Embed Size (px)
Citation preview

I l BlOTROPlCA 24(1): 86-96 1992
Sexual Selection and Sexual Dimorphism in the Harlequin Beetle Acrocinus longimanus’
David W. Zeh, Jeanne A. Zeh Smithsonian Tropical Research Institute, Unit 0948, APO AA 34002-0948 or Apartado 2072 Balboa, Panama
and
Gérard Tavakilian
ORSTOM, B.P. 165, 97323 Cayenne Cedex, France
ABSTRACT To investigate the causes of striking sexual dimorphism in the harlequin beetle (Acrocinus longimanus), we carried out a study of the behavior and morphology of two widely separated populations (in French Guiana and Panama). Males of this species possess greatly elongated forelegs which exhibit strong positive dometry with body size (elytra length). Males use their forelegs in fights with other males and in guarding females and oviposition sites. Field data on fighting and mating success suggest that sexual selection favors large size in males. However, the direct targets of selection were masked by high correlations between traits of the elytra, forelegs, and antennae. Sexual selection appears to be driven by intense male competition to monopolize suitable sites for egg deposition. Female harlequin beetles are highly selective in choosing only recently dead or dying trees for oviposition (Moraceae and Apocynaceae). Despite marked geographic variation in coloration, host trees, and dimate, the two populations did not differ in mean size and extent of sexual dimorphism. We suggest that sexual selection in this species is sufficiently intense to override any effects of differing ecological factors.
RESUMEN A fin de investigar las causas del notable dimorfismo sexual en el escarabajo arlequín (Acrocinus longimanus), realizamos un estudio de la morfología y del comportamiento de dos poblaciones muy separadas (Guayana Francesa y Panamá). Los machos de esta especie poseen patas delanteras muy alargadas que presentan una herte alometria positiva en relación con el tamaño del cuerpo (longitud de los élitros). Se utilizan dichas patas al combatir con otros machos y al guardar hembras al igual que áreas de colocación de huevos. Datos del campo sobre el Mto de combate y de apareamiento sugieren que la selección sexual favorece al tamaño grande en los machos. Sin embargo, fueron ocultados los objetos directos de la selección por correlaciones elevadas entre características de los élitros, las patas delanteras y las antenas. La selección sexual parece estar orientada fundamentalmente por una intensa competencia entre los machos para monopolizar sitios apropiados para la colocación de los huevos. Las hembras son sumamente selectivas al escoger para la oviposición sólo árboles recién muertos o aquellos a punto de perecer (Moraceae y Apocynaceae). A pesar de una pronunciada variación geográfica en la coloración, en los árboles hospedadores y en el dima, no presentaron las dos poblaciones diferencia en tamaño medio y nivel de dimorfismo sexual. Proponemos que en esta especie sea la selección sexual lo bastante intensa para anular cualquier efecto de diferencias en factores ecológicos.
IN MANY ANIMALS, MALES AND FEMALES exhibit pro- in a wide variety of taxa (Eden & Oring 1977, nounced morphological differences thought to have Zeh 1987a). evolved through male competition for, or female Some of the best known cases of sexually di- choice of, mates, i.e., sexual selection (Darwin 1871, morphic traits important in male combat are found Trivers 1972, Bradbury & Andersson 1987). Extent among beetles (Darwin 1871, Otte & Stayman of sexual dimorphism depends on an interplay be- 1979, Eberhard 1980, Thornhill & Alcock 1983, tween phylogeny, genetic architecture, ecology and Conner 1988). Male weaponry may consist of ce- life history trade-offs (Lande 1980, 1982; Cheverud phalic horns which are absent or poorly developed et al. 1985; Zeh 1987b). Despite this complexity, in females (e.g., Dynastinae) or enlarged mandibles ecological factors enabling males to monopolize fe- such as those characteristic of many Lucanidae. Far males or essential resources have dearly been-critical less common are species dimorphic in the size and in thë evolution of Zecondary sexual characteristics\) shape of the forelegs. Acrocinus longimanus (L.),
I provides a striking example. Ranging from southern Received 30 January 1991, revision accepted 15 July MexiCO to r m d ~ ~ ~ &!en *
1991. - - harlequin beetle is named for
“i \ l ~ l~ l l l~~~~~~~i l~~~~l r r 1 86
- - 1 010023 131 \ . - *-?.-- - - . - *
-
A
4 I c m
C
J!
FIGURE 1. Sexu female. C. Large m straight tibiae of tt
pattern of black orange markings of this giant long leg length may rf females of comp; occurs in the sh nearly straight ir grees of curvam the factors influi species, we carril passing behavior between male s knowledge, this i ing and agonistic
METHODS 7 Iequin beetle ? I R ~ntecayingtrees,

, $t
A
Harlequin Beetle Sexual Selection 87
B
mus), we carried na and Panama). body size (elytra
i sites. Field data 9 direct targets of selection appears harlequin beetles ynaceae). Despite iffer in mean size [tense to override
anus), realizamos ncesa y Panamá). ietría positiva en i otros machos y de combate y de fueron ocultados )atas delanteras y entre los machos
!ectivas al escoger zeae). A pesar de o presentaron las sta especie sea la
L Oring ‘1977,
of sexually di- nbat are found te & Stayman Alcock 1983,
I consist of ce- Orly developed ged mandibles Lucanidae. Far in the size and ngimanus (L.), from southern
19601, the inctive swirling
D
FIGURE 1. Sexual dimorphism and within-gender variability in Acrocinus longimanus. A. S m d female. B. Large female. C. Large male. D. S m d male. Nore the marked curvature of the large male’s tibiae compared to the nearly straight tibiae of the small male.
