Embed Size (px)
Citation preview

ABSTRACT: Skeletal muscle hyperexcitability is characteristically associatedwith denervation. Expression of SK3, a small conductance Ca2�-activated K�
channel (SK channel) in skeletal muscle is induced by denervation, and directapplication of apamin, a peptide blocker of SK channels, dramatically reduceshyperexcitability. To investigate the role of SK3 channels in denervation-induced hyperexcitability, SK3 expression was manipulated using a transgenicmouse that harbors a tetracycline-regulated SK3 gene. Electromyographic(EMG) recordings from anterior tibial (AT) muscle showed that denervatedmuscle from transgenic or wild-type animals had equivalent hyperexcitabilitythat was blocked by apamin. In contrast, denervated skeletal muscle fromSK3tTA mice lacking SK3 channels showed little or no hyperexcitability, similarto results from wild-type innervated skeletal muscle. However, innervated skel-etal muscle from SK3tTA mice containing SK3 channels did not show hyper-excitability. The results demonstrate that SK3 channels are necessary but notsufficient for denervation-induced skeletal muscle hyperexcitability.
© 2002 Wiley Periodicals, Inc. Muscle Nerve 26: 817–822, 2002
SK CHANNELS ARE NECESSARY BUT NOTSUFFICIENT FOR DENERVATION-INDUCEDHYPEREXCITABILITY
DAVID JACOBSON, BS, 2 PACO S. HERSON, PhD,1 TORBEN R. NEELANDS, PhD, 1
JAMES MAYLIE, PhD, 3 and JOHN P. ADELMAN, PhD 1
1 Vollum Institute, L-474, Oregon Health and Sciences University, 3181 S.W. Sam Jackson Park Road,Portland, Oregon 97201, USA2 Department of Molecular and Medical Genetics, Oregon Health and Sciences University,Portland, Oregon, USA3 Department of Obstetrics and Gynecology, Oregon Health and Sciences University,Portland, Oregon, USA
Accepted 26 August 2002
Spontaneous repetitive discharges of action poten-tials (hyperexcitability) are characteristic of dener-vated skeletal muscle.1,18,23 Denervation induceschanges in the expression of many genes encodingproteins that are involved in excitability,10,21,24,25 andthe SK3 gene is a prototypic example. SK3 mRNA isminimally expressed in normal innervated skeletalmuscle,20 but following denervation, SK3 mRNA lev-els increase dramatically over 4 days to a maximalsteady-state level.17 The changes in SK3 gene expres-sion are paralleled by the appearance of 125I-apaminbinding sites, and the presence of apamin-sensitivepotassium conductances decrease the threshold for
action potential generation.16,17 Interestingly, directinjection of apamin, a blocker of small conductanceCa2�-activated K� channels (SK channels), into de-nervated rat skeletal muscle26 dramatically sup-pressed the hyperexcitability.
The decreased hyperexcitability subsequent toapamin application16,26 suggests an important role forSK3 channels in denervation-induced hyperexcitabil-ity. However, the many other changes in skeletal muscleassociated with denervation have not permitted a clearassessment of whether SK3 channels are sufficient to in-duce hyperexcitability. The availability of transgenic micein which SK3 gene expression may be experimentallycontrolled5 affords a mechanism to assess the link be-tween SK3 channels and skeletal muscle hyperexcitability.In this study, electromyographic (EMG) recordings wereemployed to measure electrical activity in normal or de-nervated skeletal muscles from wild-type mice and SK3transgenic mice that either overexpressed or lacked SK3channels. The results show that SK3 channels are neces-sary but not sufficient for denervation-induced skeletalmuscle hyperexcitability.
Abbreviations: AT, anterior tibial; dox, doxycycline; e-c, excitation–contrac-tion; EK, potassium equilibrium potential; EMG, electromyography; FDB,flexor digitorum brevis; PSD, power spectral density; SK channel, small con-ductance Ca2�-activated K� channel; SK3tTA mice, SK3 tetracycline regu-latable transgenic miceKey words: electromyography; denervation; hyperexcitability; skeletal mus-cle; SK3Correspondence to: J. P. Adelman; e-mail: [email protected]
© 2002 Wiley Periodicals, Inc.
