Embed Size (px)
Citation preview

A
oivwrw
sodmat
i©
K
1
iti
W
0d
Neuropsychologia 45 (2007) 796–809
Self-awareness and the subconscious effect of personal pronouns onword encoding: A magnetoencephalography (MEG) study
Peter Walla a,b,c,d,∗, Katharina Greiner b, Cornelia Duregger b,d,Luder Deecke a, Stefan Thurner b
a Ludwig Boltzmann Institute for Functional Brain Topography, Department of Clinical Neurology, Medical University of Vienna,Waehringer Guertel 18-20, 1090 Vienna, Austria
b Complex Systems Research Group, ENT, Medical University Vienna, Waehringer Guertel 18-20, 1090 Vienna, Austriac Biological Psychology Unit, Faculty of Psychology, University of Vienna, Liebiggasse 5, A-1010 Vienna, Austria
d Neuroconsult, Applied Neuroscience Institute, Meiselstrasse 29/2, 1150 Vienna, Austria
Received 21 February 2006; received in revised form 11 August 2006; accepted 14 August 2006Available online 26 September 2006
bstract
The effect of personal pronouns such as “ein” (German for “a”), “mein” (German for “my”) and “sein” (German for “his”) on the processingf associated nouns was investigated using MEG. Three different encoding strategies were provided in order to vary the level of consciousnessnvolved in verbal information processing. A shallow (alphabetic), a deep (semantic) and a very deep (contextual) encoding instruction related toisual word presentation were given to all study participants. After the encoding of pronoun–noun pairs, recognition performances of nouns onlyere tested. The number of correctly recognized nouns previously associated with “sein” was significantly lower than the number of correctly
ecognized nouns previously associated with “ein” in the shallow encoding condition. The same trend was found for “mein” associated nounshich were also less accurately recognized compared to “ein” associated nouns.Magnetic field distributions recorded during the encoding phases revealed two significant effects, one between about 200 and 400 ms after
timulus onset and the other between about 500 and 800 ms. The earlier effect was found over occipito-parietal sensors, whereas the later effectccurred over left frontal sensors. Within both time ranges, brain activation varied significantly as a function of associated pronoun independent ofepth of word processing. In the respective areas of both time ranges, conditions including personal pronouns (“mein” and “sein”) showed higheragnetic field components compared to the control condition of no personal pronouns (“ein”). Evidence is shown that early stage processing is
ble to distinguish between no personal and personal information, whereas later stage processing is able to distinguish between information related
o oneself and to another person (self and non-self).Along with other previous reports our MEG findings support the notion that particular human brain functions involved in processing neurophys-ological correlates of self and non-self can be identified.
2006 Elsevier Ltd. All rights reserved.
vio
eywords: Self-awareness; Language processing; Magnetoencephalography
. Introduction
At some stage during evolution the strategy of social liv-
ng developed among some species as an alternative to a soli-ary living style. Consequently, communication as such becamemportant with respect to information exchange between indi-∗ Corresponding author at: ENT Clinic, Medical University of Vienna,aehringer Guertel 18-20, 1090 Vienna, Austria.
E-mail address: [email protected] (P. Walla).
co1catit
028-3932/$ – see front matter © 2006 Elsevier Ltd. All rights reserved.oi:10.1016/j.neuropsychologia.2006.08.017
iduals within one group. For highly developed organisms livingn groups it is furthermore adaptive to have an idea about whatther members of the group might think or plan to do, an abilityertainly improving the fitness of the respective species in termsf the evolutionary concept of “survival of the fittest” (Darwin,859, 1871). Certainly, human language is an effective way toonsciously exchange complex information between individu-
ls. Therefore, we were motivated to use human speech as a toolo investigate human brain functions involved in the process-ng of neural representations related to oneself (self-awareness),o someone else, or to neutral conditions not containing any
cholo
phaa(
wigruw
wtbmcbC1mTih2aitc(aahewitiarc
fiafa
edcasim(
sitlrapIrfrisfHtea
miapiicdupoemidwpnt
rmsscia
2
2
were all German native speakers, right handed, and had normal or corrected
P. Walla et al. / Neuropsy
ersonal association. In other words, we wanted to test theypothesis that neural systems involved in language processingre also engaged in functions related to self-awareness and thewareness of others (self- versus non-self-specific processing)see Kircher et al., 2002).
Numerous findings from previous studies demonstrate thatithin the human brain certain functions are dedicated to ascrib-
ng mental states to oneself or to others, or in other words, toive any kind of information a personal touch. Some previouseports even highlighted that distinct brain functions deal partic-larly with first person attributes, whereas other functions dealith third person attributes.Premack and Woodruff (1978a, 1978b) introduced the frame-
ork called “theory of mind” (TOM) in order to explainheir observations of chimpanzees that appeared to demonstrateehaviors that required them to know what another chimpanzeeight be thinking, or was possibly about to do. Later, this con-
ept was applied to the human brain; for example, autism haseen interpreted as the absence of such an ability (e.g. Baron-ohen, 1995; Baron-Cohen, Jolliffe, Mortimore, & Robertson,997; Baron-Cohen, Leslie, & Frith, 1985; Frith, 1987). Finally,odern brain imaging methods such as Positron Emissionomography (PET) and functional Magnet Resonance Imag-
ng (fMRI) were used to describe brain functions in normalumans related to “mentalising” (Fletcher et al., 1995; Frith,001; Frith & Frith, 1999) or “mind reading” (Gallagher etl., 2000; Vogeley et al., 2001). Such functional imaging stud-es implicate medial prefrontal cortex and posterior superioremporal sulcus (STS) as neuroanatomical correlates of a cir-umscribed system dedicated to those functions. Vogeley et al.2001) reported differences between human self-consciousnessnd the ability to model the mental states of others; that is, thebove-mentioned TOM capacity. They used short stories whichad to be read by their study participants. Those stories includedither another person or the reader (self) or were neutral storiesithout any personal connection. They found that TOM led to
ncreased neural activity in the anterior cingulate cortex andhe left temporopolar cortex, whereas self-consciousness led toncreased neural activity in the right temporo-parietal junctionnd in the anterior cingulate cortex. Also Gallagher et al. (2000)eported activity in the medial prefrontal cortex (paracingulateortex) related to TOM in their fMRI study.
Alongside this framework, the idea of “social intelligence”, aunction which has been separately defined apart from “generalntelligence” by Humphrey (1984) (see also Baron-Cohen etl., 1999) was created. In 1990, Brothers suggested the orbito-rontal cortex (OFC), superior temporal gyrus (STG) and themygdala as brain areas responsible for “social intelligence”.
