Embed Size (px)
Citation preview

The Mechanism of Bursting Pacemaker
Activity in Neurons of the Mollusc
Tritonia diomedia
by
Stephen J. Smith
A dissertation submitted in partial fulfillment
of the requirements for the degree of
Doctor of Philosophy
University of Washington
1978
Approved by
(Chairperson of Supervisory Committee)
Program Authorizedto Offer Degree
Date

UNIVERSITY OF WASHINGTON
Date: June 2, 1977
We have carefully read the dissertation entitled The Mechanism of Bursting PacemakerActivity in Neurons of the Mollusc Tritonia Diomedia
submitted byStephen J. Smith
in partial fulfillment of the requirements of the degree of Doctor of Philosophy and recommend its acceptance. In support of this recommendation we present the followingjoint statement of evaluation to be filed with the dissertation.
This work provides an experimental and theoretical analysis of themembrane mechanism underlying the generation of recurrent impulse "bursts"in certain re-identifiable molluscan neurons. The voltage-clamp techniquewas used to characterize the three ionic channels of major importance ininitiating and terminating a burst of impulses. Among these channels, oneis, on pharmacological grounds, a Ca-permeable channel. The two othersare, on the basis of ion substitution experiments, permeable to sodium/calciumand to potassium, respectively. On the basis of previous work by Thompson,it is suggested that this potassium channel is regulated not directly bythe membrane potential but instead by the intracellular Ca++-concentration.In a theoretical section, Ca++-inward currents, the intracellular Ca++-concentration and the properties of the Ca++-sensitive potassium channelare combined with voltage-clamp data of Thompson and of Connor and Stevensin order to reproduce the electrical activity of bursting neurons.
This work is of significance in that it explains how a single nervecell can, without synaptic input, generate an impulse pattern of greatcomplexity.
4)DISSERTATION READING COMMITTEE.

Doctoral Dissertation
In presenting this dissertation in partial fulfillment of the requirements
for , the Doctoral degree at the University of Washington, I agree that the
Library shall make its copies freely available for inspection. I further
agree that extensive copying of this dissertation is allowable only for
scholarly purposes. Requests for copying or reproduction of this disser-
tation may be referred to University Microfilms, 300 North Zeeb Road, Ann
Arbor, Michigan 48106, to whom the author has granted "the right to repro-
duce and sell (a) copies of the manuscript in microform and/or (b) printed
copies of the manuscript made from microform."
Signature
Date

University of Washington
Abstract
THE MECHANISM OF BURSTING PACEMAKER ACTIVITY
IN NEURONS OF THE MOLLUSC TRITONIA DIOMEDIA
By Stephen J. Smith
Chairperson of the Supervisory Committee: Professor Wolfhard AlmersDepartment of Physiology and
Biophysics
Bursting pacemaker activity was studied in pleural ganglion cell bodies
of the nudibranch mollusc Tritonia diomedia. The work described here is
concerned with the mechanism of the endogenous slow alternation of episodes
of repetitive firing and silence characteristic of bursting pacemaker neurons.
Membrane electrical properties were studied using microelectrode voltage
clamp techniques. Implications of the voltage clamp observations were
investigated by mathematically modelling the observed membrane currents and
then reconstructing membrane potential trajectories. This work is presented
here in three parts.
Part I is an analysis of the prolonged tails of membrane current observed
following depolarizations of voltage clamped bursting pacemaker neurons. The
dependence of these current tails on membrane potential, time, and external
ions indicates that they consist of two distinct slowly decaying components.ti
A normally inward component designated I B activates maximally in less than
one second at positive potentials and decays exponentially with a time con-
stant of 2 - 4 seconds upon return to a negative holding potential. I B is
apparently carried by both sodium and calcium ions. A normally outward cur-
rent component designated I C activates progressively during depolarizations
lasting up to several seconds. The decay of I C upon repolarization has a
very prolonged, non-exponential time course, remaining at detectable levels
for over a minute following long activating pulses. I C evidently reflects a

calcium-activated potassium permeability mechanism: the current is carried
mainly by potassium ions, but activation upon depolarization appears to be
mediated by a voltage-dependent influx of calcium ions. Inward calcium
currents were therefore studied to allow development of a model for the
linkage of calcium influx to I activation. Though the slow current IB
should contribute to calcium influx, a much larger and more rapidly gated
calcium current, designated I D , has also been identified.
Part II develops a set of equations describing the total ionic current
observed in Tritonia bursting pacemaker neurons. The ionic current is repre-
sented as the sum of seven components, distinct in their activation kinetics
and their instantaneous current-voltage relationships. In addition to the
three ionic currents discussed in part I, the seven components include a fast,
inactivating sodium current, II' two voltage-dependent potassium currents, IK
and IA, and a linear leakage current, IL . The descriptions of each component
are based quantitatively on voltage clamp measurements described in part I or
in previously published work. Five of the currents are described using vol-
tage-dependent conductance equations of the form introduced by Hodgkin and
Huxley's analysis of the axonal action potential. The activation kinetics
for I are described using a different formalism which explicitly represents
an involvement of calcium ions. Calculations based on published estimates of
parameters relating to neuronal intracellular calcium metabolism indicate
that the slow kinetics of I activation may directly reflect the kinetics of
calcium ion accumulation near the inner membrane surface.
In part III, the membrane current equations are combined and solved
numerically to predict membrane potential under conditions corresponding to
an unclamped, spatially isopotential membrane region. The resulting solution
is a spontaneous oscillation closely resembling naturally occurring bursting
pacemaker activity: both action potential firing and the slow alternation of

bursts and silent episodes are reproduced. The two slow currents, IB and IC ,
are both necessary to reconstruct bursting activity. The slow variation of
the intracellular calcium ion concentration governing I c activation determines
the long period of the reconstructed pacemaker oscillation. The mathematical
reconstruction constitutes a detailed hypothesis regarding the mechanism of
bursting pacemaker activity and shows that the current components identified
by the voltage clamp analysis are quantitatively sufficient to account for
the phenomenon.

TABLE OF CONTENTS
List of Tables iv
List of Figures
TITLE PAGE
INTRODUCTION 1
Molluscan bursting pacemaker neurons 1
Previous studies of the mechanism of the burst oscillation 4
Rationale for the present study 8
METHODS 11
Voltage clamp studies 11
Description of membrane current 14
Reconstruction of membrane potential 15
RESULTS I: SLOW RELAXATIONS OF VOLTAGE CLAMP CURRENT 17
Characteristics of slow current tails 17
The slow inward current, IB 20
The slow outward current, I C 21
The time course of slow potassium permeability changes 27
The role of calcium entry in the activation of I C 32
The requirement for calcium entry 32
The dependence of calcium entry on time and voltage 34
DISCUSSION OF SLOW CURRENT TAIL ANALYSIS 40
The slow inward current I B 40
The calcium-activated potassium current I C 41

TITLE PAGE
RESULTS II: A MATHEMATICAL MODEL OF THE MEMBRANE CURRENT 45
Multiple component description of membrane current 45
Capacity current 49
The fast sodium current, I I 50
The fast calcium current, ID 54
The slow inward current, I B 57
The delayed potassium current, IK 60
The transient potassium current, IA 63
The calcium-activated potassium current, I C 66
A calcium-coupled model for the activation of I C 69
(1)calcium influx 70
(2)calcium efflux 70
(3) diffusion and binding in cytoplasm 71
Solving for the dependence of Cai(r,t) on time and voltage 73
Comparisons of Cai (a,t) with gc 77
The linear leakage current, IL 80
RESULTS III: THE RECONSTRUCTION OF BURSTING PACEMAKER ACTIVITY 82
Conditions for the reconstruction of membrane potential 82
Features of the reconstructed potential waveform 84
The time course of ionic currents and intra-cellular calcium86during the reconstructed burst cycle
DISCUSSION OF MODEL RESULTS 90
The mechanism of bursting pacemaker-like activity in the model 90
(1)"background" excitability 90
(2)rate-limiting conductance charges 91
Bibliography 95
iii

LIST OF TABLES
TABLE TITLE PAGE
I Seven components of ionic current observed in 46bursting pacemakers.
II Outline of model for intracellular calcium ion 74metabolism.

LIST OF FIGURES
FIGURE TITLE PAGE
1 Intracellular recordings of bursting pacemaker activity 2
2 Procedure for measurement of slow tail currents 18
3 The effect of external potassium concentration on slowtail currents
23
4 The effect of holding potential on the slow currenttail waveform.
25
5 Comparison of tail current and slope conductance timecourses.
26
6 Procedure for estimating the time course of membranepotassium permeability.
28
7 Time course of potassium-dependent current differencesafter pulses of varying durations.
31
8 The effect of Cow and lowered Cam on the slow tailcurrent.
33
9 Inward currents in a bathing solution containing mad . 37
10 Gating parameters: gI . 52
11 Gating parameters: gd. 55
12 Gating parameters: gm. 59
13 Gating parameters: gm. 62
14 Gating parameters: gm. 65
15 Predictions and recordings of decays at -40 mV. 68
16 Prediction of intracellular free calcium concentrationresponse to a depolarizing pulse
76
17 The voltage dependence of and predictions ofcalcium accumulation.
79
18 Reconstructed and recorded membrane potential waveforms. 83
19 Time courses of selected variables during the recon-structed burst cycle.
87
20 Time courses of ionic currents during reconstructedaction potential firing.
89

mCmNOWLEDGMENTS
This dissertation represents a part of a larger effort undertaken
in collaboration with my good friend, Dr. Stuart H. Thompson. Stuart's
1976 University of Washington doctoral dissertation documents another
part of this same effort. moth of our thesis projects grew out of
exploratory experiments we performed together at the Friday Harbor
Laboratories in the fall of 1974. Stuart and I recognized that the
insights gained from our early joint experiments could be pursued most
effectively if we continued to coordinate our efforts. mccordingly,
we embarked on the closely related projects described in our two disser-
tations. Each document reflects the individual author's contributions
to the collaboration, but includes extensive reference to the work of
the other partner. While the experiments, mathematical formulations,
and computations presented in detail here were designed and executed
by myself, I wish to emphasize that the broad concept of the work is
fundamentally a product of the collaboration between myself and Stuart
Thompson.
This project was begun under the supervision of Dr. Charles F.
Stevens. When Dr. Stevens moved from Washington to Yale University,
Dr. Wolfhard mlmers accepted the difficult task of supervising my
efforts at bringing the ongoing project to completion. I am deeply
grateful to both men for their multifaceted advice and support, and
for the examples they each set for me. I would also like to thank Dr.
Arthur C. mrown, Dr. mertil Hille, Dr. L. Donald Partridge, and Dr.
A.O.D. Willows for their advice and support of this project. Finally,
I am grateful for especially helpful discussions to Dr. John Connor,
Phillip Lloyd, Robert Snow and Paul Taghert.

INTRODUCTION
Ever since the introduction of techniques for recording the activity of
individual nerve cells, it has been obvious that the temporal patterning of
action potentials into rhythmic sequences is of central importance to the
coding and transmission of information in nervous systems. Temporal pat-
terns of unit activity clearly related to sensory or motor functions are
observed at various levels in the nervous systems of many different species.
The study of rhythmicity in nervous systems can readily be pursued to the
level of individual cells: it has been shown that certain neurons fire in
endogenously rhythmic patterns even after complete isolation from all other
neurons and any external input. This thesis is a detailed analysis of the
mechanism underlying one such endogenous rhythmic firing pattern in some
identified giant neurons of the nudibranch mollusc, or sea slug, Tritonia
diomedia.
Molluscan bursting pacemaker neurons
The giant ganglion cells of gastropod molluscs have proven to be espe-
cially suitable material for studies of the physiology of individual central
nervous system neurons. Their large size (some are over 500 pm in diameter)
facilitates many microtechniques, including multiple micro-pipette impale-
ment, which are generally much more difficult or impossible to apply to the
smaller neurons found in most other species. mnother important advantage to
working with gastropod ganglion cells is that many identifiable neurons with
constant physiological properties can be recognized reliably from one indi-
vidual preparation to the next. The subjects of the present study are the
three identified bursting pacemaker cells, LP12, LP13 and RP12 (Willows,
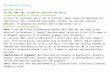
Dorsett and Hoyle, 1973) found in the pleural ganglia of Tritonia. Fig. 1

Fig. 1: Intracellular recordings from six different Tritonia
bursting pacemaker cells. The top four traces represent fully
developed bursting pacemaker activity. The two lower traces show
less common firing patterns which are transitional between bursting
and beating (regularly firing) pacemaker activity.

a
2

3
MINNS intracellular microelectrode recordings of the electrical activity in
several of these cells.
The characteristic bursting pacemaker firing pattern is distinguished
by the spontaneous occurrence of bursts of action potentials alternating
with electrically silent intervals. mctivity patterns like those exempli-
fied by Fig. 1 can persist continuously for hours or days under appropriate
experimental conditions. mursting pacemaker neurons similar to those
observed in Tritonia have been observed in many other species of gastropods.
Perhaps the most widely studied is a large, white cell in the parieto-
visceral ganglion of the sea hare, mplysia californica. This cell has been
designated variously "mr" (Arvanitaki and Chalazonitis, 1958), "the para-
bolic burster" (Strumwasser, 1965), and "R15" (Frazier et al., 1967).
Bursting pacemaker activity has also been observed in cells of terrestrial
molluscs (see Gola, 1974), although in such species the appearance of the
bursting pattern sometimes depends on the season and environmental factors
(Gainer, 1972a,b).
The endogenous nature of the bursting pacemaker rhythm was first
directly demonstrated by mlving's (1968) recordings of bursts in R15 somata
isolated by axonal ligation from all possible synaptic input. The same
conclusion was suggested earlier by Strumwasser's (1965) observation that
the phase of the burst rhythm in R15 could be changed by the injection of
current into the soma, ruling out the possibility that the cells merely fol-
low an oscillatory synaptic input. Observations confirming the endogenous
origin of bursting pacemaker rhythms in many other species, including
Tritonia, have since been reported Mater and Kaneko, 1972; Gainer, 1972b;
Thompson, 1576).

4
The physiological function of the bursting cells in Tritonia has not
Pet been determined. In mplysia and in the land snail, Otala lactea, ana-
tomical evidence strongly suggests that the cells serve a neurosecretory
function (Frazier et al., 1967; Gainer, 1972b). Several reports have indi-
cated that secretion by R15 in mplysia may be involved in body water or ion
regulation (Stinnakre and Tauc, 1966, 1969; Kupfermann and Weiss, 1976). m
bursting cell in the land-..slug, Limax, on the other hand, has been shown to
'observe a motor function, driving contractions of the salivary gland muscu-
lature (Prior and Gelperin, 1977). Burst firing patterns have been shown to
be subject to both short and long term modulation by synaptic inputs (mryan-
ltski and Chalazonitis, 1961; Pinsker and Kandel, 1967; Parnas, mrmstrong
and Strumwasser, 1974) and can also be modified by the introduction of cer-
tain specific peptides at concentrations on the order of 109 M (Barker,
Ifshin and Gainer, 1975).
Previous studies of the mechanism of the burst oscillation
The mechanism of the slow burst oscillation has been investigated
intensively in recent years, but has remained imperfectly understood. It is
clear that the observed slow variation in membrane potential is a necessary
step in the cycle of events that maintain the oscillation. It has already
been noted that the rhythm can be modified by injected current, which pre-
sumably acts only to change the membrane potential. With sufficient tonic
hyperpolarizing current the oscillation can be suppressed completely (mryan-
itaki and Chalazonitis, 1961). Similarly, no slow oscillations of membrane
current are observed when membrane potential is held constant by voltage
clamping (Carnevale, 1974; Gola, 1974; Smith, marker and Gainer, 1975).

