Embed Size (px)
Citation preview

BASIC SCIENCE
Nanomedicine: Nanotechnology, Biology, and Medicine10 (2014) 839–849
Original Article nanomedjournal.com
Secretion of intestinal goblet cells: A novel excretion pathwayof nanoparticles
Baoquan Zhao, PhDa,1, Lan Sun, PhDa,1, Wuxu Zhang, MSca,c, Yuxia Wang, MD, PhDa,Junjing Zhub, Xiaoyu Zhub, Liuzhong Yang, MMa, Chunqi Li, MD, PhDb,
Zhenzhong Zhangc, Yingge Zhang, MD, PhDa,⁎aInstitute of Pharmacology and Toxicology and Key Laboratory of Nanopharmacology and NanoToxicology, Beijing Academy of Medical Sciences,
Beijing, ChinabHangzhou Hunter Biotechnology Incorporation, Hangzhou, China
cSchool of Pharmacy; and Nanotechnology Research Center for Drugs; Zhengzhou University, Zhengzhou, China
Received 6 May 2013; accepted 18 October 2013
Abstract
Understanding the excretion pathway is one of the most important prerequisites for the safe use of nanoparticles in biomedicine.However, the excretion of nanoparticles in animals remains largely unknown, except for some particles very small in size. Here we report anovel natural pathway for nanoparticle excretion, the intestinal goblet cell (GC) secretion pathway (IGCSP). Direct live observation of thebehavior of 30-200 nm activated carbon nanoparticles (ACNP) demonstrated that ACNP microinjected into the yolk sac of zebrafish can beexcreted directly through intestinal tract without involving the hepato-biliary (hap-bile) system. Histopathological examination in mice afterligation of the common bile duct (CBD) demonstrated that the intravenously-injected ACNP were excreted into the gut lumen through thesecretion of intestinal GCs. ACNP in various secretion phases were revealed by histopathological examination and transmission electronmicroscopy (TEM). IGCSP, in combination with renal and hap-bile pathways, constitutes a complete nanoparticle excretion mechanism.
From the Clinical Editor: Nanoparticle elimination pathways are in the forefront of interest in an effort to optimize and enable nanomedicineapplications. This team of authors reports a novel natural pathway for nanoparticle excretion, the intestinal goblet cell (GC) secretionpathway (IGCSP). Direct live observation of the behavior of activated carbon nanoparticles has shown excretion directly through theintestinal tract without involving the hepato-biliary (hap-bile) system in a zebrafish model.© 2014 Elsevier Inc. All rights reserved.
Key words: Nanoparticle; Excretion; Pathway; Goblet cell; Aggregation
The understanding of the excretion pathway becomes moreand more important with the development of nanotechnologyand the application of nanomaterials in biomedicine. It isimportant not only for the biosafety issue of engineerednanoparticles but also for the practical use of nanoparticles asdiagnostic and therapeutic agents or as drug carriers, becauseexcretion is the best way to cease the action of nanoparticles ontissues and cells. Based on the understanding of the excretionpathway, appropriate protocols can be worked out to deal withthe nanoparticles once they entered the animal body. Unfortu-
This work was supported by the National Natural Science Foundation ofChina (No. 90406024), the National Basic Research Program of China (No.2010CB933904) andMajor New Drug Creations (No. 2011ZX09102-001-15).
⁎Corresponding author at: Beijing Institute of Pharmacology andToxicology, Beijing, 100850, PR China.
E-mail address: [email protected] (Y. Zhang).1 Equally contributed to the work.
1549-9634/$ – see front matter © 2014 Elsevier Inc. All rights reserved.http://dx.doi.org/10.1016/j.nano.2013.10.004
Please cite this article as: Zhao B, et al, Secretion of intestinal goblet cells: A novehttp://dx.doi.org/10.1016/j.nano.2013.10.004
nately, such pathway remains poorly understood, though thereare many literatures on the clearance of nanoparticles fromblood1,2 or tissues such as lung3,4 and liver.5 These experimentsprovide information concerning the clearance mechanism toremove particles from local tissues rather than information onsystemic excretion of nanoparticles.6 Few studies have reportedtwo main excretion pathways of intravenously-injected nano-particles, the kidney–urine pathway and hepatobiliary system(HBS)–feces pathway. Kidney excretion of nanoparticles islimited to very small ones, such as quantum dots7 andfullerines.8 HBS excretes some larger nanoparticles, but theclearance rate is no more than 1% within 24 h, and there is aninverse relation between HBS excretion and sizes.9 However,Manabe et al found that 500 nm latex particles can be clearedfrom medaca embryos,10 suggesting that there are some otherexcretion pathways for nanoparticle excretion without involvingthe HBS. Souris et al reported that the concentration ofintravenously-injected 50-100 nm silica nanoparticles in liver
l excretion pathway of nanoparticles. Nanomedicine: NBM 2014;10:839-849,

840 B. Zhao et al / Nanomedicine: Nanotechnology, Biology, and Medicine 10 (2014) 839–849
was much lower than that in intestinal wall and feces.11 Thisinconsistence in quantity does not support the hypothesis thatHBS plays main roles in the intestinal excretion of nanoparticles.On the contrary, the high concentration of nanoparticles inintestinal wall11 suggested that the intestinal wall may playimportant roles in the excretion of nanoparticles through intestinaltract. GCs are one of the four main cell types12 in the intestinalmembrane and have the ability to entrap nanoparticles.13 Recently,there have been several studies on the interactions between GCsand orally-administered nanoparticles.13–16 GCs can uptakenanoparticle,17 which makes GCs the most possible candidatecells playing roles in the intestinal excretion of nanoparticles. Ourearlier studies found that intravenously-injected nanoparticles haddistribution in intestinal GCs18 and hypothesized that intestinalwall may play roles in the excretion of nanoparticles but did notinvestigate in detail.18,19 The present work carefully examined themechanism for intestinal excretion of nanoparticles and revealed anovel pathway, IGCSP, for nanoparticle excretion.
Methods
Preparation and characterization of ACNP
ACNP were prepared from medicinal activated carbon(MAC; Haichangqing Co. Ltd, Beijing, China) by a top-downmethod (supplemental materials). The diameters of ACNP weredetermined by atomic force microscope (AFM), scanningelectron microscope (SEM) and Laser Particle Size and ZetaPotential Analyzer. The internal crystal structures were detectedby X-ray defraction (XRD). For injection, 1 mg ACNP wasadded into 100 mL normal saline and suspended in an ultrasoundfield of 40 Hz, 180 W for 20 min.
Handling of the animals
Zebrafish eggs were collected, cleaned, and washed with egg-water20 within one hour after fertilization. Eggs from differentfemales were pooled, placed in 7 cm sterile Petri dishescontaining eggwater, and incubated at 28.0 to 28.5 °C.Embryonic fish were stocked in an Aquatic Habitat re-circulatingtank system at 28.5 °C with a 14 h light/10 h dark cycle. Thewater was purified by reverse osmosis and adjusted to pH 7 andconductivity of 350 μS. The mice were handled as describedpreviously.21 Briefly, 20 Kunming mice (20-25 g in bodyweight) were constantly monitored and fed fluid nutritional dietfree from pathogens and particulate materials for 10 days beforeACNP treatment to avoid the influences of ingested particles onthe observation of ACNP. All the animal procedures wereapproved by the Animal Subject Review Committee of theBeijing Academy of Medical Science.
