Embed Size (px)
Citation preview

Eur. J. Biochem. 143, 183-187 (1984) IC FEBS 1984
Selection of a cDNA clone which contains the complete coding sequence for the mature form of ornithine transcarbamylase from rat liver : expression of the cloned protein in Escherichia coli Molecular cloning of rat ornithine transcarbamylase
Peter MclNTYRE, Julian F B MERCER, M Gregory PETERSON, Peter HUDSON, and Nichold? HOOGENRAAD
Department of Biochemistry, La Trobe University, Bundoora , Birth Defects Research Institute, Royal Children’s Hoyitdl, Pdrkvik and Howard Florey Institute of Experimental Physiology and Medicine, Melbourne Universit)
(Rcccived April 5, 1984) - EJB 84 0362
A cDNA clone corresponding to the mature form of ornithine transcarbamylase (OTCase) was selected from a rat liver cDNA library constructed in bacteriophage 2gt10. OTCase clones were selected using a synthetic DNA probe of 15 bases corresponding to the 3‘ end of the OTCase mRNA [ Horwich, A. L., Kraus, J. P., Williams, K., Kalousek, F., Konigsberg, W.& Rosenberg, L. E. (1983) Proc. Nut1 Acad. Sci. USA, 80, 4258-42621. Putative OTCase clones were subcloned into the expression vector, pUC9, and the identity of inserts confirmed by colony immunoassay and by electrophoretic transfer of cloned proteins from sodium dodecyl sulphate/polyacrylamide gels to nitrocellulose filters followed by probing with monospecific anti-OTCase antibodies and 12sI-labelled protein A. A clone corresponding to the full-length mature form of rat liver OTCase (plus 15 amino acids from Eschevichia coli P-galactosidasc) was obtained and the identity of the clone was confirmed by comparison of the 5‘ sequence with a limited N-terminal amino acid sequence [ Lusty, C., Jilka, R. L. & Nietsch, E. H. (1979) J . Biol. Chrrn. 254, 10030 - 100361. A sequence discrepancy between the published sequence (Lusty et al.) and the sequcnce predicted from the cDNA structure is noted.
Ornithine transcarbamylase (OTCase) is a mitochondria1 urea-cycle enzyme, which is expressed only in liver and intestine of ureotelic animals [I]. The enzyme is encoded by a nuclear gene and synthesized on membrane-free polysomes as a precursor which has a 3 400 -4000-Da N-terminal extension [2,3]. The precursor (pOTCase) is translocated across the mitochondrial membranes by an energy-dependent process [4] and proteolytically cleaved to a mature size of 36000 Da [5]. Pulse-labelling experiments show the pOTCase is rapidly removed from the cytoplasm with a half-life of 1 -2 min [5].
The mechanism of uptake of pOTCase by mitochondria is unknown. However, the N-terminal extension, or the three- dimensional structure it induces in pOTCase, is required for uptake since without the extension, OTCase does not enter the mitochondrion [6]. Although neither the amino acid sequence of the N-terminal extension, nor the specificity of cleavage site is known, the peptide apparently contains basic amino acids since uptake of pOTCase can be inhibited by basic proteins such as histones and protamines [6].
In order to investigate the mechanism of uptake of pOTCase by mitochondria we have isolated a cDNA clone coding for mature OTCase. Our selection procedure was based on screening a rat liver cDNA library with a synthetic DNA 15-mer based on the sequence of a short OTCase clone [7] and
Abbreviations. OTCase, ornithine transcarbamylase; pOTCase, precursor form of OTCase; cDNA, DNA complementary to RNA; pUC9, bacterial plasmid; SDS, sodium dodecyl sulphate.
Enzymes. Ornithine carbamoyltransferase (EC 2.1.3.3) EcoRI (EC 3.1.23.13); T4 polynucleotide kinase (EC 2.7.1.78); T4 DNA ligase (EC6.5.1.1); DNApolymerase I (EC 2.7.7.7); DNApolymerase I (Klenow fragment) (EC 2.7.7.7); lysozyme (EC 3.2.1.17); DNase I (EC 3.1.21.1).
subcloning putative OTCase cDNA into the expression vector pUC9 [8], thereby enabling immunological verification of the OTCase clones. The isolation of a clone corresponding to the full length of mature OTCase is described and the nucleotide sequence predicts an amino acid sequence almost identical to the N-terminal 24-amino-acid sequence of rat OTCase ob- tained by Lusty et al. 191.