pattern of black, greenish-yellow and crimson or orange markings on its thorax and elytra. In males of this giant longhorn beetle (Cerambycidae), fore- leg length may reach 150 mm, nearly twice that of females of comparable body size. Dimorphism also occurs in the shape of the foreleg tibia which is nearly straight in females and exhibits varying de- grees of curvature in males (Fig. l). To investigate the factors influencing sexual dimorphism in this species, we carried out a threepart study encom- passing behavior, morphology, and the relationship between male size and mating success. To our knowledge, this is the first published report on mat- ing and agonistic behavior in A. longimanus.
METHODS Harlequin beetle larvae undergo development with- in decaying trees, exclusively in the families Morace-
ae and Apocynaceae (Du$ 1960, pers. obs.) After 4 to 12 mo (Du@ 1960, pers. obs.), adults emerge from their pupal chambers and fly in search of suitable trees for oviposition. In this study, we lo- cated beetles on both “emergence” and “oviposi- tion” trees. Oviposition trees may be either recently fallen trees or branches, or decaying sections of standing, live trees, i.e., trees which are mosaics of living and dead tissue. On mosaic trees, eclosion of adult beetles from old, dead tissue may occur con- currently with oviposition on living or newly necrotic regions. Observations of fighting, mating, and ovi- position were made exclusively on oviposition trees. These behaviors were never seen occurring on old emergence trees.
STUDY POPULATIONS.-The research was carried out in lowland tropical forest in the Parque Nacional Soberania, Panama, and along the Piste du Kaw,

88 Zeh, Zeh, and Tavakilian
French Guiana 600
E 500-
-400- E E: O 3 300- cd 2 pc 200- -4 o
pc E 100-
0-
50
a) A 40
s
2
8 20 2 10 E z o
2 30
pc 4J
k
9
S O N D J F M A M J J A
M o
Panama 500 1
400
300
200
100
O
40 1
30
20
10
O
n t h FIGURE 2. Seasonality of precipitation and harlequin beetle abundance in French Guiana and Panama (precipitation data from Sabatier E19831 and Windsor 119901). To facilitate direct comparison, months are arranged beginning with the first month of the dry season (September in French Guiana and January in Panama).
50 km southeast of Cayenne, French Guiana. In Panama, 117 beetles were found primarily on Ficus spp. (F. insipida Willd., and other species uniden- tified due to absence of fruit and leaves). Fout beetles were taken from a Brosimum alicastram (Pitt.). In French Guiana, 38 harlequins were ob- served on Puruhancornia fusciculata (Poiret), and *
on one unidentified tree. In Panama, beetles were measured and observed
on naturally occurring, fallen, or decaying trees. Because of the scaraty of trees attractive to beetles, the study was conducted over a three-year period
from December 1987 to December 1990. Most of the beetles (87%) were found in a 3 km-long, 200 m-wide tract of forest along the C25B trail in the Parque Naaonal Soberania. Beginning in May 1988, we conducted systematic searches of the C25B site. These surveys were performed at least twice per month for 14 mo and once per month for the remainder of the study. Once located, trees were monitored twice daily until beetle activity ceased. Sixteen beetles were taken from other sites within the Park. To provide comparative data on an ama- zonian population of A. longimanus, one month of
n fi 134- A A Pani B
9 108- Q
v
M
IJ 8 2 ~
56
30 27
El
FIGURE 3. Har: by gender and POF Inset specifies the s Females: Tibia anlj
intensive field WE
in French Guian; ama site in havin forest, and in tl season (Fig. 2). metric study we along access trai lective logging. t lected by G. Ta of a comprehen French Guiana
BEHAVIOR AND h
in interactions %
removed from t ment. Foreleg fe elytra length (E measured (k O. 5 amanian beetles examined for p Zeh, in press i
returned to the angle of the dis 3B) from proje analyze geogra1 sexual dimorpl ANOVA (with

Harlequin Beetle Sexual Selection 89
S O N D
,ma (precipitation ranged beginning
1990. Most of j km-long, 200 z5B trail in the lg in May 1988, I the C25B site. least twice per month for the tted, trees were activity ceased. her sites within Lata on an ama- r, one month of
160
n fl 134 E W
9 Q 3
$
M
c;l
rl Q, k
l oa
82
56
30
A 0 fiench Guiana ? o Panama8
37 47 Elytra Length (mm)
B
-.o.. Female -A- Male
15 35 55 75
Tibia Length (mm)
FIGURE 3. Harlequin beetle dometry. A. Relationship between elytra length and foreleg length (femur + tibia) by gender and population. B. Relationship between foretibia length and curvature angle in Panamanian population. Inset specifies the angle measured. Least-squares regression results: Males: Tibia angle = 0.35(TL) - 5.83, r = 0.91; Females: Tibia angle = 0.26(TL) - 4.12, r = 0-60.
intensive field work in March 1990 was carried out in French Guiana. This region differs from the Pan- ama site in having primary as opposed to secondary forest, and in the timing and severity of its dry season (Fig. 2). Beetles for behavior and morpho- metric study were found on recently felled trees along access trails in an area of low intensity, se- lective logging. Seasonal abundance data were col- lected by G. TavakiIian from various sites as part of a comprehensive study of the cerambycids of French Guiana (pers. obs.).
