Embed Size (px)
Citation preview

1
Sequence analysis of fluoroquinolone resistance associated genes gyrA and gyrB in clinical 1
Mycobacterium tuberculosis isolates from suspected multidrug-resistant tuberculosis 2
patients in New Delhi, India 3
4
Authors: Ritu Singhala, b, Paul R. Reynoldsc, d, Jamie Marolaa, L. Elaine Eppersone, Jyoti Arorab, 5
Rohit Sarinf, Vithal Prasad Myneedu b, #, Michael Stronge, g, Max Salfinger a, h, # 6
a Mycobacteriology and Pharmacokinetics Laboratories, National Jewish Health, Denver, CO, 7
USA; b National Reference Laboratory and Centre of Excellence (WHO), Department of 8
Microbiology, National Institute of Tuberculosis and Respiratory Diseases, New Delhi, India; c 9
Department of Academic Affairs, National Jewish Health; d Department of Pediatrics, National 10
Jewish Health; e Center for Genes, Environment, and Health, National Jewish Health; f 11
Department of Tuberculosis and Chest Diseases, National Institute of Tuberculosis and 12
Respiratory Diseases, New Delhi, India; g Computational Bioscience Program, University of 13
Colorado, Denver, CO, USA; h Department of Medicine, National Jewish Health 14
15
Running Title: Sequence analysis of gyrA and gyrB genes 16
#Correspondence to: 17
Max Salfinger ([email protected]) and 18
Vithal Prasad Myneedu ([email protected]) 19
20
21
22
JCM Accepted Manuscript Posted Online 22 June 2016J. Clin. Microbiol. doi:10.1128/JCM.00670-16Copyright © 2016, American Society for Microbiology. All Rights Reserved.
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

2
Address: 23
Max Salfinger, MD 24
Mycobacteriology & Pharmacokinetics Laboratories, 25
National Jewish Health, K412a 26
1400 Jackson Street, 27
Denver, CO 80206, USA 28
29
Dr. Vithal Prasad Myneedu 30
Head of Laboratory, National Reference Laboratory and Centre of Excellence (World Health 31
Organization, WHO), 32
Department of Microbiology, National Institute of Tuberculosis and Respiratory Diseases, 33
Sri Aurobindo Marg, 34
New Delhi, 110030, India 35
36
37
38
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

3
ABSTRACT 39
Fluoroquinolones (FQ) are broad-spectrum antibiotics recommended for treatment of 40
multidrug-resistant tuberculosis (MDR-TB) patients. FQ resistance, caused by mutations in the 41
gyrA and gyrB genes of Mycobacterium tuberculosis (MTB), is increasingly reported world-42
wide; however, information on mutations occurring in strains from the Indian sub-continent is 43
scarce. Hence, in this study we aimed to characterize mutations in the gyrA and gyrB genes of 44
acid-fast bacilli (AFB) smear positive sediments or MTB isolates from AFB smear negative TB 45
suspects from India. A total of 152 samples from suspected MDR-TB patients were included in 46
the study. Of these, 146 strains detected were characterized by sequencing of the gyrA and 47
gyrB genes. The extracted DNA was subjected to successive amplification using a nested PCR 48
protocol, followed by sequencing. 49
A total of 27 mutations were observed in 25 strains in the gyrA gene, while no mutations 50
were observed in the gyrB gene. The most common mutation occurred at amino-acid position 51
94 (13/27; 48.1%) of which the D94G mutation was the most prevalent. The gyrA mutations 52
were significantly associated with rifampin (RIF) resistant-TB patients in comparison with non-53
RIF resistant patients. Heterozygosity was seen in 4/27 (14.8%) mutations, suggesting the 54
occurrence of mixed populations with different antimicrobial susceptibilities. 55
A high rate of FQ-resistant mutations (17.1%), were obtained among the isolates of TB 56
patients suspected of MDR-TB. These observations emphasize the need for accurate and 57
rapid molecular tests for the detection of FQ-resistant mutations at the time of MDR-TB 58
diagnosis. 59
60
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

4
INTRODUCTION 61
Treatment of multidrug-resistant tuberculosis (MDR-TB) patients, with strains resistant to 62
rifampin (RIF) and isoniazid (INH), is further complicated with the presence of fluoroquinolone 63
(FQ) resistance, due to prolonged, limited, and expensive treatment options (1,2). A recent 64
meta-analysis on response to treatment of 6,724 MDR-TB patients from 26 centers revealed 65
that treatment success of patients with MDR-TB was 64%, while success of patients with 66
MDR-TB plus FQ resistance was only 48% (3). This clearly indicates the need for routine 67
molecular screening for FQ resistance-associated molecular markers so that treatment of 68
such patients can be better optimized and without delay. Many of such cases develop 69
extensively drug-resistant tuberculosis (XDR-TB), defined as MDR-TB plus resistance to any 70
one FQ and any of the aminoglycosides/cyclic peptides (A-CP) (4). This poses an even more 71
serious threat in TB management since only 40% of XDR-TB patients have successful 72
treatment outcomes (2). Previous studies suggest that 5.4% (95% CI: 3.4–7.5) of MDR-TB 73
cases may actually be XDR-TB (2). 74
FQ are oral, antibacterial agents that have activity against Mycobacterium tuberculosis (MTB) 75
(5, 6). Hence, FQs are recommended for the treatment of MDR-TB patients and patients with 76
intolerance to RIF (2). FQ are also being evaluated in conjunction with first-line regimens for 77
newly diagnosed drug susceptible TB (7). Since FQ are broad-spectrum antibiotics, these 78
antimicrobials are often over-prescribed by clinicians for diverse infections (8). In many 79
resource-limited countries, FQ are readily available as over-the-counter medications, 80
propagating their misuse (9, 10). Such indiscriminate use has contributed to the increasing 81
emergence of FQ-resistant MTB world-wide, as well as India (2, 11). Newer FQ-based 82
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

5
regimens are being explored, which necessitates the knowledge of FQ resistance data from 83
high TB burden countries (12). 84
Traditionally, culture-based methods have been used for determining antimicrobial 85
susceptibility, which usually takes several weeks. Thus, a patient harboring a drug resistant 86
MTB strain may continue to spread undetected in the community (13). Molecular methods, 87
however, have decreased the time for detection of drug resistance, especially for MDR-TB 88
(14). Among these methods, reverse hybridization or line probe assays (LPA) have been 89
implemented as part of the national TB programs in high burden countries for rapid detection 90
of MDR-TB (15). These assays are directed towards the detection of the most common 91
mutations for rifampin resistance (14). In contrast to RIF resistance tests, fewer molecular 92
tests are available for diagnosing FQ resistance, and the existing methods are limited by lower 93
level of association with phenotypic drug resistance (16, 17, 18). 94
The principal target of FQ drugs in MTB is the DNA gyrase protein complex, which catalyzes 95
the ATP-dependent introduction of negative supercoils into closed circular DNA molecules. 96
The gyrA and gyrB genes encode the two subunits of DNA gyrase, which form the catalytically 97
active GyrA2 GyrB2 complex. Resistance to FQ in MTB is primarily caused by mutations in the 98
quinolone resistance-determining region (QRDR), a conserved 320bp region within the gyrA-99
gene. FQ resistance has also been shown to be associated, but less commonly attributable, to 100
mutations in a 375 bp region of the gyrB gene (5). 101
Mutations in the gyrA and gyrB genes of clinical isolates of MTB have been described in many 102
populations and studies (5, 17, 18, 19, 20, 21, 22, 23); however, the information on mutations 103
in the gyrA-gyrB genes from the Indian sub-continent is scarce (24, 25), although India has 104
one of the highest percentages of MDR-TB cases in the world (2). There is evidence that 105
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

6
different mutations are associated with different levels of phenotypic resistance, or different 106
minimum inhibitory concentrations (MIC), to FQs; moxifloxacin (MOX) has been suggested to 107
be more active with lower MICs for all isolates, compared to ofloxacin (OFL) (23, 25, 26). Thus, 108
knowledge of the specific mutation may help reveal the level of FQ resistance, which can in 109
turn inform decisions regarding the selection of OFL or higher generation FQs like MOX or 110
levofloxacin (LVX). This information can also be used to determine whether phenotypic 111
testing should be extended from OFL to MOX and LVX. In the present study, we therefore 112
aimed to characterize mutations in gyrA and gyrB genes in MTB isolates from the Indian 113
subcontinent in MDR-TB suspected patients. 114
115
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