5
These observations rule out the possibility that the rhythmic variations in
membrane potential merely reflect some underlying, autonomous (e.g., meta-
bolic) oscillation not directly involving a membrane potential-sensitive
mechanism (but see Gradmann and Slayman, 1975, for an example of such a
mechanism in a mutant strain of Neurospora crassa).
Junge and Stephens (1973) showed evidence from current injection exper-.
iments indicating that a slow variation in membrane potassium conductance
occurs during the burst cycle. Subsequent voltage clamp experiments by
Cola (1974) and T. Smith, marker and Gainer (1975) have supported this con-
clusion, and demonstrated that the potassium conductance involved reflects a
time-and voltage-dependent mechanism with intrinsically slow kinetics of
activation and decay. The slow potassium conductance is activated at depol-
arized potentials and decays after bursts on a time scale comparable to the
normal interburst silent intervals. These authors have suggested, therefore,
that this potassium conductance is fundamental to the mechanism of bursting,
its slow gating kinetics acting as the rate limiting step to determine the
low frequency of the burst oscillation.
R. W. Meech and coworkers have developed several lines of evidence sug-
gesting that membrane potassium conductance in molluscan neurons is con-
trolled partially by intracellular calcium ions. Since depolarization of
molluscan neural somata is known to produce a significant influx of calcium
ions (Geduldig and Junge, 1968; Geduldig and Gruener, 1970), Meech and
coworkers have suggested that a component of the potassium activation
observed during depolarization is activated by the accumulation of inward
current carrying calcium ions near the inner membrane surface (Meech and
Strumwasser, 1970; Meech, 1972, 1974a; Meech and Standen, 1975). Meech

6
(1974b) and Eckert and Lux (1976) have suggested that the activation kinetics
of the slow potassium conductance may directly reflect the kinetics of the
accumulation and dissipation of calcium ions inside the cell. mccording to
this view, the slow oscillation of membrane potential characteristic of
bursting activity would occur in intimate connection with an oscillation in
the level of free calcium inside the cell.
The existence of an intracellular calcium oscillation phase locked to
bursts in cell R15 has now been ascertained using both the dye mrsenazo III
(Gorman and Thomas, 1977) and the photoprotein aequorin (Zucker, personal
communication) as optical calcium indicators. In both cases, the optical
signals were of the type expected according to the postulate of a calcium-
mediated potassium conductance. In addition, Gorman and Thomas (1977)
showed that the microinjection of calcium ions into the soma of R15 produced
the expected parallel waveforms of calcium concentration, measured optically,
and potassium conductance, measured as voltage clamp current. Another test
of the calcium oscillation hypothesis, however, has been more equivocal. If
calcium entry is necessary for the activation of a potassium conductance
essential to the slow oscillation, one would expect bursting to require
external calcium ions. mursting pacemaker activity does disappear when
Tritonia neurons are bathed in media of nominally zero calcium concentration
(Smith, unpublished). In neurons of mplysia californica and Otala lactea,
on the other hand, burst-like oscillations in nominally calcium-free media
have been reported (Junge and Stephens, 1973; marker and Gainer, 1975).
Barker and Gainer (1975) showed that such oscillations disappeared when the
divalent ion chelating agent EDTA (ethylene diamine tetraacetic acid) was added
to the bathing medium, so the persistence of slow oscillations in the

7
nominal zero-calcium media might be explained as an effect of residual cal-
cium.
The slow, calcium-mediated potassium conductance may well determine the
duration of the burst cycle, but even so cannot entirely explain the
phenomenon of bursting pacemaker activity. Potassium conductances with similar
properties have been observed in many neurons that do not fire in bursts
(Connor and Stevens, 1971c; mrodwick and Junge, 1972; Meech, 1975; Gola,
1974; Partridge and Stevens, 1976), so some other factor must cooperate with
slow potassium currents to produce bursting. Several authors (Wilson and
Wachtel, 1974; Gola, 1974; Eckert and Lux, 1976) have suggested that a
region of negative or near-zero slope in the steady-state voltage clamp
current-voltage relationship may be a prerequisite for bursting. Partridge
(1975), however, has shown negative slopes, as in bursters, in neurons that
also have a slow potassium conductance similar to that of bursters (Smith,
unpublished), but themselves do not normally fire in bursts. This finding
indicates that some other factor still may be necessary to differentiate
bursters and to fully explain the bursting pattern.
my activating a voltage clamp circuit at different times during bursts
of action potentials in cell R15, Gola (1974) discovered a slowly decaying
inward current which is activated preferentially to the slow potassium cur-
rent early in the burst. Gola did not identify the ionic basis of this
inward current, but suggested that it might cooperate with the slow potas-
sium current to generate the slow oscillation characteristic of bursting
pacemaker activity. S. H. Thompson and I (Thompson and Smith, 1976) have
shown that depolarizing afterpotentials (DmPs) several seconds in duration
follow driven individual spikes in bursting pacemakers. The slow time-

8
course of DmPs suggests that they very probably reflect the same mechanism
as the slow inward current studied by Gola (1974); our analysis indicates
that this current is carried partly by sodium ions and partly by calcium
ions. DAPs were observed in the bursting pacemaker neurons of all five
gastropod species we studied, but were not observed in any non-bursting
neurons, suggesting a possible causal role in bursting pacemaker activity.
Rationale for the present study
The work described in this thesis has been aimed at advancing a com-
plete and quantitative description of bursting pacemaker activity in terms
of underlying membrane properties. The rationale for the present effort is
borrowed from Hodgkin and Huxley's (1952) analysis of the nerve action
potential. Hodgkin and Huxley showed that the main features of the action
potential could be reconstructed from a mathematical model based strictly on
the analysis of voltage clamp data. The voltage clamp technique allowed
Hodgkin and Huxley to analyse the complex ionic permeability properties of
the squid axon membrane into three relatively simple ionic current compo-
vents, with well defined ionic selectivities and activation or gating
kinetics. From voltage clamp data, they developed aset of equations to
describe the time and voltage dependencies of each ionic current component.
The equations for the three current components were then combined and solved
for the condition of constant total membrane current, corresponding to the
natural, unclamped condition of a small, isolated patch of membrane. The
resulting solutions for membrane potential closely resembled recorded mem-
brane action potentials. This mathematical reconstruction established that
Hodgkin and Huxley's voltage clamp analysis had in fact encompassed the
membrane properties essential to action potential firing, and provided a

9
detailed hypothesis as to exactly how the individual current components
cooperate to produce the action potential. The rationale for Hodgkin and
Huxley's analysis revolves around the central role played by membrane poten-
tial in linking the various processes underlying axonal excitation. Evi-
dence discussed above indicates that membrane potential plays a similar role
in the burst oscillation, so I have adopted a strategy similar to Hodgkin
and Huxley's to approach questions concerning which membrane electrical
properties are essential to bursting pacemaker activity, and how current
components identified by voltage clamp analysis could produce slow oscilla-
tions of membrane potential.
The results presented here are divided into three parts. Part I is an
analysis of the slow relaxations of voltage clamp current observed in burst-
ing pacemaker neurons. In part II, a descriptive mathematical model of the
total membrane current is developed on the basis of voltage clamp data pre-
sented in part I in addition to some previously published results. In part
III, this descriptive model is used to reconstruct membrane potential wave-
forms which reproduce many of the main features of bursting pacemaker activ-
ity. The model presented in parts II and III is similar in many respects to
that developed by Hodgkin and Huxley, particularly in that it is developed
as an empirical description of voltage clamp data. The model presented here
differs from Hodgkin and Huxley's in that it has been necessary to represent
a larger number of distinct components of ionic currents, as might be
expected considering the greater complexity of bursting pacemaker activity
compared to axonal action potential firing. It has also been necessary here
to consider the calcium mediation of potassium activation, whereas in the

10
squid axon it is apparently appropriate to describe each current component
as depending directly on membrane potential and time.
Other mathematical models reproducing bursting pacemaker activity have
now appeared in the literature (Both, Finger and Chaplain, 1976; Plant and
Kim, 1976; Gulijarani, Roberge and Matthieu, 1977). The model developed in
this thesis differs in many respects from each of these other efforts, but
the most important distinction is that only the model presented here was
developed strictly from voltage clamp measurements. The model proposed by
Both et al. (1976) does not address the question of the origin of the slow
oscillation at all, it merely assumes a slow cyclic driving function. The
models proposed by Plant and Kim (1976), and Gulijarani et al. (1977), pro-
pose membrane mechanisms which could account for slow oscillations, but the
implications of these models are limited by the fact that important kinetic
parameters were simply chosen to insure oscillation, without experimental
constraints. The major strength of the model presented in parts II and III
of this thesis is that it leads to predictions of burst-like oscillations
from parameters based entirely on voltage clamp measurements.

METHODS
Voltage clamp studies
Three identified giant cells in the pleural ganglia of the sea slug
Tritonia diomedia reliably exhibit bursting pacemaker activity; these
cells are designated RP12, LP12 and LP13 (Willows, Dorsett and Hoyle, 1973).
The three cells are similar to each other in appearance, normal firing
patterns, and all membrane properties that have been studied. The experi-
ments described below have been performed using a two microelectrode voltage
clamp applied to one of the three cells specified in an isolated ganglion
preparation.
Specimens of Tritonia were obtained by trawling in waters near the
San Juan Islands of Washington State. Animals were maintained in a
recirculating natural sea water system at 10 C prior to use. The cerebral-
pedal-pleural ganglion complex was excised with nerve root stumps 1 to 5 mm
in length and pinned to the waxed bottom of a recording and perfusion cham-
ber. The chamber was maintained at a temperature between 10 °C and 11°C.To facilitate microelectrode penetration, the epineureal sheath was exposed
for 15 min to a 0.5%, by weight, solution of collagenase (Worthington
Biochemicals) in the normal bathing solution described below. Preliminary
investigations showed no effect of this enzyme treatment on the cell's
firing patterns or on behavior under voltage clamp conditions. Cells fail-
ing to recover normal bursting pacemaker activity within 30 min after pene-
tration by the two microelectrodes required for voltage clamping were
rejected without further study.
The voltage clamp electronics employed were similar to those described
by Connor and Stevens (1971a), except for two modifications designed to

12
improve control of membrane potential during long depolarizing pulses:
(1) membrane potential was recorded differentially between an intracellular
microelectrode and a voltage reference electrode in the bathing solution,
and (2) the final output amplifier was modified to provide increased gain
at low frequencies, as described by Dionne and Stevens (1975). These
modifications were necessary because the current electrodes tend to polarize
and increase in resistance while passing large and sustained clamp currents.
The internal current and voltage electrode micropipettes were filled with
3 M KC1 and had tip resistances between 2 and 4 N2. All electrodes were
Ag-AgCl in either 3 M KC1 (micropipettes) or sea water (bath electrodes).
Membrane potential and clamp current recordings were stored on FM
magnetic tape. Clamp current was recorded on two separate tracks of the
tape: one track at low amplification to record the large currents flowing
during depolarizing pulses, the other track at high amplification to record
small tail relaxations near zero current with a minimum of noise and drift.
Tape recorded data was played back onto a pen recorder or digitized by a
computer system for display and analysis. A first order low pass filter
(time constant = 24 msec) was used to reduce noise in records of slow
current relaxations.
Solution changes were effected by introducing 10 ml of the new solution
at one end of the recording chamber, while the bath volume was maintained at
0.9 ml by a suction tube located at the opposite end of the chamber. The
normal bathing solution is similar to sea water, with the following composi-
tion: 420 mM-NaC1, 10 mM-KC1, 23.5 mM-MgC1 2 , 25.5 mM-MgSO 4 , 10 mM-CaC12 ,
2 mM-NaHCO. The various altered bathing solutions were the same except3.

13
for the following specified changes: High K+
solution, 390 mM-NaC1,
40 mM-KC1; Cobalt solution, 2.5 mM-MgC1 1 , 1 mM-CaC1 2, 30 mM-CoC1 2 ; marium
solution, OmM-NaC1, 320 mM-Tris C1,100mM-TEm Cl, 49 mM-MgC1 2, OmM-MgSO4 ,
OmM-CaC12' 10mM-maC1
2.
Tritonia bursting pacemaker neurons consist of a roughly spherical
cell body or soma, approximately 300 pm in diameter, and a slender axonal-
dendritic process which ramifies in the neuropil region of the ganglion and
also sends axons out one or more nerve roots. In the present studies,
voltage-sensing and current-passing electrodes were inserted into the soma.
Such electrode placement results in good control of membrane potential in
the soma region of the cell (see Geduldig & Gruener, 1970; Connor & Stevens,
1971a), but not in the axonal-dendritic process.
Two effects of the poor clamping of distant axonal membrane are evident
in whole 'cell voltage clamp current records: (1) capacity transients are
complex and prolonged (up to 10 msec for 90% complete settling after small
voltage steps), presumably reflecting the axoplasmic resistance in series
with part of the membrane capacitance, and (2) at clamping potentials in
the range of spike threshold, small oscillations of membrane current,
presumably due to repetitive action potential firing in electrically remote
axonal regions of the cell, can often be observed. Various schemes for
measuring membrane current across the well-clamped somatic membrane in
isolation from the axonal cable current have been described (see Connor and
Stevens, 1971a; Neher, 1971; Kado, 1973; Kostyuk, Krishtal and Pidoplichko,
1975; Connor, 1977; Lee, mkaike and mrown, 1977), but do not appear to be
necessary in the context of the present investigation. Preliminary in-
vestigations have shown that the slow current tails described below have

14
much the same appearance in intact cells and in microsurgically isolated
somata, indicating that the slow relaxations reflect mainly the behavior
of the soma membrane, which is clamped well in both the intact cell and
the isolated soma preparations. The only obvious manifestations of poor
clamping in the axon region of intact cells, the long capacity transients
and the small current oscillations at moderate depolarizations, do not
interfere with observations of slow current relaxations in the pacemaker
potential range following depolarizing activation pulses. mn intact cell
preparation was used in the present study because such cells display stable
bursting pacemaker activity more reliably than isolated somata.
Description of membrane current
The mathematical description of membrane current developed in part II
of the results below is based on the analysis and measurement of voltage
clamp data presented in part I of the results and in several previously
published studies (Thompson 1976, 1977; Connor and Stevens, 1971a,c). Two
different methods were used to determine the equations describing particular
ionic current components. In some cases, experimental data and theoretical
curves were displayed simultaneously on a computer terminal display
screen. The expressions generating the theoretical curves were then
varied to achieve a good correspondence, judged by eye, between experimental
and theoretical curves. In other cases, where the choice of an appropriate
descriptive equation (e.g. an exponential or a power of an exponential)
Lad already been made, measured data points were transformed mathematically
so that exponential parameters could be determined from a linear slope
and intercept.
Most of the ionic current mechanisms described in this paper are

15
characterized by equations for time constants and steady state activation
values as functions of membrane potential. The equations used are of the
general form:
1 K5-f(V) = K1 { + K2K3I exp , V-
K4(1)
where Kl-K5 are the parameters to fit experimental data points. Solutions
to this equation are sigmoidal in form, as is evident in Figs. 10-14. In
some cases, the time constants might be better described by an equation
giving a bell-shaped curve. Nevertheless, the equations used appear
adequate for the range of membrane potentials relevant to the reconstruction
of bursting pacemaker activity. The general expression indicated by eqn. 1
was chosen purely for convenience in fitting experimental data points,
rather than for any possible physical significance.
Reconstruction of membrane potential
The mathematical reconstruction of membrane potential described in
part III of the results was accomplished using numerical techniques,
implemented on a digital computer, to solve finite difference analogues of
the differential equations developed in part II to describe the membrane
current. The first-order differential equations defining the behavior of
the postulated voltage-dependent gating variables m and h were integrated
using the point slope formula. Approximate solutions to the partial
differential equation representing the radial diffusion of intracellular
calcium ions were found by a method equivalent to resolving the spherical
intracellular space into a series of thin, concentric shells. m first-
order transfer of ions between adjacent shells in such a system

16
approximates the diffusion process. The effects of widely varying finite
variable step sizes for the time and radius were investigated in order to
discover and avoid values which might give rise to errors in the solutions
being sought. Time increments were automatically adjusted during each
computing run, since longer increments were acceptable during quiescent
intervals than during action potentials.

RESULTS I: SLOW RELAXATIONS OF VOLTmGE CLAMP CURRENT
Previous studies have characterized the major ionic currents flowing
during voltage clamped depolarizations of Tritonia bursting pacemaker
neurons (Thompson, 1976, 1977). The results presented here are concerned
primarily with some long-lasting aftereffects of depolarization, observed
as slowly relaxing current tails following the return of the voltage
clamped membrane to negative holding potentials. These slow current
relaxations are of interest because they occur on a time scale similar to
the slow burst oscillation, and because they are observed in the pacemaker
potential range between spike threshold (typically -35 mV to -25 mV) and
the trough of the interburst hyperpolarizing wave (-55 mV to -40 mV). m
rationale for quantitatively separating the slow, subthreshold current into
two distinct components is developed here. m third component of ionic
current, believed to represent an inward calcium current, will also be
described. This putative calcium current has relatively rapid kinetics
of activation and decay, and therefore makes no direct contribution to
slow current tails, but does appear to play an important role in mediating
the activation of one of the two slower current components.
Characteristics of slow current tails
Fig. 2m illustrates the procedure we have used to record tail current
relaxations. The soma membrane is voltage clamped and held at a specified
holding potential (-40 mV in this case) for several minutes to allow the
membrane current to stabilize. The membrane then is stepped to a specified
pulse potential (+2 mV) for a specified duration (3.0 sec) and returned
to the original holding potential. The two lower traces in Fig. 2A display
the resulting membrane current signal at two different amplifications. In
the low gain record, only the large net ionic current flowing during the

Fig. 2: m. Procedure for measurement of slow tail currents. Top
trace: membrane potential controlled by voltage clamp circuitry.
Middle trace: clamp current at low gain showing larger currents
flowing during activation pulse. Bottom trace: simultaneous high-
gain record showing the smaller slow tail current. Current during
the pulse is off scale at this amplification. The flat baseline
represents the level of the steady holding current at this hold
potential. It was drawn to emphasize the waveform of the relaxa-
tion back toward this level. Similar tail current baselines are
drawn in subsequent figures. m. Slow tail currents recorded after
pulses of five different durations (indicated to the left of each
trace) by the method indicated in m. This data was recorded from
the same cell as that shown in m, using the same holding and pulse
potentials.

