Embed Size (px)
Citation preview

Journal of Cellular Biochemistry 84:100±107 (2002)
Small GTP-Binding Proteins in the Nuclei of HumanPlacenta
Phoebe Phillips,1 and Murray Thomson,2*1University of Western Sydney Nepean, PO Box 10, Kingswood, NSW, 2747, Australia2School of Biological Sciences, University of Sydney, NSW, 2006, Australia
Abstract Mitochondria and crude nuclei containing fractions from human placenta have been shown to containproteins which bind [a32P]-GTP. Prior to this study the number of GTP-binding proteins in placental nuclei and theirnucleotide speci®city was not known. Also unknown was the identity of any of the GTP-binding proteins inmitochondria of human placenta. Nuclei and mitochondria were puri®ed from human placental extracts bysedimentation. Proteins were separated by electrophoresis and transferred to nitrocellulose membranes. Overlayblot with [a32P]-GTP identi®ed two nuclei proteins with approximate molecular weights of 24 and 27 kDa. Binding of[a32P]-GTP to the 27 and 24 kDa proteins was signi®cantly displaced by guanine nucleotides but not by adenine,thymine or cytosine nucleotides or deoxy (d) GTP. Western blot with a speci®c antibody to Ran identi®ed a band at27 kDa in nuclei and in mitochondrial fractions. These data indicate that both nuclei and mitochondria contain 24 and27 kDa GTP-binding proteins. The GTP-binding proteins in nuclei display binding speci®city for guanine nucleotidesand the hydroxylated carbon 2 on the ribose ring of GTP appears essential for binding. It will be important in futurestudies to determine the functions of these small GTP-binding proteins in the development and physiology of theplacenta. J. Cell. Biochem. 84: 100±107, 2002. ß 2001 Wiley-Liss, Inc.
Key words: nuclei; GTP-binding proteins; placenta; Ran; mitochondria
GTP-binding proteins are involved in theregulation of a variety of physiological processesin many cell and tissue types. GTP-bindingproteins are activated when bound to GTP andinactivated when inherent GTPase activityconverts the bound GTP to GDP. GTP-bindingproteins are often divided into groups on thebasis of their size and quaternary structure. Theheterotrimeric GTP-binding protein group areoften referred to as G proteins and are composedof a GTP-binding a subunit, as well as, b andg subunits [Bourne, 1994]. The molecularweight of a heterotrimeric G protein a subunitusually resides within a range of 35±50 kDa[Bourne, 1994, 1997]. The heterotrimeric Gproteins are often involved in the transmission
of a cellular signal through the plasma mem-brane [Bourne, 1994]. Additionally, there is thefamily of monomeric GTP-binding proteinswhich often have a molecular weight of 14±33 kDa and due to this size have been namedsmall GTP-binding proteins [Takai et al., 2001].Small GTP-binding proteins have been foundto be involved in processes including geneexpression, cytoskeletal organization [Takaiet al., 2001] and membrane fusion [Thomson,1998].
Recently, overlay blot with [a32P]-GTP hasidenti®ed several GTP-binding proteins in thehuman placenta and it has been proposed thatthese may be important regulators of placentalphysiology [Thomson et al., 1998]. The placentais an ephemeral organ that supplies the fetuswith nutrients and the mother and the fetuswith hormones [Petraglia et al., 1996]. Becausethe healthy development and function of theplacenta is crucial to the health of both fetus andmother it would appear important to character-ise the GTP-binding proteins present in thisorgan. GTP-binding proteins of molecularweights 24, 27, 33, 37 and 52 kDa have been
ß 2001 Wiley-Liss, Inc.DOI 10.1002/jcb.1269
Grant sponsor: University of Western Sydney NepeanResearch Grant.
Phoebe Phillips's present address is Faculty of Medicine,University of New South Wales, NSW, 2052, Australia.
