Embed Size (px)
Citation preview

Shear rheology of lipid monolayers andinsights on membrane fluidityGabriel Espinosaa, Iván López-Monterob, Francisco Monroya,b,1, and Dominique Langevina,1
aLaboratoire de Physique des Solides, Université Paris Sud XI, Unité Mixte de Recherche Centre National de la Recherche Scientifique 8502, 91405 Orsay,France; and bMechanics of Biological Membranes and Biorheology, Department of Physical Chemistry I, Universidad Complutense, 28040 Madrid, Spain
Edited* by Benjamin Widom, Cornell University, Ithaca, NY, and approved February 16, 2011 (received for review December 10, 2010)
The concept of membrane fluidity usually refers to a high molecu-lar mobility inside the lipid bilayer which enables lateral diffusionof embedded proteins. Fluids have the ability to flow under anapplied shear stress whereas solids resist shear deformations. Bio-logical membranes require both properties for their function: highlateral fluidity and structural rigidity. Consequently, an adequateaccount must include, in addition to viscosity, the possibility fora nonzero shear modulus. This knowledge is still lacking as mea-surements of membrane shear properties have remained incom-plete so far. In the present contribution we report a surface shearrheology study of different lipid monolayers that model distinctbiologically relevant situations. The results evidence a large varietyof mechanical behavior under lateral shear flow.
One fundamental concept of cell biology is the recognitionthat the cell membrane is a fluid assembly of amphiphilic
molecules into a two-dimensional liquid crystalline structure (1).This structure is rigid enough to form a stable container but isfluid enough to allow lateral transport of the different membranecomponents and a differential permeability essential for cellhomeostasis (2, 3). The membrane fluidity entails lipids andmembrane proteins mobility, which is essential for their mutualinterplay and function (2, 4, 5). Indeed, adequate protein func-tion requires an energetically efficient conformational dynamics,which is only possible in a mechanically adapted membrane med-ium (6–8). Lipid bilayers are generally assumed to behave assimple Newtonian fluids, however they possess a bending anda compression elasticity (6, 9), thus the question of the existenceof a shear elasticity comes to mind. Very recently, Harland et al.showed that bilayers of single phospholipids are not purely vis-cous, but viscoelastic, with an elastic modulus that diverges atthe fluid-gel transition (10). Although the fluid mosaic pictureallows for a satisfactory description of the molecular dynamicsof the embedded objects (8, 11, 12), lacking of viscoelastic con-tributions could result in an incomplete understanding of mem-brane processes. A rigorous picture of the mechanical behavior ofa membrane includes the resistance to shear, compression, andbending. Furthermore, the concept of membrane fluidity, whenreferred to an ability of the membrane to flow under an appliedshear stress, should be distinguished from the concept of mole-cular mobility inside the membrane (13), or of the local micro-viscosity measured from changes in the mobility of a molecularfluorescent probe (14). In this sense, the high diffusion mobilitytypical of disordered lipid phases might be consistent with a finitemacroscopic shear viscosity.
The present work addresses the fundamental question of mem-brane fluidity with a systematic study of the surface shear rheol-ogy of model Langmuir monolayers. Marsh demonstrated thatthe properties of monolayers and bilayers are equivalent whenthe monolayer surface pressure equals the hydrophobic freeenergy of the bilayer per unit area (π ≈ 30–35 mN∕m) (15). How-ever, the work of Marsh focused on single component bilayers.For two components or more, interactions across the leaflets cangenerate correlations that lead to demixing in bilayers whereasmonolayers remain monophasic, as shown very recently in thecase of ternary lipid mixtures by Ziblat et al. (16). The structure
of the bilayers can then be appreciably different from that ofthe corresponding monolayers, especially at large cholesterolcontent, thus with a possible influence on their dynamics. Inour study, we focused on single phase monolayers, containingamounts of cholesterol smaller than the one leading to phase se-paration in bilayers (see ref. 17 for a study), to minimize possibledifferences with the properties of bilayers.
Membrane flow behavior is characterized by an intrinsic sur-face viscosity (η) relevant for in-plane motion and an intermono-layer friction opposing velocity gradients across the layer (18).For thermal motions, intermonolayer slippage hardly affects η,which is primarily determined by the lateral packing inside themonolayers (18). Thus, interdigitation and intermonolayer cou-pling effects might consequently impact less surface viscosity thanphase behavior, provided such a coupling does not lead to phaseseparation and situations such as formation of cholesterol crystalsreported in ref. 16.
Model Monolayers and Surface Shear RheologyHere, we focus on the shear viscoelasticity of homogeneous lipidphases spanning a large range of membrane fluidity relevant todifferent biological states (Table 1). Oscillatory rheology experi-ments were performed at a surface pressure of 30 mN∕m and at37 °C, which is the representative state of lipid packing in physio-logical conditions (6, 15).
There are two main families of membrane lipids, namely gly-cerophospholipids and sphingolipids (3, 19). Although a varietyof acyl chains are present in cells, lipids of the first class are foundpredominantly monounsaturated, the palmitoyl-oleyl-sn-glycero-phosphatidylcholine (POPC) being a main component of thefluid matrix of most eukaryote membranes. The fully saturatedcomponent, dipalmitoyl-sn-glycero-phosphatidylcholine (DPPC)melts well above room temperature. Sphingomyelin (SM) is com-posed by fully saturated sphingosine mainly localized in choles-terol-rich domains. These lipids (POPC and SM) have beenmutually mixed with cholesterol (Chol) at near physiological pro-portions (about 30% mol) (2, 3). The different mixtures werechosen to represent different states of plasma membranes in pro-karyote cells. The monolayers of a native lipid extract of the innermembrane of Escherichia coli have been studied to model prokar-yote membranes. Ceramide (Cer), an essential messenger in theapoptosis pathway, has also been studied.