SK3 and Muscle Hyperexcitability MUSCLE & NERVE December 2002 817

MATERIALS AND METHODS
Denervation. Animal care and handling were inaccordance with Institutional Animal Care andUse Committee guidelines. Mice between the agesof P8 –P10 were anesthetized with an isoflourane-oxygen mix. In one hindlimb, a section of thesciatic nerve (5 mm) was excised through a small(3–5 mm) incision over the hip. The incision wassutured with a single stitch and formulated cyano-acrylate (Vetbond; 3M, St. Paul, Minnesota) wasapplied to the wound. For tissue harvest, animalswere humanely sacrificed 4 –10 days after denerva-tion by deep isoflourane anesthesia and cervicaldislocation.
Electrophysiology. SK currents were examined inmuscle fibers from the flexor digitorum brevis mus-cle (FDB) as previously described.16 Whole-cell re-cordings were performed using an EPC9 (HekaElektronik, Mahone Bay, Nova Scotia, Canada) orAxopatch 200B (Axon Instruments, Union City, Cal-ifornia) amplifier and Pulse software (Heka Elek-tronik) interfaced to a Macintosh computer (AppleComputer, Cupertino, California); series resistance(50–80%) and whole-cell capacitance were electron-ically compensated. Data analysis was performed off-line using PulseFit (Heka Elektronik) and igor soft-ware (WaveMetrics, Lake Oswego, Oregon).
Electromyography. EMG recordings were per-formed under isoflourane anesthesia using 30-gaugeconcentric needle electrodes (Meditech, Westwood,Massachusetts) with sampling of at least three sepa-rate locations within the anterior tibial (AT) muscle.The signal was amplified through a DAM 50 ampli-fier (World Precision Instruments, Sarasota, Florida)and digitally recorded using a Mac Lab chart re-corder. Analysis was performed using the data re-corded during the 1st min and the 4th min followinginjection of either apamin (Alomone Laboratories,Jerusalem, Israel; 50 �l of 10 �g/ml) or saline. EMGactivity was analyzed using power spectral densityanalysis and significance was determined using bothpaired t-test and analysis of variance (ANOVA).Fast Fourier transforms, applied to 1,024-pointsegments, were performed on EMG traces over a60-s interval. The magnitude of the fast Fouriertransforms, sum of the squares of the real andimaginary parts, was averaged to produce a powerspectral density. The power spectral density wasintegrated over a frequency range of 5 to 50 Hz foreach trace.
Western Blot Analysis. Membrane proteins wereprepared from cultured cells or muscle tissue.Briefly, cells were dounce-homogenized in 10 mMTris (pH 7.5) with protease inhibitors and debris waspelleted at 1,000g for 10 min. The membranes werepelleted from the supernatant at 50,000g for 10 min,resuspended in 0.6M potassium iodide, repelletedfor 10 min, washed twice in 10 mM Tris (pH 7.5),and resuspended in 20 mM Tris (pH 7.5) with pro-tease inhibitors. Following electrophoresis throughan 8% denaturing polyacrylamide gel, 10 �g of pro-tein were prepared as a Western blot on nitrocellu-lose. SK3 antibody (Alomone Laboratories, Jerusa-lem, Israel) was used to probe the membranes (1:1,000 dilution), washed with phosphate-bufferedsaline containing 0.1% Tween, and incubated withgoat anti-rabbit horseradish peroxidase-coupled sec-ondary antibody (Santa Cruz Biotechnology, SantaCruz, California; 1:25,000 dilution). Horseradishperoxidase was illuminated using Pico-signal (Pierce,Rockford, Illinois) and exposed on Kodak X-omatBlue film (Kodak, Rochester, New York).