A
18
mV+21 MEMmRANE POTENTImL
-40 J I 3 sec
100 -
nm 0
-100 -
CLAMP CURRENT: LOW mMPLIFICmTION
2 ]
nm 0 --2
CLAMP CURRENT: HIGH mMPLIFICmTION
BPULSE DURmTIONS (msec) INDICATED NEmR TRmCES
100 - ••••
,w.00..:01.0.101PFeriirir.r...11.~0.44/.111.11.11.~..1ftrall.•■■•■■•■400 •••• •
2 nA
10 sec
HOLDING POTENTImL: -40 mVPULSE POTENTImL: +2 mV

19
pulse and the impulse-like capacity current transients are evident. The
main properties of the ionic currents flowing during such pulses have
been described by Thompson (1976, 1977). The higher gain trace shows the
slow tail current following the pulse: the current during the pulse is far
off scale at this amplification. The flat baseline represents the steady
level of holding current and has been drawn to emphasize the waveform of
the relaxation back to this steady level. It is with such slow current
relaxations that the present analysis is primarily concerned.
Fig. 2m shows the typical effect of varying the duration of the
activating pulse. mfter the shorter pulses, the slow current tail is
dominated by an initially inward transient. As pulse duration is increased,
the waveform undergoes a complex transition to an initially outward form.
There is reason to believe that this transition from inward to outward
current is intimately related to the mechanism of bursting pacemaker
activity. It has been possible to demonstrate such transition behavior
in virtually every active bursting pacemaker cell studied, but never in
non-bursting ganglion cells. Only slow tails of the initially outward
form are normally observed when the procedure of Fig. 2m is applied to
ganglion cells other than identified bursting pacemakers.
The ionic sensitivities and current-voltage relationships of the
inward tails after short pulses are very different from those of the out-
ward tails after long pulses, suggesting that the complex tails may be
a composite of two distinct slowly decaying ionic currents. Several
procedures for studying each of the two slow processes in isolation have
been developed. The two slow currents, designated I B and Ic , are discussed
separately below.

20
The slow inward current IB
The experimental analysis of I B is described in detail in S. H.
Thompson's doctoral dissertation (Thompson, 1976). Only a brief summary
of that analysis will be presented here.
IB decay tails can be studied with relatively little interference from
other slow transients by using only short (<50 msec) activation pulses. The
resulting relaxations are of the inward form shown by the top trace in
Fig. 2B. Such tails behave as if they resulted from the slow decay of
an increased membrane permeability to sodium and calcium ions following
the pulse: the amplitudes of inward tails are reduced by substituting
Tris for sodium or magnesium for calcium in the bathing salines, and the
tails are eliminated by making both substitutions simultaneously. The
tail currents after short pulses are unaffected by changes in the external
potassium concentration, unaffected by addition of the potassium current
blocking agents TEm (tetraethylammonium) and 4-AP (4-aminopyridine), and
unaffected by minor variations in hold potential near the reversal potential
for other components identified as potassium currents. These observations
rule out the possibilities that I B tails could reflect either changes in
potassium permeability or an extracellular accumulation of potassium
ions.
It has not been possible to observe I B activation directly at depolar-
ized potentials. This is because the currents associated with the slow
kinetics defining IB are much too small in comparison to the other currents
activated during membrane depolarization (see Fig. 2). The conductance g B
activated during various depolarizing pulses has instead been estimated by
extrapolating the slow current tails at subthreshold potentials back in

21
time to the ends of pulses and dividing by an appropriate estimate of the
driving force. It is assumed that slow tails measured near the potassium
equilibrium potential reflect I B alone, regardless of activating pulse
duration. This assumption is made in order to reject another slow current,
associated with long pulses, which appears to reflect mainly a variation in
potassium conductance (see the following section). From measurements of
IB tails, it has been possible to estimate the time course and saturating
level of gm activation at potentials ranging from -40 mV to +30 mV during
pulses lasting up to 2 sec. The activation of g m appears to increase
progressively with depolarization over the entire range of potentials
studied, reaching steady-state values in the range 0.02 to 0.04 limhos at
0 mV. The kinetics of activation and decay appear to be approximately
exponential, with the time constants ranging from 2 seconds or more near
-40 mV to 300 msec or less at +20 mV (see Fig. 12, Results II).
The slow outward current I
Observations described in this section suggest that the slow outward
current tails after long activation pulses reflect mainly a relaxation of
membrane permeability to potassium ions. The current governed by this
slowly varying potassium permeability is designated I C„ Transitions from
inward to outward slow tails like that illustrated by Fig. 2B appear to
reflect an increasing activation of the outward IC' relative to the
inward IB' as depolarizing pulse length is increased. Like potassium
currents described in neurons of mplysia and Helix (Meech, 1974b; Meech
and Standen, 1975), I c appears to be activated by the influx of calcium ions
during depolarization, rather than by depolarization per se.

22
Fig. 3 shows evidence that a variable potassium permeability contributes
increasingly to slow tail waveforms as activation pulses are lengthened.
The tail currents in the left column were recorded after pulses of four
different durations in a bathing saline of normal ionic composition. The
slow tails in the right column were recorded under conditions identical
except that the bath potassium concentration was increased from 10 mM to
40 mM by replacing a small fraction of the sodium ions with potassium. m
major effect of the solution change is evident only after the longer activa-
tion pulses. The finding that the tails after short pulses are relatively
unaffected by a change in the electrochemical gradient for potassium ions
is consistent with our previous conclusion that such tails reflect mainly
sodium and calcium currents. The outward tails after the longer pulses, on
the other hand, are completely eliminated by high external potassium. This
observation suggests that such outward tails probably reflect a slow varia-
tion of potassium permeability. The four-fold elevation of external potas-
sium should nearly eliminate the electrochemical gradient for membrane
potassium current at the -40 mV holding potential used in this experiment.
mssuming an intracellular potassium activity of 137.9 mM (equal to the
mean values measured in cell R-15 by Kunze, Walker and Brown, 1971) and
assuming an activity coefficient of 0.69 for K + in the external medium (see
Brown, 1976) the Nernst potentials for potassium are -73.2 mV in the 10 mM
K medium, and -39.3 mV in the 40 mM K+
medium, at 11°C. The slow inward
tails evident after long pulses in high potassium probably reflect mainly
the slow inward current, IB' and possibly a very small potassium current
now flowing inward.
The waveform of slow tail current after a long pulse depends on the

Fig. 3: The effect of external potassium concentration on slow tail
currents. The currents in the left-hand column were recorded in
the normal (10 mM K+) bathing medium. Each tail was recorded at a
holding potential of -40 mil after a pulse to +2 mV. The activation
pulse duration was varied as indicated near the traces (in msec).
The currents in the right-hand column were recorded under condi-
tions identical except that the bathing medium contained 40 mM K+
.
Note that the tails after the shortest pulses are nearly identical
in the two different bathing media. ms pulse length is increased,
tails in the normal medium develop initially outward components,
while the forms of the tails in high potassium remain relatively
constant. The digitized records in this and subsequent figures
are sampled at 200 msec intervals. The short vertical tick-marks
at the left of the holding current baselines in this and subsequent
figures showing digitized current tail records represent the times
of the trailing edges of the activation pulses, that is, the times
at which membrane potential was returned to the holding level.

23
25
•
FV
100 F--. 711°61611
•;
•
400 . • • •, .
•
•
• • 1 nA
10 sec••
• • .1' Al w a , ,if#104441.....,..4repimeivnarivoftentAvi.
t 2000 sibrAemie.rworanfru...).......0-1- 71mb,
HIGH (40 mM) IC+NORMmL (10 mM) +

24
holding potential at which it is measured. Fig. 4 depicts the slow
tails following identical 1 sec pulses to +1 mV at several different holding
potentials from -30 to -65 mV. mt -30 mV and -35 mV, the slow tails are
entirely outward. ms the holding potential is made more negative, the
tails undergo a reversal, but current decays are not always monotonic and
the form, as well as amplitude and sign, of the decay varies with potential.
The data shown in Fig. 4 are consistent with the identification of the
slow outward tails with a variable potassium permeability: the outward tail
current becomes smaller as the holding potential is made more negative and
thus closer to the potassium equilibrium potential. m more precise inter-
pretation of the hold potential dependence of slow tails is difficult
because these tails presumably represent a composite of several processes
with similar relaxation times. The slow tails at the more negative holding
potentials may include effects of a slow, time-dependent inward-going
rectification (see Marmor, 1971), as well as I B and IC' and the earlier
phases of the decay may reflect the recovery from an extracellular accumu-
lation of potassium during the activation pulse (see Eaton, 1972).
If the slow outward tails do in fact reflect a decline in membrane
ionic permeability, it should be possible to observe a corresponding
decrease in membrane conductance during such tails. Fig. 5 illustrates an
experiment designed to test this prediction. The upper trace shows the
slow outward current tail observed at a hold potential of -35 mV after a 4
sec pulse to -1 mV. The lower trace was recorded under conditions identical
except that a 5 Hz square wave, 3 mV peak to peak in amplitude,was superim-
posed on the -35 mV hold potential during the slow decay of outward tail
current. Two aspects of the membrane current response to the imposed voltage

Fig. 4: The effect of holding potential on the slow current tail
waveform. mll tails were recorded after 1 sec pulses to +1 mV,
in the normal bathing solution.- Holding potential was varied as
indicated near each trace. Slow tails were not recorded at more
negative or more positive potentials because the holding currents
required became excessively large and generally failed to stabilize
at steady values.

25
HOLDING POTENTIAL (mV) INDICATED NEAR TRmCES
—35 —
--40
—45 --I
—50 —I
—55 --J
—65 ---I
PULSE POTENTIAL: +1 mV PULSE DURATION: 1 sec

Fig. 5: Comparison of tail current and slope conductance time-
courses after a 4 sec pulse to -1 mV. The tail current at a con-
stant holding potential of -35 mV is shown in the top trace. The
current waveform shown in the lower trace was recorded under
conditions identical except that a voltage square wave (3 mV, p-p)
was added to the -35 mV holding potential beginning 1 sec after the
pulse. m "late" membrane slope conductance is proportional to the
height of the inner envelope of the current response to the square
wave (see text). Note that the inner envelope becomes narrower
along a time-course similar to the decay of net outward current
shown in the upper trace.

26
PULSE
HOLD POTENTIAL
4 sec -35 my-1 my 2 sec
-35 my square wave (3 my p-p)
it

27
square wave are evident in Fig. 5: capacity current spikes coincident
with the square wave voltage steps, and an inner envelope of the quasi-
steady currents approached during each half-cycle of the square wave.
The difference between the quasi-steady currents flowing during any two
successive square-wave half cycles can be divided by the amplitude of the
intervening voltage step to obtain a slope conductance giving a relative
indication of slow variations in membrane conductance. (Note, however,
that the absolute magnitude of the slope conductance so calculated has no
significance, since rapid changes in ionic permeability probably occur
between the time of each voltage step and the subsequent approach to a
quasi-steady current). Fig. 5 shows that the width of the inner current
envelope, which is proportional to the slope conductance defined above,
decreases along a time course similar to that of the slow outward current
tail. The most probable explanation of this finding is that both the
decline in outward current and the decline in slope conductance reflect
a slow decline in membrane permeability to an ion with a reversal potential
negative to -35 mV. Measurements of slope conductance therefore also
support the identification of the slow outward tail with a variable per-
meability to potassium ions, which should have an equilibrium potential
near -70 mV.
The time course of slow potassium permeability changes
The evidence presented above indicates that the slow current tails
observed in the pacemaker potential range reflect at least the two
distinct ionic currents IB and I
C . Fig. 6 illustrates a procedure to
separate the time course of the potassium permeability change underlying I c
from the simultaneous relaxation of I B and any other ionic current not

Fig. 6: Procedure for estimating the time course of membrane potas-
sium permeability. Tails are recorded in media of two different
potassium concentrations, and the differences in current are calcu-
lated point by point in time from digitized records. A. tail
current after a 700 msec pulse to +2 mV at a holding potential of
-40 mV. The normal bathing medium used has 10 mM le. B. tail
current in 40 mM K+
medium. Pulse dimensions and holding potential
the same as in A. C. tail current recorded after return to normal
10 mM le medium. Pulse dimensions and holding potential the same
as in A and B. D. Waveform assumed to represent the time-course
of potassium permeability (see text). Calculated by subtracting
points in B from the mean of points in A and C.

28
A NORMmL e B HIGH le
• o •
1 -: •.21P
.1%
•P•
1 nm
10 sec
C RETURN TO NORMmL le D NORMmL - HIGH le• DIFFERENCE
Nog,ANIONSA,40,
Illiodmmu siuglarimial.maumu,gmt. 411444/444441,WeiWoluslor..

29
involving potassium ions. The procedure is based on the observation,
illustrated in Fig. 3, that slow tail waveforms depend on the concentration
of potassium ions in the bathing solution. If this is due to a slowly
varying potassium permeability, the differences between current tails in
different potassium concentrations should directly reflect the time-course
of that permeability change. Fig. 6m-C shows slow current tails following
a 700 msec pulse to +2 mV, at a hold potential of -40 mV. The tails were
recorded in a normal bathing solution (10 mM Fe, Fig. 6A), then,in the
40 mM le solution (Fig. 6B), and then again in the normal solution(Fig. 6C). Fig. 6D shows the difference between the tail current in high
potassium and the mean of the two tails recorded in normal potassium,
calculated point by point in time during the tail relaxations. Tails
recorded in the normal medium both before and after the high potassium
run were averaged in the hopes of controlling for the gradual rundown of
tail current amplitudes generally evident during this rather lengthy
experimental procedure (20 min was required for each solution change to
equilibrate and 20 min to record tails after ten different pulses in
each solution).
The current difference shown in Fig. 6D should be directly propor-
tional to time-dependent variations in membrane potassium permeability
if one assumes: (1) that only potassium fluxes are affected by the
change in [K+Jbath from 10 mM to 40 mM, , and (2) that [K+
and [1(+
] 0, the
potassium concentrations immediately adjacent to the inner and outer
membrane surfaces, respectively, are constant for a given [le]bath'
If
one further assumes: (3) that [K+
]o
is equal to [K+]bath'
(4) that
membrane potassium permeability is independent of the external potassium

30
concentration, and (5) that [K+] is independent of [m
+]bath' it is
possible to calculate the proportionality constant relating the observed
current differences to variations in a potassium permeability coefficient
of the type expressed in the Goldman-Hodgkin-Katz flux equation (Goldman,
1943; Hodgkin & matz, 1949). If AP represents a deviation of the potassium
permeability coefficient from its steady-state level at a holding potential,
V, and Idiff represents the difference between tail current recorded in
+baths of two different potassium concentrations, [1(+]bath and bath
(expressed here in moles/cm3), we can write, from the Goldman-Hodgkin-
Katz equation,
1 eVF/RT-
Idiff F
2 V yK [K+ ]bath - y K
+] bath
(2)
where R, T, and F have their usual meanings, and yK is an activity
coefficient. It is assumed that yK has a value of 0.69 in the seawater-
like media used (see Brown, 1976). For the data shown in Fig. 6, the
constant APC/Idiff has a value of 8.7 x 10
-9cm sec
-1 . The data in Fig.
6D therefore represents a slow decay of AP c from a value of approximately
3 x 10-8 cm-sec-1 at a time 200 msec after the pulse. The applicability of
assumptions (1) -(5) is considered in the discussion section.
Fig. 7 shows current differences after activating pulses of ten
different durations, ranging from 25 msec to 3 sec. These differences
were obtained by the procedure illustrated in Fig. 6, using the same cell,
and the same holding and pulse potentials. The figure shows that the
potassium-sensitive component of slow current is activated progressively
during the depolarizing pulse up to the longest duration studied.
C RT

25
31
1 nm
10" sec
501...V44.11.potkablivArtahricsv
1000
•
•
•
100•4014/044:1,pw.l.e..".
•
•
* 1500
200 • \A, .
-.."‘1"414.440i.m4Piell.wrvo..ppialue
•
• •
•
2000
400
I) •.1"644 ftetbkicalichailimmiag
.
■so
\61.04..4.„..4.1•46...wevair,dgazoirjm
•
•• •
• 3000
•700
1.ftwoks.4.604#646400.4.4#

32
Though noise inherent in the preparation makes exact comparisons difficult,
the decay tails in Fig. 7 appear similar in form over a wide range of pulse
durations and relaxation amplitudes. This situation is in marked contrast
to the complex effect of pulse duration on the total current in normal
bathing media (cf. Fig. 2b), suggesting that the potassium change
separation procedure has successfully isolated an elementary component
of the slow current tails.
The role of calcium entry in the activation of I
m number of studies have found evidence that part of the potassium
current in bursting pacemaker neurons is activated by an influx of calcium
ions during membrane depolarization (Meech, 1974 b, c; Meech and Standen,
1975; Heyer and Lux, 1976; Thompson, 1977). Meech (1974 a, c) found that
recovery from this calcium-dependent potassium activation includes a slow
component, which appears comparable in its time-course to the decay of
IC in Tritonia neurons. mn investigation of the relationship of calcium
entry to the activation of I C has therefore been undertaken.
The requirement for calcium entry. One argument for the proposed mediating
role of calcium ions (Meech and Standen, 1975) cites the suppression of
potassium activation by operations presumed to reduce or eliminate voltage-
dependent calcium currents. Such operations include the removal of calcium
ions from the bathing solution and the addition of certain other divalent
metal cations, such as Cow and Mn++ (see Hagiwara, 1973). Fig. 8 shows
the effect of removing most of the bath calcium, and adding cobalt (Co++
),
on slow current tails in a voltage clamped Tritonia bursting pacemaker. The
upper trace shows the tail current, in a normal bathing solution, at a
holding potential of -30 mV, following a 1 sec depolarization to +1 mV.