BEHAVIOR AND MORPHOLOGY.-Whell not involved in interactions with other beetles, individuals were removed from the tree for marking and measure- ment. Foreleg femur length (FL), tibia length (TL.), elytra length (EL) and antenna length (AL) were measured (k0.5 mm), using Vernier calipers. Pan- amanian beetles were photographed. Beetles were examined for phoretic pseudoscorpions (Zeh and Zeh, in press a), numbered on the thorax, and returned to the tree. We measured the curvature angle of the distal end of the tibia (see inset Fig. 3B) from projected slides of 91 individuals. To analyze geographic variation in size and extent of sexual dimorphism, we performed a two-way ANOVA (with gender and location as factors) on
In-transformed data for each trait. This analysis was also conducted on principal component scores 1 and 2 (PC1 and PC2) computed from the covariance matrix of' In-transformed pooled site and gender data (Bookstein et al. 1985). Allometric relation- ships were quantified from separate In-transformed data regressions of AL, FL, TL and total foreleg length (LEGL = FL + TL) on elytra length (see Huxley 1932); and from normalized trait factor loadings on PC1 derived from principal components analyses by site and gender (Bookstein e t al. 1985).
Most observations of mating, oviposition, and guarding behavior were made under low light con- ditions at dawn or dusk. Nighttime observations were made under red light. Male mating success was scored by monitoring the number of females with which each male copulated. When females were present on a tree, males were allocated a score of one (paired with female) or zero (unpaired). Males observed in the absence of females were counted as missing values. Sexual selection on each trait was estimated using the fimess-regression ap- proach of Lande and Arnold (1983) and the non- paramerric cubic spline procedure of Schluter (1988). EL, FL, and TL trait values were measured for all 59 males included in the analysis, but only 49 individuals were measured for AL. Values were log-

. . , !. , , ~
’I
90 Zeh, Zeh, and Tavakilian
transformed and standardized to a mean zero, vari- ance one, so that selection differentials and gradients represented standardized values. Mating success was scaled to mean 1.0 by dividing each individual’s mating success by the overall mean (Lande & Arnold 1983).
To investigate the relationship between male morphology and fighting ability, we staged 20 fights in the field, involving 27 males (statistical degrees of freedom based on number of males, not number of fights). For each encounter, we placed two males equidistant (a25 un) from an unpaired female on a freshly fallen tree and observed interactions. Fight durations and outcomes were recorded, and a paired t-test of winner’s minus loser’s trait value was cal- culated for each trait. The effect of relative and absolute size on fight duration was examined using simple and multiple linear regression on ln-trans- formed data. Statistical analyses were performed using SAS (SAS 1988) and BMDP (Dixon 1985).
RESULTS POPULATION coMPmrsoNs.-h both French Guiana and Panama there was a strong assoaation between the onset of the rainy season and a peak in A. longimanus abundance (Fig. 2). The first 3 mo of the wet season accounted for 50.7 percent of the harlequins found in French Guiana (N = 209) and 71.9 percent of those found in Panama (N = 121). The greater seasonal variation in beetle abundance in Panama is associated with lower m u a l preup- itation (2611 mm vs 3846 mm) and a longer dry season (4 vs 3 mo).
In this study, we did not attempt to quantify geographic variation in coloration, which is best accomplished by spectrographic analysis of live spec- imens (J. A. Endler, pers. com.) . Nonetheless, the French Guianan beetles were easily distinguished by their bright crimson markings compared to the faded orange of Panamanian individuals. In a “blind” test, one investigator (JAZ) was able to idenufy, from color slides, the source population for all of 15 randomly selected harlequin beetles. By contrast, for all four length measurements, the pop- ulations exhibited near uniformity in mean values, level of dimorphism, and allometry, as discussed below.
MomioMmICS.-Mdes and females did not differ significantly in elytra length (F,,, , , = 1.33, P = 0.250) but there was pronounced dimorphism in antenna length = 22.58, P < 0.001), femur length = 63.02, P < 0.001) and tibia length
(F,,,,, = 55.00, P < 0.001). None of the traits exhibited significant geographic variation in mean value (maximum F = 0.62, mini” P = 0.432) or in extent of sexual dimorphism, which was mea- sured by the interaction between location and gender in the two-way ANOVA (maximum F = 1.08, minimum P = 0.301). The multivariate analysis revealed a similar pattern of strong sexual, but weak geographic differentiation in morphology. PC1 rep- resents general size (all factor loadings positive), and accounted for 95.2 percent of trait variance. PC2 (bipolar factor loadings) accounted for 3.7 percent of variance. Males and females differed significantly in mean value for both components (PC1: F,,,,, = 40.59, P < 0.001; PC2: F,~, , , = 68.83, P < 0.00 1). Neither component exhibited significant between-population difference in means (PC1: FI,,,, =0.29,P=0.593;PC2:F,~12,=0.01, P=O.917) or in extent of sexual dimorphism (PC1: =
Levine’s F-test on In-transformed data indicated that males exhibited greater size-scaled variability in all sexually dimorphic traits (TL. F = 42.33, P < 0,001; FL: F = 28.40, P < 0.001; AL: F = 7.38, P = 0.008). There was no significant sexual merence in elytra length variance ( F = 2.20, P = 0.140).