*Correspondence to: Murray Thomson, School of BiologicalSciences (F07), The University of Sydney, NSW, 2006,Australia. E-mail: [email protected]
Received 30 May 2001; Accepted 3 August 2001

shown to be localised in the mitochondria ofhuman placenta [Thomson et al., 1998].Whether these proteins were similar or iden-tical to known GTP-binding proteins found inother parts of the cell was not known at this time[Thomson et al., 1998]. GTP-binding proteins of24 and 27 kDa were found to be present in crudecell debris fractions containing nuclei, but it wasnot determined whether these were present inpuri®ed preparations of nuclei [Thomson et al.,1998].
In many species including Xenopus [Gorlichet al., 1994], mouse [Coutavas et al., 1994] andrat [Clarkson et al., 1996] the participation ofthe GTP-binding protein Ran is needed forprotein transport into the nucleus. While Ranhas been identi®ed in the nuclei of many tissuesand cells, there have only been a few studies todetermine whether there are small GTP-bind-ing proteins other than Ran in the nucleus andthese have focused on the rat. Overlay blot with[a32P]-GTP has shown the existence of 25 and26 kDa GTP-binding proteins in rat neuronalnuclei [Bhullar et al., 1999], while in rat livernuclei, 23, 24.5, 25 and 26 kDa proteins havebeen located [Rubins et al., 1990]. Prior to thisstudy Ran or other small GTP-binding proteinshad not been identi®ed in the nuclei of humanplacental tissue.
In the present study we determined whetherthere were [a32P]-GTP binding proteins innuclei from human placenta and whether thesewere speci®c GTP-binding proteins. Addition-ally, we used speci®c antibodies to determinewhether three known GTP-binding proteins(Ran, Rac and Rho) with molecular weights inthe 20±28 kDa range were located in mitochon-dria or nuclei from human placenta.
METHODS
Isolation of Mitochondria From Placenta
Human term placentae was obtained fromWestmead Hospital, Australia and placed inchilled phosphate buffered saline (PBS). Thematernal umbilical vein and fetal arterieswere perfused with PBS to remove blood. Thepreparation of mitochondria was carried outusing a modi®cation of the method by Adamset al. [1989]. Pieces of placental tissue approxi-mately 1 cm3 were homogenised in buffer A(Tris±HCl 20 mM, mannitol 210 mM, sucrose70 mM) using a motor driven Te¯on pestle.Next, the homogenate was centrifuged at 700g
for 10 min at 48C in a Sorvall Ultra Pro 80ultracentrifuge (Fixed angle rotor and 35 mltubes). The resulting supernatant was kept at48C and the pellet was re-extracted twice inbuffer A. The supernatants were combined andcentrifuged at 9,750g for 15 min at 48C. Theresulting mitochondrial pellet was collected andresuspended in 200 ml of buffer A. This suspen-sion was centrifuged three times at 9,750g at48C. Following this the supernatant wasremoved and the mitochondrial pellets resus-pended in buffer A.
Isolation of Nuclei From Placenta
The isolation of human placental nuclei wascarried out as described by Boivin and Beliveau[1995]. For the preparation of nuclei, 2 g ofhomogenised placental tissue prepared as abovewas re-homogenised in 20 ml of 2.4 M sucrose,3.3 mM CaCl2 and 0.04% trasylol. The homo-genate was then ®ltered through three layers ofcheesecloth and centrifuged at 40,000g for60 min. The supernatant was removed and thepellet was resuspended in 2 ml of 1 M sucrose,1 mM CaCl2 and 0.04% trasylol. The suspensionwas centrifuged three times at 3,000g for 5 min.The ®nal nuclear pellet was resuspended in250 mM sucrose, 1 mM CaCl2 and 0.04%trasylol, and stored at ÿ708C.
Assay of Protein Concentration
The concentration of each mitochondrialsample was determined via the Pierce BCAProtein Assay as per the manufacturer's in-structions. The samples were read at 562 nm onthe Biorad microplate reader (Biorad Model550 microplate reader).