RESULTS
Denervation Induces Apamin-Sensitive Hyperexcitabil-
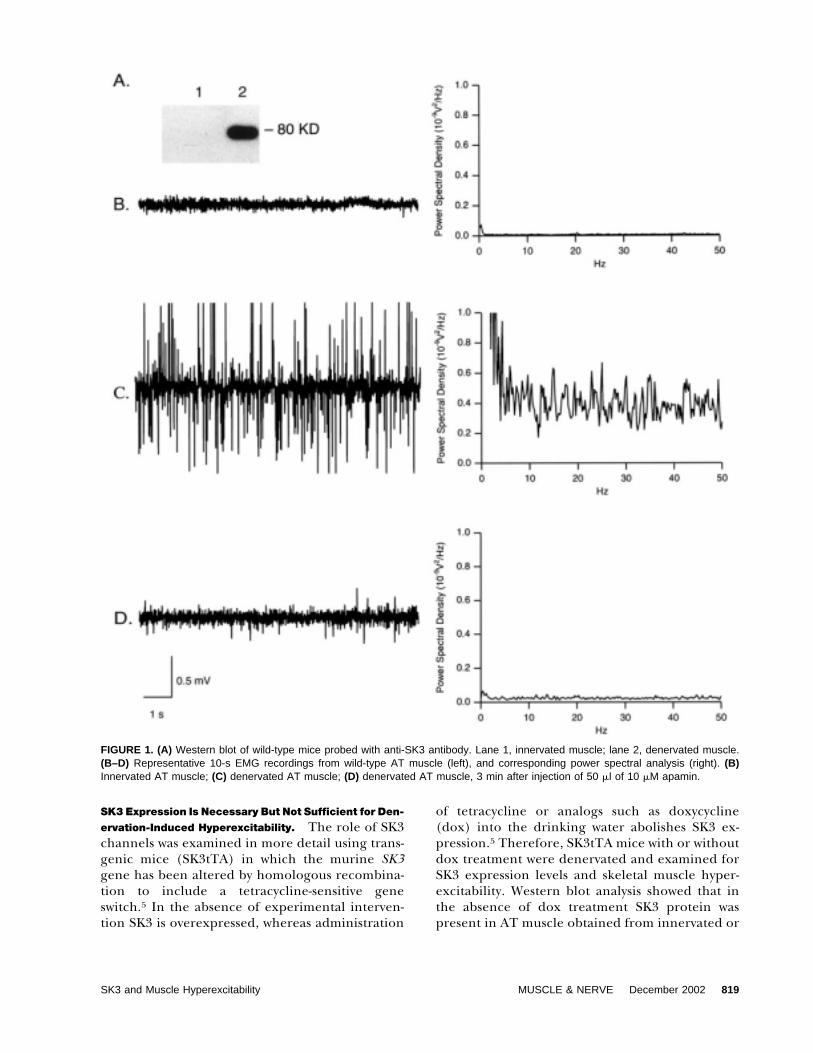
ity and SK3 Protein. Adult C57/Black6 mice weredenervated by surgically removing a 5-mm segmentof the right hindlimb sciatic nerve; the intact lefthindlimb served as a control. Electromyographic(EMG) recordings from hindlimb AT muscles wereperformed 7 to 10 days after denervation, measuringspontaneous EMG activity as an index of excitability.The EMG data from denervated and control muscleswere quantified for comparison using power spectraldensity (PSD) analysis, performed over a set 1-mininterval on all EMG recordings (see Materials andMethods). In agreement with previous studies,4,26
innervated skeletal muscle exhibited no significantspontaneous activity, as evidenced by a low PSD(2.0 � 0.3 � 10�10 V2/Hz, n � 4; Fig. 1B). Incontrast, denervated skeletal muscle exhibited ro-bust spontaneous activity, and a 100-fold higher PSD(1.9 � 0.3 � 10�8 V2/Hz, n � 4; Fig. 1C). Apamininjection into the denervated muscle resulted in a10-fold decrease in spontaneous activity within 3 min(1.9 � 0.4 � 10�9 V2/Hz, n � 4; Fig. 1D). FollowingEMG recordings, animals were sacrificed and SK3protein levels in the hindlimbs were assessed byWestern blot analysis. The results showed that SK3protein was detected only in denervated muscle (Fig.1A). These results implicate apamin-sensitive SK3channels in denervation-induced skeletal muscle hy-perexcitability.