Self-recognition has sometimes been used to define the pres-nce or absence of self-awareness in primates and young chil-ren (e.g. Gallup, 1985). Basically, the idea is that a “self-oncept” or “self-awareness” (becoming the object of one’sttention; after Duval & Wicklund, 1972; Mead, 1934) can be
een as an essential basis for self-recognition. Thus, if an organ-sm has the potential to recognize itself, then self-awarenessust exist. In Keenan, Nelson, O’Connor, and Pascual-Leone2001) showed that in humans the right hemisphere of the brain
tcsaa
gia 45 (2007) 796–809 797
eems to be preferentially involved in self-recognition. Theynterpreted their findings to indicate that neural substrates ofhe right hemisphere may selectively participate in processesinked to self-awareness. In a critical assessment Morin (2002)eported that this idea has been challenged by other authors whorgued that self-recognition is instead associated with an unso-histicated self-concept and does not require any introspection.n our opinion, the right hemisphere dominance related to self-ecognition, tested using pictures of faces, is simply due to theact that face recognition is a right hemisphere function mostlyelated to the right fusiform gyrus. In our view, at the most prim-tive level of information processing, self-recognition has to beeen simply as recognizing a person by facial features. If facialeatures resemble one’s own face, then recognition takes place.owever, this is not qualitatively different from the recogni-
ion of any other person. We therefore strongly believe that thisxperimental strategy is not adequate in order to investigate self-wareness; in fact, it can be rather misleading.
In most of the previous studies, the stimuli designed to elicitentalizing, mind reading or any functions related to social
ntelligence or concepts of self-awareness were quite complexnd difficult to define. For the present study the idea arose to sim-ly use personal pronouns as a tool to engage brain functionsnvolved in the processing of some kind of personal touch. Thedea was to let study participants read nouns together with asso-iated pronouns that were either neutral or personal. Importantly,epth of word processing (Craik & Lockhart, 1972) was variedsing different word encoding instructions. Consequently, theronoun/noun combinations were processed at different levelsf consciousness engagement (see Section 2). This approachnabled us to compare subconscious and conscious word infor-ation processing. First of all, we were interested in describ-
ng any effects related to the recognition of repeated nounsepending on the previously associated pronoun. In addition,e used magnetoencephalography (MEG) to describe the tem-oral pattern of brain activity related to the encoding of differentoun/pronoun combinations with either a neutral or a personalouch.
We wanted to test the hypothesis that the event-relatedesearch approach using an electrophysiological brain imagingethod is capable of detecting distinct brain activity repre-
enting the processing of a personal versus neutral informationimply via the association of pronouns. Our motivation was toontribute to the understanding of brain mechanisms involvedn cognitive functions related to the awareness of others and thewareness of oneself.
. Methods
.1. Participants
Twenty-two volunteers (11 females) participated in the present study. They
o normal (contact lenses) vision. All of them were university students and itan therefore be assumed that linguistic ability and years of education did notignificantly differ between them. They all gave their written informed consentnd none had any reported neuropathological or psychiatric history. The meange was 25.6 years (S.D. = 3.2).

7 ycholo
2
ava(3pws
(qsrsaToGoovttwrdoitiosr
nwnTpw
biwcia
2
mspatfivtwcepd
2
2
oodfa
2
tpadp
tI((“f
tgcndt
2
tfitl3adettdewad
3
3
3e
erd
98 P. Walla et al. / Neurops
.2. Procedure
Within a magnetically-shielded room participants were seated on a comfort-ble chair where they viewed a screen in front of them onto which words wereisually presented (distance from eyes to screen was around 1.70 m and the visualngle for words was between 1.5◦ and 3.6◦) using the software package ERTSExperimental Runtime System; http://www.erts.de). Presentation duration was00 ms with an interstimulus interval of 2.5 s. Before each verbal presentation alus (+) was shown as a fixation point. In order to avoid eye artifacts, participantsere asked to blink only during the presentation of the plus and not during or
hortly after any word presentation.Nouns consisting of 3–9 letters were taken out of a German dictionary
Knaur, Das Deutsche Worterbuch, 1985). To derive nouns with the same fre-uency of everyday occurrence a psychological pre-test was conducted. Sixtytudents were asked to classify these words with regard to their everyday occur-ence. They categorized every word into one of four groups (1 for rarely, 2 forometimes, 3 for often and 4 for very often). These classifications were averagednd all words with a mean value under 2.2 were used as stimuli (Walla, 1998).he whole experiment was divided into three sessions, each session consistingf an encoding phase and a recognition phase. During the encoding phase 96erman nouns were visually presented. Each noun was presented together withne of three different pronouns: either “ein” (German for “a”), “mein” (“my”)r “sein” (“his”). Differences between sessions consisted of different depths oferbal information processing due to different encoding tasks. In one sessionhe instruction to the subject was to decide (via button press) whether or nothe first and the last letter of each presented noun was in alphabetical order,ithout paying attention to the associated pronoun. This kind of encoding is
eferred to as perceptual encoding. In another session the instruction was toecide (via button press) whether the noun referred to a living or non-livingbject, without paying attention to the associated pronoun. This kind of encod-ng is referred to as semantic encoding. In a third session the instruction was tohink of a short meaningful sentence that included the associated pronoun, andn addition to make the living/non-living decision (via button press). This kindf encoding is referred to as contextual encoding. Contextual encoding repre-ents the only condition during which conscious processing of pronouns wasequired.
After each of the encoding phases, participants were requested to do recog-ition tests during which all previously presented nouns (without pronouns)ere shown again together with the same number of new words (no pro-ouns). All new words were also taken from the above-mentioned word list.he task was then to discriminate whether the presented word had been seenreviously in the encoding phase (old), or was a new, previously unseenord.
All responses were given by button press. There was one button to pressy the right hand and another one to press by the left hand. Counterbalanc-ng was made according to which button the subject was requested to press forhat decision, as well as concerning the order of encoding instructions (session
ounterbalancing) and words within word lists. The duration of the entire exper-ment for one study participant including all three sessions and MEG set-up wasbout 3 h.
.3. Recordings
MEG measurements were recorded with a 143 channel whole-head-systemanufactured by CTF Systems Inc. (Canada) (third order gradiometers). The
ampling rate was 250 s−1 and recordings were filtered online with a band-ass from 0.25 to 80 Hz. An offline bandpass filter from 0.3 to 50 Hz waspplied. Visual inspection of all sampled data was used to remove artefacts forhe subsequent analysis. Average event-related fields (ERFs) were calculatedor each participant and across all 22 participants for each condition. Averag-ng was done with respect to visual word presentation (time 0 s at the onset ofisual word presentation). The coordinates of the MEG data sets was based onhe subjects’ nasion and the left and right preauricular points (three electrodes
ere attached at those landmarks) in relation to the sensor locations (individualoordinate systems). Intra-individual head positions were not changed betweenxperimental sessions, and inter-individual differences were kept as small asossible. Magnetic field maps were created to visualize the magnetic fieldistributions.
woc(
gia 45 (2007) 796–809
.4. Statistics
.4.1. Behavioral dataThe effects of personal pronouns and depth of verbal information processing
n both reaction times and recognition accuracy were calculated using analysisf variance (ANOVA) by introducing the factors “depth” (three levels: shallow,eep and very deep word encoding) and “pronoun” (three levels: “ein” (Germanor “a”) associated nouns, “mein” (“my”) associated nouns and “sein” (“his”)ssociated nouns).
.4.2. Neurophysiological dataThe mean amplitudes of 100 ms intervals (overlapping for 50 ms) covering
he time range from 0 to 800 ms after stimulus onset were determined for eacharticipant, each condition and each sensor. For an additional analysis these meanmplitudes were also normalized according to McCarthy and Wood (1985) toiminish any amplitude differences between participants and instead focus onossible functional differences.