Linear Viscoelasticity and Flow DynamicsDisordered Liquid Phases: High Fluidity. Unsaturated glycerolipidsare known to assemble into bilayers in water, forming liquid
Author contributions: F.M. and D.L. designed research; G.E., I.L.-M., and F.M. performedresearch; I.L.-M. contributed new reagents/analytic tools; F.M. analyzed data; and F.M.and D.L. wrote the paper.
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.1To whom correspondence may be addressed. E-mail: [email protected] or [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1018572108/-/DCSupplemental.
6008–6013 ∣ PNAS ∣ April 12, 2011 ∣ vol. 108 ∣ no. 15 www.pnas.org/cgi/doi/10.1073/pnas.1018572108
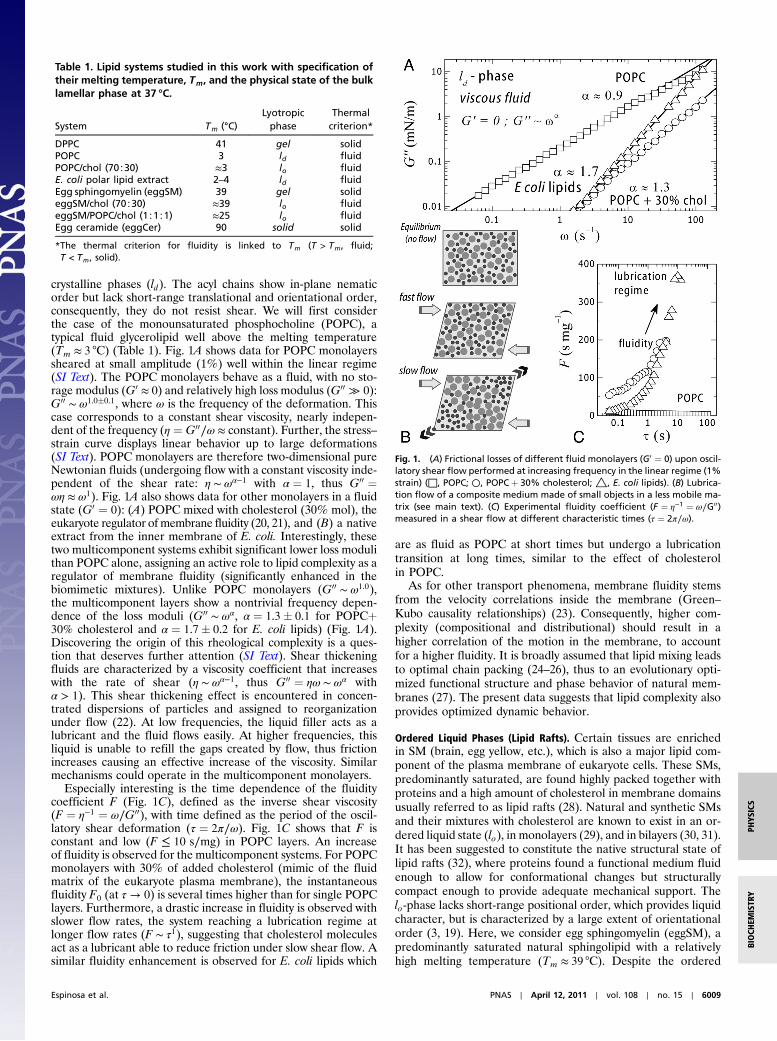
crystalline phases (ld). The acyl chains show in-plane nematicorder but lack short-range translational and orientational order,consequently, they do not resist shear. We will first considerthe case of the monounsaturated phosphocholine (POPC), atypical fluid glycerolipid well above the melting temperature(Tm ≈ 3 °C) (Table 1). Fig. 1A shows data for POPC monolayerssheared at small amplitude (1%) well within the linear regime(SI Text). The POPC monolayers behave as a fluid, with no sto-rage modulus (G0 ≈ 0) and relatively high loss modulus (G00 ≫ 0):G00 ∼ ω1.0�0.1, where ω is the frequency of the deformation. Thiscase corresponds to a constant shear viscosity, nearly indepen-dent of the frequency (η ¼ G00∕ω ≈ constant). Further, the stress–strain curve displays linear behavior up to large deformations(SI Text). POPC monolayers are therefore two-dimensional pureNewtonian fluids (undergoing flow with a constant viscosity inde-pendent of the shear rate: η ∼ ωα−1 with α ¼ 1, thus G00 ¼ωη ≈ ω1). Fig. 1A also shows data for other monolayers in a fluidstate (G0 ¼ 0): (A) POPC mixed with cholesterol (30% mol), theeukaryote regulator of membrane fluidity (20, 21), and (B) a nativeextract from the inner membrane of E. coli. Interestingly, thesetwo multicomponent systems exhibit significant lower loss modulithan POPC alone, assigning an active role to lipid complexity as aregulator of membrane fluidity (significantly enhanced in thebiomimetic mixtures). Unlike POPC monolayers (G00 ∼ ω1.0),the multicomponent layers show a nontrivial frequency depen-dence of the loss moduli (G00 ∼ ωα, α ¼ 1.3� 0.1 for POPCþ30% cholesterol and α ¼ 1.7� 0.2 for E. coli lipids) (Fig. 1A).Discovering the origin of this rheological complexity is a ques-tion that deserves further attention (SI Text). Shear thickeningfluids are characterized by a viscosity coefficient that increaseswith the rate of shear (η ∼ ωα−1, thus G00 ¼ ηω ∼ ωα withα > 1). This shear thickening effect is encountered in concen-trated dispersions of particles and assigned to reorganizationunder flow (22). At low frequencies, the liquid filler acts as alubricant and the fluid flows easily. At higher frequencies, thisliquid is unable to refill the gaps created by flow, thus frictionincreases causing an effective increase of the viscosity. Similarmechanisms could operate in the multicomponent monolayers.