7
MATERIALS AND METHODS 116
Sample selection. Samples from presumptive MDR-TB patients from districts within New 117
Delhi, sent to the Mycobacteriology Laboratory in New Delhi, India over a period from 118
November 1 to November 30, 2013, were analyzed in this study. The presumptive MDR-TB 119
patients were identified as per the national guidelines (Programmatic management of drug-120
resistant tuberculosis; PMDT), which includes smear positive failures on category I or category 121
II regimen, history of contact with MDR-TB, all re-treatment cases (smear positive and smear 122
negative) and HIV-TB (27). Category I regimen is prescribed for new patients (intensive phase 123
for 2 months and consists of INH, RIF, pyrazinamide (PZA) and ethambutol (EMB) given under 124
observation thrice weekly, followed by continuation phase of 4 months of INH and RIF given 125
thrice weekly) (27). Category II regimen is prescribed for TB patients who have had a history 126
of at least one month of anti-tuberculosis treatment. Relapses, treatment after default and 127
failures are treated with a category II regimen (Intensive phase for 2 months and consists of 128
INH, RIF, PZA, EMB and Streptomycin (SM) given under observation thrice weekly, followed by 129
INH, RIF, PZA and EMB given under observation thrice weekly for 1 month and then followed 130
by continuation phase of 5 months of INH, Rif and EMB given thrice weekly (27). 131
Sample testing. Testing was performed using Genotype MTBDRplus (Hain Lifescience, 132
Nehren, Germany), a line probe assay (LPA), and a WHO endorsed molecular test. This 133
diagnostic tool may be used for detection of MDR-TB (both RIF & INH) directly on smear 134
positive sputum samples or on culture isolates in case of smear negative samples. Smear 135
positive sputum samples were identified as MDR-TB directly by LPA, with a turn-around time 136
of approximately 5 days. Smear negative sputum were inoculated for culture, and if positive 137
for MTB, subjected to LPA (27) according to PMDT. The site of testing was at the 138
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

8
Mycobacteriology Laboratory, located in the Department of Microbiology at the National 139
Institute of Tuberculosis and Respiratory Diseases (NITRD) in New Delhi, India, and is one of 140
the National Reference Laboratories (NRL) under the Revised National Tuberculosis Control 141
Program (RNTCP) in India and a WHO center of excellence. 142
Processing of samples. The samples were processed by a standard N-acetyl-L-cysteine - 143
sodium hydroxide (NALC-NaOH) method (28), with a final concentration of 1.0% NaOH, (28). 144
The samples were stained for acid-fast bacilli (AFB) by Ziehl-Neelsen (ZN) method. All 145
processed AFB smear-positive samples were considered for DNA extraction and LPA, and all 146
AFB-smear negative samples were inoculated into BACTEC MGIT (Becton Dickinson, Sparks, 147
MD, USA) tubes, and when positive for MTB were considered for LPA. The cultures were 148
incubated for 42 days before being considered negative. 149
DNA extraction. For DNA extraction, 500 μL of NALC-NaOH processed samples or 1 mL of a 150
culture from BACTEC MGIT tubes were used. The DNA extraction was performed using 151
Genolyse version 1.0 kit (Hain Lifescience, Nehren, Germany). Supernatants containing DNA 152
was transferred into a fresh tube and stored at - 20˚C. 153
Genotype MTBDRplus assay. The amplification and hybridization using GenoType MTBDRplus 154
kit 2.0 were performed as per the manufacturer’s instructions and previously described (29). 155
For quality control of each assay, M. tuberculosis H37Rv was used as a positive control and 156
molecular grade water as negative control was put in every run. 157
Primer Design for Sequencing of FQ-resistance regions of gyrA and gyrB. The molecular 158
amplification, sequencing, and polymorphism detection was carried out at National Jewish 159
Health, Denver, CO. A consensus sequence of the target regions was created using 160
alignments of approximately 10 to 15 MTB gyrA or gyrB sequences present in the National 161
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

9
Center for Biotechnology Information (NCBI) database (http://www.ncbi.nlm.nih.gov/). Two 162
pairs of primers each for gyrA and gyrB were designed using Primer 3 163
(http://bioinfo.ut.ee/primer3-0.4.0/primer3/). The gyrA primer sets were gyrARS-164
Forward(F)/gyrAPRR-Reverse(R):5’-CAGCTACATCGACTATGCGA-3’/5’-165
ATTTCCCTCAGCATCTCCA-3’; 358 bp amplicon. And a nested set: gyrAPRR-F/gyrARS-R: 5’-166
GACTATGCGATGAGCGTGAT-3’/5’-GGGCTTCGGTGTACCTCAT-3’; 310bp amplicon. The two 167
primer sets designed for gyrB were gyrBPRR-F/gyrBPRR-R: 5’-AACAGCTGACCCACTGGTTT-168
3’/5’-CGCTGCCACTTGAGTTTGTA-3’; 556 bp amplicon and gyrBPRS-F/gyrBPRS-R:5’-169
CGCAAGTCCGAACTGTATGTCGTAG-3’/5’-GTTGTGCCAAAAACACATGC-3’; 346 bp amplicon. 170
Annealing temperature and magnesium concentrations were optimized prior to sample 171
analysis. 172
Amplification. Initial amplification of gyrA and gyrB using 40 cycles and with a single pair of 173
primer sets for each gene gave poor amplification, likely due to low DNA quantity in the 174
Genolyse-extracted DNA. Secondary purification and concentration of DNA also did not yield 175
consistent amplification. Thus, nested amplification was performed with primer sets as 176
described above. 177
gyrA amplification. PCR was performed with the Advantage-GC Genomic LA Polymerase Mix 178
(Clontech #639153, Mountain View, CA, USA) using 400 µM dNTP and 400 nM primers. The 179
first amplification employed gyrAPRR-F/RS-R in a 10 µL reaction mix at the following 180
conditions: 1 X 2 min at 94oC, 15 X [15s at 95°C, 20s at 60°C, and 75s at 72°C] 1 X 2 min at 181
72°C, hold at 4oC. The second amplification was performed with gyrARS-F/gyrAPRR-R for 35 182
cycles at the same conditions listed above, using 2 µL of the first PCR reaction mix as 183
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

10
template. Supplementary Figure S1 illustrates the positioning of forward and reverse primers 184
for gyrA and gyrB. 185
gyrB amplification: The first PCR reaction was performed with Amplitaq Gold (ThermoFisher 186
Scientific, Carlsbad, CA) using the gyrBPRR primer set with 2.5 mM Mg++, 200uM dNTP and 187
200 nM primers at the following conditions: 1 X 95oC 10min; 20 X [95oC 15s, 59.1oC 20s, 72oC 188
75s], 72oC 2min, 4oC hold. The second amplification used primer set gyrBRS at 2 mM Mg++, 189
200 uM dNTP, 200 nM primers as follows: 1 X 95oC 10min; 40 X [95oC 15s, 60oC 20s, 72oC 190
75s], 72oC 2min, 4oC hold. Supplementary Figure S1 illustrates the positioning of forward and 191
reverse primers for gyrA and gyrB. 192
Amplicon purification and sequencing. The amplicons were prepared for sequencing by 193
treatment with Exonuclease I and Shrimp Alkaline Phosphatase (Exo-SAP). Ten µL of 1X PCR 194
buffer/ 0.375 mM Mg++ containing 0.05 U SAP and 2 U Exo-nuclease I was added to each PCR 195
reaction. The mixture was incubated at 37°C for 30 minutes followed by 95°C for 10 minutes. 196
Sequencing was performed with Big Dye 3.1 (ABI, Applied Biosystems, Foster City, CA). Each 197
10 µL reaction mix contained 1.5 µL 5X buffer, 0.875 µL BDX64 (MCLAB, San Francisco, CA), 198
0.35 µL Big Dye 3.1, 1 µL 5 µM primer, and 2 µL Exo/SAP-treated amplicon. Cycling conditions 199
were 2 min at 96°C, 35 cycles as follows: 10s at 96°C, 5s at 52°C, and 2min at 60°C. 200
Sequencing products were purified with spin columns (EdgeBio, Gaithersburg, MD) according 201
to manufacturer’s directions, dried, and loaded onto an ABI 3730 Genome Analyzer (Applied 202
Biosystems, Foster City, CA). 203
Sequence analysis. Sequence traces were imported into a BioNumerics database (Applied 204
Maths, Austin, TX), and trimmed if necessary. Sequences were exported in FASTA format. The 205
genetic polymorphisms of gyrA and gyrB were compared to the reference sequence from M. 206
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