MATERIALS AND METHODS
Enzymes and reagents
Restriction endonucleases, T4 DNA ligase, T4 polynucleo- tide kinase, DNA polymerase I, DNA polymerase I (Klenow fragment), DNase I and lysozyme were purchased from Be- thesda Research Laboratories Inc. (New England) Biolabs, Boehringer Corporation Ltd and Sigma Chemical Co. Blocked deoxyribonucleotide dimers, bases linked to polystyrene and mesitylenesulphonyl-3-nitrotriazole were purchased from Ba- chem Inc. [p3*P]ATP, [u-~’P]~ATP, [35S]dATP and bacterio- phage MI3 mp9 were purchased from Amersham Internatio- nal Ltd. Deoxyribonucleotides, dideoxyribonucleotides and calf-liver t RNA were from Boehringer Mannheim. Nitrocellu- lose was purchased from Schleicher & Schull and 1251-labelled protein A was prepared by the chloramine-T method [lo].
Isolation of RNA
RNA was extracted from Wistar rats by the method of Harding and Rutter [ I l l and poly(A)-rich RNA obtained by oligo(dT)-cellulose affinity chromatography.
184
Synthesis of oligodeoxyribonucleotide
An oligonucleotide having the sequence d(C-C-T-G-T-T- A-G-T-A-C-C-G-A-C) was synthesized by the phosphotri- ester method [12]. The oligonucleotide (50 pmol) was 5'4a- belled using T4 polynucleotide kinase and 50 pCi [Y-,~P]ATP [13]. The labelled oligonucleotide was separated from ATP and inorganic phosphate on a 10-ml columnoof Sephadex G-25 (superfine) in 1 mM EDTA, pH 7.0 and 0.1 A sodium dodecyl sulphate(SDS). Fractions containing the labelled oligonucleo- tide were combined and used for screening a rat liver cDNA library.
Colony hybridization and characterization of' recombinant bacteriophages
A rat liver cDNA library, constructed by insertion of cDNA into the EcoRI site of bacteriophate LgtlO was kindly provided by Geoff Howlett, Melbourne University. This library was screened for OTCase inserts using a32P-labelled synthetic DNA probe (8.4 x lo7 dpm/250 ng). Filters for plaque hybridization were prepared according to the method of Benton and Davis [14J, hybridizations were carried out for 16 h in 4 x NaCl/Cit 0.1 A SDS at 40 "C and filters washed in the same buffer for 2 x 20 min [15]. The sizes of the DNA inserts in positive lambda clones were determined by restric- tion analysis of recombinant bacteriophage DNA. DNA was isolated from plate lysates, purified on 1 -ml DEAE-Sephacel columns [I61 and digested with EcoRI (2-pg samples of DNA, 8 units EcoRI, 37 "C for 2 h). The insert sizes were estimated by agarose gel electrophoresis by comparison with 4 x 1 74 HueIII fragments. The authenticity of the inserts was further checked by Southern blot analysis [I71 of the gel and hybridizing with the 5'-labelled oligonucleotide probe.
Nick translation
The 1450-base-pair EcoRI insert was isolated from a low- melting-temperature agarose gel and further purified on a small DEAE-Sephacel column [16]. 200 ng of this material was nick translated with [E~ 'P]~ATP to a specific activity of 4 x 10sdpm/pg [18].
Cloning and expression qf OTCase in pUC9
2 pg DNA from putative OTCase clones in AgtlO, with inserts of 1450 and 1200 base pairs, was digested with EcoRI and the DNA was ligated into the EcoRI site of the expression vector pUC9 and transformed into Escherichia coli JMlOl [8]. OTCase-positive clones were selected by filter hybridization with the nick-translated insert from the 1450-base-pair clone.
Screening recombinants in p UC9 with antibodies
Replica filters were also screened for expression of im- munoreactive OTCase as follows : recombinants were cultured on nutrient agar containing ampicillin (30 pg/ml) with the fl- galactosidase inducer, isopropylthiogalactoside (1 mM). After 16 h culture at 37"C, colonies were lysed in situ by placing nitrocellulose filters on a filter-paper (Whatman 1) soaked in 100 mM NaHCO,, 1 % Triton X-100 and 2 mg/ml lysozyme in a chloroform-saturated atmosphere. After 20 min at room temperature, filters were waspd in the same buffer minus lysozyme and incubated with 3 A (w/v) bovine serum albumin in 10 mM Tris/HCl, pH 7.4,0.9% (w/v) NaCl for 2 h at 25 "C.