A four-way repeated measures design (ANOVA) using these mean ampli-udes as dependent variables was then applied to all raw and normalized data.n the analysis, the factors “depth” (three levels) for kind of verbal encodingperceptual encoding, semantic encoding and contextual encoding), “pronoun”three levels) for kind of associated pronoun (“ein” (“a”), “mein” (“my”), orsein” (“his”)), “hemi” (two levels) for left and right hemisphere and “sensor”or all used sensor locations were introduced.
The consecutive ANOVA results (from 0 to 800 ms after stimulus onset) werehen used to demonstrate significant neurophysiological changes over time. Toraphically display the distribution of significant effects (differences betweenonditions), t-tests were calculated for every single sensor location (raw andormalized data). Using a simple color code, each sensor drawn in a two-imensional sensor map was marked according to the result of the respective-test.
.4.3. Source localizationIn order to localize neural generators underlying distinct magnetic field dis-
ributions in relation to all conditions of interest, equivalent current dipoles weretted using the CTF software “Dipole Fit”. According to the statistical results,
wo temporal ranges were significant, one between 200 and 400 ms after stimu-us onset and the other between 500 and 800 ms after stimulus onset (see Section). The clearest distribution within the earlier time window was found at 360 msnd within the later time window at 560 ms. According to the magnetic fieldistributions at these two time points, a four-dipole model was chosen for thearly interval and a five-dipole model was chosen for the later interval, to explainhe total magnetic field distributions. With respect to the statistical results fromhe t-test calculations (t-maps), only one-dipole was defined as significant andescribed for each of the two temporal ranges. All dipoles other than the onexplaining the respective area of interest as defined by significant pronoun effectsere fixed between pronoun conditions. These standardized situations enabledreliable demonstration of variations in dipole strength related to the respectiveipole of interest for both the early and the late time window.
. Results
.1. Behavioral data
.1.1. Reaction times related to task performance duringncoding
The statistical analysis revealed a significant main depthffect (F(1.946) = 19.275, p < .001; Greenhouse–Geisser cor-ected) indicating that reaction times related to task performanceuring the encoding phases differed between different levels of
ord processing. In addition, a significant main pronoun effectccurred (F(1.886) = 3.764, p = .035; Greenhouse–Geisserorrected) demonstrating that ein (a)-, mein (my)- and seinhis)-pronoun associations to nouns had an effect on reac-
P. Walla et al. / Neuropsychologia 45 (2007) 796–809 799
Table 1Reaction times (ms) during word encoding: t-test results related to reaction timesduring all three encoding sessions and for all possible pairs of pronouns
Mean S.D.
Perceptual encoding“a” nouns 1124.5 126.3“my” nouns 1136.7 153.6“his” nouns 1127 129.1
Mean (ms) S.D. t p-Value
Perceptual encoding (paired differences)“a” nouns vs. “my” nouns −12.2 68.3 −1.354 .191“a” nouns vs. “his” nouns −2.5 61.3 −0.258 .799“my” nouns vs. “his” nouns 9.7 62.3 1.23 .233
Mean S.D.
Semantic encoding“a” nouns 925 123.4“my” nouns 973.6 151.9“his” nouns 925.8 103.5
Mean (ms) S.D. t p-Value
Semantic encoding (paired differences)“a” nouns vs. “my” nouns −48.6 85.5 −2.562 .019“a” nouns vs. “his” nouns −0.8 65.2 −0.213 .834“my” nouns vs. “his” nouns 47.8 91.6 2.24 .037
Mean S.D.
Contextual encoding“a” nouns 1144.9 161“my” nouns 1145 157.1“his” nouns 1134.4 166.1
Mean (ms) S.D. t p-Value
Contextual encoding (paired differences)“a” nouns vs. “my” nouns −0.1 96.9 −0.005 .996“a” nouns vs. “his” nouns 10.5 70 0.684 .502“my” nouns vs. “his” nouns 10.6 87.7 0.551 .588
Nt
tp(
ttGipp(
ratts(
Fig. 1. Reaction times related to task performance during all three conditions ofword encoding and for all kinds of associated pronoun. In both encoding condi-t“i
wsb
3
tNwtttsco
la
ote that significant differences occurred only in the semantic encoding condi-ion (significant p-values in bold).
ion times related to task performance during the encodinghases. No significant depth × pronoun interaction occurredF(3.231) = 1.134, p = .346; Greenhouse–Geisser corrected).
ANOVAs within each level of word processing condi-ion revealed no significant pronoun effect in the percep-ual word encoding condition (F(1.962) = 1.193, p = .314;reenhouse–Geisser corrected), a significant pronoun effect
n the semantic word encoding condition (F(1.756) = 4.526,= .022; Greenhouse–Geisser corrected) and no significantronoun effect in the contextual word encoding conditionF(1.774) = 0.211, p = .785; Greenhouse–Geisser corrected).
Consequently, although the previous two-way ANOVAevealed no significant depth × pronoun interaction, follow-upnalysis demonstrated a selective pronoun effect in the seman-
ic word encoding condition. Table 1 shows all mean reactionime data and all possible pairs of conditions calculated by pairedample t-tests. As can be seen, the reaction time related to “mein”my) associated nouns in the semantic word encoding condition(ien
ions without conscious pronoun processing, nouns associated with the pronounmein” (German for “my”) elicited the longest reaction times (significant onlyn the semantic encoding condition; see Table 1).
as significantly longer than in both other conditions. Fig. 1hows all mean reaction times related to task performances asars for all three word encoding conditions.
.1.2. Recognition performance during test phasesIn the present study, recognition performance is defined as
he absolute number of hits (correctly recognized repetitions).ormally, the number of false alarms (wrongly classified newords) is subtracted from the number of hits in order to correct
he performance for guessing. However, our intention has beeno identify effects of different pronouns in a mixed experimen-al setup, instead of a blocked design. It was therefore decidedimply to compare the absolute numbers of hits between eachategory of pronoun in order to describe the relative contributionf each pronoun category to correct recognition (see Fig. 2).
