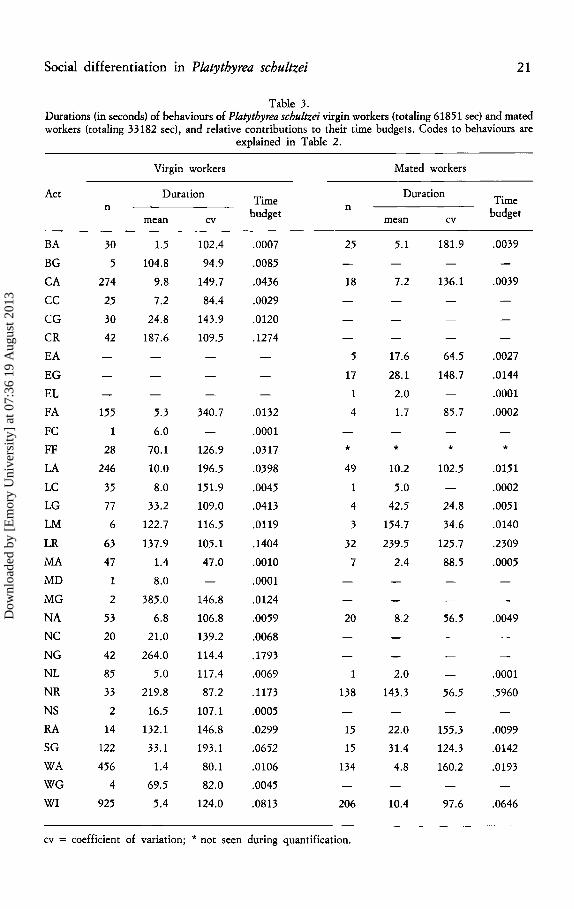
Social differentiation in Platythyrea schultzei 21
Table 3. Durations (in seconds) of behaviours of Platythyrea schultzei virgin workers (totaling 61851 sec) and mated workers (totaling 33182 sec), and relative contributions to their time budgets. Codes to behaviours are
explained in Table 2.
Virgin workers Mated workers
Act Duration Time
Duration Time n
budget n budget
mean cv mean cv
BA 30 1.5 102.4 .0007 25 5.1 181.9 .0039
BG 5 104.8 94.9 .0085
CA 274 9.8 149.7 .0436 18 7.2 136.1 .0039
cc 25 7.2 84.4 .0029
CG 30 24.8 143.9 .0120
CR 42 187.6 109.5 .1274
EA 5 17.6 64.5 .0027
EG 17 28.1 148.7 .0144
EL 1 2.0 .0001
FA 155 5.3 340.7 .0132 4 1.7 85.7 .0002
FC 1 6.0 .0001
FF 28 70.1 126.9 .0317 * * * * LA 246 10.0 196.5 .0398 49 10.2 102.5 .0151
LC 35 8.0 151.9 .0045 1 5.0 .0002
LG 77 33.2 109.0 .0413 4 42.5 24.8 .0051
LM 6 122.7 116.5 .0119 3 154.7 34.6 .0140
LR 63 137.9 105.1 .1404 32 239.5 125.7 .2309
MA 47 1.4 47.0 .0010 7 2.4 88.5 .0005
MD 8.0 .0001
MG 2 385.0 146.8 .0124
NA 53 6.8 106.8 .0059 20 8.2 56.5 .0049
NC 20 21.0 139.2 .0068
NG 42 264.0 114.4 .1793
NL 85 5.0 117.4 .0069 1 2.0 .0001
NR 33 219.8 87.2 .1173 138 143.3 56.5 .5960
NS 2 16.5 107.1 .0005
RA 14 132.1 146.8 .0299 15 22.0 155.3 .0099
SG 122 33.1 193.1 .0652 15 31.4 124.3 .0142
WA 456 1.4 80.1 .0106 134 4.8 160.2 .0193
WG 4 69.5 82.0 .0045
WI 925 5.4 124.0 .0813 206 10.4 97.6 .0646
cv = coefficient of variation; * not seen during quantification.