Pigment inhibition in zebrafish
Pigmentation genes were suppressed by adding 1-phenyl,2-thiourea (PTU) (Sigma, USA) into fish water in a finalconcentration of 75 μM in the developmental stage of28 somites. 22 This concentration produced enoughpigment inhibition with no adverse effects on the hatchingand survival.
Microinjection and observation of ACNP in zebrafish
Embryonic zebrafish at the age of 24 h were put into a 7-cmculture dish and anesthetized by addition of tricaine methane-sulfonate into the dish at a final concentration of 0.64 mM. Theanesthetized zebrafish were placed in the slanting grooves of asilica gel sheet. Excessive water around the fish wasabsorbed with filter papers to leave just enough water to bathethe fish body. Five μL suspension of ACNP at the concentrationof 5 mg/mL was injected into the yolk sac of zebrafish in an IM300 microinjection instrument (Narishige, Japan) (Figure S3).All ACNP suspensions were freshly dispersed by ultrasonicationfor 10 min before use. During the whole period ofexperiment, zebrafish that received microinjection of ACNPexhibited no significant differences in death rate, development,teratogenesis, and cardiovascular toxicity in comparison withcontrol (Figure S4).
Ligation of common bile duct and injection of ACNP intothe mice
The common bile duct was ligated with the method originallydescribed by Cameron and Oakley23 with modifications. Briefly,Kunming mice of 20-25 g were anaesthetized with pentobarbital(25 mg/kg) and fixed onto a wood surgical sheet. A mid-abdominal incision was made, and the abdominal tissues wereseparated carefully to clearly expose the CBD. Two sterile nylonmedical surgical sutures (Unic Surgical Sutures, Mfg., Co., Ltd.,Suzhou, China), 0.2 mm in diameter, were put through under theCBD, and two nodes were made at both ends of a segment ofCBD (Figure S5), and the CBD was then cut off between the twoends. After closure of abdomen, ACNP were suspended in 0.9%NaCl to a final concentration of 5 mg/mL and injected throughtail veins in a dose of 50 mg/kg (i.e., 10 mL/kg). All thesuspensions of ACNP were freshly dispersed by sonication for10 min before use. After the injection of ACNP, the animals'body weight, behavior, and number of blood cells weremonitored (Figure S6). At the end of the experiments, thesephysical parameters showed no significant differences fromthose of control (Figure S6).
Histopathological examination
On the 4th day after injection of ACNP, the micewere anaesthetized and their internal organs were taken outand fixed in 40% formaldehyde. 5 μM thick sectionswere made on a Lecia RM2135 Rotary Microtome. Thesections were stained in hematoxylin and eosin or alcianblue and observed under an Olympus BH2 phase contrastmicroscope (Japan).
Quantitative evaluation of the efficiency of IGCSP for ACNP
The quantitative evaluation was carried out with ratesoccupied by ACNP-containing GCs in the total number of theGCs seen under a phase light microscope in 10 visions in 3 HE-stained histopathological sections, which were calculated by aformula: Rates (%) = Number of ACNP containing GCs/totalnumber of seen GCs×100%. 120 Mice without CBD ligationwere randomly divided into 4 groups and 30, 60, 100 and

Figure 1. ACNP characteristics. (A) The AFM images of the prepared ACNP of 30, 60, 100, and 200 nm. (B) The SEM image of 100 nm ACNP (left), which isspheric with many micropores in the sphere (right).
841B. Zhao et al / Nanomedicine: Nanotechnology, Biology, and Medicine 10 (2014) 839–849
200 nm ACNP were intravenously injected respectively. Thedifferent organs of every 3 mice in each group were respectivelytaken out for quantitative histopathological examination on day1-180 after ACNP injection.
Results
Characteristics of ACNP
Structurally, ACNP belong to a large family of carbon-based nanoparticles composed of graphene sheets24–26. Theprepared ACNP had diameters of 30, 60, 100, and 200 nm,respectively, as imaged with AFM (Figure 1, A) and SEM(Figure 1, B), which are spheres with mesopores (Figure 1, B).XRD revealed that the graphite microcrystals of ACNP have athickness of 0.707 nm, a width of 2.007 nm and a layerdistance of 0.372 nm. The ζ electropotentials are 0.65-6 mV inpH 7.4 saline. The measurements of some structures andproperties of ACNP in different sizes are listed in Table S1.Withdrawing test demonstrated that the suspension was stablewithin 2 h.
Tissue distribution of ACNP in zebrafish
Under microscope, the black ACNP were well visible inthe transparent embryonic zebrafish. The suspension of ACNPappeared in the yolk sac as cloud-like immediately after theinjection (Figure S3, B). Within 3 h, most ACNP aggregatedinto larger recognizable particles (Figure S3, B). In 0-4 hafter injection, ACNP mainly accumulated in the yolk sac(Figure 2, A). With the development of zebrafish, ACNPgradually distributed to intestinal wall (Figures 2 and 3, A),extracranial tissues (Figure 3, B), myocardium (Figure 3, C),kidney (Figure 3, D), and blood vessels (Figure 3, E).
Because HBS is a known excretion pathway of nanoparticles,we paid special attention to the distribution of ACNP in liverand gallbladder. Surprisingly, there were little ACNP found inliver and no ACNP in gallbladder (Figure 3, D), which doesnot support the hypothesis that HBS is the main pathway forin the excretion of ACNP.
Excretion of ACNP through intestinal tract of zebrafish
Since there were little distribution of ACNP in liver and noACNP found in gallbladder, there should be no ACNP inintestinal tract. Again to our surprise, a great amount of ACNPwas observed in and excreted through intestinal tract with theperistalsis of the intestines as demonstrated in Figure 2. Aftermicroinjection, ACNP appeared as small aggregated particles(circles) in the yolk sac at the fourth hour, when the intestinaltract had not developed (Figure 2, A). On the third day, theintestinal tract (green arrow) had developed, and most yolk sachad been absorbed with a small part remaining (yellow arrow).No ACNP were in the intestinal tract; most ACNP still existed inthe residual yolk sac, and some of them appeared in the tissuesbetween intestinal wall (red arrow) and abdominal skin (Figure 2,B). On the fourth day, yolk sac was completely absorbed. ACNPbegan to appear in the intestinal wall (red arrows) and gut lumen(green arrow) (Figure 2, C). On the fifth day, more ACNPentered the intestinal tract (green arrow, Figure 2, D). On thesixth day, the quantity of ACNP in intestinal tract (green arrow)reached its peak (Figure 2, E). On the seventh day, ACNPhad moved downward to the lower segment of the gut lumenwith the intestinal peristalsis and were finally excretedthrough cloaca (yellow arrow, Figure 2, F). On the eighthday, while previously observed ACNP had been completelyexcreted out, ACNP continuously entered the intestinal tractfrom other tissues (Figure 2, G). These results suggested that

Figure 2. The dynamic changes of ACNP in gut lumen of zebrafish after ACNP microinjection in yolk sac. (A) At the fourth hour. (B) On the third day. (C) Onthe fourth day. (D) On the fifth day. (E) On the sixth day. (F) On the seventh day. (G) On the eighth day.