Sodium Dodecyl Sulfate±PolyacrylamideGel Electrophoresis
Sodium Dodecyl Sulfate±Polyacrylamide GelElectrophoresis (SDS±PAGE) was performedusing the Biorad Mini Protean II system. Theseparation of mitochondrial and nuclear pro-teins involved resuspending the samples in SDSsample buffer (50 mM Tris±HCl pH 6.8, 2% SDSw/v, 10% glycerol and 5% b-mercaptoethanol).The samples were then heated at 958C for3±5 min and then centrifuged for 5 min at3,000 rpm. Samples were loaded onto 12% SDSgels that contained a 4% stacking gel and run at100 V for approximately 1.5 h. Protein bandsobtained for mitochondrial and nuclei sampleswere detected using coomassie stain and silver
Small GTP-Binding Proteins in the Nuclei 101

stain (Biorad Silver Stain Plus Kit) as per themanufacturer's instructions.
Overlay Blot Detectionof GTP-Binding Proteins
GTP overlay assays were performed onisolated nuclei. The samples were ®rst run on12 % SDS gels and then transferred electro-phoretically to nitrocellulose. The lanes on thenitrocellulose were marked using ponceaustain. The stain was removed with TBS(20 mM Tris and 0.9% NaCl, pH 7.5). Thenitrocellulose was then blocked with 5% skimmilk in TBS for 20 min. The method describedby Zeuzem et al. [1992] was followed to performthe assay in order to characterise GTP bindingin proteins from the nuclei. The nitrocellulosemembrane was washed twice in incubatingbuffer (50 mM Tris±HCl pH 7.5, 1 mM EDTA,5 mM MgCl2, 0.3% Tween 20, 1 mM K2HPO4/KH2PO4) for 10 min. The membrane wasleft to incubate for 1 h in 1 ml of [a32P]-GTP(3� 10ÿ6mol/L) per 10 ml of buffer. The mem-brane was then washed ®ve times for 10 min inincubating buffer. The membrane was allowedto dry and then wrapped in plastic food wrapand exposed to X-ray ®lm at ÿ708C.
To investigate the nuclear fractions forspeci®city for guanine nucleotides, membranescontainingSDS±PAGEseparatedproteinswerepre-incubated for 20 min with a 10� concen-tration of the unlabelled nucleotides AMP, ADP,ATP, GMP, GDP, GTP, CTP, ITP, TTP, anddGTP. On completion, [a32P]-GTP was added tothe membranes (in the presence of unlabellednucleotides) for 1 h. The membranes were thenwashed as outlined above in incubating bufferand exposed to X-ray ®lm at ÿ708C.
Immunodetection of Ran in the Nucleus
The immunoreactivity of Ran was detected byWestern blot using the puri®ed goat polyclonalantibody Ran (C-20) obtained from Santa CruzBiotechnology (USA). Nuclei containing 50 mg ofprotein were separated by 12% SDS±PAGE andtransferred by electrophoresis to nitrocelluloseusing a Biorad Mini Trans-Blot system. Themembranes were blocked in 5% skim milk inTBS with 0.05% Tween-20 (TTBS) for 1 h. Themembranes were incubated for 1 h at roomtemperature with polyclonal anti-Ran/TC4antiserum (1 mg/ml) in 5% skim milk in TTBS.The membranes were then washed three timesfor 5 min each with TTBS and incubated with
the secondary antibody (gamma anti-goatcoupled to horseradish peroxidase obtainedfrom Santa Cruz Biotechnology; 1:2,000) for45 min. The membrane was then rinsed withTTBS (3� 5 min) and ®nally with TBS. Themembrane was then incubated in chemilumi-nescence luminol reagent (ECL Du Pont) andexposed to X-ray ®lm.
In addition, a Western blot was performedwith polyclonal antibodies for Rac and Rho(obtained from Santa Cruz Biotechnology) usingthe same procedure as above.
Analysis of SDS±PAGE Results
The molecular weight of proteins and bandintensity was estimated with the aid of thescienti®c program NIH IMAGE.
Statistical Analysis
One way analysis of variance (ANOVA) wasused to analyse the data. P< 0.05 was consid-ered to be signi®cant.
