Embed Size (px)
Citation preview

HAL Id: hal-00890892https://hal.archives-ouvertes.fr/hal-00890892
Submitted on 1 Jan 1991
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Selection for high and low, colony weight gain in thehoney bee, Apis mellifera, using selected queens and
random malesNw Calderone, Mk Fondrk
To cite this version:Nw Calderone, Mk Fondrk. Selection for high and low, colony weight gain in the honey bee, Apismellifera, using selected queens and random males. Apidologie, Springer Verlag, 1991, 22 (1), pp.49-60.�hal-00890892�

Original article
Selection for high and low, colony weight gainin the honey bee, Apis mellifera,
using selected queens and random males
NW Calderone MK Fondrk
The Ohio State University, Department of Entomology, Columbus, OH 43210, USA
(Received 13 June 1990; accepted 24 October 1990)
Summary — Seasonal colony weight gain (honey production) in the honey bee, Apis mellifera, canbe modified by selection. Two-way selection for strains of honey bees exhibiting high or low season-al colony weight gain was made for 3 generations. In each generation, daughter queens were rearedfrom selected colonies and permitted to mate naturally with unselected males. Two strains of honeybees were produced that differed significantly with respect to seasonal colony weight gain. These re-sults suggest that commercial and hobbyist beekeepers can increase the efficiency of honey produc-tion by using a simple selection and mating scheme. Several variables were also evaluated as indi-cators of seasonal colony weight gain. Short-term colony weight gain was significantly correlatedwith seasonal colony weight gain and is a useful aid to selection. Early-winter colony weight, late-winter colony weight, early-spring colony weight, and winter weight loss were not correlated withseasonal colony weight gain and do not appear to be useful aids to selection.
Apis mellifera / honey yield / colony-level selection
INTRODUCTION
The selection of improved strains of hon-ey bees is a frequently discussed topic inthe bee literature. However, despite dem-onstrations of genetic variability for a num-ber of traits, benefits resulting from selec-tion for those traits have not generally
been realized by beekeepers. The paucityof commercially available selected strainsof honey bees can be traced, in part, toseveral aspects of honey-bee biology.First, when a breeder employs colony-level selection, the queen-worker castesystem results in measurements beingbased partially on the aggregate perfor-mance of workers, rather than directly on
a Correspondence and reprints.b Current address : USDA-ARS, Bee Research Laboratory, Building 476 BARC-EAST, Beltsville, MD20705, USAc Current address : Department of Entomology, University of California, Davis, CA 95616, USA

the performance of the reproductive indi-viduals. Second, multiple mating (polyan-dry) by queens lowers the degree of ge-netic relatedness among the workersbeing evaluated and makes control overthe male gametic contributions to succeed-ing generations difficult. These factors re-duce the response to selection. Additional-
ly, the honey-bee’s sex-determining me-chanism results in a high proportion of invi-able brood and a corresponding reductionin colony viability when the effective sizeof the breeding population is restricted, asis common with many mating systems.These impediments can be overcome, inpart, through the use of isolated matingyards, instrumental insemination, and con-trolled breeding populations. However,these remedies require considerable skilland effort on the part of the breeder andhave not, in general, been implemented bythe various sectors of the beekeeping in-dustry in the United States.
An ideal system for hobbyists, commer-cial queen producers, and commercialhoney producers is one that incorporates amodest selection coefficient, produces nat-urally-mated queens, and avoids the diffi-culties associated with instrumental insem-
ination, isolated mating yards, and themaintenance of controlled breeding popu-lations. Rothenbuhler (1980) suggestedthat beekeepers might avoid these difficul-ties and still make improvements in theirstock by carefully evaluating their colonies,rearing daughter queens from the ’best’10% of those colonies, and allowing thosedaughters to mate naturally with malesfrom other than the queen mothers. Weevaluated this technique as a tool for in-creasing honey production using seasonalcolony weight gain as an indicator of hon-ey production (McLellan, 1977). We alsoevaluated short-term colony weight gain,early-winter colony weight, early-springcolony weight, and winter weight loss asindicators of seasonal colony weight gain.
MATERIALS AND METHODS
Two-way selection for high and low, seasonalcolony weight gain was conducted at The OhioState University Bee Laboratory from 1983through 1986. Seasonal colony weight gain isdefined here as the net change in colony weightfrom the first nectar flow in the spring throughthe end of the yellow sweetclover (Melilotus offi-cinalis) nectar flow in early summer. This periodgenerally includes the major nectar flow in thecentral Ohio area. In 1986, this period was ex-tended through the soybean nectar flow (Gly-cine max var) which ended in early August.
Colony management
Colonies were kept in a common apiary. Eachwas maintained in a 10-frame, standard depth,Langstroth-style hive consisting of 2 broodchambers separated from the honey-storagearea by a queen excluder. Combs for honeystorage were provided to all colonies at thesame time whenever the average colony weightindicated that there were < 10 combs availablefor additional storage. Colonies were inspectedat 10-d intervals in the spring, at which timebrood chambers were reversed, queen cells re-moved, and disease inspections performed. Col-onies with > 50 cells of chalkbrood (Ascophaeraapis) were removed from the experiment. Nocases of American or European foulbrood wereobserved. No attempt was made to equalize col-ony populations at this time (Szabo and Lefko-vitch, 1987) because variation in the size of col-ony populations may account for a significantproportion of variation in honey production (Far-rar, 1937; Cale and Gowen, 1956; Moeller,1961) and may be heritable. All colonies wereinspected in November to ensure that at least29,25 kg of honey were available for overwinter-ing.