A two-way ANOVA including factor “depth” (three levels:evel of word processing) and “pronoun” (three levels: kind ofssociated pronoun) revealed a significant main depth effect
F(1.980) = 59.614, p < .001; Greenhouse–Geisser corrected),ndicating that recognition performances varied between differ-nt word encoding conditions (Craik & Lockhart, 1972). Theumber of hits (correctly recognized nouns across pronouns)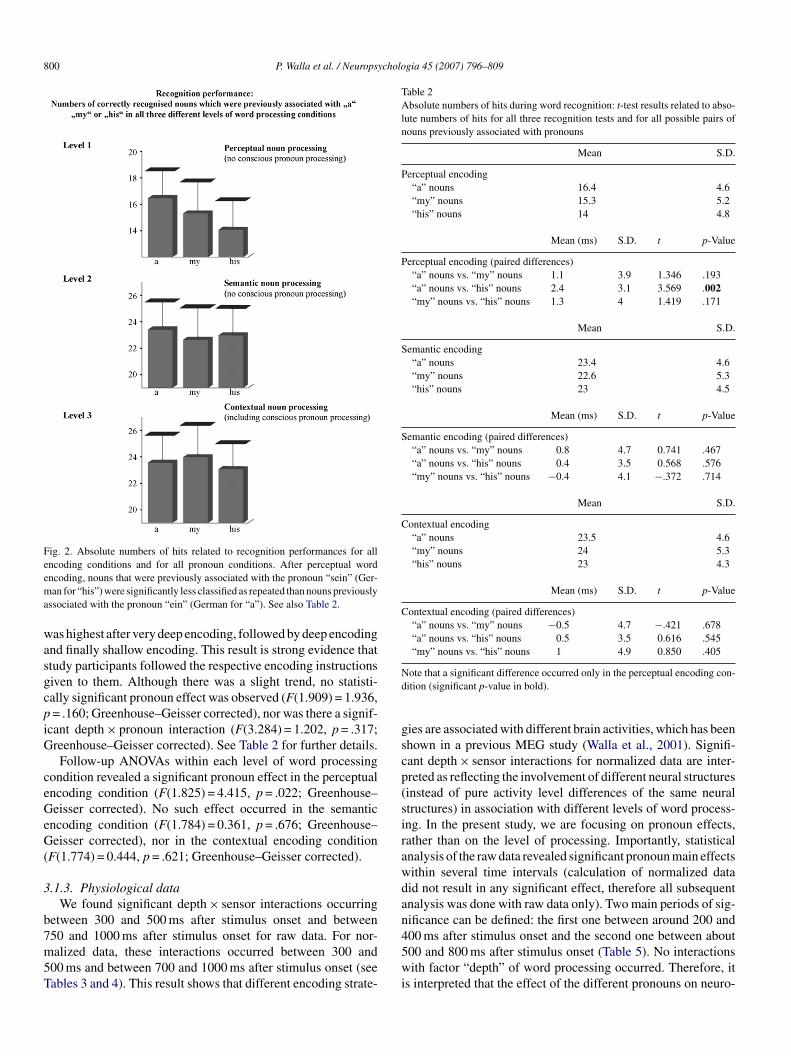
800 P. Walla et al. / Neuropsychologia 45 (2007) 796–809
Fig. 2. Absolute numbers of hits related to recognition performances for allencoding conditions and for all pronoun conditions. After perceptual wordema
wasgcpiG
ceGeG(
3
b7m5T
Table 2Absolute numbers of hits during word recognition: t-test results related to abso-lute numbers of hits for all three recognition tests and for all possible pairs ofnouns previously associated with pronouns
Mean S.D.
Perceptual encoding“a” nouns 16.4 4.6“my” nouns 15.3 5.2“his” nouns 14 4.8
Mean (ms) S.D. t p-Value
Perceptual encoding (paired differences)“a” nouns vs. “my” nouns 1.1 3.9 1.346 .193“a” nouns vs. “his” nouns 2.4 3.1 3.569 .002“my” nouns vs. “his” nouns 1.3 4 1.419 .171
Mean S.D.
Semantic encoding“a” nouns 23.4 4.6“my” nouns 22.6 5.3“his” nouns 23 4.5
Mean (ms) S.D. t p-Value
Semantic encoding (paired differences)“a” nouns vs. “my” nouns 0.8 4.7 0.741 .467“a” nouns vs. “his” nouns 0.4 3.5 0.568 .576“my” nouns vs. “his” nouns −0.4 4.1 −.372 .714
Mean S.D.
Contextual encoding“a” nouns 23.5 4.6“my” nouns 24 5.3“his” nouns 23 4.3
Mean (ms) S.D. t p-Value
Contextual encoding (paired differences)“a” nouns vs. “my” nouns −0.5 4.7 −.421 .678“a” nouns vs. “his” nouns 0.5 3.5 0.616 .545“my” nouns vs. “his” nouns 1 4.9 0.850 .405
Nd
gscp(sirawdan
ncoding, nouns that were previously associated with the pronoun “sein” (Ger-an for “his”) were significantly less classified as repeated than nouns previously
ssociated with the pronoun “ein” (German for “a”). See also Table 2.
as highest after very deep encoding, followed by deep encodingnd finally shallow encoding. This result is strong evidence thattudy participants followed the respective encoding instructionsiven to them. Although there was a slight trend, no statisti-ally significant pronoun effect was observed (F(1.909) = 1.936,= .160; Greenhouse–Geisser corrected), nor was there a signif-
cant depth × pronoun interaction (F(3.284) = 1.202, p = .317;reenhouse–Geisser corrected). See Table 2 for further details.Follow-up ANOVAs within each level of word processing
ondition revealed a significant pronoun effect in the perceptualncoding condition (F(1.825) = 4.415, p = .022; Greenhouse–eisser corrected). No such effect occurred in the semantic
ncoding condition (F(1.784) = 0.361, p = .676; Greenhouse–eisser corrected), nor in the contextual encoding condition
F(1.774) = 0.444, p = .621; Greenhouse–Geisser corrected).
.1.3. Physiological dataWe found significant depth × sensor interactions occurring
etween 300 and 500 ms after stimulus onset and between
50 and 1000 ms after stimulus onset for raw data. For nor-alized data, these interactions occurred between 300 and00 ms and between 700 and 1000 ms after stimulus onset (seeables 3 and 4). This result shows that different encoding strate-
45wi
ote that a significant difference occurred only in the perceptual encoding con-ition (significant p-value in bold).
ies are associated with different brain activities, which has beenhown in a previous MEG study (Walla et al., 2001). Signifi-ant depth × sensor interactions for normalized data are inter-reted as reflecting the involvement of different neural structuresinstead of pure activity level differences of the same neuraltructures) in association with different levels of word process-ng. In the present study, we are focusing on pronoun effects,ather than on the level of processing. Importantly, statisticalnalysis of the raw data revealed significant pronoun main effectsithin several time intervals (calculation of normalized dataid not result in any significant effect, therefore all subsequentnalysis was done with raw data only). Two main periods of sig-ificance can be defined: the first one between around 200 and
00 ms after stimulus onset and the second one between about00 and 800 ms after stimulus onset (Table 5). No interactionsith factor “depth” of word processing occurred. Therefore, its interpreted that the effect of the different pronouns on neuro-

P. Walla et al. / Neuropsycholo
Table 3ANOVA results related to magnetic field recordings (raw data) during wordencoding
ms p-Value
300–400 .037350–450 .018400–500 .020750–850 .020800–900 .007850–950 .005900–1000 .004
Significant depth × sensor interactions occurred for intervals from 300 to 500 msand from 750 to 1000 ms after stimulus onset. All values are Greenhouse–Geissercorrected.
