Embed Size (px)
Citation preview

SEGRl!~TORY ACTIVITY OF THE CITORIOTD PLEXGS I N TISSTYE CTJLTURE
GLADYS CAMERON Iwtitule for Medical Research, C e d 4 m o f Lebanon l lospi tal
Los Angeles, Califomiw
SIX FIGURES
Tissue culture studies were made of the chorioid plexus of chick, ra t and rabbit embryos and of three-day-old hatched chicks. It was found that solutions of certain colored indi- cators when incorporated in the external medium of cultures of the intact chorioid plexus were picked up and transported through the ependymal epithelium covering the plexus and ac- cumulated in the tissue spaces of the plexus. Occasionally the plexus was broken or cut in removal from the embryo, in which case the ependymal epithelium grew out in sheets from the cut ends of the villi. Vesicles filled with fluid gradually developed in these ependymal sheets. I n both cases the phe- nomenon was reversibly inhibited by cold and by oxygen lack indicating a positive vital proccss, secretory in nature, of the eperidynia of the chorioid plexus.
Divergence of opiiiiori still exists concerning the role of the chorioid plexus. Faivre (1853) and Luschka (1855) sug- gested that its function is secretory, Mestrezat ( '12) that the ependyinal covering acts as a dialyzing membrane, others that it maintains a hemoencephalic barrier, and still others that it is absorptive in function. Merritt and F. Fremont- Smith ( ' 3 7 ) discuss the findings which support the various theories.
The early workers in tissue culture obtained and reported the growth of sheets of cells which were unorganized. Now it is well established that fragmciits of tissues may be grown
115

116 GLADYS CAMERON
in tissue culture in such H niaiiner that they coiitiiiue, for a time at least, to perform their specific functions. Oiie of the most striking examples is the seci*etory activity of mcso- iiephric tubules in tissue culture. This has been the basis for the study by Chambers and his co-workers ( '33) of different phases of renal activity by observing the accumulation, under controlled conditioiis, of solutions of various acid dyes in tho luniiiia of proximal convoluted tubules.
This report pi*esents evidcnce in tissue culture that the epeiidymal cells of tlie chorioid plexus possess the ability actively to ti*ansfer material fr-orn the cistcrnal rnediuni into the tissue spaces of tlie chorioid plexus.
E X P Eli1 ME N T A 1 2
The tissue used €01. these experiments was the vasculai., fringe-like border of folded pia rniater, the chorioid plexus, which is covered with cpeiidyma and forms the roof of the third and fourth ventricles, also pai*t of the wall of the two lateral ventricles. The plexus fi.orn the various regions was used indiscriminately. It was dissected from the chick brain of from 8 to 20 days' iiicubatioii and from chicks three days after hatching. Similar tissue f iwn rat and rabhit ern1)ryos near term was also used. Occasionally, tag ends of epeiidymal epithelium were present but usually thc plexus was obtained and cultured intact.
('TIIC'K TISSVES
The fragmeiits of tissue were cultured by the douhle cover- slip method, the inediurri coiisistiiig of oiic par t fowl plasrmi arid one part Tyi-ode solution coiitaiiiiiig 10% c1rnl)ryo extract. The mixture coiitaiiiiiig ii tissue f i . ape i i t was allomcd to clot oil a srnall covei*slip which, attached to a larg:.ci. coverslip, wiis sealed with paraffiii over a depression slide aiicl the cul- ture iiicu1)atec-l at 37.5"('. The appearance and behaviol. of the cultur*al fmgrneiits f rom chicks of all the ages I X S C ~ w e i ~ ~ similar.