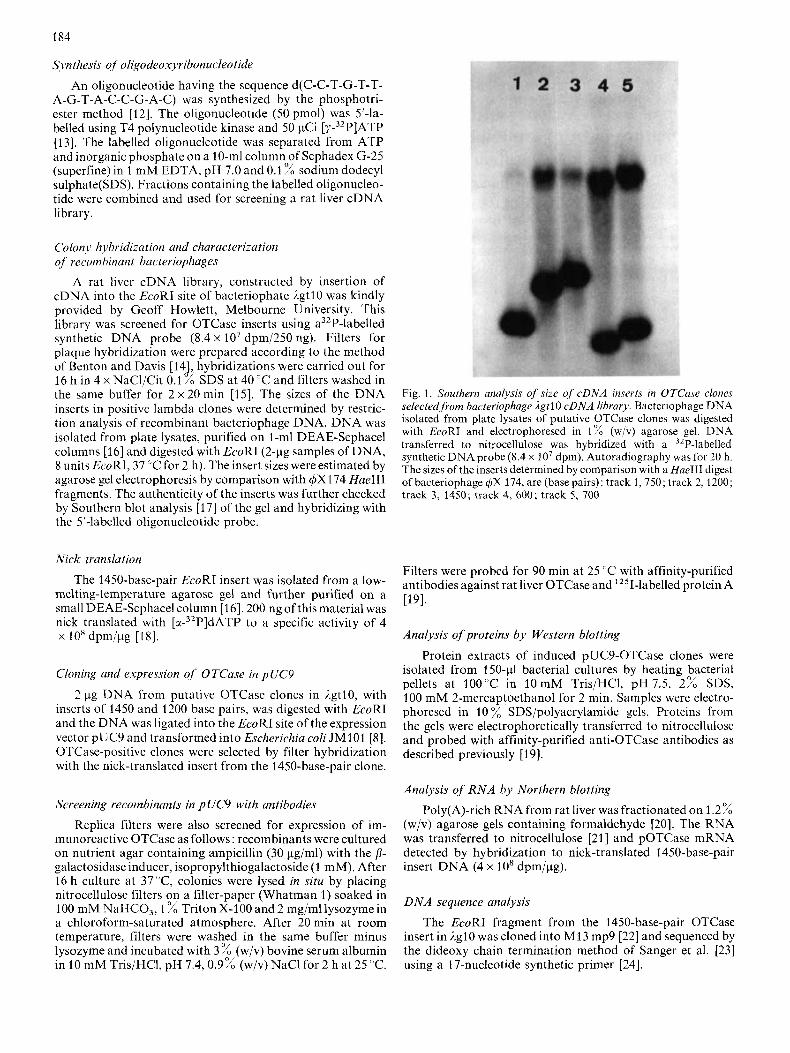
Fig. 1. Southern analysis of size of cDNA inserts in OTCase clones selected from bacteriophage lg t 10 cDNA library. Bacteriophage DNA isolated from plate lysates of putative OTCase clones was digested with EcoRI and electrophoresed in 1% (wiv) agarose gel. DNA transferred to nitrocellulose was hybridized with a 32P-labelled synthetic DNAprobe (8.4 x lo7 dpm). Autoradiography was for 20 h. The sizes of the inserts determined by comparison with a HueIII digest of bacteriophage @X 174, are (base pairs): track 1,750; track 2,1200; track 3 , 1450; track 4, 600; track 5 , 700
Filters were probed for 90 min at 25 "C with affinity-purified antibodies against rat liver OTCase and 1251-labelled protein A ~ 9 1 .
Analysis of proteins by Western blotting
Protein extracts of induced pUC9-OTCase clones were isolated from 150-p1 bacterial cultures by heating bacterial pellets at 100°C in 10mM Tris/HCl, pH 7.5, 2% SDS, 100 mM 2-mercaptoethanol for 2 min. Samples were electro- phoresed in 10 "/, SDS/polyacrylamide gels. Proteins from the gels were electrophoretically transferred to nitrocellulose and probed with affinity-purified anti-OTCase antibodies as described previously [19].