842 B. Zhao et al / Nanomedicine: Nanotechnology, Biology, and Medicine 10 (2014) 839–849
the yolk sac injected ACNP were effectively excreted throughintestinal tract.
Blood transportation of ACNP in zebrafish
The above results gave rise to the question of how ACNP gotto the intestinal tract. To answer this question, we first need toknow the transportation of ACNP in animal body. Theobservation of the behavior of ACNP in blood vesselsdemonstrated that yolk sac-injected ACNP was transportedthrough blood circulation. With the development of cardio-vascular system in zebrafish, ACNP were first found tomigrate from tissues to the wall of blood vessels. Figure 3,E demonstrated the case of ACNP in the wall of zebrafishtail veins. The yolk sac microinjected ACNP first migrated to
and then temporarily accumulated in the walls of bloodvessels, where they were finally released into the bloodstream. After injection, ACNP in the walls of blood vesselsgradually decreased with their release into the blood stream.These results indicated that the ACNP were transported bythe blood stream.
Elimination of ACNP from various tissues
To obtain the data supporting the excretion of ACNP fromvarious tissues through intestinal tract, we investigated therelations between the elimination and intestinal excretion ofACNP. Results demonstrated that the intestinal excretion ofACNP was accompanied by their elimination from varioustissues. ACNP in the tissues around the upper end of intestinal
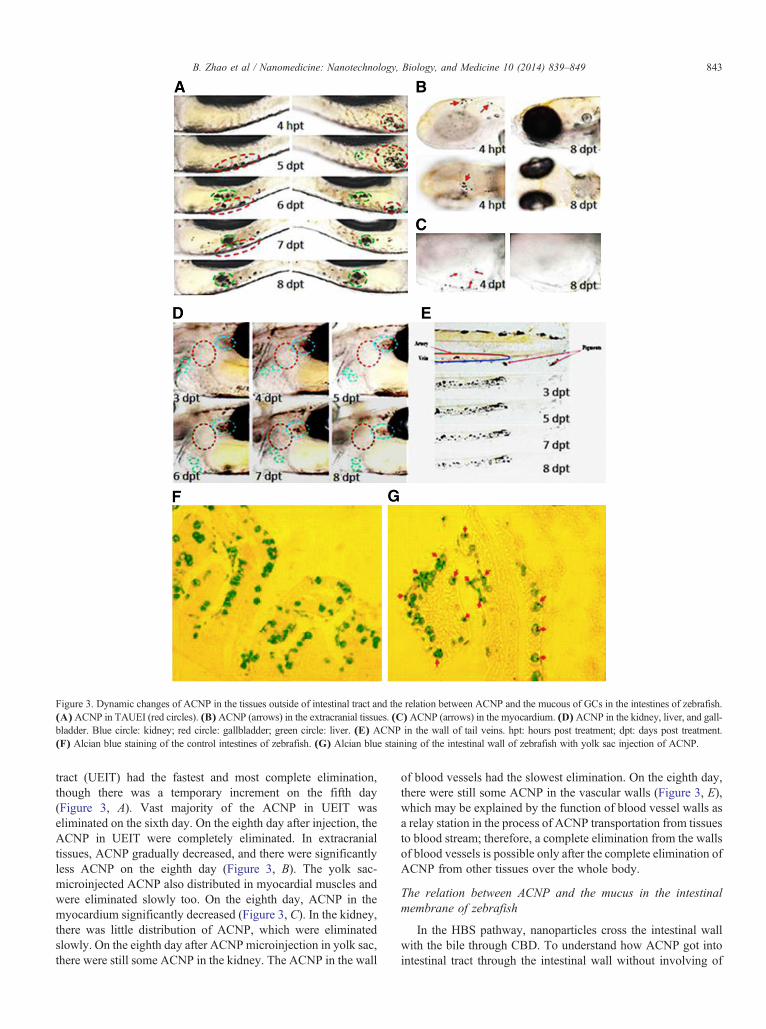
Figure 3. Dynamic changes of ACNP in the tissues outside of intestinal tract and the relation between ACNP and the mucous of GCs in the intestines of zebrafish.(A)ACNP in TAUEI (red circles). (B)ACNP (arrows) in the extracranial tissues. (C)ACNP (arrows) in the myocardium. (D)ACNP in the kidney, liver, and gall-bladder. Blue circle: kidney; red circle: gallbladder; green circle: liver. (E) ACNP in the wall of tail veins. hpt: hours post treatment; dpt: days post treatment.(F) Alcian blue staining of the control intestines of zebrafish. (G) Alcian blue staining of the intestinal wall of zebrafish with yolk sac injection of ACNP.
843B. Zhao et al / Nanomedicine: Nanotechnology, Biology, and Medicine 10 (2014) 839–849
tract (UEIT) had the fastest and most complete elimination,though there was a temporary increment on the fifth day(Figure 3, A). Vast majority of the ACNP in UEIT waseliminated on the sixth day. On the eighth day after injection, theACNP in UEIT were completely eliminated. In extracranialtissues, ACNP gradually decreased, and there were significantlyless ACNP on the eighth day (Figure 3, B). The yolk sac-microinjected ACNP also distributed in myocardial muscles andwere eliminated slowly too. On the eighth day, ACNP in themyocardium significantly decreased (Figure 3, C). In the kidney,there was little distribution of ACNP, which were eliminatedslowly. On the eighth day after ACNPmicroinjection in yolk sac,there were still some ACNP in the kidney. The ACNP in the wall
of blood vessels had the slowest elimination. On the eighth day,there were still some ACNP in the vascular walls (Figure 3, E),which may be explained by the function of blood vessel walls asa relay station in the process of ACNP transportation from tissuesto blood stream; therefore, a complete elimination from the wallsof blood vessels is possible only after the complete elimination ofACNP from other tissues over the whole body.