Both univariate and multivariate analyses dem- onstrated that sexual dimorphism is primarily a consequence of stronger positive allometry in males (,) than in females (,), particularly in the forelegs (Fig. 3A). Univariate allometric coeffiaents (&SE) were as follows: AL, = 0.96 k 0.06, ALM = 1.20 -C 0.04; = 1.21 -i- 0.05, FL, = 2.01 -t 0.04;
0.93, P = 0.337; PC2: F1.125 = 0.09, P = 0.770).
‘I& = 1.17 k 0.08, ” L M = 2.19 -i- 0.05; LEGL, = 1.20 * 0.05, E G L M = 2.11 k 0.04. Multi- variate allometric coefficients exhibited a similar pat- tern in which male tibia length was most positively allometric (Table 1).
When male and female tibia lengths were stan- dardized, using an analysis of covariance (AN- COVA), sexual dimorphism was shown to also exist in the angle of tibia curvature (least squares mean,
Although both sexes exhibited a strong increase in tibia angle with inaeasing length, the relationship was more pronounced in males (Fig. 3B).
= 7.49, least squares mean, = 8.62, P = 0.04).
BEHAvIOR.-The 32-month survey of the 60 ha C25B site yielded 8 active host trees and 103 har- lequin beetles. Three of the trees had fallen within 48 h of being discovered. In these cases and on two of the three trees in French Guiana, we were able to monitor activity from the outset of the tree’s
period of amactivt that females were oviposition sites: occurred on the maining 23 perce mosaic trees or on during the preced was positively cor visiting beetles ar which beetle mat 4).
Host trees UI viscid, milky sap fly mainly during within hours afte the copious q u a leased upon i m ~ attracting beetles investigated, ger trees does appew pers. obs.). Ovil underside of rm most heavily sha
Male compel is intense and in into fierce, prot begin when two ing their heads f tennae back and abdomens, causi two males then stretched in appa 5A). Interaction the smaller male butting (Fig. 5B sideways in oppc each has the cur his opponent’s two combatants
. TABLE 1. Trai
term score
- Trait 3
EL 44.d FL 30.( TL 31.f Al. 103.’
PC 1 PC2
le of the traits iation in mean un P = 0.432) {hich was mea- tion and gender im F-_= 1.08, variate analysis &mal, but weak )logy. PC1 rep- s positive), and variance. PC2
for 3.7 percent ed significantly
= 68.83, P < ited significant
31, P = 0.917)
9 , P = 0.770). 1 data indicated aled variability F = 42.33, P
001; AL: F = ;nifiCant sexual F = 2.20, P =
(PC1: F,,125 =
ans (PC1: F1J2,
(PC1: FI,,,, =
: analyses dem- is primarily a
lmeuy in males in the forelegs &ients (&SE)
= 2.01 k 0.04; I- 0.05; LEGL,
:d a similar pat- most positively
gths were stan- variance (AN- lwn to also exist squares mean,
52, P = 0.04). ong increase in he relationship :. 3B).
5, ALM = 1.20
t 0.04. Multi-
of the 60 ha
.d fallen within ses and on two , we were able It of the tree’s
s and 103 ha-
Harlequin Beetle Sexual Selection 91
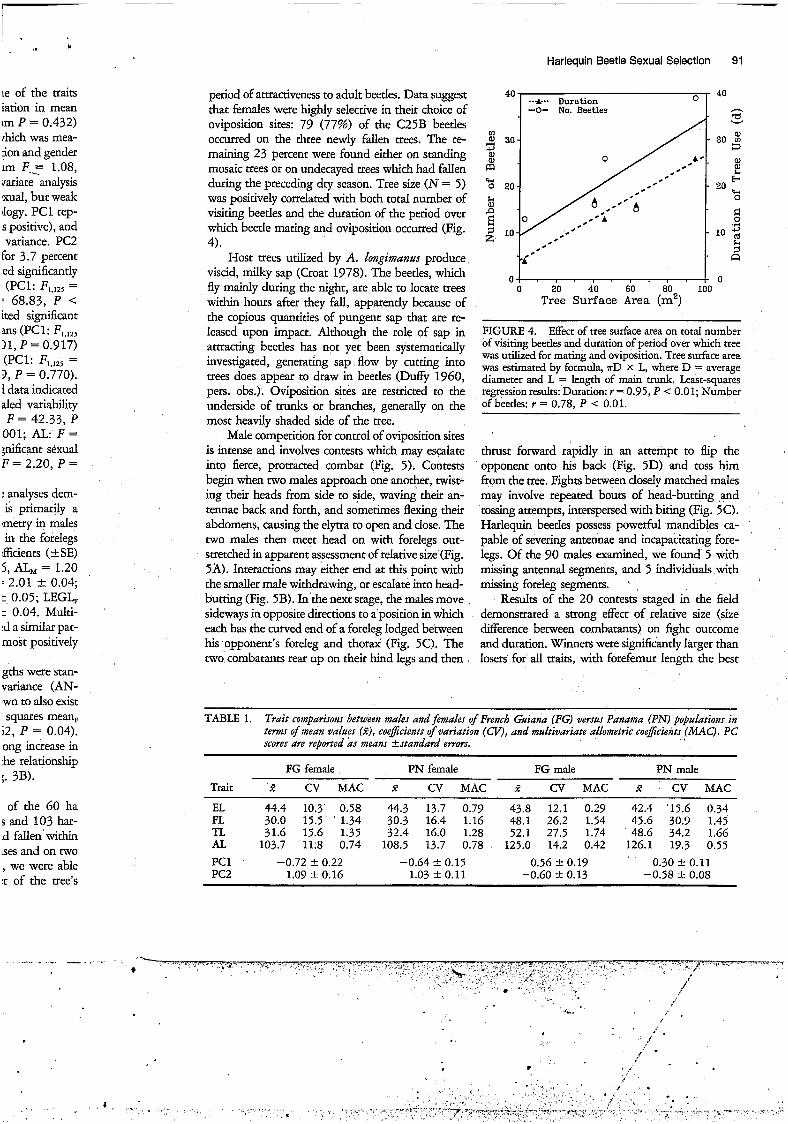
period of attractiveness to adult beetles. Data suggest that females were highly selective in their choice of oviposition sites: 79 (77%) of the C25B beetles occurred on the three newly fallen trees. The re- maining 23 percent were found either on standing mosaic trees or on undecayed trees which had fallen during the preceding dry season. Tree size (N = 5) was positively correlated with both total number of visiting beetles and the duration of the period over which beetle mating and oviposition occurred (Fig. 4).