Analysis of R N A by Northern blotting
Poly(A)-rich RNA from rat liver was fractionated on 1.2% (w/v) agarose gels containing formaldehyde [20]. The RNA was transferred to nitrocellulose [21] and pOTCase mRNA detected by hybridization to nick-translated 1450-base-pair insert DNA (4 x 1 O8 dpm/pg).
D N A sequence analysis
The EcoRI fragment from the 1450-base-pair OTCase insert in LglO was cloned into MI3 mp9 [22] and sequenced by the dideoxy chain termination method of Sanger et al. [23] using a 17-nucleotide synthetic primer [24].
185
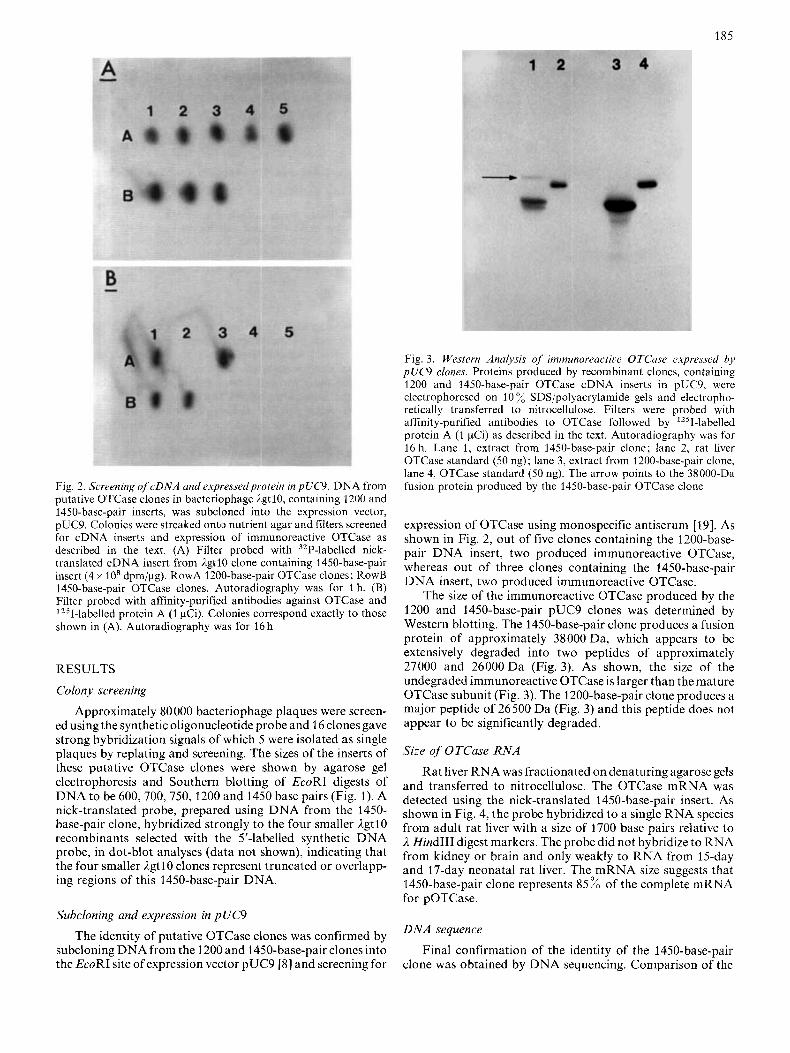
Fig. 2. Screening of cDNA and expressedprotein in pUC9. DNA from putative OTCase clones in bacteriophage IgtlO, containing 1200 and 1450-base-pair inserts, was subcloned into the expression vector, pUC9. Colonies were streaked onto nutrient agar and filters screened for cDNA inserts and expression of immunoreactive OTCdse as described in the text. (A) Filter probed with 32P-labelled nick- translated cDNA insert from AgtlO clone containing 1450-base-pair insert (4 x 10' dpm/pg). RowA 1200-base-pair OTCase clones; RowB 1450-base-pair OTCase clones. Autoradiography was for 1 h. (B) Filter probed with affinity-purified antibodies against OTCase and 1251-labelled protein A (1 pCi). Colonies correspond exactly to those shown in (A). Autoradiography was for 16 h
RESULTS
Colony screening
Approximately 80 000 bacteriophage plaques were screen- ed using the synthetic oligonucleotide probe and 16 clones gave strong hybridization signals of which 5 were isolated as single plaques by replating and screening. The sizes of the inserts of these putative OTCase clones were shown by agarose gel electrophoresis and Southern blotting of EcoRI digests of DNA to be 600,700,750,1200 and 1450 base pairs (Fig. 1). A nick-translated probe, prepared using DNA from the 1450- base-pair clone, hybridized strongly to the four smaller ilgtl0 recombinants selected with the 5'4abelled synthetic DNA probe, in dot-blot analyses (data not shown), indicating that the four smaller AgtlO clones represent truncated or overlapp- ing regions of this 1450-base-pair DNA.
