Embed Size (px)
Citation preview
TECHNICAL REPORT
www.nature.com/naturebiotechnology • JANUARY 2003 • VOLUME 21 • nature biotechnology 93
Screening for gene functionin chicken embryo usingRNAi and electroporationVladimir Pekarik, Dimitris Bourikas, Nicola Miglino,Pascal Joset, Stephan Preiswerk, and Esther T. Stoeckli*
Published online 16 December 2002; doi:10.1038/nbt770
In the postgenomic era the elucidation of the physiological func-tion of genes has become the rate-limiting step in the quest tounderstand the development and function of living organisms.Gene functions cannot be determined by high-throughput meth-ods but require analysis in the context of the entire organism.This is particularly true in the developing vertebrate nervous sys-tem1. Because of its easy accessibility in the egg, the chickenembryo has been the model of choice for developmental in vivostudies. However, its usefulness has been hampered by a lack ofmethods for genetic manipulation. Here we describe anapproach that could compensate for this disadvantage. By com-bining gene silencing by dsRNA2 (through RNA interference,RNAi) with in ovo electroporation3,4, we developed an efficientmethod to induce loss of gene function in vivo during the devel-opment of the chicken CNS. This method opens new possibilitiesfor studying gene function not only by gain-of-function but also byloss-of-function approaches and therefore represents a new toolfor functional genomics.
The mouse is by far the most widely used vertebrate model systemfor the functional analysis of genes in vivo, allowing for both gain-and loss-of-function approaches. However, many genes involved indevelopmental or functional aspects of the nervous system cannot bestudied because their additional role in early developmental process-es means that loss of function either leads to early embryonic lethal-ity or is compensated for by other genes. As a solution, interferencewith gene function in a locally and temporally defined manner isrequired, and to some extent this can be achieved by sophisticatedgene knockout techniques5. Yet there remain problems with high costand the large amount of time required to gain insight into the func-tional role of a gene of interest. In the postgenomic era, the function-al analysis of genes rather than the identification of candidate geneshas become rate limiting. Thus, a pre-selection of functionally rele-vant genes among all the candidates is crucial. Furthermore, thereare as yet no tools available for temporal control that would allowgene regulation in a very narrow time window, as is required, forinstance, to analyze developmental aspects of gene regulation.
A useful alternative is provided by in vivo studies in chickenembryos, where gene function has been successfully studied at the pro-tein level6–9. In vivo injections of antibodies can be targeted precisely intime and space. Furthermore, the development of in ovo electropora-tion techniques offers a straightforward method for conducting gain-of-function experiments involving local overexpression of a gene ofinterest3,4,10,11. So far, loss-of-function experiments have been more
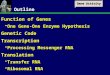
problematic and have depended on the availability of dominant-negative forms of the gene product. By combining RNAi and in ovoelectroporation, we developed a method for gene silencing in vivo.
The injection of dsRNA into cells as a means to interfere with genefunction was first applied a few years ago in the nematodeCaenorhabditis elegans2. Reports about the specificity of RNAi in ver-tebrates followed12–14. However, in both zebrafish and mouse, RNAihas been used only at the one-cell stage, and there have been noreports about the feasibility of the method in older embryos. Becausethis is a prerequisite for functional analyses of many genes, we triedto adapt the method to older chicken embryos. Electroporation isextremely efficient at enhancing uptake of DNA or RNA into verte-brate cells in ovo3,4,11 (Fig. 1). Using the expression of yellow fluores-cent protein (YFP) as a parameter to measure uptake of nucleicacids, we calculated that ∼ 60% of the cells in the electroporated areaof the spinal cord were transfected (Fig. 1B). In contrast, the injec-tion of the YFP plasmid without electroporation resulted in no oronly very few YFP-expressing cells (Fig. 1C). The concomitant injec-tion of YFP dsRNA and the plasmid encoding YFP resulted in a
University of Zurich, Institute of Zoology, Winterthurerstrasse 190, CH-8057Zurich, Switzerland. *Corresponding author ([email protected]).