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013
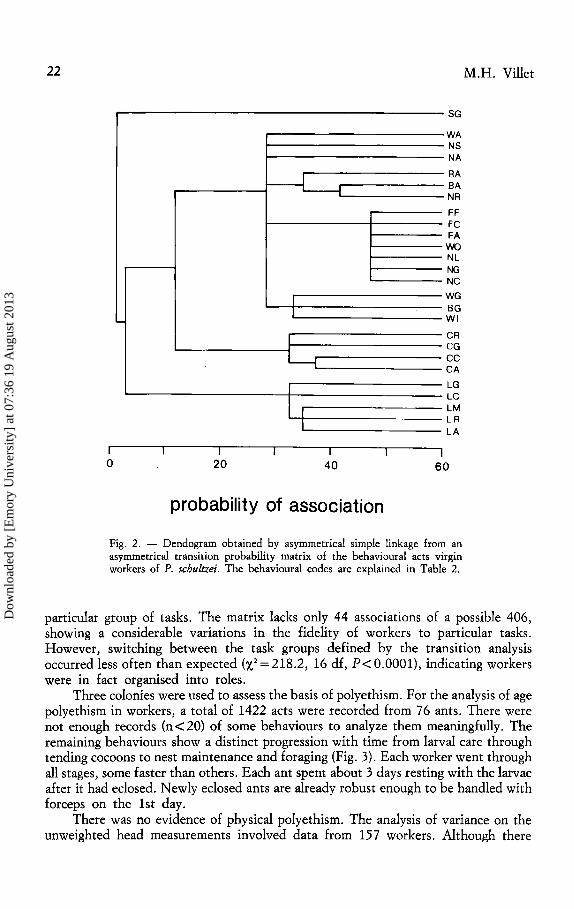
22
~
I I
I
I
li I
0 20 40
probability of association
SG
WA NS NA RA BA NR FF FC FA wo NL NG NC
WG BG WI
CR CG cc CA
LG LC LM LR LA
60
Fig. 2. - Dendogram obtained by asymmetrical simple linkage from an asymmetrical transition probability matrix of the behavioural acts virgin workers of P. schultzei. The behavioural codes are explained in Table 2.
M.H. Villet
particular group of tasks. The matrix lacks only 44 associations of a possible 406, showing a considerable variations in the fidelity of workers to particular tasks. However, switching between the task groups defined by the transition analysis occurred less often than expected (X2 =218.2, 16 df, P<O.OOOl), indicating workers were in fact organised into roles.
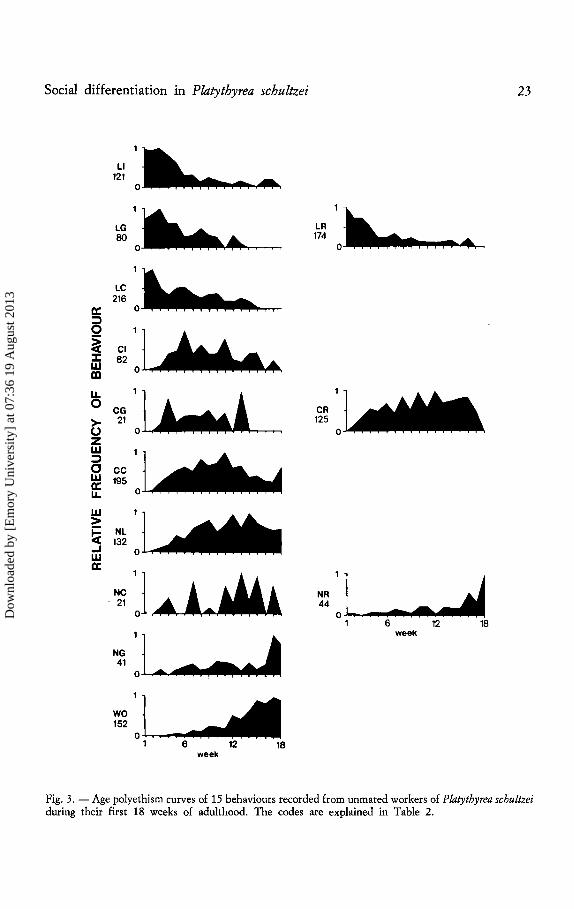
Three colonies were used to assess the basis of polyethism. For the analysis of age polyethism in workers, a total of 1422 acts were recorded from 76 ants. There were not enough records (n<20) of some behaviours to analyze them meaningfully. The remaining behaviours show a distinct progression with time from larval care through tending cocoons to nest maintenance and foraging (Fig. 3). Each worker went through all stages, some faster than others. Each ant spent about 3 days resting with the larvae after it had eclosed. Newly eclosed ants are already robust enough to be handled with forceps on the 1st day.