Subcloning and expression in pUC9
The identity of putative OTCase clones was confirmed by subcloning DNA from the 1200 and 1450-base-pair clones into the EcoRI site of expression vector pUC9 [8] and screening for
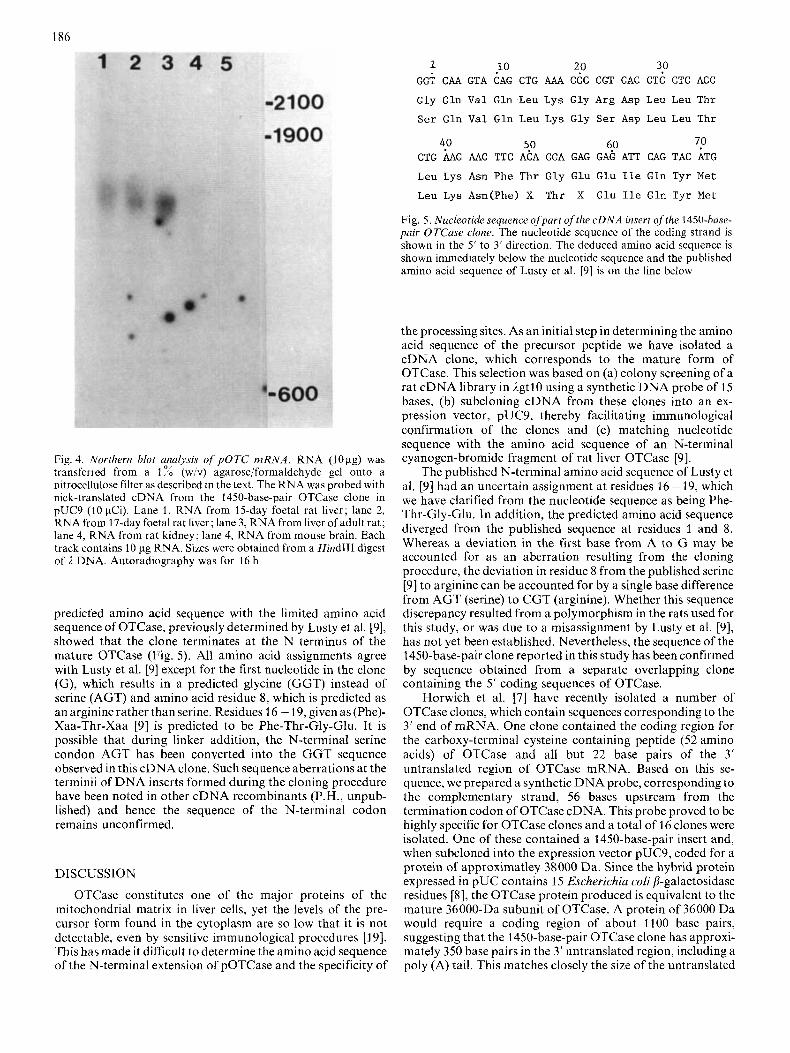
Fig. 3. Western Analysis of immunoreactive OTCuse expressed by pUC9 clones. Proteins produced by recombinant clones, containing 1200 and 1450-base-pair OTCase cDNA inserts in pUC9, were electrophoresed on 10 % SDS/polyacrylamide gels and electropho- retically transferred to nitrocellulose. Filters were probed with affinity-purified antibodies to OTCase followed by '251-labelled protein A (1 pCi) as described in the text. Autoradiography was for 16 h. Lane 1, extract from 1450-base-pair clone; lane 2, rat liver OTCase standard (50 ng); lane 3, extract from 1200-base-pair clone, lane 4, OTCase standard (50 ng). The arrow points to the 38000-Da fusion protein produced by the 1450-base-pair OTCase clone
expression of OTCase using monospecific antiserum [19]. As shown in Fig. 2, out of five clones containing the 1200-base- pair D N A insert, two produced immunoreactive OTCase, whereas out of three clones containing the 1450-base-pair DNA insert, two produced immunoreactive OTCase.