11
tuberculosis strain H37Rv (GenBank accession number NC_000962/ L27512). The 207
polymorphisms obtained were annotated as per the M. tuberculosis genome numbering 208
system. 209
Structure Analysis. Gyrase A mutations were mapped onto the X-ray crystal structure of the 210
M. tuberculosis gyrase A protein and color coded using the PyMol structure visualization 211
software package (https://www.pymol.org/ ). The coordinates of the gyrase A X-ray crystal 212
structure were obtained from the Protein Data Bank 213
(http://www.rcsb.org/pdb/home/home.do ), PDB reference code 3ILW . 214
215
216
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

12
RESULTS 217
A total of 152 samples from patients suspected of MDR-TB from the Delhi region of North 218
India were considered for the Genotype MTBDRplus assay. History of anti-tuberculosis drugs 219
under category I regimen or category II regimen or history of contact was collected for these 220
patients. Overall, 104/152 (68.4%) patients were at the start of retreatment/ category II 221
regimen due to default, incomplete treatment, treatment failures or relapse. 16/152 (10.5%) 222
patients were failures of a new TB/ category I regimen. Thirty out of 152 (19.7%) patients 223
were failures of retreatment/ category II regimen. Out of 152, 1 patient had a history of 224
contact with MDR-TB and one was HIV positive. The LPA was performed on 16 MTB isolates 225
from cultures of AFB smear-negative sputum samples, whereas remaining LPA was performed 226
directly on the AFB smear-positive sputum samples. 227
The 179 bp region between 148 to 327 bp of the gyrA gene, encoding amino-acids 49 to 109, 228
was studied for the identification of mutations. Six strains yielded poor gyrA sequences even 229
on repeat sequencing and were excluded from the analysis. A total of 27 mutations in 25 230
strains (17%) were obtained in 146 strains studied for gyrA gene, in which two strains had 231
more than one mutation. Table 1 provides the clinical details, susceptibility to INH and RIF 232
and mutations observed in gyrA. The substitution at amino acid 94 was most common 233
mutation (13/27; 48.1%). Mutations observed include A281G D94G (8/ 27; 29.6%) and C269T 234
A90V (8/ 27; 29.6%), followed by A281C D94A (2/27; 7.4%), G280T D94Y (2/27; 7.4%), C269Y 235
A90V* (2/27; 7.4%), G280A D94N (1/27;3.7%), A283G S95A (1/27;3.7%), T271C S91P 236
(1/27;3.7%), and A281R D94G* (2/27; 7.4%). Strains ID12, and ID18 had double mutations 237
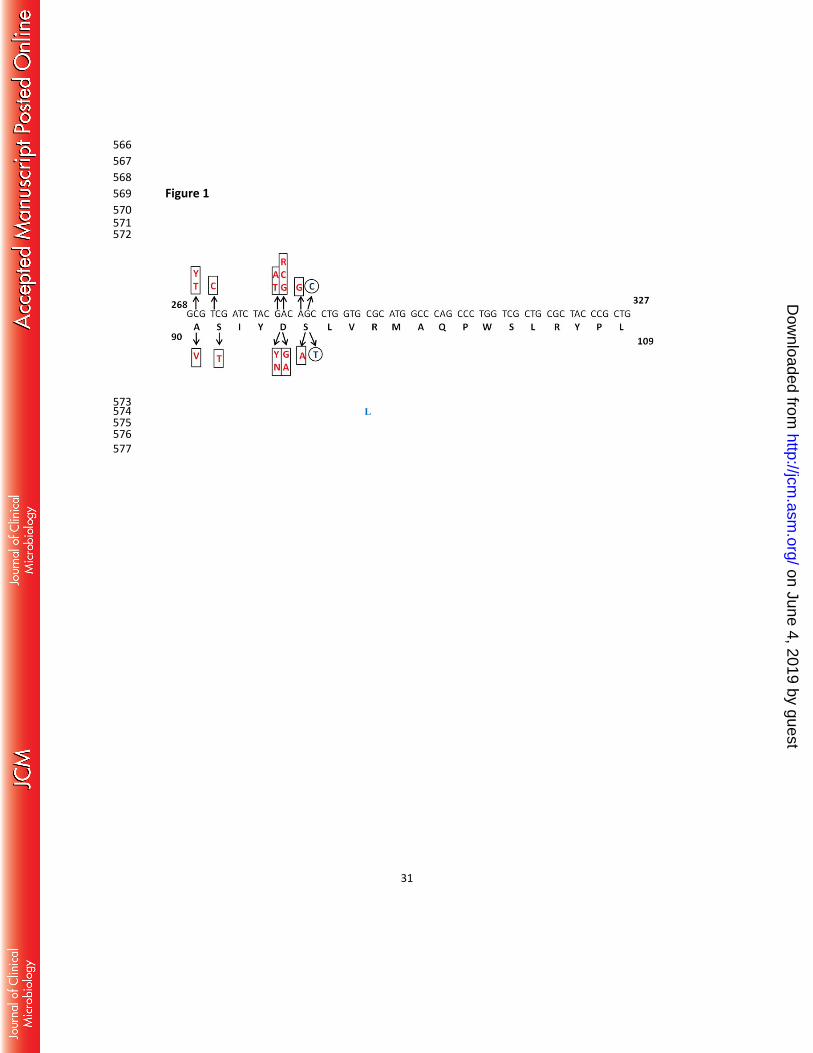
A90V/D94G and A90V/D94A respectively. The location of the mutations in the codons and 238
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

13
respective amino-acid changes in the primary sequence is depicted in Figure 1, and in the 239
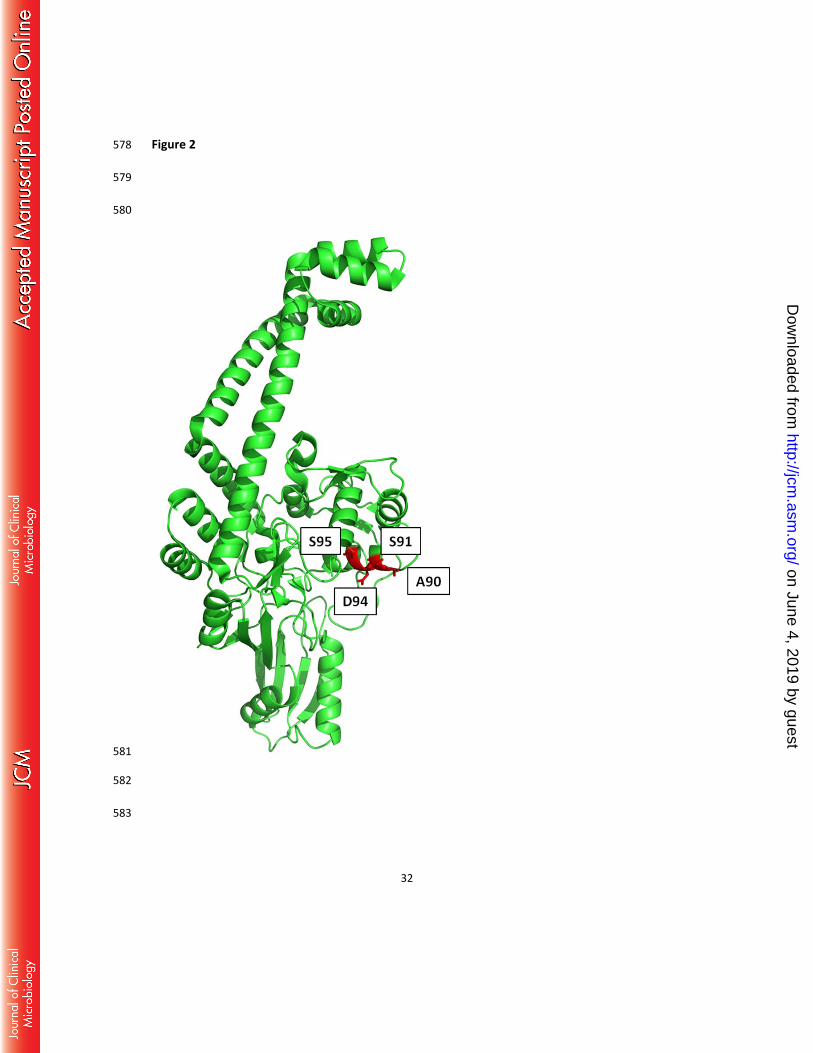
gyrA protein structure in Figure 2. 240
Co-existence of wild-type and mutated base (Heterozygosity) was seen in 4/27 (14.8%) 241
mutations which included strains ID12, ID24 and ID25 of which two resistance-associated 242
mutations were seen in ID12 detailed in Table 1, Supplementary Figure S2. Polymorphism 243
G284C, which encodes a S95T polymorphism, as compared to the reference H37Rv gyrA 244
sequence, was found in 138/146 (94.5%) of the strains. This S95T polymorphism is a common 245
lineage specific polymorphism, not likely to be involved in fluoroquinolone resistance. 246
Among the patients with the isolates having gyrA mutations, 22/25 (88%) were AFB smear-247
positive and 3/25 (12%) were AFB smear-negative when identified as MDR-TB suspects. Of 248
these isolates, the majority (24/25; 96%) were previously treated pulmonary TB cases, and 249
one (1/25; 4%) case was failure of category I regimen. Four of previously treated (4/24; 250
16.7%) cases were failures of category II regimen. 251
Of 146 MDR-TB suspects, 21/146 (14.4%) were found to be MDR-TB, 10/146 (6.8%) RIF mono-252
resistant, 5/146 (3.4%) INH mono-resistant, and 110/146 (75.3%) susceptible to both 253
antibiotics based on the well characterized mutations. In the MDR-TB strains, 14/21 (66.7%) 254
had FQ resistance mutations in the gyrA gene, and of the RIF monoresistant strains 2/10 255
(20%) had FQ resistance mutations in the gyrA gene. In the non-MDR cases, FQ resistance was 256
observed in 10/115 (8.7%) strains, the difference being statistically significant (< 0.0001). 257
Similarly, the RIF resistant strains were found in 16/25 (64%) strains with gyrA mutations in 258
comparison with 15/121 (12.4%) strains without gyrA mutations. This difference was also 259
found to be statistically significant (< 0.0001). The 418 bp region between 1229 to 1647 bp of 260
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