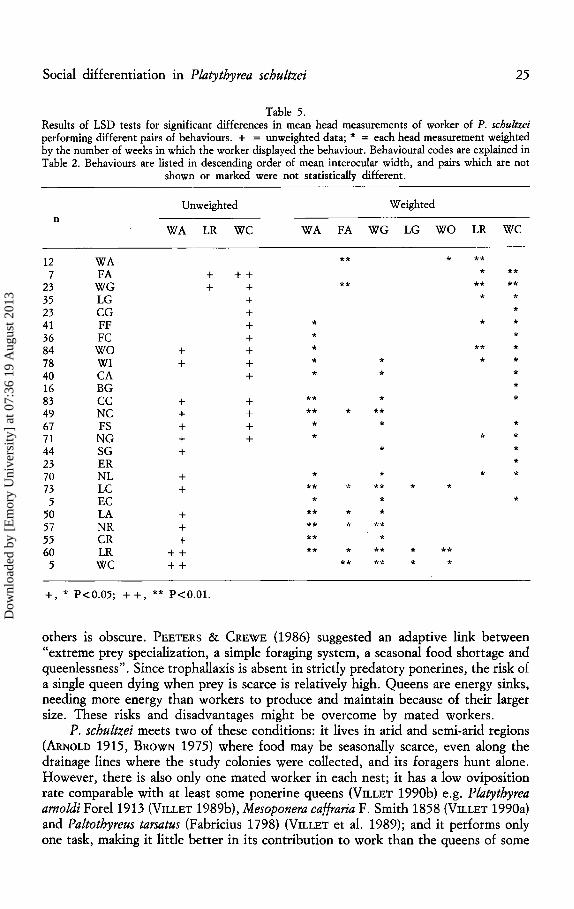
There was no evidence of physical polyethism. The analysis of variance on the unweighted head measurements involved data from 157 workers. Although there
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013
Social differentiation in Platythyrea schultzei
a: ~ 0
~ J: LU m LL 0
t> z LU ~ 0 LU a: LL
LU > fi ...1 LU a:
Ll 121
0
~~~ LC :~ 216
c•'~ 82
0
CG 21
cc 195
NL 132
NC 21
NG 41
wo 152
' l ~ 0 .A:.-r1 0
0
J~
week
23
LR ~ ...... , 174 "" .... '
CR 125
0
~<L ~--II ~ 1 6 12 18
week
Fig. 3.- Age polyethism curves of 15 behaviours recorded from unmated workers of Platythyrea schultzei during their first 18 weeks of adulthood. The codes are explained in Table 2.