The size of the immunoreactive OTCase produced by the 1200 and 1450-base-pair pUC9 clones was determined by Western blotting. The 1450-base-pair clone produces a fusion protein of approximately 38000Da, which appears to be extensively degraded into two peptides of approximately 27000 and 26000 Da (Fig. 3). As shown, the size of the undegraded immunoreactive OTCase is larger than the mature OTCase subunit (Fig. 3). The 1200-base-pair clone produces a major peptide of 26500 Da (Fig. 3) and this peptide does not appear to be significantly degraded.
Size of' OTCase RNA
Rat liver RNA was fractionated on denaturing agarose gels and transferred to nitrocellulose. The OTCase mRNA was detected using the nick-translated 1450-base-pair insert. As shown in Fig. 4, the probe hybridized to a single RNA species from adult rat liver with a size of 1700 base pairs relative to 1 Hind111 digest markers. The probe did not hybridize to RNA from kidney or brain and only weakly to R N A from 15-day and 17-day neonatal rat liver. TheomRNA size suggests that 1450-base-pair clone represents 85 4 of the complete mRNA for pOTCase.
DNA sequence
Final confirmation of the identity of the 1450-base-pair clone was obtained by DNA sequencing. Comparison of the
3 86
30 10 2.0 1 GGT CAA GTA ~ A G CTG AAA GGC CGT GAC CTC CTC ACC
G l y Gln V a l G l n L e u L y s G l y A r g A s p Leu Leu T h r
Ser G l n V a l G l n Leu Lys G l y Ser A s p Leu Leu T h r
70 5.0 6s
CTG LAG AAC TTC ACA GGA GAG GAG ATT CAG TAC ATG 40
Leu Lys A s n Phe T h r G l y G l u G l u I l e G l n T y r M e t
Leu Lys A s n ( P h e ) X T h r X G l u I l e G l n T y r M e t
Fig. 5. Nucleotide sequence ojpart of the cDNA insert ofthe 1450-base- pair OTCase clone. The nucleotide sequence of the coding strand is shown in the 5‘ to 3’ direction. The deduced amino acid sequence is shown immediately below the nucleotide sequence and the published amino acid sequence of Lusty et al. [9] is on the line below
Fig. 4. Northern blot analysis O fpOTC mRNA. RNA transferred from a 1 % (w/v) agarose/formaldehyde
(10pg) was gel onto a
nitrocellulose filter as described in the text. The RNA was probed with nick-translated cDNA from the 1450-base-pair OTCase clone in pUC9 (10 pCi). Lane 1, RNA from 15-day foetal rat liver; lane 2, RNA from 17-day foetal rat liver; lane 3, RNA from liver of adult rat; lane 4, RNA from rat kidney; lane 4, RNA from mouse brain. Each track contains 10 pg RNA. Sizes were obtained from a Hind111 digest of 1 DNA. Autoradiography was for 16 h
predicted amino acid sequence with the limited amino acid sequence of OTCase, previously determined by Lusty et al. [9], showed that the clone terminates at the N terminus of the mature OTCase (Fig. 5). All amino acid assignments agree with Lusty et al. [9] except for the first nucleotide in the clone (G), which results in a predicted glycine (GGT) instead of serine (AGT) and amino acid residue 8, which is predicted as an arginine rather than serine. Residues 16 - 19, given as (Phe)- Xaa-Thr-Xaa [9] is predicted to be Phe-Thr-Gly-Glu. It is possible that during linker addition, the N-terminal serine condon AGT has been converted into the GGT sequence observed in this cDNA clone. Such sequence aberrations at the terminii of DNA inserts formed during the cloning procedure have been noted in other cDNA recombinants (P.H., unpub- lished) and hence the sequence of the N-terminal codon remains unconfirmed.
DISCUSSION
OTCase constitutes one of the major proteins of the mitochondria1 matrix in liver cells, yet the levels of the pre- cursor form found in the cytoplasm are so low that it is not detectable, even by sensitive immunological procedures [ 191. This has made it difficult to determine the amino acid sequence of the N-terminal extension of pOTCase and the specificity of
the processing sites. As an initial step in determining the amino acid sequence of the precursor peptide we have isolated a cDNA clone, which corresponds to the mature form of OTCase. This selection was based on (a) colony screening of a rat cDNA library in AgtlO using a synthetic DNA probe of 15 bases, (b) subcloning cDNA from these clones into an ex- pression vector, pUC9, thereby facilitating immunological confirmation of the clones and (c) matching nucleotide sequence with the amino acid sequence of an N-terminal cyanogen-bromide fragment of rat liver OTCase [9].