RESULTS
SDS±PAGE of Nuclear Extracts
Proteins expressed in the nuclei and mito-chondria from human placentae were analysedusing one-dimensional SDS±PAGE and detect-ed using coomassie blue (Fig. 1). The samplesfrom nuclei and mitochondria showed bandingpatterns, which spanned the 200±20 kDamolecular weight range. Staining the gels withsilver did not result in additional bands becom-ing apparent (results not shown).
GTP Overlay of Nuclei Fractions
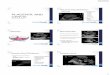
GTP overlay assay was used to determinewhether GTP-binding proteins were detectablein nuclei isolated from human placenta. Variousprotein amounts were analysed by this methodand a dose dependant identi®cation of 24 and27 kDa GTP-binding proteins was observed(Fig. 2). A 3- and 18-h exposure of the membranewith labelled proteins is shown in Figure 2a,b,respectively. The 24 and 27 kDa proteins from a50 mg loading and a 2- and 6-h autoradiographexposure is shown in Figure 2c,d.
Speci®city of GTP-Binding Proteins
Overlay blot analysis was performed in thepresence and absence of unlabeled nucleotidesto determine the speci®city of the af®nity of the24 and 27 kDa proteins for GTP. Nitrocellulose
102 Phillips and Thomson

bound proteins were incubated with the radio-label and 10� concentration of the unlabelednucleotides AMP, ADP, ATP, GMP, GDP, GTP,CTP, ITP, and TTP. Results in Figure 4 showthat AMP, ADP, ATP, CTP, ITP, and TTP didnot signi®cantly inhibit the binding of [a32P]-GTP. GMP, however, signi®cantly (P< 0.05)displaced the [a32P]-GTP from the 24 and 27kDa proteins (Fig. 3). Furthermore, both GDPand GTP completely abolished the binding oflabelled GTP (P< 0.001) (Fig. 3).
Participation of Ribose Hydroxyl Groupin GTP Binding
To determine whether the oxygen atom in thehydroxyl group attached to carbon 2 in GTP isnecessary for binding of [a32P]-GTP to 24 and27 kDa, nuclei proteins immobilised on nitro-
cellulose were preincubated with radio-labeled[a32P]-GTP with and without a 10� concen-tration of deoxy (d) GTP. There was no signi-®cant (P� 0.06) displacement of [a32P]-GTP inthe presence of dGTP (Fig. 3).
Rac, Rho and Ran Immunoreactivity
In order to obtain the identity of the smallGTP-binding proteins expressed within placen-tal nuclei, Western blots were performed usingpolyclonal antibodies for two GTP-bindingproteins Ran and Rac which have molecularweights comparable to the two GTP-bindingproteins in the nuclei of the placenta. Resultsrevealed the presence of Ran immunoreactivityin nuclei obtained from the placenta (Fig. 4).GTP-overlay performed on a membrane thathad been analysed for Ran antibody bindingshowed that Ran immunoreactivity was presentat precisely the same position as the 27 kDa[a32P]-GTP binding protein (Fig. 4). In contrast,Rac and Rho were found to be absent from thenuclei in all three placentas tested (results notshown).
The presence of ran was also investigated andfound to be present in percoll puri®ed mitochon-dria of human placentae (Fig. 5). The concen-tration of Ran did not signi®cantly (P� 0.12)differ between the mitochondria and thenucleus.
DISCUSSION
In a previous study primarily on mitochon-dria, crude fractions from human placentawhich contained nuclei were shown to containtwo small molecular weight GTP-binding pro-teins [Thomson et al., 1998]. In the presentstudy nuclei were separated from crude frac-tions and the GTP-binding proteins in the nucleifractions were analysed for their protein con-tent. As shown in Figure 1, nuclei and mito-chondria fractions stained nonspeci®cally forprotein showed a wide range of protein bandsover the 200±20 kDa range. For both nuclei andmitochondria at loadings of 50 mg, all proteinbands in the gel were sharply de®ned and nobands exhibited vertical streaking, a symptomof overloaded gels. This indicated that 50 mg ofprotein was an appropriate amount to be loadedonto the gel.