Selection criterion and mating system
A base population of 33 colonies with naturally-mated queens obtained from several commer-cial sources was evaluated for variability in sea-sonal colony weight gain during the spring and

early summer of 1983. Colonies were weighedevery 8 d (short-term weight gains) during Mayand June until the end of the sweetclover nectarflow. Selections were made on the basis of total
weight gain during that period. Queens from the3 colonies with the highest seasonal weightgains were selected to initiate 3 sublines com-prising the high strain. Queens from the 3 colo-nies with the lowest gains were selected to ini-tiate 3 sublines comprising the low strain.
Daughter queens were reared from thequeen mother of each subline, marked for identi-fication, introduced to queen-mating colonies,and permitted to mate naturally. Reproductiveisolation of the daughter queens from the parentpopulation was achieved by maintaining thequeen-mating apiary > 23 km from the parentapiary (Peer, 1957). Consequently, all daughterqueens had access to the same pool of unse-lected males. Mated queens were introduced tocolonies from the preceding generation duringthe first 10 d of August. Assignment to colonieswas randomized. Requeening in early Augustensured complete replacement of the workerpopulations by the time colonies entered winter.Colonies were evaluated the following season,and the queen from each subline heading thecolony with the highest (high-strain sublines) orlowest (low-strain sublines) seasonal weightgain was selected to continue that subline in
subsequent generations. The same selection,mating, and requeening procedures were fol-lowed each year.
In 1984 and 1985, colony weights were ob-tained in December or January, and in lateMarch (static weights). These were followed bybi-weekly weighings (short-term weight gains)during the evaluation period that extended fromearly May to late June (1984) or early July(1985), depending on the termination date of thenectar flow. In 1986, colonies were weighed at
30-d intervals (short-term weight gains) and theevaluation period was continued through earlyAugust due to a dearth of nectar in May andJune.
Statistical model
Variation in seasonal colony weight gain was an-alyzed using a nested analysis of variance(Kempthorne, 1952; Sokal and Rohlf, 1981)model with strain (high or low) as the main effectand subline as the nested effect (the composi-tion of the expected mean squares is given in ta-ble I) :
yijk = seasonal weight gain of the k th colony ofthe j th subline of the i th strain; μ = averageseasonal weight gain of all colonies; ai = aver-
age deviation of colonies of the ith strain from μ;
bij = average deviation of colonies of the jthsubline nested within the ith strain from the aver-
age seasonal weight gain of colonies of the ithstrains; ϵijk = individual error.
A 1-tailed test of significance was used totest the hypothesis that the average weight gainby colonies from the high strain was less than orequal to the average weight gain by coloniesfrom the low strain ((H0,μH < = μL; HA:μH > μL).The mean square s2b is the appropriate errorterm to test for strain effects assuming that sub-line effects are significant (table I). Subline ef-fects were evaluated using the mean square s2as the error term using a 2-tailed test of signifi-cance. The mean square s2 is also the appropri-ate error term for testing strain effects if sublineeffects are not significant (table I). We report theresults of 1-tailed tests for strain effects usingboth s2b and s2 as error terms.
Evaluation of indicatorsof seasonal colony weight gain
The ability to make effective selections for hon-ey production early in the season is importantbecause it permits the production of the nextgeneration of colonies within the same year,thereby increasing the rate of stock improve-ment. Short-term colony weight gain is one pos-sible indicator of seasonal colony weight gain.Correlation coefficients between seasonal colo-
ny weight gain and weekly (1983), bi-weekly(1984, 1985), and monthly (1986) short-termweight gains were calculated. Early-winter colo-ny weight, early-spring colony weight, and win-ter weight loss (principally honey consumption)were also evaluated as indicators of seasonal
colony weight gain. Correlation coefficientswere calculated separately for the high and lowstrains in 1984, 1985, and 1986.