Figure 1. Demonstration of the efficiency of RNAi for interference withgene function in vivo. (A) Transient loss of gene function in vivo isachieved by injection of dsRNA into the central canal of the spinal cordfollowed by electroporation to increase uptake of the injected dsRNA into cells. (B) Using the same paradigm for the injection of a plasmidencoding yellow fluorescent protein (YFP), we calculated that 60% of the cells in the electroporated area of the spinal cord expressed thetransgene. (C) Injection of the plasmid without electroporation did notresult in YFP expression. (D) The concomitant injection andelectroporation of the YFP plasmid with YFP dsRNA drastically reducedthe expression of YFP. (E–G) RNAi specifically interferes with theexpression of the corresponding protein. The injection andelectroporation of axonin-1 dsRNA reduced axonin-1 levels on theelectroporated side (indicated by open arrowheads on the right sidecompared with the normal levels indicated by white arrowheads on theleft side of the spinal cord; E). However, there was no change in theexpression of NrCAM in the same section (F) and NgCAM shown in anadjacent section (G). Embryos were injected with axonin-1 dsRNA andelectroporated. After 38 h, embryos were killed, fixed in 4% (wt/vol)paraformaldehyde, and used for cryostat sectioning. The relativefluorescence intensities for the electroporated and the control side of thespinal cord were measured. In ovo RNAi with axonin-1 resulted in a 27.7± 4.0% decrease after 38 h (n = 3). Values in different sections rangedfrom 8.6% to 40.0%. No decrease was measured in control embryosinjected and electroporated with buffer only (n = 2). Bar, 100 µm in (B–D),75 µm in (E–G).
A
B C D
E F G©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reb
iote
chn
olo
gy
TECHNICAL REPORT
nature biotechnology • VOLUME 21 • JANUARY 2003 • www.nature.com/naturebiotechnology94
substantial decrease in YFP expression (Fig. 1D). The concomitantinjection of an unrelated dsRNA did not interfere with YFP expres-sion (data not shown). Furthermore, the injection of either sense orantisense RNA together with the YFP plasmid did not have a sub-stantial effect on YFP expression (see Supplementary Fig. 1 online),although there was a slight tendency toward a lower percentage ofgreen cells in embryos that were injected with YFP and antisense RNA(see Supplementary Table 1 online). Thus, as described in C. elegans2,we found the injection of dsRNA to be much more potent than theinjection of antisense RNA in perturbing gene expression.
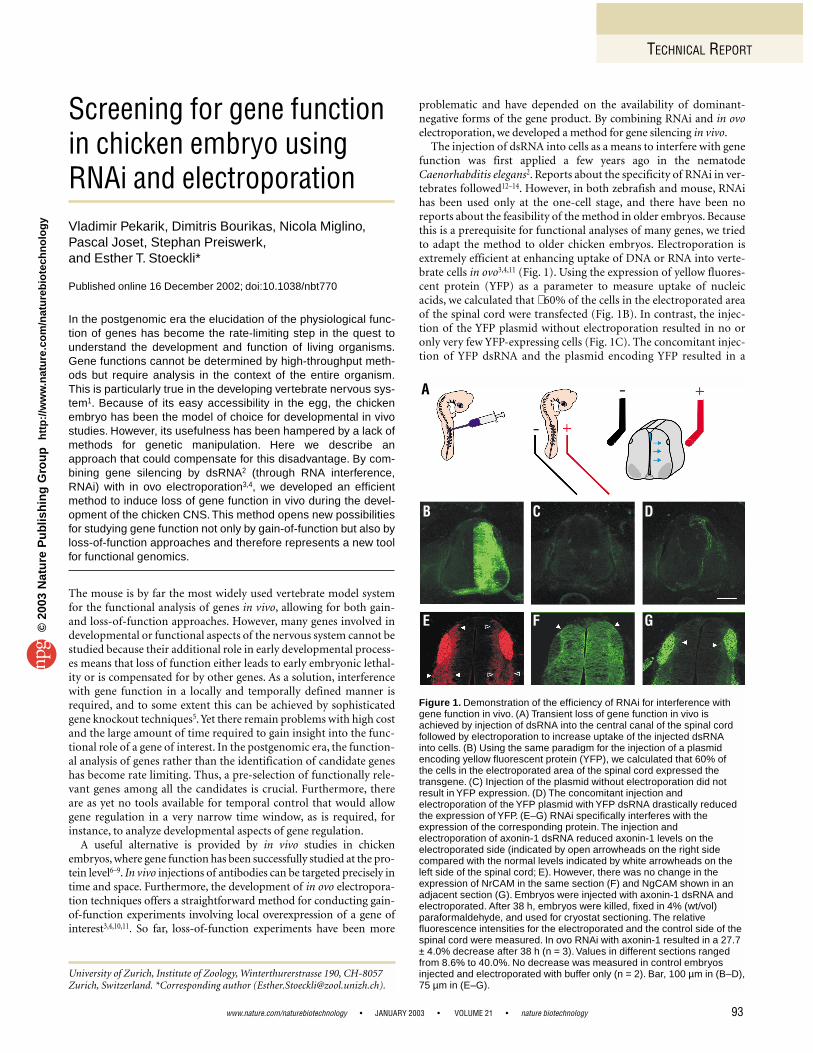
Using several short-range guidance cues and based on our knowl-edge from previous in vivo studies after loss of function at the proteinlevel7,15, we were able to test the specificity and reproducibility of ourin ovo RNAi method by interfering with known short-range guidancecues involved in commissural axon pathfinding in the embryonicchicken spinal cord. In our previous experiments, the injection of