Fig. 8: The effect of Co and lowered Ca++ on the slow tail
current after a long pulse. The two current tails were both
recorded at a holding potential of -30 mV, after a 1 sec pulse to
+1 mV. The upper trace was recorded in the normal bathing saline.
The lower trace was recorded in a solution where part of the Mgt
and all but 1 MM of the Ca++ were replaced by Co++.

33
NORMAL (10 mM Ca)
COBALT, LOW CmLCIUM (30 mM Co, imM Ca)

34
The tail is of the initially outward form presumed to reflect mainly the
slow decay of IC. The lower trace shows the very different slow tail
current recorded in the cobalt bathing solution. In this solution, the
tail is initially inward, resembling the slow tails normally observed
at this holding potential only following pulses of much shorter duration
(see Fig. 2). The suppressing effect of cobalt and low calcium on I C
shown in Fig. 8 is representative of observations over a wide range of
pulse dimensions and holding potentials at which outward tails are
normally observed. Other experiments (see Thompson, 1977) show that the
slow outward tails can also be suppressed by the addition of either
Co++
or Mn++ to solutions containing the normal (10 mM) concentration of
calcium, or by simply removing calcium in the absence of Co++
or Mn++
.
mdditional voltage clamp experiments suggesting a role of calcium
entry in the activation of potassium currents in Tritonia neurons have
been described by Thompson (1977). Thompson's study includes a description
of a faster calcium-sensitive component of the outward current flowing
during depolarizing pulses. On the basis of a common requirement for
calcium entry, it seems likely that the calcium-dependent potassium
current observed during pulses and that observed as slow tails after
pulses reflect a single mechanism of potassium transport, with complex
gating kinetics which may depend on both calcium influx and voltage.
The dependence of calcium entry on time and voltage. In considering possible
roles of calcium entry in potassium activation, it should be helpful to
characterize the dependence of calcium entry on time and membrane potential.
One question of particular interest concerns the pattern of calcium entry
during sustained depolarizations. Many published studies have suggested

35
that calcium currents are subject to a voltage dependent inactivation,
possibly similar to that governing sodium currents (Geduldig and Gruener,
1970; Connor and Stevens, 1971; Kostyuk, Krishtal and Doroshenko, 1974; Stan-
den, 1975). The rate and extent of such inactivation bears on the inter-
pretation of the continued, progressive activation of I C observed during
long depolarizing pulses (see Fig. 7). If there is a sustained influx of
calcium during long depolarizations, the progressive activation of I C
may simply reflect the cumulative entry of calcium ions. If, on the other
hand, calcium currents inactivate rapidly and completely, an alternative
explanation for the progressive activation of I C is required. Previous
voltage clamp studies of the calcium current in molluscan somata have
suggested divergent views of the kinetics of calcium inactivation, based
on different types of observations in many different types of cells (see
calcium current references above, but also Kostyuk,Krishtal and Pidoplichko,
1975; mdams and Gage, 1976; Eckert and Lux, 1976; Connor, 1977). I have
attempted, therefore, to analyze the calcium current in Tritonia neurons
for conditions of time and voltage comparable to those employed in
characterizing IC activation.
One possible source of ambiguity in a voltage clamp study of calcium
inactivation arises from the presence of a calcium-dependent potassium
activation mechanism. Calcium activation followed by a potassium activa-
tion dependent on calcium entry may sometimes be difficult to distinguish
from calcium activation followed by calcium inactivation. Both sequences
would appear as an inward current which declines or becomes outward
during sustained depolarizations, and in either case both the initial
inward current and the later outward going component would be reduced
by reductions in external calcium. Connor (1977) has shown evidence that

36
an apparent calcium inactivation in mrchidoris neurons can be attributed
mainly to a calcium dependent potassium activation. He has also shown
that barium ions appear to carry inward currents in a manner very similar
to calcium, but either block or fail to activate the calcium-dependent
potassium current. This finding is important because no other agents
which effectively block this potassium current without blocking calcium
channels have yet been described. (The calcium activated potassium current
in Tritonia neurons appears quite resistant to the effects of external
TEm, even in high doses, Thompson, 1977.)
Fig 9 illustrates an experiment designed to assess the time-course
and voltage dependence of membrane calcium permeability during long
depolarizations of a voltage clamped Tritonia bursting pacemaker cell. The
cell was bathed in a medium in which all sodium was replaced by Tris and
TEm to eliminate inward sodium currents and block the TEA-senstive
fraction of outward potassium current. Fig. 9B shows a representative
record of the membrane current when the cell is depolarized in a medium
where calcium has been replaced by barium. Fig. 9m is the current during
an identical pulse in a solution where magnesium replaces calcium. Since
barium, but not magnesium (Hagiwara, 1973), substitutes for calcium in
carrying inward current, and since calcium-activated potassium current
should be absent in both media, the difference between the currents, as
represented graphically in Fig. 9C, should give an indication of the cal-
cium current that would normally occur during such a pulse. Fig. 9C
indicates that a calcium current of about 25 nm would activate very
rapidly upon depolarization to +22 mV. The current declines only to about
one half by the end of the 5 sec depolarizing pulse, but then turns off
very rapidly on return to the holding potential. The turning on and off

Fig. 9: Inward currents in a Na+-free, Cam-free solution contain-
ing Ba++
. m. Upper trace: voltage clamped membrane potential.
Lower trace: membrane current in Na+-free, Ca
++ -free solution
before adding Ba++
. B. Same as m, except 10 mM Bad (replacing
Mgt)) was added to the bathing solution. Note development of a net
inward current during pulse. C. The current records from m and
B are superimposed. The difference between these currents is
assumed to represent an inward Bad current. D. The dependence
of inward Bad current on membrane potential during pulse. Filled
circles represent peak inward current early in pulse. Open circles
represent inward current near the end of the 3 sec pulse.

•D
•
0 Current- Voltage Relationship
Barium-dependent +22 mV -
-- 3 sec —
-38 mV -
0 nA
-25 nA -
Pulse Potential, mV'10 20 30-20 -10
0-30
A
0 0
o Late0
0
Na, Ca free
50 mM Mg
100 mM TEm
10 mM Ba Traces
added Super-
Imposed ••
Voltage Clamp Currents • • • Early
• -10 nm
-20
•

38
of the inward current evident in Fig. 9B is too rapid to be accurately
resolved in intact Tritonia neurons (because of the long capacity transients
in this preparation, see methods), but an upper limit of 20 msec for 90%
activation and decay can be safely assumed. The decline of inward current
by about one-half evident in Fig. 9C is typical of our observations in
this voltage range.
If inward barium currents accurately represent normal inward calcium
currents, as Connor's data suggests (Connor, 1977), a major influx of
calcium must persist for the duration of the longest pulses used in
investigating IC activation. It is therefore possible that the progessive
activation of I with long activation pulses may simply represent an effect
of cumulative calcium influx. The decline of inward current during the
5 sec pulse shown in Fig. 9B may indicate a partial inactivation of
calcium current, but the inactivation is far from complete. The decline of
inward current is usually even less prominent at lower pulse potentials;
currents often decline by as little as 10-20% during a 5 sec pulse to
0 mV, as evident in Fig. 9D. Fig. 9D shows current-voltage relationships
derived by subtracting currents in magnesium from currents in barium, as
indicated by Fig. 9C.
The calcium current component characterized by findings like those
illustrated in Fig. 9 will be designated I D . We have noted that the slow
inward current IB also appears to be carried partly by calcium ions but
we shall tentatively regard I B and ID as distinct transport processes.
The two currents have completely distinct dependerres on time and mem-
brane potential: ID turns on and off far more rapidly than I B and IB

39
begins to activate at more negative potentials than ID . The maximum
apparent inward calcium fluxes associated with I D are 10 to 20 times
larger than largest calcium fluxes associated with the slower I B kinetics.
It is therefore to be expected that I B will be of little importance to
the activation of calcium-dependent potassium currents whenever depolariz-
ations are large enough to activate ID.

DISCUSSION OF SLOW CURRENT TAIL ANALYSIS
The slow inward current IB
A slowly activating component of inward current in bursting pacemaker
neurons was first reported, nearly simultaneously, by Lux and Eckert (1974)
and by Gola (1974). Though less detailed, the observations reported by Gola .
are very similar to the observations of IB described by Thompson (1976).
Gola did not identify the ionic species carrying the slow inward current,
but he did describe slow tails in the pacemaker potential range'which
closely resemble the I B tails evident after short pulses in Tritonia
neurons. Eckert and Lux (1976) have now described an inward current
(Iin slow) with some properties similar to those of IB' The characteriza-
tions of Iin slow and of IB differ in that Iin slow is regarded as being
highly selective for calcium, while IB appears to be carried by sodium
as well as calcium. The properties of Iin slow' as described by Eckert and
Lux (1976), are actually intermediate between the properties of IB , the
slow inward current, and ID, the fast calcium current, as described
in this thesis and by Thompson (1976). IB and ID are distinguished here
mainly on the basis of their differing activation and decay rates. Were
IB and ID to be regarded as a single mechanism, with very complex
activation kinetics, that mechanism would coincide closely to Eckert and
Lux's Iin slow. More work on the selectivity and activation properties
of the inward current will be necessary to decide which type of characteri-
zation is the more appropriate.
IB is almost surely the ionic current which accounts for the long
depolarizing afterpotentials (DAPs) observed after single action potentials
or bursts in bursting pacemaker neurons (Thompson and Smith, 1976). Like

41
IB' DAPs are observed only in bursting pacemaker neurons. The time-course
of IB and DAPs are similar, and I B and DmPs show a similar dependence on
both sodium and calcium in the external medium.
Since the slow inward current tails persist (with reduced amplitude)
in the absence of either sodium or calcium, but not in the absence of both,
we tentatively describe I B as a single conductance component, imperfectly
selective between sodium and calcium ions. Nevertheless, we have not ruled
out the possibility that the sodium and calcium components might reflect
completely separate transport mechanisms, perhaps even depending somewhat
differently on time and membrane potential.
The'calciuMactivated - potassium current, IC
The activation of long-lasting increases in potassium conductance by
depOlarizing pulses in molluscan neural somata was first noted by Connor
-and Stevens (1971c). Subsequent studies have linked such conductance
increases to post-tetanic hyperpolarization, spike frequency adaptation,
and bursting pacemaker activity (Brodwick and Junge, 1972; Partridge and
Stevens, 1976; Junge and Stephens, 1973; Gola, 1974; T. Smith, Barker and
Gainer, 1975).
Meech and Strumwasser (1970) were the first to show that long-lasting
increases in potassium conductance could be produced by the intracellular
injection of calcium ions. Meech (1974) later showed that calcium entry
during depolarization was probably responsible for the potassium conduct-
ance increases underlying post-tetanic hyperpolarization. In this thesis
and in a preceding paper (Thompson, 1977), it has been suggested that I C
in Tritonia neurons reflects a mechanism similar to that described by

42
Meech. I have also described a voltage-dependent component of calcium
entry, ID, with properties appropriate to account for the activation of
I during membrane depolarization. In the following section (Results II),
I discuss the relationships among ID , the intracellular accumulation of
calcium ions, and the activation of I c in more detail.
The procedure used to measure the time course of the potassium
permeability change underlying I c involves five assumptions, which are
liated in the text above. Only assumptions (1) and (2) are required for the
inference that current differences in different external potassium concen-
trations are directly proportional to potassium permeability. mssumptions
(3), (4), and (5) are necessary only to determine the exact magnitude of the
related permeability change.
mssumption (1) requires that only potassium fluxes be affected by
changes in external potassium concentrations. m deviation from the
required condition would occur, for instance, if external potassium ions
block an ionic channel selective for some other species, or if an exchange
mechanism couples potassium influx to the movements of some other ion. m
deviation from assumption (1) would be misleading in the present context
only if the blocked or coupled ion flux is electrogealc and varies with
a time and voltage dependency similar to that of the potassium permeability
change. The only mechanism known at present that can be considered as a
possible source of such an error is the ouabain-sensitive sodium-potassium
exchange pump. The best available evidence indicates that this transport
mechanism is, indeed,electrogenic in molluscan neurons (Gorman and Marmor,
1970; Thomas, 1972a), and that it has a slow time and voltage dependence
due to its dependence on the internal sodium concentration [Na]..

43
Experiments by Thomas (1972b) have suggested, however, that the pump
rate is affected by [m] o only at concentrations below the physiological
level (i.e., less than 1 mM in snail neurons where the normal [K] o is
4 mM), . Thomas has also shown that sodium loads are cleared with a time
constant on the order of 10 minutes, and that the electrogenic pump
current has a similar time course. These relaxations are much slower
than the decays of I c. For both of these reasons it seems unlikely that
a [K]o - dependent variation in sodium-potassium exchange distorts
the estimates of slow potassium permeability changes presented in this
thesis.
mssumption-(2) requires that potassium concentrations near the
membrane do not vary during slow tail decays, and (3) requires further
that external potassium near the membrane be equal to bath potassium.
The lower baseline level of extracellular potassium implies that external
concentration effects are the most likely to be significant. Eaton
(1972) has presented evidence that a considerable accumulation of
potassium occurs during large depolarizing pulses. Such an effect may
be significant during the first few hundred milliseconds after large
depolarizations, but probably does not distort the estimates of the
slower conductance tails considered above. Eaton's studies and similar
preliminary investigations of my own indicate that the external potassium
levels equilibrate in times on the order of a few hundred milliseconds.
This conclusion is also substantiated by direct measurements of external
potassium near the membrane (vieher and Lux, 1973), in which relaxations
after depolarization were observed to be essentially complete within 1 sec.
Furthermore, I have purposely chosen to record slow tails after activation
pulses of relatively low amplitude to minimize the efflux of potassium

44
during the depolarization. (Peak outward currents during the pulses em-
ployed for the experiment illustrated by Figs. 6 and 7 were only about one-
third of the outward currents during the pulses employed by Eaton, 1972,-
to demonstrate accumulation).
mssumption (4) requires that membrane potassium permeability remain
constant as external potassium is varied. On this point, one can merely
note that a major dependence of potassium permeability on external
potassium concentration has been described only in connection with potas-
sium currents: showing strong inward rectification (Hodgkin and Horowicz,
1959; Noble amiTsien, 1968,1969). The calcium-activated potassium
current, on the other hand, shows an outward rectification characteristic
(Meech and Standen, 1975; Smith, unpublished).
mssumption (5) requires that [if i remain at the same value during
the tail measurements in normal and high potassium media. Though some
increase of [le] i in the high potassium medium is to be expected,
experiments using ion-selective microelectrodes to measure intracellular
potassium activities indicate that the effect would be small for the
conditions of the present study. Russell and Brown (1972) found that
changes of [e] i in mplysia neurons produced by reducing [e] bath tozero occurred with a time constant on the order of hours, and sometimes
began only after a delay comparable to the duration of each run in the
present study. It is to be expected that the smaller fractional changes
in [e]bath imposed in my experiments would produce even more gradual
effects. Miura, Hoffman and Rosen (1977) found that no significant changes
in [K]i occurred in canine cardiac purkinje fibers exposed to changes
in [e]bath comparable to those employed here.

RESULTS II: m MATHEMmTICmL MODEL OF
THE MEMBRmNE CURRENT
Multiple component description of membrane current
This section describes a mathematical model intended to summarize
the electrical behavior of the Tritonia bursting pacemaker cell soma
membrane, as observed under voltage clamp conditions. Voltage clamp
experiments described by Thompson (1976, 1977), along with the results
described in the previous section (Results I), indicate that the'membrane
current in the-physiological voltage range can be regarded as the sum of
a capacity current and seven distinguishable components of ionic current.
Some properties of these ionic current components are summarized in Table
I. The designations II , IK , - Im and IL were introduced by Connor & Stevens's
(1971 a,b) analysis of the ionic current in neurons of the dorid nudibranchs
mrchidoris montereyensis and mnisidoris nobilis. Since these species are
closely related to Tritonia, and since similar ionic currents can be
identified in each species, we adopt the same terminology for the four
analgous components. New designations are introduced for three ionic
current components, IB , IC, and ID, which were not described in the studies
by Connor & Stevens.
Each ionic current component appears to reflect a time-dependent
membrane permeability to sodium, calcium or potassium ions (except that
the ions carrying I L have not been determined) as indicated in Table I.
In order to model the electrical behavior of the ionic current components,
each will be represented as a variable conductance in series with an ionic
battery, as in the work of Connor and Stevens (1971c). mt a membrane

Table I: Summary of the properties of seven components of ionic
current observed in bursting pacemaker neurons. The information
presented in this table was derived as indicated in the more
detailed descriptions of each individual current component in the
text following this section. The column headed "Gating Kinetics"
specifies appropriate descriptive formalisms, most often in terms
of first order, voltage-dependent gating variables like the m and
h in Hodgkin and Huxley's (1952) description of the squid axonal
sodium current. The specific coefficients governing the behavior
of these m- and h-like variables are described by eqns. 6-10 below.
The column headed "T(-40 my)" indicates exponential time constants
approximating changes in activation gating observed at a membrane
potential of -40 mV. The column headed g specifies conductance
scale factors, determined for a cell with an estimated membrane
capacitance of 19.8 nF (see following text).