The relation between ACNP and the mucus in the intestinalmembrane of zebrafish
In the HBS pathway, nanoparticles cross the intestinal wallwith the bile through CBD. To understand how ACNP got intointestinal tract through the intestinal wall without involving of
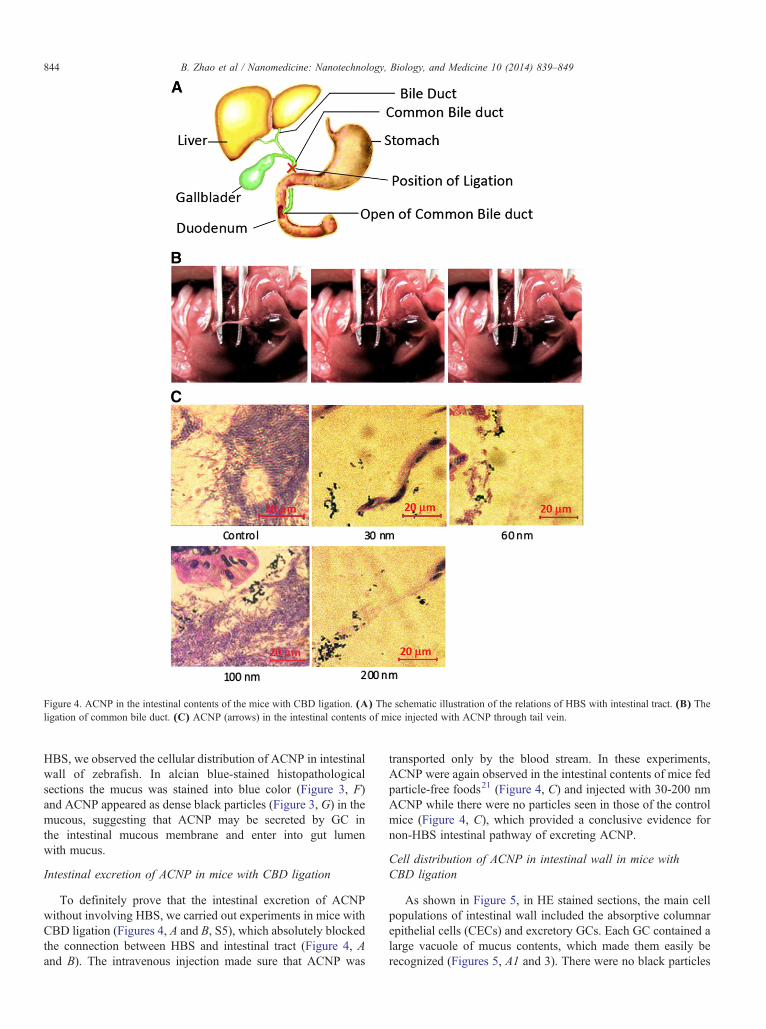
Figure 4. ACNP in the intestinal contents of the mice with CBD ligation. (A) The schematic illustration of the relations of HBS with intestinal tract. (B) Theligation of common bile duct. (C) ACNP (arrows) in the intestinal contents of mice injected with ACNP through tail vein.
844 B. Zhao et al / Nanomedicine: Nanotechnology, Biology, and Medicine 10 (2014) 839–849
HBS, we observed the cellular distribution of ACNP in intestinalwall of zebrafish. In alcian blue-stained histopathologicalsections the mucus was stained into blue color (Figure 3, F)and ACNP appeared as dense black particles (Figure 3, G) in themucous, suggesting that ACNP may be secreted by GC inthe intestinal mucous membrane and enter into gut lumenwith mucus.
Intestinal excretion of ACNP in mice with CBD ligation
To definitely prove that the intestinal excretion of ACNPwithout involving HBS, we carried out experiments in mice withCBD ligation (Figures 4, A and B, S5), which absolutely blockedthe connection between HBS and intestinal tract (Figure 4, Aand B). The intravenous injection made sure that ACNP was
transported only by the blood stream. In these experiments,ACNP were again observed in the intestinal contents of mice fedparticle-free foods21 (Figure 4, C) and injected with 30-200 nmACNP while there were no particles seen in those of the controlmice (Figure 4, C), which provided a conclusive evidence fornon-HBS intestinal pathway of excreting ACNP.
Cell distribution of ACNP in intestinal wall in mice withCBD ligation
As shown in Figure 5, in HE stained sections, the main cellpopulations of intestinal wall included the absorptive columnarepithelial cells (CECs) and excretory GCs. Each GC contained alarge vacuole of mucus contents, which made them easily berecognized (Figures 5, A1 and 3). There were no black particles
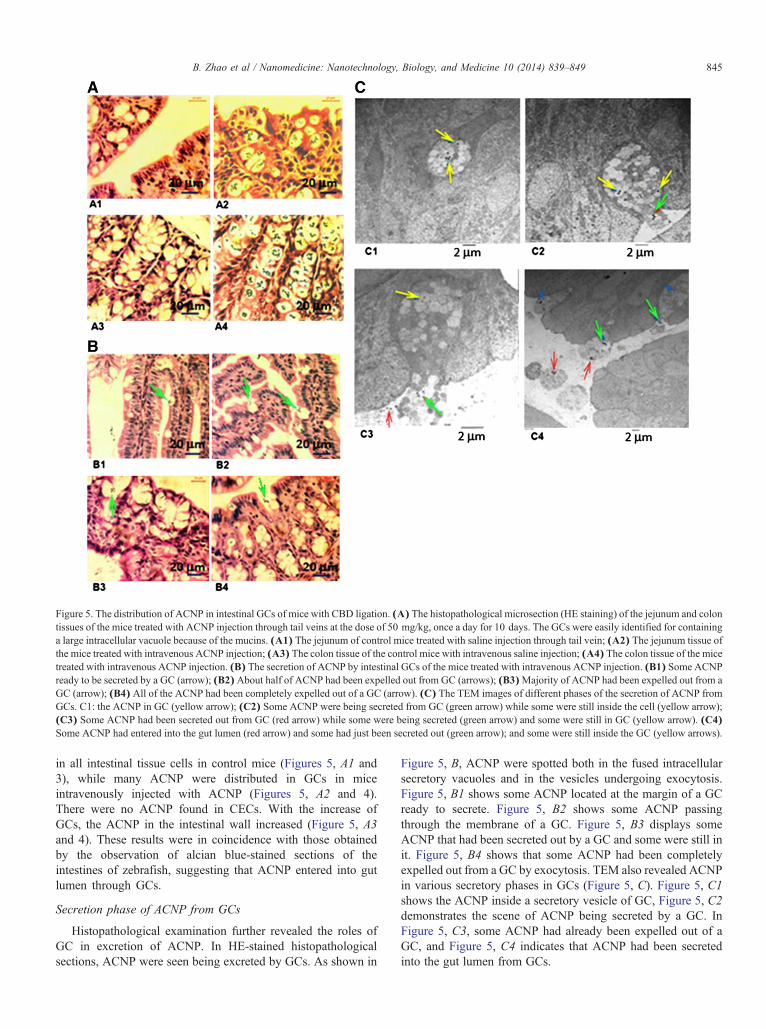
Figure 5. The distribution of ACNP in intestinal GCs of mice with CBD ligation. (A) The histopathological microsection (HE staining) of the jejunum and colontissues of the mice treated with ACNP injection through tail veins at the dose of 50 mg/kg, once a day for 10 days. The GCs were easily identified for containinga large intracellular vacuole because of the mucins. (A1) The jejunum of control mice treated with saline injection through tail vein; (A2) The jejunum tissue ofthe mice treated with intravenous ACNP injection; (A3) The colon tissue of the control mice with intravenous saline injection; (A4) The colon tissue of the micetreated with intravenous ACNP injection. (B) The secretion of ACNP by intestinal GCs of the mice treated with intravenous ACNP injection. (B1) Some ACNPready to be secreted by a GC (arrow); (B2)About half of ACNP had been expelled out from GC (arrows); (B3)Majority of ACNP had been expelled out from aGC (arrow); (B4) All of the ACNP had been completely expelled out of a GC (arrow). (C) The TEM images of different phases of the secretion of ACNP fromGCs. C1: the ACNP in GC (yellow arrow); (C2) Some ACNP were being secreted from GC (green arrow) while some were still inside the cell (yellow arrow);(C3) Some ACNP had been secreted out from GC (red arrow) while some were being secreted (green arrow) and some were still in GC (yellow arrow). (C4)Some ACNP had entered into the gut lumen (red arrow) and some had just been secreted out (green arrow); and some were still inside the GC (yellow arrows).