~licroscopic examiliatioil of the freshly p r e p a i d cultuws of the intact plexus before incubation showed a tangled skein of minute hlood vessels and capillaries entirely covered with cuboidal, eperidynial epithelium (fig. I ) . On the free external surface of this epithelium, motile cilia could he seen. The capil1ai.y tufts were distinct and the red blood cells in them plainly visilole. After 24 hours' iiicubation an area of liquc- faction of the plasma clot, such as occurs in cultures of most types of epithelium, was evidciit immediately surrounding the cxplant. Similar liquefaction has been reported by Weiss ( '33) in his tissue culture studies of mid-brain from chick embryos. At this time the plexus hecame swollen and cyst- like. Spaces had formed betweeii the hlood vessels aiid the epeiidymal epitheliuni covering the plexus xvhich appeared to he filled with fluid. These cyst-like structures followed the contour of the blood vessels. During the following several days, the cysts pi-ogi-cssively iiicreased in size a i d the cuboidal epithelium flatteiied u s these bloated structures hecame more aiid more distended (figs. 2, 3 ) . By this time tho hlood vessels were almost indiscei.nihlc, probably degeiierated, although a few intact red cells were sometimes present. A loose iiet\voi.k of coiinective tissue could b e seen within the distended cysts (figs. 2, 3), and some isolated cells wandtbi-ing freely in the surrounding medium.
Fragments of epcndyma occasioiially were found growiiig as unorganized sheets, in some areas of which cilia could be seen lmiting on the epithelial surface. After 6 or 7 days' incubation, vesicles began to form in these epeiidymal sheets. They were small at first but gradually inc1.cascd in size a s fluid accumulated in them. I n these vesicles the cilia were found to he on the inner border of the cells and the cellular components were entirely epithelial (fig. 4). This is in con- t ras t to cultures of the intact plexus in which a connective tissue network could he seen iii the bloated sacs, and the cilia were found on the outel. surface of the ependymal covering. By renewal of the n?ediuni every few days, the cultures were maititaiiiecl in active coiiditioii for two or three weeks with

118 GLADYS CAMERON
progressive increase in the size of the edematous cysts in the intact plexus and in the size and number of the epithelial vesicles.
The increasing distension of the chorioidal explant sug- gested secretory activity of the cove~ing ependyma. This finding was tested by exposing the cultures to several colored compounds which had previously heen found to he activcly picked up and accumulated in the cysts of proximal kidney tubules. The compounds used were phenol i ~ d , chlor phenol red and Orange G. Phenol red and chlor phenol red a re non- toxic, acid, sulphonated dyes which belong to the Clark and Lubs series of colorimetric pH indicators. Orange G (C.I. no. 27) is also a relatively non-toxic, acid, sulphonated dye. It does not act as a pH indicator. Neutral red is a basic dye and well known vital stain coloring the structural components of the cytoplasm. The coloration is iiidcpcndeiit of any physio- logical activity. This was found to he true also of the tissues of the chorioid plexus.
Phenol red was addcd to the medium of some of the cultures during preparation (fiirisl coriceiitration about 0.025% ). Within a few hours the cells of the epeiidymal layer were pale yellow (the acid range of phenol red) , while the fluid in the iiarrow space between the capillaries and cpendyma mas pale pink (the alkaline range of phenol red). After 24 hours a striking picture presented itself. The cells covering the plexus were still yellow and the spaces between the blood vessels and the ependyma had greatly increased in size and ~ve re filled with deep rose-colored fluid. By contrast, the surrounding medium was colorless or very pale pink. At no t h e were cells colored except those of the ependymal epithelium surrounding the tissue spaces of the plexus. Chlor phenol red and Orange G WWY also added to the medium of some of the cultures. I n the case of chlor phenol red, the epcndymal cells developed a pale lavender color while the fluid within the now swollen edematous chorioid plexus was a brilliant purple (fig. 5). I n cultures with Orange Q, the ependyma and contained fluid were yellow. Phenol red was also added to week-old cultures

in which sheets of ependynia containing vesicles were present. A faint yellow tint could be seen only in the ependymal epi- thelium which delimited the vesicles while the contained fluid became pale pink. The coloration of both cells and fluid was less intense than in the intact chorioid plexus. This reduced degree of activity of the vesicles in chorioidal ependpmal sheets is in accord with results for cysts formed in outgrowing sheets of renal epithelium where the secretory activity was found to be less than that of the originally explanted meso- iiephric tubules. The less intense color. in the vesicles and cysts which formed in outgrowing epithelial sheets was pos- sibly due to the age of the cultures. It has generally been our experience that secretory activity slows down in propor- tion to the aging of the cultures.
RAT AND RABBIT TTSSUES
Similar experiments were done with the chorioid plexus from rat and rabbit embryos near term. Formation of edema- tous cysts filled with fluid and dye occurred. For the first few days no ciliary movement was seen, but, after one week, active ciliary beating could be seen on the outer border of the ependyma covering the plexus and hy contrast on the inner border of the cells which lined newly formed vesicles in the outgrowing sheets of ependymal epithelium (figs. 4, 6) . When phenol red was added to these cultures, a eoncentration of the indicator occurred within the new vesicles, while the original, bloated cysts, as described in the chick plexus, were large and contained much fluid and dye.
To determine whether the accumulation of fluid and co~i- centration of dye in the swollen chorioid plexus were more than the result of physical diffusion, some cultures containing the dyes were left at room temperature, about 20°C., and others kept a t 10°C., for 12 hours. Those at room temperature showed a much reduced color and fluid accumulation while those at 10°C., showed none. When returned to the incubator at 37.5"C., accumulatioii of fluid with concentration of the