Host trees utilized by A. longimanus produce viscid, milky sap (Croat 1978). The beetles, which fly mainly during the night, are able to locate trees within hours after they fall, apparently because of the copious quantities of pungent sap that are re- leased upon impact, Although the role of sap in attracting beetles has not yet been systematically investigated, generating sap flow by cutting into trees does appear to draw in beetles (Dufi 1960, pers. obs.). Oviposition sites are restricted to the underside of trunks or branches, generally on the most heavily shaded side of the tree.
Male competition for control of oviposition sites is intense and involves contests which may escalate into fierce, protracted combat (Fig. 5). Contests begin when two males approach one another, twist- ing their heads from side to side, waving their an- tennae back and forth, and sometimes flexing their abdomens, causing the elytra to open and dose. The two males then meet head on with forelegs out- stretched in apparent assessment of relative size (Fig. 5A). Interactions may either end at this point with the smaller male withdrawing, or escalate into head- butting (Fig. 5B). In the next stage, the males move sideways in opposite directions to a position in which each has the curved end of a foreleg lodged between his opponent’s foreleg and thorax (Fig. 5C). The two combatants rear up on their hind legs and then
40 I --.A--.. Duration
O O 20 40 60 60 100
Tree Surface Area (m’)
FIGURE 4. Effect of tree surface area on total number of visiting beetles and duration of period over which tree was utilized for mating and oviposition. Tree surface area was estimated by formula, ?rD x L, where D = average diameter and L = length of main trunk. Least-squares regressionresults: Duration: r = 0.95, P < 0.01; Number of beetles: r = 0.78. P < 0.01.
thrust forward rapidly in an attempt to flip the opponent onto his back (Fig. 5D) and toss him from the tree. Fights between closely matched males may involve repeated bouts of head-butting and tossing attempts, interspersed with biting (Fig. 5C). Harlequin beetles possess powerful mandibles ca- pable of severing antennae and incapacitating fore- legs. Of the 90 males examined, we found 5 with missing antennal segments, and 5 individuals with missing foreleg segments.
Results of the 20 contests staged in the field demonstrated a strong effect of relative size (size difference between combatants) on fight outcome and duration. Winners were significantly larger than losers for all traits, with forefemur length the best
TABLE 1. Trait comparisons between males and females of French Guiana (FG) versus Panama (PN) populations in terms of mean values (R), coeficients of variation (W), and multivariate allometric coeficients (MAC). PC scores are reported as means &standard errors.
FG female PN female FG male PN male Trait R cv MAC R CV MAC R CV MAC R CV MAC
EL 44.4 10.3 0.58 44.3 13.7 0.79 43.8 12.1 0.29 42.4 15.6 0.34 FL 30.0 15.5 1.34 30.3 16.4 1.16 48.1 26.2 1.54 45.6 30.9 1.45 TL 31.6 15.6 1.35 32.4 16.0 1.28 52.1 27.5 1.74 48.6 34.2 1.66 AL 103.7 11.8 0.74 108.5 13.7 0.78 125.0 14.2 0.42 126.1 19.3 0.55 PC 1 -0.72 t 0.22 -0.64 -C 0.15 0.56 C 0.19 0.30 C 0.11
-0.58 & 0.08 PC2 1.09 rt 0.16 1.03 t 0.11 -0.60 C 0.13

' . .. , . . .
" * "
Ø' -.i . .
92 Zeh, Zeh, and Tavakilian
FIGURE 5. Male fight sequence. A. Opponents dose in, head on with forelegs outstretched. B. Head-butting. C. Male bites opponent's leg. Curved end of opponent's tibia is hooked around biting male's thorax. D. Male (top right) has used right foreleg to flip over opponent (bottom left).
predictor of fight outcome (FL t = 4.25, P < 0.001; TL. t = 3.84, P < 0.001; AL: t = 3.18, P c: 0.001; E L t = 2.92, P < 0.001). Fight duration varied between 20 sec and 34 min and increased as forefemur size difference between com- batants decreased (regression coefficient @] * SE = -5.45 _C 1.12, P < 0.001, t.2 = 0.567). Ab- solute size also affected fight duration. When size difference was held constant, small males fought for shorter periods than large males. This was dem- onstrated by a multiple regression of fight duration on 1) size difference and on 2) absolute size of smaller combatant @, = -2.07 +- 1.23; ß2 = 3.52 k 0.93; ? = 0.76).