Table 4ANOVA results related to magnetic field recordings (normalized data) duringword encoding
ms p-Value
300–400 .049350–450 .028400–500 .050700–800 .021750–850 .019800–900 .009850–950 .009900–1000 .007
Sac
pdsmodnofifc
TAe
m
2233455667
Srs
aStpparssp
osarirchtsifilswd(
mgtt
dm
ignificant depth × sensor interactions occurred for intervals from 300 to 500 msnd from 700 to 1000 ms after stimulus onset. All values are Greenhouse–Geisserorrected.
hysiological parameters (magnetic field distributions) did notiffer between the three word processing conditions (perceptual,emantic and contextual). Consequently, a new set of averagedagnetic field recordings was created. Across the factor “depth”
f word processing, one average was made for each pronoun con-ition, resulting in three main conditions of interest instead ofine conditions (three levels of depth of word processing for each
f three kinds of pronoun). Fig. 3 shows the respective magneticeld distributions related to every kind of pronoun separatelyor the earlier time period (200–400 ms) within which signifi-ant pronoun effects occurred. As calculated by the t-tests, thereable 5NOVA results related to magnetic field recordings (raw data) during word
ncoding
s p-Value
00–300 .01050–350 .00300–400 .00350–450 .00750–550 .04000–600 .01550–650 .00300–700 .00150–750 .02100–800 .050
ignificant main pronoun effects occurred for intervals covering an early timeange from about 200 to 400 ms and a later range from about 500 to 800 ms aftertimulus onset. All values are Greenhouse–Geisser corrected.
aowlpisncrtla(ojptMsi
gia 45 (2007) 796–809 801
re significant effects mainly over the parietal region (Fig. 3).trikingly, no significant differences at all occurred between
he “mein” (“my”)-pronoun condition and the “sein” (“his”)-ronoun condition. This indicates that this early stage of wordrocessing (200–400 ms) already shows discrimination betweenneutral and a personal touch related to a noun; but whether the
espective personal touch is related to oneself or to another per-on is not yet clear. Fig. 5 shows this effect at two representativeensor locations (MLP35 and MLP45, both in the left occipito-arietal region).
The particular neurophysiological situation at the later periodf time, between 500 and 800 ms after stimulus onset, is firsthown by MEG maps averaged for each pronoun condition sep-rately (Fig. 4). A very distinct area at the left fronto-temporalegion demonstrates significant physiological effects. First, sim-lar to the early processing stage, both pronoun conditions rep-esenting personal attribution differ from the neutral pronounondition—but here at another brain location. On the otherand, a second look at the t-test results in Fig. 4 reveals thathis stage of word processing can also distinguish between per-onal attribution to oneself compared to somebody else. Thiss even more evident in Fig. 5, in which continuous magneticeld changes over time are visualized at representative sensor
ocations (MLF33, MLF34). The two left fronto-temporal sen-or locations at the bottom of Fig. 5 indicate a short temporalindow of about 200 ms within which clear neurophysiologicalifferences between the “mein” (“my”) condition and the “sein”“his”) condition occurred.
The fact that almost no significant effects occurred in the nor-alized data sets, as can be seen in Figs. 3 and 4 (t-maps), sug-
ests that all respective neurophysiological differences relatedo the kind of pronoun associated with a noun are rather quanti-ative, instead of reflecting qualitatively different systems.
Source localization revealed that within the early time win-ow, in particular at 360 ms after stimulus onset, a four-dipoleodel is able to explain the total magnetic field pattern (aver-
ged across all participants) with a remaining average error ofnly 6.27%. According to both the MEG maps for this timeindow and to the respective statistical results, only one-dipole
ocation is subject to activity differences related to the kind ofronoun–noun pair that had to be encoded. This particular dipoles located within the left occipital area. As shown in Fig. 6, thetrength of that dipole varied as a function of the kind of pro-oun that had to be encoded, while other dipoles were fixed. Itan be seen that the strengths of the equivalent current dipoleselated to both personal pronouns are higher than the strength ofhe respective dipole related to the neutral condition. Within theater time window, in particular at 560 ms after stimulus onset,five-dipole model can explain the total magnetic field pattern
averaged across all participants) with a remaining average errorf only 3.25%. At this time point it is another dipole that is sub-ect to activity differences related to what kind of pronoun–nounair had to be encoded, namely the one located within the left
emporal area (while other dipoles were fixed). This is due to theEG maps seen in Fig. 4 and the statistical results (t-maps), alsohown in Fig. 4. At this dipole location the strength as measuredn nA m is higher in both personal pronoun conditions compared

802 P. Walla et al. / Neuropsychologia 45 (2007) 796–809
Fig. 3. Early neurophysiological effect: MEG maps (magnetic field distributions) averaged across depth of word processing and across all study participants for thetime interval from 200 to 300 ms after stimulus onset. First line: one map for each of the three conditions of pronoun (“ein” (“a”), “mein” (“my”), “sein” (“his”)).Second line: difference magnetic field distributions related to comparisons (subtractions) between each possible pair of pronoun condition (“mein” vs. “ein”, “sein”vs. “ein”, “sein” vs. “mein”). Sensor areas where t-tests resulted in significant differences are marked with a white dotted circle. Third line: t-maps showing thedistribution of significant differences for each of the above-mentioned comparisons (raw data). Note that “mein” vs. “ein” and “sein” vs. “ein” both resulted ins n” vsf any dM
tc
4
(
acs
ignificant differences, whereas no differences occurred for the comparison “seior each of the above-mentioned comparisons (normalized data). Note that hardly
EG curves recorded at selected sensor sites.
o the neutral condition, and also higher in the “mein” (“my”)ondition compared to the “sein” (“his”) condition (see Fig. 7).
. Discussion
In their framework about consciousness Crick and Koch2003) defined the NCC (Neural Correlates of Consciousness)
eana
. “mein”. Fourth line: t-maps showing the distribution of significant differencesifference occurred. See Fig. 5 demonstrating respective differences by showing
s the minimal set of neuronal events that gives rise to a spe-ific aspect of a conscious percept. They also stated that con-ciousness must be a largely private meaning that is accessible
xclusively to the owner of the brain. Confronting this idea,nd despite the principal difficulty in understanding conscious-ess functions, the use of modern brain imaging methods isble at least to describe underlying brain activity, without neces-
P. Walla et al. / Neuropsychologia 45 (2007) 796–809 803
Fig. 4. Later neurophysiological effect: MEG maps (magnetic field distributions) averaged across depth of word processing and across all study participants for thetime interval from 500 to 800 ms after stimulus onset. First line: one map for each of the three conditions of pronoun (“ein” (“a”), “mein” (“my”), “sein” (“his”)).Second line: difference magnetic field distributions related to comparisons (subtractions) between each possible pair of pronoun condition (“mein” vs. “ein”, “sein”vs. “ein”, “sein” vs. “mein”). Sensor areas where t-tests resulted in significant differences are marked with a white dotted circle. Third line: t-maps showing thedistribution of significant differences for each of the above-mentioned comparisons (raw data). Note that “mein” vs. “ein” and “sein” vs. “ein” both resulted insignificant differences. In addition, the comparison between “sein” and “mein” also resulted in significant differences at some of the sensor sites (no such differenceswere found during the early period of time, see Fig. 3). Fourth line: t-maps showing the distribution of significant differences for each of the above-mentionedc by sh
sisa
l
rd
omparisons (normalized data). See Fig. 5 demonstrating respective differences
arily addressing the above-mentioned privacy. This restrictions due to the objective nature of such methods. The present
tudy focused on particular forms of consciousness, namely self-wareness and the awareness of others.It is important to emphasize again that in the present study,anguage served as a tool to investigate information processing
ptsb
owing MEG curves recorded at selected sensor sites.
elated to a person compared to that related to a neutral con-ition. The respective person can either be oneself or another
erson—self or non-self. In principle, our study was designedo control the processing of nouns, each of which represents aemantic content being possessed by oneself (my car), by some-ody else (his car) or being neutral (a car; belonging to nobody).