818 SK3 and Muscle Hyperexcitability MUSCLE & NERVE December 2002
SK3 Expression Is Necessary But Not Sufficient for Den-
ervation-Induced Hyperexcitability. The role of SK3channels was examined in more detail using trans-genic mice (SK3tTA) in which the murine SK3gene has been altered by homologous recombina-tion to include a tetracycline-sensitive geneswitch.5 In the absence of experimental interven-tion SK3 is overexpressed, whereas administration
of tetracycline or analogs such as doxycycline(dox) into the drinking water abolishes SK3 ex-pression.5 Therefore, SK3tTA mice with or withoutdox treatment were denervated and examined forSK3 expression levels and skeletal muscle hyper-excitability. Western blot analysis showed that inthe absence of dox treatment SK3 protein waspresent in AT muscle obtained from innervated or
FIGURE 1. (A) Western blot of wild-type mice probed with anti-SK3 antibody. Lane 1, innervated muscle; lane 2, denervated muscle.(B–D) Representative 10-s EMG recordings from wild-type AT muscle (left), and corresponding power spectral analysis (right). (B)Innervated AT muscle; (C) denervated AT muscle; (D) denervated AT muscle, 3 min after injection of 50 �l of 10 �M apamin.
SK3 and Muscle Hyperexcitability MUSCLE & NERVE December 2002 819
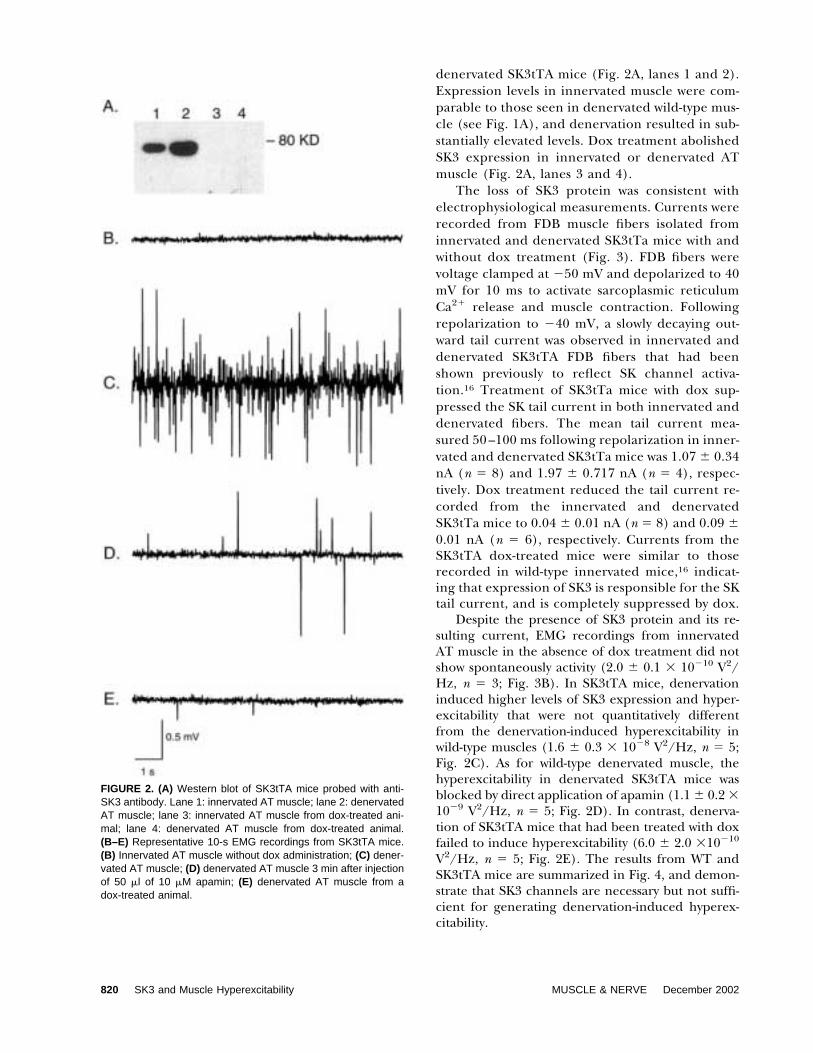
denervated SK3tTA mice (Fig. 2A, lanes 1 and 2).Expression levels in innervated muscle were com-parable to those seen in denervated wild-type mus-cle (see Fig. 1A), and denervation resulted in sub-stantially elevated levels. Dox treatment abolishedSK3 expression in innervated or denervated ATmuscle (Fig. 2A, lanes 3 and 4).