RESULTS
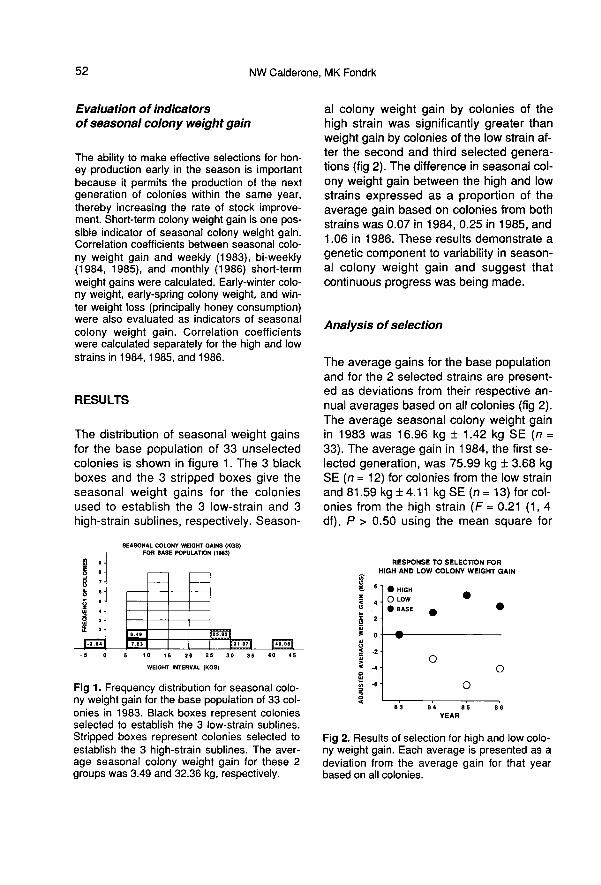
The distribution of seasonal weight gainsfor the base population of 33 unselectedcolonies is shown in figure 1. The 3 blackboxes and the 3 stripped boxes give theseasonal weight gains for the coloniesused to establish the 3 low-strain and 3
high-strain sublines, respectively. Season-
al colony weight gain by colonies of thehigh strain was significantly greater thanweight gain by colonies of the low strain af-ter the second and third selected genera-tions (fig 2). The difference in seasonal col-ony weight gain between the high and lowstrains expressed as a proportion of theaverage gain based on colonies from bothstrains was 0.07 in 1984, 0.25 in 1985, and1.06 in 1986. These results demonstrate a
genetic component to variability in season-al colony weight gain and suggest thatcontinuous progress was being made.
Analysis of selection
The average gains for the base populationand for the 2 selected strains are present-ed as deviations from their respective an-nual averages based on all colonies (fig 2).The average seasonal colony weight gainin 1983 was 16.96 kg ± 1.42 kg SE (n =33). The average gain in 1984, the first se-lected generation, was 75.99 kg ± 3.68 kgSE (n = 12) for colonies from the low strainand 81.59 kg ± 4.11 kg SE (n = 13) for col-onies from the high strain (F = 0.21 (1, 4df), P > 0.50 using the mean square for
subline, s2b, as the error term; and F =0.34 (1,19 df), P > 0.50 using the meansquare s2 as the error term). Subline ef-fects were not significant (F = 1.60 (4, 19df), P > 0.25). The average gain in 1985,the second selected generation, was 44.79kg ± 3.20 kg SE (n = 18) for colonies fromthe low strain and 55.75 kg ± 2.24 kg SE (n= 23) for colonies from the high strain (F =9.46 (1, 2 df), P < 0.05 using the meansquare for subline, s2b , as the error term;and F = 9.49 (1, 37 df), P < 0.005 usingthe mean square error s2 as the error
term). Subline effects were not significant(F = 1.00 (2,37 df), P > 0.20). The averagegain in 1986, the third selected generation,was 3.01 kg ± 1.11 kg SE (n = 16) for colo-nies from the low strain and 10.64 kg ±1.96 kg SE (n = 19) for colonies from thehigh strain (F = 6.05 (1, 3 df), P < 0.05 us-
ing the mean square for subline, s2b, asthe error term; and F = 11.33 (1, 30 df), P< 0.005 using the mean square s2 as the
error term). Subline effects were not signifi-cant (F = 1.87 (3, 30 df), P> 0.15). The dif-ference between the 2 selected strains
was greatest in 1985 (difference betweenstrains = 10.96 kg: average gain based onboth strains = 50.94 kg). However, the dif-ference between the 2 strains expressedas a proportion of the average gain for ayear based on both strains was greatest in1986 (difference between strains = 7.63 kgaverage gain based on both strains = 7.15kg).
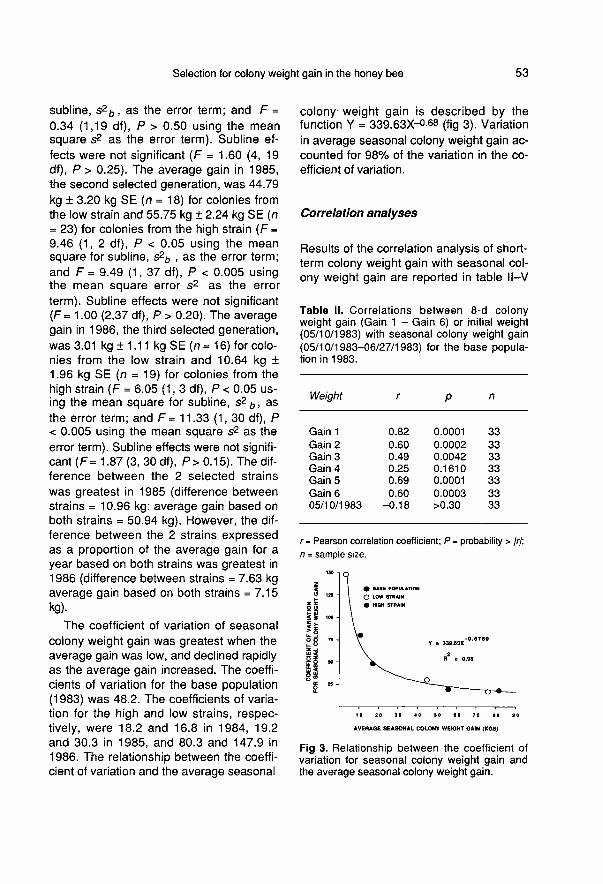
The coefficient of variation of seasonal
colony weight gain was greatest when theaverage gain was low, and declined rapidlyas the average gain increased. The coeffi-cients of variation for the base population(1983) was 48.2. The coefficients of varia-tion for the high and low strains, respec-tively, were 18.2 and 16.8 in 1984, 19.2and 30.3 in 1985, and 80.3 and 147.9 in1986. The relationship between the coeffi-cient of variation and the average seasonal
colony weight gain is described by thefunction Y = 339.63X-0.68 (fig 3). Variationin average seasonal colony weight gain ac-counted for 98% of the variation in the co-efficient of variation.