845B. Zhao et al / Nanomedicine: Nanotechnology, Biology, and Medicine 10 (2014) 839–849
in all intestinal tissue cells in control mice (Figures 5, A1 and3), while many ACNP were distributed in GCs in miceintravenously injected with ACNP (Figures 5, A2 and 4).There were no ACNP found in CECs. With the increase ofGCs, the ACNP in the intestinal wall increased (Figure 5, A3and 4). These results were in coincidence with those obtainedby the observation of alcian blue-stained sections of theintestines of zebrafish, suggesting that ACNP entered into gutlumen through GCs.
Secretion phase of ACNP from GCs
Histopathological examination further revealed the roles ofGC in excretion of ACNP. In HE-stained histopathologicalsections, ACNP were seen being excreted by GCs. As shown in
Figure 5, B, ACNP were spotted both in the fused intracellularsecretory vacuoles and in the vesicles undergoing exocytosis.Figure 5, B1 shows some ACNP located at the margin of a GCready to secrete. Figure 5, B2 shows some ACNP passingthrough the membrane of a GC. Figure 5, B3 displays someACNP that had been secreted out by a GC and some were still init. Figure 5, B4 shows that some ACNP had been completelyexpelled out from a GC by exocytosis. TEM also revealed ACNPin various secretory phases in GCs (Figure 5, C). Figure 5, C1shows the ACNP inside a secretory vesicle of GC, Figure 5, C2demonstrates the scene of ACNP being secreted by a GC. InFigure 5, C3, some ACNP had already been expelled out of aGC, and Figure 5, C4 indicates that ACNP had been secretedinto the gut lumen from GCs.
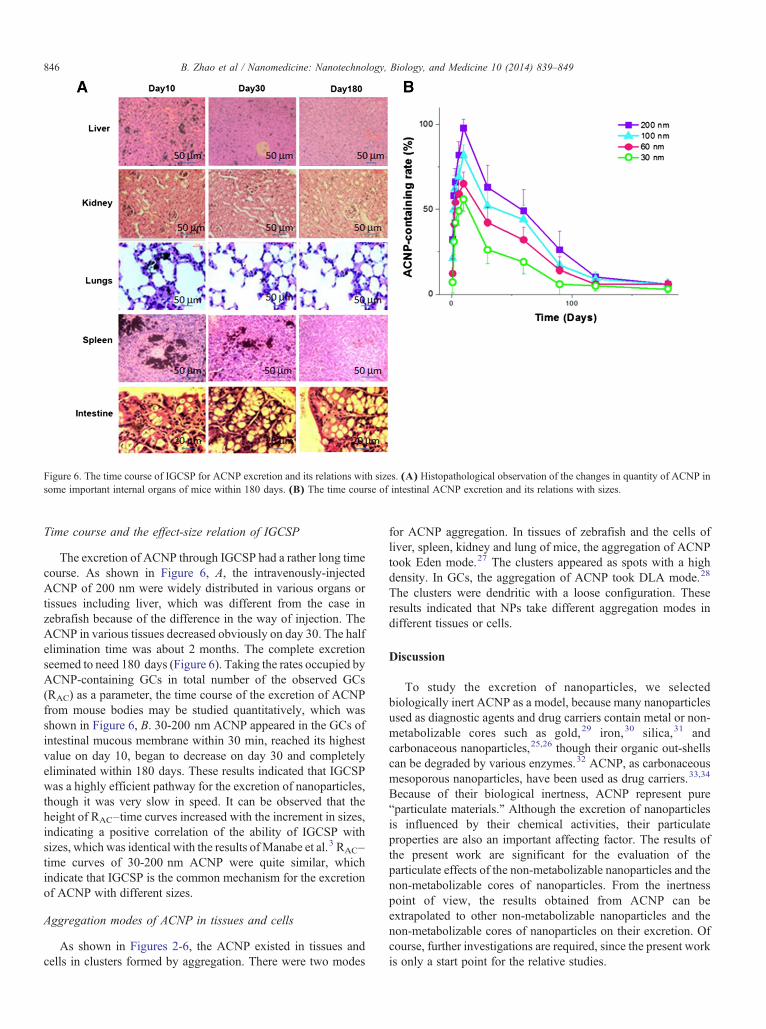
Figure 6. The time course of IGCSP for ACNP excretion and its relations with sizes. (A) Histopathological observation of the changes in quantity of ACNP insome important internal organs of mice within 180 days. (B) The time course of intestinal ACNP excretion and its relations with sizes.
846 B. Zhao et al / Nanomedicine: Nanotechnology, Biology, and Medicine 10 (2014) 839–849
Time course and the effect-size relation of IGCSP
The excretion of ACNP through IGCSP had a rather long timecourse. As shown in Figure 6, A, the intravenously-injectedACNP of 200 nm were widely distributed in various organs ortissues including liver, which was different from the case inzebrafish because of the difference in the way of injection. TheACNP in various tissues decreased obviously on day 30. The halfelimination time was about 2 months. The complete excretionseemed to need 180 days (Figure 6). Taking the rates occupied byACNP-containing GCs in total number of the observed GCs(RAC) as a parameter, the time course of the excretion of ACNPfrom mouse bodies may be studied quantitatively, which wasshown in Figure 6, B. 30-200 nm ACNP appeared in the GCs ofintestinal mucous membrane within 30 min, reached its highestvalue on day 10, began to decrease on day 30 and completelyeliminated within 180 days. These results indicated that IGCSPwas a highly efficient pathway for the excretion of nanoparticles,though it was very slow in speed. It can be observed that theheight of RAC–time curves increased with the increment in sizes,indicating a positive correlation of the ability of IGCSP withsizes, which was identical with the results ofManabe et al.3 RAC–time curves of 30-200 nm ACNP were quite similar, whichindicate that IGCSP is the common mechanism for the excretionof ACNP with different sizes.
