Embed Size (px)
Citation preview

(141), ra69. [DOI: 10.1126/scisignal.2001140] 3Science Signalingand Steven M. Smith (28 September 2010) Ping Che, John D. Bussell, Wenxu Zhou, Gonzalo M. Estavillo, Barry J. Pogson
ArabidopsisSignaling and Promotes Acclimation to Stress in Signaling from the Endoplasmic Reticulum Activates Brassinosteroid`
This information is current as of 7 January 2014. The following resources related to this article are available online at http://stke.sciencemag.org.
Article Tools http://stke.sciencemag.org/cgi/content/full/sigtrans;3/141/ra69
Visit the online version of this article to access the personalization and article tools:
MaterialsSupplemental
http://stke.sciencemag.org/cgi/content/full/sigtrans;3/141/ra69/DC1 "Supplementary Materials"
Related Content
http://stke.sciencemag.org/cgi/cm/stkecm;CMP_19131 http://stke.sciencemag.org/cgi/content/abstract/sigtrans;4/201/ec332 http://stke.sciencemag.org/cgi/content/abstract/sigtrans;6/290/ec204
's sites:ScienceThe editors suggest related resources on
References http://stke.sciencemag.org/cgi/content/full/sigtrans;3/141/ra69#BIBL
11 article(s) hosted by HighWire Press; see: cited byThis article has been
http://stke.sciencemag.org/cgi/content/full/sigtrans;3/141/ra69#otherarticlesThis article cites 54 articles, 21 of which can be accessed for free:
Glossary http://stke.sciencemag.org/glossary/
Look up definitions for abbreviations and terms found in this article:
Permissions http://www.sciencemag.org/about/permissions.dtl
Obtain information about reproducing this article:
the American Association for the Advancement of Science; all rights reserved. byAssociation for the Advancement of Science, 1200 New York Avenue, NW, Washington, DC 20005. Copyright 2008
(ISSN 1937-9145) is published weekly, except the last week in December, by the AmericanScience Signaling
on January 7, 2014 stke.sciencem
ag.orgD
ownloaded from
on January 7, 2014
stke.sciencemag.org
Dow
nloaded from

R E S E A R C H A R T I C L E
P L A N T B I O L O G Y
Signaling from the Endoplasmic ReticulumActivates Brassinosteroid Signaling and PromotesAcclimation to Stress in ArabidopsisPing Che,1,2* John D. Bussell,1† Wenxu Zhou,1,2† Gonzalo M. Estavillo,3
Barry J. Pogson,3 Steven M. Smith1,2*
(Published 28 September 2010; Volume 3 Issue 141 ra69)Dow
nloaded fr
The ability to acclimate to stresses enables plants to grow and develop under adverse environmentalconditions. Regulated intramembrane proteolysis (RIP) triggered by endoplasmic reticulum (ER) stressmediates some forms of stress signaling. Brassinosteroids (BRs) have been implicated in plant adapta-tion to stress, but no mechanisms for activation have been discovered. Here, we reveal a connectionbetween ER stress signaling and BR-mediated growth and stress acclimation. Arabidopsis transcriptionfactors bZIP17 and bZIP28 were translocated from the ER through the Golgi, where they were proteolyt-ically cleaved by site 2 protease and released to translocate into the nucleus. Stresses, including heatand inhibition of protein glycosylation, increased translocation of these two bZIPs to the nucleus. Thesenuclear-localized bZIPs not only activated ER chaperone genes but also activated BR signaling, whichwas required for stress acclimation and growth. Thus, these bZIPs link ER stress and BR signaling,which may be a mechanism by which plant growth and stress responses can be integrated.
som
on January 7, 2014tke.sciencemag.org
INTRODUCTION
Brassinosteroid (BR) regulates many growth and developmental pro-cesses, such as cell elongation, vascular development, and photomorpho-genesis (1–4). Exogenous BR treatment can also induce plant tolerance toa wide spectrum of biotic and abiotic stresses, such as pathogen attack,low and high temperatures, drought, and high salt (5–9). Although hightemperature has been reported to activate BR signaling (10), it is notknown whether endogenous BR signaling normally provides toleranceto stresses, how such stresses activate BR signaling, or which responsemechanisms are activated by such BR signaling (11).
Regulated intramembrane proteolysis (RIP) is a conserved stressresponse mechanism in eukaryotes (12) (fig. S1). In mammals, insects,and fungi, it plays a key role in sensing stresses, such as those resultingfrom changes in metabolism of lipids (13, 14) and sterols (15, 16), oxygendeficit (17–19), and the unfolded protein response (UPR) (20, 21). RIPactivates a specific subset of transcription factors that reside as trans-membrane proteins within the endoplasmic reticulum (ER) in unstressedcells. In response to stress, these transcription factors are translocated tothe Golgi where they are released into the cytosol by proteolytic cleavageby two membrane-embedded proteases (12, 20). One is a subtilisin-likeserine protease, which cleaves at one specific site (site 1 protease or S1P),and the other is a metalloprotease (site 2 protease or S2P), which cleavesat a second site within the transmembrane domain. Consequently, the
1Australian Research Council Centre of Excellence in Plant Energy Biology,University of Western Australia, Crawley, Western Australia 6009, Aus-tralia. 2Centre of Excellence for Plant Metabolomics, University of WesternAustralia, Crawley, Western Australia 6009, Australia. 3Australian ResearchCouncil Centre of Excellence in Plant Energy Biology, Research School ofBiology, Australian National University, Canberra, Australian Capital Territory0200, Australia.*To whom correspondence should be addressed. E-mail: [email protected] (S.M.S.); [email protected] (P.C.)†These authors contributed equally to this work.
www.S
transcription factors are released from the Golgi membrane and translo-cated to the nucleus where they activate the expression of their targetgenes (12, 15, 20).
In plants, RIP is implicated in responses to stresses caused by proteinmisfolding as well as excess salt and heat (22–28) (fig. S1). ArabidopsisS1P and S2P homologs have been identified (22, 29, 30). S1P providestolerance to salt stress by activating bZIP17 (22), a mechanism similar tothe ER stress response mediated by the transcription factor ATF6 in mam-mals (20, 21) (fig. S1). Two other bZIP17 homologs, bZIP28 and bZIP49,have also been identified. bZIP28 mediates tunicamycin (TM)–inducedER stress responses related to UPR (23, 27, 28) and is also activated inresponse to heat shock (31). The function of bZIP49 is unknown.
Through our investigation of the function of S2P, we discovered a con-nection among ER stress sensing, BR signaling, and stress acclimation.
RESULTS
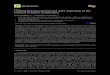
S2P mediates bZIP proteolysis andnuclear translocationWe visualized the expression of the S2P gene in transgenic plants express-ing the GUS reporter under the control of the S2P promoter. S2P::GUSwas expressed mainly in the vasculature of cotyledons, leaves, and roots ina pattern identical to that of S1P::GUS (fig. S2A). With a 35S::S2P-GFP(green fluorescent protein) transgene, we established that S2P localized tothe Golgi (Fig. 1A), as did S1P (fig. S2B). We investigated the proteolysisand subcellular localization of bZIP17 and bZIP28 in 35S::GFP-bZIPtransgenic wild-type and s2p-null mutant plants (fig. S3, A and B). Inroots of wild-type plants, GFP-bZIP17 was observed mainly in the ERas previously demonstrated (22, 27), with little fluorescence in the nucleus(Fig. 1B), although more fluorescence was observed in the nuclei of somecells, particularly in the elongation zone (fig. S2C). However, GFP-bZIP17 relocalized predominantly to the nucleus within 1 hour of heatshock or 4 hours of treatment with TM, an agent that blocks N-linked
CIENCESIGNALING.org 28 September 2010 Vol 3 Issue 141 ra69 1
R E S E A R C H A R T I C L E
protein glycosylation in the secretory pathway and induces ER stress (Fig.1B). In the s2p mutant, GFP-bZIP17 was localized predominantly in theGolgi (Fig. 1C), and heat shock or TM treatment failed to alter its local-ization (Fig. 1B). GFP-bZIP28 exhibited an S2P-dependent distributionsimilar to GFP-bZIP17 (fig. S2, D and E).
www.S
To demonstrate whether, in addition to nuclear localization, bZIP17proteolysis was also S2P-dependent, we monitored the size shift of theGFP-bZIP17 fusion protein before and after heat shock treatments in bothwild-type and s2p mutant backgrounds. In the absence of heat shock, wedetected three main bands (I, II, and III) of GFP-bZIP in wild-type roots
CIENCESIGNALING.org 28 Se
on January 7, 2014 stke.sciencem
ag.orgD
ownloaded from
(Fig. 1D). Heat shock induced a mobilityshift of band I (the most abundant bandbefore heat shock) to band III (the mostabundant band after heat shock), suggest-ing that band I was the full-length GFP-bZIP17 localized in the ER and band IIIwas the truncated N-terminal form local-ized in the nucleus (Fig. 1D). Althoughband III was about the size predicted forS2P-processed GFP-bZIP17 fusion protein(~70 kD), band I was slightly bigger thancalculated full-length GFP-bZIP17 fusionprotein (~108 kD). We determined that bandI was glycosylated GFP-bZIP17, because itwas converted to band II when the proteinextract was treated with b-N-acetylglucosa-minidase H (Endo H) (Fig. 1E). However,in s2p, only band II, the Endo H–resistantform, was observed in the presence orabsence of heat shock (Fig. 1, D and E),indicating that GFP-bZIP17 was retainedin the Golgi and was not cleaved even un-der stress conditions. Collectively, these re-sults show that proteolysis, and relocationof bZIPs from ER to the nucleus, occursthrough the Golgi and is S2P-dependent.Because of the presence of the Endo H–resistant form in unstressed cells (Fig. 1E)and accumulation of bZIP17 in the Golgiin unstressed s2p cells (Fig. 1, B and C),these results also suggest that transit ofbZIPs through the Golgi to the nucleusoccurs even in the absence of stress butis greater when plants are subjected tostress.