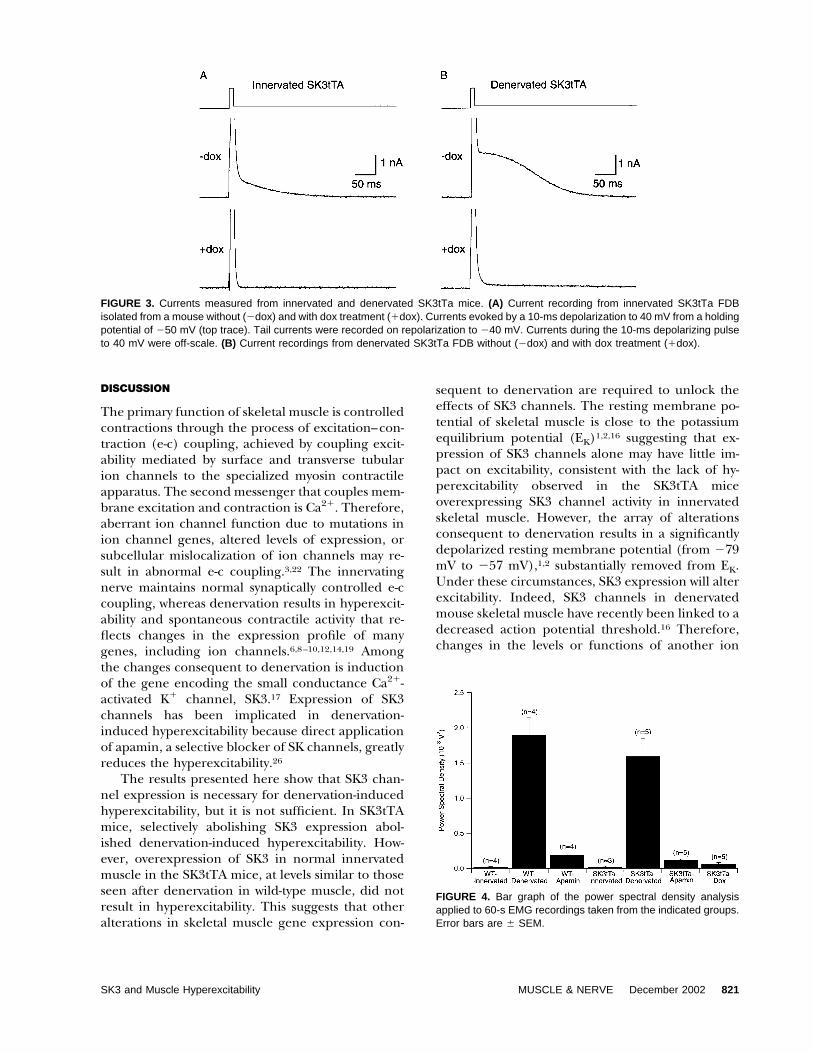
The loss of SK3 protein was consistent withelectrophysiological measurements. Currents wererecorded from FDB muscle fibers isolated frominnervated and denervated SK3tTa mice with andwithout dox treatment (Fig. 3). FDB fibers werevoltage clamped at �50 mV and depolarized to 40mV for 10 ms to activate sarcoplasmic reticulumCa2� release and muscle contraction. Followingrepolarization to �40 mV, a slowly decaying out-ward tail current was observed in innervated anddenervated SK3tTA FDB fibers that had beenshown previously to reflect SK channel activa-tion.16 Treatment of SK3tTa mice with dox sup-pressed the SK tail current in both innervated anddenervated fibers. The mean tail current mea-sured 50 –100 ms following repolarization in inner-vated and denervated SK3tTa mice was 1.07 � 0.34nA (n � 8) and 1.97 � 0.717 nA (n � 4), respec-tively. Dox treatment reduced the tail current re-corded from the innervated and denervatedSK3tTa mice to 0.04 � 0.01 nA (n � 8) and 0.09 �0.01 nA (n � 6), respectively. Currents from theSK3tTA dox-treated mice were similar to thoserecorded in wild-type innervated mice,16 indicat-ing that expression of SK3 is responsible for the SKtail current, and is completely suppressed by dox.