Especially interesting is the time dependence of the fluiditycoefficient F (Fig. 1C), defined as the inverse shear viscosity(F ¼ η−1 ¼ ω∕G00), with time defined as the period of the oscil-latory shear deformation (τ ¼ 2π∕ω). Fig. 1C shows that F isconstant and low (F ≤ 10 s∕mg) in POPC layers. An increaseof fluidity is observed for the multicomponent systems. For POPCmonolayers with 30% of added cholesterol (mimic of the fluidmatrix of the eukaryote plasma membrane), the instantaneousfluidity F0 (at τ → 0) is several times higher than for single POPClayers. Furthermore, a drastic increase in fluidity is observed withslower flow rates, the system reaching a lubrication regime atlonger flow rates (F ∼ τ1), suggesting that cholesterol moleculesact as a lubricant able to reduce friction under slow shear flow. Asimilar fluidity enhancement is observed for E. coli lipids which
are as fluid as POPC at short times but undergo a lubricationtransition at long times, similar to the effect of cholesterolin POPC.
As for other transport phenomena, membrane fluidity stemsfrom the velocity correlations inside the membrane (Green–Kubo causality relationships) (23). Consequently, higher com-plexity (compositional and distributional) should result in ahigher correlation of the motion in the membrane, to accountfor a higher fluidity. It is broadly assumed that lipid mixing leadsto optimal chain packing (24–26), thus to an evolutionary opti-mized functional structure and phase behavior of natural mem-branes (27). The present data suggests that lipid complexity alsoprovides optimized dynamic behavior.
Ordered Liquid Phases (Lipid Rafts). Certain tissues are enrichedin SM (brain, egg yellow, etc.), which is also a major lipid com-ponent of the plasma membrane of eukaryote cells. These SMs,predominantly saturated, are found highly packed together withproteins and a high amount of cholesterol in membrane domainsusually referred to as lipid rafts (28). Natural and synthetic SMsand their mixtures with cholesterol are known to exist in an or-dered liquid state (lo), in monolayers (29), and in bilayers (30, 31).It has been suggested to constitute the native structural state oflipid rafts (32), where proteins found a functional medium fluidenough to allow for conformational changes but structurallycompact enough to provide adequate mechanical support. Thelo-phase lacks short-range positional order, which provides liquidcharacter, but is characterized by a large extent of orientationalorder (3, 19). Here, we consider egg sphingomyelin (eggSM), apredominantly saturated natural sphingolipid with a relativelyhigh melting temperature (Tm ≈ 39 °C). Despite the ordered
Fig. 1. (A) Frictional losses of different fluid monolayers (G0 ¼ 0) upon oscil-latory shear flow performed at increasing frequency in the linear regime (1%strain) (□, POPC; ○, POPCþ 30% cholesterol; △, E. coli lipids). (B) Lubrica-tion flow of a composite medium made of small objects in a less mobile ma-trix (see main text). (C) Experimental fluidity coefficient (F ¼ η−1 ¼ ω∕G00)measured in a shear flow at different characteristic times (τ ¼ 2π∕ω).
Table 1. Lipid systems studied in this work with specification oftheir melting temperature, Tm, and the physical state of the bulklamellar phase at 37 °C.
System Tm (°C)Lyotropicphase
Thermalcriterion*
DPPC 41 gel solidPOPC 3 ld fluidPOPC/chol (70∶30) ≈3 lo fluidE. coli polar lipid extract 2–4 ld fluidEgg sphingomyelin (eggSM) 39 gel solideggSM/chol (70∶30) ≈39 lo fluideggSM/POPC/chol (1∶1∶1) ≈25 lo fluidEgg ceramide (eggCer) 90 solid solid
*The thermal criterion for fluidity is linked to Tm (T > Tm, fluid;T < Tm, solid).
Espinosa et al. PNAS ∣ April 12, 2011 ∣ vol. 108 ∣ no. 15 ∣ 6009
BIOCH
EMISTR
YPH
YSICS
character of these dense assemblies (19, 33), they appear fluid-like under shear with a vanishing shear rigidity (G0 ≈ 0).