14
gyrB gene encoding the amino-acids 410 to 543 for 109 strains was also investigated, and no 261
mutations were found in any of the samples. 262
DISCUSSION 263
This study aimed at the detection of mutations in gyrA and gyrB genes for FQ resistance. The 264
study population included TB patients from North India suspected to be MDR-TB patients. 265
Most of the patients had a history of irregular or defaulted TB treatment using the first-line 266
anti-tuberculosis agents like RIF, INH, ethambutol, and pyrazinamide. It has been shown 267
earlier that such patients are much more likely to develop MDR-TB. The treatment regimen 268
involving such patients includes FQ or aminoglycosides (2). 269
In the present study, 17.1% of patients as described above were found to harbor FQ 270
mutations. In India, with a population of over a billion individuals, this has significant 271
implications, due to the high potential proportion of FQ resistance among TB and MDR-TB 272
suspects. This is likely due to FQ’s being over-prescribed in certain regions of the world, 273
especially in many resource-limited countries, where they are readily available over the 274
counter (8). In the Indian subcontinent, FQ resistance for MTB has increased from 3% of 275
laboratory cultures in 1996 to 35% in 2004 in Mumbai and from 17.4% in 2005 to 42.9% in 276
2009 in Pakistan (9, 10). FQ resistance is known to be associated with poor treatment 277
outcomes among MDR-TB patients (30), and the increasing prevalence of FQ resistance TB 278
isolates is of great concern. 279
Resistance to FQ in MTB is caused mainly by the mutations in the QRDR region, a highly 280
conserved region in the gyrA protein, located at the N-terminal portion extending from amino 281
acid 74 to amino acid 113. FQ resistance in MTB is less common due to mutations in the 282
second protein of the gyrase complex, gyrB, with QRDR mutations observed from codon 461 283
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

15
to 499 of gyrB (31). In the previous review of mutations by Maruri et al, 59% and 41% of 284
mutations were in gyrA and gyrB respectively. For gyrA mutations, 81% and 19% of mutations 285
were inside and outside the QRDR respectively. Whereas for gyrB mutations, 44% of 286
mutations were inside the QRDR region, 50% were outside the QRDR, and 1% were deletions 287
(31). 288
In the present study, mutations were found only in the gyrA gene, with substitutions at codon 289
94 in gyrA being the most common at 48.1%. The substitutions in codon 94 (37%) are the 290
most common as reviewed by Maruri et al (31). In another review by Avalos et al, across 18 291
countries, mutation data of 3,846 clinical isolates with known phenotypic resistance profiles 292
to FQ was studied. The most frequent gyrA mutation was D94G which ranged from 21-32% 293
(32). The relative frequency of mutations in the QRDR region reported from different regions 294
world-wide are detailed in table 2. D94G is reported as the most common codon 94 295
substitution in most reports, as seen in studies from Pakistan, China, The Philippines, 296
Thailand, Vietnam, Uzbekistan, Russia, Germany, France and USA (33, 34, 35, 36, 37, 38, 39, 297
40, 17, 41). All mutations at codon 94 have been associated with in-vitro resistance as seen in 298
earlier studies (17, 24, 33,36, 37, 40, 41) The codon 90 mutation, A90V has also been found in 299
high frequency with studies from India, The Philippines, China, and Belgium reporting it as the 300
most common (24, 35, 42, 23). The review by Avalos et al. found 13-20% A90V mutations 301
contributing to resistance (32). In the present study, both D94G and A90V were the most 302
commonly observed mutations. Interestingly, both these mutations have been reported in all 303
gyrA mutation studies. However, other amino acid 94 substitutions, including D94A, D94Y, 304
D94N and D94H have been reported with varying frequency from none to 17.6%, with a rare 305
D94F substitution reported only in one study (35). 306
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

16
The frequency of A90V mutations has been found to be less in some studies from Germany 307
(10.3%), and the USA (5.9%), suggesting that the frequency of mutations in a gene tend to 308
vary by geographic location (40, 41, 43). Two strains in the present study had double 309
mutations ID18; A90V/D94A and ID12; A90V/D94G. The presence of double mutations has 310
been related to high-level drug resistance, as reported previously (17, 31, 39, 40, 42). Aubrey 311
et al reported high-level drug resistance on a strain with A90V/D94G mutation, which had 312
very high MIC of >160 µg/mL (5). 313
In the present study, a novel mutation was found in gyrA in ID23; S95A; within the QRDR 314
region. Figure 2 demonstrates the X-ray crystal structure of the M. tuberculosis gyrase A 315
protein (Protein Data Bank code 3ILW) with amino acids at positions A90, S91, D94, S95 316
shown in red within the QRDR. Although the S95A mutation is in close proximity to well 317
characterized fluoroquinolone resistance mutation sites, including D94, further 318
microbiological confirmation would be necessary to assess the impact of this specific 319
mutation on fluoroquinolone resistance. 320
Hetero-resistance, defined as both wild type and a mutation at specific loci, was seen in 321
14.8% of FQ mutations identified by sequencing. Presence of hetero-resistant mutations 322
suggest, mixed susceptible and resistant bacteria in the same clinical sample. Two different 323
mechanisms for hetero-resistance have been postulated, co-existence of two genotypically 324
different strains i.e. susceptible and resistant, or a single strain evolving from susceptible and 325
resistant organisms. It has been shown earlier that infection with two different strains was 326
responsible for hetero-resistance in 71% of the patients and could serve as a quality indicator 327
for TB control programs in various countries (44). This occurrence may be present in higher 328
frequencies in hyper-endemic regions (15). The evolution of a susceptible strain into a 329
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