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013
24 M.H. Villet
Table 4. Summary of the 26 X 26 matrix of associations between 26 behaviours of 34 workers of Platythyrea schultzei. The number of workers that performed a given behaviour and another, different behaviour anywhere (not necessarily juxtaposed) in a 30 min sampling period, was counted. Behaviours were then assigned to one of five groups: larval care (LA, LC, LG, LR); cocoon care (CA, CC, CG, CR); nest care (FC, NA, NC, NG, NL, NS, WO); personal care (FA, FF, LM, NR, RA, SG, WI); and social interaction (BA, BG, WA, WG). These behavioural codes are explained in Table 2. The summary was obtained by adding up the numbers in the blocks of cells defined by these groups. Since each group contains more than one behaviour, the totals will exceed 34. Expected values, calculated by contingency table methods from
Larval care
Cocoon care
Nest care
Personal care
Social interaction
the summary, are given in brackets.
Larval care
78 (38.1)
112 (101.5)
294 (253.7)
145 (101.5)
165 (177.6)
Cocoon care
50 (38.1)
255 (253.7)
68 (101.5)
112 (177.6)
Nest care
353 (285.4)
330 (253.7)
344 (444.0)
Personal care
53 (38.1)
156 (177.6)
Social interaction
60 (133.2)
were more statistically significant differences between pairs of behaviours in the latter data set, neither the unweighted nor the weighted data revealed any pattern (Table 5) related to the tasks defined by the transition analysis (Fig. 2).
DISCUSSION
No true queens have ever been recorded in P. schultzei (BROWN 1975), and it appears that the caste has been lost in this species. Queenlessness is not uncommon in the ponerines (HASKINS & WHELDEN 1965, HASKINS & ZAHL 1971, PEETERS & CREWE 1985). WHEELER & CHAPMAN (1922) suggested that Diacamma rugosum (Le Guillou 1840) might possess only one mated worker per nest, and WARD (1981) found only one in each of two very small nests, one of Rhytidoponera fulgens (Emery 1883) and one of R. confusa Ward 1980. Reproduction by a single mated worker in each colony has since been confirmed in Diacamma australe (Fabricius 1775) (FUKUMOTO et al. 1989) and D. rugosum (PEETERS & HIGASHI 1989), Platythyrea lamellosa Roger 1860 (VILLET 1989b), Platythyrea sp. cf. cribrinodis Gerstaecker 1858 (VILLET 1989b), Streblognathus aethiopicus Smith 1858 (WARE et al. 1990) and Bothroponera krugeri Forel 1910 (WILDMAN & CREW 1989). P. schultzei is unique in that reproductive differentiation has become linked to other aspects of labour partitioning within the colony (tending eggs) to the exclusion of the reproductive's nestmates.
Some species of Platythyrea and Rhytidoponera Mayr 1862 have retained the queen caste (HASKINS & WHELDEN 1965, BROWN 1975), and the reason for its loss in
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013
Social differentiation in Platythyrea schultzei 25
Table 5. Results of LSD tests for significant differences in mean head measurements of worker of P. schultzei performing different pairs of behaviours. + = unweighted data; * = each head measurement weighted by the number of weeks in which the worker displayed the behaviour. Behavioural codes are explained in Table 2. Behaviours are listed in descending order of mean interocular width, and pairs which are not
shown or marked were not statistically different.
Unweighted Weighted n
WA LR we WA FA WG LG wo LR we
12 WA ** * ** 7 FA + ++ * **
23 WG + + ** ** ** 35 LG + * * 23 eG + * 41 FF + * * * 36 Fe + * * 84 wo + + * ** * 78 WI + + * * * * 40 eA + * * * 16 BG * 83 ee + + ** * * 49 Ne + + ** * ** 67 FS + + * * * 71 NG + + * * * 44 SG + * * 23 ER * 70 NL + * * * * 73 Le + ** * ** * * 5 Ee * * *
50 LA + ** * * 57 NR + ** * ** 55 eR + ** * 60 LR ++ ** * ** * **
5 we ++ ** ** * *
+, * P<0.05; ++, ** P<O.Ol.
others is obscure. PEETERS & CREWE (1986) suggested an adaptive link between "extreme prey specialization, a simple foraging system, a seasonal food shortage and queenlessness". Since trophallaxis is absent in strictly predatory ponerines, the risk of a single queen dying when prey is scarce is relatively high. Queens are energy sinks, needing more energy than workers to produce and maintain because of their larger size. These risks and disadvantages might be overcome by mated workers.