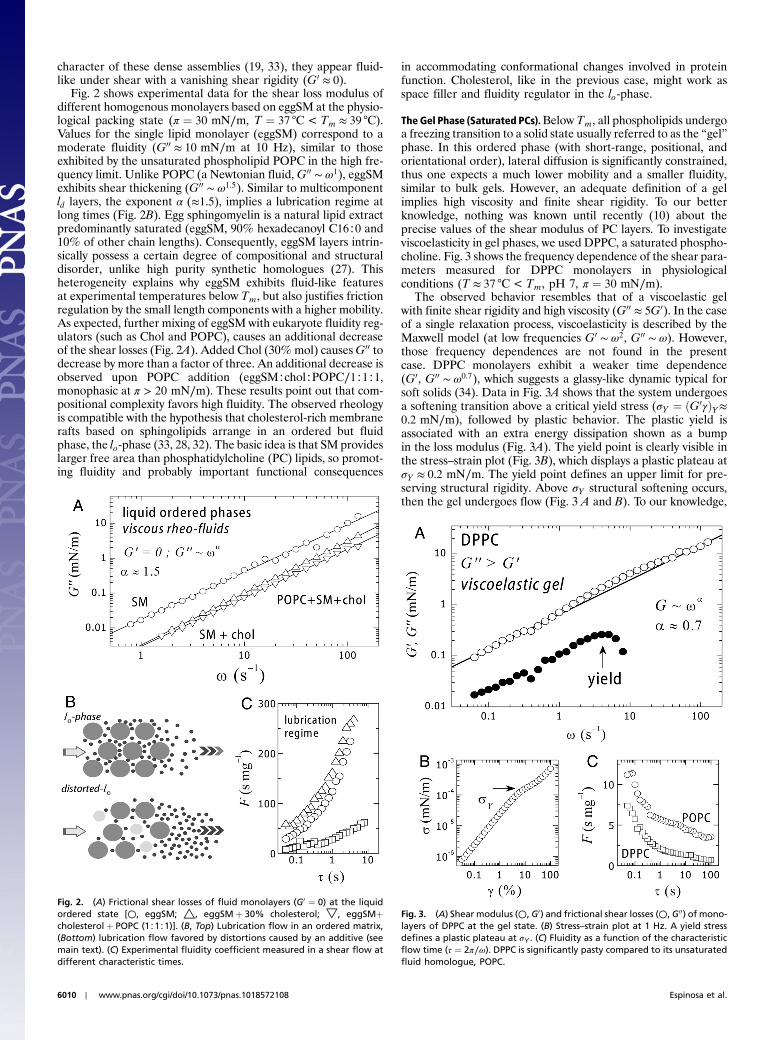
Fig. 2 shows experimental data for the shear loss modulus ofdifferent homogenous monolayers based on eggSM at the physio-logical packing state (π ¼ 30 mN∕m, T ¼ 37 °C < Tm ≈ 39 °C).Values for the single lipid monolayer (eggSM) correspond to amoderate fluidity (G00 ≈ 10 mN∕m at 10 Hz), similar to thoseexhibited by the unsaturated phospholipid POPC in the high fre-quency limit. Unlike POPC (a Newtonian fluid, G00 ∼ ω1), eggSMexhibits shear thickening (G00 ∼ ω1.5). Similar to multicomponentld layers, the exponent α (≈1.5), implies a lubrication regime atlong times (Fig. 2B). Egg sphingomyelin is a natural lipid extractpredominantly saturated (eggSM, 90% hexadecanoyl C16∶0 and10% of other chain lengths). Consequently, eggSM layers intrin-sically possess a certain degree of compositional and structuraldisorder, unlike high purity synthetic homologues (27). Thisheterogeneity explains why eggSM exhibits fluid-like featuresat experimental temperatures below Tm, but also justifies frictionregulation by the small length components with a higher mobility.As expected, further mixing of eggSM with eukaryote fluidity reg-ulators (such as Chol and POPC), causes an additional decreaseof the shear losses (Fig. 2A). Added Chol (30%mol) causesG00 todecrease by more than a factor of three. An additional decrease isobserved upon POPC addition (eggSM∶chol∶POPC∕1∶1∶1,monophasic at π > 20 mN∕m). These results point out that com-positional complexity favors high fluidity. The observed rheologyis compatible with the hypothesis that cholesterol-rich membranerafts based on sphingolipids arrange in an ordered but fluidphase, the lo-phase (33, 28, 32). The basic idea is that SM provideslarger free area than phosphatidylcholine (PC) lipids, so promot-ing fluidity and probably important functional consequences
in accommodating conformational changes involved in proteinfunction. Cholesterol, like in the previous case, might work asspace filler and fluidity regulator in the lo-phase.
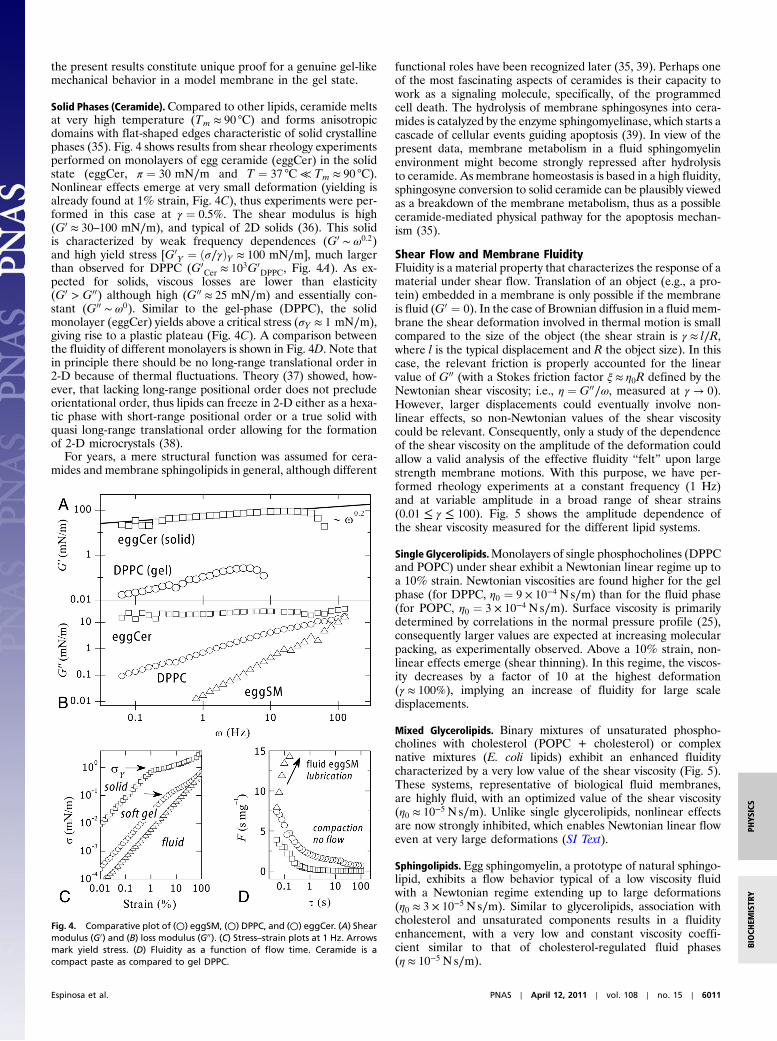
The Gel Phase (Saturated PCs).Below Tm, all phospholipids undergoa freezing transition to a solid state usually referred to as the “gel”phase. In this ordered phase (with short-range, positional, andorientational order), lateral diffusion is significantly constrained,thus one expects a much lower mobility and a smaller fluidity,similar to bulk gels. However, an adequate definition of a gelimplies high viscosity and finite shear rigidity. To our betterknowledge, nothing was known until recently (10) about theprecise values of the shear modulus of PC layers. To investigateviscoelasticity in gel phases, we used DPPC, a saturated phospho-choline. Fig. 3 shows the frequency dependence of the shear para-meters measured for DPPC monolayers in physiologicalconditions (T ≈ 37 °C < Tm, pH 7, π ¼ 30 mN∕m).
