Embed Size (px)
Citation preview

Secondary and Interactive Effects of Chronic Gaseous Pollutant Exposure of Producers, Consumers, and Decomposers
Influence of Chronic Air Pollution on Mineral Cycling in Forests1
Paul J. Zinke 2
Abstract: This paper reviews the literature concerning the impact of'chronic air pollution on mineral element cycling in forests. The concept involves the forest trees taking up essential and other elements from the soil and surrounding en-vironment eventually to return them to the soil upon mortality and decay. Chronic pollutants are considered in the context of this cycling as another form of elemental addition to the site subject to cycling in the same manner. The review is organized to assess the addition of the major elements, carbon, nitrogen, and sulfur, and the trace metallic elements. In addition, pollutant effects upon the ecosystem living components in terms of producers and decompos- ers are considered along with potential changes in the redox and pH state of the different portions of the forest. Some oriuinal data on foliar comnosition of Big Cone Spruce (Pseudotsuga macrocarpa (vasey) Mayr) in relation to pollution exposure, and the evaluation of a soil subject to increments o$ hydrogen in simu- lated acid rain leaching are presented.
I N T R O D U C T I O N The f o r e s t w i l l be cons idered t o b e t h e s tand of t r e e s and t h a t p o r t i o n of
This paper w i l l p r e sen t a review of t h e atmosphere encompassed by canopy and va r ious a s p e c t s of minera l element t runk space , and t h e p o r t i o n of t h e s o i l c y c l i n g i n f o r e s t s as a f f e c t e d by chronic encompassed by t h e r o o t space. Mineral a tmospheric p o l l u t i o n . c y c l i n g i s t h e process of c y c l i n g of e l e -
ments from t h e s o i l through uptake by r o o t s o r by f o l i a g e , t r a n s p o r t w i th in t h e t r e e s and t h e even tua l r e t u r n t o t h e f o r e s t s o i l i n t h e p roces ses of f o l i a r l each ing , r o o t exudat ion , f o l i a g e drop ,
Presented at the Symposium on Effects e t c . Return t o t h e s o i l s t o r a g e occu r s , of Air Pollutants on Mediterranean and and t h e c y c l e is cont inued by uptake Temperate Forest Ecosystems, June 22-27, aga in , o r i t may be broken by l o s s from 1980, Riverside, California, U.S.A. t h e s o i l , o r t i e u p as i n s o l u b l e p rec ip i -
t a t e s o r compounds. A i r p o l l u t i o n expo-s u r e w i l l be cons idered t o be t h e a d d i -Associate Professor of Forestry, Dep. t i o n a l a tmospheric i n p u t s which e n t e r t h eof Forestry and Resource Manage., Univer- n u t r i t i o n a l and e lementa l cyc l e ofsity of California, Berkeley, Calif. f o r e s t s due t o man made e f f e c t s on atmos-phe r i c composition.
L

ELEMENTAL CYCLIdG
The i d e a o f t h e c y c l i n g of e lements from v e g e t a t i o n t o s o i l , f o l lowing uptake from s o i l probably o u t d a t e s w r i t t e n r e c o r d s as is appa ren t from t h e p r i m i t i v e a g r i c l t u r a l p r a c t i c e of s h i f t i n g c u l t i v a - t i o n wherein a f o r e s t i s cu t and burned f o r s o i l enrichment by t h e ash. This i s fol lowed by t h e regrowth of a f a l l o w f o r e s t c rop , which i n t u r n is burned aga in . The f o r e s t vege ta t ion c y c l e s e l e - ments e s s e n t i a l f o r v e g e t a t i v e growth as w e l l as unncessary elements a c c i d e n t a l t o t h e p a r t i c u l a r environment. The 16 o r 17 e s s e n t i a l e lements ( c , d, 0, P, K , N, S, C a , Fe, Mg, Mn, Cu, Zn, Mo, B, C l , and perhaps Se) , a r e cyc led by n e c e s s i t y f o r without them t h e r e would be no f o r e s t . I n a d d i t i o n almost every o t h e r element is t r a n s l o c a t e d by p l a n t s . This concept i s used i n geochemical p rospec t ing f o r many of t h e more va luab le elements , u s i n g t h e p l a n t as a chemical prospec tor (NASA 1968). These elements which a r e cycled may be enr iched i n concen t r a t ion i n v a r i - ous p o r t i o n s of t h e c y c l e , as i n t h e f o l i a g e , t h e p l a n t d e t r i t u s on t h e ground, o r i n t h e s u r f a c e of t h e s o i l beneath t h e p l a n t s . Such enrichment i s o f t e n t h e i n d i c a t i o n of an extraneous element . S i m i l a r l y t h e inpu t of ex t rane-ous e lements i n t h e process of p o l l u t i o n may be evidenced by enrichment over t h e normal background amounts.
The s t a t e o f t h e v e g e t a t i o n and its c a p a c i t y f o r s t o r a g e of e lements w i l l determine t h e r e l a t i v e change induced by t h e a d d i t i o n of a p o l l u t a n t element. The s t a t e of t h e v e g e t a t i o n is p a r t l y de t e r -mined by t h e f a c t t h a t v e g e t a t i o n is a g i a n t r educ t ion r e a c t i o n i n which pho-t o s y n t h e s i s reduces carbon from carbon d iox ide t o carbon i n reduced carbon com-
ounds as desc r ibed by Stumm and -Morgan 71 970) . The f o r e s t t r e e s c a r r y out t h i s r e d u c t i o n r e a c t i o n , fol lowed by t h e i r dea th and t h e subsequent o x i d a t i o n of t h e carbon compounds i n t h e s o i l . The c y c l i n g of t h e elements on t h e s i t e occurs i n t h e c o n t e x t of t h i s g i a n t redox r e a c t i o n ( ~ a r r e l s , e t a l . , 1975) , and i ts s t a t e and magnitude determine t h e r a t e and amount a t which t h e o t h e r e lements w i l l be cyc l ed .
The c y c l i n g of most of t h e elements which undergo redox changes i n t h e f o r e s t n u t r i e n t c y c l e (C H 0 N S Mn ~ e )w i l l tend t o be reduced i n t h e l i v i n g vegeta ted p o r t i o n o-f t h e c y c l e and com-p l e t e t h e c y c l e t o an oxid ized form i n t h e s o i l . However, l o c a l s o i l f a c t o r s f avo r ing anaerobic c o n d i t i o n s such as water i n exces s , may cause t h e s o i l t o become a s i n k f o r some elements i n t h e reduced form, o r f o r o the r e lements i n t h e oxid ized form.
Thus p o l l u t a n t s may e n t e r t h e e l e -mental cyc l e s i n f o r e s t s and vary i n mob i l i t y depending upon t h e redox poten-t i a l at va r ious p o r t i o n s of t h e e lementa l cyc le . Some p o l l u t a n t s may a l s o b e oxi-dants i n r e l a t i o n t o t h e p o r t i o n of t h e ecosystem t h a t i s absorb ing them and thus a f f e c t t h e redox p o t e n t i a l a t t h a t po in t accord ing t o Haagen-Smit (1958) .
STORAGE POINTS AND SINKS
The soil and vegetation of the forest have several points of long term s t o r a g e of m a t e r i a l s added t o t h e elemen-ta l c y c l i n g system. It has been l ea rned from pas t f e r t i l i z a t i o n exper ience t h a t e lements added t o t h e f o r e s t , e i t h e r t o s o i l o r d i r e c t t o t r e e s , may be s t o r e d f o r vary ing pe r iods of t ime i n d i f f e r e n t p o r t i o n s of t h e f o r e s t . Obviously, t h e t r e e t runks provide s t o r a g e of t h e m a t e r i a l s conta ined i n t h e wood f o r t h e l e n g t h of l i f e of t h e t r e e and i t s subse-q.uent decomposition t ime. Deciduous por- t i o n s of t h e t r e e r e t a i n m a t e r i a l s f o r l e n g t h s of t ime p ropor t iona te t o t h e i r r e s idence t i m e on t h e t r e e ; bark f o r longer pe r iods , l e a v e s and twigs f o r l e s s e r pe r iods . The s t o r a g e t ime i n t h e decomposing o r o x i d i z i n g p o r t i o n s of t h e d e t r i t u s dropped t o t h e s o i l depends upon l o c a l environmental c o n d i t i o n s of tem-p e r a t u r e , redox s t a t e , and a v a i l a b i l i t y o r t o x i c i t y of e lements t o decomposing ( ox id i z ing ) organisms. For example, decomposition t imes f o r s u r f a c e d e t r i t u s on t h e s o i l s u r f a c e i n a f o r e s t may va ry from a f r a c t i o n of a year t o many yea r s .
The s o i l beneath t h e f o r e s t i s a g i a n t fixed-bed ion exchanger, and once elements e n t e r t h e s o i l fo l lowing r e l e a s e from s t o r a g e i n t h e v e g e t a t i v e p o r t i o n of t h e f o r e s t and i t s d e t r i t u s t h e y may be s t o r e d on t h i s exchange complex. The c a p a c i t y of t h i s i o n exchange bed as we l l as t h e n a t u r e of a s s o c i a t e d c a t i o n s w i l l determine t h e s t o r a g e p r o b a b i l i t y of e l e -ments added t o t h e f o r e s t whether as a d d i t i v e s ( p o l l u t a n t s , f e r t i l i z e r s ) , o r i n t h e normal course of mineral weather-i n g o r r a i n f a l l a d d i t i o n s . The t o t a l capac i ty of t h i s bed f o r c a t i o n exchange i n con i f e rous f o r e s t s o i l s ranges from 40 t o 300 gm e q u i v a l e n t s per meter squared t o a meter depth (Zinke , & g. , 1979). Whether t h e p o l l u t a n t added has a s i g n i -f i c a n t e f f e c t depends upon t h e n a t u r a l base l i n e composition of o t h e r c t i o n s o n t h e s o i l column ( u s u a l l y H , Ca¥f, Mg , K', N a ) maintained. by t h e f o r e s t and t h e l y o t r o p i c s e r i e s ( l e a c h i n g pre-cedence) of t h e s e elements . The f o r e s t s o i l a l s o has an ion exchange c a p a c i t y a l though u s u a l l y of l e s s e r amount. Thus, depending upon t h e n a t u r e of t h e pol lu-

t a n t a d d i t i v e , and whether i t a t t a i n s i o n i c form, it may be s u b j e c t t o scrub-b ing out a t t h e s o i l exchange complex s t a g e of t h e e lementa l cyc l e . This would apply t o t h e hydrogen and s u l f a t e of a c i d r a i n , o r t h e c a t i o n i c forms of some m e t a l l i c p o l l u t a n t s such as l ead o r z inc . Laboratory s imu la t ion through l each ing wi th succes s ive increments of t h e pol lu-t a n t such as a c i d r a i n can be made r e a d i l y t o determine t h i s c a p a c i t y , and an example o f t h i s fo l lows l a t e r .
The s o i l has o t h e r c a p a c i t i e s f o r s t o r a g e of a d d i t i v e s i n a d d i t i o n t o t h a t of t h e ion exchange c a p a c i t i e s . E i t h e r fo l lowing breakthrough o r s a t u r a t i o n of t h e exchange c a p a c i t y , an a d d i t i v e e l e -ment may be s t o r e d on t h e m e t a l l i c oxide complex of t h e s o i l , o r as i n s o l u b l e pre-c i p i t a t e s o r ox ides . The r e l e a s e and uptake of e lements i n t o s t o r a g e i n asso-c i a t i o n wi th t h e m e t a l l i c oxides ( u s u a l l y i r o n and manganese) w i l l depend upon t h e pH and redox p o t e n t i a l of t h e s o i l as reviewed by Jenne ( 1 968) . Reten t ion w i l l be l e a s t under t h e reducing cond i t i ons brought about by l a r g e amounts of organic mat te r and poor ly dra ined - wet condi-t i o n s .
Thus, t h e f o r e s t s o i l i s a major de te rminant i n t h e f a t e o f t h e chronic p o l l u t a n t element added t o t h e f o r e s t . The break through c a p a c i t y of t h e s o i l f o r t h e added m a t e r i a l w i l l depend upon t h e c u r r e n t i npu t of s i m i l a r e lements by t h e n a t u r a l c y c l i n g of c a t i o n s from t h e t r e e cove r , t h e i n p u t s by c u r r e n t minera l weather ing , t h e c u r r e n t a d d i t i o n s of hydrogen from r a i n f a l l and o rgan ic com-pounds produced by t h e f o r e s t , t h e deple-t i o n of b a s i c m e t a l l i c c a t i o n s taken up by t h e t r e e growth as t h e t r e e s age on t h e s i t e . The g r e a t e r t h e s t o r a g e capa- c i t y of t h e s o i l e i t h e r as i o n exchange o r m e t a l l i c ox ide (hydrous) c a p a c i t y t h e g r e a t e r t h e b u f f e r i n g c a p a c i t y of t h e f o r e s t on t h e inpu t of p o l l u t a n t s .
It w i l l be of i n t e r e s t now t o review some of t h e expe r i ence of how t h e va r ious p o l l u t a n t s behave i n r e l a t i o n t o t hese p roces ses of e lementa l c y c l e s i n a f o r e s t .
POLLUTANT ADDITIVES T O FOREST CYCLES
Any of t h e elements added t o t h e f o r e s t as p o l l u t a n t s can i n f l u e n c e e le -mental c y c l i n g , e i t h e r by a c t i n g as n u t r i e n t e lements r e q u i r e d f o r t h e growth o f t r e e s , by changing t h e redox s t a t e o f any p a r t of t h e f o r e s t , o r by changing t h e pH a t some p o i n t i n t h e system. Some o f t h i s i n t e r a c t i o n may be phys io log ica l i n t h e v e g e t a t i o n , o r i n t h e s o i l micro-
f l o r a , o r may be a mat te r of s o i l chemis- t r y . Growth r a t e s of t h e v e g e t a t i o n may be changed, t h u s a f f e c t i n g t h e r a t e s of e lementa l cyc l ing . Each p o l l u t a n t may have s e p a r a t e e f f e c t s depending upon i t s chemical and phys io log ica l n a t u r e , and t h e r e may be s y n e r g i s t i c e f f e c t s w i th combinations of a d d i t i v e s . Some of t h e major ch ron ic p o l l u t a n t s w i l l be con-s i d e r ed s e p a r a t e l y .
Carbon Compounds &i Accompanying Oxi-dan-fcs
Carbon a d d i t i o n s t o t h e f o r e s t may be important because of t h e r o l e t hey p l ay i n p l a n t growth and subsequent minera l cyc l ing . F in layson and P i t t s (1976) have r epor t ed t h a t carbon may account f o r 45$ of t h e mass of smog aero- s o l . Much of t h i s may be t h e r e s u l t o f secondary r e a c t i o n s c r e a t i n g ca rboxy l i c a c i d s , e s t e r s , carbonyl compounds, a l c o h o l s , pe rox id i c polymers, l ong cha in a lkanes and a lkenes , and f a t t y a c i d s . I n a d d i t i o n , carbon i n t h e forms of va r ious compounds is a world-wide atmospheric p o l l u t a n t due t o human a c t i v i t i e s ox id i z - i n g reduced carbon f o r energy, and t o i n d i r e c t e f f e c t s such as enhanced oxida-t i o n of s o i l o rganic mat te r due t o c l e a r -i n g f o r e s t l a n d f o r a g r i c u l t u r e . Thus carbon d iox ide as a ch ron ic p o l l u t a n t has been measured by obse rva t ions a t Mauna Loa (Hawaii) t o be r i s i n g a t t h e r a t e o f from .35 t o 1 .79 ppm per year t o a l e v e l of 325 ppm i n 1 974 (Hobbs, g &., 1974).Pe terson (1969) i n a review s t a t e d t h a t carbon d ioxide a t i t s p re sen t l e v e l i n t h e atmosphere i s s t i l l l i m i t i n g t o p l a n t growth and t h a t t h e p r o d u c t i v i t y of p l a n t s should i n c r e a s e as t h e C o n con ten t i n c r e a s e s . A r e s u l t i n g i n c r e a s e of p l a n t and f o r e s t growth of 5$ by t h e year 2000 is a ' p r e d i c t e d , and t h i s would i n c r e a s e t h e r a t e s of uptake of necessary growth elements from t h e s o i l , t h u s i n c r e a s i n g r a t e s of e lemental cyc l ing . This a l l assumes o t h e r e lements o r requi rements such as water a r e no t l i m i t i n g t o p l a n t growth. On t h e o t h e r hand, r educ t ion i n p l a n t growth may occur where t o x i c carbon compounds from p o l l u t i o n cause damage t o t r e e f o l i a g e , t h u s reducing r a t e s of mineral cyc l ing . Ozone produced as a secondary product from u l t r a v i o l e t radia-t i o n on hydrocarbons i n t h e atmosphere may reduce p l a n t growth. Evans, g &. ( 1 974) found ozone c o n t e n t s of 0.1 ppm a t t h e t o p of a smoke plume from a f o r e s t f i r e . However p l a n t s syn thes i ze methyl c h l o r i d e accord ing t o Lovelock (1 975) , and t h i s may d e s t r o y such added ozone. The ox idan t s formed from t h e o rgan ic pol- l u t a n t s in t roduced t o t h e atmosphere w i l l have adverse e f f e c t s upon va r ious amino a c i d s i n p l a n t s r e s u l t i n g i n damage

--
(Haagen-Smit , 1958). F a t t y a c i d s on p l a n t s u r f a c e s may be a l t e r e d by t h e s i n g l e t oxygen r e s u l t i n g from NO2, ben-zaldehyde, and polynuclear hydrocarbons, i n c r e a s i n g s a t u r a t e d a c i d con ten t of t h e f o l i a g e accord ing t o Dowty &. (1 973) . Other o x i d a n t s such as peroxyacetyl n i t r a t e (PAN) , hydroxyl OH, hydroperoxyl HO , a l s o have similar e f f e c t s on vegeta- t i $ n . Their e f f e c t s as ox idan t s a r e def ined by t h e i r c a p a b i l i t y t o ox id i ze i o d i d e ion i n aqueous s o l u t i o n of potas-sium i o d i d e accord ing t o Kuntz, e t 9. (1 973). Also, t h e i r a c t i o n may b e s y n -e r g i s t i c , s o t h a t a l though one o r another is i n low atmospheric concen t r a t ion t h e sum of t h e ox idan t s must be considered i n e f f e c t on t h e v e g e t a t i o n . Also, t h e v o l a t i l e t e r p e n e s produced by t h e f o r e s t i t s e l f must be taken i n t o account , and accord ing t o Rasmussen (1 970) , as many as 1008 of t h e s p e c i e s i n some western coni-f e r o u s f o r e s t s may produce such v o l a t i l e hydrocarbons.
Nitrogen
Nitrogen as i t c y c l e s i n t h e f o r e s t i s i n reduced form i n t h e p l a n t and f o l -lowing m i n e r a l i z a t i o n t o ammonia i n decomposing d e t r i t u s i n t h e s o i l it is g r a d u a l l y oxid ized t o n i t r a t e where it is aga in taken up by p l a n t s t o be reduced i n va r ious n i t r o g e n compounds. Nitrogen is added as a ch ron ic a i r p o l l u t a n t as ammonium, and n i t r a t e n i t r o g e n which a l s o i s t h e end product of o x i d a t i o n of 80 by ozone. Where n i t r o g e n i s l i m i t i n g i n t h e f o r e s t as an e s s e n t i a l element t h i s should s t i m u l a t e f o r e s t growth and decom- p o s i t i o n p roces ses provided o t h e r e le -ments a r e no t l i m i t i n g . However, i f o t h e r e lements a r e l i m i t i n g t h i s would tend t o emphasize t h e o t h e r d e f i c i e n c i e s . Addi t ions as a ch ron ic p o l l u t a n t should show similar symptoms t o t hose o f t h e a d d i t i o n of n i t r o g e n as a f e r t i l i z e r . Thus f o l i a r n i t r o g e n c o n t e n t s should be h ighe r . A r e s u l t similar t o t h i s encoun-t e r e d i n Big cone spruce f o l i a g e i n t h e San Bernardino mountains i s repor ted l a t e r i n t h i s paper .
S u l f u r
t h a t The s u l f u r c y c l e i n f o r e s t s t h e s u l f u r i s i n reduced
is such form i n
a s s o c i a t i o n wi th o rgan ic compounds i n t h e p l a n t s , bu t i s oxid ized upon decomposi-t i o n i n t h e o rgan ic d e t r i t u s r e tu rned t o t h e s o i l . The decomposing s o i l micro-f l o r a conve r t t h e s u l f u r t o s u l f a t e . The degree t o which t h i s t a k e s p l ace depends upon t h e redox p o t e n t i a l of t h e s o i l . For example, i f it is ve ry low, t h e s u l f u r w i l l be r e t a i n e d i n s u l f i d e form, f r e -q u e n t l y t y i n g up heavy me ta l s .
The forms of s u l f u r added i n ch ron ic atmospheric p o l l u t i o n have been SO?, SO ", w i t h a s s o c i a t e d c a t i o n s H , NH a n t HSO as determined i n t h e p o l l u t h ~ plume trim S t . Louis Missouri by Charl-son , g 9. ( 1 973) . They found t h a t t r o p i c a l a i r masses were dominated by t h e more a c i d i c NH HSO wh i l e n o r t h e r n a i r masses were l e s s a c i d i c and dominated by (NH ) SO (1 975) . Usual ly t h e s u l f u r i n the4a?mo%-phere w i l l be oxid ized t o su l -f a t e and t h i s w i l l occur e i t h e r a s ammonium o r ca lc ium s u l h a t e d u s t accord- i ng t o daagen-Snit (1959), and s i n c e t r a p p i n g s u r f a c e s o f v e g e t a t i o n a r e aero-b i c , any reduced s u l f u r w i l l be oxid ized t o s u l f a t e i n t h e f o l i a g e .
S ince s u l f u r is an e s s e n t i a l element f o r p l a n t growth, t h e a d d i t i o n s may o r may n o t be d e l e t e r i o u s . Actual d e l e t e r i-ous e f f e c t s would occur t o t r e e s i f s u l -f u r d iox ide were t h e main a d d i t i v e . This would occur only c l o s e t o t h e emiss ion source as has happened a t Kennet t , C a l i -f o r n i a ; Copper Basin, Tennessee; and Sud- bury, Ontar io. A t g r e a t e r d i s t a n c e t h i s would be oxid ized . Presumably i f t h e p o l l u t a n t were added i n extreme amounts, t h e t i t r a b l e a c i d i t y produced would break through t h e s o i l exchange c a p a c i t y a f t e r d e p l e t i n g o t h e r c a t i o n s p r e s e n t . An example of a s imu la t ion of t h i s i s offered i n t a b l e 2 expla ined l a t e r .
Trace Elements lusts ' .
Washout, P a r t i c u l a t e s ,
Most of t h e elements o t h e r t han C , N, and S w i l l be added i n dus t and r a i n as ch ron ic p o l l u t a n t s . The p a r t i c u l a t e s i n atmospheric p o l l u t i o n c o n t a i n elements such as l e a d , sodium, magnesium, aluminium, vanadium, and z i n c i n Los Angeles t ype smog accord ing t o F in layson and P i t t s ( 1 976). Sedimentary c o r e s t aken o f f s h o r e i n t h e P a c i f i c Ocean by Bruland, e t S., (1974) , i n d i c a t e d t h a t l e a d , s i l v e r , copper , z i n c , chromium, n i c k e l , molybdenum, and c o b a l t a r e ch ron ic p o l l u t a n t s i n t h e Los Angeles a r e a . Lead, i r o n , manganese, n i c k e l , copper and z i n c were analyzed i n p r e c i p i -t a t i o n as common t r a c e element p o l l u t a n t s throughout t h e U.S. accord ing t o Lazarus e t- al . (1 970) . Beryllium is common i n i n d u s t r i a l d u s t s near sources . The amount of t r a c e element f a l l out l e s s e n s as t h e square of t h e d i s t a n c e from sources accord ing t o B e r t i n e & Goldberg (1974). Cannon and Boles (1962) found t h a t high-ways r ep re sen ted l i n e a r sou rces f o r l e a d , t h e amount i n v e g e t a t i o n dec reas ing r a p i d l y wi th d i s t a n c e from t h e source. Dedolph e t ( 1 970) found t h a t t h i s d i m u n i t i o n was loga r i thmic wi th d i s t a n c e f o r p a r t i c u l a t e l e a d . However, some e l e -ments such as cadmium, n i c k e l , l e a d , and