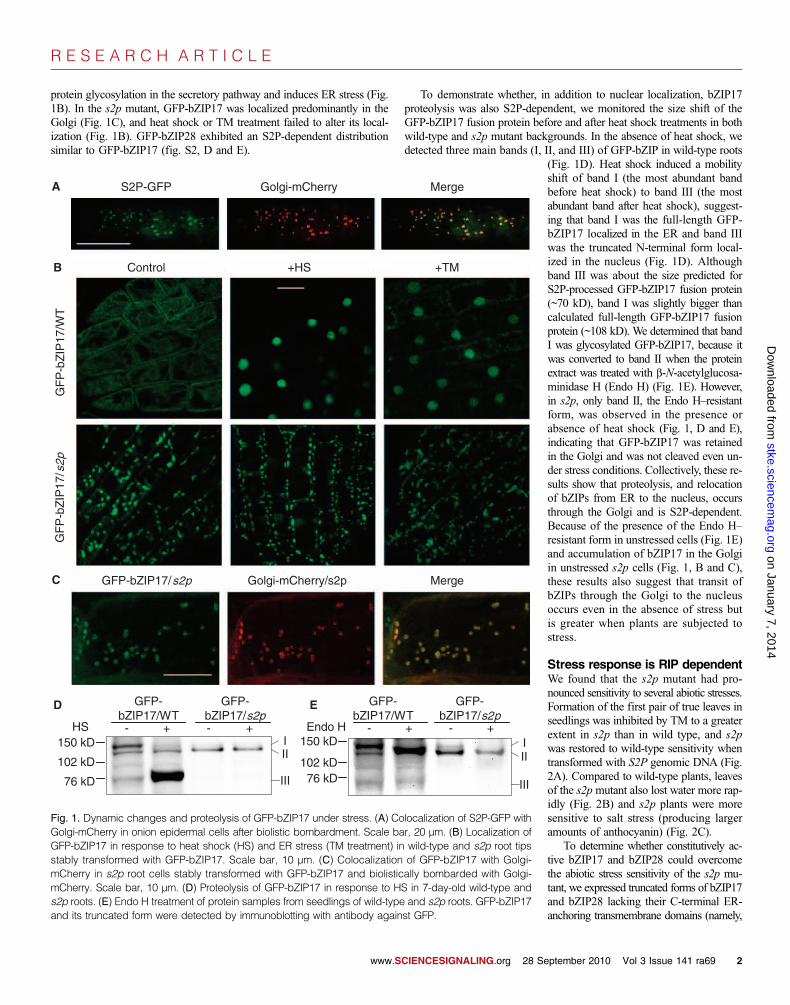
Stress response is RIP dependentWe found that the s2p mutant had pro-nounced sensitivity to several abiotic stresses.Formation of the first pair of true leaves inseedlings was inhibited by TM to a greaterextent in s2p than in wild type, and s2pwas restored to wild-type sensitivity whentransformed with S2P genomic DNA (Fig.2A). Compared to wild-type plants, leavesof the s2pmutant also lost water more rap-idly (Fig. 2B) and s2p plants were moresensitive to salt stress (producing largeramounts of anthocyanin) (Fig. 2C).
To determine whether constitutively ac-tive bZIP17 and bZIP28 could overcomethe abiotic stress sensitivity of the s2p mu-tant, we expressed truncated forms of bZIP17and bZIP28 lacking their C-terminal ER-anchoring transmembrane domains (namely,
A Golgi-mCherry Merge
Control +HS +TMB
C GFP-bZIP17/s2p Golgi-mCherry/s2p Merge
GF
P-b
ZIP
17/W
TG
FP
-bZ
IP17
/s2p
S2P-GFP
Endo H - + - +- + - +
D E
HS
76 kD
102 kD
150 kD III
III
III
III
GFP-bZIP17/WT
GFP-bZIP17/s2p
76 kD102 kD
150 kD
GFP-bZIP17/WT
GFP-bZIP17/s2p
Fig. 1. Dynamic changes and proteolysis of GFP-bZIP17 under stress. (A) Colocalization of S2P-GFP withGolgi-mCherry in onion epidermal cells after biolistic bombardment. Scale bar, 20 µm. (B) Localization ofGFP-bZIP17 in response to heat shock (HS) and ER stress (TM treatment) in wild-type and s2p root tipsstably transformed with GFP-bZIP17. Scale bar, 10 µm. (C) Colocalization of GFP-bZIP17 with Golgi-mCherry in s2p root cells stably transformed with GFP-bZIP17 and biolistically bombarded with Golgi-mCherry. Scale bar, 10 µm. (D) Proteolysis of GFP-bZIP17 in response to HS in 7-day-old wild-type ands2p roots. (E) Endo H treatment of protein samples from seedlings of wild-type and s2p roots. GFP-bZIP17and its truncated form were detected by immunoblotting with antibody against GFP.
ptember 2010 Vol 3 Issue 141 ra69 2
F
4
5
6
7
8
Col 0
bZIP17::bZIP17 C/s2p
bZIP28::bZIP28 C/s2p
35S::bZIP17 C/s2p0
0.03
ativ
e tr
ansc
ript
abun
danc
e
*
**
**
*
Col 0 (100%)
S2p (100%)
0.02 µg/ml TM
#4 (100%)
#5(98 ± 1%)S2P::S2P/s2p
Col 0 (100%)
s2p (5 ± 2%)
Col
0s2
p
bZIP
17::b
ZIP
17C
/s2p
bZIP
28::b
ZIP
28C
/s2p
B
Col 0s1p-3s2p
0
20
40
60
80
2 40 6 8 10 12 14
Leaf
wei
ght (
%)
Time (hours)
100
Co
l 0
s2
p
C
G
BiP
CRT1
bZIP28
::
bZIP28
C/s2p
Col
0
s2p
bZIP17
::
bZIP17
C/s2p
CNXs
(98 ± 2%)
(4 ± 1%)
(100%)
(98 ± 1%)
8
00
0
1.0
1.5
2.0
0.5
CNX2
1.01.52.02.53.03.5
0.50
4.0CRT1
Rel
ativ
e tr
ansc
ript
abun
danc
e
* **
*
**
1 0.5 1.5 1.9
E
101214 CNX1
46
8
2R
elat
ive
tran
scrip
tab
unda
nce
BiP1,2
51015
2025
3035
* ***
*
**
*
Col 0s2p
234567
10
CRT2
01234
5
PDIL1-167
Rel
ativ
e tr
ansc
ript
abun
danc
e
* *
*
***
2Cont.
2TM
4Cont.
4 hoursTM
2 Cont.
2 TM
4 Cont.
4 hoursTM
∆∆
∆∆
∆
∆∆
R E S E A R C H A R T I C L E
on January 7, 2014 stke.sciencem
ag.orgD
ownloaded from
Fig. 2. s2p is sensitive to various abioticstresses. (A) Phenotypes of 7-day-oldwild type (WT), s2p, and s2p trans-formed with S2P gene (S2P::S2P/s2p)grown on ½ MS with or without TM (twoindependent S2P::S2P/s2p transgeniclines were used, #4 and #5). (B) Waterloss from detached leaves. Leaves weredetached from 4-week-old soil-grownplants, allowed to dry in the growth cham-ber (23°C, 60% relative humidity), andweighed at indicated intervals. (C) Four-week-old plants were irrigated for 12days at 4-day intervals with increasingconcentrations of NaCl: 50, 100, 200,and 300mM. Stress sensitivity is indicatedby red pigmentation (anthocyanin accu-mulation). (D) TM sensitivity of s2p wasrescued by expressing bZIPDCs 7 daysafter sowing. (E) Quantitative reversetranscription PCR (qRT-PCR) analysis ofrelative gene expression of ER stressmarker genes in wild-type and s2p seed-lings treated with TM (5 mg/ml) for 2 or4 hours. Two-sample t tests were per-formed to compare expression of chap-erone genes of s2p with that of wild typeat the same time as TM treatment. *P <0.05. Cont., control. (F) Effects of bZIPDCson expression of ER stress marker genesin unstressed seedlings. Two-sample ttests were performed to compare ex-pression of chaperone genes of bZIPDCtransgenic plants with that of wild type.*P < 0.05. (G) Effects of bZIPDCs on theabundance of the indicated ER stress–related proteins in unstressed seedlings.Chaperones were immunodetected withantibodies against BiP and CRT (53).a-Tubulin (a-TUB) was probed as load-ing control. Numbers below each panelindicate the relative protein levels quan-tified by measuring band intensities withImageJ (http://rsb.info.nih.gov/ij/ ), nor-malized by dividing by the a-tubulinband intensity and presented as the val-ue relative to that of wild type. Experi-ments were performed three times withsimilar results. In (A) and (D), percentilerepresents the percentage of plants withthe first pair of leaves on the shoots(±SE from three independent counts).In (B) to (F), error bars indicate SE (n = 3).
A 0 µg/ml TM D 0.02 µg/ml TM
0
1
2
3
BiP1,2 BiP3 CNX1 CNX2 CRT1 CRT2
Rel * *
** *
**
***
α-TUB
CRT2
PDIL1-1
1 0.4 2.2 2.81 0.5 2.2 2.41 0.6 2.1 3.0
www.SCIENCESIGNALING.org 28 September 2010 Vol 3 Issue 141 ra69 3
R E S E A R C H A R T I C L E
on January 7, 2014 stke.sciencem
ag.orgD
ownloaded from
bZIP17DC and bZIP28DC) (fig. S4). The truncatedbZIPs performed their nuclear functions indepen-dently of RIP (22, 23, 27, 31). Either protein com-plemented the TM response of s2p (Fig. 2D),indicating that they have functionally redundant rolesin regulating ER stress responses and that the trun-cated bZIPs obviate the requirement for S2P-dependentproteolysis.
To further define the role of S2P in ER stress sig-naling, we examined the expression of genes encodingER chaperones. ER stress, such as that induced by TMtreatment, causes unfolded proteins to accumulate,leading to stimulation of genes encoding chaperones(23, 27, 28, 32). Quantitative real-time polymerasechain reaction (qPCR) showed that the basal expres-sion of some chaperone genes was lower in s2p seed-lings and that TM-stimulated induction of some ofthese genes (especially CRT1, CRT2, and PDIL) wasimpaired in s2p (Fig. 2E). This defective transcription-al response could explain the sensitivity of s2p to TM.The existence of other RIP-independent ER stress reg-ulatory pathways, such as that mediated by bZIP60(30) or heterotrimeric G protein signaling (33), couldexplain why the response of chaperone genes to TM isnot completely abolished in s2p. In the s2p mutantcontaining bZIP28DC, basal expression of the chaper-one genes was increased in the absence of ER stress(Fig. 2F). Although bZIP17DC triggered a weaker in-duction for most genes analyzed than did bZIP28DCwhen controlled by their native promoters, bZIP17DCcontrolled by the strong 35S promoter enhanced chap-erone gene expression to a similar amount as that in-duced by bZIP28DC under its native promoter (Fig.2F). Consistent with the analysis of transcripts, accu-mulation of chaperone proteins was lower in s2p, butenhanced by bZIPDCs in the absence of TM (Fig.2G). These data support a role for these bZIPs in reg-ulating the ER stress response and suggest that in-creased basal ER chaperones may be a mechanismfor TM tolerance of s2p plants expressing these con-stitutively active forms of the proteins. These observa-tions are consistent with the model in which ER stressleads to S2P-dependent proteolysis of bZIP proteins(hereafter referred to as S2P-RIP), which in turn acti-vates the production of ER chaperones.