Our observations of beetle behavior revealed a tendency for males to remain on newly fallen trees throughout daylight hours, with dominant males maintaining guarding positions over oviposition sites. I
Small males generally occupy peripheral locations on the tree. Females are less likely to remain on fallen trees during daylight and usually 0y in after nightfall. When a female lands on a tree and en- counters a male, he initiates mating by positioning himself over the female and inserting his aedeagus into her ovipositor (Fig. 6A). We observed no cases
in which the female rejected the male, even when small individuals intercepted females en route to oviposition sites ( N = 3). Copulation continues for 3 to 5 min during which time the female may move around, apparently assessing oviposition site quality by chewing into the bark. The male then withdraws his aedeagus but continues to straddle the female, maintaining contact by massaging her elytra with his maxillary and labial palpi. The male remains in this guarding position as the female chews an ovi- position hole (Fig. 6B). Excavation time ranges from 10 to 60 min, after which the female insetts her ovipositor and deposits a single egg. She then rotates 180" (as does the guarding male) and uses her mandibles to pack excavated wood fibers over the egg. Copulation then resumes. The cycle of copu- lation, excavation, and oviposition may continue for several hours, leaving a characteristic line of pits which penetrare the bark. The pits may be circular, crescent-shaped or efiptical and vary in maximum length or diameter between 15 and 40 mm.
Male mating success was found to be signifi- cantly positively correlated with size across all traits. The Lande & Arnold (1983) selection differentials +- SE (linear regression coefficients of mating success

, L
ead-butting. C. :. D. Male (top
le, even when :s en route to i continues for lale may move ion site quality hen withdraws Ue the female, ler elytra with lale remains in chews an ovi- ne ranges from ale inserts her he then rotates and uses her
fibers over the cyde of copu- ay continue for ìc line of pits lay be circular, 7 in maximum 40 mm.
I to be signifi- Kross all traits. 3n differentials mating success
Harlequin Beetle Sexual Selection 93
FIGURE 6. in search of oviposition site. B. Female (left) chews oviposition pit while male stands guard over female.
Mating and oviposition, A. Male (top) inserts aedeagus into female's ovipositor as she "tastes" bark
_ r .
I

94 Zeh, Zeh, and Tavakilian
5.0 I 4.0
3.0
m m u o
2.0
$ 1.0
/ 4 I I I ) I ,
.4 4.6 4.8 5.0 !
.2 Ln (Antenna Length (mm)) Ln (Elytra Length (mm)) 4
3.0
$
2
.d 2.0
cd d
1.0
0.0
3.3 3.5 3.7 3.9
3*0*
3 .O 3.5 4.0 4.5 2.75 3.25 3.75 4.25 4
L n (Tarsus Length (mm)) Ln (Femur Length (mm)) I
FIGURE 7. Mating success curves for four male morphological traits calculated using the cubic spline technique. Dotted lines indicate standard error confidence limits which were generated by resampling original data with 30 repeated bootstraps (see Schluter 1988).
on trait values) are as follows: FL = 0.663 * 0.188; AL = 0.654 -C 0.200; EL = 0.647 $ 0.189; TL = 0.566 k O. 194. Traits were highly intercorrelated with correlation coefficients ranging from 0.96 to 0.99. With such extremely high correlations, it was not possible to identify the direct targets of sexual selection (all gradients not significantly greater than zero; see Mitchell-Olds & Shaw 1987). Because ,
harlequin beetle mating success was not normally distributed, we also used the nonparametric cubic spline fitness function estimation procedure (Schlu- ter 1988). Results were consistent with the selection differentials and revealed that for three traits (EL, FL and TL) mating success peaked just below max- imum trait value (Fig. 7).
DISCUSSION The morphological and behavioral findings reported here demonstrate that sexual dimorphism in A. Jon- gimanus is expressed in foreleg traits important in male competition for scarce oviposition sites, and is a consequence of sexual differences in allometry. In combat, males use their elongated, curved forelegs is hooked levers with which they attempt to toss the opponent from the tree. In this coleopteran version of jujitsu, foreleg length is the best predictor of fight outcome. Male forelegs lengthen dispro- portionately with increasing body size, and the angle of tibial curvature exhibits similar positive allom- etry. These morphological relationships are reflected
. '
in male behavic are similarly siz the prolonged c Taken in conjun males intercepti tence of an alti Thornhill & Al
Fighting su increased matin positive relatior lection different the highly corre of this "multici 1987), none ol cant and there the so-called d Arnold 1983). bmed with hig taken to indici overall measur character (e.g., of why the tra suggest that, ir does directly ta ever, effective 1
well require a resulting in t k being masked I male phenotyl lometric relatic of this dimoq 19601, provid on forelegs in 1
species. Extent of
patterns of se: able uniformi despite large c dimate. Such : flow (Wright difference ben istence of geog doubt on thi phoretic data 1
doscorpion w elytra of harle in press a, b) The French G the Panama I
LlTERATUl BEIER, M. 19 BOOKSTEIN, F. 1

Harlequin Beetle Sexual Selection 95
.) ,
;/r , -
I
3.9 4.1
b"
pline technique. al data with 30
dings reported iism in A. lon- : important in ln sites, and is I allometry. In w e d forelegs rempt to toss
in male behavioral tactics. Even when opponents are similarly sized, small males do not engage in the prolonged combat typical of large individuals. Taken in conjunction with our observations of small males intercepting females, this suggests the exis- tence of an alternative male mating strategy (see Thornhill & Alcock 1983).