z i n c may b e v a p o r i z e d a t t h e s o u r c e and c a r r i e d l o n g e r d i s t a n c e s a c c o r d i n g t o Lagerwer f f & Spech t (1 970) .
The t r a c i n g o f i n p u t o f t r a c e e l e -ments i n t h e e l e m e n t a l c y c l e s i n vege ta - t i o n h a s been a t t e m p t e d by many i n v e s t i -g a t o r s . C h e s t e r and S t o n e r (1 973) used an e n r i c h m e n t f a c t o r i n which a r a t i o o f t h e e l e m e n t t o i r o n . i n t h e p a r t i c u l a t e b e i n g added i s d i v i d e d by t h e a v e r a g e r a t i o o f t h e e lement t o i r o n i n t h e e a r t h ' s c r u s t . They found t h a t t i n , l e a d , and z i n c most o f t e n were t h e e l e -ments e n r i c h e d b y p o l l u t i o n . P e i r s o n , e t -a l . (1 974) employed a n enr ichment f a c t o r i n t h e form o f t h e r a t i o o f t h e e lement t o Scandium c o n t e n t , compared t o t h e same f o r t h e l o c a l s o i l s . They found t h e s o i l s e n r i c h e d f o r V , Co, N i , Zn, A s , Se , Sb, and Pb t h i s way. S i m i l a r en r ichment r a t i o s f o r o t h e r p o l l u t a n t s shou ld iden-t i f y a u g m e n t a t i o n s o f t h e e l e m e n t s i n e l e m e n t a l c y c l e s i n f o r e s t s .
The c o n t e n t s o f p o l l u t a n t s i n t h e p l a n t s s h o u l d b e i d e n t i f i a b l e by a n o m a l i e s i n c o m p o s i t i o n . Thus, Schack-l e t t e and Connor (1 973) used t h e r e g i o n a l v a r i a t i o n of vanadium i n Span i sh moss ( ~ i l l a n d s i a ) a l o n g t h e g u l f c o a s t o f t h e U.S. t o i d e n t i f y a r e a s where c h r o n i c pol-l u t i o n by a i r b o r n e vanadium o c c u r s . Con-t e n t s as h i g h as 560ppm V were found i n c e r t a i n a r e a s , and t h e s e were assumed t o r e p r e s e n t t h e o u t p u t from o i l r e f i n e r i e s u s i n g Venezuelan c r u d e o i l which i s h i g h i n vanadium. I n s o i l - v e g e t a t i o n c y c l i n g , vanadium i s f r e q u e n t l y h e l d a t t h e s o i l -r o o t i n t e r f a c e due t o i m m o b i l i z a t i o n i n t h e o x i d i z e d s t a t e ( p e n t a v a l e n t f o r m ) . Thus, t h e redox s t a t e o f t h e s o i l -v e g e t a t i o n sys tem and i t s components i n t h e f o r e s t may d e t e r m i n e where a c h r o n i c p o l l u t i n g e lement may b e immobi l ized. E lements u n d e r g o i n g changes i n s o l u b i l i t y w i t h v a r i o u s o x i d a t i o n s t a t e s may b e immobi l ized a t v a r i o u s i n t e r f a c e s where pH o r Redox change. Chromium, vanadium, manganese, and i r o n a r e s u s c e p t i b l e t o t h i s .
Some o f t h e c h r o n i c p o l l u t a n t s added t o t h e e l e m e n t a l c y c l e o f f o r e s t s may l a t e r b e r e l e a s e d from t h e f o r e s t by v a p o r i z a t i o n . For example, C u r t i n e t a l . ( 1 974) found t h a t t i n i s t r a n s p i r e d i n v a p o r s from c o n i f e r s t o t h e amount o f 23-80 ppm i n t h e r e s i d u e o f t h e vapor . They found t h a t some c o n i f e r o u s t w i g s c o n t a i n e d up t o 6-40 ppm t i n i n t h e a s h . The t i n w a s added as a t m o s p h e r i c p o l l u -t a n t i n d u s t from i n d u s t r i a l a r e a s . How-e v e r , no ment ion w a s made o f t h i s as b e i n g d e l e t e r i o u s .
The i n p u t s o f d r y p a r t i c u l a t e m a t e r i a l s as d u s t s may b e washed o f f by r a i n . T h i s washoff may b e a major pro-c e s s of c y c l i n g o f t h e added e lement t o s o i l s . C a r l s o n --e t a l . (1 976) found t h a t a s i m u l a t e d r a i n removed 45% of a n a p p l i e d a e r o s o l o f PbC12, and t h a t l i g h t m i s t y r a i n i s most e f f e c t i v e . H e i c h e l & Hankin (1972) found t h a t t h e p a r t i c u l a t e s i n which l e a d adhered t o t r e e s averaged 7 mic romete r s i n d i a m e t e r . C h l o r i n e and Bromine were t h e main a s s o c i a t e d n e g a t i v e e l e m e n t s a s s o c i a t e d w i t h them. The assessment o f t h e i n t e n s i t y o f i n p u t o f c h r o n i c p o l l u t a n t s s u c h as t h e s e have been made by a n a l y z i n g c o n c e n t r a t i o n s on t r e e b a r k by L o t s c h e r t ( l 9 7 7 ) , and Grod-z i n s k a (1 977) . Some o f t h e s e accumula-t i o n s a r e washed down t h e t r e e t r u n k by s t em f l o w and may b e accumulated i n t h e s o i l a t t h e t r e e b a s e .
POLLUTANT INPUT I N PRECIPITATION
Washout as w e l l as i n p u t o f e l e m e n t s i n s o l u t i o n i n p r e c i p i t a t i o n is a major p a t h o f i n p u t t o t h e e l e m e n t a l c y c l i n g t h a t o c c u r s i n f o r e s t s . For example, t h e n i t r o g e n c o n t e n t s t o r e d i n a 1000 y e a r o l d redwood f o r e s t i s a b o u t t h e amount c o n t r i b u t e d by r a i n f a l l d u r i n g t h a t t i m e p e r i o d ( ~ i n k e ,5 g . , 1979) . The geo-c h e m i s t r y o f p r e c i p i t a t i o n w a s reviewed t h o r o u g h l y b y C a r r o l l (1 9 6 2 ) . She found t h a t t h e c a t i o n s which b a l a n c e t h e i o n s i n r a i n w a t e r were m o s t l y b a s i c m e t a l l i c e l e m e n t s n e a r c o a s t l i n e s o r a r i d r e g i o n s , b u t t h a t a l o n g t h e s t o r m v e c t o r from t h e s e s o u r c e s and over r e g i o n s w i t h more v e g e t a t i o n , t h e r e would t e n d t o b e a c i d i f i c a t i o n due t o l e s s d u s t o r ocean a e r o s o l s t o p r o v i d e t h e b a s i c m e t a l l i c c a t i o n s . The c o n t r o l o f wind e r o s i o n o v e r t h e G r e a t P l a i n s t h u s would t e n d t o add t o t h e a c i d f i c a t i o n o f r a i n f a l l downwind from t h e a r e a . However t h e c u r r e n t emphasis on p o l l u t i o n h a s added t h e c o n s i d e r a t i o n o f s o u r c e s o f a n i o n s i n p r e c i p i t a t i o n t h a t come from b u r n i n g f o s - s i l f u e l s . A s C a r r o l l ( 1 9 6 2 ) ment ioned, a pH below 5.7 i n d i c a t e s t h a t hydrogen is i o n i z i n g t o meet t h e n e c e s s a r y b a l a n c e w i t h a n i o n s .
The e f f e c t o f p r e c i p i t a t i o n i n p u t upon n u t r i e n t e l ement c y c l i n g i n a f o r e s t w i l l t e n d t o b e s p e c i f i c t o t h e s i t e con-d i t i o n s . T h i s would b e de te rmined by t h e f o l l o w i n g l o c a l f a c t o r s : 1. background n a t u r a l compos i t ion o f t h e s o i l s o l u t i o n . 2. t h e c a t i o n exchange c a p a c i t y o f t h e s o i l and t h e c o m p o s i t i o n o f c a t i o n s occu- py ing t h i s c a p a c i t y . 3. t h e volume o f wa te r e n t e r i n g as r a i n f a l l , 4. t h e a c i d b a s e b a l a n c e o f t h e r a i n f a l l , 5. t h e n a t u r e of t h e v e g e t a t i o n s p e c i e s w i t h

regard t o i n t e n s i t y of c y c l i n g b a s i c m e t a l l i c e lements , and perhaps o t h e r s unique t o t h e s i t e . The e f f e c t s of i npu t i n p r e c i p i t a t i o n w i l l most obvious on i n p r e c i p i t a t i o n w i l l be most obvious on sites wi th s o i l s having low exchange ma t t e r con ten t s o i l s ) and wi th s p a r s e slow growing v e g e t a t i o n ( p i n e s , sp ruces , h e a t h l a n d ) , and i n c l i m a t e s wi th l a r g e amounts o f p r e c i p i t a t i o n .
FOREST ECOSYSTEM EFFECTS
The minera l c y c l i n g i n a f o r e s t t a k e s p l a c e i n t h e c o n t e x t of an ecosys-tem of p roces ses l i n k i n g t h e va r ious l i v -i n g organisms and t h e components o f t h e environment. Chronic atmospheric pol lu-t i o n is one of t h e s e components. The e lements added w i l l be u t i l i z e d by t h e Producers i n t h e f o r e s t . These a r e t h e organisms which c a r r y out t h e r educ t ion of carbon t o carbon compounds which comprise t h e f o r e s t . This f i x e d carbon then s e r v e s as an energy source f o r t h e Decomposers i n t h e f o r e s t which a l s o have requi rements f o r t h e n u t r i e n t e lements e i t h e r i n t h e biomass of t h e producers o r fu rn i shed d i r e c t l y by t h e s o i l . Thus p o l l u t a n t e lements w i l l i n t e r a c t i n t h e c y c l e between Producers and Decomposers, w i th s t o r a g e of e lements and p o s s i b l e s i n k s of u n a v a i l a b l e m a t e r i a l accumulat- i n g i n t h e s o i l p o r t i o n of t h e c y c l e .
E f f e c t s -of Chronic P o l l u t i o n on Producers
These e f f e c t s can range from enhancement of p r o d u c t i v i t y t o a marked decrease depending upon whether t h e addi-
. t i v e is an e l e m e n t c u r r e n t l y l i m i t i n g growth o r is a t o x i c element. I f t o x i c , t h e e f f e c t s w i l l show up i n f o l i a r dam-age , o r i n t e r f e r e n c e wi th va r ious physio-l o g i c a l p roces ses . The syposium proceed- i n g s e d i t e d by Naegele (1973) con ta ins many d e s c r i p t i o n s of t h e s e t o x i c e f f e c t s . Mc Cune i n t h i s p u b l i c a t i o n f e l t t h a t such t o x i c e f f e c t s should be s p e c i f i c w i th regard t o t h e Receptor ( l e a f , t r e e , f o r e s t ) , t h e P o l l u t a n t , t h e Event , and t h e Environment. Acute e f f e c t s a r e r e l a -t i v e l y easy t o de te rmine , as i n t h e dea th of l e a v e s near a sme l t e r r e l e a s i n g SO2. Obviously t h i s would decrease t h e r a t e of e lementa l c y c l i n g a t t h e s i t e a long wi th a b r i e f s u r g e of added m a t e r i a l s t o t h e s o i l as t h e d e f o l i a t i o n took p l a c e and l eaves dropped t o t h e s o i l t o be decom-posed. Whether t h i s is a l o s s t o t h e s i t e depends upon t h e c a p a c i t y o f t h e unde r ly ing s o i l as d i scussed i n r e l a t i o n t o d e f o l i a t i o n by h e r b i c i d e s by Zinke (1974) . The e f f e c t s a r e more d i f f i c u l t t o a s s e s s i n t h e case o f ch ron ic low l e v e l p o l l u t i o n as d iscussed by 'Feder (1 973) . The changes i n p r o d u c t i v i t y
would need t o be s t a t i s t i c a l l y s i g n i f i -cant be fo re and a f t e r t h e i n i t i a t i o n of t h e p o l l u t i o n . Mi l l e r (1 973) has made an approach t o i n d i c e s of l e v e l of ch ron ic p o l l u t i o n i n terms of v i s u a l c h a r a c t e r i s -t i c s on t h e t r e e s ; need le r e t e n t i o n , nee-d l e l e n g t h , needle c h l o r o s i s , branch dea th i n t h e case of ponderosa p ine . Fol iage composition should i n d i c a t e such i n p u t s , and a paper by Arkley and Glauser i n t h i s symposium covers t h i s . Also an example i n t h e case of Big Cone Spruce f o l i a g e is presented i n t a b l e 1 . McBride, e t al . (1975) made measurements of a c t u a l g r o w t h r a t e r e t a r d a t i o n o f 26% f o r he ight and volume growth of ponderosa
In1974) found a g r i c u l t u r a l p l a n t s , White a decrease i n C02 uptake due
t o s y n e r g i s t i c e f f e c t s of NO + SO2 when each were p re sen t a t l e v e f s n o t con-s ide red i n h i b i t i n g t o growth. S t imu la t ion of primary producers by p o l l u t i o n is no t o f t e n r e p o r t e d due t o t h e nega t ive conno-t a t i o n of t h e t e r m . However, Schnappinger (1975) found an i n c r e a s e i n growth due t o response t o z i n c conta ined i n f l y ash .
There a r e numerous v a r i a b l e s which may a f f e c t t h e i n t e n s i t y w i th which a given l e v e l of p o l l u t a n t w i l l a f f e c t t h e producers i n an ecosystem. Thus t h e t r a p p i n g e f f i c i e n c y of t h e f o l i a g e sur -f a c e is d i f f e r e n t f o r va r ious s p e c i e s as d iscussed by Zinke (1966) , t h e v e l o c i t y of t h e a i r pas t t h e t r a p p i n g s u r f a c e as h a s been eva lua ted by Hori (1 953) and S l i n n ( 1 976). Following t r a p p i n g t h e m a t e r i a l may not be absorbed by t h e sur-f a c e as found by Motto (1970) . This is p a r t i a l l y a t t r i b u t a b l e t o t h e p a r t i c l e s i z e accord ing t o Natusch & Wallace (1974) . There may be s y n e r g i s t i c e f f e c t s a t t h e f o l i a g e s u r f a c e . iiovelock (1974) has a l l uded t o t h e p o s s i b l e c r e a t i o n of PAN on f o l i a g e s u r f a c e by t h e combination of 0 and hydrocarbons ing l i n e n .
Once absorbed by p o l l u t i n g element w i l l t a l c y c l e on t h e s i t e . from t h e p l a n t may be
as occurs on dry-
t h e producer t h e e n t e r t h e elemen-
The d i s p o s i t i o n n o t on ly through
l e a f drop , but may be by d i r e c t r o o t exu-d a t i o n as Olson e t a1 (1 962) found wi th a d d i t i o n s of r a d i o a c t i v e Cesium added t o a t r e e t runk . However upon be ing dropped t o t h e s o i l as d e t r i t u s o r e n t e r i n g i n o t h e r ways, t h e p o l l u t i n g element w i l l t hen be a f f e c t i n g t h e decomposers i n t h e f o r e s t .
E f f e c t s of- Chronic P o l l u t i o n on Decom-nosers
The e f f e c t s upon decomposers, t h e s o i l mic ro f lo ra and f auna t h a t complete

t h e e lementa l c y c l e s i n t h e s o i l , w i l l be v a r i a b l e . A s w i th t h e producers which f i x carbon d i o x i d e , t h e a d d i t i v e s t o t h e s i t e th rough a i r p o l l u t i o n may i n c r e a s e o r decrease t h e i r a c t i v i t i e s . These organisms r e q u i r e t h e same n u t r i e n t e l e -ments as t h e producers as w e l l as t h e a d d i t i o n a l ones of sodium, i o d i n e , and vanadium f o r t h e s o i l fauna. If t h e pol-l u t a n t s a r e added i n t o x i c q u a n t i t i e s t h e y would presumably r e t a r d t h e decompo-s i t i o n of organic ma t t e r i n t h e s o i l and thus slow o r b lock e lementa l c c l i n g .They may be s u b l e t h a l o r even mutagenic t o t h e s e organisms accord ing t o S to t zky 1 9 7 4 ) .
The d e t r i t u s o r l i t t e r l a y e r on t h e f o r e s t f l o o r is t h e f i r s t major a r e a of a c t i v i t o f t h e decomposers. Wittkamp & Frank (1969) found t h a t l i t t e r samples impregnated w i t h in t roduced elements such as Cobal t 60 and Cesium 137 had loga-r i t h m i c r e l a t i o n s h i p wi th t ime i n t h e f r a c t i o n remaining, and t h a t l o s s c o e f f i - c i e n t s were d i f f e r e n t f o r d i f f e r e n t e l e -ments. These v a r i e d w i t h t h e s p e c i e s of t r e e y i e l d i n g t h e d e t r i t u s . The s t o r a g e per iod of t h e a d d i t i v e element depended upon r e l a t i v e chemical m o b i l i t y of t h e element , composi t ion of o the r e lements p r e s e n t , and t h e mois ture and tempera ture c o n d i t i o n s o f t h e s i t e .
If a p o l l u t a n t i s an element which i s l i m i t i n g r a t e of decompostion, t h e e f f e c t on t h e decomposers may be posi-t i v e . For example l e a f l i t t e r wi th t o o h igh a ~ a r b o n / ~ i t r o g e nr a t i o ( n i t r o g e n l i m i t i n g ) decomposes ve ry s lowly . Adding n i t r o g e n as a ch ron ic p o l l u t a n t would a c c e l e r a t e t h i s decomposition and r e l e a s e s t o r e d elements more r a p i d l y .
The s o i l f a u n a s e r v e a major t a s k i n e lementa l c y c l i n g i n a f o r e s t ecosystem by p h y s i c a l l y decomposing organic d e t r i t u s , reducing i t i n s i z e and i n c r e a s i n g s u r f a c e a r e a t o b r i n g about f a s t e r r a t e s of funga l and b a c t e r i a l decomposition. Earthworms of va r ious s p e c i e s were found by Gish & Chris tensen (1973) t o accumulate t r a c e elements from p a r t i c u l a t e p o l l u t i o n i n o rde r of i n c r e a s i n g atomic weight : N i < Zn < Cd < Pb. The s o i l f auna may s e r v e as a l i n k f o r t r a n s f e r of n u t r i e n t e lements t o v a r i o u s p r e d a t o r s i n t h e f o r e s t ecosys-tem.
S to t zky ( 1 974) c h a r a c t e r i z e s t h e s o i l as bo th an a b i o t i c as w e l l as b i o t i c s i n k f o r p o l l u t a n t s . Ethylene and C O a r e degraded by s o i l micro organisms. Fungi i n s o i l metabol ize v o l a t i l e s from p l a n t f o l i a g e such as e thane , b u t a n e , , e t c . Thus s o i l by v i r t u e o f t h e v a r i o u s organ- i s m s p r e s e n t has a s t r o n g c a p a b i l i t y t o
d e t o x i f y p o l l u t a n t s . A t low concentra-t i o n s p o l l u t a n t s may b r i n g about an enrichment o f t h e micro organisms which can u t i l i z e them.
The u l t i m a t e r o l e of t h e decomposers i s t h u s t o c y c l e elements from t h e vege-t a t i o n t o t h e s o i l , u s u a l l y o x i d i z i n g them as energy s o u r c e s , and u l t i m a t e l y r e l e a s i n g them t o t h e chemical and physi-c a l p roces ses of t h e s o i l o r renewed uptake by t h e r o o t s of t h e producers .
Chronic P o l l u t a n t s i n t o S o i l
The f i n a l f a t e of a d d i t i v e s t o t h e ecosystem may be t o remain i n t h e s o i l i n i n s o l u b l e form. Thus once processed by t h e decomposers t h e elements may become minera l p r e c i p i t a t e s which may a c t as s i n k s (Nat . Res. Council 1977) . Thus f l u o r i n e added through p o l l u t i o n may be p r e c i p i t a t e d i n t h e s o i l a s i n s o l u b l e CaF2. Page and Ganje (1970) showed t h a t t he t o p 2.5 cm. of s o i l w a s most a c t i v e i n t r a p p i n g l e a d e n t e r i n g t h e s o i l -v e g e t a t i o n system i n sou the rn C a l i f o r n i a . S o i l s sampled i n 1967 were compared wi th previous samples obta ined i n 191 9. An average inc rease of 15-36 ppm l e a d accounted f o r 1/5 of t h e l e a d contained i n t h e 47 b i l l i o n g a l l o n s of g a s o l i n e which had been burned du r ing t h e per iod encompassed. Keaton ( 1937) added l a r g e amounts of l ead n i t r a t e t o s o i l (2784 ppm) and found t h a t very l i t t l e ( 1 7 ppm) remained i n s o l u b l e form. Lee & Tallis (1973) found it p o s s i b l e t o d a t e p e r i o d s of l e a d p o l l u t i o n i n a p e a t bog i n Grea t B r i t a i n , f i n d i n g a peak i n 500 A.D. w i t h t h e o p e r a t i o n of Roman l e a d mines, and a subsequent i n d u s t r i a l peak beginning about 1460 AD. S imi l a r b u t r e c e n t h i s -t o r i c a l evidence of l e a d c o l l e c t i o n i n ecosystem s i n k s w a s found i n wood con ten t of l ead dated by annual r i n g s by Sheppard & Funk (1975) .
S o i l o rganic mat te r g e n e r a l l y enhances t h e r o l e of s o i l as a s i n k f o r added t r a c e elements . Purr e t a1 (1971) have noted t h a t pH w i l l determine whether s o i l i s a sou rce o r s i n k . Thus, a t neu-t r a l t o s l i g h t l y a l k a l i n e pH; A s , B, Cd, Go, Cu, F, Fe, Pb, Mn, N i , Sn, and Zn w i l l tend t o be immobilized, bu t Ca, Cr, K Mg, Mo, Rb, Se, and Sr may become more a v a i l a b l e t o p l a n t s . Thus anyth ing caus-i n g a change i n s o i l pH may r e l e a s e pre-vious p o l l u t a n t s t h a t had t empora r i l y become t rapped i n t h e s o i l as a s i n k . Various p r e c i p i t a t e s such as s o i l car-bonates may form, a l s o depending upon pH.
The s o i l i on exchange c a p a c i t y w i l l r e t a i n elements i n a form ready t o be
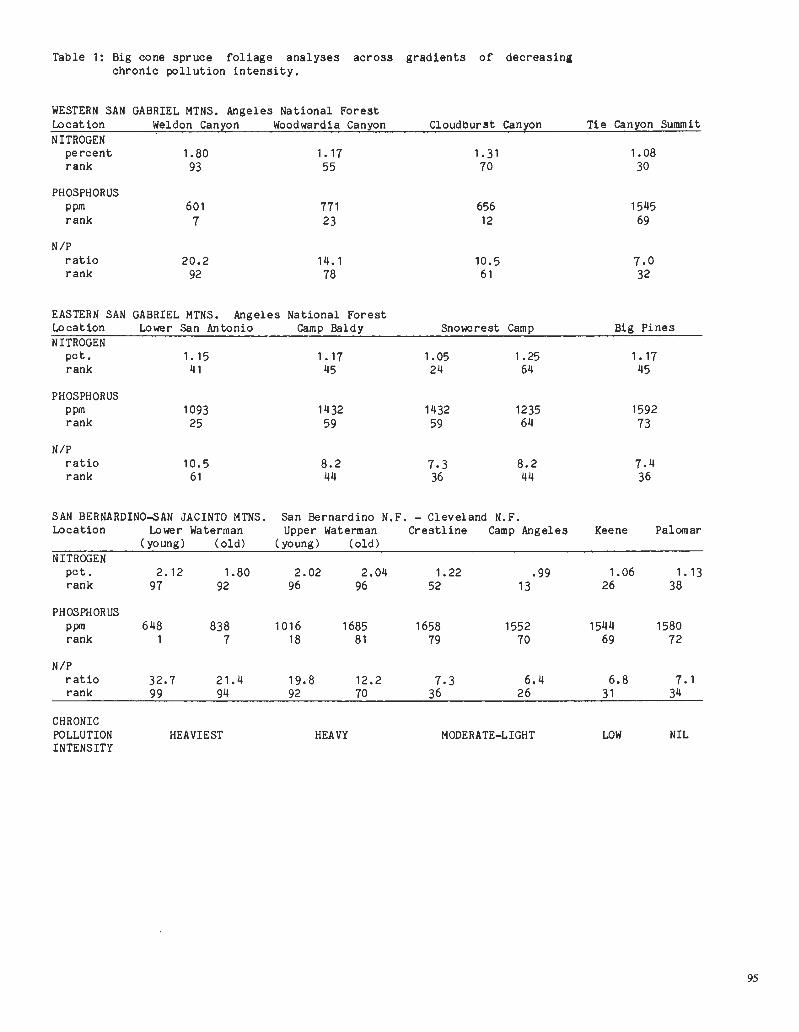
Table 1: Big cone sp ruce f o l i a g e a n a l y s e s a c r o s s g r a d i e n t s o f dec reas ing ch ron ic p o l l u t i o n i n t e n s i t y .
WESTERN SAN GABRIEL MTNS. Angeles Nat ional F o r e s t Location Weldon Canyon Woodwardia Canyon Cloudburst Can yon T i e Canyon Summit NITROGEN
p e r c e n t 1.80 r ank 93
PHOSPHORUS PPm 601 r ank 7
N /P r a t i o 20.2 r ank 92
EASTERN SAN GABRIEL MTNS. Angeles Nat ional F o r e s t Locat ion Lower San Antonio Camp Baldy Snowcrest Camp Big P i n e s NITROGEN
p e t . rank
PHOSPHORUS Ppm 1093 rank 25
N /P r a t i o r ank
SAN BERNARDINO-SAN JACINTO MTNS. San Bernardino N.F. - Cleveland N.F. Locat ion Lower Waterman Upper Waterman C r e s t l i n e Camp Angeles Keene Palomar
(young) ( o l d ) (young) ( o l d ) NITROGEN
p o t . 2.12 1.80 2.02 2.04 1.22 .99 1.06 1.13 rank 97 92 96 96 52 13 26 38
PHOSPHORUS PPm 648 838 1016 1685 1658 1552 1544 1580 r ank 1 7 18 . 81 79 70 69 72
N /P r a t i o 32.7 21.4 19.8 12.2 7.3 6.4 6.8 7.1 rank 9 9 94 92 70 36 26 3 1 34
CHRONIC POLLUTION HEAVIEST HEAVY MODERATE-LIGHT LOW NIL INTENSITY

Table 2: S imula t ion of a c i d r a i n f a l l a d d i t i o n by l each ing a sur-f a c e s o i l developed under t h e i n f luence of a mature pon- de rosa p ine a t t h e a r i d l i m i t o f t h e occur rence of t h e s p e c i e s . ( i n m i l l i e q u i v a l e n t s per 100 m s . f i n e e a r t h p o r t i o n ex-c e p t last column as i n d i c a t e d )
Exchangeable Cat ions S o i l Treatment Exchange Remaining on S o i l
Capac i ty %
Leached s o i l s :
Leachate Composition- 3 I 500 m l H20
+5.71 meq. H+
+11 .40 meq. H+
+17.09 meq. H+
+22.9 meq. H+
1' Fine e a r t h p o r t i o n , 0-2.5 i nch sampling hor izon .
D i f f e r ence between t h e sum ( ~ a " + M ~ + + + K' + ~ a ' ) and t h e Exchange Capaci ty .
^-IThe H' w a s added as 0.1N H C 1 t o the.500 m l of d i s t i l l e d wa te r , s i n c e 1 m l of 0.1N H C 1 = 0.1 meq. H .
recyc led back t o t h e l i v i n g organisms on t h e s i t e , and a l s o keep them from leach- i n g away from t h e s i t e . Depending upon t h i s c a p a c i t y , e lements added through ch ron ic p o l l u t i o n may be r e t a i n e d i n t h e e l emen ta l c y c l e of a f o r e s t provided t h e y a r e n o t t rapped i n a s i n k o r l eached away.
FOLIAGE ANALYSES I N D I C A T I N G FOLLrnION
The e f f e c t o f ch ron ic air p o l l u t i o n of t h e t ype t y p i c a l of t h e Los Angeles -San Bernardino b a s i n on f o l i a r ana lyses of t h e Cone Spruce (Pseudotsuga macrocarpaBi. Vasey) Mayr) w a s determined. Samples o f t h e f o l i a g e were ga thered throughout t h e range of t h e s p e c i e s , and a l s o on g r a d i e n t s o f e l e v a t i o n o r d i s -t a n c e from t h e a r e a s t y p i f i e d by "smog" t o a r e a s r e l a t i v e l y f r e e o f "smog". Ana-l y s e s were made of 9 major e lements i n t h e v a r i o u s age c l a s s e s of t h e f o l i a g e . The e lements most obvious ly r e l a t e d t o t h e i n t e n s i t y o f t h e smog and t h e i r rat-i n g i n a cumula t ive p r o b a b i l i t y d i s t r i b u -
t i o n from lowest t o h ighes t va lue a r e shown i n t a b l e 1.
Nitrogen con ten t of t h e f o l i a g e and t h e r a t i o of n i t r o g e n t o phosphorus con-t e n t were t h e most obvious i n d i c a t o r s of t h e e f f e c t o f ch ron ic air p o l l u t i o n o f t h e Los Angeles t ype . Where t h e r e w a s ambigui ty i n t h e n i t r o g e n v a l u e s as i n San Antonio Canyon, t h e n i t r o g e n t o phos-phorus r a t i o was t h e b e s t i n d i c a t o r i n t h i s s p e c i e s .
SIMULATED A C I D RAINFALL LEACHING
A l a b o r a t o r y experiment w a s con-ducted i n which samples o f a n e a r l y c a t i o n s a t u r a t e d s o i l under t h e i n f l u e n c e of ponderosa p ine were leached wi th so lu- t i o n s c o n t a i n i n g s u c c e s s i v e increments of H The s o i l samples were ob ta ined a t t h e lower r a i n f a l l l i m i t o f ponderosa p ine where t h e s o i l was most l i k e l y t o be base s a t u r a t e d . F ive s e p e r a t e t e n gram samples of t h i s s o i l were leached by s o l u t i o n s which were r e s p e c t i v e l y 500 m l of d i s t i l l e d wa te r , and s o l u t i o n s t o