function-blocking antibodies to perturb the interaction of the short-range guidance cues axonin-1 (Fig. 2A) and NrCAM (Fig. 2B), butnot NgCAM (Fig. 2C), induced pathfinding errors of commissuralaxons at the floor plate, the ventral midline of the spinal cord7,8. Herewe used different fragments of the cDNAs encoding these cell adhe-sion molecules to generate dsRNA. The injection of axonin-1 dsRNAinto the central canal of the spinal cord followed by electroporationin ovo resulted in a decrease of axonin-1 expression in the dorsalspinal cord (Fig. 1E) and in pathfinding errors and defasciculation ofcommissural axons (see Fig. 2D). The injection of NrCAM dsRNAresulted in pathfinding errors without effects on fasciculation (Fig. 2E).These phenotypes are identical to the ones observed after interferencewith axonin-1 and NrCAM function at the protein level7 (Fig. 2A, B).As expected from these previous in vivo studies, the interference withNgCAM function using RNAi induced a defasciculated growth pat-tern but did not impair midline crossing by commissural axons(Fig. 2F). The phenotypes were highly reproducible and occurredwith high penetrance (Table 1). The same phenotypes were obtainedfor all dsRNAs derived from three different cDNA fragments of eachgene with no detectable difference in efficiency (see SupplementaryTable 2 online).
Encouraged by these results, we used in ovo RNAi to test prospec-tive candidate guidance cues. For this purpose, we produced dsRNAfrom cDNA fragments that were identified in a subtractive hybridiza-tion screen for guidance cues involved in commissural axon guidance.One example is shown in Figure 3. In the absence of this candidategene’s function, most commissural axons stalled in the floor plate anddid not reach the contralateral border. This phenotype was highlyreproducible (n = 9 out of 12 embryos) and different from phenotypesobserved after perturbation of axonin-1, NrCAM, or NgCAM func-tion. In a BLAST search of the NCBI databank we identified the cDNAfragment as Rab-GDP dissociation inhibitor16 (GDI; accession no.AF076291). Chick Rab-GDI was identified in a library derived fromE18 chick ciliary ganglia16 and found to be upregulated by synapse
Table 1. Efficiency of in ovo RNAi method in producing loss-of-function phenotypes
dsRNA na No effectb <10% 10–20% >20% errors errors errors
Axonin-1 41 0 7.1 28.6 64.3NrCAM 42 6.8 33.0 34.1 26.1NgCAM 23 92.5 7.5 0 0Controlc 23 95.7 4.2 0 0
aFor quantification of the phenotypes after axonin-1, NrCAM, or NgCAM per-turbation by in ovo RNAi, we used only embryos in which we could analyze atleast five injection sites. n, number of embryos analyzed for each condition.Because the effects induced by the different dsRNAs derived from individualcDNAs did not differ, we have pooled the results.bPathfinding errors were classified into four groups: <1% of the fibers turningipsilaterally (no effect), <10%, between 10 and 20%, and >20% of the fibersturning along the ipsilateral floor plate border.cThe control group was injected with PBS containing BSA.