Overlay blot with [a32P]-GTP showed majorbands of 24 and 27 kDa (Fig. 2) and bindingdisplayed a dose responsive manner (Fig. 2).
Fig. 1. SDS±PAGE of proteins from human placenta. Fiftymicrograms of protein were added to each lane. Lane 1 showsnuclear proteins, lane 2 shows mitochondrial proteins. The gelwas stained using coomassie blue.
Small GTP-Binding Proteins in the Nuclei 103

This pattern is similar to the results reported formitochondria [Thomson et al., 1998] and a 50 mgloading was in the linear range of response.Higher doses with an autoradiograph exposuretime of 18 h showed some GTP-binding activityin the 30±70 kDa range although signal to noiseratio in this region was low and discrete bandswere not apparent. Figure 1 demonstrates thatthere are a number of discrete protein bands inthe 30±200 kDa range, however, the [a32P]-GTP binding properties of the proteins in thismolecular weight range are absent or negligible.It would appear therefore that the 24 and 27kDa GTP-binding proteins in the nuclei are inthe small GTP-binding protein class and appearlikely to be monomeric [Bourne, 1997; Thomson,1998].
Because the 24 and 27 kDa proteins are closein molecular weight and the overlay blot
technique creates a `halo' effect around thebands, discrimination of the 24 and 27 kDabands from each other is sometimes problematic[Thomson et al., 1998]. The exposure time of theoverlay blot in autoradiography can also effectresolution between the 24 and 27 kDa bands asshown in Figure 2.
GTP-binding proteins have af®nity for bothGDP and GTP and much lower af®nity fornonguanosine nucleotides [Denslow et al.,1991]. The speci®city of GTP-binding by the 24and 27 kDa proteins in placental nuclei wasexamined by pre-incubating blots with 10� theconcentration of unlabelled nucleotide (Fig. 3).The pyrimidine nucleotides tested (CTP andTTP) did not signi®cantly effect the binding of[a32P]-GTP to 24 and 27 kDa proteins. Itappears therefore that a purine is essential fora nucleotide to interact with the binding site of
Fig. 2. Binding of [a32P]-GTP to placental nuclei proteins. In (a) and (b) Lanes 1, 2, 3, 4, 5 and 6 contain 5,10, 20, 50, 75 and 100 mg of protein respectively. Figures a and b show exposure times of 6 and 18 hrespectively. The effect of time of exposure on detection of the 24 and 27 kDa [a32P]-GTP binding proteinsis shown in (c) and (d) which show 2 and 6 h autoradiograph exposure times, respectively. The experimentwas repeated a total of three times using different placentae with similar results.
104 Phillips and Thomson

24 and 27 kDa GTP-binding proteins. Theadenosine nucleotides also did not show anysigni®cant displacement of [a32P]-GTP. Thisindicated that a) the amino group in the purinemust be at carbon 2 rather than carbon 6 and/orb) an oxygen atom double bonded to carbon 6 isrequired for binding to 24 and 27 kDa proteins.In addition, ITP had no signi®cant effect on thebinding of [a32P]-GTP. ITP resembles GTPexcept for the absence of the amino group atposition 2, thus the amino group at carbon 2must by itself be important in GTP binding. Incontrast, both GDP and GTP completely abol-
ished the binding of [a32P]-GTP. GMP lacks theb-phosphate group, and the low af®nity for GMPdisplayed by the 24 and 27 kDa proteinsindicates that the b-phosphate contributes tothe binding interaction [Denslow et al., 1991].
In order to investigate the importance ofthe ribose ring in binding of [a32P]-GTP toGTP-binding proteins, deoxy-GTP was used as acompetitor (Fig. 3). The results showed thatdGTP had no signi®cant effect on the binding of[a32P]-GTP to 24 and 27 kDa nuclear proteins.This indicates that the 20-hydroxyl group pre-sent on GTP is needed for binding.