which increments of H' as H C 1 equ iva l en t t o 20$, 40$, 808, of t h e exchange capa- c i t y of t h e sample were added. The r e s u l t s presented i n t a b l e 2 show t h e l e a c h i n g of t h e exchange complex of s o i l s t h a t t a k e s p l a c e wi th added increments of a c a t i o n . Presumably t h e a d d i t i o n s of increments of H i n a c i d r a i n f a l l would have similar e f f e c t s which would have t o be cons idered i n t h e con tex t of o t h e r c a t i o n s be ing cycled by t h e f o r e s t , and by t h e composi t ion of c a t i o n s a l r e a d y on t h e exchange complex of t h e s o i l . Follow- i n g t h i s l e a c h i n g , each s o i l p o r t i o n w a s analyzed f o r remaining exchangeable m e t a l l i c c a t i o n s .
These d a t a show t h a t given t h e s to- chiometry of t h e l e a c h i n g of c a t i o n s from t h e s o i l exchange complex by H t h e e f f e c t s of increments of a c i d r a i n f a l l t o a g iven s o i l may be s imula ted wi th so lu- t i o n s having composi t ions s i m i l a r t o t h a t of t h e r a i n f a l l . Also, as t h e increment of H i n t h e p r e c i p i t a t i o n inc reased , e i t h e r because of concen t r a t ion o r of volume of r a i n f a l l l e a c h i n g t h e s o i l would tend t o become hydrogen s a t u r a t e d , d i s p l a c i n g b a s i c m e t a l l i c c a t i o n s i n pro- p o r t i o n . In t h e f o r e s t s i t u a t i o n t h e s e would be leached out o f t h e s o i l p r o f i l e i n a base s a t u r a t e d s o i l , o r taken up by t h e v e g e t a t i o n .
CONCLUSIONS
This review of t h e l i t e r a t u r e and t h e p r e s e n t a t i o n of some o r i g i n a l work i s a p p l i c a b l e t o t h e e f f e c t of ch ron ic a i r p o l l u t i o n on minera l element c y c l i n g of f o r e s t s . The l i t e r a t u r e i s g e n e r a l , i l l u s t r a t i n g p roces ses , but i s no t s i t e s p e c i f i c except f o r t h e l o c a t i o n of t h e s t u d i e s . S ince e f f e c t s a t any s p e c i f i c s i t e may be i n small increments each y e a r , but s eve re i n cumulat ive e f f e c t s over a number o f y e a r s , a major remaining problem is t o f i n d ways of determining s i t e s p e c i f i c i n d i c a t i o n s of d e t r i m e n t a l e f f e c t s of such ch ron ic p o l l u t i o n . Some p e r t i n e n t conc lus ions from t h e l i t e r a t u r e reviewed i n t h i s paper a r e :
1 . The f o r e s t v e g e t a t i o n as is t r u e of any v e g e t a t i o n r e q u i r e s many e s s e n t i a l growth e lements which i t w i l l cyc l e i n a d d i t i o n t o o t h e r s mobilized by l o c a l minera l weather ing o r i npu t through pre- c i p i t a t i o n .
2. To t h e e x t e n t t h a t ch ron ic p o l l u t i o n adds e lements normally r equ i r ed by p l a n t s , o r chemica l ly mobile i n t h e p l a n t , t h e s e w i l l be cycled much as t h e o t h e r s on t h e s i t e .
3. The f o r e s t i s c r e a t e d i n a v a s t reduc-
t i o n r e a c t i o n wherein CO i s reduced t o organic compounds by Producers i n t h e ecosystem, and subsequent ly oxid ized i n energy r e l e a s i n g s t e p s by t h e s e p l a n t s o r by t h e Decomposers i n t h a t ecosystem.
4. The cyc l ing of e lements t a k e s p l ace i n t h e con tex t of t h e s e redox r e a c t i o n s , and those elements which change s t a t e w i th redox p o t e n t i a l change a r e u s u a l l y i n t h e reduced form i n t h e producer v e g e t a t i o n and subsequent ly oxid ized i n energy r e l e a s i n g s t e p s by t h e s e p l a n t s o r by t h e decomposers on t h e s i t e .
5. Chronic air p o l l u t i o n a d d i t i v e s t o t h e f o r e s t a f f e c t the c y c l i n g of e lements n o t on ly by d i r e c t a d d i t i o n s of t h e p o l l u t i n g elements t o t h e c y c l e s , bu t a l s o by a f f e c t i n g t h e redox s t a t u s of t h e absorb- i ng p l a n t t i s s u e , and o the r p o r t i o n s of t h e s o i l - v e g e t a t i o n system.
6 . Concentrat ion p o i n t s and s i n k s f o r t h e elements added i n chronic p o l l u t i o n e x i s t i n t h e va r ious p a r t s of t h e v e g e t a t i o n , t h e s u r f a c e d e t r i t u s on t h e s o i l , and i n t h e s o i l c a p a c i t i e s f o r abso rp t ion on ion exchange; hydrous ox ide complexes, and as i n s o l u b l e p r e c i p i t a t e s .
7. The e f f e c t s of chronic p o l l u t a n t s added t o a given f o r e s t and i t s e lementa l c y c l i n g processes w i l l be h igh ly s i t e s p e c i f i c ; depending upon c l i m a t e , degree of weathering development of t h e s o i l , t h e type of mine ra l s from which t h e s o i l is forming, t h e c h a r a c t e r i s t i c s of t h e f o r e s t s p e c i e s and t h e i r s tock ing densi- t i e s , t h e topographic s i t u a t i o n wi th regard t o a i r f l o w , and t h e n a t u r e of t h e p o l l u t a n t .
8. Local s i t e s p e c i f i c i n d i c a t o r s o f i n t e n s i t y of p o l l u t i o n e f f e c t may be v i s u a l as i n appearance and s i z e of f o l i - age, f o l i a r ana lyses f o r p o l l u t a n t e l e - ments and t h e i r r a t i o s t o background s i t e e lements , and s o i l assessment of capaci- t i e s t o absorb t h e added elements.
LITERATURE CITED
B e r t i n e , K. & E. Goldberg. 1977. H i s to ry of Heavy meta l p o l l u t i o n i n sou the rn C a l i f o r n i a Coas ta l Region. Env. Sc i & Tech. 1 1 : 3 297-99 .
Bruland, K.W., K. B e r t i n e , M. Koides, & E. Goldberg. 1974. H i s to ry of metal p o l l u t i o n i n sou the rn C a l i f o r n i a Coa- s ta l Zone. Env. S c i . & Tech. 8:5 425- 432
Cannon, H.L. & J . Bowles. 1962. Sc ience 137, 765-766

Car lson , R. , F. Bazzaz, and J . S tukel . 1976. Phys io log ica l e f f e c t s , wind reen-t ra inment and rainwash of Pb a e r o s o l p a r t i c l e s depos i t ed on p l a n t l e a v e s . Env. Sc i & Tech. 10:12 11391142.
C a r r o l l , D. 1962. Rainwater a s a chemical agent o f geologic processes . U.S. G. S. Water Supply Paper 1535-G 18pp + 1 p l a t e .
C u r t i n , G . C . , H. King, & E.L. Nosier . 1974. Movement of e lements i n t o t h e at-mosphere from con i f e rous t r e e s i n suba lp ine f o r e s t s of Colorado and Idaho. J. Geochem. Explor. 3:245-263.
Dowty, B. J. L a s e t e r , G. G r i f f i n , I Pol-i t z e r , & C. Walkinshaw. 1973. Reac t ions of s i n g l e t oxygen wi th p ine po l l en . Sc ience 181 1 7 Aug. 1973 669-670.
Evans, L.F., N. King, D. Packham & E.T. Stephens. 1974. Ozone Measurements i n smoke from f o r e s t f i r e s . Env. Sc i & Tech. 8: 1 75-76.
Evans,L. I.Weeks, A. Ecc l e s ton , & D. Packham. 1977. Photochemical ozone i n smoke from p resc r ibed burn ing of f o r e s t s . Env. S c i & Tech. 1 1 : 9 896-900.
Feder , W.A. 1973. Cumulative e f f e c t s of Chronic Exposure o f p l a n t s t o low lev- e l s of a i r p o l l u t i o n . pp. 21-30 i n Air P o l l u t i o n Damage t o Vegeta t ion . Adv. i n Chem. S e r i e s #I22 Amer Chem. Soc. Wash. D . C .
F in layson , B. & J. P i t t s . 1976. Photo-chemis t ry of t h e p o l l u t e d t roposphere . Sc ience v. 192 111-119.
F r i e d l a n d e r , S. K. 1973. Small p a r t i c l e s i n a i r pose a b i g c o n t r o l problem. Env. S c i . & Tech. 7:13 1115-1118.
P u r r , K . , T. Park inson , R. H in r i chs , D. Van Campen, C. Bache, W.Gutenann, L. St John, I Pakkala , & D . L isk 1977. Na-t i o n a l Survey of Elements and Radioac-t i v i t y i n F l y ashes . Absorption of e l e -ments by cabbage grown i n f l y a sh s o i l mix tures . Env. S c i & Tech. 11:13 1194-1 201 .
G a r r e l s , R.M., F. McKenzie, C. Hunt. 1975. Chemical Cycles and t h e Global Environment. 206pp. Wm. Kaufmann Inc . Los A l t o s , Calif.
Gish, C. & R. Chr i s tensen . 1973. Cadmium, Nicke l , Lead, and Zinc i n earthworms from roads ide s o i l . Env. S c i . & Tech. 7:11 1060-1062.
Goldberg, E. D. 1971 . Atmospheric d u s t ,
t h e sedimentary c y c l e , and man. Geophy-s i c s 1 :117-1 32.
Grodzinska, K. 1977. Changes i n t h e f o r e s t environment i n sou the rn Poland as a r e s u l t o f s t e e l m i l l emissions. Pp. 207-21 5 i n Vegeta t ion , Sc ience , and Environmental P r o t e c t i o n . Maur zen , Ltd. , Tokyo.
Haagen-Smit , A . J. 1 958. Air Conservat ion. Science 128 no. 3329 869-878.
Heiche l , G.H. & L. Hankin. 1972. P a r t i -c l e s con ta in ing l e a d , c h l o r i n e , and bromine d e t e c t e d on t r e e s w i th an e l ec -t r o n microprobe. Env. S c i . & Tech.6: 13 1121-1 122.
Hobbs, P.V., Har r i son , H. and E. Robin-son. 1974. Atmospheric e f f e c t s of pol-l u t a n t s . Science 183(41 28) :909-91 5.
Hori , J. 1953. On t h e d i f f u s i o n by t u r -bu len t motion near a f o r e s t and i t s e f -f e c t upon t h e c a p t u r e of f o g s by t h e f o r e s t . Low Temperature S c i . , Ser . A , Phys. Sc i . pp. 75-86. Abstr . i n Trans. Amer. Geophys. Union 35:2 327.
Jenne , E.A. 1968. Con t ro l s on Mn, Fe, Co, N i , Cu, and Zn c o n c e n t r a t i o n s i n s o i l s and water . The s i g n i f i c a n c e of hydrous Mn and Fe oxides . Ad.v. Chem. Se r . 73: 337-367.
Lagerwerff , J. & A. Specht. 1970. Contam-i n a t i o n of roads ide s o i l and v e g e t a t i o n wi th Cadmium, Nickel , Lead, and Zinc. Env. S c i & Tech. 4:7 583-585.
Lazarus, A.L., E. Larange, & J.P. Lodge. 1930. Lead and o the r meta l i o n s i n Un-i t e d S t a t e s P r e c i p i t a t i o n . Env. S c i . & Tech. 4:l 5 - 58 .
Lee, J. & J. T a l l i s . 1973. Regional and H i s t o r i c a s n e c t s of l e a d ~ o l l u t i o n i n B r i t a i n . ~ a t u r e , Vol. 245 sept. 28 1 973 21 6-21 8.
Lin ton , R . , A. Loh, D. Natusch, C. Evans, & P. Williams. 1976. Surface predomi-nance of t r a c e elements i n a i r b o r n e p a r t i c l e s . Science v o l . 191 27Feb. 1976 852-854.
L o t s c h e r t , W. 1977. Bark of 'deciduous t r e e s as an i n d i c a t o r f o r a i r po l lu -t i o n . Pp. 247-255 i n Vegeta t ion , Sci-ence, and Environmental P r o t e c t i o n . Maur zen, Lt d ., Tokyo.
Lovelock, J. 1975. Natura l halocarbons i n t h e a i r and i n t h e sea . Nature Vol. 256 Ju ly 17 1975 pp.193-194.
98

McBride, J . , V. Semion, P. M i l l e r . 1975. Impact of a i r p o l l u t i o n on t h e arowth o f ponderosa p ine . C a l i f . ~ ~ r i c k t u r e Dec. 1975 8-9.
M i l l e r , Paul R . 1973. Oxidant induced community change i n a mixed c o n i f e r f o r e s t . pp. 101-117 i n A i r P o l l u t i o n Damage t o Vegetat ion. Adv. i n chem. S e r i e s #122. Amer. Chem. Soc. Wash. D . C .
Motto, H.L. R Daines, D. Chilko, C. Mot- t o . 1970. Lead i n P l a n t s and S o i l s . Env. S c i & Tech. 4:3 231 -237.
Nat iona l Aeronaut ics and Space Author i ty NASA. 1968. Appl ica t ion of Biogeochem- i s t r y t o minera l p rospec t ing . NASA SP- 5056 134pp. O f f i c e of Technology U t i l i - z a t i o n . NASA Wash. D . C .
Nat iona l Research Council . 1977. Geochem- i s t r y and t h e Environment. Vols I & 11.
Matusch, D . , J. Wallace. 1974. Urban a e r o s o l t o x i c i t y ; t h e i n f l u e n c e of ¥oar t i d e s i z e . s c i e n c e v . 186: no. 4165 695-699
Olson, J .S . , D.A. C r o s s l ~ ~ & M. Witkamp. 1962. Mob i l i t y of C s t r a c e r i n a f o r e s t o f T u l i p Poplar (Li r iodendron t u l i i f e r a ) . Heal th Phys ics Div. Oak h i o n a l Laboratory. Ann. Pro- g r e s s r e p o r t . No. ORNL 3347 I962 ( 6 6 ) . Oak Ridge Tenn.
Page, A. & T . J . Ganje. 1970. Accumula- t i o n s o f l e a d i n s o i l s f o r r eg ions of h igh and low motor v e h i c l e t r a f f i c den- s i t y . Env. Sc i . and Tech. 4:2 140-142.
Pe t e r son , E. 1969. Carbon d iox ide a f f e c t s g l o b a l ecology. Env. S c i & Tech. 3:11 1 162-1 169.
Rasmussen, R. 1970. Isoprene: I d e n t i f i e d as a F o r e s t t ype emission t o t h e atmo- sphere . Env. Sc i & Tech. 4: 8 667-671 .
S c h a c k l e t t e , H . J . & J . J. Connor. 1973. Airborne chemical e lements i n Spanish Moss. U.S.G.S. Prof . Paper 574s 46pp. U.S. Gov. P r i n t i n g O f f i c e , Wash. D.C.
Schnappinger, M. D. Martens, & C . Plank. 1975. Zino a v a i l a b i l i t y as inf luenced by, a p p l i c a t i o n of f l y ash t o t h e s o i l . Env. S c i . & Tech. 9:3 258-261 .
Shephard, J. & W. Funk. 1975. Trees as Environmental s e n s o r s moni tor ing long term heavy meta l contaminat ion of t h e Spokane R ive r , Idaho. h v . Sc i . & Tech. 9:7 638-642.
S l i n n , W. 1976.' Some approximations f o r t h e wet and d r y removal of p a r t i c l e s and gases from t h e atmosphere. Jou r . O f A i r , Water, and S o i l P o l l u t i o n . Accept- ed fo'r Publ.
Smith, W. 1973. Metal contaminat ion of Woody P l a n t s . Env. S c i & Tech. 7:7 631 -636.
S to t zky , G. 1974. A c t i v i t y , Ecology, & Popula t ion dynamics of microorganisms i n S o i l . i n Microbial Ecology by Las- k i n , A . I . & H. Lecheval ie r . CRC P r e s s Cleveland Ohio. pp. 73-1 35.
Stumm, W. & J . J . Morgan. 1970. Aquatic Chemistry. 583pp. Wi ley- In te rsc ience N.Y.
White, K.L., A.C. H i l l & J. Bennet t . 1974. S y n e r g i s t i c i n h i b i t i o n of ap- pa ren t pho tosyn thes i s r a t e o f a l f a l f a by combinat ions of s u l f u r d iox ide and n i t r o g e n d ioxide . Env. S c i & Tech. 8:6 574-575
Witkamp, M. & M. Frank. 1969. Loss of weight of Co 60 & C s 137 from t r e e l i t t e r i n t h r e e subsystems of a watershed. Env. S c i & Tech. 3:11 1195- 1198.
Zinke, P. J. 1966. Fo res t i n t e r c e p t i o n s t u d i e s i n t h e United S t a t e s . pp. 137- I61 i n Fo res t Hydrology Proc. o f a Na- t i o n a l Science Foundation advanced Sci- ence Seminar Penn. S t a t e Univ. Penn. Pergamon P res s . N.Y.
Zinke, P. J. 1974. E f f e c t of Herb ic ides on s o i l s of South Vietnam.. P a r t B. Working Papers -The e f f e c t s of h e r b i c i d e s i n South Vietnam. 39pp. Nat iona l Ac. of S c i . , N a t . Res. Council . Wash. D.C.
Zinke, P . J . , A. S tangenberger , W. Colwell . 1979. The f e r t i l i t y of t h e f o r e s t . C a l i f o r n i a A g r i c u l t u r e 33: 5 1 0-1 1 .

Forest Genetics and Air Pollutant stress'
2Stefan Bialobok
Abstract : The breeding of t r e e s , p a r t i c u l a r l y c o n i f e r s , f o r c u l t i v a t i o n in regions under t h e in f luence of indus t ry i s of considerable economic importance. The r o l e of gene t ic polymorphism in t h e process of adap ta t ion t o such environ- ments i s discussed. Resu l t s a r e presented of research on t h e g e n e t i c v a r i a b i l i t y in Scots pine populat ions growing under t h e in f luence of indus t ry and i n regions f r e e of gaseous po l lu t ion . Methods of s e l e c t i n g t r e e populat ions t o l e r a n t t o i n d u s t r i a l emissions a r e discussed aimed a t t h e i d e n t i f i c a t i o n of gene t ic markers of to le rance .
I n o r d e r t o reduce l o s s e s in f o r e s t ecosystems a s wel l a s i n some spec ies of broadleaf trees caused by i n d u s t r i a l emissions a t tempts were made (Gerhold et al , . , 1977). The g r e a t e r to le rance of t o determine t h e s e n s i t i v i t y of trees (depending Scots pine and Norway spruce t o t h e a i r p o l l u t i o n on t h e i r g e n e t i c p r o p e r t i e s ) t o acu te i n j u r y of within a population i s sometimes assoc ia ted with t h e a s s i m i l a t i o n apparatus by t h e most common t h e g r e a t e r r e s i s t a n c e t o low temperatures d i seases gaseous p o l l u t a n t s . The degree of p l a n t i n j u r y a i d a xerothermic adap ta t ion (Bialobok, 1979). A by gases i s dependent a l s o on t h e system of suggestion a l s o e x i s t s t h a t t h e cause of t o l e r -eco log ica l f a c t o r s opera t ing in t h e environment. ance t o some gases may depend on mechanisms t h a t
reduce t h e access of gases t o t h e p l a n t organism GENETIC VARIATION OF TOLERANCE BETWEEN AND WITH- (Gerhold et a l . , 1977). I N SPECIES
However, i t has been experimentally e s t a b l i s h e d So f a r a wide s c a l e of g e n e t i c v a r i a b i l i t y was o r deduced from r e l i a b l e evidence t h a t t h e r e
observed wi th in var ious spec ies of t r e e s and e x i s t g e n e t i c mechanisms condi t ioning higher shrubs i n t h e i r s e n s i t i v i t y t o a i rborne po l lu t ion . to le rance t o i n d u s t r i a l emissions (Niemtur, 1979). The r e s u l t s l i s t e d i n reviews (Davis et a l . , 1976, The l a t t e r i n v e s t i g a t o r s tud ied 58 progenies of Bialobok 1979) a r e only approximate. It appears Scots pine from 4 provenances obtained from seeds t h a t most t o l e r a n t a r e some spec ies of broadleaf c o l l e c t e d on remnant l i v i n g t r e e s in t h e h igh ly trees. Coniferous t r e e s which a r e of g r e a t e r po l lu ted condi t ions of t h e Upper S i l e s i a n indus-economic importance a r e genera l ly charac te r i sed t r i a l region. He has shown t h a t these progenies by low to le rance . have a g r e a t e r to le rance than those c o l l e c t e d
from t r e e s growing beyond t h e range of p o l l u t i o n . It was a l s o found t h a t t h e r e is g e n e t i c
v a r i a b i l i t y in response t o SO^, O3 and HF POLYMORPHISM AND ITS ROLE wi th in one populat ion of Pinus s y l v e s t r i s , P. s t robus , P. ponderosa and within Picea a b i e s S tud ies on t h e g e n e t i c v a r i a b i l i t y of enzymes
in f o r e s t t r e e s developed s t r o n g l y a f t e r S t e m , (1974) pointed o u t t h e g r e a t importance of gen-
p r e s e n t e d a t t h e Symposium on E f f e c t s of A i r e t i c polymorphism f o r t h e development of ecolog-
P o l l u t a n t s on Mediterranean and Temperate Fores t i c a l g e n e t i c s , f o r e s t gene t ics and t r e e breeding.
Ecosystems, June 22-27, 1980, Rivers ide, Cal i for-n i a , U.S.A. Thanks t o t h i s method i t i s poss ib le t o
recognize t h a t g e n e t i c s t r u c t u r e of a populat ion
2 ~ r e eB i o l o g i s t , I n s t i t u t e of Dendrology, P o l i s h ( a l l e l e f requencies and degree of heterozygosi ty)
Academy of Sciences , 63-120 Kornik, Poland. in space and time, t o conduct s t u d i e s on i n h e r i t -ance and l inkage , t o determine t h e gene t ic d i s t a n c e

between ind iv idua l s and populat ions , t o i d e n t i f y mechanisms of adap ta t ion t o an environment and t o employ i t in p r a c t i c a l f o r e s t r y .
Polymorphism of f o r e s t trees which a r e long- l i v i n g organisms r e p r e s e n t s adapt ive s t r a t e g i e s of populat ions t o a changing environment (Stern e t a l . , 1974). In t h i s gene t ic system hetero- zygotes a r e favored and thus t h e population is capable of e x i s t i n g i n a heterogenic environment surviving numerous and major changes.
, Tree populat ions in i n d u s t r i a l regions grow under s i m i l a r condi t ions a s t h e herbaceous p l a n t s t h a t co lon ize m i n e s p i l l s having high concen t ra t ions of heavy elements (Pb, Zn, Cu). This phenomenon S t e m (1974) a l s o cons iders a s being a good example of t h e developing poly- morphism.
I n condi t ions where a t l e a s t a p a r t of t h e t r e e s can surv ive i n an i n d u s t r i a l environment t h a t has n o t been too s t r o n g l y po l lu ted , the populat ion adopts t h e optimal s t r a t e g y i n reac-t i o n t o t h e changes t ak ing place. This, of course , i s t h e r e s u l t of t h e evolut ionary adap ta t ion process t o t h e new environmental condi t ions where t h e g e n e t i c system i s under a cons tan t s e l e c t i o n pressure . Tolerant geno- types appear t o have a c e r t a i n l e v e l of f i t n e s s t o t h e i n d u s t r i a l environment. What is most important t o us i s t h a t t h e d r a s t i c changes in the i n d u s t r i a l environment should n o t exceed the l e v e l of g e n e t i c f l e x i b i l i t y of t h e popula- t i o n s of t h e most important f o r e s t forming spec ies . I n t h e acute" condi t ions of change i n an i n d u s t r i a l environment, t h e coded gene t ic system wi th in a populat ion, t h a t condi t ions i t s adapt ive capac i ty , gradual ly ceases t o opera te s i n c e t h e p o t e n t i a l of ind iv idua l genotypes f o r adap ta t ion has been surpassed.
GENETIC VARIABILITY OF ENZYMES I N SCOTS PINE POPULATIONS
The use of e l e c t r o p h o r e t i c a n a l y s i s of enzymes f o r t h e i d e n t i f i c a t i o n of v a r i a b i l i t y determining g e n e t i c to le rance t o i n d u s t r i a l p o l l u t i o n i s so f a r only i n an in t roduc tory phase. The r e s u l t s obtained so f a r a r e encouraging. The s t u d i e s conducted on Scots p ine a r e t r e a t e d only a s a model.
Mejnartowicz (1978) has s tud ied t h e var ia - b i l i t y of izozymes of leucyloaminopeptidase (LAP) and a c i d phosphotase (APH) in 19 populat ions of Sco ts p ine from t h e whole range of t h e spec ies in Eurasia . The Poland populat ions were chosen fromregions under i n d u s t r i a l in f luence and from reg ions beyond p o l l u t i o n . I n a l l t h e populat ions APH was coded in locus B wi th 15 a l l e l e s , and t h e r e was considerable polymorphism in t h e s t u d i e d populat ions , and LAP was coded in 2 l o c i , LAP-A having 5 a l l e l e s and LAP-B 6 a l l e l e s .
It i s i n t e r e s t i n g t h a t t h e population of Scots pine from Babki ( c e n t r a l Poland--strong indus-t r i a l po l lu t ion) has had t h e lowest degree of heterozygosi ty on locus LAP-A, ha0.1124 and t h e h ighes t va lue i n LAP-B, h=0.3970. Among individ- u a l s s e n s i t i v e t o SO2 t h e genotype APH-B4/B6 dominated and among ind iv idua l s t o l e r a n t t o t h i s gas t h e a l l e l e APH-B5 appeared commonly (Mejnartowicz, 1977-78). Szmidt (1978) when studying t h e gene t ic polymorphism of c a t a l a s e i n t h r e e populat ions of Scots pine from i n d u s t r i a l regions and 5 populat ions from regions f r e e of p o l l u t i o n , has found 7 phenotypes of t h e ca ta lase . When comparing t h e populat ions on t h e b a s i s of heterozygosi ty he has shown t h a t popula- t i o n s from regions f r e e of p o l l u t i o n were charac-t e r i z e d by g r e a t e r p a r t i c i p a t i o n of homomorphic ind iv idua l s of t h e type C-l /C-1 compared t o populat ions under t h e in f luence of emissions.
Mejnartowicz, e t a l . , (1978) having taken m a t e r i a l from p l u s t r e e s in a Scots p ine seed orchard e s t a b l i s h e d a l a c k of c o r r e l a t i o n between t h e degree of heterozygosi ty of t r e e s i n LAP l o c i and t h e degree of t h e i r s e n s i t i v i t y t o t h e a c t i o n of SO2 in a l abora to ry t e s t .
These in t roduc tory r e s u l t s i n d i c a t e t h a t t h e r e i s d i f f e r e n t i a t i o n in t h e frequency of some a l l e l e s in a populat ion depending on t h e inclu- ence of i n d u s t r i a l emissions. The important th ing i s t h a t an enzymatic marker b e found f o r to le rance which would be independent of environ-mental in luences . INHERITANCE OF TOLERANCE TO INDUSTRIAL EMISSIONS
Not many i n v e s t i g a t o r s have attempted determin- ing in genera t ive progeny t h e combining a b i l i t y , h e r i t a b i l i t y and r e p e a t a b i l i t y of to le rance t o i n d u s t r i a l emissions. I n t h e case of Scots p ine h e r i t a b i l i t y and r e p e a t a b i l i t y of to le rance t o Oq is t o be found under a low o r medium gene t ic c o n t r o l . However, some provenances have a h igher h e r i t a b i l i t y of to le rance t o 03. Needle i n j u r y in these pines was p o s i t i v e l y c o r r e l a t e d wi th t h e i n j u r i e s caused by SO2 (Gerhold e t a l . , 1977, Demeritt, 1977). Bialobok e t a l . , (1978) have a l s o found a s i g n i f i c a n t c o r r e l a t i o n between t h e degree of needle i n j u r y on mother t r e e s and on t h e i r progenies following treatment wi th SO?. However, t h e s e c o r r e l a t i o n s were n o t found follow- ing treatment with 03 o r a mixture of 03 and SO2.
The s e l e c t i o n of t r e e ind iv idua l s t o l e r a n t t o i n d u s t r i a l emissions i s being done by t h e r e l a - t i v e l y simple methods of mass s e l e c t i o n , without understanding t h e gene t ic b a s i s f o r t h e popula- t i o n s t r u c t u r e . This s e l e c t i o n i s usua l ly being conducted on seed l ings in labora to ry condi t ions . Sometimes p o s i t i v e r e s u l t s were obtained f o r t h e p r a c t i c e which with some p r o b a b i l i t y a r e being u t i l i z e d in e s t a b l i s h i n g tree p l a n t a t i o n s o r producing t r e e s f o r ornamental purposes (Demeritt , 1977).