46
Table I: Seven components of ionic current observed in bursting pacemakers
Current Component MajorPermeantIons
Vrev
(mV)
GatingKinetics
t(-40 mV)
(msec)
g
(umhos)
II Fast sodium Na+
+45 m3h 4 29.7note
IB Fast calcium Cam +130 m 7 0.259
IB Slow inward++
Na+ ,Cay +85 1800 0.026
note
IK Delayed outward -52
notem2
24 2.55
IA Transient outward K+
-63 m4h 18 11.7note
Slow outward K+
-73 Cai depnote
1 to 50seconds
6.7x10-5
note
IL Leakage +7 none none 0.039
notes:
I • subject to complete or nearly complete inactivation at potentialspositive to 0 mV.
IB: gm = gB (0 mV,40). See p. 60.
figure is V axis intercept of instantaneous I K(V), extrapolatedIK: Erevfrom the interval -40 mV < V < -20 mV. See p. 63.
IA' subject to complete or nearly complete inactivation at potentials•
IC : membrane potential dependence assumed to be mediated by changes inintracellular concentration of ionized calcium. The figure for grexpresses the sensitivity to intracellular calcium ions in umhos/riM.
positive to -30 mV.

47
potential V, any component current, I, will be given by
I = g (7-vrev)
(3)
where g is a variable with the dimensions of a conductance and Vrev
is a
reversal, or zero-current, potential. Variations in g represent the
gating characteristics of the particular ionic current component. The
gating process may depend on membrane potential, but it is assumed that
at least on the macroscopic level gating variations occur in a fashion
that is graded and continuous in time. This assumption and eqn. 3 imply
that each compOnent will have a linear instantaneous current-voltage
relationship. While this condition has not yet been verified in a
strict way for each component observed in Tritonia neurons, neither has
there been any evidence of pronounced rectification within the limited
potential range (-50 mV to -30 mV) over which the slow pacemaker
oscillation occurs. In the case of each current component active in this
pacemaker potential range, the quantitative models presented below have
been based mainly on tail, currents measured in this same potential
range. The linear conductance defined by eqn. 3 should therefore be
adequate to approximate the electrical behavior of:each ionic current
component during the slow pacemaker oscillation of pribery interest here.
The ionic transport mechanisms associated with each current component
will sometimes be referred to as the ionic conductances g I , gm , gm, g
gc , g , or gL .
Formalisms of two different types will be used to describe the gating
of the various ionic current components. For all the conductances
except g,,, gating is presumed to depend directly on membrane potential and

48
will be described by equations of the type introduced by Hodgkin & Huxley's
(1952) analysis of the axonal ionic currents. m general form for this
type of equation is
g = imx h (4),
where g is a constant with the dimensions of a conductance and m and h
are dimensionless variables governed by first order differential equations
whose coefficients depend on membrane potential. The m term is used to
represent the activation of conductance by depolarization, and may be
raised to the power x (greater than 1) where necessary to describe a
delay in the increase of conductance. The h term is used to represent
the inactivation of a conductance by depolarization, if such a process
must be described. Both m and h vary between 0 and 1, so the conductance
scale factor g corresponds to the maximum possible, or fully-activated,
conductance associated with a given current component (in one case these
terms have a different significance, see section on IB below). Since the
gating of the conductance g appears to depend on calcium entry, rather
than depending directly and exclusively on membrane potential, the
formalism of eqn. 4 is inappropriate for this component. m new formalism,
explicitly representing the involvement of calcium ions is developed in
a later section.
Measured conductance scale factors and reversal potentials for each
ionic current are specified in Table I, along with some information on
the gating kinetics governing each conductance. The ideal situation of
characterizing all current components completely in a single exemplar
cell has not been achieved. First, the experimental lifetimes of

49
individual neurons are too short for the large number of measurement
operations that would be required. Second, for the fast sodium current
II' Connor and Stevens's(1971) data has been used in preference to the less
reliable measurements available for Tritonia neurons. (The voltage
clamp technique they used is superior to that employed in the present
studies for the resolution of fast current changes, see Methods). In
order to minimize the errors introduced by pooling data on individual
currents from different cells, 1) g I was scaled in direct proportion to
membrane capacitance from Connor and Stevens's measurements_ and 2) gm and gK
were estimated from representative pulses in the same cell used for the
final determinations of kinetics and gc . Estimates of gL , gd, and gm ,
and all reversal potentials remain subject to errors due to individual
variations among bursting pacemaker neurons.
Capacity current
If the somatic membrane is assumed to behave as an ideal capacitor,
and to be spatially isopotential, the somatic capacity current, Icap' will
be given by
I = C dVcap s dt (5)
where Cs is the soma capacitance.
Thompson (1976) used a voltage clamp procedure to estimate Cs in
Tritonia bursting pacemakers, and found values ranging from 9 nF to 46 nF.
When referred to the apparent spherical surface area of the somata, these
values imply a specific membrane capacitance, C m, upwards of 5 pf/cm2
.
This figure greatly exceeds the accepted value for nerve membranes of

50
1 pf/cm2 . mnatomical studies (Mirolli andTalbott, 1972; Graubard, 1975) of
molluscan somata provide an obvious basis for this discrepancy: the sur-
face membrane appears to be highly infolded, forming numerous narrow
clefts ranging up to 15 micrometers in depth.
Electrical measurements of Cs in intact cells are subject to errors
due to poor clamping of the axonal cable. An estimate of Cs based on
anatomical findings is employed here to scale the capacity current.
Anatomical studies of molluscan nerve cells similar in size to the
Tritonia bursters have found that surface infolding results in a 7.5-
fold (4imlliandTalbott,1972) or a 6-fold (Graubard, 1975) increase in
membrane area over that expected from the overall dimensions of the
soma. The somata of the Tritonia bursting cells from which data in the
model have been selected appeared under the dissecting microscope to be
spheres approximately 300 micrometers in diameter. The surface area of
such a sphere is 0.00283 cm2. Assuming an infolding factor of 7, and a
true membrane capacitance of 1.0 pF/cm 2, a value of 19.8 nF has been
adopted for C s .
The fast sodium current, •.„
The bursting pacemaker neurons of Tritonia possess a sodium current
mechanism resembling that first described in the squid giant axon by
Hodgkin and Huxley (1952). Connor and Stevens (1971a,c) found that equations
retaining Hodgkin and Huxley's m3 h formalism could adequately describe
the inward currents in Dorid neurons, though in certain other details
these currents differed from the axonal sodium currents. The sodium
current in Tritonia neurons appears to be similar to the inward current

51
measured by Connor and Stevens (Thompson, 1976). Connor and Stevens's
(1971c) descriptive equations, in their exact form, have therefore been
adopted to represent the sodium current in Tritonia bursting pacemakers.
The sodium conductance g will be described by
gI = gI mI3(V't) hI (V , t)
(6a)
where gI is the fully-activated sodium conductance, and m I (V,t) is an
activation variable and hI(V,t) an inactivation variable analagous to
Hodgkin and Huxley's m and h, respectively. The value of gI in Table I
was obtained by scaling the value measured by Connor and Stevens (1971c) to
maintain the same conductance per unit membrane capacitance. The value
of EI in Table I is equal to that measured by Connor and Stevens (1971a).
The variables mi (V,t) and hI (V,t) are governed by the first order
differential equations:
dmI (V , t)T (V) MIdt
MI(V,t) = mi (V,03)
and
dhI'(V t)
ThI(V) dt+ hI (V ' = hI (V co)
where TmI (V), 1111,(V, 00), ThI (V), and h (V,01) are estimated from the
experimental data in figure 10. The data points were obtained from
voltage clamp measurements by Connor and Stevens (1971c); the smooth curves
in Fig. 10 were fit by eye and are given by
11.5TmI (V) = V + 16 + 0.5 cosec (6d)
1 + exp( 5.75 )
(6b)
(6c)

Fig. 10: The dependence of g i gating parameters on membrane poten-
tial. The symbols represent values estimated from voltage clamp
data by Connor and Stevens (1971a,c). The solid curves were fit by
eye (but see text, p. 84). A. mctivation time constant, TmI
(V).
Eqn. 6d specifies the solid curve. B. Steady-state activation,
m1 3 (V,00). Eqn. 6e specifies the solid curve. C. Inactivation
time constant,ThI(V) . Eqn. 6f specifies the solid curve.
D. Steady-state inactivation, h 1 (V,00). Eqn. 6g specifies the
solid curve.

ml3(V ' m))
53
1 30
V - 11 + exp( )
(6e)
V' = 7 ln(214 - 1) - 21.5 = -30.1 mV
= 2.7
85thI(V) = V + 24 + 4.5 msec (6f)
1 = exp( 3.75 )
1 6hI(V,°°
) 1 + exp(V + 19.5 )
}
4.5
(6g)
The parameter 0 / is incorporated to adjust the steepness of the exponential
foot of m3i (V,00), with only minor effects on the rest of the curve. The
estimation.of this parameter is subject to rather large errors: the data
points at the foot of the curve are unreliable, as they necessarily
represent very small and potentially contaminated inward currents.
One apparent discrepancy in adopting Connor and Stevens's description
of II must be discussed. Noting that some inward current persisted in the
absence of external sodium ions, provided calcium was present, Connor and
Stevens described II as a mixed sodium and calcium current. It has since
been reported that the activation of the sodium and calcium inward currents
can be temporally separated (Kostyuk, Krishtal andftdoplichko,1975; mdams
& Gage, 1976; Connor, 1977). These recent findings indicate that the
sodium current has rapidly activating kinetics like those described by
Connor and Stevens for I I' while the calcium current either does not
inactivate or does so only very slowly. Connor and Stevens's method
of measuring I I neglects any inward current which does not inactivate
rapidly. so their data should correctly represent the sodium component

54
of inward current.
The fast calcium current, ID
Observations discussed earlier in this thesis (see Results I) indi-
cate that Tritonia bursting pacemaker neurons-have a time- and voltage-
dependent inward calcium current. The largest and fastest component of
this calcium current is designated ID: its properties are assumed to
correspond to the prominent inward currents observed in barium-substitUted,
sodium-free bathing salines, as discussed previously. Inward calcium
currents similar to ID have been reported in neurons of several other
gastropod species (Kostyuk, et al., 1975; Eckert and Lux, 1976; mdams
and Gage, 1976; Connor, 1977).
The data points in Fig. 11 represent estimates of the peak calcium
conductance activated by various step depolarizations of a voltage
clamped Tritonia neuron. ms noted previously, the activation and decay
of ID
occur too rapidly to be resolved accurately in the intact cell prep-
aration used in obtaining the data shown in Fig. 11. Connor (1977, per-
sonal communication) has used an isolated cell voltage clamp technique,
however, and achieved a more precise temporal resolution of the calcium
currents in mrchidoris neurons. His preliminary observations indicate that
both the activations and decays are approximately exponential, with time
constants of 10 msec or less. Since the activation and decay of I D
appears to be exponential at all potentials, the gating kinetics can be
approximated by a single potential-dependent first-order variable:
= gD MD, t)
(7a)

Fig. 11: The dependence of steady-state activationon membrane
potential, mD (V,c0). The symbols represent the relative activation
of calcium conductance at various membrane potentials, estimated
from the data shown in Fig. 9. Conductance values were calculated
from the peak inward currents specified in that figure, assuming
ED = +130 mV. The solid curve (specified by eqn. 7C) was fit by
eye.

55
1.00
0.75
0. 50
0.25
0.00-30.-25. -20. -15. -10. -S. 0. S. 10. 15. 20. 25.
MILLIVOLTS

56
where gD is the fully-activated calcium conductance. The value for iD
listed in Table I was determined from the data shown in Fig. 9, assuming
a reversal potential, ED, of +130 my. This value for ED is approximately
the Nernst potential for calcium ions, if it is assumed that the intra-
cellular calcium concentration near the membrane approaches 5 x 10 -7
during pulses such as those imposed in Fig. 9 (see section on intra-
cellular calcium concentrations below). The activation variable is
governed by the differential equation
dri1D(V,t)TmD (V)
MD(V't) = MD (V 'm)dt(7b)
The function mD(V,07) has been determined from the experimental data
points shown in Fig. 11. The curve in that figure corresponds to
mD (V,0*) -1 11.7V + 3.5
)1 + exp(-7.25
(7c)
m fixed value of 7 msec will be adopted for the time constant T D . This
value is in the range suggested by Connor's data, and is just below the
upper limit of 10 to 20 msec. estimated from voltage clamp studies of
Tritonia neurons (see Results I, the dependence of calcium entry on time
and voltage).
The possibility that mayy be subject to some degree of potential-
dependent inactivation has already beet discussed (Results I, calcium
entry). There has also been a suggestion (Heyer and Lux, 1976) that
somatic calcium currents may be subject to a frequency-dependent
facilitation when activated repetitively. Neither inactivation nor

57
facilitation is represented in our description of I D since these phenomena
are probably of secondary quantitative importance in the present context,
and because there remains considerable ambiguity in the experimental
observations of these processes.
The slow inward current IB
m component of inward current characterized by very slow decays
after depolarizing pulses was originally observed in bursting pacemaker
neurons by Gola (1974) and by Eckert and Lux (1975). m voltage clamp
analysis of the analogous inward current in Tritonia neurons, desig-
nated IB' has been described by Thompson (1976). Of all the ionic
current components distinguished in voltage clamp experiments on
Tritonia neurons, IB is the one component which is observed exclusively
in identified bursting pacemaker neurons. Ion substitution experiments
have indicated that IB is carried by both sodium and calcium ions. The
fraction carried by calcium appears to be approximately one-third, and that
by sodium approximately two-thirds (Thompson, 1976).
IB is the smallest of the ionic currents described here. In a given
cell, the largest observed values of IB are less than one hundredth the
maximal values of the sodium current II and less than one tenth the
calcium current ID' IB is, nonetheless, readily distinguished from the
other inward currents by its slow kinetics of decay at negative hold
potentials: II and ID relax to steady values within a few milliseconds
following a depolarizing pulse, whereas IB persists for seconds as an
inward tail. Because of the simultaneous activation of other much
larger ionic currents, I B cannot be observed directly in the positive
potential range. The kinetics of activation at positive potentials

58
must be inferred from observations of the slow tails at negative holding
potentials following depolarizing pulses of various dimensions. The
lack of more direct information on the gating of I B at depolarized
potentials is probably not important for the present purposes: I B
constitutes a significant fraction of the total ionic current only at
the subthreshold potentials where the larger ionic conductances are
gated off.
The gating kinetics of gB , as inferred from tail current measurements,
appear to be approximately exponential in time at all potentials, and I B
does not appear to inactivate markedly with prolonged depolarization
(Thompson, 1976). The gating of gm can, therefore, be adequately described
by an equation with a single voltage-dependent, first order activation
variable, which need not be raised to a power:
gm =
B (V, t)
(8a)
where gm is the estimated steady-state conductance at 0 mV. The value
for "B listed in Table I was determined using a method described by Thompson
(1976), assuming a value of 85 mV for E B . The activation variable m B(V,t)
is governed by
dmB (V,t)T (V) mB + mB (V,t) = mB (V,00)
dt(8b)
The functions TB (V) and mB (V,00) have been estimated from the experimental
data in Fig. 12. The smooth curve and straight line in that figure were
fit by eye and are given by

Fig. 12: The dependence of g m gating parameters on membrane poten-
tial. The symbols represent values estimated from voltage clamp
data as described by Thompson (1976). The solid lines were fit by
eye. m. mctivation time constant, TmB (V). Eqn. 8c specifies the
solid curve. B. Steady-state activation, m B (V, 00). Eqn. 8d
specifies the straight line. (See text, p. 60.)

0. 10. 20.0.00 I1 I
-60.. -SO. -40. -30. -20. -10.MILLIVOLTS
B
e0. a CI
4000.
2000.
6
00. L I I
-60. -SO. -40. -30. -20. -10.MILLIVOLTS
0 .
A

60
3000TmB (V) = V + 40 + 300 msec
1 + exp( 8.5(8c)
mB (7,03) = 1 + V/60 (8d)
A straight line has been used to represent the function mB (V,00) because
the available data did not seem adequate to determine a curve of the
more familiar sigmoidal form. It is difficult to imagine a physical
interpretation for the linear relationship indicated. While it•is not
likely that this function could be strictly correct, it should be
adequate for the present descriptive purposes, over the physiological
voltage range from -50 mV to +30 mV.
The delayed potassium current, IK
The potassium current in Tritonia neurons has been resolved into three
kinetically and pharmacologically distinct components (Thompson, 1977).
mlthough none of these components have properties identical to the squid
axon potassium current, the component we designate I m is the most similar
to the axonal current. IK
activates along a sigmoidal time course follow-
ing step depolarizations, and it decays exponentially after depolarization.
Like the axonal current (EhrensteinandGilbert, 1966), I K undergoes some
inactivation during prolonged depolarization, but the process is slow and
incomplete. IK differs from the axonal potassium current in its response
to certain pharmacological agents, and the activation times of I m are
slower than those of the axonal current by approximately a factor of
ten (Thompson, 1977).
If we neglect the slow inactivation process, the activation of g K
under voltage clamp conditions can be approximated by an equation with a

Fig. 13: The dependence of g K gating parameters on membrane
potential. The symbols represent values estimated from voltage
clamp data, as described by Thompson (1977). The solid curves
were fit by eye (but see text, p. 84). m. mctivation time
constant, Tmm (V). Eqn. 9c specifies the solid curve. B. Steady-
state activation, mm2 (V,0*). Eqn. 9d specifies the solid curve.