Aggregation modes of ACNP in tissues and cells
As shown in Figures 2-6, the ACNP existed in tissues andcells in clusters formed by aggregation. There were two modes
for ACNP aggregation. In tissues of zebrafish and the cells ofliver, spleen, kidney and lung of mice, the aggregation of ACNPtook Eden mode.27 The clusters appeared as spots with a highdensity. In GCs, the aggregation of ACNP took DLA mode.28
The clusters were dendritic with a loose configuration. Theseresults indicated that NPs take different aggregation modes indifferent tissues or cells.
Discussion
To study the excretion of nanoparticles, we selectedbiologically inert ACNP as a model, because many nanoparticlesused as diagnostic agents and drug carriers contain metal or non-metabolizable cores such as gold,29 iron,30 silica,31 andcarbonaceous nanoparticles,25,26 though their organic out-shellscan be degraded by various enzymes.32 ACNP, as carbonaceousmesoporous nanoparticles, have been used as drug carriers.33,34
Because of their biological inertness, ACNP represent pure“particulate materials.” Although the excretion of nanoparticlesis influenced by their chemical activities, their particulateproperties are also an important affecting factor. The results ofthe present work are significant for the evaluation of theparticulate effects of the non-metabolizable nanoparticles and thenon-metabolizable cores of nanoparticles. From the inertnesspoint of view, the results obtained from ACNP can beextrapolated to other non-metabolizable nanoparticles and thenon-metabolizable cores of nanoparticles on their excretion. Ofcourse, further investigations are required, since the present workis only a start point for the relative studies.

847B. Zhao et al / Nanomedicine: Nanotechnology, Biology, and Medicine 10 (2014) 839–849
Aggregation is a common phenomenon for nanoparticles. Tofulfill the needs of injection and blood transportation, aggrega-tion must be prevented. Ultrasonication just before injection canmake ACNP disperse in suspension. After they were injectedinto yolk sac or veins, the concentration of ACNP was greatlydecreased by the dilutive effects of yolk sac fluid and blood,which prevent ACNP from aggregation. At the same time, themovement of animals and the flow of blood are also helpful forprevention of the aggregation. However, the case in tissues andcells was completely different from that in blood. Theconcentration of ACNP in tissues would be much higher thanthat in blood because of EPR effects, and the flow of tissue fluidwas much slower than that of blood. The case in cells that canphagocytose ACNP was the same as that in tissues: high ACNPconcentration and low flow rate, which explain the aggregationof ACNP in tissues and cells.
There were two aggregation modes for ACNP in tissues andcells: the Eden mode27 and the DLA mode,28 which can be moreclearly seen in hitopathological sections of mice, because themagnification of the direct observation of ACNP clusters inliving zebrafish was too low to recognize the details. In tissuespace and cells of liver, spleen, lungs and kidney of mice, theaggregation took Eden mode, in which the clusters were spotswith high density.27 In intestinal GCs, the aggregation took DLAmode, in which the clusters were dendritic with a looseconfiguration.28 These results suggested that the aggregationof nanoparticles is a rather complex process and the growth ofclusters can take different modes in different microenvironments.GCs had unique mucin contents, and, therefore, ACNPaggregation in them took unique DLA mode.
Whether biological systems can remove or excrete clusters ofnanoparticles is another question requiring answers for practicaluse of nanoparticles. The results of this study indicate that GCshave the capability to secrete clusters of nanoparticles. As for theclusters in tissues and cells, the removing process may be morecomplex. They must disaggregate into dispersed nanoparticles orclusters small enough for blood transportation, which istheoretically possible. The concentration of ACNP in the blooddecreased gradually with time after the injection. When bloodconcentration was low enough, the ACNP in tissues began toenter into blood, and hence the concentration of ACNP in tissuesbegan to decrease. When tissue concentration was low enough,some of ACNP at the edge of cluster tended to separate; leadingto the decrease in size and finally disaggregation of the cluster. Inspite of the difficulty, this process must happen, sooner or later,faster or slower. As discussed above, the aggregation onlyinfluences the speed but cannot prevent the excretion ofnanoparticles.
To understand how the ACNP cross the intestinal wall andare released into the gut lumen, HBS was firstly consideredbecause it is a well-known excretion system. There have beenmany literatures that believe nanoparticles can be excretedthrough HBS based on the results that nanoparticles can besimultaneously seen in liver and intestinal tract. In the late1990s, Renaud et al found that the low-density lipoprotein-20 nm colloidal gold (CG) could be excreted through HBS butalbumin- or polyvinylpyrrolidone-CG could not.35 Thisselective excretion was rationally attributed to the function of
the lipoprotein receptor of liver cells. In other words, thenanoparticles without low-density lipoproteins could not beexcreted through HBS. Later studies found that negativelycharged gold nanoparticles9 and polystyrene carboxylatednanoparticles36 could also be excreted through HBS, althoughit was limited to no larger than 20 nm and the excretionquantity was very low. Souris et al11 reported that 80 nmnanoparticles can be excreted through intestinal tract and theconcentration in feces was very high while the concentration inliver was very low. In comparison with liver, the nanoparticleconcentration was much higher in intestinal wall. It seemseasier to explain the intestinal excretion of nanoparticles byintestinal secretion than by HBS pathway. More recently, inone study by Manabe et al, it was found that that nanoparticlesas large as 500 nm, far beyond the ability of HBS, can beeliminated from fish body and the elimination of 500 nmparticles was faster and more efficient than 50 nm particles nomatter whether modified or not,10 which cannot be explainedby HBS pathway. Obviously, there were paradoxes between theresults of the experiments on HBS for nanoparticle excretion.These paradoxes are unexplainable based on the belief thatHBS is the only pathway for excretion of nanoparticles throughintestinal tract. Especially, the results of Manabe et al stronglysuggest the existence of another pathway for intestinalexcretion of nanoparticles without involving HBS, and theresults of Souris et al suggest that intestinal secretion is apossible pathway for nanoparticle excretion, since the concen-tration of nanoparticles was more identical with that in fecesthan that in liver.11
Based on previous reports, we also believed that HBS is themost possible pathway for nanoparticle intestinal excretion.However, surprisingly, there was only a little distribution ofACNP in the liver and no ACNP in gallbladder during the wholeperiod of experiments in zebrafish. These observations do notsupport HBS pathway because gallbladder is the only organ tostore bile and CBD is the only way for excretion of ACNP fromliver to intestinal tract. Thus, we believe that there must be analternative mechanism for ACNP to enter the intestinal tract,which does not involve HBS. To confirm the existence of thispathway, we carried out the experiments in mice with ligation ofCBD in mice, which completely cut off the way that ACNP mustpass through to get to gut lumen from liver. As it was anticipated,ACNP were still observed in the feces of these mice with CBDligation. The histopathological and TEM examination revealedthat the mechanism of such non-HBS pathway for nanoparticleexcretion was through the secretion of intestinal GCs. Thedistribution of ACNP only in GCs suggested IGCSP, which wasthen proved by the secretion phase observed in histopathologicalsections and TEM ultrasections.
IGCSP had a long time course, which is quite in favor ofthat nanoparticles are used as drug carriers for long lastingtreatments of chronic diseases.37–39 As demonstrated inzebrafish and mice, most intravenously-injected ACNP canbe excreted through IGCSP, which greatly ameliorates ourconcern about the eternal existence of biologically inertnanoparticles in animal body.