So f a r i t was n o t poss ib le t o so lve t h e prob- l e m of breeding var ious pine spec ies f o r cu l t iva - t i o n in f o r e s t s under t h e in f luence of i n d u s t r i a l emissions, and t h e r e i s no cheap way of propaga-t i o n of s e l e c t e d ind iv idua l s .
Gerhold, (1977) has discussed s e v e r a l methods of breeding trees more t o l e r a n t t o i n d u s t r i a l emissions , i n which t h e use of seeds orchards e s t a b l i s h e d from t o l e r a n t c lones i s envisaged.
It appears, however, t h a t it would be va luab le t o expand t h e i n v e s t i g a t i o n s of gene t ic va r ia - b i l i t y of enzymes in populat ions of coniferous trees endangered by i n d u s t r i a l po l lu t ion . W e have considerable information a l ready on t h e course of metabol ic c y c l e s in t h e tree organism exposed t o t h e a c t i o n of some gases. I b e l i e v e these two sources of information could be combined, (biochemical gene t ics and metabolic changes) in order t o i d e n t i f y t h e enzymatic gene t ic marker t h a t would be use fu l i n t h e s e l e c t i o n of t o l e r a n t populations.
LITERATURE CITED
Bialobok, Stefan. 1979. I d e n t i f i c a t i o n of r e s i s t a n t o r t o l e r a n t
s t r a i n s and a r t i f i c i a l s e l e c t i o n o r produc- t i o n of such s t r a i n s in order t o p r o j e c t vege ta t ion from a i r po l lu t ion . Symp. on t h e e f f e c t s of a i r b o r n e p o l l u t i o n on vege ta t ion , Warsaw (Poland) 20-24 Aug. 1-16, 1979.
Demeritt, Maurice E. JK. 1977. Genetic Evaluation of Two-Year Height
and Ozone Tolerance in Scotch Pine (Pinus s y l v e s t r i s L.) . A Thesis i n Genetics. The Pennsylvania S t a t e Univers i ty .
Davis, Donald D. and Raymond G. Wilhour. 1976. S u s c e p t i b i l i t y of woody p l a n t s t o su l -
f u r d iox ide and photochemical oxidants . EPA-600/3-76-102. p. 72. U. S. Environmental Pro tec t ion Agency, C o r v a l l i s , Oregon 97330.
Gerhold, Henry G. , and Raymond G. Wilhour. 1977. E f f e c t of a i r p o l l u t i o n on Pinus
s t r o b u s L. and g e n e t i c r e s i s t a n c e . A Li te ra -t u r e Review. EPA-600/3-77-002 p. 45 Environ- mental P r o t e c t i o n Agency, Corva l l i s , Oregon, 97330.
Mejnartowicz, Leon. 1977-1978. Genetic c h a r a c t e r i s t i c of some
Scots pine t r e e s suscep t ib le or somewhat r e s i s t a n t t o t h e a c t i o n of SO .̂ Annual Report from P r o j e c t PL-Fs-74 from U. S. Department of Agricul ture under PL-480.
Mejnartowicz, Leon. 1978. S t ruk tura genetyczna populac j i sosny
zwyczajnej z terenow przemyslowych zanieczy- szczonych przez S02. In Proc. from: Reakcje biologiczne drzew na e m i s je przemyslowe. Kornik 4-5 maj 1978.
Mejnartowicz, L., S. Bialobok and P. Karolewski.
1978. Genetic c h a r a c t e r i s t i c of Sco ts p ine specimens r e s i s t a n t and s u s c e p t i b l e t o SO2 ac t ion . Arboretum Komickie . 23: 233-238.
Mejnartowicz, Leon. 1979. Genetic v a r i a t i o n i n some isoenzyme
l o c i i n Scots p ine (Pinus s i l v e s t r i s L.) population. Arboretum Komickie . 24: 91-104.
Niemtur, S tanislaw. 1979. Badania nad wewnatrzgatunkowym
zroznicowaniem odpomosci sosny zwy- c z a j n e j (Pinus s i l v e s t r i s L.) na emisje przemyslowe. Doctoral Thesis , Uniwersytet &ski , Katowice.
S t e m , K. and P e t e r M. A. T i g e r s t e t . 1974. Okologische Genetic. 98-p. 75 p. G.
Fischer. S t u t t g a r t .
Szmidt, Alfred. 1978. Zmiennosc ka ta lazy w populacjach
sosny zwyczajnej znajdujacych s i e pod wplywem zanieczyszczen przemyslowych. I n Proc. from: Reakcje biologiczne drzew na emisje przemyslowe. Komik 4-5 maj 1978.

Interactions of Air Pollutants and Plant
Michael reshow^
Abstract: Each in te rac t ion between an a i r pollu-t a n t and disease is unique. Pol lutants may m i t i -gate disease response or in tens i fy it; and the presence of ce r t a in diseases can modify the p lan ts response t o a pol lutant . Air po l lu tan ts can a c t d i r e c t l y on a fungus or bac te r ia inh ib i t ing parasi- t i s m . They may a l so ac t most s t rongly on the hos t , modifying i t s physiology and rendering it e i t h e r more or l e s s sens i t ive t o a plant pathogen. Where t he plant is weakened by the po l lu tan ts , it tends t o be more s ens i t i ve t o weak pathogens, but l e s s s ens i t i ve t o obl igate parasi tes . Where the pollu-t an t physical ly in jures the host, in fec t ion may be f a c i l i t a t e d . Pol lutant in te rac t ions have been demonstrated both i n the laboratory and the f i e l d a t ambient pol lutant concentrations. In te rac t ions between pol lu tan ts and ab io t i c s t r e s s a r e part icu-l a r l y c r i t i c a l . Water regime, temperature rela-t ions , mineral nu t r i t i on and other parameters of the physical environment play a major ro le i n the expression of a i r pol lut ion injury.
Air po l lu t ion can k i l l plants . Even when the concentrations of an a i r po l lu tan t a re not d i r e c t l y l e t h a l , they may be harmful, adversely a f f e c t i n g growth, reproduction and myriads of other b io log ica l processes (Mil ler and McBride 1975). One such process is the i n t e r ac t i on t ha t r e s u l t s between the po l lu tan t and plant pathogens.
There a r e many ways i n which such i n t e r -ac t ions might take place. But do they? The most obvious would be i f the pol lutant d i r ec t l y a f fec ted the fungus, v i rus , bac te r ia ,
p r e s e n t e d a t t he Symposium on Effec ts of A i r Po l l u t an t s on Mediterranean and Temperate Forest Ecosystems, June 22-27, 1980, Riverside, Ca l i forn ia , U.S.A.
2 ~ r o f e s s o r of Biology, University of Utah, S a l t Lake City, Utah 84112.
i n sec t or other pathogen. I f some stage of an organism's l i f e cycle , o r some s t age of paras i t -i s m , were adversely impaired by a po l lu tan t , the pol lutant e s sen t i a l l y would be ac t ing as a fungicide, bacter iocide or other biocide and could be expected t o bring about some degree of control.
The second way an a i r pol lutant might a c t would be i n adversely a f f ec t i ng the growth or reproduction of the host plant and thereby influencing i t s s u i t a b i l i t y as a host. There a r e two ways i n which such act ion could be brought about. Most apparently, the po l lu tan t might cause some physical in jury t o the plant leaving les ions t ha t could serve a s i n f ec t i on cour t s for a fungus or bacteria. O r , the plant might simply be weakened, thus lowering i t s res i s tance t o a prospective pathogen. Presum-ably the weaker pathogens, the f acu l t a t i ve pa ra s i t e s , would benef i t most by such

--
conditions. Obligate parasites, on the other hand, preferring a vigorous host, might respond quite differently and their development might even be discouraged in the weakened host.
Conversely, the impact of various diseases or plant stresses on the plant's response to air pollutants has also been considered. It should be quite apparent that adverse climatic or edaphic factors might affect a plant's sensi- tivity to a pollutant, but the presence of certain biotic diseases may also increase or decrease a plant's sensitivity.
A major question is not so much do such interactions take place, but what concentrations of an air pollutant are required to precipitate such effects, and how widely do such concentra- tions occur?
Many of the interactions that will be dis- cussed have been demonstrated under laboratory conditions, often at pollutant concentrations known to be fairly widespread. Some of the interactions have been observed in the field.
The literature treating such air pollutant- plant pathogen interactions has been reviewed most recently in 1978 (Laurence), 1975 (Treshow), and 1973 (Heagle); it is not the intent of this paper to once again provide an exhaustive review of this literature. Rather, I should like to first bring the literature up to date, briefly including that delving into the interactions of abiotic stresses on pollutant effects, and discuss the implication and pos- sible practical relevance of such interactions, especially concerning forest ecosystems. Pri-marily I should like to develop some of the principles in pollutant-pathogen interactions.
INFLUENCE OF POLLUTANTS ON DISEASE EXPRESSION
Sulfur Dioxide
There is no question that sulfur dioxide interacts with fungi, suppressing their growth and acting fungicidally. The only question concerns the SO2 concentrations at which such interactions occur.
A few examples exist where SO2 has acted in such a way in the field, limiting the develop- ment of certain plant diseases. The examples provided by Scheffer and Hedgcock (1955) in which parasitic fungi generally appeared to be retarded or inhibited near smelters where SO2 was present is most often cited (Treshow 1975). Suppressed disease development was especially apparent where rust fungi, Melampsora and Puccinia, or needle cast fungi were involved. Linzon (1973) also discusses the reduced heart rot and blister rust (Cronartium ribicola) near a smelter.
But such incidents are becoming more infre- quent as ground level SO2 concentrations become reduced by modern control technology or use of tall stacks. Higher SO2 concentrations, often exceeding 1 ppm for extended periods, now tend to be of only historic record. We are now interested in the possible impact of SO2 concen- trations in the 0.1 to 0.5 ppm range or even lower in cases of prolonged duration. Where such concentrations still exist near smelters, power plants, or large urban centers, some pollutant-pathogen interaction might be sought.
There is some evidence from laboratory and <
field research that certain pathogenic fungi respond to SO2 in this concentration range. Weinstein and others (1975) for instance, have demonstrated that SO2 concentrations of 0.14 to 0.15 ppm, continuous for 7 days, while not visibly injuring plant foliage, affected the bean rust fungus, Uromyces. The incidence and severity of the rust as measured by the abun- dance of uredia was decreased as were both the size and percent germination of the uredo- spores. Hence the reproductive potential of the pathogen was reduced. The results suggested that the effect on the pathogen was indirect; perhaps involving an 802-induced chemical change in the resistance of the host and its suitabil- ity as a habitat for the fungus. The authors speculated that production of phytoalexins or supply of metabolites might have been altered, or compounds derived from SO2 might have accumu- lated.
One of the most comprehensive field studies of the interactions of industrial air pollu- tants, especially SO2, with plant disease was conducted by A. Grzywacz and J. Wazny (1973). The study was notable since it dealt with eco- nomically significant diseases of a coniferous forest, specifically Scots pine (Pinus sylvestris L.). Interactions with six fungi were considered: Armillaria mellea, Fomes annosus, Lophodermium pinastri, Microsphaera alphitoides, Melampsora pinitorqua, and Cronartium flaccidium.
In an overall study, comparisons were made of disease presence in industrial areas and forests throughout Poland. In a more detailed approach, disease presence was compared at increasing distances from an industrial town, Torun, with one major SO2 source.
The general comparisons showed that Armillaria was present in 3.7 percent of the trees in the area damaged by SO2 compared with only 1.38 percent in all forests. Lophodermium also tended to be more serious in the industrial areas, but the results were indefinite. This was consistent with earlier findings of Donabauer (1966) that high SO2 concentrations increased the intensity of Armillaria root rot.
Fomes showed the reverse trend and occurred

in 0.87 percent of the areas affected by indus- trial pollution compared with 1.5 percent in the country as a whole. Fomes, as in the overall study, was least prevalent in areas of highest SO2 concentrations. Grzywacz and Wazny (1973) found that stumps close to the SO2 source were sound. Related, decomposition fungi were also less abundant closest to the source. Only 50 percent of the stumps showed signs of decom- posing, and 10 percent strongly decomposing, compared with 30 percent strongly decomposing farther out. However, Gryzwacz (1978) later reported that stands influenced by air pollution were more subject to decomposition by brown, white or soft rot fungi. Sufficiently high SO2 concentrations might suppress the fungal growth directly. Such was the case where Mejstrik (1978) showed SO2 concentrations as low as 84 ug/m3 (.03 ppm) inhibited growth of Flamulina velopes, Nematoloma fasciculare, and Pleurotus ostreatus from 34 to 39 percent.
Lophodermium injury was least significant near the SO2 source. At distances beyond 1 km injury appeared on up to 70 percent of the needles. There was a slight decrease beyond this. When annual SO2 concentrations averaged below 0.2 ppm, disease incidence was essentially the same as in the control group. Fungus devel- opment was noticeably affected. Within 1 km, reproduction was largely by pycnidia; further distant, apothecia were common. The numbers of needles with both the sexual and asexual stages increased with distances. Closest to the SO2 source, apothecia were only 15 percent the length of controls deformed, collapsed, under- developed, closed or even dried up and with ill- developed asci. Chiba and Tanaka (1968) however reported high SO2 concentrations of 2 ppm for 14 hours caused increased infection of injured pine needles (P. densiflora) by Rhizosphoera kalkhoffii.
Weidensaul and Darling (1979) inoculated Scots pine seedlings with the fungus Scirrhia acicola. The plants were inoculated either 5 days before or 30 minutes after fumigation for 6 hours with 533 ug/m3 (0.20 ppm) SO2). After 8 weeks, the seedlings inoculated 5 days before fumigations had more lesions incited by the fungus than those inoculated 30 minutes af ter fumigations in the control group. It was postu- lated that since the needle blight fungus pene- trates through the stoma, the ability of the SO2 to keep the stoma open facilitated entry of the fungus.
The pollutant-disease interaction may vary with the host species. Ham (1971) found that when loblolly pine served as the host, the brown spot fungus (5. acicola) was not measurably affected by SO2 in the 0.5-0.9 ppm range for 2-3 hours even when SO2 caused visible injury.
Simulated rain, acidified with sulfuric acid has also been shown to influence pollutant-
pathogen interactions (Shriver 1974). Although not involving forest species, the work did demonstrate the way in which plant species might be affected.
Kidney beans (Phaseolus vulgaris var. red kidney) and willow oak (Quercus phellus) were exposed to simulated rain acidified to a pH of 6.0 or 3.2 for 10 minutes each day for 30 to 60 days. The acid "rain" caused an 86 percent inhibition in the number of telia produced by Cronartium fusiforme on willow oak, a 66 percent inhibition in the reproduction of the root knot nematode (Meloidogyne hapla) on beans in the field, and a 29 percent decrease in the percent of leaf area of field erown beans affected by "-- -- - - - - - -
Uromyces phaseoli. Exposures to pH 3.2 "rain" completely destroyed the integrity of the cuti- -cular waxes in both oak and bean. Exposures to the pH 6.0 solution did not visibly alter the cutin. Shriver postulated that damage to the cutin facilitated leaching of carbohydrates from the leaf including those that might inhibit hyphal growth of Botrytis cinerea thus predis- posing plants to infection.
This is consistent with the findings of F. A. Last and 0. D. Fowler at the Institute of Terrestrial Ecology at Penicuik, Scotland (per- sonal communication) who showed that the cuticle of Scots pine was gradually broken down by low concentrations, of S02.
Sulfur dioxide, perhaps largely as acid rain, may have an effect on soil microorganisms (Wainwright 1978). Sulfur-oxidizing micro- organisms, including Alternaria tenuis, Auriobasidium pullulans and Cephalosporium sp., were isolated from leaves, litter and soils polluted with high levels of S02. The predomi- nant autotroph on leaf surfaces was Thiobacillin thioparis.
Hibben and Taylor (1975) found that S02- pathogen interaction depended on the infection stage. Conidial gemination and the appres- sorial stages of Microsphaera =were most sensitive to S02. Since infection was not reduced when leaves were previously fumigated, the effect was thought to be mostly fungicidal.
Ozone
There is considerable evidence that photo- chemical pollutants, most notably ozone, affect plant pathogens and fungus-host interactions. The interaction may be positive or negative depending on the 03 concentrations and the 03 sensitivity of the host.
In one such study, Miller and Elderman (1977) showed that fumigation of ponderosa and Jeffrey pine seedlings with ozone enhanced infection by Fomes annosus. Average infection of the two

species increased from 57 percent to 78 percent when the seedlings were exposed to 431.2 pg/~3 (.25 ppm) ozone. They also found that ozone injury increased the susceptibility of pine stumps to colonization by Fomes annosus. In the laboratory, wood from trees slightly damaged by oxidants was more decay-susceptible than wood from severely damaged trees.
Far more research has involved powdery mildew (Heagle and Strickland 1972, Heagle 1975, Scheutte 1971), and the negative impact of ozone on disease. Even ozone concentrations in the 5 to 50 pphn range reduced infection when conidia were exposed. According to Scheutte, appresoria formation was largely affected. Also, some epidermal cells, hypersensitive to ozone, were killed thereby reducing infection. Eight hour exposure to 50 pphm ozone reduced formation of secondary hyphae 66 percent and 4 hour exposure reduced 41 percent. Ozone impaired formation of the penetration peg thus inhibiting subsequent development of a functional host-parasite rela- tionship.
While ozone impacted certain critical stages of infection under controlled conditions, in the field such effects would be less apparent since all stages of parasitism would be present simul- taneously.
Interactions of facultative parasitic and saprophytic fungi with their hosts may also occur and are most pronounced when the plants are weakened or injured by ozone (Manning 1975). Obligate parasitism appears to be retarded by ozone and ozone-injured host tissue. Colonization of ozone-in jured white pine needles by Lophodermium pinastri was reduced while colonization by the saprophyte Aureobasidium pullulans was increased. Ozone-injured lilac leaves were rarely infected by powdery mildew Micosphaera a.
When the fungus is especially sensitive to ozone the impact may be mostly fungicidal there- by reducing infection. Sporulation and germina- tion of the weak parasite Botrytis cinerea was significantly inhibited by 2, 6 hour ozone exposures at a concentration of 30 pphm (Krause and Weidensaul 1978). There was also less infection, based on total lesion area, when plants were exposed to 15 pphm. Ozonation of conidia produced in vivo and in vitro decreased germination of conidia, germ tube length, and pathogenicity. The authors postulated that the mode of action was to alter the permeability of the conidia membrane. Ozone may increase the conidiophore respiration causing prematurely- formed and non-viable conidia. Ozone also may inhibit, directly or indirectly, enzyme activity of the fungus and cause less maceration of the host cells resulting in a decreased infection.
Curtis and others (1976) found that plants exposed to ozone produced peroxidases and iso-
flavonoids that are toxic to microorganisms. According to Laurence and Wood (1978), ozone inhibited infection of soybean by Pseudomonas glycinea presumably because of a bacteriostatic compound, possibly an isoflavinoid compound, produced in response to ozone.
Fluoride
Fluoride is accumulated in the foliage of plants and affects many metabolic processes. Consequently, it is reasonable to presume that pollutant-pathogen interactions can be influenced, perhaps to a greater extent than with any other pollutant. This was confirmed with viruses by some of the early work reviewed by Heagle (1973) and Treshow (1975).
Host-fluoride interactions also have been demonstrated for fungi (McCune and others 1973) although not on forest species. Studying the interactions of 7 to 10 with 3 diseases of bean and 2 diseases of tomato, HF consist- ently reduced uredial formation as the foliar fluoride content increased. Tender green beans exposed to HF and inoculated with powdery mildew conidia developed fewer foliar lesions than non- fumigated control plants; 4.4 compared with 48.7 on fumigated leaves having an average of 399 ppm F. Powdery mildew was reduced proportionate to the length of the exposure period. Early blight lesions on tomato were similarly reduced by the presence of fluoride.
INFLUENCE OF DISEASE ON POLLUTANT SEVERITY
Not only may the presence of a pollutant influence pathogenicity, the reverse may be true~presence of diseases influence the degree of pollutant injury. This has best been demon- strated with viruses. None of these involved forest species though, and there has been no new results published since last reviewed by Laurence (1978).
Also, in the forest, variation in pollutant resistance among individual trees would far exceed any disease and pollutant interaction, and any impact of a disease on pollutant sever- ity probably wouldn't be measurable.
ABIOTIC STRESS
Severity of air pollution injury is deter- mined not only by the dose of the pollutant to which plants are exposed but on the predisposi- tion of the plant. Predisposition is determined by any environmental parameter to which the plant is subjected. It has been discussed more in passing than as a major concern (Heck and others 1965, Heck 1968, Treshow 1970, Taylor 1974), but the interactions of abiotic stress or disorders with any pollutant is far more

significant than with biotic pathogens. Moisture stress, the light regime, edaphic factors and temperature relations all influence the sensitivity of plants to every pollutant.
Unusually high or low temperatures prior to exposure tend to reduce plant sensitivity to ozone, for instance. A normal high light inten- sity renders plants more sensitive. High humid- ity, by encouraging open stomates , also increases sensitivity. In dry air, plants are more tolerant of ozone than at higher humidi- ties, and PAN injury is most severe in the Los Angeles area when the relative humidity exceeds 50 percent.
Any factor favoring moisture stress and reduced water uptake would cause stomatal clo- sure and favor resistance to pollutant uptake and injury. In this way, plants in more saline soils are more tolerant of certain pollutants (Oertli 1958) and there is little to no ozone or PAN injury when soil moisture is deficient.
Ozone injury may be negligible when ambient air temperatures during and after exposure are below 90Â F., yet may be severe when tempera- tures exceed 90' F. (Taylor 1974).
Effects of low atmospheric and soil moisture on tolerance or sensitivity of plants to SO2 injury are well known; suffice it to note that plants are far more sensitive under higher moisture conditions.
Mineral nutrition also plays arole in pollu- tant sensitivity. Vegetable crops are known to be most susceptible to ozone when nitrogen nutrition is adequate although some reports suggest injury is greatest when nitrogen is low. Sulfur dioxide sensitivity is reported to be greatest with increased sulfur nutrition. Stomata1 behavior may be involved since a defic- iency of nitrogen, potassium or phosphor,us may decrease stomatal opening. Plants low in sulfur had the lowest stomatal capacity, thus less capacity for gas absorption (Leone and Brennan 1972).
Toxicity from heavy metals also plays an interesting role with air pollutants. Ormrod (1977) has found that ozone toxicity was en- hanced on pea plants growing in elevated concen- trations of cadmium or nickel. However, when cadmium concentrations were sufficiently high to adversely affect growth,100 umol of CdS04, then ozone injury was less than that of control plants.
Harkov and others (1979) found that the amount of ozone injury depended on the cadmium and ozone levels as well as other environmental conditions. Where environmental conditions were not conducive to ozone in jury (e .g. bright sunlight), cadmium had little effect on the amount of leaf damage. When environmental
conditions allowed only slight to moderate foliar injury on tomato leaves, damage was enhanced by cadmium treatment especially at higher concentrations.
Along the same line, low concentrations of SO2 affected metal uptake, increasing zinc and cadmium susceptibility of bean plants. Foliar injury caused by heavy metals was significantly enhanced by SO2 (Krause and Kaiser 1977).
Lamoreaux and Chaney (1978) also found sig- nificant interactions between cadmium and S02. While cadmium or SO2 alone reduced net photosyn- thesis and transpiration of excised silver maple leaves, the reduction in cadmium-treated leaves was greatest in the presence of S02.
CONCLUSIONS
Virtually every environmental parameter, biotic and abiotic can interact with air pollu- tants to aggravate or mitigate the extent of pollution damage. The significant question then becomes, to what extent and at what concentra- tions? Is the net impact real, and if so, how meaningful is it under field conditions?
All pollutants don't interact the same. Sulfur dioxide, for instance, appears primarily to act fungicidally reducing disease and decom- position activity. It is also postulated though that SO2 induces chemical changes imparting resistance to certain pathogens. Ozone tends to act more on the host, weakening it and accentu- ating disease activity. Research with vegetable crops at low ozone concentrations indicate that pollutant-disease interactions do occur with some diseases and very likely occur in forest stands weakened by photochemical pollutants. Ozone has increased susceptibility to Fomes and rendered trees more susceptible to decay. Each interaction is unique since every fungus has its own sensitivity as does each plant. Thus the potential for interaction is enormous.
Pollutants such as fluoride that accumulate significantly in plants would likely have the greatest interaction. While the trend is to inhibit disease development, the concentrations at which this has been demonstrated are now infrequent in the field.
The significance of abiotic stress and pollu- tant interactions is often taken for granted, but it should be emphasized that this is of paramount importance. Climatic and edaphic factors are often determinants in the expression of pollutant damage. While the abiotic environ- ment strongly influences the severity of air pollution damage, air pollutants influence the severity of plant diseases caused by biotic pathogens. It would appear that as in all life systems, every environmental parameter has some influence on every other. In the case of

pollutants and pathogens, this interaction can be significant.
LITERATURE CITED
Chiba, 0. and K. Tanaka. 1968. The effect of sulfur dioxide on the development of pine needle blight caused by Rhizosphaera kalkhoffii Bubak (I). J. Jap. For. Soc. 50: 135-139.
Curtis, C. R., R. K. Howell and D. F. Kremer. 1976. Soybean peroxidase from ozone injury. Environ. Pollut. 11:189-194.
Donabauer, E. 1966. Secondary damage to forest caused by industrial exhaust fumes. Mitt. Forst. Bundesvers. 73:lOl.
Grzywacz, A. 1978. Interactions between air pollutants and decaying fungi of pine wood. In. 3rd Internat. Congress of Plant Pathology. Munchen. 1978.
Grzywacz, A. and J. Wazny. 1973. The impact of industrial air pollutants on the occurrence of several important pathogenic fungi of forest trees in Poland. Eur. J. For. Path. 3:129-141.
Ham, Donald L. 1971. The biological interactions of sulfur dioxide and Scirrhia acicola in loblolly pine. Ph.D. Thesis, Duke Univ., No. Carol. 75 p.
Harkov, R., B. Clarke and Eileen Brennan. 1979. Cadmium contamination may modify re- sponse of tomato to atmospheric ozone. J. Air Poll. Control Assoc. 29: 1247-1249.
Heagle, A. S. 1973. Interaction between air pollutants and plant parasites. Ann. Rev. Phytopathol. 11:365-388.
Heagle, A. S. 1975. Response of three obligate parasites to ozone. Environ. Pollut. 9:91-95.
Heagle, A. S. and Alice Strickland. 1972. Reaction of Erysiphe graminis f. sp. hordei to low levels of ozone. Phytopathol. 62:1144-1148.
Heck, W. W. 1968. Factors influencing expression of oxidant damage to plants. Ann. Rev. Phytopathol. 6:165-188.
Heck, W. W., J. A. Dunning and I. J. Hindawai. 1965. Interactions of environmental factors on the sensitivity of plants to air pollution. J. Air Poll. Control Assoc. 15:511-515.
Hibben, C. R. and M. P. Taylor. 1975. Ozone and sulfur dioxide effects on the lilac powdery mildew fungus. Environ. Pollut. 9:105-114.
Krause, G. H. M. and H. Kaiser. 1977. Plant response to heavy metals and sulfur dioxide. Environ. Pollut. 12:63-71.
Krause, C. R. and T. C. Weidensaul. 1978. Effects of ozone on the sporulation, germination, and pathogenicity of Botrytis cinerea. Phytopathol. 68:195-198.
Lamoreaux, R. J. and W. R. Chaney. 1978. Photosynthesis and transpiration of excised silver maple leaves exposed to cadmium and sulphur dioxide. Environ. pollut. 17:259-268.
Laurence, J. A. 1978. Effects of air pollutants on plant- pathogen inteactins. 71st Ann. Mtg. Air Poll. ControlAssoc. Houston, Texas. 1978.
Laurence, J. A. and F. A. Wood. 1978. ~f fects of ozone on infection of soy bean by Pseudomonas glycinea. Phytopathol. 68:441-445.
Leone, I. A. and Eileen Brennan. 1972. Sulfur nutrition as it contributes to the susceptibility of tobacco and tomato to SO2 injury. Atmos. Environ. 6:259-266.
Linzon, S. N. 1973. The effects of air pollution on forests. Pap. 4th It. Chem. Cong. Conf. 1973 pp. 1-18.
Manning, W. J. 1975. Interactions between air pollutants and fungal, bacterial and viral plant pathogens. Environ. Pollut. 9: 87-90.
McCune, D. C., L. H. Weinstein, J. F. Mancini, and P. van Leuken. 1973. Effects of hydrogen fluoride on plant- pathogen interactions. Internat. Clean Air Congress, Dusseldorf.
Mejstrik, V. 1978. Influence of sulfur dioxide on the growth of some fungus species. Zbl. Bakt. I1 Abt. 133: 464-467.
Miller, Paul R., and Joe R. McBride 1975. Effects of air pollutants on forests. In Responses of plants to air pollution. J.-Brian Mudd and T. T. Kozlowski, eds. p. 192- 235. Academic Press, New York.