60. 1.00
0.75
40.
6
❑
0.50
U❑ 0.25
U. I I I I—60. —40.. —20. 0. 20.
MILLIVOLTS
0.00-40. 20.0 .—20. —10.
MILLIVOLTS10.
❑

63
This descriptidn of gk is quantitatively quite similar to Connor and
Stevens's (1971a,c) description of a potassium current in Archidoris neurons.
It differs in that Connor and Stevens included a secondary activation term
describing a slower component of potassium activation. In Tritonia neurons,
these faster and slower components of potassium conductance can be
pharmacologically separated, so they are described here as two separate
components, andd gc (Thompson, 1977; see also Meech and Standen, 1975).
The value of -52 mV for EK specified in Table I is not the reversal
potential for Ik. II( actually reverses at a potential closer to -60 mV.
The value of -52 mV is used because it provides a much more accurate
description of Ik at the potentials where this component would be expected
to be active under physiological conditions (-40 to +30 mV). The
discrdpancy between this figure and the true reversal potential is due
to a marked outward rectification in the instantaneous current-voltage
relationship for this component (Smith and Thompson, unpublished). The
measured reversal potential of -60 mV is still considerably more positive
than the estimated potassium equilibrium potential of -73.2 mV (see
Results I, the slow outward current, ThisThis difference is presumed to
reflect- an imperfect selectivity of the transport mechanism for potassium
ions, an effect of extracellular potassium accumulation, or both.
The transient potassium current, IA
m current component in Tritonia bursting pacemakers has properties
almost identical to those of the transient potassium current I m described
in Dorid neurons by Connor and Stevens (1969, 1971b,c;see Thompson, 1977).
m is evident in voltage clamp experiments only when holding potentials
or prepulses negative to -40 mV are employed: This component is almost

64
completely inactivated in the range of normal resting potentials. On
depolarization from a sufficiently negative potential, the conductance
gm
activates rapidly, but then declines to zero during maintained depolari-
zations (Thompson, 1977). The sequence of activation and inactivation
observed is analogous to the behavior of the axonal sodium current, except,
of course, that Im is normally outward, rather than inward, in the
physiological potential range.
Connor and Stevens (1971b,c) found that a formalism similar to the m3h
description of sodium current (Hodgkin and Huxley; 1952) could adequately
describe the potential-dependent activation and inactivation of g m. The
rising phase of gm, however, appears to be best fit by the fourth power,
rather than the third power, of a first order activation variable. Like
the conductance studied by Connor and Stevens, gm in Tritonia neurons can
be described by the equation
..._gm = g
m mm4 (V'
hm(V '(10a
wherem is the fully-activated conductance. The value for gm listed in
Table I was determined assuming a reversal potential, E m , of -63 mV, accord-
ing to the method described by Thompson (1977). The activation variable
mm (V,t) and the inactivation variable hm (V,t) are governed by
dmA(V,t)Tmm (V) mm(V,t) = - mm (V,c0) (10b)
dt
dhm(V
't)
Thm (V) hA(V , t) = h
m'CV 00)
dt(10c)

(10e)
and
1 + exp(V 4.'473.9
)
1 10.5
66
The steady-state activation and inactivation functions, m m (V,00) and hm (V,00),
have been determined from the experimental data represented in Fig. 14.
The smooth curves in that figure are given by:
mm4(V ' 1 V + 59 41 + exp( -10.5)
(10d)
The activation and inactivation time constants governing g m appear to have
only a minor dependence on membrane potential (Thompson, 1977; see also
Connor and Stevens, 1971b). For modelling purposes, rmm will be assigned
a fixed value of 18 msec, and Thm will be assigned a fixed value of 260
msec. These values are near the middle of the range of values measured
by Thompson (1977).
The calcium-activated potassium current, I C
m calcium-dependent component of potassium conductance in molluscan
neurons was first described by Meech and coworkers (Meech and Strumwasser,
1970; Meech, 1974a,b,c; Meech and Standen:, 1975). They showed that part of
the potassium activation normally occurring during depolarization is
actually mediated by an influx of calcium through voltage-sensitive
calcium channels. The evidence for such a mechanism includes the
susceptibility of a kinetically distinct component of potassium current to
supression by a variety of different operations known to prevent calcium
entry. In addition, Meech (1974a) has shown that microinjection of calcium
into cells can mimic certain effects of depolarization on the membrane

67
potassium conductance.
m component of potassium current observed in Tritonia neurons has
properties similar in many respects to the currents described by Meech
and his associates. This component, designated I C, is described in detail
in previous sections of this thesis (see Results I), and in Thompson
(1977). The chief characteristic distinguishing I from other components
of potassium conductance is its dependence on calcium entry. I C accounts for
the slowest variations of potassium current in voltage clamped Tritonia
neurons: it activates progressively for several seconds during step
depolarizations and the decays on return to a negative holding potential
require tens of seconds.
Waveforms representing the slow, decay of the conductance g after
various depolarizing voltage clamp pulses are shown in Fig. 15. The data
points in this figure are chord conductances derived from estimates of slow
potassium permeability changes presented above (see Results I, the time-
course of slow potassium permeability changes). Increments in I over
steady-statevalues (Ay, were calculated from increments in permeability
(APC' see eqn. 2) using an appropriate form of the Golman-Hodgkin-Katz
current equation:
F v y •
+ o_y[e ] eVF /RTK
SIC = APC RT 1 - eVF/RT
As in eqn. 2, yK is an activity coefficient with the value 0.69 and the
concentrations are in moles/cm3. m value of 1.379 x 10
-4 moles/cm
3 for
the intracellular potassium acitivty (y[e] i) was determined from the
direct measurements by Kunze,WalkerandBrown(1971). m value of 10_5 moles/
cm3 (10mM) was used for [K1 so the calculated values for AI_ should

Fig. 15: Slow decays of the potassium conductance g c at -40 mV
after pulses to +2 mV for the durations indicated. The times
designated 0 sec correspond to the trailing edges of the specified
activation pulses. The data points are chord conductances derived
from the same data as the points shown in Fig. 7 (see text, p. 67,
for method). The solid curves were calculated from a theory
represented by equs. 7, 8, 12, 13, 16-18. These curves are pro-
portional (see eqn. 18, p. 77) to predictions of the free calcium
concentration near the inner membrane surface after pulses of the
same dimensions used in obtaining the corresponding experimental
data. Both points and curves represent transient conductance
changes only, i.e., the pulse dependent increases over the steady-
state conductance at the holding potential of -40 mV.

60. 3000 msec
4010 20 30
SECONDS
SO.
0.310 30
SECONDS
700 msec
40.
40
e.
100 msec200 msec
40 SO
SIMONDS SECCVDS0 10 30 40 SO 10 20
30
e.
68

69
approximate the increments in I c following the specified pulses in a normal
potassium bathing medium at the holding potential V = -40 mV. The con-
ductance values plotted in Fig. 15 were obtained by dividing the values
for AI by the net driving force on potassium ions at -40 mV (assuming
ER = -73.2 mV, see Results I, the slow outward current, I c). The chord
conductances so calculated are used below to estimate the conductance
scaling factor "gc necessary to quantitatively model I c .
m calcium-coupled model for the activation of I c . Meech and coworkers
(see Meech and Thomas, 1977) have described evidence suggesting that the
calcium-activated potassium conductance may be controlled directly by
the concentration of free calcium at the inner surface of the cell mem-
brane. This hypothesis provides the basis for the description of I
activation kinetics developed in this section. m model incorporating
experimental measurements of membrane calcium transport and cytoplasmic
calcium diffusion and binding is shown to predict kinetics of intracellular
accumulation with a close resemblance to the observed kinetics of I c .
Factors capable of influencing intracellular calcium levels may
conveniently be divided into three categories: (1) the influx across
the .cell surface membrane, (2) the efflux across the surface membrane,
and (3) the intracellular diffusion and binding of calcium. It will be
assumed that the voltage clamp data on inward calcium currents accurately
characterize the calcium influx. Experimental data on efflux, diffusion
and binding in gastropod somata, on the other hand, are scant, but a great
deal of information on these processes has recently become available from
studies on cephalopod giant axons. The model developed here therefore
makes use of data from the squid axon, where necessary, by scaling

70
intensive properties to a geometry representing the approximately spherical
gastropod neural soma.
(1) Calcium influx. The influx of calcium as a function of time and
membrane potential is assumed to occur as implied by eqns. 7 and 8, which
describe the calcium inward currents I D and IB , respectively. The molar
influx of calcium, mnf1ux is then given by:
(ID + 0.33 133)
minf lux z F(12)
where F is Faraday's constant and z = +2 for the divalent calcium ion. The
factor 0.33 represents the observation that only about one third of I B
appears to be carried by calcium ions (see Thompson, 1976).
(2) Calcium efflux. Since the electrochemical gradient for calcium ions
across nerve membranes is large and inwardly directed, the efflux respons-
ible for maintenance of the steady state must be an active process. The
active efflux of calcium has been best characterized by experiments on
internally dialysed squid axons (see Brinley, 1976). Such experiments
indicate that efflux is in approximately direct proportion to the con-
centration of calcium at the inner membrane surface, over most of the range
of possible physiological calcium concentrations (when other factors, such
as internal mTP and external Na are held constant at their normal levels,
see DiPolo, 1973, 1977; Brinley, Spangler andMullins, 1975). It is assumed
that active efflux across the soma membrane is similar to,the process
studied in axons. mt the low internal concentrations expected in neuronal
cytoplasm, the molar efflux of calcium, me fflux' is then given approximately
by

=mefflux
K e 1
whereKe isaconstantx.Aditheunitsofm/secar 1
t) is a variable
representing the free calcium concentration near the membrane (see below).
Some numerical values for Ke determined from published experimental
data on squid axons are indicated in Table II below.
(3) Diffusion and binding in cytoplasm. It will be assumed that calcium
ions entering across the surface membrane may diffuse into the cytoplasm.
It is known, however, that calcium ions in neuronal cytoplasm are subject
to binding and sequestration reactions and that those reactions occur on
a scale sufficient to effect a high degree of buffering to imposed calcium
loads and to restrict diffusion drastically compared to that which would
otherwise be expected in aqueous solution (Luxoro andYaiiez, 1968; Blaustein
amdHodgkia,1969, Baker and Crawford, 1972; see also Rose and Loewenstein,
1975). If it is assumed that free calcium ions diffuse as in dilute
solution, but that bound calcium is immobilized, the differential equation
governing diffusion is:
8[Ca++]2 , ++, 9[CaX] - D V icaat 9t (3)
where [Cam] denotes the concentration of free calcium in some small
volume element, and [CaX] the concentration of the bound form (or forms) of
calcium in that same volume element. D is ,a diffusion coefficient with
the value 6.4x10-6
cm2/sec (see BlausteinandHodgkin, 1969). To use eqn.
14, it is necessary to characterize the kinetics of CaX formation, and to
define specific geometrical and boundary conditions.
71
(13)

72
It is assumed that calcium binding in somatic cytopolasm can be
approximated by a single class of reactive sites, uniformly distributed
throughout the volume of the cell, and that these sites bind calcium in
the manner described by Baker and Schlaepfer's (1975) studies on isolated
squid axoplasm. Those authors described a component of calcium binding,
probably to axoplasmic proteins, which could be approximated by first
order Michaelis-Menten kinetics with a dissociation constant between 300
and 500 nM (the value 400 nM is adopted here) and a total concentration
of sites between 20 and 40 1114/4 axoplasm (30 414 is adopted here). Since
calcium binding'to other proteins with similar estimated affinities (e.g.,
troponin C) is known to be extremely rapid compared to the slow kinetics
of IC' it will be assumed that the binding reaction equilibrates
instanteneously. For the formation of CaX, we can now write:
[X][CaX] - (15)
1 + m /[Ca++
]
where [X] = 30 4M, the total concentration of binding sites, and m =
400 nM, the dissociation constant. It is probable that the uptake of
calcium by mitochondria (Meechandlhomas, 1977; Tiffert, BrinleyaniScarpa,
1977) or other organelles (Baur et al., 1977) may account for the majority ,
of calcium binding under conditions of large calcium loads or high local
concentrations (e.g., microinjection), but such effects will be neglected
here. (There are indications that mitochondrial binding may be less sig-
nificant at low, physiological calcium concentrations than a simple extra-
polation of high dose behavior to low calcium levels would imply, see
Carafoli and Crompton, 1976. m similar conclusion emerges from consideration
of the effects of mitochondrialinhibitorsin axoplasm reported by Baker and

73
Schlaepfer, 1975).
The somata of Tritonia bursting pacemaker neurons are approximately
spherical in overall shape and about 300 p in diameter. There are folds
several microns deep in the surface membrane which may restrict diffusion
under some conditions, but for purposes of calculating slow concentration
relaxations after impulse-like calcium influxes, it will be assumed that
soma geometry can be approximated as a smooth and symmetrical sphere. This
also neglects any effects of the relatively small axonal process'from the
soma. By assuming spherical symmetry, eqns. 14 and 15 can be combined and
rewritten in the form:
3Cai(r,t) D [Cel (r,t)+mx ] 2 a 2 aCa(r,t)fr
at X r2 9r- 3r (16)
where Cai (r,t) represents the concentration of free calcium ions at a
radius r within the cell. The value of r varies from 0 at the center of
the cell to a (150 pm) at the surface membrane.
Solving for the dependence of Cai (r,t) on time and voltage The main features
of the intracellular calcium model developed above are summarized in Table
II. The intracellular calcium distribution will depend in general on
time and membrane potential, due to the time- and voltage-dependence of the
calcium inward currents I D and IB . Depolarizing pulses will result in
pulse-like influxes of calcium ions. After activating pulses, accumulations
of calcium near the membrane will dissipate due to the diffusion of calcium
into the internal volume of cytoplasm, due to the intracellular binding of
calcium ions, and due to active extrusion at the cell surface membrane.

Table II: Summary of factors considered in predictions of intra-
cellular free calcium concentration transients. References and
numerical values of parameters taken from studies on squid axonal
cytoplasm or membrane are included, as the presently available data
on calcium metabolism in gastropod somata are very limited.

74
Table II: Outline of model for intracellular calcium ion metabolism
1. Calcium Entry
Observations: Voltage clamp of calcium inward currents in Tritonia neurons.
Representation: Eqns. 7 and 8; Table I. Time- and voltage-dependent gatingof two conductances, gm and gd .
2. Extrusion of Calcium from Cell
Observations: Tracer calcium efflux from internally dialysed squid axons(work cited immediately below).
Representation: Eqn. 13. Efflux approximated by a direct proportionalityto the instantaneous concentration of calcium ions near the innermembrane surface. Pump sensitivity constants derived from the datapoints at low calcium concentrations presented in several publishedstudies are listed below.
Ke (cm/sec) Reference
1 x 10-4
3 x 10-4
1 x 10 3
3.2 x 103
Blaustein and Russell (1975)
Brinley, Spangler and Mullins (1975)
Di?olo (1973)
Blaustein (1976)
3a. Calcium Binding in Cytoplasm
Observations: Equilibrium dialysis of extruded squid axoplasm with tracercalcium (Baker and Schlaepfer, 1975).
Representation: Eqn. 15. Michaelis-Menten binding to a single class ofsaturable sites (dissociation constant 0.4 I1M) distributed throughoutthe intracellular volume (total concentration 30 4M). Must be evalu-ated simultaneously with 3b below.
3b. Diffusion of Free Calcium in Cytoplasm
Observations: Binding as described in 3a above. Fick's law diffusion ofunbound calcium ions is assumed (see Hodgkin and Keynes, 1957, andBlaustein and Hodgkin, 1969).
Representation: Eqn. 16. Partial differe9ti4 equation for simultaneousdiffusion (diffusion constant 6 x 10 -° cm (sec) and bingli-tigNsitl-■ a_sphere (radius 0.015 cm) approximating the neuronal soma geometry.