The published N-terminal amino acid sequence of Lusty et al. [9] had an uncertain assignment at residues 16 - 19, which we have clarified from the nucleotide sequence as being Phe- Thr-Gly-Glu. In addition, the predicted amino acid sequence diverged from the published sequence at residues 1 and 8. Whereas a deviation in the first base from A to G may be accounted for as an aberration resulting from the cloning procedure, the deviation in residue 8 from the published serine [9] to arginine can be accounted for by a single base difference from AGT (serine) to CGT (arginine). Whether this sequence discrepancy resulted from a polymorphism in the rats used for this study, or was due to a misassignment by Lusty et al. [9], has not yet been established. Nevertheless, the sequence of the 1450-base-pair clone reported in this study has been confirmed by sequence obtained from a separate overlapping clone containing the 5’ coding sequences of OTCase.
Horwich et al. [7] have recently isolated a number of OTCase clones, which contain sequences corresponding to the 3’ end of mRNA. One clone contained the coding region for the carboxy-terminal cysteine containing peptide (52 amino acids) of OTCase and all but 22 base pairs of the 3’ untranslated region of OTCase mRNA. Based on this se- quence, we prepared a synthetic DNA probe, corresponding to the complementary strand, 56 bases upstream from the termination codon of OTCase cDNA. This probe proved to be highly specific for OTCase clones and a total of 16 clones were isolated. One of these contained a 1450-base-pair insert and, when subcloned into the expression vector pUC9, coded for a protein of approximatley 38000 Da. Since the hybrid protein expressed in pUC contains 15 Escherichia coli P-galactosidase residues [8], the OTCase protein produced is equivalent to the mature 36000-Da subunit of OTCase. A protein of 36000 Da would require a coding region of about 1100 base pairs, suggesting that the 1450-base-pair OTCase clone has approxi- mately 350 base pairs in the 3’ untranslated region, including a poly (A) tail. This matches closely the size of the untranslated

187
region found by Horwich et al. in their 3’ OTCase clone (356 base pairs) [7]. The size of mRNA for pOTCase was found by RNA blot analysis t o be 1700 nucleotides, consistent with a migration of 1 4 S i n sucrose gradients [7]. By comparing the size of the mRNA with the 1450-base-pair c D N A , which encompasses the mature OTCase subunit, it can be predicted that the precursor peptide extension on pOTC is accom- modated by a 250-nucleotide length of m R N A . Since the peptide extension is about 3400 -4000 Da in size, it would appear that this extra length of mRNA comprises about 100 nucleotides of translated sequence and perhaps 15 0 nucleo- tides of untranslated m R N A . Thus, it would appear that mRNA for this X-linked enzyme has a relatively small untranslated 5’ region and a more extensive 3’ untranslated region.
Although Horwich et al. [7] recently reported the isolation of rat liver OTCase clones, these all contained relatively short c D N A inserts corresponding t o the 3‘ end of the OTCase mRNA. An additional disadvantage t o these clones is that the longest of them, which encodes the carboxy-terminal52 amino acids of OTCase, also conta.ined a c D N A fragment corre- sponding t o an unrelated mRNA [7]. The OTCase c D N A clone we have isolated corresponds to the full length of the mature OTCase subunit and should, therefore, be useful a s a hybridization probe t o select clones containing c D N A encod- ing the 3400 -4000-Da N-terminal extension of pOTCase and to characterize the structure of the X-linked gene coding for OTCase.
We wish to thank Lynda Graf for her skilful technical assistance and Jeremy Nathans and Dennis Scanlan for valuable advice. This work was supported by a grant from the Australian National Health and Medical Research Council.
REFERENCES
1. Jones, M.E., Anderson, A.D., Anderson, C. & Hodes, S . (1961)
2. Conboy, J.G., Kalousek, F. & Rosenberg, L.E. (1979) Proc. Nutl Arch. Biochem. Biophys. 95, 499 -507.
Acud. Sci. USA 76, 5724 - 5727.