Miller, Paul, R. and M. J. Elderman. 1977. Photochemical oxidant air pollution effects on a mixed conifer ecosystem. Ecological Research Series EPA 60013-77-104.
Oertli, J. J. 1958. Effect of salinity on susceptibility of sunflower plants to smog. Soil Sci. 87:249- 251.
Ormrod, D. P. 1977. Cadmium and nickel effects on growth and ozone sensitivity of pea. Water, Air and Soil Pollut. 8:263-270.
Saunders, P. J. W. 1971. & Ecology of Leaf Surface Microorganisms, pp. 81-89, T. F. Preece and C. H. Dickinson. Academic Press, London.
Scheffer, T. C., and G. G. Hedgcock. 1955. Injury to northwestern forest trees by sulfur dioxide from smelters. U.S. Forest Serv. Tech. Bull. 1117, 1-49.
Scheutte, L. R. 1971. Response of the primary infection process of Erysiphe graminis f. sp. hordei to ozone. Ph.D. Thesis, Univ. Utah, Salt Lake City, 71 p.
Shriver, D. S. 1974. Effects of simulated rain acidified with sulfuric acid on host-parasite inter- actions. Ph.D. Thesis, N. Carolina State, Raleigh, N.C.
Taylor, 0. C. 1974. Air pollution effects influenced by plant-environmental interaction. Amer. Chem. Soc. Sympt. Ser. 3:l-7.
Treshow, M. 1970. Environment and plant response. 422 p. McGraw-Hill Co., New York.
Treshow, M. 1975. Interactions of Air Pollutants and Plant Disease. InResponses of plants to air pollution. J. Brian Mudd and T. T. Kozlowski, eds. p. 307-334. Academic Press, New York.
Wainwright, M. 1978. Sulphur-oxidizing microorganisms on vegetation and in soils exposed to atmos- pheric pollution. Environ. Pollut. 17:167- 174.
Weidensaul, T. C. and S. L. Darling. 1979. Effects of ozone and sulfur dioxide on the host-pathogen relationship of Scotch pine and Scirrhia acicola. Phytopathol. 69:939-941.
Weinstein, L. H., D. C. McCune, A. L. Aluisio and P. van Leuken. 1975. The effect of sulphur dioxide on the incidence and severity of bean rust and early blight of tomato. Environ. Pollut. 9:145-155.

Studies on Relationship Between Air Pollutants and Microorganisms in
Japan1
Kiyoshi ~ a n a k a ~
Abstract: A review of the literature on the interaction of air pollutants with parasitic and epiphytic microorganisms is pre- sented with a brief outline of air pollution problems in Japan. Some fungi such as Cercospora sequoiae, Pucciniastrum styracinum, Puccinia kusanoi, Melampsora coleosporioides, Trichoroma matsutake, and some epiphytic bacteria disappeared in the areas affected by air pollutants, and Rhizosphaera kalkhoffii increased its activity due to SO2 in the air.
Fungi, bacteria, viruses and nematodes, as well as insects are all responsible for bringing a cer- tain species of higher plants into equilibrium with its environment (Treshow 1968). The effect of air pollutants on population trends of various micro- organisms, therefore, is extremely important in all ecological systems. Further, air pollutants may affect plant disease development in different ways. Pathogenicity may be influenced through a direct effect of the pollutants on the parasite, or the effects may be indirect through pollution-induced changes in physiology of the host plant eagle 1973).
Prior to 19601s, the interaction of microorgan- isms with pollutants in the air had received limit- ed attention in Japan. But during the past decades, a series of experiments has been conducted in order to explain the effect of air pollution on dis-ease development, and only recently, some attempts have been made to understand the change of popula- tions of microorganisms for use as biological indi- cators of air pollution.
From the beginning of recognition of the signifi- cant and sometimes devastating effects of air pol- lutants on vegetation, plant pathologists have been
presented at the Symposium on Effects of Air Pollutants on Mediterranean and Temperate Forest Ecosystems, June 22-27, 1980, Riverside, California, U.S.A.
2~orest Pathologist , Kansai Branch, Forestry and Forest Products Research Institute, Momoyama, Fushimi, Kyoto 612 Japan.
confused with the similarities between the symptoms of many types of pollution damage and those of in- fectious diseases sometimes leading to incorrect diagnoses. Fortunately, each of the common air pollutants produces a characteristic injury pattern, and the sensitivity of many species differs suffi- ciently for the trained eye to determine the caus- ative agent dams and Salzbach 1961) .
This paper will review the literature on: 1)the modification of parasitism and the population changes of epiphytic microorganisms induced by air pollutants; 2) attempts to make reliable guides in distinguishing among symptoms caused by air pollu- tants and parasites, and those occasionally pro- duced by saprophyfcic fungi. These subjects are considered in conjunction with a brief outline of air pollution effects on forest ecosystems in Japan
SULFUR DIOXIDE
In Japan, towards the end of 19th century, air pollution was reported to be harmful to forests around ore smelters that emitted large amounts of SO2. Plume behavior resulted in pollutants disper- sion in the surrounding area, killing all the veg- etation (mainly Cryptomeria japonica D.Don and Pinus densiflora Sieb. et Zucc.) and causing ero- sion of topsoils following the death of the plants (Doi 1919). The concentration of SO2 of the smoke was reported to be 1 ppm at 1 mile from a large ore smelter located in Ashio, Tochigi Prefecture, and occasionally reached as high as 8 to 10 ppm at the ground level (Doi 1919 ) .

Raising the height of stacks in order to reduce the concentration of SO at the ground level was
2the first and primitive control measure, new tech- nology involves the installation of sulfur recov- ery devices. In 1915, one ore smelter in Hitachi, Ibaraki Prefecture, built a new stack on a hill rising about 350 m above sea level; the stack was nearly 170 m high, so that the smoke was discharged at an elevation of approximately 520 m. Although smelter smoke is released from high stacks, it tends to diffuse rather slowly, and the SO., there-fore, reaches the ground in harmful concentrations even after a considerable length of time and dis- tance. Observations made three years after the replacement of stacks revealed the fact that the severity of damage in the area near to the smelter was markedly reduced,while the total damaged area increased as large as 24,000 ha ( ~ o i 1919).
The reduction of the concentration of S O in the area in the vicinity of the ore smelter on account of the change of stack height was good enough to resume the seedling production of Cryptomeria Japonica in nurseries. Kaburagi (1930) was im- pressed with the absence of a needle blight of Cryptomeria ,japonica caused by Cercospora sequoiae Ell. et Ev. in the nurseries located in the recov- ered area close to the higher stack. The needle blight is one of the most destructive diseases in forest nurseries producing Cryptomeria seedlings, and has spread rapidly throughout Japan except Ho- kkaido. The causal fungus was introduced from North America in the end of 19th century, and Iba- raki Prefecture including the ore smelter is con- sidered to be the epicenter of the introduced dis- ease (Ito 1976). Kaburagi (1930) suggested that a strong correlation existed between SO in the air 2and the absence of the disease. This is believed to be the first report on the effect of air pollu- tion on diseases caused by parasites in Japan. Ka- buragi's observation is strongly supported by the following hypotheses that: 1)SO
? might act direct-
ly upon the fungus on the surfaces of host plants; and 2) the reduction of disease severity resulted in a decrease of inoculum for new infection.
Some of the acute type of damage by the emis- sions from smelters still continues in areas near to sources. In the 19601s, widespread SO, air pol- lution damage to ornamental trees had become appa- rent to the public; this increase was due to urban- ization and industrialization. The one most appa- rent episode in and around Tokyo was the stunting and death of a large number of Zelkova serrata Mak., widely used as shade trees throughout Japan (yambe 1973).
With a considerable increase in public awareness of the chronic type of pollution in many parts, automatic SO 2 recorders were set up by the national and the prefectural agencies to determine the con- centrations, frequency, and the duration of atmo- spheric SO exposures. The number of measuring locations $ad increased from the beginning at the 15 strategic locations in 1965 to as many as 1473 locations in 1979, forming an extensive sampling system for SO2 (The Japanese Environmental Agency
The analysis of air provides a framework of knowledge, and a foundation for developing air quality management strategies, and for prompting the enactment of the Clean Air Act in 1967, and its amendments made in 1970.
Figure 1.Mean value of annual concentrations at 15 strategic locations.
Figure 1 shows annual changes in average concen- trations of SO
2 at 15 strategic locations between
1965 and 1977, the concentration increased until 1967, and then declined gradually. The decline has proved the continuing effectiveness of regula- tion of air pollution sources by the national and the prefectural governments based on the act.
During the summers in the middle of 19601s, the worst average concentrations of SO 2 were experienc- ed. In this period, an unrecorded needle blight of Japanese red pine, Pinus densiflora, occurred throughout the normal range of that species, pre- dominantly in central Japan. Up to that time the disease had been little known and of little concern to forest pathologists. A specific fungus domi- nantly found on blighted needles was investigated by Kobayashi (1967), and he identified it as Rhizo- sphaera kalkhoffii Bubak which had been known as a causal fungus of a needle blight of spruce and fir in northern Europe and North America.
The results of inoculation experiments to Japa- nese red pine seedlings proved that the pathogen- icity of the causal fungus was considerably weak for pines, and the disease would scarcely occur if pines grew under normal conditions anak aka and Chi- ba 1971). The unusual incidence of the disease occurring in the early s m e r of 1.965 might be re- lated to abnormal weather conditions--extremely little rain fell in early spring followed by heavy rain in May (Chiba and Tanaka 1968). Although the disease has not occurred so widely as in 1965, severe damage has been observed in the vicinity of ore smelters or industrial areas. In addition, Japanese red pine is highly sensitive to S O (Ino- ue 1973), and the distinction between the needle blight symptoms caused by the disease and those caused by SO is exceedingly difficult (Tanaka 1975, Tanita 1976)? Theref ore exhaustive experiments were conducted to examine the relationship of SO,,
c-with disease development.

A t o t a l of 163 two-year-old seedlings of Japan- ese red pine were exposed t o 2.0 ppm SO
2 f o r 1,2,3, and 4 hours i n a fumigation chamber before o r a f t e r inoculation i n t h e middle of Ju ly , with a water suspension of conidia of t h e fungus produced on PDA p la t e s .
Damage was more severe i n t he treatment combin-ing t h e fumigation with inoculation, especial ly inoculat ion before fumigation, compared with those i n t h e treatment of fumigation without inoculation. Numerous pycnidia of t h e fungus were produced i n l a rge l e s ions on needles t r ea t ed by fumigation a f t e r inoculat ion a s well a s fumigation f o r longer duration before inoculat ion, no obvious change i n t he appearance of needles was detected on t h e seed- l i ngs inoculated with t h e fungus without fumigation. Symptoms produced on t h e SO t r ea t ed needles, name-
2l y , l a rge les ions with numerous pycnidia, were i den t i ca l t o symptoms of t h e needle 'blight occur-r ing i n t h e f i e l d ( ~ h i b aand Tanaka 1968).
Most pine seedlings displayed acute symptoms of in jury following exposure t o 2.0 ppm SO f o r 2 hours. The SO concentration used i n this study
2 was several times higher than t h a t found i n t h e ambient a i r . It was necessary t o elucidate t he re la t ionship between SO
2 and disease development a t
lower concentrations because such conditions a r e frequently encountered i n r u r a l and fo re s t regions. Therefore, three-year- o ld seedlings of Japanese red pine were exposed f o r 2 weeks i n chambers t o 0.2 ppm SO t h e l e v e l of SO i s l i k e l y t o be found
2i n t h e ambient a i r (Tanaka 1576a).
In t h i s experiment, spec ia l emphasis was placed on t h e e f f ec t of SO fumigation a f t e r inoculation i n comparison with $he treatment before inoculation. The t r ea t ed needles with SO2 fumigation a f t e r inocu- l a t i o n were more severely affected by the fungus, compared with those fumigated by SO gas before inoculat ion. This r e s u l t i s consis tent with find- ings of t h e 2.0 ppm SO
2 fumigation (Chiba and Tana-
ka 1968). S O in jury appeared t o f a c i l i t a t e t he spread and reproduction of t h e fungus within tis-sues of needles r a the r than t o increase t h e i n i t i a l r a t e of in fec t ion by t h e fungus.
To fu r the r examine these findings from a r t i f i - c i a l SO exposure of seedlings i n chambers, f i e l d
2experiments were planned and conducted during the summers of 1972 and 1973 by using a t ransplant ing method anak aka e t a l . 1974a) and a f i l t e r e d a i r method (Tanaka e t a l . 1974b) i n Hiroshima Prefec- t u r e . In t h e t ransplan t ing study, two experimental p lo t s were establ ished. One was i n Fukuyama c i t y close t o a l a rge s t e e l re f inery . Especially i n t h i s a rea , d i s t i n c t SO damage markings on t h e needles of Japanese reg pine were v i s ib l e . The other was i n a r u r a l a rea i n Miyoshi c i t y . It was loca ted about 60 km north of a highly indus t r ia l ized area along the Seto Inland Sea including Fukuyama c i t y . I n t h e r u r a l a rea , t h e disease caused by Rhizosphaera ka lkhoff i i had not been noted i n recent years . A t o t a l of 900 pot ted seedlings of Japanese red pine used i n t h e t ransplant ing study were cultivated, previously i n t h e nurseries i n t he
r u r a l area i n Miyoshi c i t y . Seedlings were divided in to 12 groups as shown i n f igure 2 according t o t he combination of t ransplant ing and inoculation.
SO concentrations of both areas were determined2by means of t h e lead peroxide method. Needles were analyzed f o r sulfate-S and f o r total-S dry bas is i n order t o determine the accumulation of su l fu r i n t h e needles. Fol ia r symptoms were r a t ed from 0 t o 5 , according t o t h e degree of injury,and t h e average of these individual indexes provides t h e disease sever i ty f o r each group.
After being t ransported from t h e r u r a l a rea t o t h e indus t r ia l ized area, several seedlings exhib- i t e d t yp i ca l symptoms and signs of t he disease, while others remaining i n t h e r u r a l a rea displayed no evidence of in jury , even though they received the conidial spraying of t h e causal fungus ( f i gu re 2 ) .
In t he experiment using a f i l t e r e d a i r method, an experimental p lo t was establ ished i n an a rea i n Mihara c i t y close t o a powdered-coal-burning power p lan t . In t h i s a rea , a l a rge number of nee-d ie bl ighted Japanese red pines were found, and they displayed gradual reduction i n shoot and wood growth. The previous year 's annual mean contents of su l fu r compounds i n the a i r was 1.04 mg SO3 1100 cm2 PbO2 per day.
Plants were grown i n p l a s t i c covered chambers. One chamber ( 2 x 4 ~ 2 m ) was equipped with carbon f i l - t e r s and blowers, and t h e control o r ambient a i r chamber was provided with blowers. The a i r passed through each chamber a t a r a t e of about one a i r change / min. SO2 concentrations of t h e ambient a i r were determined by means of an automatic re-corder, and SO concentrations i n chambers were 2measured, by t h e lead peroxide method.
The experimental design was s imi la r t o t h e t rans- plant ing experiment i n Fukuyama and Miyoshi, t h a t i s , a t o t a l of 300 seedlings were divided i n t o 12 groups according t o t h e combination of exposure t o SO2 and inoculation of t h e causal fungus, then seed- l i ngs were placed i n chambers t o determine whether some protect ion from infec t ion could be obtained when the a i r drawn i n t o t he chamber was f i l t e r e d . There was a la rge reduction i n t h e sever i ty of t h e disease i n t h e f i l t e r e d a i r compared with t h a t of t he disease i n t h e ambient a i r . These findings by f i e l d s tudies furnished t h e evidence t h a t SO2 i n t he a i r was responsible f o r t h e development of t h e disease. The su l fu r content espec ia l ly sulfate-S i n t h e needles of t h e t r e e s exposed t o SO2 was s ig-n i f i can t ly grea te r than t h a t of untreated needles both i n t h e t ransplant ing and t h e f i l t e r ed -a i r ex-periments.
An increasing i n disease:severity may.be expected when a i r po l lu tan ts weaken t r e e s , making them more susceptible t o infec t ion by weak paras i tes , o r re-duced vigor t o a degree t h a t p lan ts become predis- posed t o a t tack by f acu l t a t i ve pa ra s i t e s , such as Rhizosphaera ka lkhoff i i , and Botryt is cinerea Pers. ( ~ a n n i n ge t a1. 1970) . Conversely, Linzon (1958)

--- 4
On July 18, 1970, Zelkova serrata trees in a
i! 2j,6;/(U 2sp jCO
0-0-0-0-
0 0
J A S D J A S D J A S D
5.11
transportat ion inoculation
PJ A S J T !
J : Jun 20
A:Aug 1
S: Sep 13
JASDJ A S D J- J-0
Figure 2. Effect of SO2 on the development of needle blight of pine caused by Rhizosphaera kalkhoffii
found that smelter emissions decreased the inci- dence of Cronartium ribicola C.J.Fis.,and there are additional reports that aggressive or obligate para- sites, such as Microsphaera alphitoides Griff. et Maubl. (~och 1935) , and Melampsorella cerastii Wint , Peridermium coloradense Arth. et Kern, and some species of Melampsora and Gynmosporangium (Scheffer and Hedgcock 1955) were less abundant in areas ad- jacent to smelter zones or to industrialized areas. ltol found a similar case in Tokyo that a" close re- lationship existed between the increase of distance from the metropolitan area and the increase of the incidence of willow rust disease caused by Melampsora coleosporioides Dietel and a leaf rust disease of Styrax japonicum Sieb. et Zucc. caused by Pucciniastrum styracinum Hirat. The complex of air pollutants was brought about by urbanization and industrialization, typical of the mixture of pollutants occurring in and around Tokyo, but SO2 was considered to be principally responsible for reducing the abundance of these fungi.
OXIDANTS MAINLY OZONE ( 0 )
Beginning in the 19701s, changes from solid to liquid and gas fuels, and especially increasing use of motor vehicles, have caused a reduction of the SO type of urban pollution and an increase of "ozone" pollution.
Personal communication from S. Ito, University of Tokyo, January 15, 1980.
yard of a senior high school located in Suginami Ward in Tokyo showed unusual shedding of their leaves, and strawberry leaves showed white spots clearly indicating the effects of oxidants. In addition to the damage to plants, 45 students of the school were hospitalized for treatment of irri- tated eyes, sore throats and difficulty in breath- ing. From this evidence, the Metropolitan Govern- ment issued the statement that the damage to humans and vegetation was caused by photochemical air pol- lution (The Tokyo Metropolitan Government 1971), and this was thought to be the first episode of 'Los Angeles type smog" in Japan. But as a result of later investigations it was found that there were differences between Tokyo and Los Angeles air pollution, since SO2 concentration and humidity in Tokyo were much higher than in Los Angeles; fur- thermore, it was confirmed that there was forma- tion of sulfuric acid in the air (The Tokyo Metro- politan Government 1971). These findings suggest that the complex of photochemical oxidants and sulfuric acid might exert a far greater influence on humans and vegetation than any other alone at that time.
In the latter half of 19701s, with continuous increasing industrialization and transportation, every major city in Japan has experienced an in- creasing in photochemical oxidants even though there was a slow but steady decline in the concen-. tration of SO2, in the ambient air (figure 1).
Photochemical oxidants , mainly ozone (0 ) ,con-tinue to cause the defoliation of broad leaf trees. Especially, in the Kanto district, at the center of Tokyo, unusual defoliation of Zelkova serrata still continues, since it is sensitive to both SO 2(Inoue 1973) and 0 (Kadota and Ohta 1972). While, in the Kansai dist?ict,at the center of Osaka, there is a high incidence of defoliation of poplar trees in the early summer in the parks, the gardens or the yards of schools throughout the district.
In addition to the measuring system of SO 2throughout the country, a number of automatic oxi- dant recorders were also set up by the national and the prefectural agencies, but there is still a shortage of satisfactory apparatus for monitoring the low ambient concentrations. To make up for the shortage of an instrumented system, plants have been used extensively as bioindicators of air pollu- tion. The use of bioinaicators has long been ac- cepted, since damage to green plants is usually one of the first signs that air pollution is becoming a serious problem e err^ 1964); their usefulness in this capacity is based primarily on the sensitivity of selected plant species and varieties for speci- fic pollutants (Heck 1966).
Many clones, species and varieties of poplars were selected as bioindicators because of their high sensitivity to oxidants and their wide distri- bution, especially in the Kansai district. They also have enounnous advantages in that they are fast growing, easy to propagate vegetatively by

cutt ings, and are uniformely sens i t ive t o oxidant injury.
Despite these advantages, poplars are frequently plagued by various kinds of insects and diseases t h a t degrade the usefulness of poplar clones as pollut ion indicators . For example, in te rpre ta t ion of t he response t o oxidants i s made d i f f i c u l t by l ea f diseases caused by Marssonina brunnea (El1.et Ev.) Magnus, Septot inia podophyllina Mhetz. and Melampsora lar ici-poplul ina Kleb. A r e l i ab l e guide t o d is t inguish among symptoms caused by a i r pollu- t a n t s , insec ts , and diseases has been developed and published (Tanaka 1975 ) . The defol iat ion caused by Marssonina brunnea i s one of the most serious dis- eases. It occurs wherever poplars grow, and i s not r e s t r i c t e d t o urban regions, but frequently occurs with sever i ty i n urban areas. The disease develops t i n y spots with l i g h t margins about 1mm i n dia- meter over the e n t i r e surface of t he l ea f . These superf ic ia l spots a r e not so d i f f i c u l t t o d is t in- guish from the symptoms caused by ozone by plant pathologists with a t ra ined eye, but many people who want t o use poplars as indicators a re confused with the s imi lar symptoms produced by ozone and the disease (Tanaka 1975). In addition, t he disease causes premature lower l e a f abscission,and the de- fo l i a t i on progresses upward toward the t i p s of the shoots from the l a t e spring through the summer months. The unsightly appearance of affected t r ee s discourages poplars ' use as indicators of oxidants.
A number of invest igators suspected t h a t abscis- s ion was not due d i r ec t ly t o t he pathogen but re-su l ted instead from increased r a t e s of ethylene production from the infected p lants (Abeles 19731, fur ther , ozone-induced abscission of leaves of vary- ing maturity was closely correlated with r a t e s of ethylene production ( ~ b e l e s 1973). From these
reasons, Tanaka (1976~)) hypothesized t h a t increased r a t e s of ethylene production caused by the syner- g i s t i c e f f ec t s of t he infect ion of t he fungus and ozone was responsible fo r t he incidence of the dis- ease i n t he urban area.
Fortunately, there i s s t r i k ing v a r i a b i l i t y i n suscept ib i l i ty of poplar clones t o the fungus and ozone. Clones were selected tha t a re sens i t ive t o ozone and r e s i s t an t t o t he disease. The se lec t ion of fers one of t he best means of increasing t h e usa- b i l i t y of poplar clones as ozone indicators (Tanaka 1977a). Table 1presents t he re la t ionship between ozone sens i t i v i ty and disease resis tance of poplar clones (Tanaka 1 9 ~ ~ a ) . The data of resis tance of poplar clones t o t he disease are based on the three years' observation made i n Osaka using 38 clones of young rooted cut t ings t an aka 1 9 7 ~ a ) , and t h e data of ozone sens i t i v i ty of them are based on the re-s u l t s from the exposure experiments with 0.15 ppm ozone fo r 40 hours i n chambers conducted by Enoki (1977).
Populus maximowiczii cv. O J 115 and cv. OJP 1, P.m a x i m o ~ i ~ c h o c a r ~ acv. OP 41, P .maxino-& z i i x 2.nigra pl&tierens cv.0P 52, and P.nigra i t a l i c a x ~.maximowiczii,cv.Kamabuchiare promising clones for ozone indicators because they are sensi-t i v e t o ozone and r e s i s t an t t o the disease ( tab le 1).
Yambe (1978) a l so attempted t o f ind a possible usefulness of the change of microbial f l o r a on bam- boo leaves as indicators fo r a i r pol lut ion. He found t h a t Puccinia kusanoi Die te l and epiphytic bac ter ia on the l ea f surfaces of bamboo decreased sharply i n number i n the urbanized and indus t r ia l - ized areas.
Table 1. Relationship between clonal suscep t ib i l i t y of poplars t o 0 and t o Marssonina brunnea. 3
Suscept ib i l i ty t o Marssonina brunnea I Highly Resistant Moderately Susceptible Highlyres i s t an t r e s i s t an t susce'otible
r e s i s t a n t
1R e s i s t a n t ( R ) HE 2, OP 26, NR 19, I 45/51Serotina MR 32
Moderately r e s i s t an t
Gerlica I 262I 154, I 214
I 72/51, Kamabuchi, Highly O J 115, OJP 1, OJP 2, I 77/51,
I 488susceptible(ss^ OJP 3, OJP 4, OJP 5 , . OP 4 1
I

OTHE8 POLLUTANTS pa ra s i t ey Rhizosphaera ka lkhoff i i (Chiba and Tana- ka 1968).
In addit ion t o SO and oxidants, s ign i f icant and 2
sometimes devastating e f f ec t s of other a i r pollu- t a n t s have long been recognized i n Japan. Fluoride compounds have a long h i s to ry as pol lutants i n t h e v i c in i ty of ceramic industry, and more recent ly, plant in jury resu l ted from f luor ides has gradually increased because of t h e expansion of indus t r ies such a s aluminum re f ine r i e s m d f e r t i l i z e r manu- factures ( ~ a t u u r a and Kokubu 1972). Chlorine (Ta- naka 1977b), ethylene, acidic dust from stacks, and a lka l ine dust from cement k i l n s (Inoue 1972a and 1972b) have a l so been recognized as agents of dm- age t o vegetat iony but t he damaged areas a r e re la - t i v e l y r e s t r i c t e d because most of them a re emitted from point sources o r by accidental s p i l l s .
The in te rac t ion of these minor pol lu tan ts with fungi and bac ter ia has received only minor at ten- t i o n , but Inoue (1972a and 1972b) found t h a t t he number of f r u i t i n g bodies of Tricholoma matsutake Singer was s ign i f i can t ly reduced i n t he heavily dusted fo re s t s i n t h e v i c i n i t y of cement works com- pared t o l e s s severely dusted o r non-dusted por- t i ons of fo re s t s . According t o t he r e s u l t s of hand-dusted experiments,he concluded t h a t t h e alka- l i n e so lu t ion from t h e dust severely decreased the formation of t h e f r u i t i n g bodiesy and reduced the development of t h e fungus colonies.
CONCLUSIONS
The in te rac t ion of po l lu tan ts with pa ra s i t i c and epiphytic microorganisms has received l imi ted a t - t en t ion i n Japan. The ex is t ing reports a r e only associated with t r e e s and bamboos and no report with annual p lan ts was found i n a search of avai l- able l i t e r a t u r e .
Some pol lu tan ts a r e d i r e c t l y tox ic t o microor- ganisms on the leaf surface. They might possibly impair t h e i r growth o r reproductiony and change the population and community composition of mCcroorgan- i s m s on the host p lan ts , s ince t he s ens i t i ve spe- c i e s t o a i r po l lu tan ts would be gradually replaced by more t o l e r an t species (Treshow 1968). Especial- l y y aggressive o r ob l iga te paras i tes such a s Cerco- spora sequoiae (Kaburagi 1930), Pucciniastrum m- racinum and Melampsora coleosporioides (1 to1)~ and Puccinia kusanoi (Yambe 1978) a r e l e s s abundant i n polluted a r easy and the disease sever i ty they induce? is sharply reduced.
On t h e other hand, i n fo re s t stands under inf lu- ence of i n d u s t r i a l and urban a i r pol lut ion, some f o l i a r pathogens counted by pathologists i n t h e group of so-called ' f acu l t a t i ve pa ra s i t e ' a r e of grea te r importance because a i r po l lu tan ts i n ju re l ea f t i s s u e and predispose t r e e s t o a t tack by f acu l t a t i ve paras i tes . A close cor re la t ion has been found between SO i n t h e a i r and a high inc i - dence of needle bl ighz of pine caused by a weak
' Personal communication from S. It0 University of Tokyo, January 1980.
Although plant in jury caused by spec i f ic pollu- t a n t s such a s SOpy 0 and f luor ides has gradually been abated by enginJering improvements o r by-pro- ducts recovery, chronic in jury caused by a complex of urban a i r po l lu tan ts has assumed addi t iona l im- portance a s urbanization and t h e use of motor ve- h i c l e s have increased.
Injury t o green p lan ts i s usually one of t h e f i r s t s igns t h a t a i r po l lu t ion is becoming a ser ious problem, and sever i ty of spec i f ic diseases i n pol- l u t ed areas must be regarded a s an addi t iona l ex- pression of unbalanced na tura l conditions. These spec i f ic diseases and t h e change of composition of l ea f surface microorganisms provide a ids f o r moni- to r ing a i r po l lu tan tsy espec ia l ly i f they a r e sen- s i t i v e t o very low concentration of a i r po l lu tan ts . These diseases and microorganisms should be consid- ered a useful supplement t o t h e more expensive techniques and instruments used f o r t h e i den t i f i - cat ion of chronic a i r po l lu tan t e f f ec t s .
Acknowledgments: I thank D r . Paul R. Mi l le r , Pac i f ic Southwest Forest and Range Experiment Sta- t ion , U.S. Dep. Agr icaY f o r h i s c r i t i c a l readings of the manuscript.
L1T.ERATURE CITED
Adms $D ,F . and C . W. Sulzbach . 1961. Nitrogen deficiency and f luor ide suscepti-
b i l i t y of bean seedlings. Science 133: 1425-1426.
Abeles ,F.B. 1973. Ethylene i n plant biology. 302pp. New York.
Berry,C.R. 1964. Eastern white pine, a t o o l t o de tec t a i r
pol lut ion. Southern Lumberman 209(2609):164,166
Chiba,O. and K.Tanaka. 1968. The e f f ec t of sulphur dioxide on t h e develop-
ment of pine needle b l igh t caused by Rhizosph- aera ka lkhoff i i Bubk. I. J.Jap.For.Soc. 50: - 135-139
Doi ,T. 1919. orest st p ro tec t ion) . b09pp. Tokyo.
Enoki ,M . 1977. (Clonal suscep t ib i l i t y of poplars t o ozone). Ann. ~ e p . 1976. ,Osaka Agr . Centr . : 117-119.
Heagle,A.S. 1973. Interact ions between a i r po l lu tan ts and
plant paras i tes . Ann.Rev.Phytopath. 11:365-388.
Heck,W.W. 1.966. The use of p lan ts a s indicators of a i r pol-
lu t ion . A i r Water Poll.1nt.Nat.J. 10:gg-111.