804 P. Walla et al. / Neuropsychologia 45 (2007) 796–809
Fig. 5. Magnetic field changes over time (MEG curves) recorded at selected sensor sites showing the early effect (top) between about 200 and 400 ms after stimulusonset (at MLP35 and MLP45; left occipito-parietal area). As can be seen in the magnified curve sections on the right of both curve diagrams, both curves reflectingpersonal engagement (“mein” (“my”) and “sein” (“his”)) deviate from the curve reflecting the neutral pronoun condition “ein” (“a”). The two sensor sites at thebottom (MLF33 and MLF34, left fronto-temporal) demonstrate the later effect between about 500 and 800 ms after stimulus onset. As can be seen in the magnifiedc al ent the tic
somapptlptbfpi
4w
t“n“nte
urve sections on the right of both curve diagrams, both curves reflecting personhe neutral pronoun condition “ein” (“a”). In addition, for about two thirds ofurve reflecting somebody else (“sein” (“his”)).
For a better understanding of the rationale of the presenttudy, it is important to consider the idea that controlling depthf noun processing was our way of manipulating the involve-ent of neural systems engaged in the processing of the noun
s a visual stimulus. In the present study, three levels of nounrocessing were provided. Level 1 was meant to elicit nounrocessing only with respect to alphabetical information (let-er strings) without consciously eliciting semantic meaning;evel 2 was meant to include conscious semantic informationrocessing related to nouns; and finally level 3 was meanto elicit conscious semantic information processing related to
oth nouns and their associated pronouns within a meaning-ul context. As a consequence of increasing depth of wordrocessing, the involvement of consciousness also graduallyncreased.nedp
gagement (“mein” (“my”) and “sein” (“his”)) deviate from the curve reflectingme period, the curve reflecting onself (“mein” (“my”)) also deviates from the
.1. Reaction times related to task performances duringord encoding
The present study revealed a significantly longer reactionime during word encoding if the pronoun “mein” (German formy”) was associated with the noun in the case of semanticoun processing (word processing level 2). The phenomenontime on task” seemed to be manipulated by the personal pro-oun “mein” (“my”), even though it actually was not part ofhe task to process associated pronouns at all in this kind ofncoding condition (semantic). Although not statistically sig-
ificant, we found the same situation in the perceptual wordncoding condition. In both conditions the encoding instructionsid not include conscious intentional processing of associatedronouns. This fact could therefore already be seen as a first
P. Walla et al. / Neuropsychologia 45 (2007) 796–809 805
Fig. 6. Source localization for the earlier time range (200–400 ms): For the averaged data set (across 22 participants) the magnetic field distributions relatedto all three kinds of pronoun are displayed for a particular time point within the time window from 200 to 400 ms, which was found to include significantdifferences over left occipital and left occipito-parietal sensors depending on the kind of pronoun. The localized dipole in this respective region shows stronger braina s como
stdbao
ottti
stltc
it
ctivity as reflected by dipole strength (nA m) related to both personal pronounccipital.
ign of a subconscious neural representation of a “person” (inhis case “oneself”) reflected by longer reaction times. In theeepest encoding condition, during which each pronoun had toe consciously included in a short sentence together with itsssociated noun, not the slightest difference between conditionsf pronouns occurred (see Fig. 1).
As calculated by a two-way ANOVA, a main “depth” effectccurred, reflecting the fact that reaction times were shortest in
he semantic word encoding condition. Since it was this condi-ion within which significant effects occurred, one might arguehat the lack of significant effects in both of the other conditionss due to a ceiling effect. Although our data are not conclu-smar
pared to the neutral pronoun. The location of this dipole is interpreted as left
ive regarding this question, we tend to believe that this is nothe case: in the perceptual word encoding condition we see ateast a qualitatively similar effect (although the absolute reactionimes resemble the reaction times from the contextual encodingondition).
We therefore interpret that the personal association of a mean-ngful noun to “oneself” prolongs the response time in a seman-ic decision task even if it is not intentionally processed. We
uggest that a kind of subconscious (automatic) semantic infor-ation processing system “noticed” the occurrence of a personalssociation to oneself, which resulted in a prolongation of theespective response time.

806 P. Walla et al. / Neuropsychologia 45 (2007) 796–809
Fig. 7. Source localization for the later time range (500–800 ms): for the averaged data set (across 22 participants) the magnetic field distributions related to all threekinds of pronoun are displayed for a particular time point within the time window from 500 to 800 ms, which was found to include significant differences over lefttemporal and left fronto-temporal sensors depending on the kind of pronoun. The localized dipole in this respective region shows stronger brain activity as reflectedb utral pd
4o
wnvptwfw
Tib(tcpn
y dipole strength (nA m) related to both personal pronouns compared to the neipole is interpreted as left temporal.
.2. Recognition performance related to nouns dependingn previously associated pronouns
Only in the most superficial word processing condition dide find a significant pronoun effect related to noun recog-ition. The recognition performance of nouns that had pre-iously been associated with a pronoun relating to anothererson (“sein” (German for “his”)) was worst out of the
hree conditions, followed by nouns previously associatedith “mein” (“my”). The best recognition performance wasound related to nouns that had previously been associatedith a neutral pronoun (“ein” (German for “a”)) (see Fig. 2).
si“
ronoun (strongest activity in the “mein”(“my”) condition). The location of this
he fact that such a significant pronoun effect occurred onlyn the word encoding condition – not requiring any contri-ution of conscious semantic verbal information processingjust letter strings) – also indicates that subconscious seman-ic information processing automatically took place. Higheronscious word information processing was not sensitive toersonal or neutral associations regarding the recognition ofouns.
Overall, the behavioral results can be interpreted as demon-trating that subconscious language (semantic) processing cannclude the processing of information related to “self” and tonon-self”. Such subconscious conceptual information process-

cholo
iH
4
rngaiptitptoarcfiu8stqp
4
isenraTpacewsa“
btpiasisn
aip
wffpisots–Wbpnriluosptac
4
oniasbopbTbsrbetr
cal
P. Walla et al. / Neuropsy
ng has been reported previously (e.g. Henke, Treyer et al., 2003;enke, Mondadori et al., 2003).
.3. Neurophysiological findings
The finding of significant depth × sensor interactions for bothaw and normalized data is interpreted as evidence that differenteural structures were involved in the different encoding strate-ies. Together with our behavioral results about time on tasknd recognition performance related to depth of word process-ng, we see this as neurophysiological confirmation that studyarticipants truly varied their encoding strategies according tohe instructions given to them. However, as already mentionedn the results section, statistical analysis revealed that the fac-or “pronoun” did not interact with the factor “depth” of wordrocessing. Therefore, neurophysiological phenomena relatedo different pronouns should be seen as independent from depthf word processing. With this in mind, we can use the statisticalnalysis to draw a distinction between two distinct time windowsepresenting two different stages of serial word information pro-essing during which significant pronoun effects occurred. Therst time window was between about 200 and 400 ms after stim-lus onset, with the latter stage occurring between about 500 and00 ms after stimulus onset. Moreover, only the raw data showedignificant effects, which is interpreted as demonstrating thathe differences are quantitative in nature, instead of reflectingualitative differences due to separate systems involved in therocessing of different pronouns.