Fighting success appears to be responsible for increased mating success. Our data show a strong positive relationship (positive, highly significant se- lection differentials) between mating success and all the highly correlated measures of male size. Because of this "multicollinearity" (Mitchell-Olds & Shaw 1987), none of the selection gradients was signifi- cant and therefore it was not possible to identify the so-called direct targets of selection (Lande & Arnold 1983). Strong selection differentials com- bined with high trait intercorrelation are generally taken to indicate that selection is acting on some overall measure of size and not on any particular character (e.g., Grant 1987). This begs the question of why the traits are so highly intercorrelated. We suggest that, in the case of A. longimanus, selection does directly target traits of the male forelegs. How- ever, effective utilization of elongated forelegs may well require a corresponding increase in body size, resulting in the direct targets of sexual selection being masked by selection to maintain an integrated male phenotype (Crespi 1990). Our results on al- lometric relationships, together with the uniqueness of this dimorphism in the Cerambycidae (Dufi 1960), provide evidence for strong, direct selection on forelegs in the recent evolutionary history of this species.
Extent of sexual dimorphism, mean sizes, and patterns of seasonal abundance exhibited remark- able uniformity in Panama and French Guiana, despite large differences in host tree utilization and dimate. Such similarity could be attributable to gene flow (Wright 1931). However, the marked color difference between the two populations and the ex- istence of geographic barriers (Eisenberg 1989) cast doubt on this explanation. Comparative electro- phoretic data on Cordylocbemes scorpioides, the pseu- doscorpion which disperses exclusively under the elytra of harlequin beetles (Beier 1948, Zeh & Zeh, in press a, b), also make this explanation unlikely. The French Guiana population is distinguished from the Panama population by a nearly fixed allele dif-
ference at the LDH locus (Zeh & Zeh, pers. obs.). We hypothesize that the two populations exhibit strong similarity because sexual selection is suffi- ciently intense to override any effects of differing .ecological factors.
Harlequin beetles display a pervasive but poorly understood pattern characteristic of highly sexually dimorphic species: extreme variance in male mor- phology (e.g., Eberhard 1983, Grimaldi 1987, Zeh & Zeh, in press c). This pattern seems to contradict the theory that strong selection should act to erode genetic variability (Fisher 1930). The argument that high variability represents purely environmental ef- fects acting on the phenotype (e.g., Zeh & Zeh 1988) does not adequately explain the patterns of variability documented in A. longimanus. While variance for the sexually monomorphic trait (EL) did not vary between the sexes, males were signif- icantly more variable than females in all the sexually dimorphic traits.
Recent research has linked the maintenance of genetic variability to selection which fluctuates in space and time (Hairston & Dillon 1990). We propose that there may be a similar effect of varying
, environments on selection in harlequin beetles. In this study, we demonstrated a significant benefit of
, large male size in terms of fighting and mating success. However, our results are based primarily on observations of beetles at high density on the prime oviposition habitats provided by newly fallen trees. Little is known of the relationship between male morphology and mating success in other con- texts, e.g., when females oviposit in the canopies of standing mosaic trees. Further research is required to establish whether the relationship between male size/fighting ability and mating success holds in these environments or whether the key to mating success lies in an alternative ability, such as the capacity to locate females.
ACKNOWLEDGMENTS
The study was supported by a Smithsonian Postdoctoral Fellowship to DWZ. We thank the Panamanian Instituto Nacional de Recursos Naturales Renovables (INREN- ARE) for permission to carry out this research, G. Cremers (ORSTOM) for host tree identification in French Guiana, and T. Mitchell-Olds and an anonymous reviewer for helpful comments on the manuscript. We are epecially grateful to J.-M. and O. Baloup for logistical support.
is coleopteran best predictor gthen dispro- and the angle
ositive allom- 1s are reflected
LITERATURE CITED BEIER, M. 1948. Phoresie und Phagophilie bei Pseudoscorpionen. Österreich. Zool. Z. 1: 441-497. BOOKSTEIN, F. L., B. CHERNOFF, R. L. ELDER, J. M. HUMPHRIES, G. R. SMITH, AND R. E. Smuss. 1985. Morphometrics
. .

96 Zeh, Zeh, and Tavakilian
in evolutionary biology: the geometry of size and shape change with examples from fishes. Acad. Nat. Sci. Philadelphia Spec. Publ. 15.
BRADBURY, J. W., AND M. B. ANDERSSON (Eds.). 1987, Sexual selection: testing the alternatives. J. Wiley and Sons, Ltd., Chichester, England.
CHEVERUD, J. M., M. M. Dow, AND W. LEUTENEGGER. 1985. The quantitative assessment of phylogenetic constraints in comparative analyses: sexual dimorphism in body weight among primates. Evolution 39: 1335-1351.
CONNER; J. 1988. Field measurements of natural and sexual selection in the fungus beetle, Boliiotberus cornutus- Evolution 42: 736-749.