The observed behavior resembles that of a viscoelastic gelwith finite shear rigidity and high viscosity (G00 ≈ 5G0). In the caseof a single relaxation process, viscoelasticity is described by theMaxwell model (at low frequencies G0 ∼ ω2, G00 ∼ ω). However,those frequency dependences are not found in the presentcase. DPPC monolayers exhibit a weaker time dependence(G0, G00 ∼ ω0.7), which suggests a glassy-like dynamic typical forsoft solids (34). Data in Fig. 3A shows that the system undergoesa softening transition above a critical yield stress (σY ¼ ðG0γÞY≈0.2 mN∕m), followed by plastic behavior. The plastic yield isassociated with an extra energy dissipation shown as a bumpin the loss modulus (Fig. 3A). The yield point is clearly visible inthe stress–strain plot (Fig. 3B), which displays a plastic plateau atσY ≈ 0.2 mN∕m. The yield point defines an upper limit for pre-serving structural rigidity. Above σY structural softening occurs,then the gel undergoes flow (Fig. 3 A and B). To our knowledge,
Fig. 2. (A) Frictional shear losses of fluid monolayers (G0 ¼ 0) at the liquidordered state [○, eggSM; △, eggSMþ 30% cholesterol; ▽, eggSMþcholesterol þ POPC (1∶1∶1)]. (B, Top) Lubrication flow in an ordered matrix,(Bottom) lubrication flow favored by distortions caused by an additive (seemain text). (C) Experimental fluidity coefficient measured in a shear flow atdifferent characteristic times.
Fig. 3. (A) Shear modulus (○, G0) and frictional shear losses (○, G00) of mono-layers of DPPC at the gel state. (B) Stress–strain plot at 1 Hz. A yield stressdefines a plastic plateau at σY . (C) Fluidity as a function of the characteristicflow time (τ ¼ 2π∕ω). DPPC is significantly pasty compared to its unsaturatedfluid homologue, POPC.
6010 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1018572108 Espinosa et al.
the present results constitute unique proof for a genuine gel-likemechanical behavior in a model membrane in the gel state.
Solid Phases (Ceramide).Compared to other lipids, ceramide meltsat very high temperature (Tm ≈ 90 °C) and forms anisotropicdomains with flat-shaped edges characteristic of solid crystallinephases (35). Fig. 4 shows results from shear rheology experimentsperformed on monolayers of egg ceramide (eggCer) in the solidstate (eggCer, π ¼ 30 mN∕m and T ¼ 37 °C ≪ Tm ≈ 90 °C).Nonlinear effects emerge at very small deformation (yielding isalready found at 1% strain, Fig. 4C), thus experiments were per-formed in this case at γ ¼ 0.5%. The shear modulus is high(G0 ≈ 30–100 mN∕m), and typical of 2D solids (36). This solidis characterized by weak frequency dependences (G0 ∼ ω0.2)and high yield stress [G0
Y ¼ ðσ∕γÞY ≈ 100 mN∕m], much largerthan observed for DPPC (G0
Cer ≈ 103G0DPPC, Fig. 4A). As ex-
pected for solids, viscous losses are lower than elasticity(G0 > G00) although high (G00 ≈ 25 mN∕m) and essentially con-stant (G00 ∼ ω0). Similar to the gel-phase (DPPC), the solidmonolayer (eggCer) yields above a critical stress (σY ≈ 1 mN∕m),giving rise to a plastic plateau (Fig. 4C). A comparison betweenthe fluidity of different monolayers is shown in Fig. 4D. Note thatin principle there should be no long-range translational order in2-D because of thermal fluctuations. Theory (37) showed, how-ever, that lacking long-range positional order does not precludeorientational order, thus lipids can freeze in 2-D either as a hexa-tic phase with short-range positional order or a true solid withquasi long-range translational order allowing for the formationof 2-D microcrystals (38).
For years, a mere structural function was assumed for cera-mides and membrane sphingolipids in general, although different
functional roles have been recognized later (35, 39). Perhaps oneof the most fascinating aspects of ceramides is their capacity towork as a signaling molecule, specifically, of the programmedcell death. The hydrolysis of membrane sphingosynes into cera-mides is catalyzed by the enzyme sphingomyelinase, which starts acascade of cellular events guiding apoptosis (39). In view of thepresent data, membrane metabolism in a fluid sphingomyelinenvironment might become strongly repressed after hydrolysisto ceramide. As membrane homeostasis is based in a high fluidity,sphingosyne conversion to solid ceramide can be plausibly viewedas a breakdown of the membrane metabolism, thus as a possibleceramide-mediated physical pathway for the apoptosis mechan-ism (35).