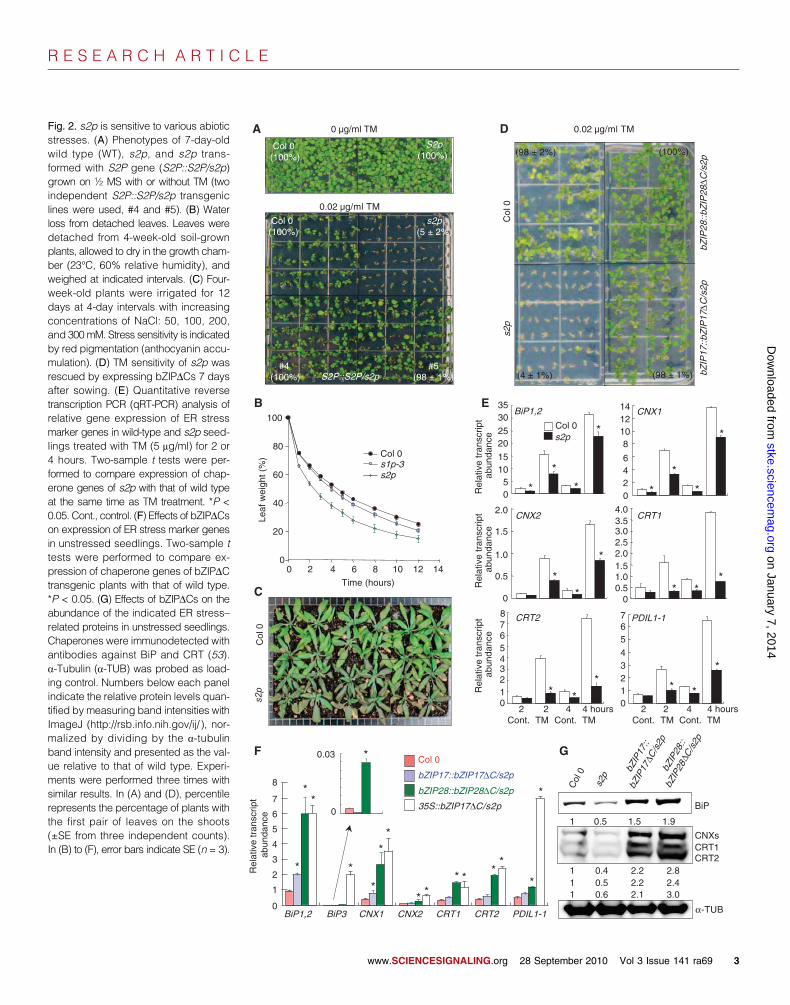
S2P-RIP stimulates BR responsesAlthough shoot growth appeared normal (Fig. 2A),the s2p mutant exhibited impaired root growth (70%shorter than that of wild type and more branched thanthat of wild type) when grown on the surface of agarmedium (Fig. 3A). When grown in well-watered soil,again s2p root growth was impaired, whereas shootsappeared the same as those of the wild type (Fig. 3, Band C). Root growth was restored when s2p was trans-formed with the S2P genomic DNA (Fig. 3A). The s1pmutants (22) also had short roots (Fig. 3, D and E).Consistent with bZIP17 and bZIP28 mediating responsesof S2P-RIP, expression of bZIP17DC or bZIP28DC ins2p restored root growth (Fig. 3A).
H
Prim
ary
root
leng
th (
mm
)
10
0
20
30
40
50
0.60 0.2 0.4[BL] nM
Col 0
bzip17
bzip28
AbZ
IP17
::
bZIP
17C/s2
p
bZIP
28::
bZIP
28C/s
2p
s2p
Col 0
S2P::S
2P/s2
p
G
0
50
100
150
200
Prim
ary
root
leng
th(%
of c
ontr
ol)
0.5 1
Col 0
s2p
bZIP17::bZIP17 C/s2p
bZIP28::bZIP28 C/s2p
[BL] nM
F
Col 0 s2p
0 0.5 0 0.5[BL] nM
s2pCol 0B
s2pCol 0C
Col 0 s1p-2s1p-3D
E
Prim
ary
root
leng
th (
mm
)
0 1 2 3 4 5 6 7 80
10
20
30
40
50
Time (days)
Col 0s1p-3
s2ps1p-2
* *
**
**
∆ ∆
∆
∆
Fig. 3. Roles of S2P-RIP in plant development and BR response. (A) Complementation of
root development of s2p with S2P genomic DNA or bZIPDC constructs. (B) Shoot pheno-types of 4-week-old wild type and s2p grown in soil under constant light. (C) Root phenotypesof 4-week-old wild type and s2p grown in soil under constant light. (D) Root phenotypes of1-week-old wild type, s1p-2, and s1p-3 mutants grown vertically on ½ MS plates. (E) Wildtype (Col 0), s1p-2, s1p-3, and s2p mutants were grown vertically on ½ MS plates; rootlengths were measured at intervals after germination. (F) Effect of 0.5 nM BL on wild-type(Col 0) and s2p root growth. (G) Root growth of seedlings after 5 days on 0.5 or 1 nM BLrelative to control without BL. (H) Root growth of seedlings after 7 days on different concen-trations of BL, comparing wild type with bzip17 (SALK_104326) and bzip28 (SALK_132285C)mutants. Two-sample t tests were performed to compare root elongation of bzip17 and bzip28to that of wild type at the same concentration of BL. *P < 0.01 (t test). In (E), (G), and (H), errorbars indicate SE (n = 30).www.SCIENCESIGNALING.org 28 September 2010 Vol 3 Issue 141 ra69 4
R E S E A R C H A R T I C L E
BR signaling activates genes involved in cell wall loosening contribut-ing to increased cell expansion (34–36), which is a key aspect of root andshoot development. Disruption of BR signaling causes short root anddwarf shoot phenotypes (4). To better understand the function of S2P inplant development, we tested the sensitivity of root elongation to exoge-nous BR or in plants with mutations that compromise BR signaling. Inwild-type roots, low concentrations of brassinolide (BL) stimulate growth,but at higher concentrations, BL inhibits root growth. To distinguish be-tween wild-type and s2p root response, we used a high concentration ofBL that repressed wild-type root growth. Whereas wild-type root growthwas repressed by a superoptimal concentration of exogenous BL, s2p rootsgrew longer in response to this concentration of BL (Fig. 3, F and G).Knockout mutants bzip28 and bzip17 (22, 27, 31) both showed reducedinhibition of root growth by superoptimal BL (Fig. 3H). Expression of
www.S
bZIP17DC or bZIP28DC in s2p restored the inhibition of root growthmediated by BL to that of wild type (Fig. 3G).
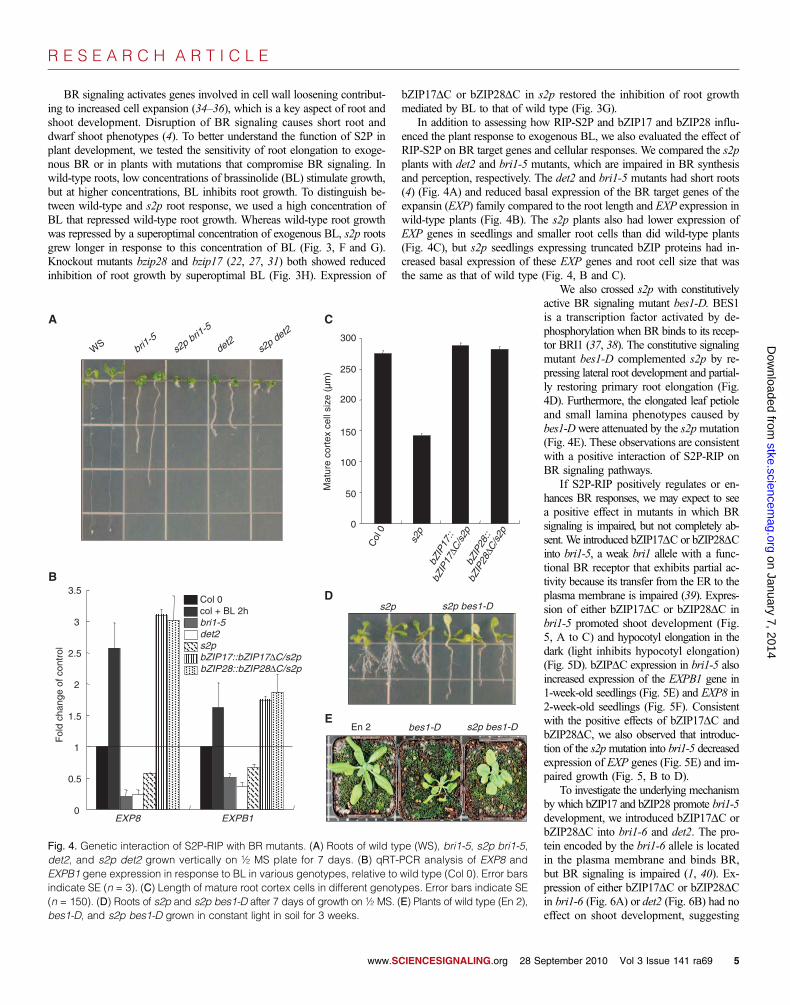
In addition to assessing how RIP-S2P and bZIP17 and bZIP28 influ-enced the plant response to exogenous BL, we also evaluated the effect ofRIP-S2P on BR target genes and cellular responses. We compared the s2pplants with det2 and bri1-5 mutants, which are impaired in BR synthesisand perception, respectively. The det2 and bri1-5 mutants had short roots(4) (Fig. 4A) and reduced basal expression of the BR target genes of theexpansin (EXP) family compared to the root length and EXP expression inwild-type plants (Fig. 4B). The s2p plants also had lower expression ofEXP genes in seedlings and smaller root cells than did wild-type plants(Fig. 4C), but s2p seedlings expressing truncated bZIP proteins had in-creased basal expression of these EXP genes and root cell size that wasthe same as that of wild type (Fig. 4, B and C).
CIENCESIGNALING.org 2
on January 7, 2014 stke.sciencem
ag.orgD
ownloaded from
We also crossed s2p with constitutivelyactive BR signaling mutant bes1-D. BES1is a transcription factor activated by de-phosphorylation when BR binds to its recep-tor BRI1 (37, 38). The constitutive signalingmutant bes1-D complemented s2p by re-pressing lateral root development and partial-ly restoring primary root elongation (Fig.4D). Furthermore, the elongated leaf petioleand small lamina phenotypes caused bybes1-Dwere attenuated by the s2pmutation(Fig. 4E). These observations are consistentwith a positive interaction of S2P-RIP onBR signaling pathways.
If S2P-RIP positively regulates or en-hances BR responses, we may expect to seea positive effect in mutants in which BRsignaling is impaired, but not completely ab-sent. We introduced bZIP17DC or bZIP28DCinto bri1-5, a weak bri1 allele with a func-tional BR receptor that exhibits partial ac-tivity because its transfer from the ER to theplasma membrane is impaired (39). Expres-sion of either bZIP17DC or bZIP28DC inbri1-5 promoted shoot development (Fig.5, A to C) and hypocotyl elongation in thedark (light inhibits hypocotyl elongation)(Fig. 5D). bZIPDC expression in bri1-5 alsoincreased expression of the EXPB1 gene in1-week-old seedlings (Fig. 5E) and EXP8 in2-week-old seedlings (Fig. 5F). Consistentwith the positive effects of bZIP17DC andbZIP28DC, we also observed that introduc-tion of the s2pmutation into bri1-5 decreasedexpression of EXP genes (Fig. 5E) and im-paired growth (Fig. 5, B to D).