Despite the presence of SK3 protein and its re-sulting current, EMG recordings from innervatedAT muscle in the absence of dox treatment did notshow spontaneously activity (2.0 � 0.1 � 10�10 V2/Hz, n � 3; Fig. 3B). In SK3tTA mice, denervationinduced higher levels of SK3 expression and hyper-excitability that were not quantitatively differentfrom the denervation-induced hyperexcitability inwild-type muscles (1.6 � 0.3 � 10�8 V2/Hz, n � 5;Fig. 2C). As for wild-type denervated muscle, thehyperexcitability in denervated SK3tTA mice wasblocked by direct application of apamin (1.1 � 0.2 �10�9 V2/Hz, n � 5; Fig. 2D). In contrast, denerva-tion of SK3tTA mice that had been treated with doxfailed to induce hyperexcitability (6.0 � 2.0 �10�10
V2/Hz, n � 5; Fig. 2E). The results from WT andSK3tTA mice are summarized in Fig. 4, and demon-strate that SK3 channels are necessary but not suffi-cient for generating denervation-induced hyperex-citability.
FIGURE 2. (A) Western blot of SK3tTA mice probed with anti-SK3 antibody. Lane 1: innervated AT muscle; lane 2: denervatedAT muscle; lane 3: innervated AT muscle from dox-treated ani-mal; lane 4: denervated AT muscle from dox-treated animal.(B–E) Representative 10-s EMG recordings from SK3tTA mice.(B) Innervated AT muscle without dox administration; (C) dener-vated AT muscle; (D) denervated AT muscle 3 min after injectionof 50 �l of 10 �M apamin; (E) denervated AT muscle from adox-treated animal.
820 SK3 and Muscle Hyperexcitability MUSCLE & NERVE December 2002
DISCUSSION
The primary function of skeletal muscle is controlledcontractions through the process of excitation–con-traction (e-c) coupling, achieved by coupling excit-ability mediated by surface and transverse tubularion channels to the specialized myosin contractileapparatus. The second messenger that couples mem-brane excitation and contraction is Ca2�. Therefore,aberrant ion channel function due to mutations inion channel genes, altered levels of expression, orsubcellular mislocalization of ion channels may re-sult in abnormal e-c coupling.3,22 The innervatingnerve maintains normal synaptically controlled e-ccoupling, whereas denervation results in hyperexcit-ability and spontaneous contractile activity that re-flects changes in the expression profile of manygenes, including ion channels.6,8–10,12,14,19 Amongthe changes consequent to denervation is inductionof the gene encoding the small conductance Ca2�-activated K� channel, SK3.17 Expression of SK3channels has been implicated in denervation-induced hyperexcitability because direct applicationof apamin, a selective blocker of SK channels, greatlyreduces the hyperexcitability.26
The results presented here show that SK3 chan-nel expression is necessary for denervation-inducedhyperexcitability, but it is not sufficient. In SK3tTAmice, selectively abolishing SK3 expression abol-ished denervation-induced hyperexcitability. How-ever, overexpression of SK3 in normal innervatedmuscle in the SK3tTA mice, at levels similar to thoseseen after denervation in wild-type muscle, did notresult in hyperexcitability. This suggests that otheralterations in skeletal muscle gene expression con-
sequent to denervation are required to unlock theeffects of SK3 channels. The resting membrane po-tential of skeletal muscle is close to the potassiumequilibrium potential (EK)1,2,16 suggesting that ex-pression of SK3 channels alone may have little im-pact on excitability, consistent with the lack of hy-perexcitability observed in the SK3tTA miceoverexpressing SK3 channel activity in innervatedskeletal muscle. However, the array of alterationsconsequent to denervation results in a significantlydepolarized resting membrane potential (from �79mV to �57 mV),1,2 substantially removed from EK.Under these circumstances, SK3 expression will alterexcitability. Indeed, SK3 channels in denervatedmouse skeletal muscle have recently been linked to adecreased action potential threshold.16 Therefore,changes in the levels or functions of another ion
FIGURE 4. Bar graph of the power spectral density analysisapplied to 60-s EMG recordings taken from the indicated groups.Error bars are � SEM.