Shear Flow and Membrane FluidityFluidity is a material property that characterizes the response of amaterial under shear flow. Translation of an object (e.g., a pro-tein) embedded in a membrane is only possible if the membraneis fluid (G0 ¼ 0). In the case of Brownian diffusion in a fluid mem-brane the shear deformation involved in thermal motion is smallcompared to the size of the object (the shear strain is γ ≈ l∕R,where l is the typical displacement and R the object size). In thiscase, the relevant friction is properly accounted for the linearvalue of G00 (with a Stokes friction factor ξ ≈ η0R defined by theNewtonian shear viscosity; i.e., η ¼ G00∕ω, measured at γ → 0).However, larger displacements could eventually involve non-linear effects, so non-Newtonian values of the shear viscositycould be relevant. Consequently, only a study of the dependenceof the shear viscosity on the amplitude of the deformation couldallow a valid analysis of the effective fluidity “felt” upon largestrength membrane motions. With this purpose, we have per-formed rheology experiments at a constant frequency (1 Hz)and at variable amplitude in a broad range of shear strains(0.01 ≤ γ ≤ 100). Fig. 5 shows the amplitude dependence ofthe shear viscosity measured for the different lipid systems.
Single Glycerolipids.Monolayers of single phosphocholines (DPPCand POPC) under shear exhibit a Newtonian linear regime up toa 10% strain. Newtonian viscosities are found higher for the gelphase (for DPPC, η0 ¼ 9 × 10−4 Ns∕m) than for the fluid phase(for POPC, η0 ¼ 3 × 10−4 Ns∕m). Surface viscosity is primarilydetermined by correlations in the normal pressure profile (25),consequently larger values are expected at increasing molecularpacking, as experimentally observed. Above a 10% strain, non-linear effects emerge (shear thinning). In this regime, the viscos-ity decreases by a factor of 10 at the highest deformation(γ ≈ 100%), implying an increase of fluidity for large scaledisplacements.
Mixed Glycerolipids. Binary mixtures of unsaturated phospho-cholines with cholesterol (POPC + cholesterol) or complexnative mixtures (E. coli lipids) exhibit an enhanced fluiditycharacterized by a very low value of the shear viscosity (Fig. 5).These systems, representative of biological fluid membranes,are highly fluid, with an optimized value of the shear viscosity(η0 ≈ 10−5 Ns∕m). Unlike single glycerolipids, nonlinear effectsare now strongly inhibited, which enables Newtonian linear floweven at very large deformations (SI Text).
Sphingolipids. Egg sphingomyelin, a prototype of natural sphingo-lipid, exhibits a flow behavior typical of a low viscosity fluidwith a Newtonian regime extending up to large deformations(η0 ≈ 3 × 10−5 Ns∕m). Similar to glycerolipids, association withcholesterol and unsaturated components results in a fluidityenhancement, with a very low and constant viscosity coeffi-cient similar to that of cholesterol-regulated fluid phases(η ≈ 10−5 Ns∕m).
Fig. 4. Comparative plot of (○) eggSM, (○) DPPC, and (○) eggCer. (A) Shearmodulus (G0) and (B) loss modulus (G00). (C) Stress–strain plots at 1 Hz. Arrowsmark yield stress. (D) Fluidity as a function of flow time. Ceramide is acompact paste as compared to gel DPPC.
Espinosa et al. PNAS ∣ April 12, 2011 ∣ vol. 108 ∣ no. 15 ∣ 6011
BIOCH
EMISTR
YPH
YSICS

Ceramide. This highly hydrophobic lipid self-assembles in a verydense solid state (G0 > 0). Flow is highly restricted by structuralstiffness and high friction, resulting in a behavior typical of plasticsolids (34). In the linear regime (γ < 0.9%) the system behaves ashard solid (G0 ≈ 85 mN∕m) with a practically null fluidity (theshear viscosity takes values as high as η0 ≈ 4 × 10−3 Ns∕m, threeorders of magnitude higher than typical fluid phases). Above a1% strain, nonlinear effects emerge with viscous shear thinningfollowed by a sharp structural softening characterized by a dropof the storage modulus (from the linear value, G0
0 ≈ 85 mN∕m,typical of a hard solid down to a vanishing value, G0 ≈ 0 atγ ≥ 10%, typical of a viscous fluid). Consequently, in an en-riched-ceramide membrane, Brownian motion should be largelyimpeded.
Molecular Foundations. In molecular terms, the shear viscosityis expected to increase with the strength of the net attractivepotential (mainly dependent on tail length) and to decrease withincreasing molecular area (18). Data show this general trend:The condensed phospholipid DPPC is more viscous than the un-saturated homologue POPC, whereas charged E. coli lipids(dominated by the electrostatic repulsions) represent the highestfluidity case. Sphingosine is larger than the glycerol head thussphingolipids might, in general, exhibit smaller viscosities thanglycerolipids, as experimentally observed. Ceramide polar headsare less bulky than those of sphingomyelin, because they do nothave phosphate groups, thus enabling much higher lateral cohe-sion and closer packing. Ceramide lipids can therefore pack into asolid structure characterized by high shear stiffness and extremelyhigh viscosity. In general, lipid mixing increases fluidity, even con-comitantly with significant structural condensation (e.g., inducedby cholesterol) (19–21). Molecular disorder and lubricationeffects induced by mixing are thus believed to underlie the func-tional dichotomy—compact, but fluid—of optimized biologicalmembranes.
Biological Implications. This elastic dichotomy may have profoundconsequences in protein function: Protein lateral translation in afluid membrane is not impeded although it takes place at con-stant area, but proteins undergoing conformational transitionsshould exert net forces on the lipid surrounding, which reacts withan equivalent compression force. Systematic studies of surfacerheology might contribute to a better understanding of thismechanical interplay existing in biological membranes, althoughit is secularly ignored in protein energetics.