To investigate the underlying mechanismby which bZIP17 and bZIP28 promote bri1-5development, we introduced bZIP17DC orbZIP28DC into bri1-6 and det2. The pro-tein encoded by the bri1-6 allele is locatedin the plasma membrane and binds BR,but BR signaling is impaired (1, 40). Ex-pression of either bZIP17DC or bZIP28DCin bri1-6 (Fig. 6A) or det2 (Fig. 6B) had noeffect on shoot development, suggesting
,
,
bZIP28
::
bZIP28
C/s2p
CM
atur
e co
rtex
cel
l siz
e (µ
m)
0
50
100
150
200
250
300
Col
0
s2p
bZIP17
::
bZIP17
C/s2p
B
0
0.5
1
1.5
2
2.5
3
3.5
Fol
d ch
ange
of c
ontr
ol
EXP8 EXPB1
Col 0col + BL 2hbri1-5det2s2pbZIP17::bZIP17 C/s2pbZIP28::bZIP28 C/s2p
s2p bri1
-5
bri1-5
det2s2
p det2A
WS
Es2p bes1-Dbes1-DEn 2
s2p bes1-Ds2pD
∆ ∆
∆∆
Fig. 4. Genetic interaction of S2P-RIP with BR mutants. (A) Roots of wild type (WS), bri1-5, s2p bri1-5det2, and s2p det2 grown vertically on ½ MS plate for 7 days. (B) qRT-PCR analysis of EXP8 andEXPB1 gene expression in response to BL in various genotypes, relative to wild type (Col 0). Error barsindicate SE (n = 3). (C) Length of mature root cortex cells in different genotypes. Error bars indicate SE(n = 150). (D) Roots of s2p and s2p bes1-D after 7 days of growth on ½ MS. (E) Plants of wild type (En 2)bes1-D, and s2p bes1-D grown in constant light in soil for 3 weeks.
8 September 2010 Vol 3 Issue 141 ra69 5
R E S E A R C H A R T I C L E
www.SCIENCESIGNALING.org 28 Se
on January 7, 2014 stke.sciencem
ag.orgD
ownloaded from
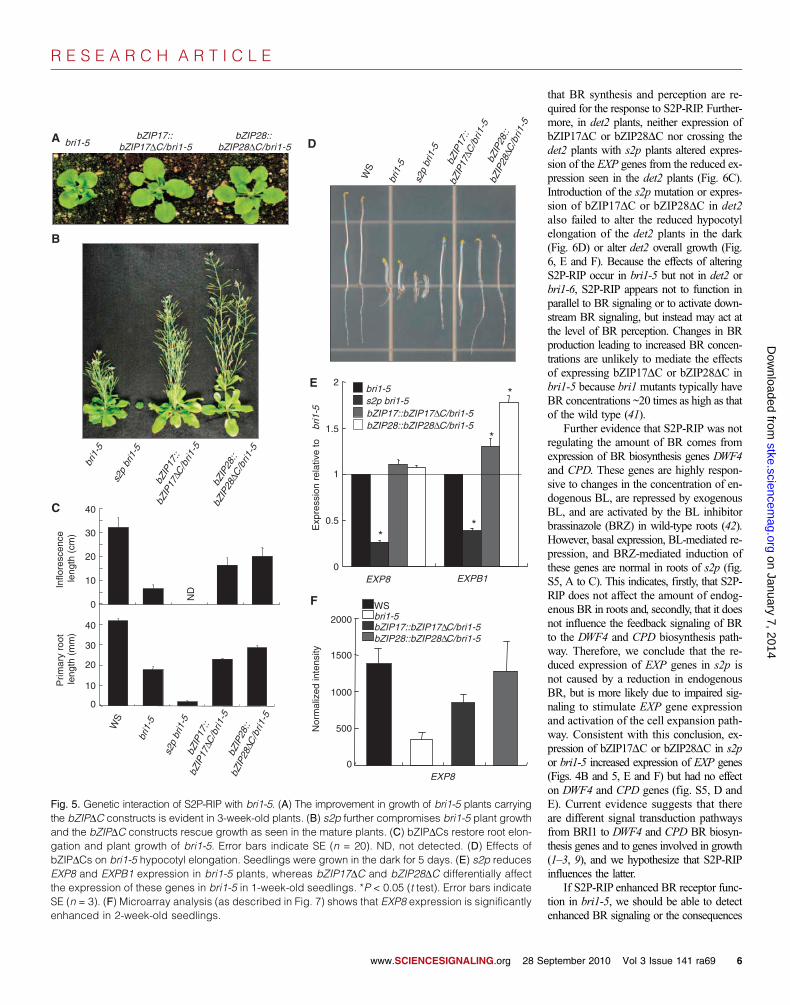
that BR synthesis and perception are re-quired for the response to S2P-RIP. Further-more, in det2 plants, neither expression ofbZIP17DC or bZIP28DC nor crossing thedet2 plants with s2p plants altered expres-sion of the EXP genes from the reduced ex-pression seen in the det2 plants (Fig. 6C).Introduction of the s2p mutation or expres-sion of bZIP17DC or bZIP28DC in det2also failed to alter the reduced hypocotylelongation of the det2 plants in the dark(Fig. 6D) or alter det2 overall growth (Fig.6, E and F). Because the effects of alteringS2P-RIP occur in bri1-5 but not in det2 orbri1-6, S2P-RIP appears not to function inparallel to BR signaling or to activate down-stream BR signaling, but instead may act atthe level of BR perception. Changes in BRproduction leading to increased BR concen-trations are unlikely to mediate the effectsof expressing bZIP17DC or bZIP28DC inbri1-5 because bri1 mutants typically haveBR concentrations ~20 times as high as thatof the wild type (41).
Further evidence that S2P-RIP was notregulating the amount of BR comes fromexpression of BR biosynthesis genes DWF4and CPD. These genes are highly respon-sive to changes in the concentration of en-dogenous BL, are repressed by exogenousBL, and are activated by the BL inhibitorbrassinazole (BRZ) in wild-type roots (42).However, basal expression, BL-mediated re-pression, and BRZ-mediated induction ofthese genes are normal in roots of s2p (fig.S5, A to C). This indicates, firstly, that S2P-RIP does not affect the amount of endog-enous BR in roots and, secondly, that it doesnot influence the feedback signaling of BRto the DWF4 and CPD biosynthesis path-way. Therefore, we conclude that the re-duced expression of EXP genes in s2p isnot caused by a reduction in endogenousBR, but is more likely due to impaired sig-naling to stimulate EXP gene expressionand activation of the cell expansion path-way. Consistent with this conclusion, ex-pression of bZIP17DC or bZIP28DC in s2por bri1-5 increased expression of EXP genes(Figs. 4B and 5, E and F) but had no effecton DWF4 and CPD genes (fig. S5, D andE). Current evidence suggests that thereare different signal transduction pathwaysfrom BRI1 to DWF4 and CPD BR biosyn-thesis genes and to genes involved in growth(1–3, 9), and we hypothesize that S2P-RIPinfluences the latter.
If S2P-RIP enhanced BR receptor func-tion in bri1-5, we should be able to detectenhanced BR signaling or the consequences
bri1-5A bZIP28::bZIP28 C/bri1-5
bZIP17::bZIP17 C/bri1-5
bZIP
17::
bZIP
17 C
/bri1
-5
bri1
-5
s2p
bri1
-5
bZIP
28::
bZIP
28 C
/bri1
-5
C
Inflo
resc
ence
le
ngth
(cm
)
40
WS
10
0
20
30
ND
Prim
ary
root
le
ngth
(m
m)
0
10
20
30
40
F WSbri1-5bZIP17::bZIP17 C/bri1-5bZIP28::bZIP28 C/bri1-5
Nor
mal
ized
inte
nsity
0
500
1000
1500
EXP8
2000
B
bri1-5
s2p bri1-5
bZIP28
::
bZIP28
C/bri1
-5
bZIP17
::
bZIP17
C/bri1
-5bZ
IP17
::bZ
IP17
C/b
ri1-5
bZIP
28::
bZIP
28 C
/bri1
-5
bri1
-5
WS
s2p
bri1
-5D
E
EXP8 EXPB10
0.5
1
1.5
2
Exp
ress
ion
rela
tive
to
bri1
-5
bri1-5s2p bri1-5bZIP17::bZIP17 C/bri1-5bZIP28::bZIP28 C/bri1-5
*
*
**
∆∆
∆ ∆∆ ∆
∆∆
∆∆
∆∆
Fig. 5. Genetic interaction of S2P-RIP with bri1-5. (A) The improvement in growth of bri1-5 plants carryingthe bZIPDC constructs is evident in 3-week-old plants. (B) s2p further compromises bri1-5 plant growthand the bZIPDC constructs rescue growth as seen in the mature plants. (C) bZIPDCs restore root elon-gation and plant growth of bri1-5. Error bars indicate SE (n = 20). ND, not detected. (D) Effects ofbZIPDCs on bri1-5 hypocotyl elongation. Seedlings were grown in the dark for 5 days. (E) s2p reducesEXP8 and EXPB1 expression in bri1-5 plants, whereas bZIP17DC and bZIP28DC differentially affectthe expression of these genes in bri1-5 in 1-week-old seedlings. *P < 0.05 (t test). Error bars indicateSE (n = 3). (F) Microarray analysis (as described in Fig. 7) shows that EXP8 expression is significantlyenhanced in 2-week-old seedlings.
ptember 2010 Vol 3 Issue 141 ra69 6
R E S E A R C H A R T I C L E
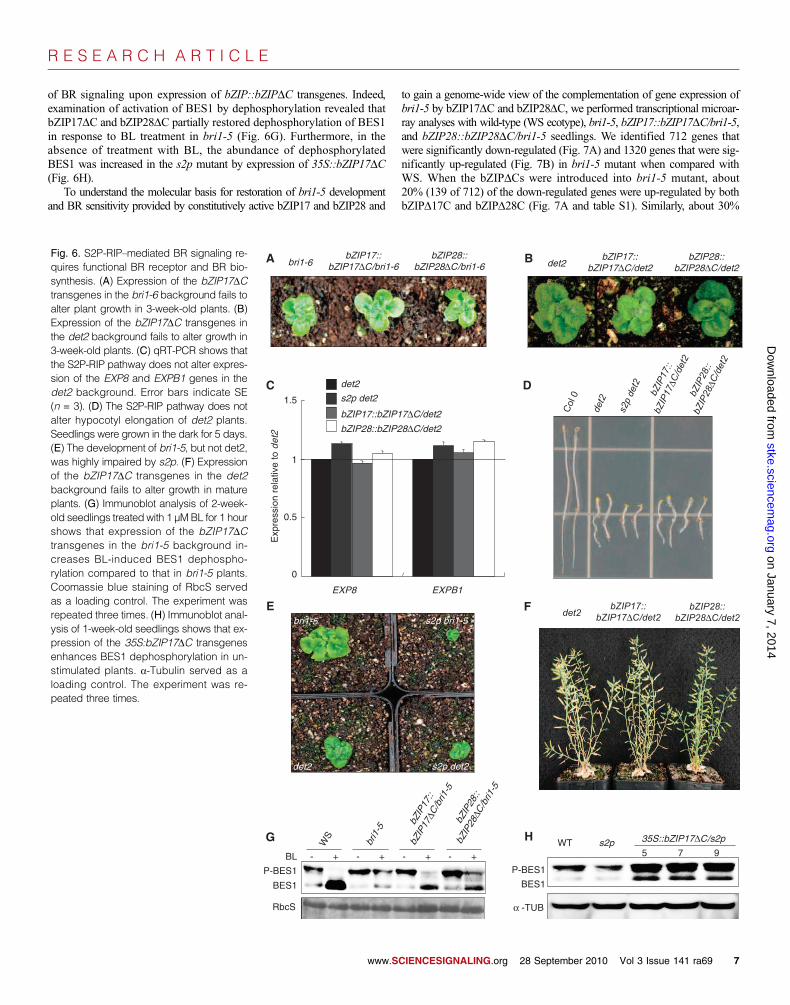
of BR signaling upon expression of bZIP::bZIPDC transgenes. Indeed,examination of activation of BES1 by dephosphorylation revealed thatbZIP17DC and bZIP28DC partially restored dephosphorylation of BES1in response to BL treatment in bri1-5 (Fig. 6G). Furthermore, in theabsence of treatment with BL, the abundance of dephosphorylatedBES1 was increased in the s2p mutant by expression of 35S::bZIP17DC(Fig. 6H).