Figure 2. In ovo RNAi produces specific and reproducible phenotypes in the embryonic chicken spinal cord. The phenotypes obtained afterperturbation of short-range guidance cues at the protein level (A–C) and after RNAi (D–F) were identical. Pathfinding errors are indicated byarrows. Changes in the fasciculation of commissural axons are indicatedby arrowheads. The open arrowheads point to the turning point ofcommissural axons at the contralateral floor plate border, where theynormally turn into the longitudinal axis of the spinal cord. To demonstratethe specificity of in ovo RNAi, we chose to perturb the function of the short-range guidance cues axonin-1, NrCAM, and NgCAM incommissural axon pathfinding, because their loss-of-function phenotypeswere known from our earlier in vivo perturbation studies at the proteinlevel7. (A, D) After perturbation of axonin-1 interactions, somecommissural axons failed to cross the floor plate, the ventral midline ofthe spinal cord, and turned into the longitudinal axis along the ipsilateralfloor plate border (arrows). In addition, commissural axons showed adefasciculated growth pattern indicated by a wider trajectory along thefloor plate border compared to control embryos (arrowheads).(B, E) The perturbation of NrCAM interactions had only a small effect on the fasciculation of commissural axons but induced pathfinding errors(arrows) similar to those seen after axonin-1 perturbation. (C, F) Theinterference with NgCAM interactions resulted in a defasciculated growthpattern of commissural axons (arrowheads) but did not cause anypathfinding errors. (G, H) The trajectories of commissural axons in controlembryos. (I) Compared with the control embryo in (H), the defasciculationafter injection of NgCAM dsRNA was even more obvious in transversesections of the spinal cord. (J) Schematic drawing indicating the two types of preparations used for the analysis of commissural trajectories,transverse sections of 250 µm thickness (H, I) on the left and whole-mount (open-book) preparations (A–G) on the right. In both casescommissural axons were visualized with Fast-DiI applied to the cellbodies of commissural neurons. For details see Perrin and Stoeckli19.The floor plate is indicated by dashed lines in (A) through (G). Bar, 50 µm in (A) through (G); 100 µm in (H) and (I).
A B C
D E F
G H I
J
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reb
iote
chn
olo
gy

TECHNICAL REPORT
www.nature.com/naturebiotechnology • JANUARY 2003 • VOLUME 21 • nature biotechnology 95
formation. Consistent with this observation, Rab-GDI was notdetectable in commissural neurons during early phases of axongrowth, but was upregulated at stage 23 when commissural axonshave contacted the floor plate (see Supplementary Fig. 2 online).Rab GTPases are involved in vesicle docking17. The role of Rab-GDIsis the recovery of Rab-GDP from the acceptor membrane and itstransport back to the donor membrane, where Rab-GDP will be reac-tivated by GDP-GTP exchange factors (GEFs) to be ready for a newcycle of vesicle fusion18. A model for Rab-GDI’s function that is con-sistent with our results postulates that vesicle fusion needs to be regu-lated closely when axons contact their intermediate target, the floorplate, representing a choice point along their trajectory15. The interac-tion between the axon and the choice point induces changes in pro-tein expression that allow the axon to navigate the next segment ofthe trajectory toward its final target. Loss of GDI function compro-mises the recovery of Rab-GDP after vesicle fusion and thus interferesindirectly with axonal pathfinding by preventing the display of newguidance cues on the growth cone’s surface.