Correlation analyses
Results of the correlation analysis of short-term colony weight gain with seasonal col-ony weight gain are reported in table II-V
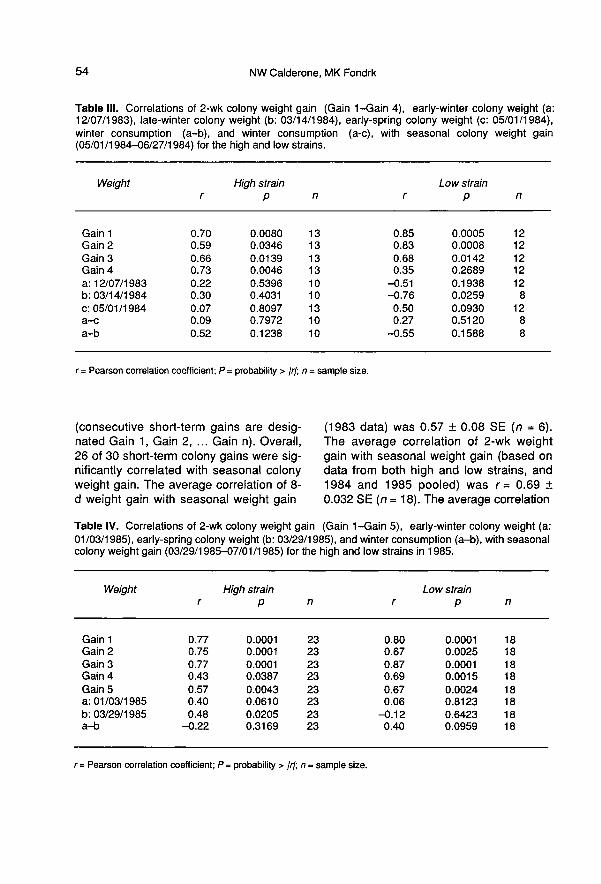
(consecutive short-term gains are desig-nated Gain 1, Gain 2, ... Gain n). Overall,26 of 30 short-term colony gains were sig-nificantly correlated with seasonal colonyweight gain. The average correlation of 8-d weight gain with seasonal weight gain
(1983 data) was 0.57 ± 0.08 SE (n = 6).The average correlation of 2-wk weightgain with seasonal weight gain (based ondata from both high and low strains, and1984 and 1985 pooled) was r = 0.69 ±0.032 SE (n = 18). The average correlation
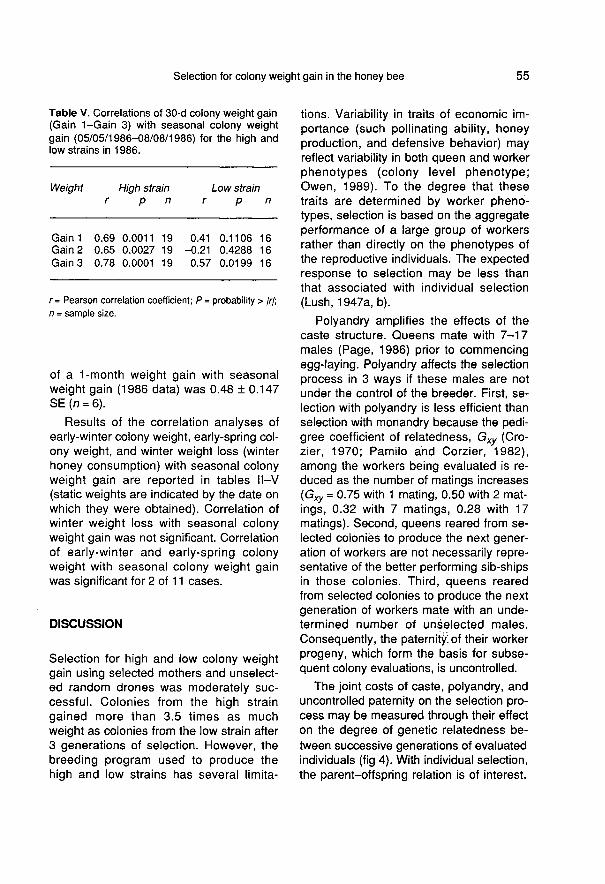
of a 1-month weight gain with seasonalweight gain (1986 data) was 0.48 ± 0.147SE (n = 6).