17
resistant strain could be the result of poor treatment in terms of dose and/or duration. Other 330
studies have also reported hetero-resistance with a frequency varying from 4 to 37.1% (17, 331
37, 39, 40). 332
An interesting observation in the QRDR region of gyrA of MTB is the presence of G to C 333
polymorphism at position 284; S95T as compared to the reference H37Rv gene. This encodes 334
either serine (in M. tuberculosis H37Rv and H37Ra) or threonine (in M. tuberculosis Erdman, 335
M. bovis BCG, M. africanum) (19). In the present study, this polymorphism was found in 336
94.5% isolates. The S95T polymorphism has not been implicated to FQ resistance since it has 337
been found in strains with MIC values to ofloxacin at less than the critical concentration, <2 338
µg/mL (6, 33, 35, 42). An exception was an earlier Indian study, which found S95T in 88.2% of 339
ofloxacin-resistant isolates with no other mutation in QRDR gyrA or gyrB (24). 340
We looked for gyrB mutations in the QRDR region, as several worldwide studies have 341
reported mutations associated with in vitro resistance such as D472H, R485C, D495N, D500N, 342
D500H, N510D, N533T, N538D, N538I, T539N, 546M (5, 23, 36, 45). However, in our study, we 343
did not find any gyrB mutations among our samples, which agrees with other studies 344
performed in Indian and in China (24, 34). 345
Interestingly, we found a significant association of MDR-TB with the gyrA mutations, in 346
comparison with strains without the gyrA mutations. Over half of the MDR-TB strains 347
examined in this study also had mutations in gyrA (66.7%). The rate of FQ resistance among 348
MDR-TB patients also has been found to be high in other countries in the region, The 349
Philippines; 51.4% (46), Taiwan; 22.2% (47), and in China (Shanghai); 25.1% (20). The rates 350
from countries in other continents are lower, with 4.1% in the United States and Canada (48) 351
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

18
and 4.3% in MDR-TB in Russia (49). It has been hypothesized that resistance to other anti-TB 352
drugs could potentially influence resistance to FQ (13). 353
In summary, our study revealed a high rate, 17.1%, of FQ mutations among the isolates of 354
Indian TB patients suspected of MDR-TB. The FQ mutations in sequence verified MDR strains 355
was 66.7% whereas, in non-MDR cases, it was 18.7%. All the mutations were found 356
exclusively in gyrA gene, with A90V and D94G being the most common. One novel mutation 357
S95A was also observed. The data strongly suggests for conducting expanded studies, 358
examining both phenotypic and genotypic data on a larger number of isolates from India and 359
surrounding countries. This would further help in developing, refining, and improving rapid 360
molecular diagnostic tests for determining resistance (50). This would also aid in the control 361
and identification of pre-XDR-TB and XDR-TB in a timelier and effective manner, preferably at 362
the time of detection of MDR-TB. 363
364
Acknowledgement 365
This work was supported by the NIH/FIC AIDS and TB International Training and Research 366
Program (D43TW001409) at NYU School of Medicine, Principle Investigator: Dr. Suman Laal 367
(PhD). Michael Strong acknowledges support from the Boettcher Foundation Webb-Waring 368
program. 369
370
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

19
REFERENCES 371
1. Centers for Disease Control and Prevention. 2006. Emergence of Mycobacterium 372
tuberculosis with extensive resistance to second-line drugs world-wide, 2000-2004. 373
Morb Mortal Wkly Rep MMWR 55: 301-305. 374
2. World Health Organization. 2011. Guidelines for the programmatic management of 375
drug-resistant tuberculosis. WHO/HTM/TB/2011.6. World Health Organization, 376
Geneva, Switzerland 377
3. Falzon D, Gandhi N, Migliori GB, Sotgiu G, Cox HS, Holtz TH, Hollm-Delgado MG, 378
Keshavjee S, DeRiemer K, Centis R, D'Ambrosio L, Lange CG, Bauer M, Menzies D; 379
Collaborative Group for Meta-Analysis of Individual Patient Data in MDR-TB. 2013. 380
Resistance to fluoroquinolones and second-line injectable drugs: impact on multidrug-381
resistant TB outcomes. Eur Respir J 42:156-168. 382
4. Sotgiu G, Ferrara G, Matteelli A, Richardson MD, Centis R, Ruesch G. 2009. 383
Epidemiology and clinical management of XDR-TB: a systematic review by TBNET. Eur 384
Respir J 33:871–881. 385
5. Aubry A, Veziris N, Cambau E, Truffort-Pernot C, Jarlier V, Fisher LM 2006. Novel 386
gyrase mutations in quinolone-resistant and hyper-susceptible clinical isolates of 387
Mycobacterium tuberculosis: functional analysis of mutant enzymes. Antimicrob 388
Agents Chemother 50:104–112. 389
6. Giannoni F, Iona E, Sementilli F, Brunori L, Pardini M, Migliori G. 2005. Evaluation of a 390
new line probe assay for rapid identification of gyrA mutations in Mycobacterium 391
tuberculosis. Antimicrob Agents Chemother 47:2928-2933. 392
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

20
7. Peloquin CA, Hadad DJ, Molino LPD, Palaci M, Boom WH, Dietze R, Johnson JL. 2008. 393
Population pharmacokinetics of levofloxacin, gatifloxacin, and moxifloxacin in adults 394
with pulmonary tuberculosis. Antimicrob Agents Chemother. 52:852–857 395
8. Dooley KE, Golub J, Goes FS, Merz WG, Sterling TR. 2002. Empiric treatment of 396
community acquired pneumonia with fluoroquinolones and delays in treatment of 397
tuberculosis. Clin Infect Dis 34:1607–1612. 398
9. Agrawal D, Udwadia ZF, Rodriguez C, Mehta A. 2009. Increasing incidence of 399
fluoroquinolone-resistant Mycobacterium tuberculosis in Mumbai, India. Int J Tuber 400
Lung Dis 13(1):79–83. 401
10. Jabeen K, Shakoor S, Chishti S, Ayaz A, Hasan R. 2011. Fluoroquinolone-resistant 402
Mycobacterium tuberculosis, Pakistan, 2005–2009. Emerg Infect Dis 17 (3): 564-565. 403
11. Devasia AR, Blackman A, May C, Eden S, Smith T, Hooper N. 2009. Fluoroquinolone 404
resistance in Mycobacterium tuberculosis: an assessment of MGIT 960, MODS and 405
nitrate reductase assay and fluoroquinolones cross resistance. J Antimicrob 406
Chemother 63:1173-1178. 407
12. Isaakidis P, Das M, Kumar AM, Peskett C, Khetarpal M, Bamne A, Adsul B, Manglani 408
M, Sachdeva KS, Parmar M, Kanchar A, Rewari BB, Deshpande A, Rodrigues C, Shetty 409
A, Rebello L, Saranchuk P. 2014. Alarming levels of drug-resistant tuberculosis in HIV-410
infected patients in metropolitan Mumbai, India. PLoS One 21;9(10):e110461. 411
13. Jabeen K, Shakoor S, Hasan R. 2015. Fluoroquinolone-resistant tuberculosis: 412
implications in settings with weak healthcare systems. Int J Infect Dis 32:118-123. 413
14. Ling DI, Zwerling AA, Pai M. 2008. GenoType MTBDR assays for the diagnosis of 414
multidrug-resistant tuberculosis: a meta-analysis. Eur Respir J 32:1165-1174. 415
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

21
15. Singhal R, Myneedu VP, Arora J, Singh N, Sah GC, Sarin R. 2014. Detection of drug 416
resistance and characterization of mutations in Mycobacterium tuberculosis isolates 417
from North-eastern states of India using GenoType MTBDRplus assay. Indian J Med 418
Res 140:501-506. 419
16. Ritter C, Lucke K, Sirgel FA, Warren RW, van Helden PD, Böttger EC, Bloemberg GV. 420
2014. Evaluation of the AID TB resistance line probe assay for rapid detection of 421
genetic alterations associated with drug resistance in Mycobacterium tuberculosis 422
strains. J Clin Microbiol. 52:940-946 423
17. Brossier F, Veziris N, Aubry A, Jarlier V, Sougakoff W. 2010. Detection by GenoType 424
MTBDRsl test of complex mechanisms of resistance to second-line drugs and 425
ethambutol in multidrug-resistant Mycobacterium tuberculosis complex isolates. J Clin 426
Microbiol 48:1683–1689. 427
18. Huang WL, Chi TL, Wu MH, Jou R. 2011. Performance assessment of the GenoType 428
MTBDRsl test and DNA sequencing for detection of second-line and ethambutol drug 429
resistance among patients infected with multidrug-resistant Mycobacterium 430
tuberculosis. J Clin Microbiol 49:2502–2508. 431
19. Takiff HE, Salazar L, Guerrero C, Philipp W, Huang WM, Kreiswirth B. 1994. Cloning 432
and nucleotide sequence of Mycobacterium tuberculosis gyrA and gyrB genes and 433
detection of quinolone resistance mutations. Antimicrob Agents Chemother 38:773–434
780. 435
20. Xu P, Li X, Zhao M, Gui X, DeRiemer K, Gagneux S, Mei J, Gao Q. 2009. Prevalence of 436
fluoroquinolone resistance among tuberculosis patients in Shanghai, China. 437
Antimicrob Agents Chemother 53(7):3170-3172. 438
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