CRESPI, B. J. 1990. Measuring the effect of natural selection on phenotypic interaction systems. Am. Nat. 135:
CROAT, T. B. 1978. Flora of Barro Colorado Island. Stanford University Press, Stanford, California. DARWIN, C. 1871. The descent of man and selection in relation to sex. John Murray (Publishers) Ltd., London,
DIXON, W. J. (Ed.). 1985. BMDP statistical software. University of California Press, Berkeley, California. DUFFY, E. A. J. 1960. A monograph of the immature stages of neotropical timber beetles. Brit. Mus. Nat. Hist.,
EBERHARD, W. G. 1980. Horned beetles. Sci. Am. 242: 166-182.
32-47.
England.
London, England.
. 1983. Behavior of adult bottle brush weevils (Rhinostomus barbirostris) (Coleoptera: Curculionidae). Rev. Biol. Trop. 3 1: 233-244.
EISENBERG, J. F, 1989. Mammals of the neotropics, volume 1, the northern neotropics. University of Chicago Press, Chicago, Illinois.
EMLEN, S. T., AND L. W. ORING. 1977. Ecology, sexual selection, and the evolution of mating systems. Science
FISHER, R. A. 1930. The genetical theory of natural selection. Clarendon Press, Oxford, England. GRANT, P. R. 1987. Ecology and evolution of Darwin's finches. Princeton University Press, Princeton, New Jersey. GRIMALDI, D. A. 1987. Phylogenetics and taxonomy of Zygotbrica (Diptera: Drosophilidae). Bull. Am. Mus. Nat.
Hist. 186(2)1: 268. HAIRSTON, N. G., AND T. A. DILLON. 1990. Fluctuating selection and response in a population of freshwater
copepods. Evolution 4 4 1796-1805. HUXLEY, J. S. 1932. Problems of relative growth. Methuen, London, England. LANDE, R. 1980. Sexual dimorphism, sexual selection, and adaptation in polygenic characters. Evolution 34: 292-
197: 215-223.
307. . 1982. A quantitative genetic theory of life history evolution. Ecology 63: 607-615. , AND S. J. ARNOLD. 1983. The measurement of selection on correlated characters. Evolution 37: 1210- 1226.
MITCHELL-OLDS, T., AND R. G. SHAW. 1987. Regression analysis of natural selection: statistical inference and biological interpretation. Evolution 41: 1149-1 161.
OTTE, D., AND K. STAYMAN. 1979. Beetle homs: some patterns in functional morphology. In M. S. Blum, and M. A. Blum (Eds.). Sexual selection and reproductive competition in insects. Academic Press, New York, New York.
SABATIER, D. 1983. Fructification et dissémination en forêt guyanaise. Thèse de Sime cycle. USTL Montpellier, France.
SAS INSTITUTE INC. 1988. SAS/STAT user's guide, release ¿.O3 edition. SAS Institute Inc., Cary, North Carolina. SCHLUTER, D. 1988. Estimating the form of natural selection on a quantitative character. Evolution 42: 849-861. THORNHILL, R., AND J. ALCOCK. 1983. The evolution of insect mating systems. Harvard University Press, Cambridge,
Massachusetts. TRIVERS, R. L. 1972. Parental investment and sexual selection. In B. Campbell (Ed.). Sexual selection and the
descent of man, 1871-1971, pp. 136-179. Aldine, Chicago. WINDSOR, D. 1990. Climate and moisture variability in a tropical forest: long-term records from Barro Colorado
Island, Panama. Smithson. Contrib. Earth Sci., No. 29. WRIGHT, S. 1931. Evolution in mendelian populations. Genetics 16: 97-159. ZEH, D. W. 1987a. Aggression, density and sexual dimorphism in chemetid pseudoscorpions (Arachnida: Pseu-
doscorpionida). Evolution 41: 1072-1087. . 1987b. Life history consequences of sexual dimorphism in a chemetid pseudoscorpion. Ecology 68: 1495- 1501. , AND J. A. ZEH. 1988. Condition-dependent sex omaments and field tests of sexual-selection theory. Am. Nat. 132: 454-459. , AND - , In press (a). Dispersal-generated sexual selection in a beetle-riding pseudoscorpion. Behav. Ecol. Sociobiol. , AND - . In press (b). On the function of harlequin beetle-riding in the pseudoscorpion Cordylocbernes scorpioides (Pseudoscorpionida: Chernetidae). J. Arachnol.
ZEH, J. A., AND D. W. ZEH. In press (c). Are sexually-selected traits reliable species characters?: implications of intra-brood variability in Semeiocbemes armìger (Balzan) (Pseudoscorpionida: Chernetidae). Bull. Br. Arachnol. soc.
f
BlOTROPlCA 2
NOTES
Herbivory i
Most herbivore number and di there. Leaf dan of leaf damage Dirzo 1987, E impossibility tc have been usec 1982). Wht ( canopy. A new consisting of a dirigible ballo0 (Hallé & Blanc
We tried t we could find Central Americ zone, is writtei of different for were lower t h z Amazonian reg forest canopies Bongers, pers. forest are relau
In the pres Guyana, indu 53'04'W) and October 1989 was used to rc 105 m long w of trees, vines, using a telesco height of bem the following I
80-90 and 9( life forms and
Overall da between life fi (Peat Saut) ar
Landsberg and 15 p e r a canopy CO"
values. They r relatively scarc due to drough accounting for was not menu sampling peric Unfortunately also exists that in the tropics.
commonly, wl





