P. schultzei meets two of these conditions: it lives in arid and semi-arid regions (ARNOLD 1915, BROWN 1975) where food may be seasonally scarce, even along the drainage lines where the study colonies were collected, and its foragers hunt alone. However, there is also only one mated worker in each nest; it has a low oviposition rate comparable with at least some ponerine queens (VILLET 1990b) e.g. Platythyrea arnoldi Forel1913 (VILLET 1989b), Mesoponera caffraria F. Smith 1858 (VILLET 1990a) and Paltothyreus tarsatus (Fabricius 1798) (VILLET et al. 1989); and it performs only one task, making it little better in its contribution to work than the queens of some
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013

26 M.H. Villet
species. In addition, the species is an unsp~cialised hunter and scavenger and thus less likely to be short of food if conditions deteriorate. Food reserves could also be obtained by milking the larvae.
If milking removes nutrients from the larvae, it might cause them to fall short of a caste-determining developmental threshold (WHEELER 1986), resulting in an apparent loss of the queen caste in this species. This mechanism needs investigation, since other queenless ants in the Ectatommini and Ponerini do not milk their larvae (VILLET 1989b). It also does not explain why only one mated worker was found in each nest.
Three mechanisms may regulate the numbers of reproductives in hymenopteran colonies (FLETCHER & Ross 1985). Unwanted reproductives may be killed or driven out either by the workers or by the dominant reproductive, or their reproductive behaviour may be regulated by pheromones emitted by the dominant reproductive.
When mated P. schultzei workers were removed from their colonies and their roles filled by virgin workers, no executions or aggressive interactions amongst the remaining workers were seen. Even the youngest worker from which the replacement reproductive was drawn did not show signs of interaction, and it is likely that the number of reproductives is not regulated by behavioural means. Trophallaxis, a potential source of subtle dominance interactions, is absent in this species.
The ability of workers which retrieve eggs to find the mated worker even if she is not carrying eggs implies that the mated worker emits a chemical cue by which she can be identified. This suggests that the egg-laying individual may also produce a pheromone that directly inhibits oogenesis in her nestmates. The production of sufficient pheromone to inhibit the entire colony may be facilitated by the small size of colonies. Bothroponera krugeri also has a single mated worker per colony, and a single virgin worker replaces a lost reproductive in this species too (VrLLET 1989b, WILDMAN & CREWE 1989). The regulatory mechanism is unknown. When the sole mated worker of a Streblognathus aethiopicus colony is removed, several virgin workers undergo ovarian development (WARE et al. 1990).
In the ponerine Ophthalmopone berthoudi Forel 1890, young workers form the pool from which mated workers are produced during the season of male activity (PEETERS & CREWE 1985). The functional replacement of experimentally removed mated workers of P. schultzei by young virgin workers resembles this pattern, but with the important difference that only one virgin worker in each nest assumed this role. This underlines the fact that, developmentally, the reproductives are workers and not morphologically cryptic queens. In queenright species many workers become reproductively active when the queen is removed (BouRKE 1988), i.e. inhibition is removed; in P. schultzei reproductive inhibition is maintained, and a virgin worker assumes the function of the removed reproductive. Similar results were obtained from Bothroponera krugeri (WILDMAN & CREWE 1989, VILLET 1989b).