To understand the molecular basis for restoration of bri1-5 developmentand BR sensitivity provided by constitutively active bZIP17 and bZIP28 and
EXP8
C
0
0.5
1
1.5
Exp
ress
ion
rela
tive
to d
et2
det2
s2p det2
bZIP17::bZIP1
bZIP28::bZIP2
G
P-BES1
BES1
BL - + - +
WS
bri1-5
RbcS
E
det2
bri1-5
www.S
to gain a genome-wide view of the complementation of gene expression ofbri1-5 by bZIP17DC and bZIP28DC, we performed transcriptional microar-ray analyses with wild-type (WS ecotype), bri1-5, bZIP17::bZIP17DC/bri1-5,and bZIP28::bZIP28DC/bri1-5 seedlings. We identified 712 genes thatwere significantly down-regulated (Fig. 7A) and 1320 genes that were sig-nificantly up-regulated (Fig. 7B) in bri1-5 mutant when compared withWS. When the bZIPDCs were introduced into bri1-5 mutant, about20% (139 of 712) of the down-regulated genes were up-regulated by bothbZIPD17C and bZIPD28C (Fig. 7A and table S1). Similarly, about 30%
EXPB1
7 C/det2
8 C/det2
det2
bZIP
17::
bZIP
17∆C
/det2
Col 0
s2p d
et2
bZIP
28::
bZIP
28∆C
/det2
D
F det2
bZIP28::
bZIP28∆C/det2
bZIP17::
bZIP17∆C/det2s2p bri1-5
∆∆
on January 7, 2014 stke.sciencem
ag.orgD
ownloaded from
Fig. 6. S2P-RIP–mediated BR signaling re-quires functional BR receptor and BR bio-synthesis. (A) Expression of the bZIP17DCtransgenes in the bri1-6 background fails toalter plant growth in 3-week-old plants. (B)Expression of the bZIP17DC transgenes inthe det2 background fails to alter growth in3-week-old plants. (C) qRT-PCR shows thatthe S2P-RIP pathway does not alter expres-sion of the EXP8 and EXPB1 genes in thedet2 background. Error bars indicate SE(n = 3). (D) The S2P-RIP pathway does notalter hypocotyl elongation of det2 plants.Seedlings were grown in the dark for 5 days.(E) The development of bri1-5, but not det2,was highly impaired by s2p. (F) Expressionof the bZIP17DC transgenes in the det2background fails to alter growth in matureplants. (G) Immunoblot analysis of 2-week-old seedlings treated with 1 µMBL for 1 hourshows that expression of the bZIP17DCtransgenes in the bri1-5 background in-creases BL-induced BES1 dephospho-rylation compared to that in bri1-5 plants.Coomassie blue staining of RbcS servedas a loading control. The experiment wasrepeated three times. (H) Immunoblot anal-ysis of 1-week-old seedlings shows that ex-pression of the 35S:bZIP17DC transgenesenhances BES1 dephosphorylation in un-stimulated plants. a-Tubulin served as aloading control. The experiment was re-peated three times.
det2bZIP28::
bZIP28 C/det2bZIP17::
bZIP17 C/det2BA bri1-6
bZIP17::bZIP17 C/bri1-6
bZIP28::bZIP28 C/bri1-6 ∆ ∆∆∆
bZIP28
::
bZIP28
C/bri1
-5
bZIP17
::
bZIP17
C/bri1
-5
- + - +
WT s2p 35S::bZIP17 C/s2p
5 7 9
P-BES1
BES1
H
s2p det2
∆
∆∆
α -TUB
CIENCESIGNALING.org 28 September 2010 Vol 3 Issue 141 ra69 7
R E S E A R C H A R T I C L E
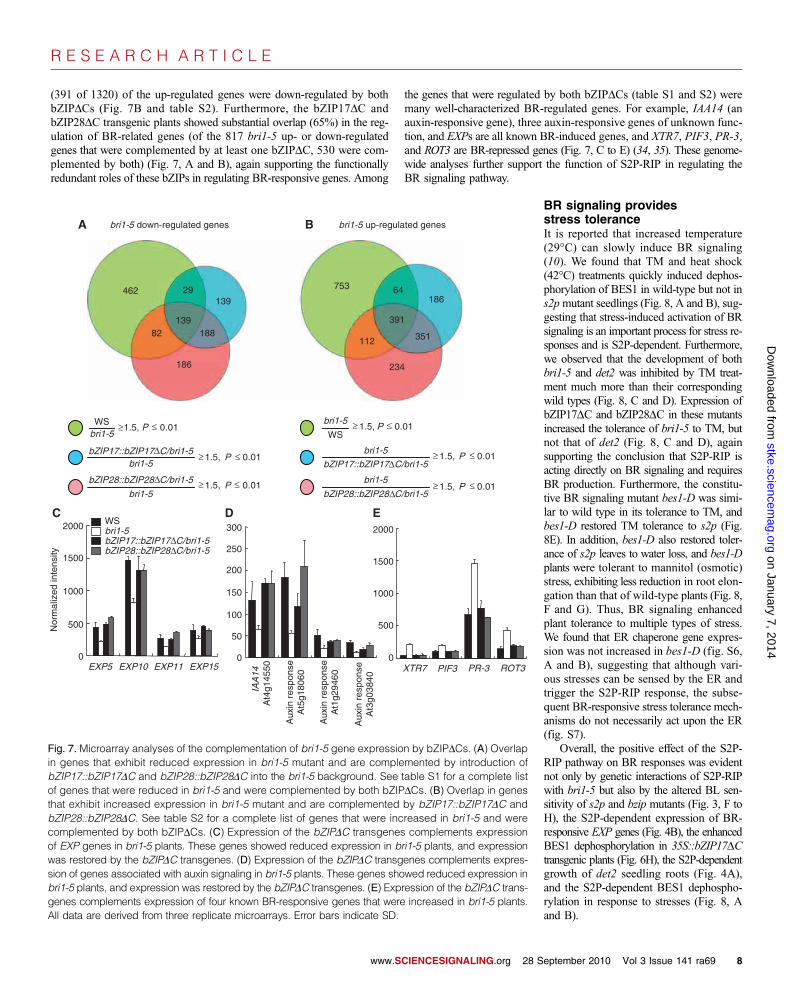
(391 of 1320) of the up-regulated genes were down-regulated by bothbZIPDCs (Fig. 7B and table S2). Furthermore, the bZIP17DC andbZIP28DC transgenic plants showed substantial overlap (65%) in the reg-ulation of BR-related genes (of the 817 bri1-5 up- or down-regulatedgenes that were complemented by at least one bZIPDC, 530 were com-plemented by both) (Fig. 7, A and B), again supporting the functionallyredundant roles of these bZIPs in regulating BR-responsive genes. Among
www.S
the genes that were regulated by both bZIPDCs (table S1 and S2) weremany well-characterized BR-regulated genes. For example, IAA14 (anauxin-responsive gene), three auxin-responsive genes of unknown func-tion, and EXPs are all known BR-induced genes, and XTR7, PIF3, PR-3,and ROT3 are BR-repressed genes (Fig. 7, C to E) (34, 35). These genome-wide analyses further support the function of S2P-RIP in regulating theBR signaling pathway.
CIENCESIGNALING.org 28 S
on January 7, 2014 stke.sciencem
ag.orgD
ownloaded from
BR signaling providesstress toleranceIt is reported that increased temperature(29°C) can slowly induce BR signaling(10). We found that TM and heat shock(42°C) treatments quickly induced dephos-phorylation of BES1 in wild-type but not ins2pmutant seedlings (Fig. 8, A and B), sug-gesting that stress-induced activation of BRsignaling is an important process for stress re-sponses and is S2P-dependent. Furthermore,we observed that the development of bothbri1-5 and det2 was inhibited by TM treat-ment much more than their correspondingwild types (Fig. 8, C and D). Expression ofbZIP17DC and bZIP28DC in these mutantsincreased the tolerance of bri1-5 to TM, butnot that of det2 (Fig. 8, C and D), againsupporting the conclusion that S2P-RIP isacting directly on BR signaling and requiresBR production. Furthermore, the constitu-tive BR signaling mutant bes1-D was simi-lar to wild type in its tolerance to TM, andbes1-D restored TM tolerance to s2p (Fig.8E). In addition, bes1-D also restored toler-ance of s2p leaves to water loss, and bes1-Dplants were tolerant to mannitol (osmotic)stress, exhibiting less reduction in root elon-gation than that of wild-type plants (Fig. 8,F and G). Thus, BR signaling enhancedplant tolerance to multiple types of stress.We found that ER chaperone gene expres-sion was not increased in bes1-D (fig. S6,A and B), suggesting that although vari-ous stresses can be sensed by the ER andtrigger the S2P-RIP response, the subse-quent BR-responsive stress tolerance mech-anisms do not necessarily act upon the ER(fig. S7).