120 GLADYS CANEROX
dyes in the tissue spaces was resumed. The cilia which had been motionless in the cold i*esumed beating.
Xitrogcn atmosphere was also used arid found to be i111
inhibiting agent. For these esperinieiits thc intact chorioid plexus was cultivated in 1-ollcr tubes. The tissue w a s fastened to the wall of the tube by a filni of blood plasma and a super- natant fluid coiitaiiiing phenol iaed added. The control culture tubes had free access to ail. while thc experiniciital tubes were filled with nitrogen. I n the control cultures the tissue spaces, a s pi*cvlously described, becanic swollen and edema- tous. Thcy m i . e bounded by yellow colored epeiidyrnal epi- thcliurn and coiitained i.ose-colored fluid. Actively lwating cilia wci*e seen. On the other hand, cultures exposed to nitro- gen showed no edema n o r any coloration of the cells and no ha t i i i g cilia. Several hours aftei. 1.etu1.n to a normal at- mosphere, accuniulatioti of fluid atid phenol i*ed was secii in the tissue spaces of the plexus aiicl the cilia bccanic active.
1) IS( ’1 ‘ ss TO N
Val-ious investigators have suggested that the chorioid plexus possesses a s c c i ~ t o r y function. Pettit and Girarcl (’02) showed that drugs which stirnulate secretion, such as pilocarpine, muscariiie, otc., inci*case cei.ehi*o-spinal fluid and also CRUSC histological changes with the development of gi’aii- ulcs in the cells of the chorioid plexus suggestive of secretory activity. Wced ( ’Xi?) showed that, during development, the histological difierentiation of the ependynial cells of the chori- oid plexus w a s accompanied by pi*oduction of spiiial fluid and IIogue (’47, ?49) from her study of the cytology of the cells of human fetal chorioid plexus iii tissue cu1tui.e con- cluded that the epctidyrnal cells hare a secretory function. I n some cultures slio observed “hranching vesicles” lined with high ciliated columnar cells which contained gi-anules “probably secreted by the cpendynial cells. ” She also noted the “pinching off of parts of cpendynial cells” which she interpreted as apocrinci secretion. The formation of “glob- ules ” which wei-c not pinched off hut which discharged their

121 CHORIOII) I'LEXPS IS TISSI-E: CULTI'RE
clear contents iiito the surrounding iriedium suggested still another type of secretion.
The observations described in this paper 1.eveal conclu- sively that the chorioidal epithelium possesses a. secretory function. In tissue culture the intact chorioid plexus becomes swollen and edematous with fluid and acid sulphoiittted dyes which accuiiiulate in the tissue spaces hetween the ependpmwl covering and the blood vessels. From cut fragments of the plexus, sheets of epciidyi-rial epithelium gi*o\\- in which fluid filled vesicles develop. In both cases the phenonieiion is re- versibly inhibited by nietaholic depressants such as low tem- perature and the lack of oxygen. The results a re similar to those obtained using isolated, proximal, coiivoluted kidney tubules in which the accumulation of sulphonated dycs and fluid within the lurriina of the tuhulcs and in cysts which de- veloped in outgrowths of renal epithelium was shown to be a secretory process. It is still a question whethey the trans- port of fluid and dye into the tissue spaces of the plexus is made possible by the removal of osrriotically active substances f rom the tissue spaces or whether it is due to the secretion of fluid into them by the ependyma. It is evideiit, however, that the functional activity resides in the ependynial cells. They absorb the acid sulphoriated dyes from the esterrial medium aiid transfer them into the tissue spaces of the chorioid plexus. The same also occurs in the case of the vesicles which develop in outgrowths of epcndymal epithelium.
When the intact plexus is removed from the emhi-yo, the cilia are seen to be on the outer, free surface of the cuboidal epitlielium covering i t and the dyes and fluid a re transferred from the external medium through the outer ciliated surface of the epithelial cell and thence through the inner lion-ciliated surface into the tissue spaces of the plexus. However, in the case of the vesicles which develop in the outgrowth of the ependymal epithelium, the cilia line the internal surface of the vesicles. I n this case the dye and fluid a re transported through the outer non-ciliated surface and thence through the ciliated surface of the cells to the vesicular cavity. Because