22
21. Chang KC, Yew WW, Chan CY. 2010. Rapid assays for fluoroquinolone resistance in 439
Mycobacterium tuberculosis: a systematic review and meta-analysis. J Antimicrob 440
Chemother 65:1551–1561. 441
22. Campbell PJ, Morlock GP, Sikes RD, Dalton TL, Metchock B. 2011. Molecular 442
Detection of mutations associated with first- and second-line drug resistance 443
compared with conventional drug susceptibility testing of Mycobacterium 444
tuberculosis. Antimicrob Agents Chemother 55:2032–2041. 445
23. Groll AV, Martin A, Jureen P, Hoffner S, Vandamme P, Portaels F. 2009. 446
Fluoroquinolone resistance in Mycobacterium tuberculosis and mutations in gyrA and 447
gyrB. Antimicrob Agents Chemother 53(10): 4498–4500. 448
24. Siddiqi N, Shamim M, Hussain S, Choudhary RK, Ahmed N, Prachee. 2002. Molecular 449
characterization of multidrug-resistant isolates of Mycobacterium tuberculosis from 450
patients in North India. Antimicrob Agents Chemother 46:443–450. 451
25. Sulochana S, Narayanan S, Paramasivan CN, Suganthi C, Narayanan PR. 2007. 452
Analysis of fluoroquinolone resistance in clinical isolates of Mycobacterium 453
tuberculosis from India. J Chemother 19(2):166-171. 454
26. Lau RW, Ho PL, Kao RYT, Yew WW, Lau TCK, Cheng VCC, Yuen KY, Tsui SKW, Chen X, 455
Yam WC. 2011. Molecular characterization of fluoroquinolone resistance in 456
Mycobacterium tuberculosis: functional analysis of gyrA mutation at position 74. 457
Antimicrob Agents Chemother 55 (2): 608–614. 458
27. Revised National Tuberculosis Control Programme (RNTCP). 2012. Guidelines on 459
Programmatic Management of Drug Resistant TB (PMDT) in India. Central TB Division, 460
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

23
Directorate General of Health Services, Ministry of Health & Family Welfare, New 461
Delhi, India 462
28. Kent PT, Kubica GP. 1985. Public health mycobacteriology, a guide for level III 463
laboratory. Atlanta, GA: U.S. Department of Health and Human Services, U.S. 464
Government Printing Office, Washington, DC. 465
29. Albert H, Bwanga F, Mukkada S, Nyesiga B, Ademun JP, Lukyamuzi G. 2010. Rapid 466
screening of MDR-TB using molecular line probe assay is feasible in Uganda. BMC 467
Infect Dis 10(41):1-7. 468
30. Yew WW, Chan CK, Chau CH. 2000. Outcomes of patients with multidrug-resistant 469
pulmonary tuberculosis treated with ofloxacin/levofloxacin-containing regimens. 470
Chest 117:744–751. 471
31. Maruri F, Sterling TR, Kaiga AW, Blackman A, Mayer C, Cambau E, Aubrey A. 2012. A 472
systematic review of gyrase mutations associated with fluoroquinolone-resistant 473
Mycobacterium tuberculosis and a proposed gyrase numbering system. J Antimicrob 474
Chemother 67:819–831. 475
32. Avalos E, Catanzaro D, Catanzaro A, Ganiats T, Brodine S, Alcaraz J, Rodwell T. 2015. 476
Frequency and geographic distribution of gyrA and gyrB mutations associated with 477
fluoroquinolone resistance in clinical Mycobacterium tuberculosis isolates: A 478
systematic review. PLoS One 27;10(3):e0120470. 479
33. Ali A, Hasan R, Jabeen K, Jabeen N, Qadeer E, Hasan Z. 2011. Characterization of 480
mutations conferring extensive drug resistance to Mycobacterium tuberculosis isolates 481
in Pakistan. Antimicrob Agent Chemother 55(12):5654–5659. 482
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

24
34. Chena J, Chenb Z, Lia Y, Xiaa W, Chena X, Chena T, Zhouc L, Xua B, Xua S. 2012. 483
Characterization of gyrA and gyrB mutations and fluoroquinolone resistance in 484
Mycobacterium tuberculosis clinical isolates from Hubei Province, China. Braz J Infect 485
Dis 16(2):136-141. 486
35. Bravo LT, Tuohy MJ, Ang C. 2009. Pyrosequencing for rapid detection of 487
Mycobacterium tuberculosis resistance to rifampin, isoniazid, and fluoroquinolones. J 488
Clin Microbiol 47:3985–3990. 489
36. Pitaksajjakul P, Wongwit W, Punprasit W. 2005. Mutations in the gyrA and gyrB 490
genes of fluoroquinolone-resistant Mycobacterium tuberculosis from TB patients in 491
Thailand. Southeast Asian J Trop Med Public Health 4:228–237. 492
37. Kiet VS, Ngoc Lan NT, An DD, Dung NH, Hoa DV, Vinh Chau NV, Chinh NT, Farrar J, 493
Caws M. 2010. Evaluation of the MTBDRsl test for detection of second-line drug 494
resistance in Mycobacterium tuberculosis. J Clin Microbiol 48:2934–2939. 495
38. Feuerriegel S, Cox HS, Zarkua N. 2009. Sequence analyses of just four genes to detect 496
extensively drug-resistant Mycobacterium tuberculosis strains in multidrug-resistant 497
tuberculosis patients undergoing treatment. Antimicrob Agents Chemother 53:3353–498
3356. 499
39. Mokrousov I, Otten T, Manicheva O. 2008. Molecular characterization of ofloxacin-500
resistant Mycobacterium tuberculosis strains from Russia. Antimicrob Agents 501
Chemother 52:2937-2939. 502
40. Hillemann D, Rusch GS, Richter E. 2009. Feasibility of the GenoTypeMTBDRsl assay for 503
fluoroquinolone, amikacin-capreomycin, and ethambutol resistance testing of 504
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

25
Mycobacterium tuberculosis strains and clinical specimens. J Clin Microbiol 47:1767–505
1772. 506
41. Devasia R, Blackman A, Eden S, Li H, Maruri F, Shintani A, Alexander C, Kaiga A, 507
Stratton CW, Warkentin J, Tang YW, Sterling TR. 2012. High proportion of 508
fluoroquinolone-resistant Mycobacterium tuberculosis isolates with novel gyrase 509
polymorphisms and a gyrA region associated with fluoroquinolone susceptibility. J Clin 510
Microbiol 50:1390–1396. 511
42. Shi R, Zhang J, Li C. 2006. Emergence of ofloxacin resistance in Mycobacterium 512
tuberculosis clinical isolates from China as determined by gyrA mutation analysis using 513
denaturing high-pressure liquid chromatography and DNA sequencing. J Clin Microbiol 514
44:4566–4568 515
43. Bartfai Z, Somoskovi A, Kodmon C, Szabo N, Puskas E, Kosztolanyi L, Farago E, 516
Mester J, Parsons LM, Salfinger M. 2001. Molecular characterization of rifampin-517
resistant isolates of Mycobacterium tuberculosis from Hungary by DNA sequencing 518
and the line probe assay. J Clin Microbiol 39:3736-3739. 519
44. Thiel SH, Ingen JV, Feldmann K. 2009. Mechanisms of heteroresistance to isoniazid 520
and rifampin of Mycobacterium tuberculosis in Tashkent, Uzbekistan. Eur Respir J 521
33:368–374. 522
45. Malik S, Willby M, Sikes D, Oleg VT, James EP. 2012. New insights into 523
fluoroquinolone resistance in Mycobacterium tuberculosis: Functional genetic analysis 524
of gyrA and gyrB mutations. PLoS one 6(7) e39754. 525
46. Grimaldo ER, Tupasi TE, Rivera AB, Quelapio MI, Cardano RC, Derilo JO, Belen VA. 526
2001. Increased resistance to ciprofloxacin and ofloxacin in multidrug-resistant 527
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