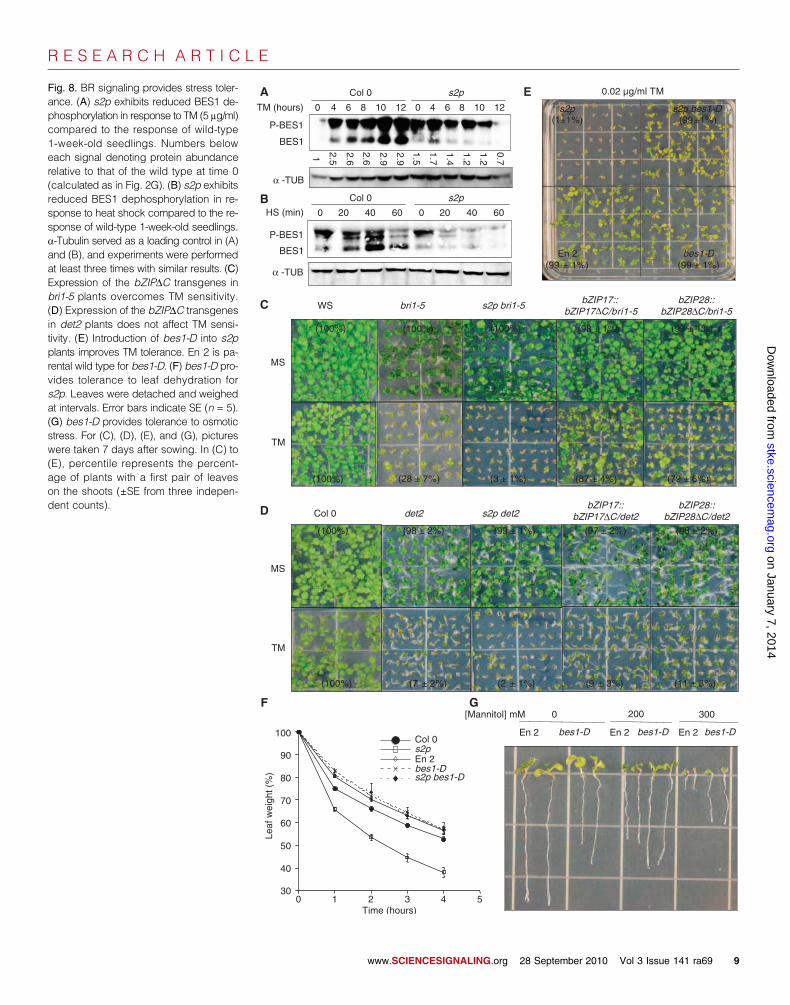
Overall, the positive effect of the S2P-RIP pathway on BR responses was evidentnot only by genetic interactions of S2P-RIPwith bri1-5 but also by the altered BL sen-sitivity of s2p and bzip mutants (Fig. 3, F toH), the S2P-dependent expression of BR-responsive EXP genes (Fig. 4B), the enhancedBES1 dephosphorylation in 35S::bZIP17DCtransgenic plants (Fig. 6H), the S2P-dependentgrowth of det2 seedling roots (Fig. 4A),and the S2P-dependent BES1 dephospho-rylation in response to stresses (Fig. 8, Aand B).
0
50
100
150
200
250
300
IA
A14
At4
g145
50
Aux
in r
espo
nse
At5
g180
60
Aux
in r
espo
nse
At3
g038
40
DCWSbri1-5bZIP17::bZIP17 C/bri1-5bZIP28::bZIP28 C/bri1-5
Nor
mal
ized
inte
nsity
0
500
1000
1500
2000
EXP10EXP5 EXP11 EXP15
Aux
in r
espo
nse
At1
g294
60
0
500
1000
1500
2000
XTR7 PIF3 PR-3 ROT3
E
A B
WSbri1-5
1.5, P 0.01
bZIP17::bZIP17 C/bri1-5bri1-5
1.5, P 0.01
bri1-5
WS1.5, P 0.01
bri1-5 down-regulated genes bri1-5 up-regulated genes
462 29139
13982 188
186
753 64186
391
112 351
234
bZIP28::bZIP28 C/bri1-5
bri1-51.5, P 0.01
bri1-5bZIP17::bZIP17 C/bri1-5
1.5, P 0.01
bri1-5
bZIP28::bZIP28 C/bri1-51.5, P 0.01
≥
≥ ≥
≥
≥
≥
≥ ≥
≥
≥≥
∆
∆
∆∆
∆
∆
≥
Fig. 7. Microarray analyses of the complementation of bri1-5 gene expression by bZIPDCs. (A) Overlapin genes that exhibit reduced expression in bri1-5 mutant and are complemented by introduction ofbZIP17::bZIP17DC and bZIP28::bZIP28DC into the bri1-5 background. See table S1 for a complete listof genes that were reduced in bri1-5 and were complemented by both bZIPDCs. (B) Overlap in genesthat exhibit increased expression in bri1-5 mutant and are complemented by bZIP17::bZIP17DC andbZIP28::bZIP28DC. See table S2 for a complete list of genes that were increased in bri1-5 and werecomplemented by both bZIPDCs. (C) Expression of the bZIPDC transgenes complements expressionof EXP genes in bri1-5 plants. These genes showed reduced expression in bri1-5 plants, and expressionwas restored by the bZIPDC transgenes. (D) Expression of the bZIPDC transgenes complements expres-sion of genes associated with auxin signaling in bri1-5 plants. These genes showed reduced expression inbri1-5 plants, and expression was restored by the bZIPDC transgenes. (E) Expression of the bZIPDC trans-genes complements expression of four known BR-responsive genes that were increased in bri1-5 plants.All data are derived from three replicate microarrays. Error bars indicate SD.
eptember 2010 Vol 3 Issue 141 ra69 8
C
MS
bri1-5WSbZIP17::
bZIP17 C/bri1-5s2p bri1-5 bZIP28::
bZIP28 C/bri1-5
D bZIP17:: bZIP28::
s2p(1±1%)
s2p bes1-D
bes1-D(99
En 2
B Col 0 s2p
HS (min) 0 20 40 60 0 20 40 60
P-BES1
BES1
P-BES1
TM (hours) 0 4 6 8 0
BES1
10 12 4 6 8 10 12
α -TUB
(100%)
(100%)
(100%) (100%)
1 2.5
2.6
2.6
2.9
2.9
1.5
1.7
1.4
1.2
1.2
0.7
TM
∆ ∆
± 1%)(99 ± 1%)
(99±1%)
α -TUB
(28 ± 7%) (3 ± 1%) (87 ± 4%) (79 ± 6%)
(98 ± 1%) (99 ± 1%)
R E S E A R C H A R T I C L E
stke.sciencemD
ownloaded from
Fig. 8. BR signaling provides stress toler-ance. (A) s2p exhibits reduced BES1 de-phosphorylation in response to TM (5 mg/ml)compared to the response of wild-type1-week-old seedlings. Numbers beloweach signal denoting protein abundancerelative to that of the wild type at time 0(calculated as in Fig. 2G). (B) s2p exhibitsreduced BES1 dephosphorylation in re-sponse to heat shock compared to the re-sponse of wild-type 1-week-old seedlings.a-Tubulin served as a loading control in (A)and (B), and experiments were performedat least three times with similar results. (C)Expression of the bZIPDC transgenes inbri1-5 plants overcomes TM sensitivity.(D) Expression of the bZIPDC transgenesin det2 plants does not affect TM sensi-tivity. (E) Introduction of bes1-D into s2pplants improves TM tolerance. En 2 is pa-rental wild type for bes1-D. (F) bes1-D pro-vides tolerance to leaf dehydration fors2p. Leaves were detached and weighedat intervals. Error bars indicate SE (n = 5).(G) bes1-D provides tolerance to osmoticstress. For (C), (D), (E), and (G), pictureswere taken 7 days after sowing. In (C) to(E), percentile represents the percent-age of plants with a first pair of leaveson the shoots (±SE from three indepen-dent counts).
E 0.02 µg/ml TMA Col 0 s2p
Col 0 det2 s2p det2 bZIP17 C/det2 bZIP28 C/det2
MS
Time (hours)0 1 2 3 4 5
90
40
30
50
60
70
80
Leaf
wei
ght (
%)
Col 0s2pEn 2bes1-Ds2p bes1-D
F
En 2 bes1-D En 2 bes1-D bes1-D
[Mannitol] mM 0 200 300
En 2
G
100
(100%)
(100%)
TM
∆ ∆(98 ± 2%) (99 ± 1%) (97 ± 2%) (98 ± 2%)
(7 ± 2%) (2 ± 1%) (9 ± 3%) (11 ± 3%)
www.SCIENCESIGNALING.org 28 September 2010 Vol 3 Issue 141 ra69 9
on January 7, 2014 ag.org

R E S E A R C H A R T I C L E
on January 7, 2014 stke.sciencem
ag.orgD
ownloaded from
DISCUSSION
BRI1 is synthesized in the ER and then translocated through the Golgi tothe plasma membrane (43). One way in which BR signaling might bestimulated under stressful conditions is by S2P-RIP–dependent ER chap-erone synthesis, which could enhance the correct folding or modificationof BRI1 and promote its translocation from the endomembrane to the plas-ma membrane (3, 39, 40, 43–45). Naturally occurring abiotic stresses, suchas heat shock, salinity, or osmotic stress, may impair BRI1 maturation andtranslocation; in the laboratory, agents such as TM or mutations in BRI (forexample, bri1-5) may impair BRI1 maturation and translocation. We pro-pose that S2P-RIP can overcome such constraints, enhance BR signaling,and induce acclimation to stress by promoting BRI1 delivery to the plasmamembrane and potentially by directly enhancing regulation of some BR tar-get genes (fig. S7). We believe it will be important in the future to in-vestigate the effect of the S2P-RIP pathway on BRI1 synthesis, assembly,and translocation, and its effects on the interaction of BRI1 with othermembrane-associated proteins, such as BAK1 and BSKs (46–48), which po-tentially provide specificity to BRI1.
We considered the possibility that bZIP17DC and bZIP28DC mightinteract directly with transcription factor BES1 in the nucleus, but ob-tained no evidence for interaction either in yeast two-hybrid experimentsor with bimolecular fluorescence (BiFC) in planta. Elucidating details of themolecular interactions linking ER signaling to BR signaling will be an areafor future research.
Previous studies indicated that bZIP28 is involved in TM stress (27),whereas bZIP17 is involved primarily in salt stress (22, 27). We have con-firmed and extended these observations to show that both are involved inER stress and heat stress. These proteins may have overlapping sensitivityto different stresses and partially redundant functions, providing flexibilityin perception and response to multiple environmental challenges.
The activation of bZIPs in response to stress, as well as their ability toinduce BR signaling (fig. S7), is unexpected because BR is typicallyviewed as a positive regulator of plant growth, and yet plant growth isoften impaired by abiotic stresses, such as heat shock or salt stress. How-ever, in natural environments, plants are constantly subjected to stresses,including extreme fluctuations in temperature and water supply. Differ-ential activation of growth processes under stressful conditions may berequired to optimize resource allocation or plant development during ac-climation. As examples, moderate water stress can stimulate primary rootelongation and the rate of cell production (49), and high temperature canstimulate hypocotyl elongation (10). Integration of stress responses andgrowth processes through a common BR signaling pathway may providethe means to achieve optimal growth under challenging environmentalconditions.
MATERIALS AND METHODS
Plant materials and growth conditionsThe det2 mutant is in Arabidopsis thaliana Columbia ecotype, bes1-Dand bri1-6 are in Enkheim-2 (En 2), and bri1-5 is in the WS ecotype.The s2p homozygous mutant (NASC N444004) was identified by PCRwith gene-specific and left border (LB) T-DNA primers (table S3). s2pbri1-5, s2p det2, and s2p bes1-D double mutants were generated bycrossing single mutants and allowing the F1 to self-fertilize, and thencandidate double F2 plants were genotyped through a combination of phe-notype and PCR genotype examination. All seeds were surface-sterilizedand placed on half-strength Murashige and Skoog (½ MS) medium with1% (w/v) sucrose or in soil under continuous light at 23°C. For root growthinhibition assay, seeds were placed on culture medium containing different
www.SC
concentrations of BL and then grown vertically. Root length was measuredon the fifth or seventh day after germination. For TM sensitivity assay,seeds were placed on culture containing TM (0.02 mg/ml) and then grownhorizontally for 7 days. The inhibition of formation of the first pair oftrue leaves in seedlings was the measure for sensitivity. For transientBL, BRZ, or TM treatments, 7-day-old seedlings or roots were transferredinto liquid MS culture containing either mock solution, 5 nM BL, 3 mMBRZ, or TM (5 mg/ml) for different times as indicated.