.4. Early processing stage
At the early stage (200–400 ms) there was a clear and signif-cant physiological signature over left occipito-parietal sensors,howing that “mein” (“my”) and “sein” (“his”) associated nounslicited higher femto Tesla values than “ein” (“a”) associatedouns (Figs. 3 and 5). The respective source localization resultsevealed a left occipital location for the underlying neural gener-tor eliciting this pronoun dependent activity pattern (see Fig. 6).he location and the early timeframe related to this physiologicalhenomenon both lead to the idea that underlying neural activitylready reflects some kind of subconscious personal engagementompared to a neutral condition (yet it does not show any differ-nce between “self” and “non-self”). By personal engagement,e mean possession. As mentioned above, the idea of the present
tudy was to use language (pronouns combined with nouns) astool to investigate human brain functions related to the “self”,non-self” or a neutral condition.
It is important to emphasize again that such a distinctionetween personal and neutral does not seem to depend at all onhe level of information processing. In other words, even if aronoun associated with a noun is not consciously perceived,t elicits brain activity that can distinguish between personalnd neutral information. Although it might be surprising that
ubconscious brain activity related to language processing cannclude information related to personal engagement, our datahow strong evidence that this is the case. In addition to oureurophysiological data, the above-mentioned response datasceI
gia 45 (2007) 796–809 807
lso strongly support the notion that subconscious brain activ-ty can include personal information processing (concept of aerson).
At this point we would like to introduce a neurobiologicalay of thinking which is, in our opinion, an important basis
or an appropriate understanding of this situation. The crucialact to understand is that neither light nor tones nor any otherhysical or chemical modality related to a stimulus is presentn the brain. There is no odor molecule in the brain; insteadtimuli coming from the outside world are coded in the formf electrical signals containing all relevant information as dis-inct frequencies (action potentials) and/or amplitudes (gradedynaptic potentials). The respective coding – signal transductionis the major job of a sensory organ within a single modality.ithin a functional unit of a neural network, a certain distri-
ution of signal frequencies and amplitudes can represent veryarticular information. At some stage of information processingeurophysiological metarepresentations start to exist. Metarep-esentation in this case means that the respective content, whichs coded in form of signal frequencies and amplitudes, is noonger reflecting any physical or chemical properties of a stim-lus, but rather the meaning (semantic information) of a wholebject perceived through sensory organs. Once such a metarepre-entation exists it can be used for any kind of further informationrocessing (maybe even dreaming!). Now, along with this viewhe present study shows evidence that the concept of a “person”lso exists as such a metarepresentation in the human brain andan be elicited by personal pronouns.
.5. Later processing stage
Between about 500 and 800 ms after stimulus, onset sensorsver the left frontal and left temporal areas showed significanteurophysiological differences among all three conditions ofnterest. According to the respective source localization results,neural generator located in the left temporal area appears to
how varying brain activity depending on the kind of pronouneing encoded (see Fig. 7). As can be seen in Fig. 5, at this stagef word processing not only do both conditions with personalronouns (“mein” and “sein”) deviate from the neutral condition,ut the “mein” condition also deviates from the “sein” condition.his neurophysiological phenomenon seems evident for only arief period, roughly 200 ms in duration. This qualitative resultuggests that at this stage of verbal information processing, neu-ophysiological indices can be distinguished that differentiateetween “self” and “non-self”. We want to emphasize that thisffect is rather small, and so should be treated with some cau-ion. However, as argued below, this result agrees with previousesults related to self-awareness and the awareness of others.
In summary, it is concluded that an early stage of word pro-essing (200–400 ms) already discriminates between personalnd neutral associations to nouns. The involved activities areocated roughly at the left occipital area. Furthermore, at a later
tage of word processing (500–800 ms), neural activity even dis-riminates between associations to oneself and to somebodylse. This effect is located roughly at the left temporal area.mportantly, we note that magnetic field distributions as recorded
8 ycholo
istmowpimbpaasiarrttq
pstaswrAithoinfmabostttbartli
iidtr
tc
4
bItIsafna
4
resliawissTainp
crmsoaoab
A
phMp
08 P. Walla et al. / Neurops
n the present study at sensors outside the skull are not neces-arily attributable to neural generators located directly beneathhem. Instead, any neural generator with a detectable elicited
agnetic field actually results in two field maxima (positive forutcoming and negative for ingoing), roughly on either side ofhere the neural generator itself is located in the brain. In theresent study, we find significant effects mainly for one max-mum both in the early and in the later time range. Therefore,
ore detailed information about where exactly the underlyingrain activities occurred related to the effects we found cannot berovided. We can only use rather rough anatomical terms suchs occipital and temporal. In addition, we used a grand aver-ge approach, which means that we collapsed the data from 22tudy participants. In doing so, neurophysiological data includenter-individual differences related to slight differences in brainnatomy and to the exact head position during magnetic fieldecording. Thus, our anatomical descriptions should be seen asather rough estimates, not at all accurate locations. However,here are other methods known for their excellent spatial resolu-ion, in particular fMRI, which will be used in future to answeruestions related to spatial properties.
The present neurophysiological results may be compared torevious results reported by Farrer and Frith (2002). In a fMRItudy they compared two conditions: experiencing oneself ashe cause of an action (the sense of agency), and experiencingnother person as being the cause of that action. Their resultshowed that being aware of causing an action was associatedith activation in the anterior insula, suggesting an important
ole of this cortical area for functions involved in self-awareness.more recent study by Farrer et al. (2003) revealed similar find-
ngs. They conducted a PET study and used a method allowinghem to modify their subject’s degree of control of a virtualand. Insular activation was found to correlate with the degreef movement control. The more the subjects were aware of hav-ng control of the virtual hand movements, the higher was theeural activation in the insula, thus providing more evidenceor this region’s importance for self-awareness. Immediate orinimal self-awareness has been related to motor action. Two
spects were defined: self-ownership, as in the sense that it is myody that is moving; self-agency, the sense that I am the initiatorr source of the action (see Gallagher, 2000). This idea relateself-awareness to the motor system in the human brain. Despitehe fact that in our study a motor response was required to followhe encoding instructions, we believe that our results show thathe left insular cortex is somehow involved in discriminatingetween personal association compared to no personal associ-tion, and furthermore between oneself and somebody else inelation to verbal information processing. The lateralization tohe left side, coming out of the t-test calculations (Fig. 4) for theater effect, could be due to the fact that language processing wasnvolved. This would certainly be expected to be left lateralized.
The present results are related to the above-mentioned find-ngs because the left fronto-temporal sensor sites where signif-
cant pronoun effects were found to show a neurophysiologicaliscrimination between “self” and “non-self” could be consis-ent with insular activation. As mentioned above, we can onlyoughly estimate the location of neural generators underlyingR
B
gia 45 (2007) 796–809
he physiological effects found, but insular activity could be aandidate for the later effect.
.6. The anterior cingulate gyrus
As mentioned in Section 1, the anterior cingulated gyrus haseen found to be involved in processes related to self-awareness.n our study, we did not find significant neurophysiological signshat can be linked to brain activity in the anterior cingulate area.f such activity is expected to be bilateral, this situation is noturprising because magnetic field patterns related to bilateralctivity in this area can easily be deleted. This is due to theact that both cortices in this area are very close together, andeurons potentially contributing to the outside magnetic fieldre generally parallel on both hemispheres.