Results of the correlation analyses ofearly-winter colony weight, early-spring col-ony weight, and winter weight loss (winterhoney consumption) with seasonal colonyweight gain are reported in tables II-V
(static weights are indicated by the date onwhich they were obtained). Correlation ofwinter weight loss with seasonal colonyweight gain was not significant. Correlationof early-winter and early-spring colonyweight with seasonal colony weight gainwas significant for 2 of 11 cases.
DISCUSSION
Selection for high and low colony weightgain using selected mothers and unselect-ed random drones was moderately suc-cessful. Colonies from the high strain
gained more than 3.5 times as much
weight as colonies from the low strain after3 generations of selection. However, thebreeding program used to produce thehigh and low strains has several limita-
tions. Variability in traits of economic im-portance (such pollinating ability, honeyproduction, and defensive behavior) mayreflect variability in both queen and workerphenotypes (colony level phenotype;Owen, 1989). To the degree that thesetraits are determined by worker pheno-types, selection is based on the aggregateperformance of a large group of workersrather than directly on the phenotypes ofthe reproductive individuals. The expectedresponse to selection may be less thanthat associated with individual selection
(Lush, 1947a, b).Polyandry amplifies the effects of the
caste structure. Queens mate with 7-17males (Page, 1986) prior to commencingegg-laying. Polyandry affects the selectionprocess in 3 ways if these males are notunder the control of the breeder. First, se-lection with polyandry is less efficient thanselection with monandry because the pedi-gree coefficient of relatedness, Gxy (Cro-zier, 1970; Pamilo and Corzier, 1982),among the workers being evaluated is re-duced as the number of matings increases(Gxy = 0.75 with 1 mating, 0.50 with 2 mat-ings, 0.32 with 7 matings, 0.28 with 17matings). Second, queens reared from se-lected colonies to produce the next gener-ation of workers are not necessarily repre-sentative of the better performing sib-shipsin those colonies. Third, queens rearedfrom selected colonies to produce the nextgeneration of workers mate with an unde-termined number of unselected males.Consequently, the paternity of their workerprogeny, which form the basis for subse-
quent colony evaluations, is uncontrolled.The joint costs of caste, polyandry, and
uncontrolled paternity on the selection pro-cess may be measured through their effecton the degree of genetic relatedness be-tween successive generations of evaluatedindividuals (fig 4). With individual selection,the parent-offspring relation is of interest.
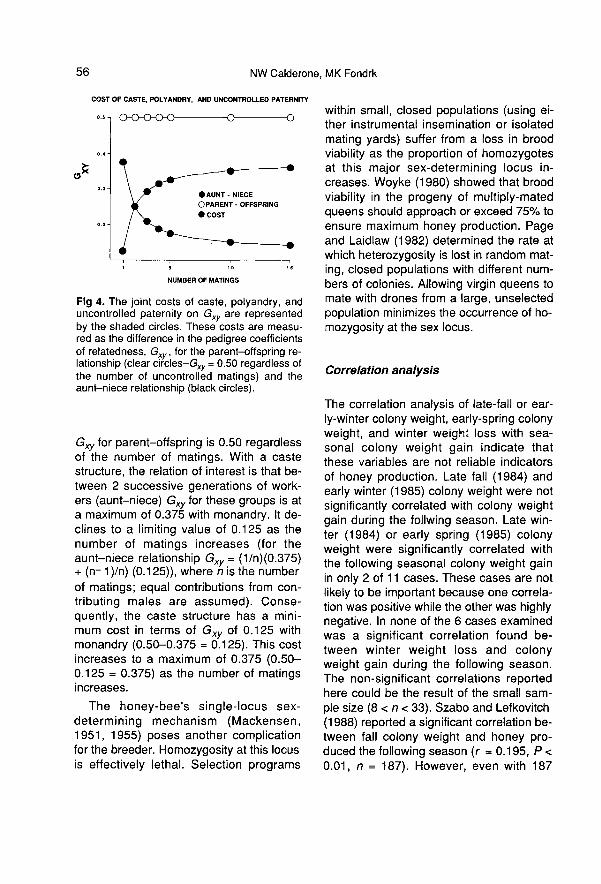
Gxy for parent-offspring is 0.50 regardlessof the number of matings. With a castestructure, the relation of interest is that be-tween 2 successive generations of work-ers (aunt-niece) Gxy for these groups is ata maximum of 0.375 with monandry. It de-clines to a limiting value of 0.125 as thenumber of matings increases (for theaunt-niece relationship Gxy = (1/n)(0.375)+ (n- 1 )/n) (0.125)), where n is the numberof matings; equal contributions from con-tributing males are assumed). Conse-quently, the caste structure has a mini-mum cost in terms of Gxy of 0.125 with
monandry (0.50-0.375 = 0.125). This costincreases to a maximum of 0.375 (0.50-0.125 = 0.375) as the number of matingsincreases.
The honey-bee’s single-locus sex-
determining mechanism (Mackensen,1951, 1955) poses another complicationfor the breeder. Homozygosity at this locusis effectively lethal. Selection programs
within small, closed populations (using ei-ther instrumental insemination or isolated
mating yards) suffer from a loss in broodviability as the proportion of homozygotesat this major sex-determining locus in-
creases. Woyke (1980) showed that broodviability in the progeny of multiply-matedqueens should approach or exceed 75% toensure maximum honey production. Pageand Laidlaw (1982) determined the rate atwhich heterozygosity is lost in random mat-ing, closed populations with different num-bers of colonies. Allowing virgin queens tomate with drones from a large, unselectedpopulation minimizes the occurrence of ho-mozygosity at the sex locus.