Plasmid constructsThe coding region of S2P with its own promoter was amplified withgenomic DNA template and cloned into pCAMBIA3300 vector andnamed as S2P::S2P. For bZIP17::bZIP17DC and bZIP28::bZIP28DCconstructs, the truncated coding regions of these bZIPs with their own pro-moters were amplified from genomic DNA template and cloned intopCAMBIA3300 vector (see http://www.cambia.org/daisy/cambia/home.html). pGREEN180-GFP was constructed by introduction of CAMV 2 ×35S promoter and CAMV terminator into pGREEN179 (50) and insertionof enhanced GFP into either the Bam HI–Xba I sites (which createspGREEN180-GFPC for C-terminal GFP tagging) or the Eco RI–BamHI sites (which creates pGREEN180-GFPN for N-terminal GFP tagging).N-terminal GFP fusions of bZIPs were constructed by inserting openreading frames of bZIPs into pGREEN180-GFPN. C-terminal fusions ofGFP to S1P and S2P were made in pGREEN180-GFPC. Biolistic bombard-ment into onion epidermal cells or Arabidopsis roots was performed asdescribed previously (51). The plasmid G-RB, expressing mCherry targetedto Golgi (52), was used as a Golgi localization control. For stable transgeniclines, the resulting constructs were introduced into Agrobacterium tume-faciens strain C58, which was used to generate transgenic plants by thefloral dip method. Primer sequences for these constructs are provided intable S4.
Protein extraction and Western blot analysisProteins were extracted with 2 × SDS sample buffer, boiled for 5 min,and centrifuged for 10 min at 10,000g. The resulting supernatants weretransferred into a new microfuge tube. SDS–polyacrylamide gel electro-phoresis (SDS-PAGE) was performed in 10% (w/v) polyacrylamide gels.After electrophoresis, proteins were electrophoretically transferred to a ni-trocellulose membrane (Amersham) and immunodetected with antibodiesagainst GFP (Invitrogen), immunoglobulin heavy-chain–binding protein(BiP; Santa Cruz Biotechnology), calreticulin (CRT) (53), BES1 (38),or a-tubulin (Sigma).
qPCR and microarray analysisTotal RNAwas isolated with an RNeasy kit, treated with RNase-free DNaseI (Qiagen) according to the manufacturer’s instructions, and quantified by aNanoDrop (ND-1000) spectrophotometer.
For qPCR analysis, 1 mg of total RNAwas reverse-transcribed with theSuperScript III RT kit (Invitrogen). qPCR was performed on a RocheLightCycler 480 qPCR instrument. Transcript abundance was calculatedrelative to the actin-2 gene as described by Czechowski et al. (54). AllqPCR data represent the average of three biological replicate experiments.Primer sequences are provided in table S3.
For microarray analysis, three independent biological replicates foreach sample (2-week-old WS, bri1-5, bZIP17::bZIP17DC/bri1-5, andbZIP28::bZIP28DC/bri1-5 seedlings) were analyzed with Affymetrix Ar-abidopsis gene chips (ATH1). For each microarray, overall intensity nor-malization for the entire probe sets was performed with Avadis 4.3 (StrandLife Sciences Pvt Ltd). Using the t test packaged in Mev 4.0 (http://www.tigr.org/software/tm4/), we estimated a P value (adjusted Bonferroni cor-
IENCESIGNALING.org 28 September 2010 Vol 3 Issue 141 ra69 10

R E S E A R C H A R T I C L E
on January 7, 2014 stke.sciencem
ag.orgD
ownloaded from
rection was applied) with 100 permutations to correct for multiple com-parisons; P ≤ 0.01 was considered significant. An additional, arbitrarycriterion of a 1.5-fold change cutoff was applied to select genes withup- or down-regulation relative to bri1-5.
Microscopy and imagingFor GUS staining, whole seedlings and excised plant organs and tissueswere incubated in 5-bromo-4-chloro-3-indole glucuronide (X-gluc) solu-tion {X-gluc (0.5 mg/ml) in 50 mM tris-NaCl buffer (pH 7.0), 0.5% (v/v)Triton X-100, 0.5 mM K3[Fe(CN)6], 0.5 mM K4[Fe(CN)6], 10 mMNa2EDTA}. X-gluc stock solution (100 mM) was prepared by dissolving26.1 mg of X-gluc in 0.5 ml of dimethyl sulfoxide just before use. Vacuuminfiltration was carried out for 10 min. Tissue was then incubated at 37°Cin the dark for 16 hours or until color developed. To improve the con-trast, we removed soluble pigments by incubating the stained material inseveral changes of 70% (v/v) ethanol until the chlorophyll was clearedfrom the tissue. The stained tissue was examined under bright-field mi-croscopy with an Olympus SZX7 microscope. For fluorescence imaging,5- or 6-day seedlings were subjected to heat shock (42°C for 60 min) orTM (5 mg/ml for 4 hours) treatments. To measure root hair zone cortexcells, we stained 5-day seedlings for at least 10 min with propidium iodide(0.1 mg/ml) to visualize root cell walls. A Leica TCS SP2 AOBS multi-photon confocal microscope was used with laser excitation of GFP at488 nm and red fluorescent protein (RFP) or propidium iodide at 561 nm.Emission collection was in the ranges of 495 to 525 nm and 590 to 700nm, respectively. Confocal images were captured with Leica ConfocalSoftware and manipulated with ImageJ or Photoshop. Root cell lengthmeasurements were made with ImageJ.
SUPPLEMENTARY MATERIALSwww.sciencesignaling.org/cgi/content/full/3/141/ra69/DC1Fig. S1. A diagram of regulated intramembrane proteolysis.Fig. S2. Localization of S1P, S2P, and bZIP28.Fig. S3. Characterization of s2p mutant.Fig. S4. Structures of full-length and truncated bZIPs.Fig. S5. S2P-RIP does not affect BR biosynthesis gene expression.Fig. S6. BES1 does not induce chaperone gene expression in wild-type or s2p backgrounds.Fig. S7. Possible interactions of stress-induced S2P-RIP with BR signaling pathway.Table S1. Complementation of bri1-5 down-regulated genes by bZIPDC.Table S2. Complementation of bri1-5 up-regulated genes by bZIPDCs.Table S3. Oligonucleotides used for PCR, RT-PCR, and qRT-PCR.Table S4. Oligonucleotides used for plasmid constructs.
REFERENCES AND NOTES1. J. Li, H. Jin, Regulation of brassinosteroid signaling. Trends Plant Sci. 12, 37–41
(2007).2. Y. Belkhadir, J. Chory, Brassinosteroid signaling: A paradigm for steroid hormone
signaling from the cell surface. Science 314, 1410–1411 (2006).3. J. M. Gendron, Z. Y. Wang, Multiple mechanisms modulate brassinosteroid signaling.
Curr. Opin. Plant Biol. 10, 436–441 (2007).4. S. D. Clouse, J. M. Sasse, Brassinosteroids: Essential regulators of plant growth and
development. Annu. Rev. Plant Physiol. Plant Mol. Biol. 49, 427–451 (1998).5. A. Bajguz, S. Hayat, Effects of brassinosteroids on the plant responses to environmental
stresses. Plant Physiol. Biochem. 47, 1–8 (2009).6. R. Karlova, S. C. de Vries, Advances in understanding brassinosteroid signaling.
Sci. STKE 2006, pe36 (2006).7. S. Kagale, U. K. Divi, J. E. Krochko, W. A. Keller, P. Krishna, Brassinosteroid confers
tolerance in Arabidopsis thaliana and Brassica napus to a range of abiotic stresses.Planta 225, 353–364 (2007).
8. P. Krishna, Brassinosteroid-mediated stress responses. J. Plant Growth Regul. 22,289–297 (2003).
9. U. K. Divi, P. Krishna, Brassinosteroid: A biotechnological target for enhancing crop yieldand stress tolerance. N. Biotechnol. 26, 131–136 (2009).
10. J. A. Stavang, J. Gallego-Bartolomé, M. D. Gómez, S. Yoshida, T. Asami, J. E. Olsen,J. L. García-Martínez, D. Alabadi, M. A. Blázquez, Hormonal regulation of temperature-induced growth in Arabidopsis. Plant J. 60, 589–601 (2009).
www.SC
11. L. L. Haubrick, S. M. Assmann, Brassinosteroids and plant function: Some clues, morepuzzles. Plant Cell Environ. 29, 446–457 (2006).
12. M. S. Brown, J. Ye, R. B. Rawson, J. L. Goldstein, Regulated intramembrane proteolysis:A control mechanism conserved from bacteria to humans. Cell 100, 391–398 (2000).
13. A. C. Seegmiller, I. Dobrosotskaya, J. L. Goldstein, Y. K. Ho, M. S. Brown, R. B. Rawson,The SREBP pathway in Drosophila: Regulation by palmitate, not sterols. Dev. Cell 2,229–238 (2002).
14. D. Eberlé, B. Hegarty, P. Bossard, P. Ferré, F. Foufelle, SREBP transcription factors:Master regulators of lipid homeostasis. Biochimie 86, 839–848 (2004).
15. M. T. Bengoechea-Alonso, J. Ericsson, SREBP in signal transduction: Cholesterolmetabolism and beyond. Curr. Opin. Cell Biol. 19, 215–222 (2007).
16. P. J. Espenshade, A. L. Hughes, Regulation of sterol synthesis in eukaryotes. Annu.Rev. Genet. 41, 401–427 (2007).
17. A. L. Hughes, B. L. Todd, P. J. Espenshade, SREBP pathway responds to sterols andfunctions as an oxygen sensor in fission yeast. Cell 120, 831–842 (2005).
18. A. L. Hughes, C. Y. Lee, C. M. Bien, P. J. Espenshade, 4-Methyl sterols regulate fissionyeast SREBP-Scap under low oxygen and cell stress. J. Biol. Chem. 282, 24388–24396(2007).
19. S. D. Willger, S. Puttikamonkul, K. H. Kim, J. B. Burritt, N. Grahl, L. J. Metzler, R. Barbuch,M. Bard, C. B. Lawrence, R. A. Cramer Jr., A sterol-regulatory element binding proteinis required for cell polarity, hypoxia adaptation, azole drug resistance, and virulence inAspergillus fumigatus. PLoS Pathog. 4, e1000200 (2008).
20. D. Bailey, P. O’Hare, Transmembrane bZIP transcription factors in ER stress signalingand the unfolded protein response. Antioxid. Redox. Signal. 9, 2305–2321 (2007).
21. K. Kohno, How transmembrane proteins sense endoplasmic reticulum stress. Antioxid.Redox. Signal. 9, 2295–2303 (2007).