122 GLADYS CAMERON
of the reversal of the position of the cilia in the cysts of the original explant as compared with those of the newly formed vesicles, it would appear that the functional polarity of the epithelial cells is not necessarily related to the location of the cilia, but is dependent upon the orientation of epithelial cells around a cavity.
I n the present experiments no attenlpt is being made to specify the nature of the fluid accumulated in the edematous cysts of the intact chorioid plexus or in the vesicles formed in outgrowing ependymal sheets. The findings demonstmte thc accumulation of fluid and concentration of colored com- pounds under pressui*e in the tissue spaces of the chorioid plexus. This phenomenon is obviously secretory in nature since it is reversibly inhihited by metabolic depressants such as cold and oxygen lack.
1. Chorioid plexus from rat and rabbit embryos near term, from chick embryos of from 8 to 20 days’ incubation and from chicks three days after hatching were grown in tissue culture for about three weeks.
2. Closed cysts, distended with fluid, developed in the tis- sue spaces of the plcxus and in vesicles in the outgrowing ependymal sheets.
3. Active cilia were seen on the external border of cells covering the plexus and on the inner border of those limiting the vesicles formed in outgrowing sheets of ependyma. 4. Phenol red, chlor phenol red and Orange G (C.I. no.
27) when added to the cstcviial medium passed into and accumulated in the tissue spaces of the intact plexus and also in vesicles which developed in sheets of ependymal epithelium.
5. The accumulation of fluid and of thc colored compounds was reversibly inhibited by cold and by oxygen lack.
6. These results constitute evidence of the physiological activity of the ependyma of the chorioid plexus and indicate that it is of a secretory nature.

CHORIOID PLEXl‘S I N TISSUE CULTURE 123
ACKNOWLEDGMENTS
The author wishes to thank Dr. Harry Goldblatt, Director of the Institute f o r Medical Research of Cedars of Lebanon Hospital, Los Angeles, for his cooperative interest in this work, and Mrs. Virginia Sheehan for skillful technical as- sistance. Gratitude is expressed to Dr. Robert Chambers for his advice and aid in the preparation of the manuscript, and to A h . C. G. Grand f o r his kindness in preparing the photo- micrographs.
LITERATURE CITED
CHAMBERS, R., AND R. T. KEMPTON 1933 Indications of function of the chick nieso-nephros in tissue culture with phenoI red. J. Cell, and Comp. Pliysiol., 3: 131.
FAIVRE, E. Rcrheiches sur la structuie du conarium e t des plexus chorioides chez l’homme et les animaux. Compt. rend, Acad. d. Sci., 39: 424.
HOGUE, M ~ R Y JANE 1947 Hunian fetal brain (*ells in tissue cultures, their identi fieation and motility. J. Exp. Zool., 206: 85.
_____ 1949 Human fetal clioroid plexus cells in tissue cultures. Anat. Rcc., 103: 381.
LVSCIIKA, H. 1855 Die Adergeflrclite des mensclilicl~en Gehirns, Berlin. MERRITT, 11. I%., AND F. FREMONT-SMITH 1937 The Cerebrospinal Fluid. w. B.
Saunders Company, Philadelphia. METRFZAT, W. 1912 Le liquicle rephalorachidien normal ct pathologique. Valcur
cliniyue dc l’examine cliimique. Syndromes liuinoraux dnns les diverscs affections. A . Malone, Paris.
8ur la b’onction Sccretoire et la nmrphologir des Plexus Cho~ioirles des Ventriculcs Latereaux du Systcine Nerveux Centiale. Arch. d’anat. Microsc., 373: 151.
1934
1853
P E T w r , A., AND J. GIRARD 1902
WEED, L. H. 1922 The cerebrospinal fluid. Physiol. Rev., 3: 171. WEISS, PAIJL Secretory ac tk i ty of the inner layer of the rnibryonic mid-
brain of tlic cliick as revealed by tissue eulture. Anat. Rer., 58: 299.