Correlation analysis
The correlation analysis of late-fall or ear-ly-winter colony weight, early-spring colonyweight, and winter weight loss with sea-sonal colony weight gain indicate thatthese variables are not reliable indicatorsof honey production. Late fall (1984) andearly winter (1985) colony weight were notsignificantly correlated with colony weightgain during the follwing season. Late win-ter (1984) or early spring (1985) colonyweight were significantly correlated withthe following seasonal colony weight gainin only 2 of 11 cases. These cases are notlikely to be important because one correla-tion was positive while the other was highlynegative. In none of the 6 cases examinedwas a significant correlation found be-tween winter weight loss and colonyweight gain during the following season.The non-significant correlations reportedhere could be the result of the small sam-
ple size (8 < n < 33). Szabo and Lefkovitch(1988) reported a significant correlation be-tween fall colony weight and honey pro-duced the following season (r = 0.195, P <0.01, n = 187). However, even with 187

colonies, the correlation of fall colonyweight with honey production was weak.Fall colony weight is probably most signifi-cant as a determinant of honey productionwhen it is inadequate to sustain the build-up of a colony’s population the followingspring.
The correlation analysis of short-termcolony weight gain with seasonal colonyweight gain revealed that this variable is amoderately useful indicator of seasonal
colony weight gain and confirms previousfindings by Szabo (1982) and Oldroyd et al(1985). However, data from each of thesestudies suggest that there is considerablevariability in this relationship. In the
present study, the average correlation co-efficients were 0.57 (1983), 0.69 (1984 and1985), and 0.48 (1986); Szabo’s (1982)correlation coefficients for 24-h gain withseasonal gain ranged from -0.17 to 0.94(from 0.32 to 0.94 without the single nega-tive coefficient). The correlations calculat-ed by Oldroyd et al (1985) for 1-wk colonyweight gain with seasonal colony weightgain ranged from 0.68-0.81.
The moderate correlation of short-term
colony weight gain with seasonal colonyweight gain, the frequent occurrence of lowand non-significant correlations, and theoccasional negative correlation, suggestthat the identification of additional predic-tive variables would considerably enhancethe effectiveness of selection. Several oth-er variables are correlated with seasonal
colony weight gain or honey production.These include colony population (Farrar,1937; Szabo, 1982; Szabo and Lefkovitch,1989), 24-h pollen collection (Szabo,1982), area of sealed brood (Soller andBar-Cohen, 1967; Szabo, 1982; Szaboand Lefkovitch, 1989), weight of bees in
colony (Szabo, 1982), daily flight activity(Szabo, 1980), queen weight (Nelson andGary, 1983), and several morphologicalcharacteristics (Milne, 1980, 1985; Milne
and Pries, 1984; Szabo and Lefkovitch,1988). Szabo (1982) reported multiple cor-relation coefficients ranging from 0.90-0.98 for 3 variables related to honey pro-duction (24 h pollen collection, 24 h weightgain, and area of capped brood). Szabo’smodel should be evaluated further for its
predictive value and applicability in differ-ent environments.
Analysis of the coefficient of variation
The coefficient of variation for seasonal
colony weight gain was high when the av-erage gain was low, but declined rapidly asthe average gain increased. When the av-erage gain is small, changes in colonyweight are likely to be as indicative of
changes in the amount of stored pollen,brood, and adult population as they are ofchanges in stored honey. Even if stored
honey is important, variability in colonyweight gain may still reflect stochasticevents as well as genetic differencesamong colonies. For example, variability inforaging success in a resource limited en-vironment could result in genetically similarcolonies having large differences in pheno-types. Consequently, when the averagegain is low, the large variance among colo-nies may be primarily the result of environ-mental effects and changes in hive compo-nents other than stored honey. Heritabilitywill be low and the response to selectionwill be small.
We demonstrated successful selectionunder the most difficult of circumstances:no control over male parentage. Withoutselection of the contributing male gametes,both the rate and amount of progress thatcan be achieved is reduced. Completecontrol of parental contributions can be ac-complished through the use of instrumen-tal insemination (Laidlaw, 1987a, b) or iso-lated mating yards (Ruttner, 1988; Waller

et al, 1989). Szabo and Lefkovitch (1987)reported successful selection for increasedhoney production using isolated matingyards. However, these techniques havenot been assimilated into the routines ofmost commercial operations in the UnitedStates. Recently, Hellmich and Waller
(1990) demonstrated that control over themale gametic contributions can beachieved with natural mating through theuse of a drone saturation technique thateliminates the need to identify isolatedmating stations. The technique evaluatedin this study is readily available to any bee-keeper requiring only a knowledge of
queen rearing techniques and the ability toaccurately weigh colonies.
ACKNOWLEDGMENTS
We thank RE Page, WC Rothenbuhler, H Shim-anuki and SSY Young for helpful discussionsconcerning this project. We are grateful to RHellmich, G Hoffman, and two anonymous re-viewers for commenting on the manuscript. Wealso thank The Ohio Agricultural Research andDevelopment Center for constructing and donat-ing the mechanical device required to weigh col-onies. Additional support was provided by theOhio State University James Hambleton Apicul-tural Memorial Award Fund.