GC is a main cell type of the intestinal mucosalepithelium, which undergoes continuous cycles of renewal.40

848 B. Zhao et al / Nanomedicine: Nanotechnology, Biology, and Medicine 10 (2014) 839–849
The major function of intestinal GCs and their mainsecretory product, mucin, is the formation of mucusoverlying layers on intestinal membranes, which promotesthe elimination of gut contents and provides the first line ofdefense against physical and chemical injury caused byingested food, microbes and the microbial products.13,41
There have been no reports on the roles of GC in theintestinal excretion of nanoparticles, let alone the mechanismfor ACNP to enter into GC. In our experiments withzebrafish, the yolk sac-injected ACNP were distributed invarious tissues with the formation of organs and then wastransported by blood flow to intestines and excreted throughintestinal tract, which suggested that the primary tissuedistribution was related to the development of tissues andACNP may enter GC during the differentiation. Because GCcontinuously undergo cycles of renewal, it is considered thatthe mechanism for ACNP getting into GC may be related tothe differentiation of GC. The ACNP in GC combined ormixed with mucus and then were excreted into gut lumen byGC along with the secretion of mucus. There are alsostudies on the interaction between intra-tracheal nanoparticlesand GCs in the airway42,43; however, we didn't examine thedistribution of ACNP in airway GC because the airway isnot a main excretion organ for solid substance. So, IGCSPwas the main pathway for the excretion of nanoparticles.
In the present work, zebrafish and mice were used as animalmodels. Zebrafish has been used for drug screening.19,44,45 Micehave gene sequences 80% identical with human and arefrequently used as animal models in basic life science ofhuman and preclinical studies on therapeutic agents andmethods.46 Both of them are vertebrate animals with similarintestinal tissues composed of mainly columnar and gobletcells. Therefore, the results of the present study are of greatsignificance for evaluating the biosafety of nanoparticlesin human.
To date, IGCSP for nanoparticles has been found notonly in marine animals but also in mammals. This pathwayis especially important for those larger nanoparticles withoutmodification, because the kidney7 and HBS are only ableto excrete nanoparticles modified with various chemicalstructures in small sizes.8,11 Considering that manynanoparticles used as diagnostic and therapeutic agentshave metal cores and are relatively large,47–49 in the sizesthat are beyond the excretory capability of kidney andHBS, this novel IGCSP provided greater promise for futureapplication of nanoparticles in biomedical fields byeliminating the concerns about the persistent effects ofnanoparticles. Obviously, the present results are only astarting point for the studies on IGCSP. The details, suchas how ACNP cross the cell membrane and get into thecells and how they are secreted by the intestinal GCs,warrant further investigations.
Appendix A. Supplementary data
Supplementary data to this article can be found online athttp://dx.doi.org/10.1016/j.nano.2013.10.004.
References
1. Dreaden EC, Austin LA, Mackey MA, El-Sayed MA. Size matters: goldnanoparticles in targeted cancer drug delivery.Ther Deliv 2012;3(4):457-78.
2. Wahajuddin, Arora S. Superparamagnetic iron oxide nanoparticles:magneticnanoplatforms as drug carriers. Int J Nanomedicine 2012;7:3445-71.
3. Zarogoulidis P, Chatzaki E, Porpodis K, Domvri K, Hohenforst-SchmidtW, Goldberg EP, Karamanos N, Zarogoulidis K. Inhaled chemotherapyin lung cancer: future concept of nanomedicine. Int J Nanomedicine2012;7:1551-72.
4. Elder A, Gelein R, Finkelstein JN, Driscoll KE, Harkema J, OberdorsterG. Effects of subchronically inhaled carbon black in three species. I.Retention kinetics, lung inflammation, and histopathology. Toxicol Sci2005;88:614-29.
5. Feng Y, Zong Y, Ke T, Jeong EK, Parker DL, Lu ZR. Pharmacokinetics,biodistribution and contrast enhanced MR blood pool imaging of Gd-DTPA cystine copolymers and Gd-DTPA cystine diethyl ester co-polymers in a rat model. Pharm Res Aug. 2006;23:1736-42.
6. Hagens WI, Oomen AG, de Jong WH, Cassee FR, Sips AJ. What do we(need to) know about the kinetic properties of nanoparticles in the body?Regul Toxicol Pharmacol 2007;49:217-29.
7. Choi HS, Liu W, Misra P, Tanaka E, Zimmer JP, Ipe BI, Bawendi MG,Frangioni JV. Renal clearance of quantum dots. Nat Biotecnol2007;25:1165-70.
8. Singh R, Pantarotto D, Lacerda L, Pastorin G, Klumpp C, Prato M,Bianco A, Kostarelos K. Tissue biodistribution and blood clearance ratesof intravenously administered carbon nanotube radiotracers. Proc NatlAcad Sci U S A 2006;103(9):3357-62.
9. Hirn S, Semmler-Behnke M, Schleh C, Wenk A, Lipka J, Schäffler M,Takenaka S, Möller W, Schmid G, Simon U, KreylingWG. Particle size-dependent and surface charge dependent biodistribution of goldnanoparticles after intravenous administration. Eur J Pharm andBiopharm 2011;77:407-16.
10. Manabe M, Tatarazako N, Kinoshita M. Uptake, excretion and toxicityof nano-sized latex particles on medaka (Oryzias latipes) embryos andlarvae. Aquat Toxicol 2011:105576-81.
11. Souris JS, Lee CH, Cheng SH, Chen CT, Yang CS, Ho JA, Mou CY, LoLW. Surface charge-mediated rapid hepatobiliary excretion of mesopor-ous silica nanoparticles. Biomaterials 2010;31:5564-74.
12. Kim YS, Ho SB. Intestinal goblet cells and mucins in health and disease:recent insights and progress. Curr Gastroenterol Rep 2010;12:319-30.
13. Behrens I, Pena AIV, Alonso MJ, Kissel T. Comparative uptake studiesof bioadhesive and non-bioadhesive nanoparticles in human intestinalcell lines and rats: the effect of mucus on particle adsorption andtransport. Pharm Res 2002;19:1185-93.
14. Jin Y, Song YP, Zhu X, Zhou D, Chen CH, Zhang ZR, Huang Y. Gobletcell-targeting nanoparticles for oral insulin delivery and the influence ofmucus on insulin transport. Biomaterials 2012;33:1573-82.
15. des Rieux A, Fievez V, Garinot M, Schneider YJ, Préat V. Nanoparticlesas potential oral delivery systems of proteins and vaccines: a mechanisticapproach. J Control Releas 2006;116:1-27.
16. Dong YC, Feng SS. Poly(d, l-lactide-co-glycolide)/montmorillonitenanoparticles for oral delivery of anticancer drugs. Biomaterials2005;26:6068-76.