PEETERS & CREWE (1985) also suggested that insemination regulated ovarian activity and reproductive differentiation in 0. berthoudi. In P. schultzei insemination is not a prerequisite for ovarial development or the establishment of reproductive dominance, although the three are linked. The following mechanism is suggested to regulate the number of reproductives. Soon after a mated worker dies, eclosing workers that have not been exposed to her influence compete passively for guardianship of the eggs. Whichever ant gains behavioural ascendancy undergoes ovarial development and gains the ability to suppress ovarial activity in its nestmates, probably by the same pheromonal mechanism. Behavioural ascendancy may arise from
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013

Social differentiation in Platythyrea schultzei 27
an initial advantage in reproductive physiology, such as greater ovarial development, or from an increased ability to synthesize regulatory pheromones. The onset of ovarial activity makes the behaviour of the dominant worker queen-like, and its continuation may be stimulated by possession of the packet of eggs. This worker also retains the ability to attract and mate with males when they enter the nest, and suppresses this behaviour in its fellows.
There was no evidence that obligate queenlessness caused important changes in the organization and division of labour. The behavioural catalogue of P. schultzei is no less complex than that of ants in more advanced subfamilies which exhibit 20-40 comparable behaviours (CoLE 1985), spread over two or three morphological castes in some cases (CoLE 1981). The basis of labour organisation within the repertoire of P. schultzei is clearly one of age rather than size differentiation, and even incipient behavioural differentiation amongst the workers on the basis of size is absent.
Although qualitative evidence of age polyethism in ponerine ants has been published (e.g. BONAVITA & POVEDA 1970, FRESNEAU et al. 1982, FRESNEAU 1984, FRESNEAU & DuPUY 1988), the subject has not been quantified in fine detail in a ponerine before. The age polyethism schedule of P. schultzei is a foraging-last one, as predicted when workers face the greatest risk of death during foraging (JEANNE 1986). The gradual transitions between roles may explain the amount of task switching revealed by the association matrix. A similar continuum has been reported in the ponerines Neoponera foetida (Olivier 1791) and Ectatomma ruidum Roger 1860 (FRES
NEAU et al. 1982). These data show clearly that there is sufficient behavioural and physiological
flexibility in the organisation of ponerine societies to allow the retention of eusocial division of labour and reproduction in the absence of a queen caste.
ACKNOWLEDGEMENTS
Thanks are extended to Sue Risi, Peter Hawkes and Mark Wildman for their help with field and laboratory work. The work is part of Ph.D. Thesis which was supervised by Prof. R. Crewe. Financial support was granted by the Freda Lawenski Trust, the University of the Witwatersrand and the Council for Scientific and Industrial Research.
REFERENCES
ALTMANN]. 1974. Observational study of behaviour: sampling methods. Behaviour 49: 227-267. ARNOLD G. 1915. A monograph of the Formicidae of South Africa. Annals of the South African
Museum 14: 1-766. BoNAVITA A. & PovEDA A. 1970. Mise en evidence d'une division du travail chez une fourmi
primitive. Comptes Rendus de l'Academie de Science, Paris 270: 515-518. BouRKE A. 1988. Worker reproduction in the higher eusocial Hymenoptera. Quarterly Review of
Biology 63: 291-311. BRANDAO C. 1987. Queenlessness in Megalomyrmex (Formicidae: Myrmicinae), with a discussion on
the effects of the loss of true queens in ants, pp. 111-112. In: Eder]. & Rembold H., Edits. Chemistry and biology of social insects. Miinchen: Verlag J. Peperny, XXXIII+ 757 pp.
BROWN W.L. 1975. Contributions towards a reclassification of the Formicidae. V. Ponerinae, tribes Platythyreini, Cerapachyini, Cylindromymecini, Acanthostichini, and Aenictogitini. Search, Cornell University 5: 1-115.
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013

28 M.H. Villet
CALABI P. 1988. Behavioural flexibility in Hymenoptera: a re-examination of the concept of caste, pp. 237-258. In: Trager E.]., Edit. Advances in myrmecology. Leiden: Brill Press, XXVII+551 pp.
CHILKO D.M. 1983. The IPFPHC procedure, pp. 131-144. In: Joyner S., Edit. SUGI Supplemental Library User's Guide 1983 Edition. Cary, North Carolina: SAS Institute Incorporated, XI+ 466 pp.