Inoue,I. lg72a. Studies on the influence of smoke-dust from a cement plant on the vegetation of pine forests, especially on Tricholoma matsutake. I. Trans .Mycol. SOC. Jap. 13: 196-214.
Inoue,I. 1972b. Ditto 11. M:215-288.
Inoue T . 1973. (Tree injury caused by sulfur dioxide). Plant Protection 27:8-12.
It0 ,K. 1976. Blight and canker of Cryptomeria Japonica D.Don. caused by Cercospora se uoiae Ellis et Everhart. Ann.Phytopath. Soc .-%-2:234-236.Jap
Japanese Environmental Agency. 1979. (White paper on environment for 1979) . 2163pp. Tokyo.
Kaburagi,T. 1930. (Smoke damage to plants). Iwanami Lectures for Bioloa. 1-39.
Kobayashi >T. 1967. Critical revision on the genera Rhizosphaera Mangin et Harriot and Rhizophoma Petrak et Sydow, a little-known fungus group associated with needle disease of conifers. Bul1.Gov.For.Exp. Sta. 204 :91-112,
Kadota,M. and K.Ohta. 1972. Ozone injury to some Japanese woody plant species in summer. J. Jap. For. Soc . 54:226-229.
K6ch9G. 1935. Eichenmehltau und Rauchgasschaden. Z.Pf-lanzenkr. 45:44-45.
Linzon,S.N. 1958. The influence of smelter fumes on the growth of white pine in the Sudbury region. Can.Dep. Agr.Publ. ,Ontario Dep. Lands Forests. 45pp.
Matuura,S. and N.Kokubu. 1972. Fluorine, its distribution and role in nature. 150pp. Tokyo.
Scheffer,T.C. and G.G.Hedgcock, 19.55. Injury to northern forest trees by sulfur dioxide from smelters. UDSA,For.Serv.Tech.Bull. 1117:49pp.
Tanaka,K. 1975. The similarity between symptoms caused by air pollutants, insects, and diseases. Agr. and Hort. 5O:Preface.
Tanaka ,K. 1976~~.m e effect of low-level SO fumigation on the development of needle bligh$ of pine caused by Rhizosphaera kalkhoffii. Anr1.Rep.1975.~ m e range of tolerance of biological indicators to pollutants:5-9.
Tanaka,K. 1976b. Tree diseases in air polluted areas and their usability as indicators. a : 51-56.
Tanaka,K. 1977a. Pest problems in the use of poplar clones as indicator plants for photochemical oxidnats . Trans. 88th Mtg .Jap.For. Soc . :419-421.
Tanaka,K. 1977b. Accidental chlorine gas damage to ornamen- tal woody plants. w:4l5-4l7.
TanakayK.,T.Okada and K.Hako. 1974a. Effect of SO2 on the development of needle blight of pine caused by Rhizosphaera kalkhoffii, Transplanting experiment. Trans. 86th Mtg .Jap .For. Soc.:290-292.
Tanaka,K.,M.KuwakiBT.Okada and K.Hako. 1974b. A plastic covered chamber with activated carbon filters. =:211-213.
Tanaka,K. and 0.Chiba. 1971. On a needle blight ~f pine caused by Rhizo- sphaera kalkhoffii--Life historyy physiological characteristics and pathogenicity of the causal fungus. J. Jap .For, Soc . 53: 279-286.
Tanita, S . 1976. Histological effects of air pollutants and pathogens on Pinus thunbergii. Trans 88th Mtg. Jap .For. Soc . :373-37k.
Tokyo Metropolitan Government. 1971. ~okyo fights pollution. 2 6 7 ~ ~ . Tokyo.
Treshow, M. 1968. m e impact of air pollutants on plant popu- lations. Phytopathology 58 :1108-1113.
Ymbe, Y. 1973. decline of trees in Tokyo. Bul1.Gov.For. Exp.Sta. 257:lOl-107.
Ymbe, Y. 1978. Decline of trees and microbial florae as the index of pollution in some urban areas. Bull. For.& For.Prod.Res.1nst. 301:llg-129.

Sensitivity of Lichens to Air Pollution with an Emphasis on Oxidant Air
Pollutants1
^ Thomas H. Mash I11 and Lorene L. Sigal
Abstract: The hypothesis that lichens are sensitive indicators of air pollution is now well established for oxidants, sulfur dioxide, hydrogen fluoride and trace ele- ments. From field studies differential sensitivity of dif- ferent lichen species around pollution sources is evident. Laboratory studies with the particular air pollutant general- ly confirm the same degree of differential sensitivity.
LICHENS AND THEIR ECOLOGY
Lichens, which strictly speaking are called lichenized fungi, are composite plants represent- ing a symbiotic union of algae and fungi to form morphological entities unlike either component (Ahmadjian 1967, Hale 1967). Although a few species survive in aquatic environments, most are found in terrestrial habitats, occurring on soil, rocks and as epiphytes on other plants. All lichens are perennial plants with life spans varying from a few decades to reportedly thousands of years (Weber and Andrews 1973). Essentially all lichens are autotrophs because of the photo- synthetic activity of the algae. Metabolic activity in general is limited to period's when the lichens are moist, a condition which varies in concert with atmospheric moisture conditions as lichens have no roots by which moisture might be absorbed from the substrate. Thus lichens are prominent examples of poikilohydric plants (Larcher 1973) which also include bryophytes and some lower vascular plants, such as desert species of Selaginella and ferns. Because of moisture
presented at the Symposium on Effects of Air Pollutants on Mediterranean and Temperate Forest Ecosystems, June 22-27, 1980, Riverside, California, U.S.A.
'~r. Nash is associate professor of botany, Dept. of Botany, Arizona State University, Tempe, Arizona 85281 USA; Dr. Sigal is a research scien- tist, Environmental Sciences Division, Oak Ridge National Laboratory, P.O. Box X, Oak Ridge,. Tennessee 37830 U.S.A.
constraints on photosynthetic activity, primary productivity assignable to lichens is generally small compared to higher plants. Nevertheless lichens may be important in ecosystem functioning. For example, lichens are generally recognized as important pioneer plants of xeric successional sequences because of their role in biologically controlled weathering (Syers and Iskandar 1973). In addition, lichens are important in mineral cycling, not only because of their ability to alter precipitation chemistry (Ianq and others 19761, but also from the ability of blue-green algae contain- ing lichens to fix atmospheric nitrogen. Den son (.1973) has estimated that lichens are respons ble for 50 percent of the nitrogen fixed in the Douglas-fir forests of the Pacific Northwest.
Lichens are morphologically divided into 3 major growth forms: crustose, foliose and fruticose. Crustose species are generally small, tightly attached to (or imbedded in) their substrate and poorly differentiated. Foliose species are gen- erally larger, loosely attached to their substrate and stratified into several distinct layers. In contrast to the basically 2-dimensional foliose species, fruticose s~ecies are strongly 3-dimen- sional and grow out from their substrate. Most fruti'cose species are also differentiated into distinct layers. Of these morphological forms fruticose species are generally considered to be the most sensitive to air pollution (Fenton 1964), a fact which may be related to the high surface- to-volume ratio of this form.
LICHENS IN RELATION TO AIR POLLUTION
Different species of lichens are well-known to

be differentially sensitive to air pollution (Nash 1976). A few species actually grow better in urban environments where air pollution levels are high in contrast to their performance in rural areas. Most species, however, exhibit varying degrees of sensitivity to air pollution and some are more sensitive than higher plants. Thus by documenting variations in abundance and species richness of lichen communities, it is frequently possible to identify poliution sources and to document the magnitude of the air pollution probla, Patterns within lichen communities may be attribut- able to many environmental factors other than air pollution, of course, and consequently it is extremely important that studies be made within a multivariate context, including both pollution factors and non-pollution factors.
The fact that lichens are differentially sensi- tive to air pollution is based on a series of field observations and laboratory experiments extending back over the past 130 years. Grindon (1859) and Nylander (1867) noted that lichens were disappearing from city centers. Later Arnold (1892) documented that lichens transplanted from rural areas to the center of Munich were not able to survive. Subsequently this "city effect" has been reported for many cities in Europe and North America (Hawksworth 1971). By air pollution most workers of the 1800's generally meant coal soot and related particulates. However, over the past decades a number of invisible gases, such as sulfur dioxide and ozone, have been identified as the major causes of air pollutant injury to plants. Experiments with these air pollutants over the past two decades have also demonstrated differential sensitivity among different lichen species. In general, the sensitivity patterns demonstrated in the laboratory have remarkably Corresponded to the patterns observed in the field around pollution sources. Perhaps the strongest evidence supporting the contention that lichens are sensitive to air pollution comes from studies documenting lichen recovery following pollution abatement. For example, Seaward (19761 has docu- mented that Lecanora muralis fairly rapidly reinvaded an urban complex in southern England following amelioration of air pollution in the area.
In the following sections specific evidence related to various types of air pollutants w211 be reviewed.
Oxidant Air Pollutants
Oxidant air pollutants, including ozone and peroxyacetplnitrate (PANI, are a group of gases characteristic of the oxidizing atmospheres found in the Los Angeles type smog (Pitts and Finlayson 1975). Although the effects of these compounds have been studied extensively with higher plants (National Academy of Sciences 19771, relatively few studies have been conducted with lichens. Probably the only field study in which lichens have been shown to be sensitive to oxidants is that of Sigal and Nash (1980) for the southern
Californian mountains* The study was centered in the San Bernardino Mountains, utilizing the sites mployed by the University of California (Riverside and Davis] ecosystem study of air pollutant effects in contrast to sites in a control region, Cuyamaca Rancho State Park in southern San Diego County. In addition, there was an excellent historical record of the lichens of the San ~ernardinos based on collections made by H. E. Hasse at the turn of the century. Of the 91 foliose and fruticose lichen species reported by Hasse (1913) in his lichen flora of southern California, only 34 species were found during three sunnners of study throughout southern California. Thus there has been a significant reduction in the number of species present. To obtain quantitative data on lichen variation, both conifers (Ab2es concolor, Pinus ponderosa, Pinus jeffreyi and Pseudotsuqa macrocarpa) and California black oak (Quercus kelloggii) were sampled for lichen cover at breast height.
In the case of conifers Hasse (1913) reported the presence of 16 foliose and fruticose lichen species in the San Bernardino Mountains. In our recent study [Sigal and Nash 19801 only 8 of these species were found in the San Bernardino Mountains and 4 of these were only present in vestigial quantities. In contrast, at sites to the north and to the south K'uyamacasl 15 of the 16 species reported by Hasse were found. Thus in the San Bernardinos which lie adjacent to the Los Angeles Bash, there has been a SO percent decline in species richness of lkhens on conifers. Of the species which do occur in the San Bernardinos only two (Letharia Vulpina and Hypogymnia entero- momha) occur commonly. The latter species, although it is common, is definitely showing signs of deterioration (Table 1) when randomly selected thalli from the San Bernardinos are compared with a similar set from the Cuyamacas. The San Ber- nardino population has a much higher percentage
Table 1-- Morphological characteristics of recent collections of Kypogymnia enteromorpha from the San Bernardinos and the Cuyamacas.
Percent Bleaching
Forest
38 59 41
Percent Convoluted
Forest
38 38 47 15
of bleached and convoluted thalli. In addition, there was a decrease of approximately 50 percent in overall thallus dimensions in the San Bernardino population and a decrease in fertility of 42

percent. Thus there are marked trends between the San Bernardinos and the Cuyamacas of reduced species richness and species vitality in the form- er area. Within the San Bernardinos themselves, significant variation exists. Below approximately 150 ppm-h oxidant dosage, cover of both Letharia and Hypogymnia is normal in comparison with other areas. At higher dosages of up to 285 ppm-h lichens are almost completely eliminated from conifers.
Comparisons of trends of lichens occurring on black oak between the San Bernardinos and Cuyamacas also indicated that a deterioration was occurring in the San Bernardinos, but the trends were not as dramatic as in the case of lichens occurring on conifers. No direct comparison with the Hasse material was possible because Hasse did not note the species of oak in his collections. From our sampling 39 species of lichens were found on black oak in the Cuyamacas and 30 specie3 in the San Bernardinos. Fourteen of the species found in the Cuyamacas were not found in the San Bernardinos whereas three species were found exclusively in
13.5 12.1 . ucc
7.6 COO. 12.9
10.2 12.1
TOTAL % COVER ALL L ICHENS
15.7
BF.
Bp CA GVC 4 230.8 186.7 DWA T2 145.8
184.9. ,,cc i33.2
COO. 134.7
175.0
OXIDANT DOSE ESTIMATES 183.2 BF.
the San Bernardinos. Among the 20 species which occurred in common there was a general shift from high frequency in the Cuyamacas to relatively low frequency in the San Bernardinos. In the case of cover of the 6 most common species, no significant difference was observed between the two mountain ranges except for Collema nigrescens, the only nitrogen fixing lichen in the groupi which was completely absent from the San Bernardinos. Fur- ther analysis revealed that Parmelia subolivacea, the most common lichen, was a successional species. Because older trees occurred in the Cuyamacas, a significantly higher value for this species in the Cuyamacas was predictable. Further evidence sup- porting the hypothesis that the dominant species was in fact respondhg to oxidants was obtained by running ordinations (princiwal component analy- sis) of the sites within the San Bernardinos and relating the observed variation in species com- position to oxidant dose estimates and other environmental variables (fig. 1). An initial ordination (not shown) included all sites sampled within the San Bernardinos. It exhibited a small cluster of low cover sites which occurred at the
I % COVER
PARMELIA SUBOLIVACEA 8.2 BF
D
SAN BERNARDINO NAT'L FOREST STUDY -SITE LOCATIONS
Fig. 1--Principal component analysis ordination of 10 sites in the San Bernardino Moun- tains using cover data for the 5 most important lichen species occurring on Quercus kelloggii. Eighty percent of the variation is explained by axis one (the abscissa). Figure 1A shows the positioning of the sites on the ordination plot with respect to to- tal cover of all lichens; figure lBf the same ordination but with cover values for =- melia subolivacea; figure lC, the same ordination, but with oxidant dose estimates (ppm-h); figure IDf the geographic location of the sites within San Bernardino Nation- al Forest.

highest elevations where summer fog is infrequently observed. Consequently these sites and one other site on the desert side of the San Bernardinos were excluded from the subsequent analysis. The 10 sites plotted in figure 1 are thus homogeneous with respect to altitude and position along the Los Angeles side of the San Bernardinos (fig. 1Dl. In the subsequent ordination (fig. lA, 1B and lC), 80 percent of the variation is explained by axis 1 (the horizontal axis). From figures 1A and 1B it is clear that the primary source of variation is the percent cover of all lichens, which in turn corresponds closely to the percent cover of E. subolivacea. Furthermore, there is an inverse relationship between percent cover of E. s-olivacea (fig. 1 ~ ) and oxidant dose estimate (1C). This inverse relationship was substantiated by running a Speaxman rank correlation between the two variables. The test was significant with an alpha value of 0.007. For the other 4 species no relationship between oxidant dose estimates and cover values were found. Thus some species have been completely eliminated from the San Bernardinos, other species have probably declined in abundance and a few species are apparently unaffected.
Preliminary ozone fumigations have shown dif- ferential responses in photosynthesis reduction between Pamelia sulcata and Hypogymnia entero- morpha with the former species exhibiting greater sensitivity (Nash and Siyal 19791. The Parmelia, which grows on black oak, is absent from the San Bernardinos whereas the Hypogymnia is present, but is exhibiting signs of deterioration. For these species, at least, the pattern of sensitivity observed in the field and the laboratory are com- plementary. A similar pattern of response for the two species was observed after fumigations with PAN (Sigal and Taylor L979). Further stud2es with both ozone and PAN should be run, but the current evidence strongly supports the assertion that oxidants are a major cause of the decline of the lichen flora in the San Bernardino Mountains.
Sulfur Dioxide
Sulfur dioxide is released into the atmosphere by combustion reactions involving products con- taining the ubiquitous element sulfur, such as coal and oil. It has long been recognized as a phytotoxic agent in general (Daines 1968). Lichens are well known to be sensitive to sulfur dioxide (Nieboer and others 19761. Most of the field studies in northern Europe and North America have involved sites where sulfur dioxide was a major factor (Hawksworth 1971). One of the classic studies by Rao and LeBlanc (1967) demon- strated a severe decline in lichen abundance along a 70+ km transect NNE of Wawa, Ontario where an iron sintering plant is present. Because there is essentially no human development in the affected region, air pollution, and particularly sulfur dioxide, is undoubtedly a major cause of the de- cline in the reqionts lichens. Other studies have shown that lichens accumulate sulfur (Olkkonen and Takala 1975, Laaksovirta and Olkkonen 1977) in situations where the presumed
source of sulfur is sulfur dioxide. The gas readily dissolves in water forming a sulfite or bisulfite solution depending on the pH. As a con- sequence acidification of the substrate frequently occurs, as has been shown in Stockholm where Skye (19681 found that urban tree bark had a pH of more than two units less than corresponding trees from the country-side. Acidification strongly affects the ability of lichens to survive, in part because sulfur dioxide is much more toxic at lower pH's (Tuerk and Wirth 1975). Gilbert (1965 and 1970) has clearly demonstrated the ability of lichens to penetrate into central Newcastle on basic sub- strates when they are absent from acidic ones. In the laboratory studies, photosynthetic decline in response to short term sulfur dioxide exposures as low as 0.2 ppm have been demonstrated (Tuerk and others 19741. In aqueous experiments with reputedly lower concentrations, effects have also been documented Baddeley and others 1973, Puckett and others 19741.. In addition to reduction of photosynthesis, bleaching of lichen thalli may occur due to phaeophytinisat2on of the chloro- phyll~ (Nash 1973, Puckett and others 19731. More recently potassium efflux from lichens exposed to sulfur dioxide has been shown to be the most sen- sitive criterion of response (Tomassini and others 1977). Alteration of membrane permeability may thus be an immediate response to sulfur dioxide exposure. Subsequently it is suggested (Nieboer and other 1976, Puckett and others 1974) that interference in electron flow in photosystem I and I1 will occur'and that proteins will be affected through sulfur dioxide$ ability to cleave disul- phide linkages. Thus there is very strong evi- dence from both field and laboratory studies that lichens are sensitive to sulfur dioxide.
Hydrogen Fluoride
Hydrogen fluoride is extremely volatile and is released into the atmosphere during aluminum and rare earth metal refining, fertilizer production and glass and ceramic manufacturiny. Patterns of lichen decline are well demonstrated around alumi- num factories (Martin and Dacquard 1968, Gilbert 1971, LeBlanc and others 1972, Horntredt 1975) and around a titanbn plant (Nash 19711 and around a phosphate plant (Takala and others 19781. In these areas fluoride accumulat~on by the lithens has been demonstrated with decline in the lichen cornunities being associated with elevated fluo-ride levels. Transplants of healthy lichens into the impoverished zones resulted in fluoride accu- mulation and subsequent death of the transplants. Gilbert suggested that the critical fluoride level lay between 20 and 50 ppm and Nash independently suggested that the value lay between 30 and 80 ppm. Schoenbeck (1969) found that his transplants died near a fluoride source, but that new transplants survived well for several months following cessa- tion of the fluoride emissions. In a laboratory study with exposure to approximately 5 ppb fluoride over a 9-day period Nash (1971) found that lichens accumulated 84-115 ppm fluoride in contrast to control levels of 14-25 ppm and that the fumigated samples developed chlorotic and necrotic symptoms

similar to the injury observed in the field trans- plants. Thus fluoride as the probable cause of the injury was confirmed.
Trace Elements
Lichens have a well demonstrated ability to accumulate elements from dilute aqpeous solutions (Nieboer and others 1977)# a fact probably related to their apparent dependence on atmospheric sources for mineral nutrition. As a consequence of this ability to retain elements# the study of trace element concentrations in lichens is fre- quently useful in demonstrating "fallout" patterns of particulates and aerosols. For exampleI mercury is found in elevated concentrations near a chlor- alkali works in Finland (Lodenius and LaaJcsovirta 1979); lead is elevated in lichens adjacent to highways (Laaksovirta and other 1976# Lawrey and Hale 1979); and a number of elements are present in high concentrations in citiesI such as Tel Aviv (Garty and others 1977) and Sendalp Japan (Saeki and others l977lI near the nickel complex at SudburyI Ontario (Tomassini and others 1976) I near a coal-fired power plant (Gough and Erdman 1977) and in coal mine ecosystems of Ohio (Lawrey and Rudolph 19751. In none of these cases is toxicity of the accumulated elements demonstrated. Toler-ance to high concentrations of elements may occur if the elements are relatively insoluble and are localized extracellularly. For exampleI Noeske and others (1970) showed that iron and copper were encrusted on the surface of lichens which grew on metal rich substrates in the Harz Moun- tains of Germany. FurthermoreI Garty and others (1979) have shown that particulates in Tel-Aviv are incorporated extracellularly within the lichen thallus.
In contrast to the above studies "trace" ele- ments do occasionally occur in toxic concentra- tions. Lawrey and Hale a9791 have shown accumu- lation of approximately 1000 ppm lead in a lichen growing near an expressway in the vicinity of WashingtonI D.C. They demonstrated that.lead accumulation is correlated with reduction in growth of the speciesI but they have neither demonstrated experimentally that lead was the cause of the reduced growth nor demonstrated that other factors associated with automobile emissions were unimportant. Probably the most convincing study documenting trace element toxicity is that of Nash (1975) around a zinc smelting complex in eastern Pennsylvania. In the vicinity of the smelting complexI lichen species richness was reduced to 7 in contrast to the 77 species found in a control area (Nash 1972). The zone of lichen impoverishment extended for a distance of 15 km W and 10 km E of the smelting complex (fig- 2). Although detectable and potentially toxic levels of sulfur dioxide were found adja- cent to the smeltersI the distribution of elevated levels of sulfur dioxide did not extend as far as the lichen impoverishment zone extended- The relative unirnprtance of sulfur dioxide was further documented by the lack of acidification of the bark of oak trees and by the fact that the
2oE 10 0 low DISTANCE FROM EAST SMELTER (kml
Fig. 2 ~ T h e distribution of the lichen-impover- ished zone (Nash 1972) at Palmerton# PennsylvaniaI in relation to the zones of modified forest (Jordan 1975)# to detectable levels of ambient sulfur dioxide (Nash l975lI and to elevated levels of cadmium and zinc in surface soils (Buchauer 1973).
company had installed and subsequently contin- uously used an acid plant at the time that zinc sulfide ores were initially processed. In con- trast# metal concentrations in the soil duff were as high as 1351000 ppm zinc and 1750 ppm cadmium at a site adjacent to the smelting complex Duchauer 1973). The concentration of zinc and cadmium decreased exponentially with distance from the smelters until background concentrations were found at 20 to 25 km E and 16 km W of the smelters (fig. 21. Physiological studies demonstrated that zinc and cadmium were toxic to the lichens when concentrations reached 300-500 ppm in the thallus. Since zinc was present in concentrations 100 times that of cadmium and since zinc is approximately as toxic as cadmiumI zinc was in- ferred to be the most important pollutant. ¶i points beyond the li'chen impoverishment zone zinc concentrations ranged from 20-200 ppm and cadmium from 1-30 ppmI where both sets of values were demonstra5ly non-toxic.
LITERATURE CITED
Ahmadjian, V. 1967. The lichen symbiosis. 152 p, Ginn Co.
(Blaisdell ) Boston.
Arnoldy F. 1892. Zur 1 ichenflora von Muenchen. Ber. Bayer Bot. Ges. 2: 1-76.
Baddeley, M. S., B. W. Ferry and E. ,.I.Finegan. 1973. Sulphur dioxide and respiration in
lichens. In Air pollution and lichens. B. bl. ~ e r r xII. S. Baddeley and D. L. Hawks- worthy eds. p. 299-313. Athlone Presc, Lon- don.