Furthermore, biological membranes are heterogeneous com-plex media where the different lipids are locally organized
depending on the precise membrane function (2–4). Indeed, or-ganisms invest substantial resources in synthesizing tens ofdifferent lipids and selectively arranging them to produce the dif-ferent membrane structures forming the cell (3). Contrary to arandomized lipid distribution driven by simple entropic sponta-neity, specific interactions are assumed to construct a complexorganization of lipids. The biological principle that structureattends for function implies that there must be evolutionary func-tional advantages for such a complex lipid repertoire and distri-bution. Since their existence proposed by Simons and Ikonen,lipid rafts have been frequently invoked as specialized platformsnecessary for optimal protein function and envisaged to consti-tute a unique element of membrane structure superposed tothe well-known protein mosaicity (28–40). A fundamental ques-tion arises in this context: What is the subtle interplay ofinteractions between the different lipids that endows such a un-ique mechanical behavior of biological membranes (8)? If lipiddomains are present in real membranes, not only energetic con-tributions arising from line tension and long-range domain inter-actions must be explicitly considered in the mechanical kernelof the membrane (25, 41) but also kinetic effects related to themultiphase transport (42, 43) could nontrivially shape the func-tional dynamics in such heterogeneous membranes.
ConclusionsA systematic study of the surface flow properties has beenperformed with monolayers of different membrane lipids. Wepropose that the concept of membrane fluidity is linked to theability to flow under shear, to the absence of shear modulus(G0 ¼ 0) and to low frictional losses characterized by a low visc-osity coefficient. Such a fluid-like behavior has been evidencedfor lipid layers mimetic of different biological membrane struc-tures: prokaryote (E. coli polar lipid extract) and eukaryote(POPC and SM mixed with cholesterol). These fluid layers havebeen evidenced to undergo Newtonian flow characterized by alow shear viscosity (η0 ≈ 10−5 Ns∕m). In only two particular casessolid-like rigidity was encountered: (a) the gel-phase of singlesaturated phospholipids (viscoelastic, G00 > G0 > 0) and (b) solidceramide (typically solid,G0 > G00). Consequently, only under thestrong fluidity requirement (G0 ¼ 0), membrane proteins mightbe able to undergo lateral translation/rotation in diffusion pro-cesses. Although caution is necessary before generalizing ourresults to biologically relevant situations, they capture the essen-tials of mechanical behavior for the different lipid phases found inbiological membranes. They might influence the current wisdomat three levels: (a) molecular, adequate lipid mixing and conse-quent phase behavior enables functional rheology; (b) biophysi-
Fig. 5. Surface shear viscosity of the different lipid monolayers studied in this work. A low shear viscosity (η ≤ 10−5 N s∕m) defines the system as a highlyfluid membrane (glycerolipids and sphingolipids mixed with cholesterol and E. coli lipids). Pasty fluids (POPC), viscoelastic gels (DPPC), or solids (eggCer) arecharacterized by much higher (non-Newtonian) viscosities.
6012 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1018572108 Espinosa et al.

cal, highly fluid, moderately fluid, viscoelastic or solid-like rheol-ogy can be observed for different membranes or membrane sites;and (c) biological, specific membrane function entails adequate—evolutionally optimized—membrane rheology.
MethodsMonolayer Preparation. Lipids were purchased from Avanti Polar Lipids andused without further purification. Lipid monolayers are spread dropwise on abuffered aqueous subphase (50 mM Hepes pH7, 0.1 mM EDTA in Milli-Qwater) from a chloroform solution (ca. 1 mg∕mL). All substances werepurchased from Sigma Aldrich at the highest purity.
Surface Shear Rheology. We used an Anton Paar Physica model MCR-301rheometer, equipped with a biconical bob (68.3 mm diameter, 5° cone angle).A plane shear strain is applied and normal axial torque is measured in the
oscillatory mode, γ ¼ γ0 sinωt (γ0 is the strain amplitude). Using the theore-tical treatment of interfacial flow established by Oh and Slattery (44), aftercorrection from bulk water contributions, G0 and G00 are calculated as thematerial parameters defining the interfacial stress σðtÞ ¼ γ0 (G0 sinωtþG00 cosωt), the surface shear viscosity being η ¼ G00∕ω. Temperature wascontrolled with a Peltier element.
ACKNOWLEDGMENTS. This work was supported by Grants FIS2009-14650-C02-01 and CSD2007-0010 (Consolider-Ingenio 2010: NanocienciaMolecular) from Ministerio de Ciencia e Innovación and S2009MAT-1507from Comunidad Autonoma de Madrid. G.E. thanks Consejo Nacional deCiencia y Technología (Mexico) for a doctoral fellowship. I.L.M. is supportedby the Juan de la Cierva program. F.M. acknowledges support fromUniversidad Complutense de Madrid and Triangle de la Physique for aResearch Fellowship during a sabbatical stay at Laboratoire de Physiquedes Solides.
1. Singer SJ, Nicolson GL (1972) The fluid mosaic model of the structure of cellmembranes. Science 175:720–731.
2. Alberts B, et al. (2002) Molecular Biology of the Cell (Garland Science, New York), 4thEd.,, pp 617–650.
3. van Meer G, Voelker DR, Feigenson GW (2008) Membrane lipids: Where they are andhow they behave. Nat Rev Mol Cell Biol 9:112–124.
4. Sprong H, van der Sluijs P, vanMeer G (2001) How proteins move lipids and lipids moveproteins. Nat Rev Mol Cell Biol 2:504–513.
5. Philips R, Ursell T, Wiggins P, Sens P (2009) Emerging roles for lipids in shapingmembrane-protein function. Nature 459:379–385.
6. Boal DH (2002) Mechanics of the Cell (Cambridge University Press, Cambridge, U.K.),pp 135–204.
7. Clegg RM, Vaz WLC (1985) Progress in Protein-Lipid Interactions, eds A Watts andJJHHM de Pont (Elsevier, Amsterdam), pp 173–229.
8. Marguet D, Lenne PF, Rigneault H, He HT (2006) Dynamics in the plasma membrane:How to combine fluidity and order. EMBO J 25:3446–3457.
9. RawiczW, Olbrich K, McIntosh T, NeedhamD, Evans E (2000) Effect of chain length andunsaturation on lipid bilayer elasticity. Biophys J 79:328–339.