FIGURE 3. Currents measured from innervated and denervated SK3tTa mice. (A) Current recording from innervated SK3tTa FDBisolated from a mouse without (�dox) and with dox treatment (�dox). Currents evoked by a 10-ms depolarization to 40 mV from a holdingpotential of �50 mV (top trace). Tail currents were recorded on repolarization to �40 mV. Currents during the 10-ms depolarizing pulseto 40 mV were off-scale. (B) Current recordings from denervated SK3tTa FDB without (�dox) and with dox treatment (�dox).
SK3 and Muscle Hyperexcitability MUSCLE & NERVE December 2002 821

channel following denervation, that induce a de-creased resting membrane potential, are necessaryfor the SK3-mediated hyperexcitability. Althoughthe proteins underlying the decreased resting mem-brane potential have not yet been identified, lowerexpression levels of chloride channels, inwardly rec-tifying potassium channels, or sodium channel iso-form switches are possible candidates.10,24,25
Potassium channel activity generally causes a hy-perpolarizing effect on membrane potential, raisingthe apparent paradox of K� channel-mediated hy-perexcitability: why do SK3 channels make skeletalmuscle hyperexcitable? This may reflect the uniquecytoarchitecture of skeletal muscle. Expression ofSK3 channels in the transverse tubules may result inan accumulation of extracellular K� that does notreadily diffuse from the restricted tubular space, re-sulting in a depolarizing, hyperexcitable condition.Support for this model comes from measurements ofSK current reversal potentials in denervated skeletalmuscle in different extracellular K� concentra-tions.16
Many different inherited myotonic disorders re-sult in skeletal muscle hyperexcitability. However,only the myotonia associated with myotonic dystro-phy exhibits apamin sensitivity4 and increased SK3expression.11 Interestingly, patients with that disor-der develop varying degrees of neuropathy.7,13,15
Therefore it is possible that compromised nervefunction or nerve death consequent to myotonicdystrophy may lead to hyperexcitability that mirrorsthe changes induced by acute denervation.
We thank Ms. Chris Bond for the SK3tTA mice and criticalreading of the manuscript. We also thank Dr. Jane MacPhersonfor expert assistance with EMG recordings. This work was sup-ported by a Tartar Trust Fellowship (D.J.) and NIH grants (J.M.and J.P.A.).
REFERENCES
1. Albuquerque EX, Thesleff S. A comparative study of mem-brane properties of innervated and chronically denervatedfast and slow skeletal muscles of the rat. Acta Physiol Scand1968;73:471–480.
2. Almers W. Potassium concentration changes in the transversetubules of vertebrate skeletal muscle. Fed Proc 1980;39:1527–1532.
3. Beam KG, Knudson CM, Powell JA. A lethal mutation in miceeliminates the slow calcium current in skeletal muscle cells.Nature 1986;320:168–170.
4. Behrens MI, Jalil P, Serani A, Vergara F, Alvarez O. Possiblerole of apamin-sensitive K� channels in myotonic dystrophy.Muscle Nerve 1994;17:1264–1270.
5. Bond CT, Sprengel R, Bissonnette JM, Kaufmann WA, Prib-now D, Neelands T, Storck T, Baetscher M, Jerecic J, Maylie J,Knaus HG, Seeburg PH, Adelman JP. Respiration and partu-rition affected by conditional overexpression of the Ca2�-activated K� channel subunit, SK3. Science 2000;289:1942–1946.
6. Brenner HR, Witzemann V, Sakmann B. Imprinting of ace-tylcholine receptor messenger RNA accumulation in mamma-lian neuromuscular synapses. Nature 1990;344:544–547.