Buchauer* M. 1973. Contamination of soi 1 and vegetation
near a zinc smelter by z incy cadmiumy copper and lead. Environ. Sci. Technol. 7: 131-135.
Dainesy R. H . 1968. Sulfur dioxide and plant response.
J . Occup. Med. 10: 516-524.
Denisony W. C. 1973. Life in t a l l t r e e s , Sc i , h e r , 228:
74-80.
Fenton* A. F. 1964. Atmospheric pollution of Belfast and
i t s relat ionship t o the lichen f lo ra . I r i sh Nat. J . 14: 237-245.
Garty* J e Y M. Galun* C. Fuchs and N. Zisapel. 1977. Heavy metals i n the lichen Caloplaca
aurantia from urbany suburban and rural regions in Israel ( a comparative study).
Hawksworthy D. L. 1971. Lichens a s litmus f o r a i r pollution:
A h is tor ica l review. Intern. J . Environ. Stud. 1: 281-296.
Horntvedty R. 1975. Epiphytic macro1 ichens on Scots pine
related t o a i r pollution from industry in Odds* western Norway. Meddr. Norsk Ins t ' Skogforsk. 31: 581-604.
Jordan, M. J . 1975. Effects of zinc smelter emissions and
f i r e on a chestnut-oak woodland. Ecology 56: 78-91.
Laaksovirtay F. and H. 01 kkonen. 1977. Epiphytic 1 ichen vegetation and e le-
ment contents of Hypogymnia physodes and pine needles examined as indicators of a i r pollution a t Kokk01a~ W. Finland. Ann. Bot. Fennici 14: 112-130.
Water* Air and Soil Pollut . 8: 171-188. -> -and P. Alakuijala.1976. Observations on the lead content of
and M. Kessel . -;.-a1 iza t ion of heavy metals and other
elements accumulated in the lichen thal lus . New Phytol . 82: 159-168.
Gi lber ty 0. L . 1965. Lichens as indicators of a i r pollution
i n the Tyne Valley. & Ecology and the indus t r ia l society. G . T. Goodman* ed. p. 35-47. Oxford Univ. Press* London.
1976. Further s tudies on the e f f ec t of sul- phur dioxide on 1 ichens and bryophytes. New Phytol . 69: 605-627,
-1971. The e f f e c t of airborne f luor ides on
, 1 ichens. The Lichenologist 5: 26-32.
Goughy L. P. and J . A. Erdman. 1977. Influence of a coal - f i red ~ o w e r ~ l an t
on the element content of parme: i a chloro- chroa. The Bryologist 80: 492-501.
Grindon* L. H. 1859. The Manchester Flora. London.
Haley M. E , y J r . 1967. The biology of l ichens. 176 p. Edward
Arnold Ltd. London.
Hassey tt. E. 1913. The lichen f l o r a of southern Califor-
nia. Contrib. U. S. Nat. Herb. 17: 1-132.
lichen and bark adjacent t o a highway in southern Finland. ~ n v i r o n .POI 1u i . li : 247-255.
Lang* G. E a Y W. A. Reiners and R . K. Heier. 1976. Potential a l t e r a t ion of prec ip i ta t ion
chemi s t r y by epiphytic 1 ichens. Oecologia (Berl .) 25: 229-241.
Larcher, W. 1973. Physiological plant ecology. 252 p.
Springer-Verlag, Berlin. ( t rans la ted 1975 M.A. Biederman-Thorson) .
Lawrey* J . D. and M. E. Hale* J r . 1979. Lichen growth responses t o s t r e s s in-
duced by automobi 1e exhaust pol 1 ution. Science 204: 423-424.
and E. D. Rudolph. 1975. Lichen accumulation of some heavv met-
a l s from ac id ic surface substrates of" coal mine ecosystems in southeastern Ohio. Ohio J . Sci. 75: 113-117.
LeBlanc. F.* D. N. Rao and G. Comeau. 1972. Indices of atmospheric puri ty and f luor-
ide pollution pat tern in Arviday Quebec. Can. J . Bat. 50: 991-998.
Lodeniusy M. and K. Laaksovirta. 1979. Mercury content of Hypogymnia physodes
and pine needles affected by a chlor-a1 ka1 i works a t Kuusankoski, SE Finland. Ann. Bot. Fennici 16: 7-10.

Martin, J. F. and F. Jacquard. 1968. Inf luences des fumges d'usines sur l a
d i s t r i b u t i o n des l ichens dans l a va l l ge de l a Romanche (IS&-e) . P o l l u t Atmos. 19: 95-99.
Nash, T. H., I 1 1 1971. Lichen s e n s i t i v i t y t o hydrogen f l u o r -
ide. Bu l l . Torrey Bot. Club 98: 103-106.
7 2 . S i m p l i f i c a t i o n o f the Blue Mountain l i chen communities near a z inc factory. The Bryo log is t 75: 315-324.
1973. S e n s i t i v i t y o f l ichens t o s u l f u r dioxide. The Bryol og i s t 76 : 333-339.
1975. In f luence o f e f f l uen t s from a z inc fac- t o r y on 1 ichens. Ecol . Monogr. 45: 183-198.
1976. Lichens as ind ica to rs o f a i r po l l u t i on . Die Naturwissenschaften 63: 364-367.
and L. L. S igal . 1979. Gross photosynthet ic response o f 1 i ch -
ens t o short- term ozone fumigations. The Bryol og i s t 82 : 280-285.
National Academy o f Sciences. 1977. Ozone and other photochemical oxidants.
719 p. Washington, D. C.
Nieboer, E., D. H. S. Richarson, K. J. Puckett and F. D. Tomassini.
1976. The phy to tox i c i t y o f sulphur d iox ide i n r e l a t i o n t o measurable responses i n l ichens. In T. A. Mansfield, ed. p. 61-85.
, K. J. Puckett, D. H. S. Richardson, F. 9. Tomassini and B. Grace.
1977. Ecological and physiochemical aspects o f the accumulation o f heavy metals and su l - phur i n l ichens. I n Proc. In tern. Conf. on heavy metals i n theEnv i ron . (Oct. 27-31, 1975, Toronto, Ontar io) , p. 331-352.
Noeske, O., A. Laeuchli, 0. L. Lange, G. H. Vieweg and H. Z iegler .
1970. Konzentrat i on und Lokal is i erung von Schwermetall en i n Flechten der Erzschl acken- halden des Harzes. Dtsch. Bot. Ges. Neue Folge 4: 67-79.
Nylander, W. 1867. Les l ichens du j a r d i n du Luxembourg.
Bu l l . Soc. Bot Fr. 13: 364-372.
Olkkonen, H. and K. Takala. 1975. Total sulphur content o f an ep iphy t i c
l i chen as an index o f a i r p o l l u t i o n and the usefulness o f the x-ray fluorescence method i n sulphur determinations. Ann. Bot. Fen-n i c i 12: 131-134.
P i t t s , J. N. and B. J. Finlayson. 1975. Mechanisms o f photochemical a i r pol l u -
t i on . Angew. Chem. 14: 1-15.
Puckett, K. J., E. Nieboer, W. P. F lo ra and D. H. S. Richardson.
1973. Sulphur d ix ide : i t s e f f e c t on photosyn- t h e t i c C f i x a t i o n i n l ichens and suggested mechanisms o f phy to tox ic i t y . New Phytol . 72: 141-154.
D. H. S. Richardson, W. P. F lora and E. T'I eboer.
1974. Photosynthetic 14c f i x a t i o n by the 1ichen Umbi 1 i c a r i a muhl enbergi i (Ach. ) Tuck. fo l low ing shor t exposures t o aqueous sulphur dioxide. New Phytol . 73: 1183-1192.
Rao, D. N. and F. LeBlanc. 1967. In f luence o f an i r o n s i n t e r i n g p l a n t on
cor t i co lous epiphytes i n Wawa, Ontario. The Bryo log is t 70: 141-157.
Saeki, M., K. Kunii, T. Seki, K. Sugiyama, T. Suzuki and S. Shishido.
1977. Metal burden o f urban l ichens. Environ. Res. 13: 256-266.
Schoenbeck, H. 1969. Eine Methode zur Erfassung der b io log-
ischen Workung von Luftverunreiningungen durch t r ansp lan t i e r t e Flechten. Staub 29: 14-18.
Seaward, M. R. D. 1976. Performance o f Lecanora mural i s i n an
urban environment. I n D. H. Brown, D. L. Hawksworth and R. ~ . B a i l e y , eds. Lichenology: progress and problems. p. 323-357. Academic Press, Inc., London.
Sigal, L. L. and T. H. Nash 111. 1980. Lichen communities o f southern Cal i f o r -
n i a mountains: an ecological survey r e l a t i v e t o oxidant a i r po l l u t i on . Ecol. Monog. 50: ( i n press).
and 0. C. Taylor. 1 9 7 9 . Pre l iminary s tudies o f the gross photo-
syn the t i c response o f l ichens t o peroxyacetyl- n i t r a t e fumigations. The B ry l og i s t 82: 564-
Skye, E. 1968. Lichens and a i r po l l u t i on . Acta Phyto-
geogr. Suec. 52: 1-114.

- 9
Syers, J. K. and I.K. Iskandar. Tuerk, R. and V. Wirth. 1973. Pedogenetic s ign i f i cance of 1ichens. 1975. The pH dependence o f SOn damage t o -I n The l ichens. V. Ahmadjian and M. E. Hale, l ichens. Oecologia (Be r l i n ) 19: 285-291. eds. p. 225-248. Academic Press, Inc., New York. -9 -and 0. L. Lange.
1974. CO,,~Gaswechsel-Untersuchunqen z u r SOo- Takala, K., P. Kauranen and H. 01 kkonen. ~ e s i s t e d z von Flechten. o e c o l o ~ i a (Ber l .?
1978. F luor ine content o f two 1 ichen species 15: 33-64. i n t he v i c i n i t y o f a f e r t i l i z e r factory . Ann. Bot. Fennici 15: 158-166. Webber, P. J. and J. T. Andrews.
1973. Lichenometry : a commentary. Arct. Tomassini, F. D., P. Lavoie, K. J. Puckett, E. Alp. Res. 5 (4 ) : 295-302.
Nieboer and D. H. S. Richardson. 1977. The e f f e c t o f t ime o f exposure t o sulphur d iox ide on potassium loss from and photosynthesis i n t he l ichen, Cladina rang i f e r i na (L.) Harm. New Phytol . 79: 147-155.
K. J. Puckett, E. Nieboer, D. H. S. Rich- Acknowledgments : This research was supported ardson and B. Grace. by the U.S. Nat ional Science Foundation
1976. Determinat ion of copper, i ron, n icke l , grants DEB-7610244 and DEB-7921953. and sulphur by X-ray fluorescence i n l i chens from the Mackenzi e Val 1 ey, Northwest T e r r i -t o r i es , and the Sudbury D i s t r i c t , Ontario. Can. J. Bot. 54: 1591-1603.

Influence of Air Pollution on Population Dynamics of Forest Insects and on
Tree Mortality1
Donald L. Dahlsten and David L. ~owney2
Abstract: Weakened trees are often predisposed to injury or death by insects, and in forest ecosystems particularly by bark beetles. In the San Bernardino National Forest the interaction between photochemical oxidant weakened ponderosa pine and the western pine beetle (WPB) was examined in de- tail. The major results from this study suggest that oxi- dant damaged trees attacked by WPB produce about the same total brood with lower initial attacks compared to healthier trees. This higher productivity trend is most evident in generation 1 trees Generation 2 trees, both damaged and healthy, are under much greater moisture stress and produce much less WPB brood than generation 1 trees regardless of oxidant damage.
The implication of these results is that in stands with a higher proportion of damaged trees, a given population of WPB could kill more trees and increase at a greater rate than in a stand with a lower proportion of damaged trees. Simulation modelling with these results and other factors that affect ponderosa pine mortality should provide a basis for predicting long term effects of air pollution on the WPB population and pine mortality.
Most of the work on air pollution damage has There have been some studies on the direct focused on direct injury to the plant and effects of air pollution on insects. Feir (1978) Kozlowski (1980) gives a good review of the im- studied the effects of air pollutants on insect pact of air pollution on forest ecosystems. By growth and reproduction and Hillmann and Benton comparison, little has been done on the indirect (1972) looked at the reactions of honey bees to effects such as the predisposing of plants to sulfur dioxide. insects or pathogens. Watt (1969) speculated on the effects of air pollution on population fluc- Bromenshenk (1976, 1978) has studied the tuations of insects and Heagle (1973) reviewed effects of coal-fired power plant emissions on a the interaction between air pollutants and plant variety of insects. In another study, Gilbert parasites. More recently the occurrence of (1971) looked at the indirect effects of air diseases and insect pests of trees in air pollution on several bark inhabiting insects. polluted regions of North America has been Air pollution may be the cause for the scarcity recorded (Dominik 1978). of all orders of insects in New Jersey (Muller
1971).
The effects of insects in pine stands influ- 'presented at the Symposium on Effects of Air enced by air pollution, particularly xylophagous
Pollutants on Mediterranean and Temperate Forest insects, have been studied (Sierpinski 1972, Ecosystems, June 22-27, 1980, Riverside, 1977). In California on the San Bernardino California, U.S.A. Mountains, ponderosa pine shown to have advanced
professor of Entomology and Statistician in symptoms of oxidant injury were most frequently
Entomological Sciences, respectively, University infested and killed by western pine beetle
of California, Berkeley, Calif. (Dendrocbonias'-brevicabis), and mountain pine

- -
beetle (2. ponderosae) (Stark et al. , 1968). The purpose of this study was to look a little closer at the interaction between the western pine beetle and oxidant affected ponderosa pine, to see what effect diseased trees would have on the dynamics of bark beetle populations and to look for dif- ferences between beetle population parameters in diseased and healthy ponderosa pine.
MATERIALS AND METHODS
Field Procedures
Beginning about the first week in July, the San Bernardino mountain areas with substantial ponderosa pine stands were searched for first generation western pine beetle attacked trees. These areas are generally on the southern side of the mountains at about 1500-1800 m elevation, and are areas where high oxidant air pollution levels occur. Second generation trees were located in mid-August to raid-September, depending on the timing of the first generation. Trees under attack were detected by pitch tubes or frags in bark crevices. Attacked trees were checked with an axe at the base to see which species were attacking and to check the staqe. Trees with mixed brood (2. ponderosae and 2. brevicomis) and trees with strip attacks (one side only) were relatively uncommon and were not selected for this study. Trees with spp. in the tops were selected, but sampling for western pine beetle was stopped at the base of infestation of the
z. Each suitable tree located was used in the study until the required number were found (12 per generation in 1973 and 1974, 6 per generation in 1975 and 1976). In the time period allowed for sampling the initial WPB stages there was no practical way to locate all the attacked trees in the area and thenpick a random subsample for the study.
Sampling
Prior to sampling, the major tree character- istics were recorded: location, height, DBH, height of first green branch, and estimated tree damage due to air pollution. The air pollution damage was rated by rating upper and lower crown needle retention, needle condition, needle length, and branch mortality. Needle condition and re- tention were difficult to rate on the sample trees because in many cases, fading and needle loss due to the bark beetle attacks had begun. For 1973 only one needle length rating was made for the entire tree, so the needle length rating for the other years was combined to be consistent in the analysis ( in nearly all cases the upper and lower crown needle length was the same).
Samples for all procedures except where noted later were taken at 1.5 meter intervals over the length of the western pine beetle infestation. Two 88 cm2" discs taken on opposite sides of the tree at 1.5 meter intervals gave the desired level of precision.
The samples were taken from the tree using a modified ortable 4.5 kg gas powered drill D l P r e c o n Multiple Control, Ridgewood, New Jersey). A standard circular hole saw with an 11 can diameter blade was used in the chuck of the drill. This saw cuts an area of approximately 88 cm2.
At each sample height the circumference was measured, then the two discs were cut. With the egg discs it was desirable to take a portion of the xylem with each disc to protect the galleries. After a disc was removed it was examined by the climber for insects, which were identified and called down to the notetaker.
During the last larval sample emergence car- tons were placed on the trees at each sample height. These cartons were not removed until well after brood emergence.
Laboratory Procedures
Once the samples had been returned to Berkeley, they were placed in cool storage until analyzed. X-rayed samples were placed into rearing imme- diately after the radiographs were taken. Basically there were four laboratory analysis procedures for sample discs.
Egg Disc Analysis
To determine egg mortality discs were taken at 3.0 meter intervals approximately two to three weeks after the mass attack period. Sampling at every 3.0 m interval gave 4 or more heights for each tree. Egg discs were taken only once, at the time of the first larval sample. This avoids an additional climbing of the sample tree, assures that maximum egg hatch has occurred and mortality can still be determined.
If xylem tissues still remained on the disc it was carefully removed so that the galleries in the phloem tissue were not disturbed. Gallery length was measured with a metric map reader and all attacks were recorded. Each disc was then exam- ined with a dissection scope (10-20x) along the length of all the adult galleries. Each egg niche was evaluated asempty or as containing a viable egg or nonviable egg in addition to re- cording larval eclosion. The difference between the total number of egg niches recorded and the total number of larvae hatched was calculated as egg mortality.
X-ray Analysis
The first samples to be x-rayed were taken con- currently with the egg discs (egg discs are des- troyed during analysis so they can not be placed into rearing). For generation one, an additional x-ray sample was taken late in the brood develop- ment period, usually in mid to late July. For the second generation, x-ray samples were removed on two later occasions: once in mid-September to October to check for early emergence of brood and

adult re-emergence, and finally in the following spring (March to April) after brood development was nearly complete, but before spring emergence.
The discs to be x-rayed were tkken from cold storage as soon as possible and removed from the plastic bags. The average bark thickness of each disc was measured with a metric caliper and the percentage woodpeckering on each disc was re- corded. The two 88 cm2 discs from each height were placed on one sheet of 8" x 10" Industrial ~odak@AA-2 film and exposed with a picker@ radiograph machine. The samples were then re- bagged and transported to a large storage shed f a rearing (see below).
X-rays were interpreted for western pine beetle, live and dead parent adults, brood adults, pupae, and larvae, parasitoids, predators and miscellaneous. The miscellaneous category included buprestid and cerambycid larvae, weevils, *,and unknowns, of which there were many judging from the rearing of these discs. A light table was used for interpretation and all inclusions were marked with a grease pencil by the interpreter, counted and then erased. A sec- ond interpreter then repeated the process, thereby reducing error and bias. The x-rays from each tree were then stored in envelopes as a per- manent record.
Laboratory Rearing
Each x-rayed disc was placed in a 1/2 gallon ice cream carton with a small 2 or 3 dram glass vial pushed through the lid. Since the insects in the discs are positively phototropic at least in the adult stage, and since very few beetles or other insects bore through the sides of the car- tons, this was a suitable rearing procedure. The cartons were stored in racks in a large unheated warehouse where Berkeley's mild, cool climate provided an ideal rearing environment.
The emerging insects were collected from the vials three to five times per week. Most of the specimens could be identified with a lox hand lens and then recorded on a data sheet by collec- tion date. Only the parasitoids and the western pine beetle were sexed. All specimens have been identified by taxonomic experts and specimens were sent for verification regularly; the col- lection is updated as necessary. Approximately 130 different insects were recorded. In this paper, only WPB, 4 predators (Enoclerus lecontei, Temnochila chlorodia, Aulonium longum, Medetera aldrichii) and 4 parasites (Roptrocerus XJ&-phagorum, Dinotiscus burkei, Eurytoma conica, Coeloides sp. nr. brunneri) are considered.
Sample discs were kept in rearing for nine months and at the end of this period the cartons were examined for any remaining insects; the discs were then discarded. The gallery length and number of attacks were recorded for each disc prior to discarding. Attack holes can be dis- tinguished from emergence holea as they are
oblique to the surface of the bark and often have pitch as a remnant of a pitch tube.
Field Emergence Cartons
As an additional check on emergence, a pro- cedure for rearing in situ on the sample trees was also used.
One quart squat ice cream cartons that cover 88 cm2 of bark were painted silver on the outside and prepared with a screened ventilation hole. Stikem specialawas placed on the inside of the cartons to discourage insects from boring out or from going back into the bark. A groove was cut in the bark with the sampling saw into which the lip of the carton was placed; a nail was driven through the bottom of the carton and into the tree to increase stability.
The cartons were placed on the tree at the time the last x-ray or brood sample was taken (to coincide with the pupal and/or callow adult stages of the brood) and were left in place for up to three months, until well after beetle emergence and the collection of all associated insects had taken place. The second generation cartons were nofc placed on the tree until early spring since cartons do not weather well and therefore cannot be left ort the trees for four to six months over winter. The cartons were split open and examined under the microscope and then discarded. The same insects that-were recorded for laboratory rearing were recorded for the sticky emergence carton.
RESULTS AND DISCUSSION
Attacked tree characteristics and variation by year and generation:
The tree heights of the 71 attacked trees sampled over the four year period varied from 13 to 44 meters, but most trees were in the 20-30 meter range (Fig. 1). Tree DBH, stem volume, and infested bark area varied more widely, but exhibited no significant trends with regard to year and generation. Western pine beetle attack and emergence density, however, both showed sig- nificant differences (P < -05) by generation. Year differences were not significant for these variables except for a lower attack density for generation one trees of 1976 compared to the gen- eration one trees of the other three years.
Effect of tree oxidant damage on beetle popu- lation:
In order to determine if damage to the sample trees had an effect on the western pine beetle populations, the individual components of the damage score were analyzed with respect to beetle attack density. Each individual component of the score can be regarded as an interval level vari- able, so multiple regression techniques were used to determine their relationship to attack density. The technique used was a full screen analysis
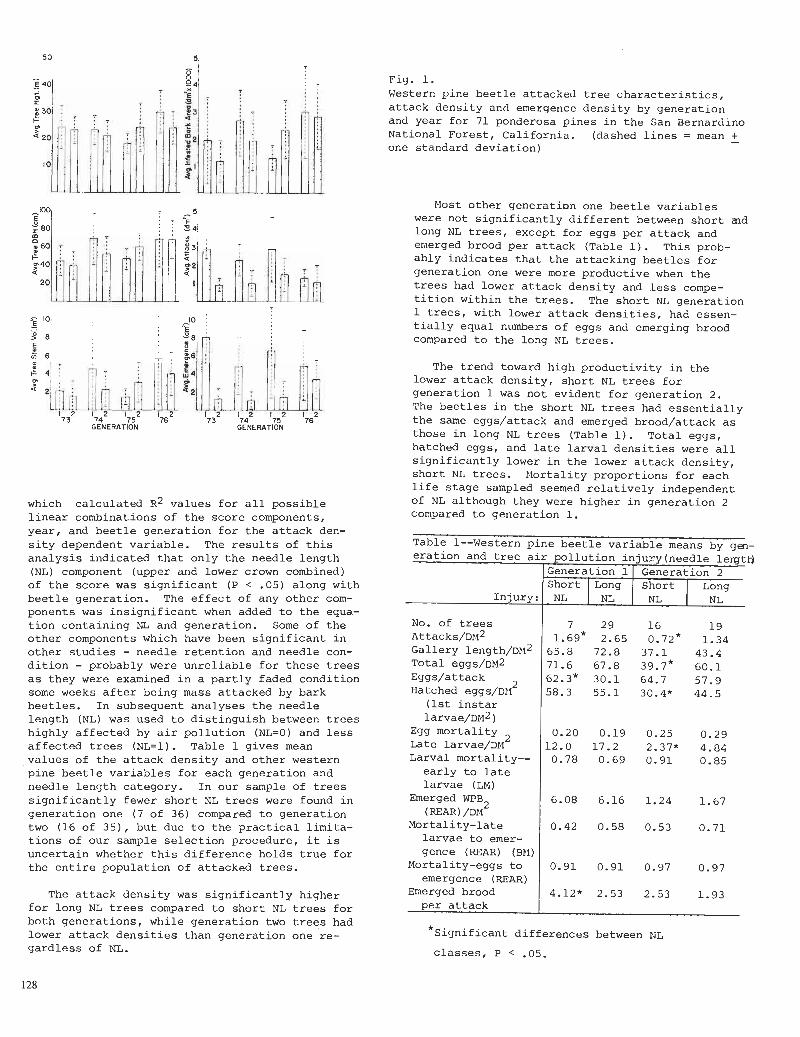
/,* L : ,, ,, , -
1 2 1 2 1 2 1 274 75 74 75
GENERATION GENERATION
which calculated p2 values for all possible linear combinations of the score components, year, and beetle generation for the attack den-sity dependent variable. The results of this analysis indicated that only the needle length (NL) component (upper and lower crown combined) of the score was significant (P < -05) along with beetle generation. The effect of any other com-ponents was insignificant when added to the equa-tion containing NL and generation. Some of the other components which have been significant in other studies - needle retention and needle con-dition - probably were unreliable for these trees as they were examined in a partly faded condition some weeks after being mass attacked by bark beetles. In subsequent analyses the needle length (NL) was used to distinguish between trees highly affected by air pollution (NL=O) and less affected trees (NL=l). Table 1 gives mean values of the attack density and other western pine beetle variables for each generation and needle length category. In our sample of trees significantly fewer short NL trees were found in generation one (7 of 36) compared to generation two (16 of 35), but due to the practical limita-tions of our sample selection procedure, it is uncertain whether this difference holds true for the entire population of attacked trees;
The attack density was significantly higher for long NL trees compared to short NL trees for both generations, while generation two trees had lower attack densities than generation one re-gardless of ML.
Fig. 1. Western pine beetle attacked tree characteristics, attack density and emergence density by generation and year for 71 ponderosa pines in the San Bernardino National Forest, California. (dashed lines = mean + one standard deviation)
Most other generation one beetle variables were not significantly different between short aid long NL trees, except for eggs per attack and emerged brood per attack (Table 1). This prob-ably indicates that the attacking beetles for generation one were more productive when the trees had lower attack density and less compe-tition within the trees. The short NL generation 1 trees, with lower attack densities, had essen-tially equal numbers of eggs and emerging brood compared to the long NL trees.
The trend toward high productivity in the lower attack density, short NL trees for generation 1 was not evident for generation 2. The beetles in the short NL trees had essentially the same eggs/attack and emerged brood/attack as those in long NL trees (Table 1). Total eggs, hatched eggs, and late larval densities were all significantly lower in the lower attack density, short NL trees. Mortality proportions for each life stage sampled seemed relatively independent of NL although they were higher in generation 2 compared to generation 1.
Table I--Western pine beetle variable means by gen-eration and tree a: pollution injury(needle lerytH
Generation 1 Generation 2 Short Long Short Long
Injury: NL ML NL NL
No. of trees ~ t t a c k s / ~ ~ ~ Gallery length/^^^ Total eggs/~M2 Eggs/attack Hatched eggs/Dll
(1st instar larvae/~~2)
Egg mortality Late larvae/DM2
Larval mortality--early to late larvae (LM)
Emerged WPB (REAR)/Dd
Mortality-late larvae to emer-gence (REAR) (BPI)
Mortality-eggs to emergence (REAR)
Emerged brood per attack
*significant differences between NL
classes, P < .05.

Needle length was also used to compare differ- ences in predator and parasite densities for each generation (Table 2). Predators included the 4 common species: Enoclerus lecontei, Temnochila chlorodia, Au1oniw.1 longum, and Medetera aldrichii. Parasites included Roptrocerus e-phagorum, Dinotiscus burkei, Eurytoma conica and -Coeloides sp. nr. brunneri. For generation 1, only emerged (REAR) predators/dm2 were signifi- cantly higher for the long NL trees. For genera- tion 2, initial parasitized larvae/dm2 were higher for short NL trees, but late parasitized larvae/ dm2 were higher for long NL trees. A much larger increase in numbers of parasitized larvae occurred for the gen. 2, long NL trees. Increased woodpeckering, along with a higher final larval density for the long NL trees may explain the increased numbers of parasitized larvae. A similar proportion (about 1%) of initial larvae were parasitized in both cases.
Table 2--WPB predator and parasite means by gen- eration and tree air pollution injury class (needle length).
Generation 1 Generation 2 Short Long Short Long
Injury: NL NL NL NL 2
No. trees 7 29 16 19 Initial pred./~~2 0.962 1.910 0.53 0.30 Initial para- 0.20 0.36 0.18* 0.08 sitized lar./DM
Late predators/~~2 1.83 2.75 0.80 0.84 Late parasitized 0.56 0.61 0.26* 0.48 larvae/~ll^
Emerged (REAR) 0.73* 1.50 0.25 0.27 predators/~~^
Eraer.(REAR) para/D~2 0.48 0.42 0.22 0.28 % area samples wood-0.1% peckered
0.8% 7.0% 11.0%
*Significant difference between means of smog injury classes, P < .05 .
Table 3--Ozone effects table: western pine beetle in ponderosa pine.
tree mort.
low long 4.0 3.9 consbat constant or de- or de- creasing creasing
high short 2.4 3.7 increaskq increasing
Motes: (1) Effect depends on individual tree character-
istics. (2) Based on both generations combined and multi-
plying female adult attacks by two to account for males.
(3) Based on both generations combined. (4) Assuming constant no. of damaged pines/stand.
The consequences of bark beetle activity in an area are summarized in Table 3 and show that attack rates are lower on oxidant affected trees, but the output of brood is essentially the sane in healthy and diseased trees. Bark beetles, assuming no direct deleterious effects of air pollution,should increase in areas with high ozone damage and tree mortality will also in- crease in these areas. The possible interactions of air pollution, ponderosa pine, and western pine beetle and the effects on forest succession are shown in Table 4.
Table 4~Interaction table: air pollution, ponderosa pine, and western pine beetle.
Causes a change in VPB 1 Ponderosa Forest pop. pine mortal- succes- level ity caused sion
by WPB An increase In: air pollution + + + soil water ? ? ? foliar injury ++ + + soil nutrients ? ? ? mature tree growth ? ? ? root disease + + + forest succession - - 0 WPB pop. level 0 ++ + pine mort. caused by WPB ? 0 +
Key: 0 = unrelated or not possible + = moderate increase
++ = large increase - = a large decrease ? = unknown, needs more investigation
LITERATURE CITED
Kozlowski, T. T. 1980. Impacts of air pollution on forest eco- systems. BioScience 30:88-93.
Watt, K.E.F. 1969. Prospective effects of air pollution on insects. Can. Entomol. 101:1235-1238.
Heagle, A. S. 1973. Interactions between air pollutants and plant parasites. Annu. Rev. Phytopathol. 11: 365-388.
Dominik, J. 1978. Investigations of the occurrence of diseases and insect pests of North American trees in the regions of different degrees of air pollution caused by the industry. Final Report of Investigations covering the period of August 1, 1973-June 30, 1978. Warsaw: Sin., 1978. 91, 21 leaves: 111.