Fig. 3. Speci®city of [a32P]-GTP binding to the 24 and 27 kDa proteins. Equal amounts (50 mg protein) ofnuclear extract from three different placentae were subjected to overlay blot in the presence of a 10-foldexcess of unlabelled nucleotide (competitor) or saline vehicle only (control). The values are themeans� SD. (a) Shows the autoradiograph of the proteins while (b) shows densitometry of the bands.
Small GTP-Binding Proteins in the Nuclei 105

The GTP-binding protein Ran was ®rstidenti®ed in isolated rat liver nuclear envelopesby photo-af®nity labelling with [a32P]-GTP[Seydel and Gerace, 1991]. In a range of tissuesnot including placenta, Ran has been found to bean essential factor in import of proteins contain-ing a nuclear localization sequence into thenucleus [Melchioir et al., 1993; Moore andBlobel, 1994]. The current nuclear import model[Cole and Hammell, 1998] suggests that importcomplexes are formed when Ran is primarilybound to GDP. This complex is then transportedthrough the nuclear pore complex (NPC).Because Ran has been shown to be so importantfor nuclei function in other tissues and specieswe determined whether Ran was present in thenuclei of human placentae as discussed below.
Using antisera to Ran, a major band wasidenti®ed at 27 kDa (Fig. 4). It appears possiblethat the 27 kDa [a32P]-GTP binding protein isRan and future studies will be needed toinvestigate this possibility. It will also beinteresting to determine in future studieswhether Ran is functioning to import biomole-cules through the nuclear membrane in humanplacenta. There have been very few studies thathave determined whether there are GTP-bind-ing proteins other than Ran in the nucleus, inone other study using overlay blot Bhullar et al.[1999] have identi®ed 25 and 26 kDa proteinsthat bind [a32P]-GTP. Bhullar et al. [1999] alsoused Ran antisera to detect a major protein ofapproximately 26 kDa as well as a minor band of32 kDa. Photoaf®nity labelling has been used todetect an 80 kDa GTP-binding protein in the
nuclei of rabbit liver nuclei [Singh et al., 1995].In the present study we show that in addition tothe 27 kDa protein, nuclei from human placentacontain a 24 kDa GTP-binding protein that doesnot cross-react with a antibody to Ran (Fig. 4). Itwill be important in future studies to determinethe biological properties of the 24 kDa protein,and whether it shows structural similarities toknown small GTP-binding proteins.
Interestingly, Ran was also identi®ed inmitochondria from the human placenta(Fig. 5). Mitochondria have been shown tocontain a GTP-binding protein of approximately27 kDa [Thomson et al., 1998], however, prior tothe present report the identity of this proteinwas not known. The mitochondrion shares somecommon features with the nucleus, both havetwo membranes and import biological mole-cules. Ran has been shown to function in importfunctions in the nucleus and it has beenproposed that in the mitochondria GTP-binding
Fig. 4. Western blot with Ran antibody shown in lane 1.Overlay blot with [a32P]-GTP performed on the same piece ofnitrocellulose shown in lane 2. The experiment was repeated atotal of three times using three separate placentae with similarresults.
Fig. 5. Ran immunoreactivity in proteins from (1) mitochon-dria and (2) nuclei. The experiment was repeated a total of threetimes using three separate placentae with similar results.
106 Phillips and Thomson

proteins modulate the import of proteins andcholesterol through outer and inner membranecontact points. This poses the question, doesmitochondrial Ran regulate the mitochondrialimport of biological molecules?
The presence of GTP-binding proteins in thenucleus of human placental cells raises numer-ous intriguing possibilities. The 24 and 27 kDaproteins may function to regulate proteinsynthesis in the organ and may control thegrowth and development and function of theplacenta. As such these proteins may be a vitalpart of the myriad of placental functions whichsupports the growth and development of thefetus.
ACKNOWLEDGMENTS
The authors thank The Delivery Suite, West-mead Hospital Australia, for their assistance instudies on the human placenta.