Résumé — Sélection d’abeilles (Apismellifera L) ayant un facteur «gain depoids saisonnier de la colonie» fort oufaible, à l’aide de reines sélectionnéeset de mâles pris au hasard. On a effec-tué sur 3 générations une sélection 2-voies pour obtenir des lignées d’abeilles,Apis mellifera, présentant un gain de poidssaisonnier de la colonie (GPSC) fort oufaible. Trente trois colonies formant la po-pulation de base ont été pesées à inter-valles réguliers (gains de poids à courtterme) du début du printemps jusqu’à la finde la miellée principale. Les sélections ont
été faites sur la base du gain total de poidsdurant cette période. Les reines des 3 co-lonies ayant le GPSC le plus élevé ont étéchoisies pour être à l’origine d’une soucheélevée formée de 3 sous-lignées. Lesreines des 3 colonies ayant le GPSC le
plus faible ont été choisies pour être à l’ori-gine d’une souche faible formée de 3 sous-lignées. Des reines filles ont été élevées àpartir de ces colonies, et on les a laisséess’accoupler librement avec des mâles nonsélectionnés dans un rucher isolé de la po-pulation mère. Les colonies obtenues ontété évaluées la saison suivante et la reinede chaque sous-lignée à la tête de la colo-nie ayant le GPSC le plus élevé (sous-lignées les plus élevées) ou le plus faible(sous-lignées les plus faibles) a été sélec-tionnée pour perpétuer cette sous-lignéedans les générations suivantes. Deuxsouches d’abeilles ont ainsi été produites,dont le GPSC différait significativement.Ces résultats suggèrent que les apicul-teurs professionnels ou amateurs peuventaccroître l’efficacité de la production demiel grâce à un schéma simple de sélec-tion et d’accouplement. Néanmoins les
progrès possibles sont limités par le sys-tème d’accouplement utilisé dans ce pro-gramme de sélection. Ces limites sont une
conséquence des effets de la caste, de lapolyandrie et de la paternité non contrôlée.Plusieurs variables ont été évaluées entant qu’indicateurs du PGSC. Le gain depoids de la colonie à court terme est signi-ficativement corrélé avec le PGSC et peutêtre un outil utile pour la sélection. Le gainde poids de la colonie au début de l’hiver,à la fin de l’hiver, au début de printemps etla perte de poids hivernale ne sont pascorrélés avec le PGSC et n’apparaissentdonc pas comme des outils utiles pour lasélection.
Apis mellifera / sélection / rendementde miel

Zusammenfassung — Selektion fürhohe und niedrige Zunahmen desVolksgewichts durch Auswahl der
Königinnen und freie Paarung. EineZweiweg-Selektion mit Bienenvölkern ho-her oder niedriger Gewichtszunahmewährend der Saison wurde drei Genera-tionen lang durchgeführt. Die Ausgangs-population von 33 Völkern wurde vom zeiti-gen Frühjahr bis zum Ende der Haupt-tracht in regelmäßigen Abständen gewo-gen; auf diese Weise kamen wir zu denKurzzeit-Zunahmen. Die Selektion wurdeauf Grund der Gesamtzunahme währendder Saison vorgenommen. Königinnen vonden drei Völkern mit den höchsten Saison-zunahmen wurden zur Gründung von dreiUnterlinien ausgewählt ("Hoch"-Stamm).Aus den drei Völkern mit den niedrigstenGewichtszunahmen während der Saisonwurden Königinnen für drei Unterlinien des"Niedrig"-Stammes gezogen. Die aus die-sen ausgewählten Völkern aufgezogenenJungköniginnen wurden auf isoliertenStänden frei gepaart. In der folgenden Sai-son wurden die nachgezogenenKöniginnen geprüft und die Königinnen mitden jeweils höchsten, bzw niedrigsten Ge-wichtszunahmen des "Hoch"- oder "Nie-
drig"-Stammes für die Nachzucht zur
Erhaltung des Stammes in der nachfolgen-den Generation ausgewählt.
Auf diese Weise wurden zwei Bie-nenstämme erzeugt, die sich signifikant inder Gewichtszunahme während derSaison unterschieden. Diese Ergebnisseweisen darauf hin, daß Erwerbs- undFreizeitimker den Honigertrag durch eineinfaches Selektions- und Paarungs-schema steigern können. Das in diesemSelektionsprogramm benutzte Paarungs-schema schränkt jedoch die erreichbareGeschwindigkeit wie das Gesamtausmaßdes Fortschrittes ein. Diese Begrenzungenergeben sich aus Einflüssen der Kaste, derMehrfachpaarung sowie der unkontrollier-ten Wahl der Väter.
Darüber hinaus wurden mehrere Fakto-ren als Hinweise auf Gewichtszunahmenüber die Saison geprüft. Die Kurzzeit-Zunahmen waren signifikant mit der Ge-samtzunahme während der Saison korre-
liert; sie sind deshalb nützliche Hilfen fürdie Selektion. Das Volksgewicht im
Frühwinter, im Spätwinter, im zeitigenFrühjahr und der Gewichtsverlust währenddes Winters waren hingegen mit der Ge-wichtszunahme während der Trachtzeitnicht korreliert und sie ergeben deshalbkeine brauchbaren Hinweise für die Selek-tion.