26
Mycobacterium tuberculosis isolates from patients seen at a tertiary hospital in the 528
Philippines. Int J Tuberc Lung Dis 5(6):546-550. 529
47. Huang TS, Kunin CM, Shin-Jung LS, Chen YS, Tu HZ, Liu YC. 2005. Trends in 530
fluoroquinolone resistance of Mycobacterium tuberculosis complex in a Taiwanese 531
medical centre: 1995-2003. J Antimicrob Chemother 56(6):1058-1062. 532
48. Bozeman L, Burman W, Metchock B, Welch L, Weiner M. 2005. Fluoroquinolone 533
susceptibility among Mycobacterium tuberculosis isolates from the United States and 534
Canada. Clin Infect Dis 40(3):386-391. 535
49. Balabanova Y, Ruddy M, Hubb J, Yates M, Malomanova N, Fedorin I, Drobniewski F. 536
2005. Multidrug-resistant tuberculosis in Russia: Clinical characteristics, analysis of 537
second-line drug resistance and development of standardized therapy. Eur J Clin 538
Microbiol Infect Dis 24(2):136-139. 539
50. Somoskovi A, Salfinger M. 2015. The race is on to shorten the turnaround time for 540
diagnosis of multidrug-resistant tuberculosis. J Clin Microbiol 53(12):3715-3718. 541
542
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