Fig. 16: Prediction of intracellular free calcium concentrations
when membrane potential of a spherical cell is pulsed for one sec
(indicated by heavy bar below traces) to +4 mV from a holding
potential of -50 mV. Ke was assigned a value of 4 x 10
4 cm/sec,
which is within the range of values indicated in Table II. mll
other parameters are as indicated in connection with equs. 7, 8,
12, 13, 16 and 17, and were obtained either from voltage clamp
measurements or from the studies on squid axon indicated in Table

2000.Lti
1000.
0. e.
82a--1--.65a
8. 10.2. 4. 6.
5000.
FREE CALCIUM CONCENTRmTIONS
mT SPHERICAL RmDII INDICmTED
RmDIUS OF CELL. = a = 150 um
4000.
3000.
TIME (SECONDS)
76

77
cannot be followed to completion on the time scale used in Fig. 16.
Comparisons of Cai (a,t) with gc . The variable designated Ca i (a,t)
corresponds in the model to the free calcium concentration near the inner
membrane surface and therefore represents the physiological parameter
hypothesized to control or gate g c in some way. The solid curves in
Fig. 15 were drawn from the equation
g = Ca (a 0C C i '
(18)
where Cai (a,t) was calculated in each panel for conditions of time and
membrane potential identical to the corresponding experimental measurements
of g , and g is a constant with the value 0.067 nmho/nM calcium. The
solid curves in Fig. 15 thus represent the simple case where the conductance
g is assumed to be in direct proportion to the calculated calcium con-
centration at the inner membrane surface. This might be approximately the
case, for instance, if calcium ions bind in a one-to-one fashion with
potassium channels, and only a small fraction of the number of possible
calcium binding sites are occupied at the highest concentrations of calcium
considered.
A very close similarity between the measured time-course of g and the
calculated time-course of Ca i(a,t) is evident in Fig. 15. This agreement
is striking since all but one of the parameters used in calculating Ca i(a,t)
where fixed, in advance, from independent experimental data, as described
above. Ke was treated as a free parameter because the available experiment-
al literature indicates a wide range of values under slightly varying condi-
tions. The final value for Ke (4 x 10-4 cm/sec) is within the range of

78
experimentally determined values (see Table II).
It should be emphasized that eqn. 18 and the Ca i (a,t) model predict
not only the time course of individual g c relaxations, but also the progres-
sive activation of larger tails by longer pulses, over the entire 30-fold
range of pulse durations shown in Fig. 15. The constanti c was arbitrarily
chosen to bring about a vertical scaling between Ca i(a,t) and the measure-
ments of gc, but once chosen, the same value of "gc was used in each panel
of Fig. 15.
Fig. 17 shows that the model also predicts a good agreement between
Cai (a,t) and gC when the activating pulse amplitude, rather than duration,
is varied. The data shown in Fig. 17 was recorded from a different cell
than that shown in Fig. 15, so a different value of ; was used for vertical
scaling, but all parameters in the Ca i (a,t) calculation had values the same
as those used in calculating the curves shown in Fig. 15.
The activation and decay kinetics of aree obviously complex. The
fact that these kinetics are naturally predicted by the model developed
here definitely seems to support the hypothesis that g c is controlled by
intracellular calcium ions. Regardless of the ultimate fate of this
mechanistic hypothesis, however, the Ca i (a,t) model and eqn. 18 are
demonstrably adequate to reproduce the observed time- and voltage-dependence
of gc tails. Though the adequacy of the Ca i(a,t) model in predicting other
aspects of activation has not been tested here, the suitability of the
model for tails in the pacemaker potential range implies that it should be
useful in reconstructing the role of g c in the bursting pacemaker oscillation.

3.0 100
2.5
75
3.5
7/
25
0-30. -20. -10. 0. 10. 20.
0.5
1.0
0.0 -40.
50
79
Pulse Potential (mV)

80
The linear leakage current, IL
The voltage clamp analysis upon which the ionic current descriptions
above are based has relied heavily on the measurement of the current
relaxations following step changes in membrane potential. m linear leak-
age current, by definition, undergoes no such relaxation and is therefore
not subject to analysis by the methods employed for the other current
components. The direct experimental measurement of such a linear current
is in fact very difficult in bursting pacemakers, due to the fact that
larger non-linear currents appear to be active in every voltage range.
(A non-linear, inward-going rectification, not described above, becomes
prominent when the membrane is hyperpolarized below the physiological
potential range, see Marmor, 1971). These difficulties notwithstanding,
the effects of a linear leakage component must be considered. It is
known, for instance, that molluscan neural somata have a permeability
to chloride (Barker and Levitan, 1975) which is not represented by any
of the conductance components described above.
Parameters appropriate to represent the linear leakage current in
Tritonia bursting pacemakers have been deduced from voltage clamp data
by a process of elimination. It has been assumed that any membrane con-
ductance evident in the immediate subthreshold potential range -45 mV to
-35 mV, not already accounted for by the variable conductances described
above, is linear leakage described by
IL = gL (V -VL ) (19)
where gL and VL are constants. To estimate the values of gL and VL , the
equations representing the other six ionic currents were solved for

81
the steady state at subthreshold potentials and subtracted from observation
of the steady holding currents measured at these same potentials in an
actual cell. m value of 0.039 pmho was estimated for gL and +7 mV for Vt .
The value of 0.039 imho for gL is close to the value of 0.049 pmho
estimated by Connor and Stevens (1971c). The positive value for VL suggests
that a steady inward sodium or calcium current may contribute to IL.

RESULTS III: THE RECONSTRUCTION OF BURSTING
PACEMmKER ACTIVITY
Conditions for the reconstruction of membrane potential
For the purposes of reconstructing membrane potential it is assumed
that the soma membrane is isopotential and that the effects of cable
currents from the axon region are negligible. The total membrane
current, IM, can now be expressed as the sum of all the membrane current
components described above:
dVIm = Cs + II + IK + Im + IB + Ic + IO + IL (20)
The normal unclamped condition of such a cell corresponds to I m=0. The
reconstruction of membrane potential can thus be accomplished by finding
solutions for voltage as a function of time which satisfy this condition
in eqn. 20 and the subsidiary eqns. 3, 5-1Q, 12, 13, and 16-19.
Fig. 18m shows such a solution which was found using numerical
approximation techniques, implemented on a digital computer. mn intra-
cellular recording of the electrical activity from a typical Tritonia
bursting pacemaker neuron is included for comparison in Fig. 18B. The
computation represented in Fig. 18m was initialized by setting all
variables to values corresponding to the steady state at a clamped holding
potential of -38 mV. The oscillatory pattern evident in the figures was
attained within 150 sec after the simulated release from voltage clamp.
The oscillation appears to persist indefinitely in the form represented
here.

1 1 1 1 1 1 1 10
ta
Membrane Potential (mV)
. . . . . . . . . . .
Membrane Potential (mV)
I I I I I I I 1 I I
H
cm8
roC)ca
ID
r.0
CV

84
The many parameters necessary for the calculation of membrane potential
were as indicated in Tables I dild II, and in connection with the individual
membrane current equations. Values for all parameters were estimated
initially, in advance of any reconstruction calculations, strictly from the
analysis of voltage clamp data. The earliest reconstruction attempts
showed that burst-like solutions were a very characteristic feature of the
model developed here: bursting persists over a wide range of particular
parameter values. It was also discovered, however, that the form of
the predicted burst was strongly dependent on the exact values of
certain parameters which could not be estimated with commensurate accuracy
from voltage clamp data. In order to show that the model could reasonably
reproduce a burst topography close to that observed most typically in
actual cells, minor adjustments to four parameters were made after the
fact of preliminary reconstructions. The principal parameters adjusted
were (I)I and q)K' which describe the steepness at the exponential feet of
the steady-state activation curves governing g I and gk. Compensatory
adjustments to gL and EL , the leakage conductance and reversal potential,
were then required (see section on IL above). mll four parameters remained
within the range of experimental error after adjusting them to achieve the
solution shown in Fig. 18m.
Features of the reconstructed potential waveform
There are striking similarities between the reconstructed potential
waveform and the accompanying recording of actual bursting pacemaker
activity. The firing during the active phase of the burst cycle begins
in both cases with a slight acceleration, the second interspike interval
being somewhat shorter than the first, followed by a gradual deceleration

85
lasting the rest of the burst. The forms of the interburst voltage
trajectories are also quite similar. There is a small depolarized after-
potential lasting a few seconds after the final action potential (see
Thompson and Smith, 1977), then a gradual hyperpolarization, followed by
a long slow depolarization leading up to the next burst. These features
of the interburst interval vary quantitatively from cell to cell, but
their presence in absolutely characteristic of bursting pacemaker activity
in Tritonia neurons. These features are always reproduced by the model
under any conditions which allow slow oscillations.
One important discrepancy between the calculated and naturally
occurring burst waveforns is that the model does not reproduce the
observed tendency for action potential undershoots to reach less negative
values after each successive spike in a burst. This tendency, evident
in the intracellular recording shown in Fig. 18B, is a quite typical
feature observed in the bursting pacemaker neurons of many gastropod
species. mnother very likely related discrepancy, not evident in Fig. 18,
is the failure of the model to reproduce the broadening of successive
action potentials commonly observed in actual bursting cells (Stinnakre
and Tauc, 1973; Eckert and Lux, 1977). These shortcomings may be
explained by the fact that the formal description of I K used in the
model disregards a slow inactivation process evident in voltage clamp
experiments (see Connor and Stevens, 1971a).
Other evident differences between the calculated and recorded bursts
in Fig. 18 are of less concern, since the differing dimensions are highly
variable from one actual cell to another, rather than constant features
of bursting pacemaker activity. Thus, though the waveforms in Fig. 18

86
differ in the durations of the firing bursts, the frequencies of action
potential firing during bursts, the forms of the action potential under-
shoots, and the depths of the interburst hyperpolarizing waves, the
dimensions of the computed oscillation are still well within the range
observed in actual cells (see Fig. 1). On the whole, the similarity
between the two waveforms shown in Fig. 18m and 18B suggests that the
features of the membrane current essential to bursting pacemaker activity
must have been described by the model developed above.
The time-course of ionic currents and intracellular calcium during the
reconstructed burst cycle.
The time-courses of some of the major variables computed in recon-
structing bursting pacemaker activity are shown in Fig. 19. The top
panel shows the same membrane potential waveform as in Fig. 18m, but on
an expanded vertical scale. The middle panel shows the variations of
individual ionic current components, displayed to emphasize variations
on the time scale of the whole burst cycle. The bottom panel in Fig.
19 represents intracellular calcium concentrations calculated during
the burst cycle. One trace is labelled to indicate that it represents
the calcium concentration adjacent to the inner membrane surface, i.e.,
Cai (a,t). It is this concentration to which the conductance g is
assumed to be in direct proportion, as expressed by eqn. 18. The other
trace in the bottom panel represents the mean calcium concentration over
theentireintracellularvolume.ThelargedifferencesbetweenCa.(a,t)
and the mean concentration are indicative of large radial gradients of
calcium concentration within the cell. Because of diffusional delays,
which are greatly augmented by the binding of calcium to fixed sites, the

FIg. 19: Time courses of selected variables calculated in recon-
structing the bur .gt cycle. Time axes in each panel are identical
and simultaneous. Top panel shows same membrane potential waveform
as Fig. 18m. Expanded vertical scale truncates spikes but empha-
sizes subthreshold potential variations. Middle panel shows
variations of individual ionic current components, identified by
capital letters near arrows. For clarity, all traces except I B and
I have been suppressed during the firing burst (but see Fig. 20).
ID is not shown in this figure since it is within the trace width
of zero at all times, except during action potentials. Bottom
panel shows variations of intracellular calcium concentrations
near the inner membrane surface and as a volume average throughout
the cell, as indicated.

87
IONIC CURRENT (NA)3. r---.
C
/B
2.
1.
0 .
-2. -
jK
TIME (sec)
- 34.
-36.
- 38.
-40.
-42.
-44.
MEMBRANE POTENTIAL (MV)
1

Fig. 20: Time courses of ionic currents during reconstructed action
potential firing. Time axes in each panel are identical and simultaneous.
Values shown are from the same computing run as those in Fig. 19, but
with the time scale expanded as indicated on the abscissae. The top
panel shows the reconstructed membrane potential waveform; the lower
two panels show membrane currents plotted with two different vertical
scale factors to accommodate the differing magnitudes of the seven
components. The behavior of ionic currents is essentially the same
during each action potential of the burst, except for the changes in
IB and I evident in Fig. 19.

6.
4.
2
0.
-2.
7.1 6.17.87.0 7.2 7.9 8.07.3 7.4I 7.77.5 7.6
TIME (sEc)
89
MEMBRANE POTENTIAL (MV)40.
30.
0.
10.0.
-ie.
-20.
-30.
-40.
-50. 1

DISCUSSION OF MODEL RESULTS
The mechanism of bursting pacemaker-like activity in the model
The model developed in this thesis has been shown to reproduce the
main features of bursting pacemaker activity. An attempt will be made
here to succinctly summarize the mechanism of the slow pacemaker
oscillation, as it is manifested by the model. The explanation offered
below is based on the assumption that the slow oscillation can be under-
stood in terms of two sets of factors: (1) a "background" of membrane
excitability, including the ability to fire action potentials, an
action potential firing threshold, and the stationary sub-threshold
current-voltage relationship, and (2) conductance changes with slow
time-dependence, which act as rate-limiting steps governing slow variations
in membrane potential. Though such a dichotomy cannot be made in an
entirely rigorous fashion, it is nevertheless proposed here for its
possible heuristic value.
(1) "Background" excitability. ms noted above, the model appears
to fire action potentials by a mechanism much like that described by
Connor and Stevens's (1971c) model of repetitive firing in non-bursting
neurons. It is only in long-term, subthreshold behaviot that the
distinctive behavior of the present model becomes evident. Several
authors have proposed that bursting pacemaker neurons are distinguished
by the form of the subthreshold stationary, or steady-state, current-
voltage relationship (see Introduction, previous studies of the mechanism
of the burst oscillation). While there is reason to doubt that this
current-voltage relationship alone is sufficient to differentiate bursting
and non-bursting neurons, the form of this current-voltage relationship

91
is. indeed important to the behavior of the model developed here. Spontan-
eous bursting is observed only when the parameters g/ and VL are adjusted
to values such that the calculated steady-state values of the total ionic
current are between zero and a fraction of a nanoamp negative over the
entire sub-threshold, pacemaker voltage range. In other words, the steady-
state current-voltage relationship must be nearly flat and just below the
zero-current axis. The effects of I L parameters have been mentioned here
because they provide a simple means of manipulating the steady-state current-
voltage relationship in modelling experiments. mctually, all the ionic
current components but I D contribute to the total subthreshold current.
The currents IK, Ic , and IL add positive conductance in the steady-state,
while the currents II' IB' and Im
contribute negative conductance in the
steady-state (negative because of the voltage-dependence of activation
for the inward currents and because of inactivation for I A). m condition
for bursting in the model appears to be that the positive and negative
conductance contributions are nearly equal in the true steady state.
(2) Rate-limiting conductance changes. The relaxation times of the slow
inward current, IB'
and of the calcium-activated potassium current, IC' are
longer by approximately an order of magnitude (or more), than the relaxation
times of all other current components, when observed under voltage clamped
conditions. It will therefore be assumed that the relaxations of these
two currents are rate-limiting for the slow burst oscillation. The follow- ,
ing simplified account of burst formation kinetics is proposed on the
basis of this assumption.
The time-courses of IB and I during the complete calculated burst
cycle are shown in Fig. 19. Just before the first action potential in a

92
burst, both IB and I are relatively small. The positive membrane
potentials associated with the first spike produce an increment in the
inward IB . The calcium influx associated with that spike leads to an
accumulation of calcium inside the membrane and a consequent increment
in the outward I c . The increment in IB is larger: that current
activates more rapidly than Ic. The net effect of the first spike is
thus a lasting inward current. The membrane quickly fires another
action potential, and even more quickly a third as successive increments
of IB are added. This tendency is soon checked as g m reaches a saturation
value, and reversed as I c activates progressively with successive action
potentials and the accumulation of more and more calcium inside the cell.
Though IB
activates more rapidly, I can attain considerably larger values.
mction potentials occur at a decreasing rate until I c becomes large enough
to prevent the membrane potential from reaching spike threshold. In the
absence of spiking and the associated large influx of calcium, both g m
and g now begin to decay, but gm
decays the more rapidly. The small
depolarizing afterpotential following the final action potential reflects
the residual activation of IB. ms IB decays, the membrane reaches the
trough of the interburst hyperpolarizing wave. The decay of g c proceeds
as calcium which had accumulated near the inner membrane surface diffuses
away and is extruded. mfter sufficient decay of gc, the membrane begins
to depolarize. Depolarization results in a gradual reactivation of I B ,
and the small calcium influx associated with IB
leads to a gradual
reactivation of Ic. The slow interburst depolarization appears to reflect
a delicate balance between a regenerative effect of I B activation and a
degenerative effect of I activation. Eventually, the membrane potential
approaches spike threshold, depolarization becomes rapid, and the burst

93
cycle repeats.
This interpretation of bursting as an interplay of the two currents
IB and I is supported by the comparative experimental observations
discussed previously (see Results I and Thompsom and Smith, 1976). Over
a wide range of cell types, species and experimental conditions, it
appears that bursting is observed only in cells where processes like I B
and IC can both be observed. I
B has not been observed in cells that are
not bursting pacemakers, though I often is. Since IB
appears to be a
unique and distinguishing feature of bursting pacemaker neurons, it is
reasonable to suppose that it may play a pivotal role in burst production.
Though IB may be the unique attribute of bursting pacemaker neurons,
it is clear that the calcium-activated potassium current I c is also
necessary for the reconstruction of bursting pacemaker activity from
voltage clamp data like ours. Meech and Standen (1975) were the first to
suggest that the coupling of calcium and potassium activation may be
fundamental to bursting pacemaker oscillation. Eckert and Lux (1976)
studied the properties of calcium current activation in bursting pace-
makers and suggested the possibility that a slow accumulation and dissi-
pation of intracellular calcium is the rate-limiting process in the
slow oscillation. The calcium-dependent model for gc activation developed
in this thesis shows that these suggestions, in conjunction with estimates
based on known properties of neuronal calcium metabolism, can lead to a
simple and self-consistent explanation of bursting, provided that I B is
also taken into consideration. More direct studies of calcium metabolism
in bursting pacemaker neurons will be necessary to directly test the
calcium hypothesis for slow potassium gating. One recent report is

94
encouraging. In a study of the mplysia bursting pacemaker cell R15,
Thomas and Gorman (1977) used the calcium indicator dye mrsenazo III
to record a spatial average of the intracellular calcium concentration.
They observed an oscillation closely resembling that predicted by the
model developed here.