Apis mellifera / Selektion / Honigertrag
REFERENCES
Cale GH Jr, Gowen JW (1956) Heterosis in thehoneybee (Apis mellifera L). Genetics 41,292-303
Crozier RH (1970) Coefficients of relatednessand the identity of genes by descent in theHymenoptera. Am Nat 104, 216-217
Farrar CL (1937) The influence of colony popu-lations on honey production. J Agric Res 54,945-954
Hellmich RL Jr II, Waller GD (1990) Preparingfor Africanized honeybees: evaluating controlin mating apiaries. Am Bee J 130, 537-542
Kempthorne O (1952) The Design and Analysisof Experiments. John Wiley and Sons, NY
Laidlaw HH (1987a) Instrumental inseminationof honeybee queens: its origin and develop-ment. Bee World 68, 17-36
Laidlaw HH Jr (1987b) Instrumental insemina-tion of honeybee queens: its origin and devel-opment . Bee World 68, 71-88
Lush JL (1947a) Family merit and individual se-lection as bases for selection. Part I. Am Nat81, 241-261
Lush JL (1947b) Family merit and invidual selec-tion as bases for selection. Part II. Am Nat81, 362-379
Mackensen O (1951) Viability and sex determi-nation in the honey bee (Apis mellifera L).Genetics 36, 500-509

Mackensen O (1955) Further studies on a lethalseries in the honey bee. J Hered 46, 72-74
McLellan AR (1977) Honeybee colony weightgain as an index of honey production andnectar flow: a critical evaluation. J Appl Ecol14, 401-408
Milne CP Jr (1980) Laboratory measurement ofhoney production in the honeybee. III. Pupalweight of the worker. J Apic Res 19, 176-178
Milne CP Jr (1985) The need for using labora-tory tests in breeding honeybees for im-proved honey production. J Apic Res 24,237-242
Milne CP Jr, Pries KJ (1984) Honeybee corbicu-lar size and honey production. J Apic Res23, 11-14
Moeller FE (1961) The relationship between col-ony populations and honey production.USDA-ARS and WI Agric Exp Stat Prod ResRep No 55
Nelson DL, Gary NE (1983) Honey productivityof honeybee colonies in relation to bodyweight, attractiveness and fecundity of thequeen. J Apic Res 22, 209-213
Oldroyd BP, Moran C, Nicholas FW (1985) Dial-lel crosses of honeybees. I. A genetic analy-sis of honey production using a fixed effectsmodel. J Apic Res 24, 243-249
Owen RE (1989) The effects of colony-level se-lection on social behavior. In: Genetic and
Social Evolution (Breed MD, Page RE, eds)Westview Press, Boulder, CO
Page RE Jr (1986) Sperm utilization in social in-sects. Annu Rev Entomol 31, 297-320
Page RE Jr, Laidlaw HH Jr (1982) Closed popu-lation honeybees breeding. I. Population ge-netics of sex determination. J Apic Res 21,30-37
Pamilo P, Crozier RH (1982) Measuring geneticrelatedness in natural populations: methodol-ogy. Theor Popul Biol 21, 171-193
Peer DF (1957) Further studies on the matingrange of the honey bee, Apis mellifera L. CanEntomol 89, 108-110
Rothenbuhler WC (1980) Necessary links in thechain of honey bee stock improvement. AmBe J 120, 223-225, 304, 305
Ruttner F (1988) Breeding Techniques and Se-lection for Breeding of the Honeybee. TheBritish Isles Bee Breeders Association, Mu-nich
Sokal RR, Rohlf FJ (1981) Biometry. WH Free-man and Company NY, 2nd edn
Soller M, Bar-Cohen R (1967) Some observa-tions on the heritability and genetic correla-tion between honey production and broodarea in the honeybee. J Apic Res 6, 37-43
Szabo T (1980) Effect of weather factors onhoneybee flight activity and colony weightgain. J Apic Res 19, 164-171
Szabo T (1982) Phenotypic correlations be-tween colony traits in the honey bee. Am BeeJ 122, 711-716
Szabo T, Lefkovitch LP (1987) Fourth genera-tion of close-population honeybee breeding.I. Comparison of selected and control strains.J Apic Res 26, 170-180
Szabo T, Lefkovitch LP (1988) Fourth genera-tion of closed population honeybee breeding.II; Relationship between morphological andcolony traits. Apidologie 19, 259-274
Szabo T, Lefkovitch LP (1989) Effect of broodproduction and population size on honey pro-duction on honeybee colonies in Alberta,Canada. Apidologie 20, 157-163
Waller GD, Hoopingarner RA, Martin JH, LoperJM, Fierro M Meliton (1989) Controlled natu-ral matings of honey bee queens in southernArizona. Am Bee J 129, 187-190
Woyke J (1980) Effect of sex allele homo-
heterozygosity on honeybee colony popula-tions and on their honey production. I. Fa-vourable development conditions and unre-stricted queens. J Apic Res 19, 51-63