REFERENCES
Adams V, Bosch W, Schlegel J, Wallimann T, Brdiczka D.1989. Further characterization of contact sites frommitochondria of different tissues: topology of peripheralkinases. Biochim Biophys Acta 981:213±225.
Bhullar RP, McCartney DG, Kanfer JN. 1999. Hetero-trimeric and small molecular mass GTP-binding proteinsof rat brain neuronal and glial nuclei. J Neurosci Res55:80±86.
Boivin D, Beliveau R. 1995. Subcellular distribution andmembrane association of Rho-related small GTP-bindingproteins in kidney cortex. Am J Physiol 269:F180±F189.
Bourne HR. 1994. G proteins. The importance of beingGTP. Nature 369:611±612.
Bourne HR. 1997. How receptors talk to trimeric Gproteins. Curr Opin Cell Biol 9:134±142.
Clarkson WD, Kent HM, Stewart M. 1996. Separatebinding sites on nuclear transport factor 2 (NTF2) forGDP-Ran and the phenylalanine-rich repeat regions ofnucleoporins P62 and Nsp1p. J Mol Biol 263:517±524.
Cole CN, Hammell CM. 1998. Nucleocytoplasmic transport:driving and directing transport. Current Biol 8:R368±R372.
Coutavas EE, Hsieh CM, Ren M, Drivas GT, Rush MG.D'Eustachio D. 1994. Tissue-speci®c expression of Ranisoforms in the mouse. Mammalian Genome 5:623±628.
Denslow ND, Anders JC, O'Brien TW. 1991. Bovinemitochondrial ribosomes possess a high af®nity bindingsite for guanine nucleotides. J Biol Chem 26:9586±9590.
Gorlich D, Prehn S, Laskey RA, Hartmann E. 1994.Isolation of a protein that is essential for the ®rst stepof nuclear protein import. Cell 79:767±778.
Melchioir F, Paschal B, Evans E, Gerace L. 1993. Inhibitionof nuclear protein import by non-hydrolyzable analogs ofGTP and identi®cation of the small GTPase Ran/TC4 asan essential transport factor. J Cell Biol 123:1649±1659.
Moore MS, Blobel G. 1994. A G protein involved innucleocytoplasmic transport: the role of Ran. TrendsBiochem Sci 19:211±216.
Petraglia F, Florio P, Nappi C, Genazzani AR. 1996.Peptide signaling in human placenta and membranes:autocrine, paracrine, and endocrine mechanisms. EndocrRev 17:156±186.
Rubins JB, Benditt JO, Dickey BF, Riedel N. 1990. GTP-binding proteins in rat liver nuclear envelopes. Proc NatlAcad Sci USA 87:7080±7084.
Seydel U, Gerace L. 1991. A 28,000-Da GDP/GTP-bindingprotein speci®c to the nuclear envelope. J Biol Chem266:7602±7608.
Singh US, Erickson JW, Cerione RA. 1995. Identi®cationand biochemical characterization of an 80 kDa GTP-binding/transglutaminase from rabbit liver nuclei. Bio-chemistry 34:15863±15871.
Takai Y, Sasaki T, Matozaki T. 2001. Small GTP-bindingproteins. Physiol Rev 81:153±208.
Thomson M. 1998. What are guanosine triphosphate-binding proteins doing in mitochondria? Biochim Bio-phys Acta; Mol Cell Res 1403:211±218.
Thomson M, Lim G, Hall PF, Kuyznierewicz I. 1998.Overlay blot identi®cation of GTP-binding proteins inmitochondria from human placenta. Placenta 19:209±215.
Zeuzem S, Zimmermann P, Schulz I. 1992. Activation of 19-and a 21 kDa GTP binding protein to pancreaticmicrosomal vesicles is regulated by the intravesicularpH established by a vacuolarÐtype H (�)ÐATPase. JMembr Biol 125:231±241.
Small GTP-Binding Proteins in the Nuclei 107