Feir, D. 1978. Effects of air pollutants on insect growth and reproduction. Physiologist 21:36.
Hillman, R. C., and A. W. Benton. 1972. Biological effects of air pollution on insects emphasizing the reactions of the honey bee, e m e l l i f e r a to sulfur dioxide. J. Elisha Mitchell Sci. SOC. 88:195.
Muller, J. 1971. Is air pollution responsible for melanism in Lepidoptera and for scarcity of all orders of insects in New Jersey, U.S.A. J. Res. Lepid. 10:189-190.
Gilbert, 0. L. 1971. Some indirect effects of air pollution on bark living invertebrates. J. Appl. Ecol. 8: 77-84.
Bro~~ienshenk,J. J. 1976. Biological impact of air pollution on insects. Ecol. Res. Ser. 600/3-76-013: 295-312.
Bromenshenk, J. J. 1978. Investigation of the impact of coal-fired power plant emissions upon insects: entomo- logical studies at the zonal air pollution systems. Ecol. Res. Ser. 600/3-78-021:473-507.
Sierpinski, 2. 1972. The economic importance of secondary noxious insects of pine on territories with chronic influence of industrial air pollution. Mitt. Forstl. Bundes-Versuchsanst Wein. 97: 609-615.
Sierpinski, Z. 1977. Economic significance of noxious insects in pine stands under the permanent impact of the industrial air pollution. Slyvan. .64:59-71.
stark, R.W., P.R. Miller, F.W. Cobb, Jr., D.L. Wood, and J.R. Parmeter, Jr. 1968. Photochem- ical oxidant injury and bark beetle (Coleop- tera: Scolytidae) infestation of ponderosa pine. I. Incidence of bark beetle infestation in injured trees. Hilgardia 39:121-126.

Air Pollutants and Their Effects on Wildlife with Particular Reference to the House Wren (Delichon urbica)'
James R. Newman, P ~ . D . ~
Abstract : In ju ry and death t o w i l d l i f e from a i r emissions have been recorded s ince before the t u r n of the century. A i r po l lu t ion can d i r e c t l y a f f e c t w i l d l i f e ( i . e . , cause dea th ) , or can i n d i r e c t l y a f f e c t w i l d l i f e ( i . e . , cause h a b i t a t l o s s ) . A recent inves t iga t ion on the chronic and s u b l e t h a l e f f e c t s of a i r po l lu t ion on the house mart in (Delichon u rb ica ) has shown t h a t a i r v
emissions can s i g n i f i c a n t l y a f f e c t the nest ing ecology of t h i s spec ies .
INTRODUCTION emissions ( P r e l l 1936; Hais and Masek 1969). Eighty-five percent o f the w i l d l i f e inc iden t s
Since the end of the 19th cen tu ry , over have been recorded i n the l a s t 25 y e a r s . The 100 a i r p o l l u t i o n episodes have been repor ted inc rease i n the number o f w i l d l i f e inc iden t s involving i n j u r y and death t o animals (Newnan appears t o be due no t on ly t o an increase in a i r 1975, 1979). Although the major i ty o f the emissions but t o a b e t t e r understanding of a i r i n c i d e n t s involved domesticated animals, a number p o l l u t i o n e f f e c t s , broader communications, and a o f i n c i d e n t s have been repor ted f o r w i l d l i f e g r e a t e r i n t e r e s t i n repor t ing such problems. s i n c e the 1880's ( t a b l e 1 ) . Overal l , the few r e p o r t s involving w i l d l i f e appear t o be more a Table I-The number of reported a i r p o l l u t i o n func t ion o f economic b i a s t o repor t i n j u r y and i n c i d e n t s involving w i l d l i f e and af f ec ted groups dea th o f domestic animals than some innate (adapted Newman 1979) r e s i s t a n c e o f w i l d l i f e t o a i r p o l l u t i o n . For example, i n the e a r l y 1900's a r sen ic emissions from a smelter caused the i n j u r y and death o f Number of I s e v e r a l thousand c a t t l e and horses in r u r a l Wildl i fe Groups consumer Groups Montana arkin ins and Swain 1908). No mention was Period I n c i 1 Game \ one game IPrim. [secondary made o f i n j u r y o r death t o deer o r o ther w i l d l i f e den t s 1 1 p l e n t i f u l i n the a r e a , a 1 though deer and r a b b i t s a r e known t o be ve ry s e n s i t i v e to a r sen ic Before 1 1 1
1900
1900-1950
p r e s e n t e d a t t h e Symposium on E f f e c t s of Air P o l l u t a n t s on Mediterranean and Temperate Fores t 1951-1970
Ecosystems, June 22-27, 1980, Rivers ide , 1971 t o
C a l i f o r n i a , U.S.A. Present
^sen2ior Eco log i s t , P ro jec t Operations, Environmental Science and Engineering, Inc ., TOTAL P.O. Box ESE, Ga inesv i l l e , F lo r ida 32602.

Recent Federal l e g i s l a t i o n , namely the 1977 Clean A i r Act Amendments, now r e q u i r e formal c o n s i d e r a t i o n o f the e f f e c t s o f a i r emissions on w i l d l i f e and o the r components o f eco log ica l systems.
The purpose o f t h i s paper i s t o summarize some o f the known e f f e c t s o f a i r emissions on w i l d l i f e and present p re l iminary f i n d i n g s on the e c o l o g i c a l e f f e c t s o f a i r emissions on the house m a r t i n ( ~ e l i c h o n u r b i c a ) , an insec t ivorous b i r d . For t h i s paper , d i s c u s s i o n i s l imi ted t o t e r r e s t r i a l wild1 i f e and v e r t e b r a t e consumers . Previous speakers h e r e discussed the e f f e c t s on i n v e r t e b r a t e consumers.
GENERAL EFFECTS OF A I R EMISSIONS ON WILDLIFE
One of the e a r l i e s t w i l d l i f e inc iden t s (1887) involves t h e death of f a l low deer (Dama dama) from a r s e n i c emissions from a s i l v e r foundry i n Germany (Tendron, 1964). As e a r l y a s 1900 t h e Royal Soc ie ty of London was charged to i n v e s t i -g a t e t h e phenomenon of i n d u s t r i a l melanism. This i n c i d e n t i s recognized more f o r i t s demonstration o f "natura l" s e l e c t i o n than a s an environmental problem. Recently, g e n e t i c changes i n small mammals because of a i r p o l l u t i o n have been repor ted (Newman 1980).
One of t h e ' e a r l i e s t d e t a i l e d d e s c r i p t i o n s of t h e harmful e f f e c t s of a i r emissions on w i l d l i f e i s from Germany. Arsenic emissions were the cause o f widespread death of game animals i n the Tharandt f o r e s t of Germany i n 1936. S i x t y t o seventy percent of t h e red deer (Cervus e iephus ) , r o e dee r (Capreolus capreo lus ) , and wild r a b b i t s ( ~ r ~ c t o l a g u scun icu lus ) d ied. The deer exhibi ted d e f e c t i v e h a i r growth and a n t l e r formation, c i r r h o s i s of t h e l i v e r and spleen, and emaciation ( P r e l l 1936).
Adverse e f f e c t s of a i r emissions on w i l d l i f e have been repor ted from North America, Euro-pe, and Asia , and have included game and nongame animals; b i r d s and mammals; primary and secondary consumers; and he rb ivores , omnivores, and ca rn ivores ( t a b l e 1 ) . The e f f e c t s on w i l d l i f e have included d ie -o f f s and o the r population r e d u c t i o n s , phys io log ica l abnormal i t i e s , d i s e a s e , phys ica l i n j u r y , and bioaccumulation. The m a j o r i t y of w i l d l i f e i n c i d e n t s repor ted involve dee r and small b i r d s (~ewman 1979). These groups a r e not n e c e s s a r i l y more s e n s i t i v e but a r e more l i k e l y t o be monitored, e s p e c i a l l y dee r , which i s a widely managed game spec ies . The s t a t u s of smal l b i r d s i s not only of s c i e n t i f i c concern, bu t a l s o of more widespread publ ic i n t e r e s t .
Pathways of contamination by a i r emissions f o r w i l d l i f e a r e : i n h a l a t i o n , adsorp t ion , and i n g e s t i o n . Wellings (1970) repor ted the occurrence of pulmonary an th racos i s i n urban sparrows (Passe r domesticus) compared to r u r a l popu la t ions i n C a l i f o r n i a . One of t h e most s t a r t l i n g examples of contamination by i n h a l a t i o n involved the die-off of 200 t o 500 songbirds near
a B r i t i s h Columbia pulp m i l l which emitted high concen t ra t ions of H2S and o the r p o l l u t a n t s . The dead b i r d s showed i n t e r n a l hemorrhaging i n t h e lungs and l i v e r . 3
Adsorption of a i r emissions involves the adhesion of gases o r p a r t i c u l a t e s t o the e x t e r n a l s u r f a c e s o r e x t e r n a l membranes, e .g . , cornea of eyes. Light (1973) r e p o r t s a high incidence of b i i n d n e s s i n bighorn sheep (*canadensis) found i n mountain a r e a s of Ca l i fo rn ia with heavy oxidant l e v e l s . Oxidants a r e known eye i r r i t a n t s . I n v e s t i g a t o r s i n ~ z e c h o s l o v a k i a ~ have observed e a r l y aging of the cornea i n ha res ( ~ e p u s europaeus) found i n a reas with heavy SO2 and p a r t i c u l a t e depos i t ion from power p l a n t s and o the r i n d u s t r i e s .
Inges t ion i s the most commonly repor ted mode o f contamination f o r w i l d l i f e . I n j u r y and death t o w i l d l i f e from inges t ion o f contaminated food and water have been repor ted f o r numerous animal spec ies including r a b b i t s and deer from a r s e n i c emissions ( P r e l l 19361, dee r from f l u o r i d e emissions (Karstad 1967; Robinette and o t h e r s 1956; Newman and Yu 19761, and sparrows from cadmium emissions (Nishino and o t h e r s 1973). There a r e many r e p o r t s of the bioaccumulation of a i r p o l l u t a n t s i n w i l d l i f e . With a few excep t ions , the e f f e c t s of t h i s bioaccumulation a r e not known (Newman 1980).
A i r emissions may not be l e t h a l t o w i l d l i f e but may lower the h e a l t h and r e s i s t a n c e of w i l d l i f e so t h a t n a t u r a l s t r e s s such a s cold and shor tage of food may r e s u l t i n f u r t h e r i n j u r y o r dea th . Studies on the h e a l t h of wild hares (Lepus europaeus) l i v i n g i n a reas of h igh a i r p o l l u t i o n i n Czechoslovakia showed physiological responses s i m i l a r t o animals with i n f e c t i o n s o r a l l e r g i c r e a c t i o n s . There were a l s o changes i n t h e normal age s t r u c t u r e of the ha re popula t ions ( ~ o v a k o v a 1969). The general h e a l t h of deer popula t ions a f f e c t e d by f l u o r i d e emissions i s poor ( ~ o b i n e t t e and o t h e r s 1957; Karstad 1967; Newman and Yu 1976; Newman and Murphy 1979). The s u r v i v a l of these deer dur ing times of n a t u r a l s t r e s s i s quest ionable . Hais and Masek (19691, r e p o r t t h a t a r s e n i c contamination of red and roe dee r caused emaciation and loss of h a i r so t h a t many of the animals f roze t o death dur ing the win te r .
I n j u r y o r death t o vege ta t ion caused by a i r emissions can have s i g n i f i c a n t i n d i r e c t e f f e c t s on w i l d l i f e . The l o s s of food resources and h a b i t a t can occur a s the r e s u l t of i n j u r y o r
3 ~ n p u b l i s h e d r e p o r t , R.D. Harris 1971. Birds Collected (Die Off) a t Pr ince Rupert , B.C. Canadian Wi ld l i f e Service , Pr ince Rupert , B r i t i s h Columbia.
4 ~ e r s o n a l communication from E l i s k a Novakova, I n s t i t u t e o f Landscape Ecology, Prague, Czechoslovakia.

- -
dea th t o v e g e t a t i o n which provides cover , r ep roduc t ive h a b i t a t , o r food f o r w i l d l i f e . Many examples e x i s t o f e x t e n s i v e damage t o ecosystems from a i r emiss ions . Large a r e a s o f the mixed con i fe rous f o r e s t o f southern Ca l i fo rn ia (over 40,000 h e c t a r e s ) have been k i l l e d o r in jured from photochemical ox idan t s ( ~ a y l o r 1973). No o v e r a l l assessment has been made a s t o the l o s s o f w i l d l i f e whose f o r e s t h a b i t a t has been des t royed. S imi la r l a r g e w i l d l i f e h a b i t a t l o s s e s have occurred i n Montana, Tennessee, B r i t i s h Columbia, and Onta r io , Canada (~ewman 1980).
ECOLOGICAL EFFECTS OF AIR EM IS S IONS ON THE HOUSE MARTIN
With few excep t ions , t h e r e i s l i t t l e informat ion a v a i l a b l e on the e f f e c t s o f chronic exposure o f w i l d l i f e t o a i r emissions (Newman 1980). Since 1976, RNDr. E l i ska Novakova, CSC, from the I n s t i t u t e o f Landscape Ecology, Prague, and myself have been s tudying the eco log ica l responses o f the house mart in (Delichon u r b i c a ) t o chronic a i r p o l l u t i o n . In t h i s por t ion of the t a l k I would l i k e t o p resen t some o f our p re l iminary f i n d i n g s .
The house mar t in belongs t o the family Hirundinidae . It i s an insec t ivorous b i r d lAiich feeds p r imar i ly on aphids and d i p t e r e a n s . A migra to ry s p e c i e s , t h e mar t in summers throughout Europe and win te r s i n Af r i ca . As a c o l o n i a l n e s t i n g s p e c i e s , i t b u i l d s mud n e s t s on the w a l l s o f b u i l d i n g s . The n e s t i n g l o c a t i o n s a r e used each y e a r , o f t e n by t h e same b i r d s .
Previous s t u d i e s have ind ica ted t h a t 2. u r b i c a i s s e n s i t i v e t o a i r p o l l u t i o n . Fer iancova- Masarova and Kal ivodova (1965) observed changes i n the s p e c i e s d i v e r s i t y o f b i r d s , including the house m a r t i n , i n the a r e a o f an aluminum p lan t i n Czechoslovakia. Cramp and Gooders (1967) observed a c o r r e l a t i o n with increased nes t ing of D . u r b i c a and a decrease i n smoke p o l l u t i o n in London. A follow-up s tudy (~ewman 1977) in the v i c i n i t y o f t h e same aluminum p lan t s tudied by Feriancova-Masarova and Kalivodova i n 1965 showed t h a t 2. u r b i c a decreased i t s n e s t i n g d e n s i t y with inc reased a i r emiss ions .
In 1978 d i r e c t coun t s o f a c t i v e n e s t s were made i n 141 v i l l a g e s and towns loca ted i n i n d u s t r i a l i z e d (contaminated) and non- indus t r i a l i zed ( c o n t r o l or. background) a r e a s . Of t h e 141 n e s t i n g l o c a l i t i e s censused, 101 v i l l a g e s and towns were inf luenced by a i r emiss ions from var ious i n d u s t r i a l sources . Power p l a n t s and assoc ia ted su r face mines, chemical p l a n t s , l o c a l hea t ing p l a n t s ( coa l burn ing) , cement p l a n t s , and general urban emiss ions sources were the major a i r emission i n f l u e n c e s . Censusing was conducted a t pre-determined upwind and downwind l o c a t i o n s from an emiss ion source . This census represented obse rva t ions o f over 20,000 b u i l d i n g s . Records were kept o f t h e number o f a c t i v e n e s t s . A
s tandard n e s t i n g d e n s i t y measure was der ived a s we l l a s measures o f t h e occupancy o f a n e s t i n g l o c a t i o n and colony s i z e . The environmental a t t r i b u t e s o f each n e s t i n g l o c a l i t y was c h a r a c t e r i z e d . Comparisons o f background n e s t i n g l o c a l i t i e s were then made t o e c o l o g i c a l l y s i m i l a r , bu t contaminated, n e s t i n g l o c a l i t i e s .
The mean n e s t i n g d e n s i t y o f 2. u r b i c a from c o n t r o l a r e a s was 0.56521.94 n e s t s per s i d e . The range i n colony s i z e was 1 t o 33 a c t i v e n e s t s per occupied s i d e , wi th an average colony s i z e o f 2.721.1 a c t i v e n e s t s per occupied s i d e . The average occupancy ( t h e number o f occupied s i d e s per s u i t a b l e s ide ) f o r the 32 background n e s t i n g a r e a s was 2 1 . 0 3 . 1 pe rcen t .
The o v e r a l l n e s t i n g d e n s i t y f o r 1.u r b i c a from contaminated a r e a s was 0 .3933.734 a c t i v e n e s t s per s u i t a b l e n e s t i n g s i d e . This n e s t i n g d e n s i t y (30 pe rcen t ) was s i g n i f i c a n t l y lower
= 3.674, df = 8968) when compared t o^0.002 background nes t ing s i t e s . Occupancy was on ly 13.6 pe rcen t . Attempts a t n e s t i n g were a l s o lower , wi th on ly 0.74 a t t empts per contaminated v i l l a g e compared t o 1.12 a t t empts per background v i l l a g e . Colony s i z e was s i g n i f i c a n t l y lower (2.1+1.5 a c t i v e n e s t s per occupied s i d e ) . -
The house mart in responds a l s o t o the l e v e l of a i r emissions. The o v e r a l l nes t ing d e n s i t y f o r downwind l o c a t i o n s was 0.23720.378 (n = 4891) n e s t s per s i d e compared t o 0.41521.603 (n = 1969) n e s t s per s i d e f o r upwind l o c a l i t i e s . This d i f f e r e n c e was s i g n i f i c a n t (P < 0.001; t = 7.263; d f = 6808). The o v e r a l l upwind nes t ing d e n s i t y was not s i g n i f i c a n t l y d i f f e r e n t from background a r e a s .
Colony s i z e and occupancy a l s o changed with proximity t o emission sources . Upwind colony s i z e s averaged 2.521.0 n e s t s per occupied s i d e compared t o 1.820.8 n e s t s per occupied s i d e f o r downwind loca t ions . These d i f f e r e n c e s were s i g n i f i c a n t (P < 0.001; t = 3.492 d f = 99). Upwind occupancy (18.828.9 percent) was a l s o s i g n i f i c a n t l y h igher (P < 0.001, t = 5.70, d f = 99) when compared t o downwind occupancy (9.1+7.7 pe rcen t ) . -
The n e s t i n g d e n s i t y f o r downwind loca t ions dec reases s i g n i f i c a n t l y a s the d i s t a n c e to the emission source decreases ( t a b l e 2) . For upwind l o c a t i o n s , the c l o s e r t o the source , the higher t h e nes t ing d e n s i t y ; f o r increased d i s t a n c e s , n e s t i n g d e n s i t i e s decl ined. The upwind nes t ing d e n s i t i e s c l o s e s t t o emission sources were not s i g n i f i c a n t l y d i f f e r e n t from background nes t ing d e n s i t i e s . Colony s i z e was lower a t a l l downwind d i s t a n c e s , but only s i g n i f i c a n t l y lower a t t h e two c l o s e s t d i s t a n c e s . Occupancy was s i g n i f i -c a n t l y lower f o r a l l d i s t a n c e s censused downwind. For the two c l o s e s t upwind d i s t a n c e groups (ou t t o 6 tan) , bo th colony s i z e and occupancy were no t s i g n i f i c a n t l y d i f f e r e n t from background cond i t ions .

- -
- -
Table 2--Comparison of nes t ing dens i ty of D.- -u r b i c a a t var ious d i s t ances upwind and downwind from emission sources
Sample S i z e Me an Percent
Locat i o n ( , N ~ . of Sides)
Nesting Densi ty
Difference From Background
Downwind I
Upwind I
S i g n i f i c a n t , P < 0.025. ^ot S i g n i f i c a n t , P<O.O5
The a i r d i s p e r s i o n p a t t e r n s a s soc ia ted with t h e s t a c k h e i g h t s inf luenced the observed e f f e c t s . For sources w i t h h igh s t a c k s ( g r e a t e r than 100 meters) the adverse e f f e c t s on n e s t i n g were observed i n both downwind and upwind l o c a l i t i e s . For downwind l o c a t i o n s , t h e lowest n e s t i n g d e n s i t y d id not occur c l o s e s t t o the emission source , bu t a t an in termediate d i s t a n c e ( 3 t o 6 km). Nesting d e n s i t y was s i g n i f i c a n t l y . depressed out t o 12 km. For low s t a c k sources ( l e s s than 100 m e t e r s ) , t he lowest n e s t i n g d e n s i t y , colony s i z e , and occupancy occurred a t t h e c l o s e s t d i s t a n c e s (0 t o 3 km). Average n e s t i n g d e n s i t i e s were 20 pe rcen t o f background. In c o n t r a s t t o h i g h s t a c k sources , t h e n e s t i n g d e n s i t y inf luenced by low s t a c k sources was near normal a t t h e 9 .1 t o 12 km d i s t a n c e c a t e g o r i e s . For upwind l o c a l i t i e s , emiss ions from high s t a c k sources have an adverse in f luence , e s p e c i a l l y c l o s e t o the source . For low s t a c k l o c a l i t i e s , upwind n e s t i n g d e n s i t i e s were e i t h e r a t o r above background l e v e l s a t a l l d i s t a n c e s censused.
When the house mar t in n e s t s i n optimum nes t ing c o n d i t i o n s , such a s apartment b u i l d i n g s and f e e d l o t s i n proximity t o wa te r , the e f f e c t s o f a i r emiss ions were m i t i g a t e d . The g r e a t e s t adverse e f f e c t s on the n e s t i n g ecology o f D . u r b i c a were observed surrounding power p l a n t s and open s u r f a c e mines. Chemical p l a n t emissions had the n e x t most s i g n i f i c a n t e f f e c t on the house m a r t i n .
In summary, s u b l e t h a l and chronic l e v e l s o f a i r emissions from i n d u s t r i a l sources , such a s power p l a n t s and chemical p l a n t s , adverse ly a f f e c t the n e s t i n g ecology o f 2. u r b i c a , inc lud ing a r educ t ion o f the n e s t i n g d e n s i t y , colony s i z e , and occupancy o f t h i s s p e c i e s . The g r e a t e s t e f f e c t s on the n e s t i n g p a t t e r n o f D . u r b i c a were observed i n a r e a s downwind from t h e emission sources and a t decreas ing d i s t a n c e s from the a i r emission sources ( f i g . 1 ) . For low s t a c k emission sources , g r e a t e s t e f f e c t s were observed i n the immediate v i c i n i t y o f the s t acks where the h ighes t emission l e v e l s a r e expected t o occur . For h igh s t a c k s o u r c e s , g r e a t e s t e f f e c t s were observed a t in te rmedia te d i s t a n c e s from the s t a c k s and a g r e a t e r d i s t a n c e than observed i n low s t a c k l o c a l i t i e s .
Evidence suggests t h a t w i l d l i f e popula t ions l i v i n g i n optimal h a b i t a t c o n d i t i o n s a r e l e s s a f f e c t e d by a i r emissions than w i l d l i f e popula- t i o n s l i v i n g i n l e s s than optimal cond i t ions . This obse rva t ion has p a r t i c u l a r importance f o r t h e management and p rese rva t ion o f endangered spec ies .
Overal l the e f f e c t s o f a i r p o l l u t a n t s on w i l d l i f e can be dramat ic , such a s t h e d i e - o f f s , o r q u i t e s u b t l e , such a s s h i f t s i n the age s t r u c t u r e o f popula t ions . In many c a s e s the e f f e c t s have been d e b i l i t a t i n g i n j u r i e s , o f t e n c o n t r i b u t i n g to the dea th o f animals dur ing pe r iods o f n a t u r a l s t r e s s . A i r emissions can a l s o reduce w i l d l i f e popula t ions n o t on ly d i r e c t l y , bu t i n d i r e c t l y through l o s s o f h a b i t a t .
Acknowledgments: Specia l r ecogn i t ion is given t o RNDr. El i ska Novakova, CSc f o r c o l l a b o r a t i o n on the mar t in s tudy . I wish t o thank National Academy o f Science and the Czechoslovak Academy o f Science f o r support o f the b i r d s tudy and Environmental Sc ience and Engineering f o r a s s i s t a n c e i n the manuscript p repara t ion .
LITERATURE CITED
Cramp, S. , and J. Gooders. 1967. The r e t u r n of the house mart in .
Lond. Bird Rep., No. 31:93-98.
Feriancova-Masarova, Z . , and E. Kalivodova. 1965. Niekolko poznamok vplyve fluorovych
exhala tov v o k o l i Hl inikarne v Z i a r i nad Hronom na k v a n t i t u hn iezd iac ich vtakov (The e f f e c t s o f exha la t ions from the aluminum p l a n t i n Ziar nad Hronom on the spectrum of b i r d ' species i n the v i c i n i t y of the p l a n t ) . Biologia ( B r a t i s l a v a ) 20: 341-346.
Hais, K., and J. Masek. 1969. Vcinky nekterych exha lac i na hospodarska
z v i r a t a ( E f f e c t s of some exha la t ions on a g r i c u l t u r a l animals) . Ochr. Ovzduzi 3: 122-125.

Harkins, W.D., and R.E. Swain. on a Western Coniferous Forest Ecosystem. 1908. The chronic a r s e n i c a l poisoning of O.C. Taylor, ed. P a r t B, pp. 1-12. Task B
herbivorous animals. Rep. A i r P o l l u t i o n Research Center, J. Am. Chem. Soc. 30: 928-946. Unive r s i ty of C a l i f o r n i a , Rivers ide ,
C a l i f o r n i a . Kars tad, L.
1967. F luoros i s i n deer . Newman, J. R, ( ~ d o c o i l e u s v i r g i n i a n u s ) . 1977. S e n s i t i v i t y o f the house mart in
Bul l . 'Wildl. Dis. Assoc. 3:42-46. ( ~ e l i c h o n u rb ica ) t o f l u o r i d e emissions. F luor ide 100:'73-76.
L igh t , J.T. 1973. The e f f e c t s of oxidant a i r p o l l u t i o n on Newman, J.R.
f o r e s t ecosystems of t h e San Bernardino 1979. E f f e c t s of i n d u s t r i a l a i r p o l l u t i o n on Mountains. 2 Oxidant Air P o l l u t i o n E f f e c t s w i l d l i f e . Biol . Conserv. 15:181-190.






![Gaseous exchange [all]](https://img.pdfslide.us/doc/110x75/54bdd34f4a79592e1a8b463d/gaseous-exchange-all.jpg)