7. Coers C. Les variations structurelles normales etpathologiques de la junction neuromusculair. Acta NeurolPsychiatr Belg 1955;55:741–866.
8. Gonoi T, Hasegawa S. Postnatal induction and neural regu-lation of inward rectifiers in mouse skeletal muscle. PflugersArch 1991;418:601–607.
9. Hartzell HC, Fambrough DM. Acetylcholine receptors. Distri-bution and extrajunctional density in rat diaphragm afterdenervation correlated with acetylcholine sensitivity. J GenPhysiol 1972;60:248–262.
10. Heathcote RD. Acetylcholine-gated and chloride conduc-tance channel expression in rat muscle membrane. J Physiol1989;414:473–497.
11. Kimura T, Takahashi MP, Okuda Y, Kaido M, Fujimura H,Yanagihara T, Sakoda S. The expression of ion channel mR-NAs in skeletal muscles from patients with myotonic musculardystrophy. Neurosci Lett 2000;295:93–96.
12. Lupa MT, Caldwell JH. Sodium channels aggregate at formersynaptic sites in innervated and denervated regenerating mus-cles. J Cell Biol 1994;124:139–147.
13. MacDermot V. The histology of neuromuscular junction inmyotonic dystrophy. Brain 1961;84:75–84.
14. Mishina M, Takai T, Imoto K, Noda M, Takahashi T, Numa S,Methfessel C, Sakmann B. Molecular distinction between fetaland adult forms of muscle acetylcholine receptor. Nature1986;321:406–411.
15. Mondelli M, Rossi A, Malandrini A, Della Porta P, Guzaai GC.Axonal motor and sensory neuropathy in myotonic dystrophy.Acta Neurol Scand 1993;88:141–148.
16. Neelands TR, Herson PS, Jacobson D, Adelman JP, Maylie J.Small-conductance calcium-activated potassium currents inmouse hyperexcitable denervated skeletal muscle. J Physiol2001;536:397–407.
17. Pribnow D, Johnson-Pais T, Bond CT, Keen J, Johnson RA,Janowsky A, Silvia C, Thayer M, Maylie J, Adelman JP. Skeletalmuscle and small-conductance calcium-activated potassiumchannels. Muscle Nerve 1999;22:742–750.
18. Robbins N. Cation movements in normal and short-termdenervated rat fast twitch muscle. J Physiol 1977;271:605–624.
19. Rogart RB, Regan LJ. Two subtypes of sodium channel withtetrodotoxin sensitivity and insensitivity detected in dener-vated mammalian skeletal muscle. Brain Res 1985;329:314–318.
20. Roncarati R, Di Chio M, Sava A, Terstappen GC, Fumagalli G.Presynaptic localization of the small conductance calcium-activated potassium channel SK3 at the neuromuscular junc-tion. Neuroscience 2001;104:253–262.
21. Sellin LC, Thesleff S. Alterations in membrane electricalproperties during long-term denervation of rat skeletal mus-cles. Acta Physiol Scand 1980;108:243–246.
22. Takekura H, Nishi M, Noda T, Takeshima H, Franzini-Arm-strong C. Abnormal junctions between surface membraneand sarcoplasmic reticulum in skeletal muscle with a muta-tion targeted to the ryanodine receptor. Proc Natl Acad SciUSA 1995;92:3381–3385.
23. Tower SS. The reaction of muscle to denervation. Physiol Res1939;19:1–48.
24. Trimmer JS, Cooperman SS, Agnew WS, Mandel G. Regula-tion of muscle sodium channel transcripts during develop-ment and in response to denervation. Dev Biol 1990;142:360–367.
25. Venosa RA, Kotsias BA. Potassium movements in denervatedfrog sartorius muscle. Am J Physiol 1985;248:219–227.
26. Vergara C, Ramirez B, Beherens MI. Colchicine alters apaminreceptors, electrical activity, and skeletal muscle relaxation.Muscle Nerve 1993;16:935–940.
822 SK3 and Muscle Hyperexcitability MUSCLE & NERVE December 2002