3. Mori, M., Miura, S . , Tatibana, M. & Cohen, P.P. (1980) J . Biochem. (Tokyo) 88, 1829 - 1836.
4. Mori, M., Morita, T., Miura, S . & Tatibana, M. (1981) J . Biol. Chem. 256, 8263 -8266.
5. Mori, M., Morita, T., Ikeda, F., Amaya, Y., Tatibana, M. & Cohen, P.P. (1981) Proc. Natl Acad. Sci. USA 78,6056-6060.
6. Mori, M., Miura, S . , Morita, T., Takiguchi, M. & Tatibana, M. (1982) Mol. Cell. Biochem. 49, 99 - 13 1.
7. Horwich, A.L., Kraus, J.P., Williams, K., Kalousek, F., Konigs- berg, W. & Rosenherg, L.E. (1983) Proc. Nutl Acad. Sci. USA 80, 4258 -4262.
8. Vieira, J. & Messing, J. (1982) Gene 19, 259 -268. 9. Lusty, C., Jilka, R.L. & Nietsch, E.H. (1979) J . Biol. Chem. 254,
10. Bernard, C.C.A., Randell, V.B., Horvath, L.B., Carnegie, P.R. &
11. Harding, J.D. & Rutter. W.J. (1978) J . Biol. Chem. 253,
12. Ike, Y., Ikuta, S . , Sato, M., Huang, J. & Itakura, K. (1983) Nucleic
13. Sgaramella, V. & Khorana, H.G. (1972) J . Mol. Biol. 72,
14. Benton, W.D. & Davis, R.W. (1977) Science (Wush. DC) 196,
15. Montgomery, D.L., Hall, B.D., Gillam, S. & Smith, M. (1978) Cell 14, 673 -680.
16. Maniatis, T., Fritsch, E.F. & Sambrook, J. (1982) in Moleculur Cloning, a Laboratory Manual, Cold Spring Harbour Labora- tory, New York.
10030-10036.
Mackay, I.R. (1981) Immunology 43, 447 -451.
8736 -8740.
Acid Res. 11, 477 -488.
427 -444.
180-182.
17. Southern, E. (1975) J. Mol. Bid. 98, 503 -517. 18. Maniatis, T., Jeffrey, A. & Kleid, D.G. (1973) Proc. Nut1 Acad.
19. McIntyre, P., de Martinis, M.L. & Hoogenraad, N. (1983)
20. Rave, N., Crkvenjakov, R. & Boetker, H. (1979) Nucleic Acid.
21. Thomas, P.S. (1980) Proc. Nut1 Acad. Sci. USA 77, 5201 -5205. 22. Messing, J. & Vieira, J. (1982) Gene 19, 269-276. 23. Sanger, F., Nicklen, S . & Coulsen, A.R. (1977) Proc. Natl Acad.
Sci. USA 74, 5463 -5467. 24. Duckworth, M.L., Gait, M.J., Goelet, P., Hong, G.F., Singh, M.
& Titmas, R.C. (1981) Nucleic Acid Res. Y, 1691 -3706.
Sci. USA 72, 1184-1188.
Biochem. Int. 6, 365 -373.
Res. 6, 3559-3567.
P. McIntyre and N. Hoogenraad, Department of Biochemistry, La Trobe University, Bundoora, Victoria, Australia 3083 J. F. B. Mercer and M. G. Peterson, Birth Defects Research Institute, Royal Children’s Hospital Research Foundation, Flemington Road, Parkville, Victoria, Australia 3052 P. Hudson, Division of Protein Chemistry, Commonwealth Scientific and Industrial Research Organization, 343 Royal Parade, Parkville, Victoria, Australia 3052




![Ultraviolet Radiation Induction of Ornithine …...[CANCER RESEARCH 50, 2631-2635, May 1, 1990] Ultraviolet Radiation Induction of Ornithine Decarboxylase in Rat Keratinocytes1 Cheryl](https://img.pdfslide.us/doc/110x75/5f96afeee057bb0804298361/ultraviolet-radiation-induction-of-ornithine-cancer-research-50-2631-2635.jpg)
![t e c h n ol gy Journal of Biotechnology & Biomaterials · argF proB kgd) for L-ornithine production, which could produce 4.62 g/L of L-ornithine [13]. The level of L-ornithine production](https://img.pdfslide.us/doc/110x75/5e22e2c1220ab9163b5a39e7/t-e-c-h-n-ol-gy-journal-of-biotechnology-biomaterials-argf-prob-kgd-for-l-ornithine.jpg)