1 Liviiig culture of 1 0 ~ d a y vliick chorioid plexus, 1 2 Iiours in culture. Sate tlrptli of cuboidal cpelidyiiial epithcliiuii eiiclosing blood capillaries. KO peri- r:iscul:ir spaces visible. X 50.
2 S:iiiic culture :IS figurtt 1 10 days later, sliowing swollen edeiiiatous plcsus xvitli fla tteiictl e~ i r i idy i i i :~~ el)it,licliuiii. Loose i ic t~rork of coiinectirc tissue visililc in cysts. X 50.
3 Living ciilture ~f l4-tlay (+hick cliorioitl plexus, 5 clays in culture. Mwli distciitlcd with fluid aiid plienol red. El~eiiclyiii:~ p;ii-tiaIly flatteiietl. X 100.
4 Fixed and stained culture of cliorioid plexus from rat embryo i i w r tcriii. Twelve clays' growth. Sheet is coinposed entire17 of epe i idp ia l cl~itlieliuiii wit11 foriiiation of rcsicles a t lower left. x 200.
6 Living culture of 18-d:ry chick cliorioiil ~ ~ l e x u s , three d:iys in culture. 'l'issuc spaces distended with fluid and clilor pliciiol red (slio\rs black). l3peiidpia bordering cysts is colorless aiid partially flatteireil. W:iiidering cells i i i stir- rouiidiiig Iiiediuiii. X 100.
(i Fixed and staiiictl cul ture of cliorioid plexus froni ra1)bit einl~ryo iirar t c r~n . 9' ,. i.xtecii I
resicles. X 300. (lays' yrowtli of sliett of cpeiir1yin:iI rpithcliuni with foririntioii of 1:ti.ge
124
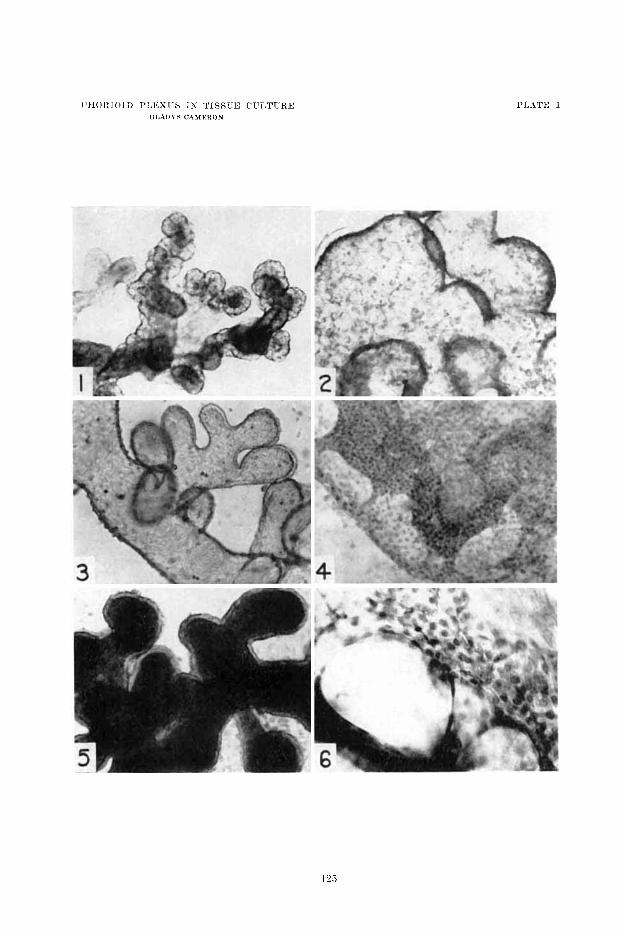
PLATE 1