.7. Left hemisphere dominance
A number of studies have concluded that left (rather thanight) hemisphere activity is associated with self-awareness. Forxample, Craik et al. (1999) found the left medial region of theuperior frontal gyrus and the left inferior frontal gyrus to beinked with self-awareness. Morin (2002), who challenged thedea that self-awareness is related to right hemisphere activity,lso discusses the notion that inner speech, which is associatedith left hemisphere activity, is linked to self-awareness. This
dea combines the notion that left hemisphere activity produceself-awareness with the suggestion that inner speech itself isomehow strongly related to the phenomenon of self-awareness.his idea comes close to our approach, since we used languages a tool to investigate self-awareness and found brain activ-ty to vary as a function of what pronoun was associated to aoun even in the absence of conscious semantic informationrocessing.
We conclude that the semantic information of visually per-eived possessive pronouns alone elicits brain activity that caneflect personal attachment in the absence of explicit infor-ation processing. This suggests a kind of semantic repre-
entation related to the concept of a person. This basic stagef information processing (200–400 ms) is most likely locatedt the left occipito-parietal area. Later on during the coursef further information processing, the left fronto-temporalrea becomes engaged, providing the potential to discriminateetween another person and oneself (500–800 ms).
cknowledgements
This study was supported by the Austrian Science Fund (FWFroject P16022-B02). Many thanks to all reviewers for their trulyelpful comments and suggestions. Finally, we want to thank Dr.ichael Landolfa (dude) for correcting English grammar and for
roviding highly valuable semantic adaptations!
eferences
aron-Cohen, S. (1995). Mindblindness: An essay on autism and theory ofmind/Simon Baron-Cohen. Cambridge, MA: MIT Press.

cholo
B
B
B
C
C
C
D
D
D
F
F
F
F
F
FG
G
G
H
H
H
K
K
M
M
M
P
P
V
W
P. Walla et al. / Neuropsy
aron-Cohen, S., Jolliffe, T., Mortimore, C., & Robertson, M. (1997). Anotheradvanced test of theory of mind: Evidence from very high functioning adultswith autism or Asperger syndrome. Journal of Child Psychology and Psy-chiatry, 38(7), 813–822.
aron-Cohen, S., Leslie, A. M., & Frith, U. (1985). Does the autistic child havea “theory of mind”? Cognition, 21, 37–46.
aron-Cohen, S., Ring, H. A., Wheelwright, S., Bullmore, E. T., Brammer, M.J., Simmons, A., et al. (1999). Social intelligence in the normal and autisticbrain: An fMRI study. European Journal of Neuroscience, 11, 1891–1898.
raik, F. I. M., & Lockhart, R. S. (1972). Levels of processing: A frameworkfor memory research. Journal of Verbal Learning and Verbal Behavior, 11,671–684.
raik, F. I. M., Moroz, T. M., Moscovitch, M., Stuss, D. T., Winocur, G., Tulving,E., et al. (1999). In search of the self: A positron emission tomography study.Psychological Science, 10, 26–34.
rick, F., & Koch, C. (2003). A framework for consciousness. Nature Neuro-science, 6(2), 119–126.
arwin, C. (1859). On the origin of species by means of natural selection, orthe preservation of favoured races in the struggle for life. London, UK.
arwin, C. (1871). The descent of man and selection in relation to sex. London,UK: Murray.
uval, S., & Wicklund, R. A. (1972). A theory of objective self-awareness. NewYork: Academic Press.
arrer, C., Franck, N., Georgieff, N., Frith, C. D., Decety, J., & Jeannerod, M.(2003). Modulating the experience of agency: A positron emission tomog-raphy study. NeuroImage, 18(2), 324–333.
arrer, C., & Frith, C. D. (2002). Experiencing oneself versus another personas being the cause of an action: the neural correlates of the experience ofagency. NeuroImage, 15, 596–603.
letcher, P. C., Happe, F., Frith, U., Baker, S. C., Dolan, R. J., Frack-owiak, R.S. J., et al. (1995). Other minds in the brain: A functional imaging study of‘theory of mind’ in story comprehension. Cognition, 57, 109–128.
rith, C. D., & Frith, U. (1999). Interacting minds, a biological basis. Science,286, 1692–1695.
rith, U. (1987). Metarepresentation and autism: How not to lose one’s marbles.Cognition, 27(3), 291–294.
rith, U. (2001). Mind blindness and the brain in autism. Neuron, 32, 969–979.allagher, H. L., Happe, F., Brunswick, N., Fletcher, P. C., Frith, U., & Frith,
C. D. (2000). Reading the mind in cartoons and stories: An fMRI study
W
gia 45 (2007) 796–809 809
of ‘theory of mind’ in verbal and non-verbal tasks. Neuropsychologia, 38,11–21.
allagher, S. (2000). Philosophical conceptions of the self: Implications forcognitive science. Trends in Cognitive Sciences, 4(1), 14–21.
allup, G. G., Jr. (1985). Do mind exist in species other than our own? Neuro-science and Biobehavioral Reviews, 9, 631–641.
enke, K., Mondadori, C. R. A., Treyer, V., Nitsch, R. M., Buck, A., & Hock, C.(2003). Non-conscious formation and reactivation of semantic associationsby way of the medial temporal lobe. Neuropsychologia, 41, 863–876.
enke, K., Treyer, V., Turi Nagy, E., Kneifel, S., Dursteler, M., Nitsch, R. M.,et al. (2003). Active hippocampus during non-conscious memories. Con-sciousness and Cognition, 12, 31–48.
umphrey, N. (1984). The social function of the intellect. In N Humphrey (Ed.),Consciousness regained (pp. 14–28). Oxford: Oxford University Press.
eenan, J. P., Nelson, A., O’Connor, M., & Pascual-Leone, A. (2001). Self-recognition and the right hemisphere. Nature, 409, 305.
ircher, T. T. J., Brammer, M., Bullmore, E., Simmons, A., Bartels, M., &David, A. S. (2002). The neural correlates of intentional and incidental self-processing. Neuropsychologia, 40, 683–692.
cCarthy, G., & Wood, C. C. (1985). Scalp distributions of event-relatedpotentials: An ambiguity associated with analysis of variance models. Elec-troencephalography and Clinical Neurophysiology, 62, 203–208.
ead, G. H. (1934). Mind, self and society. Chicago: University of ChicagoPress.
orin, A. (2002). Right hemispheric self-awareness: A critical assessment. Con-sciousness and Cognition, 11, 396–401.
remack, D., & Woodruff, G. (1978a). Chimpanzee problem-solving: A test forcomprehension. Science, 202(4367), 532–535.
remack, D., & Woodruff, G. (1978b). Does the chimpanzee have a “theory ofmind?”. Behavioural and Brain Sciences, 4, 515–526.
ogeley, K., Bussfeld, P., Newen, A., Herrmann, S., Happe, F., Falkai, P., etal. (2001). Mind reading: Neural mechanisms of theory of mind and self-perspective. NeuroImage, 14(1), 170–181.
alla, P. (1998). Neurologische grundlagen von gedachtnisfunktionen beim
menschen, Dissertation. University of Vienna, Austria.alla, P., Hufnagl, B., Lindinger, G., Imhof, H., Deecke, L., & Lang, W. (2001).Left temporal and temporo-parietal brain activity depends on depth of wordencoding: A magnetoencephalographic (MEG) study in healthy young sub-jects. NeuroImage, 13, 402–409.





