CoLE B. 1981. Dominance hierarchies in Leptothorax allardycei ants. Science 212: 83-84. COLE B .. 1985. Size and behavior in ants: constraints on complexity. Proceedings of the National
Academy of Science, USA 82: 8548-8551. CORN M.L. 1980. Polymorphism and polyethism in the neotropical ant Cephalotes atratus L. Insectes
Sociaux · 27: 29-42. FAGEN R.M. &'GoLDMAN R.N'. 1977. Behavioural catalogue analysis methods. Animal Behaviour 25:
261-274. FLETCHER D.J.C. & Ross K.G. 1985. R~gulation of reproduction in eusocial Hymenoptera. Annual
Review of Entom,ology 30: 319-343. • FRESNEAU D. 1984. Developpement ovarien et statut social chez une fourmi primitive Neoponera
obscuricornis Emery (Hym. Formicidae, Ponerinae). Insectes Sociaux 31: 387-402. FRESNEAU D. & DuPUY P. 1988. A study of polyethism in a ponerine ant: Neoponera apicalis
(Hymenoptera: Formicidae). Animal Behaviour 36: 1389-1399. FRESNEAU D., GARCIA PEREZ J. & ]AISSON P. 1982. Evolution of polyethism in ants: observational
results and theories, pp. 129-155. In: Jaisson P., Edit. Social insects in the tropics. Paris: University Paris Nord, 280 pp.
FuKUMOTO Y., ABET. & TAKI A. 1989. A novel form of colony organisat-ion in the "queenless" ant Diacamma rugosum. Physiological Ecology, Japan 26: 55-61.
HANRAHAN S.A., VILLET M.H. & WALTHER C. 1987. Preliminary studies of a trophallactic gland from larvae of a ponerine ant. Proceedings of the Electron Microscopy Society of Southern Africa 17: 49-50. .
HASKINS C.P. & WHELDEN R.M. 1965. 'Queenlessness', worker sibship and colony vs population structure in the formicid genus Rhytidoponera. Psyche 92: 217-236.
HASKINS C.P. & ZAHL P·.l).. 1971. The reproductive pattern of Dinoponera grandis Roger (Hymenoptera: Ponerinae) with notes on the ethology of the species. Psyche 78: 1-11.
HERBERS J.M. & CUNNINGHAM M. 1983. Social organisation in Leptothorax longispinosus Mayr. Animal Behaviour 31: 759-771.
HUBERT L. 1973. Min and Max hierarchical clustering using asymmetric similarity measures. Psychometrika 38: 63-72.
}EANNE R.L. 1986. The evolution of the organization of work in social insects. Monitore Zoologico Italiano (Nuova Serie) 20: 119-133.
KuGLER C. 1976. A new species of Platythyrea (Hymenoptera: Formicidae) from Costa Rica. Psyche 83: 216-221.
MASUKO K. 1986. Larval haemolymph feeding: a nondestructive parental cannibalism in the primitive ant Amblypone silvestrii Wheeler (Hymenoptera, Formicidae). Behavioral Ecology and Sociobiology 19: 249-255.
MASUKO K. 1989. Larval haemolymph feeding in the ant Leptanilla ;aponica by use of a specialised duct organ, the 'larval haemolymph tap' (Hymenoptera: Formicidae). Behavioral Ecology and Sociobiology 24: 127-132.
PEETERS C.P. & CREWE R.M. 1985. Worker reproduction in the ponerine ant Ophthalmopone berthoudi: an alternative form of eusocial organisation. Behavioral Ecology and Sociobiology 18: 29-37.
PEETERS C.P. & CREWE R.M. 1986. Queenright and queenless breeding systems within the genus Pachycondyla (Hymenoptera: Formicidae). Journal of the Entomological Society of Southern Africa 49: 251-255.