22. J. X. Liu, R. Srivastava, P. Che, S. H. Howell, Salt stress responses in Arabidopsis utilizea signal transduction pathway related to endoplasmic reticulum stress signaling. Plant J.51, 897–909 (2007).
23. H. Tajima, Y. Iwata, M. Iwano, S. Takayama, N. Koizumi, Identification of an Arabidopsistransmembrane bZIP transcription factor involved in the endoplasmic reticulum stressresponse. Biochem. Biophys. Res. Commun. 374, 242–247 (2008).
24. Y. N. Chen, E. Slabaugh, F. Brandizzi, Membrane-tethered transcription factors in Ara-bidopsis thaliana: Novel regulators in stress response and development. Curr. Opin.Plant Biol. 11, 695–701 (2008).
25. P. J. Seo, S. G. Kim, C. M. Park, Membrane-bound transcription factors in plants. TrendsPlant Sci. 13, 550–556 (2008).
26. J. S. Kang, J. Frank, C. H. Kang, H. Kajiura, M. Vikram, A. Ueda, S. Kim, J. D. Bahk,B. Triplett, K. Fujiyama, S. Y. Lee, A. von Schaewen, H. Koiwa, Salt tolerance of Ara-bidopsis thaliana requires maturation of N-glycosylated proteins in the Golgi appara-tus. Proc. Natl. Acad. Sci. U.S.A. 105, 5933–5938 (2008).
27. J. X. Liu, R. Srivastava, P. Che, S. H. Howell, An endoplasmic reticulum stress re-sponse in Arabidopsis is mediated by proteolytic processing and nuclear relocationof a membrane-associated transcription factor, bZIP28. Plant Cell 19, 4111–4119(2007).
28. J. X. Liu, S. H. Howell, bZIP28 and NF-Y transcription factors are activated by ER stressand assemble into a transcriptional complex to regulate stress response genes in Arabi-dopsis. Plant Cell 22, 782–796 (2010).
29. L. N. Kinch, K. Ginalski, N. V. Grishin, Site-2 protease regulated intramembrane pro-teolysis: Sequence homologs suggest an ancient signaling cascade. Protein Sci. 15,84–93 (2006).
30. Y. Iwata, N. V. Fedoroff, N. Koizumi, Arabidopsis bZIP60 is a proteolysis-activatedtranscription factor involved in the endoplasmic reticulum stress response. Plant Cell20, 3107–3121 (2008).
31. H. Gao, F. Brandizzi, C. Benning, R. M. Larkin, A membrane-tethered transcription factordefines a branch of the heat stress response in Arabidopsis thaliana. Proc. Natl. Acad.Sci. U.S.A. 105, 16398–16403 (2008).
32. S. Kamauchi, H. Nakatani, C. Nakano, R. Urade, Gene expression in response to endo-plasmic reticulum stress in Arabidopsis thaliana. FEBS J. 272, 3461–3476 (2005).
33. S. Wang, S. Narendra, N. Fedoroff, Heterotrimeric G protein signaling in the Arabidopsisunfolded protein response. Proc. Natl. Acad. Sci. U.S.A. 104, 3817–3822 (2007).
34. H. Guo, L. Li, H. Ye, X. Yu, A. Algreen, Y. Yin, Three related receptor-like kinases arerequired for optimal cell elongation in Arabidopsis thaliana. Proc. Natl. Acad. Sci. U.S.A.106, 7648–7653 (2009).
35. H. Goda, Y. Shimada, T. Asami, S. Fujioka, S. Yoshida, Microarray analysis ofbrassinosteroid-regulated genes in Arabidopsis. Plant Physiol. 130, 1319–1334 (2002).
36. S. D. Clouse, Brassinosteroid signaling: Novel downstream components emerge.Curr. Biol. 12, R485–R487 (2002).
37. Y. Yin, Z. Y. Wang, S. Mora-Garcia, J. Li, S. Yoshida, T. Asami, J. Chory, BES1 accu-mulates in the nucleus in response to brassinosteroids to regulate gene expression andpromote stem elongation. Cell 109, 181–191 (2002).
38. Y. Yin, D. Vafeados, Y. Tao, S. Yoshida, T. Asami, J. Chory, A new class of transcrip-tion factors mediates brassinosteroid-regulated gene expression in Arabidopsis. Cell120, 249–259 (2005).
IENCESIGNALING.org 28 September 2010 Vol 3 Issue 141 ra69 11

R E S E A R C H A R T I C L E
stke.scienceD
ownloaded from
39. Z. Hong, H. Jin, T. Tzfira, J. Li, Multiple mechanism-mediated retention of a defectivebrassinosteroid receptor in the endoplasmic reticulum of Arabidopsis. Plant Cell 20,3418–3429 (2008).
40. H. Jin, Z. Hong, W. Su, J. Li, A plant-specific calreticulin is a key retention factor for adefective brassinosteroid receptor in the endoplasmic reticulum. Proc. Natl. Acad.Sci. U.S.A. 106, 13612–13617 (2009).
41. T. Noguchi, S. Fujioka, S. Choe, S. Takatsuto, S. Yoshida, H. Yuan, K. A. Feldmann,F. E. Tax, Brassinosteroid-insensitive dwarf mutants of Arabidopsis accumulatebrassinosteroids. Plant Physiol. 121, 743–752 (1999).
42. K. Tanaka, T. Asami, S. Yoshida, Y. Nakamura, T. Matsuo, S. Okamoto, Brassinosteroidhomeostasis in Arabidopsis is ensured by feedback expressions of multiple genesinvolved in its metabolism. Plant Physiol. 138, 1117–1125 (2005).
43. N. Geldner, D. L. Hyman, X. Wang, K. Schumacher, J. Chory, Endosomal signaling ofplant steroid receptor kinase BRI1. Genes Dev. 21, 1598–1602 (2007).
44. H. Jin, Z. Yan, K. H. Nam, J. Li, Allele-specific suppression of a defective brassi-nosteroid receptor reveals a physiological role of UGGT in ER quality control. Mol. Cell26, 821–830 (2007).
45. Z. Hong, H. Jin, A. C. Fitchette, Y. Xia, A. M. Monk, L. Faye, J. Li, Mutations of an a1,6mannosyltransferase inhibit endoplasmic reticulum–associated degradation of defec-tive brassinosteroid receptors in Arabidopsis. Plant Cell 21, 3792–3802 (2009).
46. W. Tang, T. W. Kim, J. A. Oses-Prieto, Y. Sun, Z. Deng, S. Zhu, R. Wang, A. L. Burlingame,Z. Y. Wang, BSKs mediate signal transduction from the receptor kinase BRI1 inArabidopsis. Science 321, 557–560 (2008).
47. L. Song, Q. M. Shi, X. H. Yang, Z. H. Xu, H. W. Xue, Membrane steroid-binding pro-tein 1 (MSBP1) negatively regulates brassinosteroid signaling by enhancing the en-docytosis of BAK1. Cell Res. 19, 864–876 (2009).
48. T. W. Kim, S. Guan, Y. Sun, Z. Deng, W. Tang, J. X. Shang, Y. Sun, A. L. Burlingame,Z. Y. Wang, Brassinosteroid signal transduction from cell-surface receptor kinases tonuclear transcription factors. Nat. Cell Biol. 11, 1254–1260 (2009).
49. C. M. van der Weele, W. G. Spollen, R. E. Sharp, T. I. Baskin, Growth of Arabidopsisthaliana seedlings under water deficit studied by control of water potential in nutrient-agar media. J. Exp. Bot. 51, 1555–1562 (2000).
50. R. P. Hellens, E. A. Edwards, N. R. Leyland, S. Bean, P. M. Mullineaux, pGreen: Aversatile and flexible binary Ti vector for Agrobacterium-mediated plant transforma-tion. Plant Mol. Biol. 42, 819–832 (2000).
51. A. A. Wiszniewski, W. Zhou, S. M. Smith, J. D. Bussell, Identification of two Arabidopsisgenes encoding a peroxisomal oxidoreductase-like protein and an acyl-CoA synthetase-
www.SC
like protein that are required for responses to pro-auxins. Plant Mol. Biol. 69, 503–515(2009).
52. B. K. Nelson, X. Cai, A. Nebenführ, A multicolored set of in vivo organelle markers for co-localization studies in Arabidopsis and other plants. Plant J. 51, 1126–1136 (2007).
53. S. Pagny, M. Cabanes-Macheteau, J. W. Gillikin, N. Leborgne-Castel, P. Lerouge,R. S. Boston, L. Faye, V. Gomord, Protein recycling from the Golgi apparatus to the en-doplasmic reticulum in plants and its minor contribution to calreticulin retention. Plant Cell12, 739–756 (2000).
54. T. Czechowski, M. Stitt, T. Altmann, M. K. Udvardi, W. R. Scheible, Genome-wide iden-tification and testing of superior reference genes for transcript normalization in Arabi-dopsis. Plant Physiol. 139, 5–17 (2005).
55. Acknowledgments: We thank R. Boston (North Carolina State University, Raleigh,NC) for providing the antibody against CRT, Y. Yin (Iowa State University, Ames, IA)for BR mutant seeds and antiserum against BES1, Arabidopsis Biological ResourceCenter (Ohio State University, Columbus, OH) and Nottingham Arabidopsis StockCentre (University of Nottingham, Nottingham, UK) for Arabidopsis T-DNA mutants,and G. Ingram (University of Edinburgh, Edinburgh, UK) and D. Nelson (University ofWestern Australia, Perth, Western Australia, Australia) for critical comments on themanuscript. We acknowledge the Australian Microscopy and Microanalysis ResearchFacility at the Centre for Microscopy, Characterisation and Analysis, University of West-ern Australia, a facility funded by The University, State and Commonwealth Governments.Funding: This work was supported by the Australian Research Council (grantsFF0457721 and CE0561495) and the Government of Western Australia Centres of Ex-cellence scheme. Author contributions: P.C., W.Z., and S.M.S. initiated the project.P.C. performed all the experiments except the confocal microcopy and microarray,which were performed by J.D.B., and leaf dehydration experiments by G.E. and B.J.P.All authors were involved in the interpretation of data and manuscript content. P.C.and S.M.S. wrote the manuscript. Competing interests: The authors declare thatthey have no competing interests.
Submitted 30 April 2010Accepted 10 September 2010Final Publication 28 September 201010.1126/scisignal.2001140Citation: P. Che, J. D. Bussell, W. Zhou, G. M. Estavillo, B. J. Pogson, S. M. Smith, Signalingfrom the endoplasmic reticulum activates brassinosteroid signaling and promotes acclimationto stress in Arabidopsis. Sci. Signal. 3, ra69 (2010).
ma
IENCESIGNALING.org 28 September 2010 Vol 3 Issue 141 ra69 12
on January 7, 2014 g.org




![Reactive Oxygen Species Are Involved in Brassinosteroid-Induced … · Reactive Oxygen Species Are Involved in Brassinosteroid-Induced Stress Tolerance in Cucumber1[W] ... Vert and](https://img.pdfslide.us/doc/110x75/60ebd8840c3a8322ad22a20e/reactive-oxygen-species-are-involved-in-brassinosteroid-induced-reactive-oxygen.jpg)