10. Harland CW, Bradley MJ, Parthasarathy R (2010) Phospholipid bilayers are viscoelastic.Proc Natl Acad Sci USA 107:19146–19150.
11. Saffman PG, Delbrück M (1975) Brownian motion in biological membranes. Proc NatlAcad Sci USA 72:3111–3114.
12. Gambin Y, et al. (2006) Lateral mobility of proteins in liquid membranes revisited.Proc Natl Acad Sci USA 103:2098–2102.
13. Edidin M (1974) Rotational and translational diffusion in membranes. Annu Rev Bio-phys Bioeng 3:179–201.
14. Rubenstein JLR, Smith BA, McConnell HM (1979) Lateral diffusion in binary mixtures ofcholesterol and phosphatidylcholines. Proc Natl Acad Sci USA 76:15–18.
15. Marsh D (1996) Lateral pressure in membranes. Biochim Biophys Acta Biomembr1286:183–223.
16. Ziblat R, Leiserowitz L, Addadi L (2010) Crystalline domain structure and cholesterolcrystal nucleation in single hydrated DPPC:cholesterol:POPC bilayers. J Am Chem Soc132:9920–9927.
17. Safouane M, et al. (2010) Lipid cosorting mediated by shiga toxin induced tubulation.Traffic 11:1519–1529.
18. den Otter WK, Shkulipa SA (2007) Intermonolayer friction and surface shear viscosityof lipid bilayer membranes. Biophys J 93:423–433.
19. Feigenson GW (2006) Phase behavior of lipid mixtures. Nat Chem Biol 2:560–563.20. Finegold L (1993) Cholesterol in membrane Models (CRC, Boca Raton, FL), pp 1–12.21. Demel RA, de Kruyff B (1976) The function of sterols in membranes. Biochim Biophys
Acta 457:109–132.22. Cheremisihoff NP, ed. (1988) Encyclopedia of Fluid Mechanics: Rheology and
Non-Newtonian Flows (Gulf Publishing Company, Houston), pp 89–134.23. Kubo R (1986) Brownian motion and nonequilibrium statistical mechanics. Science
233:330–334.
24. Elliott R, Szleifer I, Schick M (2006) Phase diagram of a ternary mixture of cholesteroland saturated and saturated lipids calculated from a microscopic model. Phys Rev Lett96(9):098101.
25. McConnell HM (1991) Structures and transitions in lipid monolayers at the air-waterinterface. Annu Rev Phys Chem 42:171–195.
26. Feigenson GW (2007) Phase boundaries and biological membranes. Annu Rev BiophysBiomol Struct 36:63–77.
27. Mouritsen OG (2005) Life–As a Matter of Fat (Springer, Berlin), pp 9–19.28. Simons K, Ikonen E (1997) Functional rafts in cell membranes. Nature 387:569–572.29. Stottrup BL, Srevens DS, Keller SL (2005) Miscibility of ternary mixtures of phospho-
lipids and cholesterol in monolayers and applications to bilayer systems. Biophys J88:269–276.
30. Veatch SL, Keller SL (2003) Separation of liquid phases in giant vesicles of ternarymixtures of phospholipids and cholesterol. Biophys J 85:3074–3083.
31. Almeida RFM, Loura LMS, Fedorov A, Prieto M (2005) Lipid rafts have different sizesdepending on membrane composition: A time resolved FRET study. J Mol Biol346:1109–1120.
32. Edidin M (2003) The state of lipid rafts: From model membranes to cells. Annu RevBiophys Biomol Struct 32:257–283.
33. Brown DA, London E (2000) Structure and function of sphingolipid- and cholesterol-rich membrane rafts. J Biol Chem 275:17221–17224.
34. Sollich P, Lequeux F, Hébraud P, Cates ME (1997) Rheology of soft glassy materials.Phys Rev Lett 78:2020–2023.
35. López-Montero I, Monroy F, Vélez M, Devaux PF (2010) Ceramide: From lateralsegregation to mechanical stress. Biochim Biophys Acta 1798:1348–1356.
36. Zang D, Langevin D, Binks BP, Wei B (2010) Shearing particle monolayers: Strain-ratefrequency superposition. Phys Rev E Stat Nonlinear Soft Matter Phys 81(1):011604.
37. Strandburg KJ (1988) Two-dimensional melting. Rev Mod Phys 60:161–207.38. Helm CA, Möhwald H, Kjaer K, Als-Nielsen J (1987) Phospholipid monolayers between
fluid and solid states. Biophys J 52:381–390.39. Kolesnick R, Hannun YA (1999) Ceramide and apoptosis. Trends Biochem Sci
24:224–225.40. Simons K, Gerl MJ (2010) Revitalizing membrane rafts: New tools and insights. Nat Rev
Mol Cell Biol 11:688–699.41. Ursell T, KlugWS, Philips R (2009) Morphology and interaction between lipid domains.
Proc Natl Acad Sci USA 106:13301–13306.42. McConnell HM (1996) Equilibration rates in lipid monolayers. Proc Nat Acad Sci USA
93:15001–15003.43. Arriaga LR, López-Montero I, Rodríguez-García R, Monroy F (2008) Nonlinear dila-
tional mechanics of Langmuir lipid monolayers: A lateral diffusion mechanism. PhysRev E Stat Nonlinear Soft Matter Phys 77(6):061918.
44. Oh SG, Slattery JC (1978) Disk and biconical interfacial viscosimeters. J Colloid InterfaceSci 67:516–525.
Espinosa et al. PNAS ∣ April 12, 2011 ∣ vol. 108 ∣ no. 15 ∣ 6013
BIOCH
EMISTR
YPH
YSICS





