27
Legends 543
544
Figure 1 545
Location of the mutations in the analyzed strains. Sequence polymorphisms likely to be 546
involved in fluoroquinolone resistance are indicated in the boxes above (nucleotide changes) 547
and below (amino acid changes) the primary sequence. A common lineage specific 548
polymorphism is indicated by the circles, and is not likely to be involved in resistance. 549
550
Figure 2 551
Location of the identified MTB gyrA mutations. Mutations identified in this study are 552
indicated in red on the gyrA protein structure, and cluster at the quinolone resistance-553
determining region. 554
555
556
557
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from
28
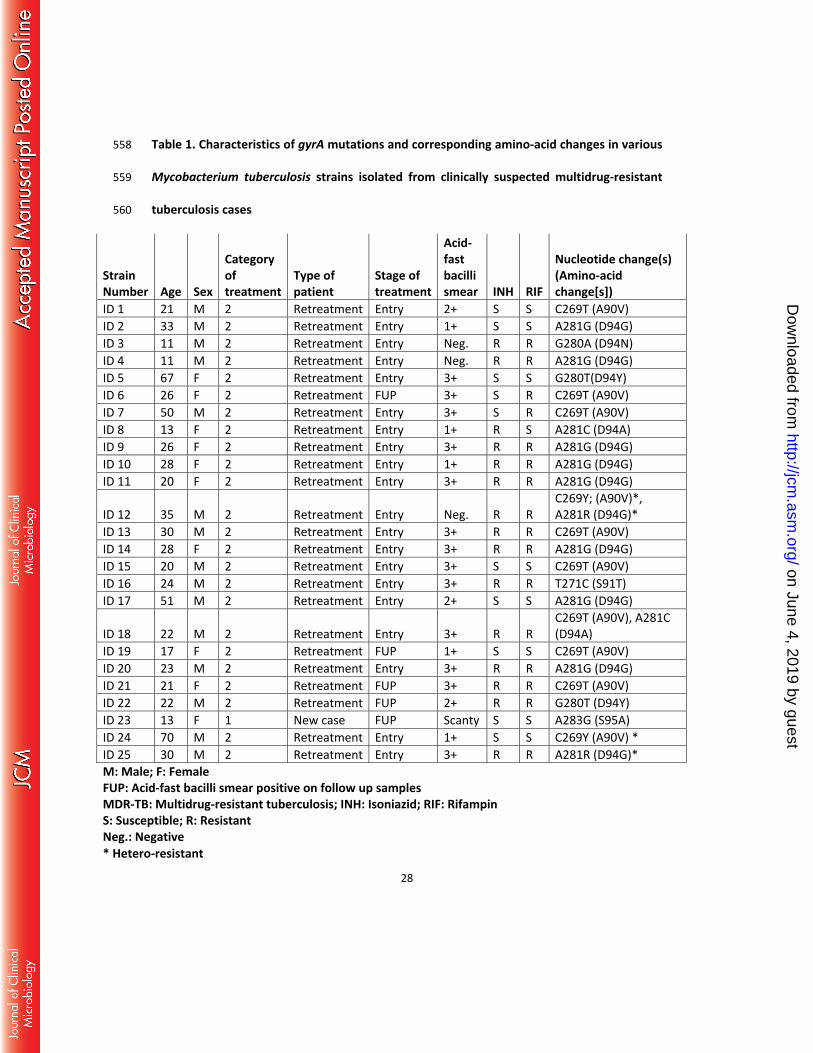
Table 1. Characteristics of gyrA mutations and corresponding amino-acid changes in various 558
Mycobacterium tuberculosis strains isolated from clinically suspected multidrug-resistant 559
tuberculosis cases 560
Strain Number
Age
Sex
Category of treatment
Type of patient
Stage of treatment
Acid-fast bacilli smear INH RIF
Nucleotide change(s) (Amino-acid change[s])
ID 1 21 M 2 Retreatment Entry 2+ S S C269T (A90V) ID 2 33 M 2 Retreatment Entry 1+ S S A281G (D94G) ID 3 11 M 2 Retreatment Entry Neg. R R G280A (D94N) ID 4 11 M 2 Retreatment Entry Neg. R R A281G (D94G) ID 5 67 F 2 Retreatment Entry 3+ S S G280T(D94Y) ID 6 26 F 2 Retreatment FUP 3+ S R C269T (A90V) ID 7 50 M 2 Retreatment Entry 3+ S R C269T (A90V) ID 8 13 F 2 Retreatment Entry 1+ R S A281C (D94A) ID 9 26 F 2 Retreatment Entry 3+ R R A281G (D94G) ID 10 28 F 2 Retreatment Entry 1+ R R A281G (D94G) ID 11 20 F 2 Retreatment Entry 3+ R R A281G (D94G)
ID 12 35 M 2 Retreatment Entry Neg. R R C269Y; (A90V)*, A281R (D94G)*
ID 13 30 M 2 Retreatment Entry 3+ R R C269T (A90V) ID 14 28 F 2 Retreatment Entry 3+ R R A281G (D94G) ID 15 20 M 2 Retreatment Entry 3+ S S C269T (A90V) ID 16 24 M 2 Retreatment Entry 3+ R R T271C (S91T) ID 17 51 M 2 Retreatment Entry 2+ S S A281G (D94G)
ID 18 22 M 2 Retreatment Entry 3+ R R C269T (A90V), A281C (D94A)
ID 19 17 F 2 Retreatment FUP 1+ S S C269T (A90V) ID 20 23 M 2 Retreatment Entry 3+ R R A281G (D94G) ID 21 21 F 2 Retreatment FUP 3+ R R C269T (A90V) ID 22 22 M 2 Retreatment FUP 2+ R R G280T (D94Y) ID 23 13 F 1 New case FUP Scanty S S A283G (S95A) ID 24 70 M 2 Retreatment Entry 1+ S S C269Y (A90V) * ID 25 30 M 2 Retreatment Entry 3+ R R A281R (D94G)* M: Male; F: Female FUP: Acid-fast bacilli smear positive on follow up samples MDR-TB: Multidrug-resistant tuberculosis; INH: Isoniazid; RIF: Rifampin S: Susceptible; R: Resistant Neg.: Negative * Hetero-resistant
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from
29
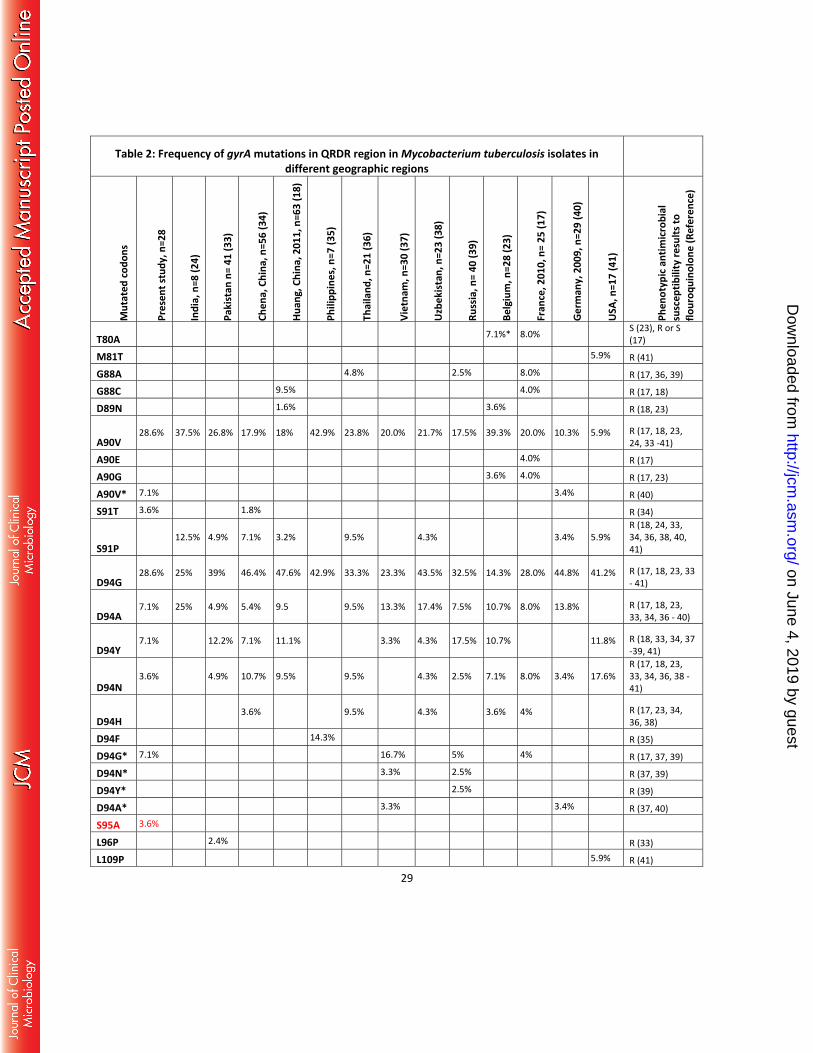
Table 2: Frequency of gyrA mutations in QRDR region in Mycobacterium tuberculosis isolates in different geographic regions
Mut
ated
codo
ns
Pres
ent s
tudy
, n=2
8
Indi
a, n
=8 (2
4)
Paki
stan
n=
41 (3
3)
Chen
a, C
hina
, n=5
6 (3
4)
Huan
g, C
hina
, 201
1, n
=63
(18)
Phili
ppin
es, n
=7 (3
5)
Thai
land
, n=2
1 (3
6)
Viet
nam
, n=3
0 (3
7)
Uzbe
kist
an, n
=23
(38)
Russ
ia, n
= 40
(39)
Belg
ium
, n=2
8 (2
3)
Fran
ce, 2
010,
n=
25 (1
7)
Germ
any,
200
9, n
=29
(40)
USA,
n=1
7 (4
1)
Phen
otyp
ic an
timicr
obia
l su
scep
tibili
ty re
sults
to
flour
oqui
nolo
ne (R
efer
ence
)
T80A 7.1%* 8.0% S (23), R or S (17)
M81T 5.9% R (41)
G88A 4.8% 2.5% 8.0% R (17, 36, 39)
G88C 9.5% 4.0% R (17, 18)
D89N 1.6% 3.6% R (18, 23)
A90V 28.6% 37.5% 26.8% 17.9% 18% 42.9% 23.8% 20.0% 21.7% 17.5% 39.3% 20.0% 10.3% 5.9% R (17, 18, 23,
24, 33 -41)
A90E 4.0% R (17)
A90G 3.6% 4.0% R (17, 23)
A90V* 7.1% 3.4% R (40)
S91T 3.6% 1.8% R (34)
S91P 12.5% 4.9% 7.1% 3.2% 9.5% 4.3% 3.4% 5.9%
R (18, 24, 33, 34, 36, 38, 40, 41)
D94G 28.6% 25% 39% 46.4% 47.6% 42.9% 33.3% 23.3% 43.5% 32.5% 14.3% 28.0% 44.8% 41.2% R (17, 18, 23, 33
- 41)
D94A 7.1% 25% 4.9% 5.4% 9.5 9.5% 13.3% 17.4% 7.5% 10.7% 8.0% 13.8% R (17, 18, 23,
33, 34, 36 - 40)
D94Y 7.1% 12.2% 7.1% 11.1% 3.3% 4.3% 17.5% 10.7% 11.8% R (18, 33, 34, 37
-39, 41)
D94N 3.6% 4.9% 10.7% 9.5% 9.5% 4.3% 2.5% 7.1% 8.0% 3.4% 17.6%
R (17, 18, 23, 33, 34, 36, 38 - 41)
D94H 3.6% 9.5% 4.3% 3.6% 4% R (17, 23, 34,
36, 38)
D94F 14.3% R (35)
D94G* 7.1% 16.7% 5% 4% R (17, 37, 39)
D94N* 3.3% 2.5% R (37, 39)
D94Y* 2.5% R (39)
D94A* 3.3% 3.4% R (37, 40)
S95A 3.6%
L96P 2.4% R (33)
L109P 5.9% R (41)
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from
30
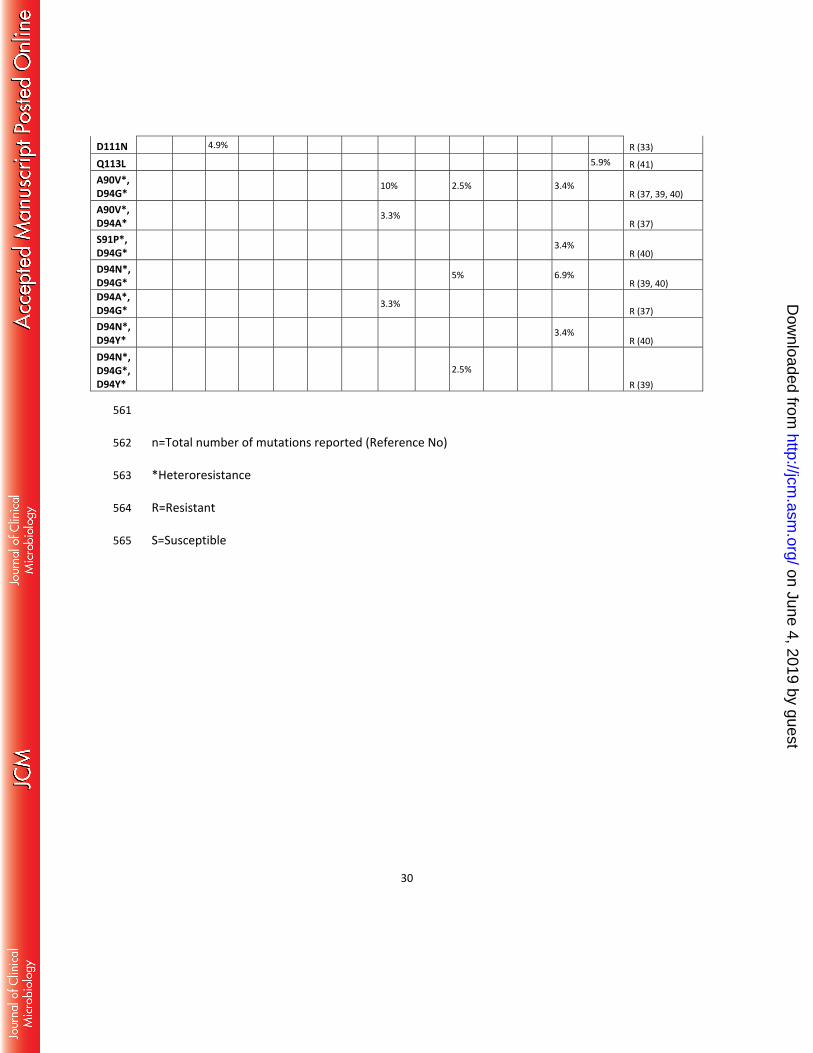
D111N 4.9% R (33)
Q113L 5.9% R (41) A90V*, D94G*
10% 2.5% 3.4% R (37, 39, 40)
A90V*, D94A*
3.3% R (37)
S91P*, D94G*
3.4% R (40)
D94N*, D94G*
5% 6.9% R (39, 40)
D94A*, D94G* 3.3%
R (37) D94N*, D94Y*
3.4% R (40)
D94N*, D94G*, D94Y*
2.5% R (39)
561
n=Total number of mutations reported (Reference No) 562
*Heteroresistance 563
R=Resistant 564
S=Susceptible 565
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from
31
566 567
568 Figure 1 569 570
571 572
573 L 574 575 576 577
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from
32
Figure 2 578
579
580
581
582
583
on June 4, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from





