PEETERS C.P. & HIGASHI S. 1989. Reproductive dominance controlled by mutilation in the queenless ant Diacamma australe. Naturwissenschaften 76: 177-179.
SEELY T.D. 1982. Adaptative significance of the age polyethism schedule in honeybee colonies. Behavioral Ecology and Sociobiology 11: 287-293.
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013

Social differentiation in Platythyrea schultzei 29
TSUJI K. 1988. Obligate parthenogenesis and reproductive division of labour in the Japanese queenless ant Pristomyrmex pungens. Behavioral Ecology and Sociobiology 23: 247-255.
VILLET M.H. 1989a. A syndrome leading to ergatoid queens in ponerine ants (Hymenoptera: Formicidae). Journal of Natural History 23: 825-832.
VILLET M.H. 1989b. Division of labour in several African ponerine ants. Johannesburg: Ph.D. Thesis University of the Witwatersrand, XIV+ 202 pp.
VILLET M.H. 1990a. Colony foundation in a ponerine ant, Mesoponera caffraria F. Smith (Hymenoptera: Formicidae). South African Journal of Zoology 25: 39-40.
VILLET M.H. 1990b. Qualitative relations of egg size, egg production and colony size in some ponerine ants (Hymenoptera: Formicidae). Journal of Natural History 24: 1321-1331.
VILLET M.H., CREWE R.M. & RoBERTSON H. G. 1989. Mating behaviour and dispersal in Paltothyreus tarsatus Fabr. (Hymenoptera: Formicidae). Journal of Insect Behaviour 2: 413-417.
VILLET M.H., HANRAHAN S.A. & WALTHER C. 1990. Larval structures associated with larva-toadult trophallaxis in Platythyrea ants (Hymenoptera: Formicidae). International Journal of Insect Morphology and Embryology 19: 243-256.
WARD P.S. 1981. Ecology and life history of the Rhytidoponera impressa group (Hymenoptera: Formicidae) II colony origin, seasonal cycles and reproduction. Psyche 88: 109-126.
WARE A., COMPTON S. & RoBERTSON H. 1990. Gamergate reproduction in the ant Streblognathus aethiopicus Smith (Hymenoptera: Formicidae: Ponerinae). Insectes Sociaux 37: 189-199.
WHEELER D.E. 1986. Developmental and physiological determinants of caste in social Hymenoptera: evolutionary implications. American Naturalist 128: 13-34.
WHEELER W.M., BEQUART ]., BAILEY I.W., SANTSCHI F. & MANN W.M. 1922. Ants of the American Museum Congo expedition: a contribution to the myrmecology of Africa. Bulletin of the American Museum of Natural History 45: 1-1055.
WHEELER W.H. & CHAPMAN J.S. 1922. The mating of Diacamma. Psyche 29: 203-211. WILDMAN M.H. & CREWE R.M. 1989. Gamergate number and control over reproduction in
Pachycondyla krugeri (Hymenoptera: Formicidae). Insectes Sociaux 35: 217-225. WILSON E.O. 1984. The relation between caste ratios and division of labour in the ant genus
Pheidole (Hymenoptera: Formicidae). Behavioral Ecology and Sociobiology 16: 89-98.
Dow
nloa
ded
by [
Em
ory
Uni
vers
ity]
at 0
7:36
19
Aug
ust 2
013








![[XLS]biodiversityadvisor.sanbi.orgbiodiversityadvisor.sanbi.org/wp-content/uploads/2014/07/... · Web viewEntella orba Stål, 1856|Hapalomantis meridionalis|Mantis orba schultzei](https://img.pdfslide.us/doc/110x75/5ae8e6fa7f8b9ae1579114a8/xls-viewentella-orba-stl-1856hapalomantis-meridionalismantis-orba-schultzei.jpg)















![Chinese language forel[2]](https://img.pdfslide.us/doc/110x75/5496a262b4795991148b4674/chinese-language-forel2.jpg)