BIBLIOGRmPHY
mdams, D.J. and P.W. Gage (1976). Gating currents associated with sodiumand calcium currents in an mplysia neuron. Science 192:783.
mlving, B.O. (1968). Spontaneous activity in isolated somata of mplysia neurons. J. gen. Physiol. 51:29.
mrvanitaki, m. and N. Chalavinitis (1958). Configurations modales del'activite propres a different neurons d'un mgme centre. J.Physiol. (Paris) 50:122.
mrvanitaki, m. and N. ChalazOnitis (1961). Slow waves and associatedspiking in nerve cells of mplysia. Bull. Inst. oceanogr., Monaco58:1.
Baker, P.F. and m.C. Crawford (1972). Mobility and transport of magnesiumions in squid giant axons. J. Physiol. 227:855.
Baker, P.F. and W. Schlaepfer (1975). Calcium uptake by axoplasm extrudedfrom giant axons of Loligo. J. Physiol. 249:37.
Barker, J.L. and H. Gainer (1975). Studies on bursting pacemaker potentialactivity in molluscan neurons. I. Membrane properties and ioniccontributions. Brain Res. 84:461.
Barker, J.L., M.S. If shin and H. Gainer (1975). Studies on bursting pace-maker potential activity in molluscan neurons. III. Effects ofhormones. Brain Res, 84:501.
Barker, J.L. and H. Levitan (1975). Mitochondrial uncoupling agents:Effects on membrane permeability of molluscan neurons. J. MembraneBiol. 25:361.
Baur, P.S., m.M. Brown, T.D. Rogers, and M.E. Brower (1977). Lipochondriaand the light response of mplysia giant neurons* J. Neurobiol.8:19.
Blaustein, M.P. (1976). The ins and outs of calcium transport in squidaxons: Internal and external ion activation of calcium efflux.Fedn Proc. 35:2574.
Blaustein, P.M. and m.L. Hodgkin (1969). The effect of cyanide on theefflux of calcium from squid axons. J. Physiol. 200:497.
Blaustein, M.P. and J.M. Russell (1975). Sodium-calcium exchange andcalcium-calcium exchange in internally dialysed squid giant axons.J. Membrane Biol. 22:285.

96
Both, R., W. Finger and R.m. Chaplain (1976). Model predictions of theionic mechanisms underlying the beating and bursting pacemakercharacteristics of molluscan neurons. Biological Cybernetics23:1.
Brinley, F.J. Jr. (1976). Calcium and magnesium metabolism in cephalopodaxons. Fedn Proc. 35(14):2572.
Brinley, F.J. Jr., S.G. Spangler, and L.J. Mullins (1975). Calcium andEDTm fluxes in dialyzed squid axons. J. gen. Physiol. 66:223.
Brodwick, M.S. and D. Junge (1972). Post-stimulus hyperpolarization andslow potassium conductance increase in mplysia giant neurone. J.Physiol. 223:549.
Brown, H.M. (1976). Intracellular Na+, K
+, and Cl activities in Balanus
photoreceptors. J. gen. Physiol. 68:281.
Carafoli, E. and M. Crompton (1976). Calcium ions and mitochondria. In:Calcium in Biological Systems, C.J. Duncan (ed.). Cambridge Univer-sity Press, Cambridge, (1976), p. 89.
Carnevale, N.T. (1974). Voltage clamp analysis of the slow oscillationsin bursting neurons reveals two underlying current components. Ph.D.Dissertation; Durham, Duke University, (1974).
Connor, J.m. (1977). Time course separation of two inward currents inmolluscan neurons. Brain Res. 119:487.
Connor, J.m. and C.F. Stevens (1971a). Inward and delayed outward membranecurrents in isolated neural somata under voltage clamp. J. Physiol.213:1.
Connor, J.m. and C.F. Stevens (1971b). Voltage clamp studies of a tran-sient outward current in gastropod neural somata. J. Physiol.213:21.
Connor, J.m. and C.F. Stevens (1971c). Prediction of repetitive firingbehaviour from voltage clamp data on an isolated neurone soma.J. Physiol. 213:31.
Dionne, V.E. and C.F. Stevens (1975). Voltage dependence of agonisteffectiveness at the frog neuromuscular junction: Resolution of aparadox. J. Physiol. 251:245.
DiPolo, R. (1973). Calcium efflux from internally dialyzed squid giantaxons. J. gen. Physiol. 62:575.
DiPolo, R. (1977). Characterization of the mTP-dependent calcium effluxin dialyzed squid giant axons. J. gen. Physiol. 69:795.

97
Eaton, D.C. (1972). Potassium ion accumulation near a pace-making cell ofmplysia. J. Physiol. 224:421.
Eckert, R. and H.D. Lux (1975). m non-inactivating inward current recordedduring small depolarizing voltage steps in snail pacemaker neurons.Brain Res. 83:486.
Eckert, R. and H.D. Lux (1976). m voltage-sensitive persistent calciumconductance in neuronal somata of Helix. J. Physiol. 254:129.
Eckert, R. and H.D. Lux (1977). Calcium-dependent depression of a lateonward current in snail neurons. Science 197:472.
Ehrenstein, G. and D.L. Gilbert (1966). Slow changes of potassium per-meability in the squid giant axon. Biophys. J. 6:553.
Frazier, W.T., E.R. Kandel, I. mupfermann, R. Waziri, and R.E. Coggeshall(1967). Morphological and functional properties of identifiedneurons in the abdominal ganglion of mplysia californica. J.Neurophysiol. 30:1288.
Gainer, H. (1972a). Effects of experimentally induced diapause on theelectrophysiology and protein synthesis of identified molluscanneurons. Brain Res. 39:387.
Gainer, H. (1972b). Electrophysiological behavior of an endogenouslyactive neurosecretory cell. Brain Res. 39:403.
Geduldig, D. and R. Gruener (1970). Voltage clamp of the Aplysia giantneurons: Early sodium and calcium currents. J. Physiol. 211:217.
Geduldig, D. and D. Junge (1968). Sodium and calcium components of actionpotentials in the mplysia giant neurone. J. Physiol. 199:347.
Gola, M. (1974). Neurones a ondes-salves des mollusques, variationscrcliques lentes des conductances ioniques. Pfiugers mrch. 352:17.
Goldman, D.E. (1943). Potential,impedance, and rectification in membranes.J. gen. Physiol. 27:37.
Gorman, m.L.F. and M.F. Marmor (1970). Contributions of the sodium pumpand ionic gradients to the membrane potential of a molluscan neurone.J. Physiol. 210:897.
Gorman, m.L.F. and M.V. Thomas (1977). Internal calcium changes in a ,
bursting pacemaker neuron measured with arsenazo III. Science 196:531.

98
Gradmann, D. and C.L. Slayman (1975). Oscillations of an electrogenicpump in the plasma membrane of Neurospora. J. Membrane Biol. 23:181.
Graubard, K. (1975). Voltage attenuation within mplysia neurons: Theeffect of branching pattern. Brain Res. 88:325.
Gulijarani, R.M., F.m. Roberge, P.m. Matthieu (1977). The modelling of, a burst generating neuron with a field effect transistor analog.
Biological Cybernetics 25:227.
Hagiwara (1973). Ca Spike. mdv. Biophys. 4:71.
Heyer, C.B. and H.D. Lux (1976). Properties of a facilitating calciumcurrent in pace-maker neurones of the snail, Helix pomatia. J.Physiol. 262:319.
Hodgkin, m.L. and P. Horowicz (1959). The influence of potassium andchloride on the membrane potential of single muscle fibres. J.Physiol. 148:127.
Hodgkin, m.L., and m.F. Huxley (1952). m quantitative description ofmembrane current and its application to conduction and excitationin nerve. J. Physiol. 117:500.
Hodgkin, m.L. and B. matz (1949). The effect of sodium ions on the elec-trical activity of the giant axon of the squid. J. Physiol. 108:37.
Hodgkin, m.L. and R.D. meynes (1957). Movements of labelled calcium insquid giant axons. J. Physiol. 138:253.
Junge, D. and C.L. Stephens (1973). Cyclic variation of potassium con-ductance in a burst-generating neurone in mplysia. J. Physiol.235:155.
mado, R.T. (1973). mplysia giant cell: Soma-axon voltage clamp currentdifference. Science 182:843.
Rater, S.B. and C.R.S. maneko (1972). mn endogenously bursting neuron inthe gastropod mollusc, Helisoma trivolvis: Characterization in vivo.J. comp. Physiol. 79:1.
Kostyuk, P.G., O.m. Krishtal, and P.m. Doroshenko (1974). Calcium currentsin snail neurones. I. Identification of Calcium current. PflugersArch. 348:83.
mostyuk, P.G., O.m. Kristal and V.I. Pidoplichko (1975). Effect of internalfluoride and phosphate on membrane currents during intracellulardialysis of nerve cells. Nature 257:691.

99
munze, D., T.L. Walker, H.M. Brown (1971). Potassium and chloride activ-ities in identifiable mplysia neurons. Fed. Proc. Fed. mm. Soc. Exp.Biol. 30:255.
Kupfermann, I. and K.R. Weiss (1976). Water regulation by a presumptivehormone contained in identified neurosecretory cell R15 of mplysia.J. gen. Physiol. 67:113.
Lee, K.S„ N. mkaike, and m.M. Brown (1977). Trypsin inhibits the actionof tetrodotoxin on neurones. Nature 265:751.
Lux, H.D. and R. Eckert (1974). Inferred slow inward current in snailneurones. Nature 250:574.
Luxoro, M. and E. Yanez (1968). Permeability of the giant axon ofDosidicus gigas to calcium ions. J. gen. Physiol. 51:115.
Marmor, M.F. (1971). The effects of temperature and ions on the current-voltage relation and electrical characteristics of a molluscanneurone. J. Physiol. 218:573.
Meech, R.W. (1972). Intracellular calcium injection causes increasedpotassium conductance in mplysia nerve cells. Comp. Biochem.Physiol. 42m:493.
Meech, R.W. (1974a). The sensitivity of Helix aspersa neurones to injectedcalcium ions. J. Physiol. 237:259.
Meech, R.W. (1974b). Calcium influx induces a post-tetanic hyperpolariza-tion in mplysia neurones. Comp. Biochem. Physiol. 48m:387.
Meech, R.W. (1974c). Prolonged action potentials in mplysia neuronesinjected with EGTA. Comp. Biochem. Physiol. 48m:397.
Meech, R.W. and N.B. Standen (1975). Potassium activation in Helix aspersa neurones under voltage clamp; a component mediated by calcium influx.J. Physiol. 249:211.
Meech, R.W. and F. Strumwasser (1970). Intracellular calcium injectionactivates potassium conductance in mplysia nerve cells. Fedn Proc.29:834.
Meech, R.W. and R.C. Thomas (1977). The effect of calcium injection on theintracellular sodium and pH of snail neurones. J. Physiol. 265:867.
Mirolli, M. and S. Talbott (1972). The geometrical factors determiningthe electrotonic properties of a molluscan neurone. J. Physiol.227:19.

100
Miura, D.S., B.F. Hoffman, and M.R. Rosen (1977). The effect of extra-cellular potassium on the intracellular potassion ion activity andtransmembrane potentials of beating canine cardiac Purkinje fibers.J. gen. Physiol. 69:463.
Neher, E. (1971). Two fast transient current components during voltageclamp on snail neurones. J. gen. Physiol. 58:36.
Neher, E. and H.D. Lux (1973). Rapid changes of potassium concentrationat the outer surface of exposed single neurons during membranecurrent flow. J. gen. Physiol. 61:385.
Noble, D. and R.W. Tsien (1968). The kinetics and rectifier properties ofthe slow potassium current in cardiac Purkinje fibres. J: Physiol.195:185.
Noble, D. and R.W. Tsien (1969). Outward membrane currents activated inthe plateau range of potentials in cardiac Purkinje fibres. J.Physiol. 200:205.
Parnas, I., D. mrmstrong, and F. Strumwasser (1974). Prolonged excitatoryand inhibitory synaptic modulation of a bursting pacemaker neuron.J. Neurophysiol. 37(4):591.
Partridge, L.D. (1975). Pentylenetetrazol-induced bursting activity inthe absence of a negative conductance characteristic. Brain Res.94:161.
Partridge, L.D. and C.F. Stevens (1976). m mechanism for spike frequencyadaptation. J. Physiol. 256:315.
Pinsker, H.M. and E.R. mandel (1967). Contingent modification of anendogenous bursting rhythm by monosynaptic inhibition. Physiologist,10:279.
Plant, R.E. and M. mim. (1976). Mathematical description of a burstingpacemaker neuron by a modification of the Hodgkin-Huxley equations.Biophys. J. 16:227.
Prior, D.J. and m. Gelperin (1977). mutoactive molluscan neuron: Reflexfunction and synaptic modulation during feeding in the terrestrialslug Limax maximus. J. comp. Physiol. 114:217.
Rose, B. and W.R. Loewenstein (1975). Calcium ion distribution in cytosolvisualized by mequorin: Distribution in the cytosol is restricteddue to energized sequestering. Science 190:1204.
Russell, J.M. and m.M. Brown (1972). mctive transport of potassium bythe giant neuron of the mplysia abdominal ganglion. J. gen. Physiol.60:519.

101
Smith, T.G. Jr., J.L. Barker and H. Gainer (1975). Requirements forbursting pacemaker potential activity in molluscan neurons. Nature253:450.
Standen, N.B. (1975). Voltage-clamp studies of the calcium inward currentin an identified snail neurone: Comparison with the sodium inwardcurrent. J. Physiol. 249:253.
Stinnakre, J. and L. Tauc (1966). Effets de l'activation osmotique del'osphradium sur les neurones du systeme nerveux central de l'mplysie.J. Physiol. (Paris) 58:266.
Stinnakre, J. and L. Tauc (1969). Central neuronal response to the acti-vation of osmoreceptors in the osphradium of mplysia. J. Exp. Biol.51:347.
Stinnakre, J. and L. Tauc (1973). Calcium inflex in active mplysia neurones detected by injected aequorin. Nature New Biol. 242:113.
Strumwasser, F. (1965). The demonstration and manipulation of a circadianrhythm in a single neuron. Circadian Clocks, mschoff, J. (ed.). NorthHolland Publishing, mmsterdam, p. 42.
Tiffert, J.T., F.J. Brinley, Jr. and m. Scarpa (1977). Regulation of freecalcium concentration by mitochondria and cytoplasmic buffering insquid axons. Fed. Proc. 36:485.
Thomas, R.C. (1972a). Electrogenic sodium pump in nerve and muscle cells.Physiol. Rev. 52:563.
Thomas, R.C. (1972b). Intracellular sodium activity and the sodium pumpin snail neurones. J. Physiol. 220:55.
Thompson, S.H. (1976). Membrane currents underlying bursting in molluscanpacemaker neurons. Ph.D. Dissertation, University of Washington,Seattle.
Thompson, S.H. (1977). Three pharmacologically distinct potassiumchannels in molluscan neurones. J. Physiol. 265:465.
Thompson, S.H. and S.J. Smith (1976). Depolarizing afterpotentials andburst production in molluscan pacemaker neurons. J. Neurophysiol.39:153.
Willows, m.O.D., D.A. Dorsett and G. Hoyle (1973). The neuronal basis ofbehavior in Tritonia. I. Functional organization of the centralnervous system. J. Neurobiol. 4:207.
Wilson, W.m. and H. Wachtel (1974). Negative resistance characteristicessential for the maintenance of slow oscillations in burstingneurons. Science 186:932.

VITA
mUTHOR: Stephen J. Smith
DATE OF BIRTH: March 22, 1947
PLmCE OF BIRTH: Santa Monica, California
MOTHER: Lois Marie Gairich
FATHER: Donald J. Smith
SECONDmRY EDUCATION: Mercer Island High SchoolMercer Island, Washington
DEGREES: Reed CollegePortland, OregonB.m. Psychology, 1970
University of WashingtonSeattle, WashingtonPh.D. Physiology-Psychology, 1978