17. Wang YY, Lai SK, Suk JS, Pace A, Cone R, Hanes J. Addressing thePEG mucoadhesivity paradox to engineer nanoparticles that “slip”through the human mucus barrier. Angew Chem 2008;47:9575-781.
18. Sun L, Yang LZ, Zhang YG. Study on the excretion pathway of nano-particles. Chin Pharm J 2006;41:366-7.
19. Wang K, Ma JB, He M, Gao G, Xu H, Sang J, Wang YX, Zhao BQ, CuiDX. Toxicity assessments of near-infrared upconversion luminescentLaF3:Yb, Er in early development of zebrafish embryos. Theranostics2013;3:258-66.
20. Westerfield M. The zebrafish book: a guide for the laboratory use ofzebrafish (Brachydanio rerio). Eugene, OR: University of Oregon Press;1993.

849B. Zhao et al / Nanomedicine: Nanotechnology, Biology, and Medicine 10 (2014) 839–849
21. Yang Z, Zhang Y, Yang Y, Sun L, Han D, Li H, Wang C.Pharmacological and toxicological target organelles and safe use ofsingle-walled carbon nanotubes as drug carriers in treating Alzheimerdisease. Nanomedicine:NBM 2010;6:427-41.
22. Karlsson J, von Hofsten J, Olsson P-E. Generating transparent zebrafish:a refined method to improve detection of gene expression duringembryonic development. Mar Biotechnol 2001;3:522-7.
23. Cameron GR, Oakley CL. Ligation of the common bile duct. J PatholBacteriol 1932;35:769-98.
24. Geim AK, Novoselov KS. The rise of graphene. Nat Mat 2007;6:183-91.25. Buseck PR, Tsipursky SJ, Hettich R. Fullerenes from the geological
environment. Science 1992;257:215-7.26. Allen MJ, Tung VC, Kaner RB. Honeycomb carbon: a review of
graphene. Chem Rev 2010;110:132-45.27. Herrmann HJ. Geometrical cluster growth models and kinetic gelation.
Phys Rep 1986;136:517-30.28. Meakin P, Tolman S. Diffusion-limited aggregation. Proc R Soc Lond A
1989;423:620-5.29. Sadauskas E, Danscher G, Stoltenberg M, Vogel U, Larsen A, Hide HW.
Protracted elimination of gold nanoparticles from mouse liver. Nano-medicine: NBM 2009;5:162-9.
30. Kumar A, Jena PK, Behera S, Lockey RF, Mohapatra, Hide SM.Multifunctional magnetic nanoparticles for targeted delivery. Nanome-dicine: NBM 2010;6:64-9.
31. Liu S, Chen G, Ohulchanskyy TY, Swihart MT, Prasad PN. Facilesynthesis and potential bioimaging applications of hybrid upconvertingand plasmonic NaGdF4: Yb(3+), Er(3+)/silica/gold nanoparticles.Theranostics 2013;3:275-81.
32. Stanishevsky AV, Styres C, Yockell-Lelievre H, Yusuf N. Nanostruc-tured carbon beads—properties and biomedical applications. J NanosciNanotechnol 2011;11:8705-11.
33. Yang Z, Ma S, Zhang Y. Using activated carbon nanoparticles todecrease the genotoxicity and teratogenicity of anticancer therapeuticagents. J Nanosci Nanotechnol 2010;10:8603-9.
34. Hu B, Zhao Y, Zhu HZ, Yu SH. Selective chromogenic detection ofthiol-containing biomolecules using carbonaceous nanospheres loadedwith silver nanoparticles as carrier. ACS Nano 2011;5:3166-71.
35. Renaud G, Hamilton RL, Havel RJ. Hepatic metabolism of colloidalgold-low density lipoprotein complexes in the rat: evidence forbulk excretion of lysosomal contents into bile. Hepatology 1989;9:380-92.
36. Johnston HJ, Semmler-Behnke M, Brown DM, Kreyling W, Tran L,Stone V. Evaluating the uptake and intracellular fate of polystyrene
nanoparticles by primary and hepatocyte cell lines in vitro. Toxicol ApplPharmacol 2010;242:66-78.
37. Dong J, Zhou G, Tang D, Chen Y, Cui B, Dai X, Zhang J, Lan Q, HuangQ. Local delivery of slow-releasing temozolomide microspheres inhibitsintracranial xenograft glioma growth. J Cancer Res Clin Oncol2012;138:2079-84.
38. Ratanajiajaroen P, Ohshima M. Synthesis, release ability and bioactivityevaluation of chitin beads incorporated with curcumin for drug deliveryapplications. J Microencapsul 2012;29:549-58.
39. Lee MJ, Jin SE, Kim CK, Choung HK, Kim HJ, Hwang JM. Effect ofslow-releasing all-trans-retinoic acid in bioabsorbable polymer ondelayed adjustable strabismus surgery in a rabbit model. Am JOphthalmol 2009;148:566-72.
40. Lievin-LeMoal V, ServinAL. The front line of enteric host defense againstunwelcome intrusion of harmful microorganisms: mucins, antimicrobialpeptides, and microbiota. Clin Microbiol Rev 2006;19:315-37.
41. Dharmani P, Srivastava V, Kissoon-Singh V, et al. Role of intestinalmucins in innate host defense mechanisms against pathogens. J InnateImmun 2009;1:123-35.
42. Tada Y, Yano N, Takahashi H, Yuzawa K, Ando H, Kubo Y, NagasawaA, Ogata A, Nakae D. Acute phase pulmonary responses to a singleintratracheal spray instillation of magnetite (fe(3)o(4)) nanoparticles inFischer 344 rats. J Toxicol Pathol 2012;25:233-9.
43. Jang S, Park JW, Cha HR, Jung SY, Lee JE, Jung SS, Kim JO, Kim SY,Lee CS, Park HS. Silver nanoparticles modify VEGF signaling pathwayand mucus hypersecretion in allergic airway inflammation. Int JNanomedicine 2012;7:1329-43.
44. Vlasits AL, Simon JA, Raible DW, Rubel EW, Owens KN. Screen ofFDA-approved drug library reveals compounds that protect hair cellsfrom aminoglycosides and cisplatin. Hear Res 2012;294:153-65.
45. Rovira M, Huang W, Yusuff S, Shim JS, Ferrante AA, Liu JO, ParsonsMJ. Chemical screen identifies FDA-approved drugs and targetpathways that induce precocious pancreatic endocrine differentiation.PNAS 2011;108:19264-9.
46. Pennisi E. Sequence tells mouse, human genome secrets. Science2002;298:1863-5.
47. Alexis F, Pridgen E, Molnar LK, Farokhzad OC. Factors affecting theclearance and biodistribution of polymeric nanoparticles. Mol Pharm2008;5:505-15.
48. Almeida JPM, Chen AL, Foster A, Drezek R. In vivo biodistribution ofnanoparticles. Nanomedicine:NBM 2011;6:815-35.
49. Cho K, Wang X, Nie S, Chen Z, Shin DM. Therapeutic nanoparticles fordrug delivery in cancer. Clin Cancer Res 2008;14:1310-6.