Our approach did not induce unspecific effects on axon guidanceand did not affect the expression levels of genes other than the one cor-responding to the dsRNA (Fig. 1E–G), making in ovo RNAi a valuabletool for screens in functional genomics. In contrast to our previousin vivo studies that depended on the availability of function-blockingantibodies, in ovo RNAi requires only partial cDNAs for loss-of-function assays. Furthermore, there are apparently no specific require-ments regarding their position or length, because all fragments that weproduced from the full-length cDNAs of axonin-1, NrCAM, andNgCAM resulted in the respective phenotypes without any detectabledifference in efficiency. The lengths of the injected dsRNAs rangedfrom 451 to 2,175 base pairs (see Supplementary Table 2 online).
Thus not only do our results provide evidence for the specificity of inovo RNAi, but they bring the chicken embryo back on stage as aninvaluable model system for developmental studies. In ovo RNAi inchicken embryos is a fast and inexpensive tool that can provide evi-dence for a gene’s function in vivo very quickly and without the needfor full-length cloning of the candidate gene.
Experimental protocolIn ovo RNAi. Research on chicken embryos was carried out according to thelocal guidelines for animal research (Kantonales Veterinäramt, Basel,Switzerland). DsRNA (200–500 ng/µl) dissolved in PBS was injected asdescribed19. Briefly, fertilized eggs were windowed after 3 d of incubation at39 °C. The extraembryonic membranes were partially removed to provideaccess to the embryo. Depending on the age of the embryos, 0.1–0.5 µl PBScontaining either dsRNA or plasmids encoding YFP (under control of theβ-actin promoter) with 0.04% (vol/vol) Trypan Blue (Invitrogen, Carlsbad, CA)was injected into the central canal of the spinal cord at the leg level using glasscapillaries with a tip diameter of ∼ 5 µm. Before and after electroporation afew drops of sterile, cold PBS were added to cool the embryo. Platinum elec-trodes (BTX, Genetronics, San Diego, CA) of 4 mm length with a distance of4 mm between anode and cathode were used. The number of pulses and theapplied voltage must be adapted to the age of the embryos. For 3-d-oldembryos we routinely used five pulses of 50 ms duration at 26 V. After 2 d,embryos were killed and the trajectory of commissural axons was analyzed bythe application of Fast-DiI19 (Molecular Probes, Eugene, OR).
The efficiency of in ovo electroporation was assessed on the basis of expres-sion of YFP. At stage 26, embryos were killed and the percentage of green cellsin the electroporated area was determined in a Neubauer chamber. The per-centage of YFP-expressing cells was 59.1 ± 3.6% (average of 4 embryos ±s.e.m.) and 63.1 ± 2.0% (n = 3 embryos) in two different experiments.Similarly, the percentage of green cells was determined when the YFP-encoding plasmid was co-injected with either sense or antisense RNA (seeSupplementary Table 1 online). Embryos treated the same way were cut witha tissue slicer (250-µm-thick slices) and mounted between coverslips to visu-alize YFP expression (see Supplementary Fig. 1 online).
To show a decrease in axonin-1 expression but no change in NrCAM orNgCAM expression, respectively, after axonin-1 dsRNA injection and elec-troporation, we stained 20-µm-thick cryostat sections9 with rabbit anti-bodies against axonin-1 and goat antibodies against NgCAM or NrCAM(all 1:1,000), followed by donkey antibodies against goat IgG–Alexa 488(1:250; Molecular Probes) and donkey antibodies against rabbit IgG–Cy3(1:250; Jackson ImmunoResearch, West Grove, PA).
Production of dsRNA. DsRNA was produced from different fragments ofcDNAs encoding axonin-1, NrCAM, NgCAM, or candidate guidance cuescloned in pSP72 or pCRII plasmids. After linearization with appropriaterestriction endonucleases (see Supplementary Table 1 online), RNA in vitrosynthesis was carried out from T7 and SP6 promoters. For each direction, 2 µgplasmid DNA was mixed with a final concentration of 4 mM dNTPs (Roche,Basel, Switzerland), 2 µl SP6 (10–20 U/µl; Roche) or T7 polymerase (15 U/µl;Promega, Madison, WI), 0.5 µl RNasin (30 U/µl; Promega) in the appropriatetranscription buffer (total volume was 20 µl).After transcription at 37 °C for 2 h,DNase I (Roche) was added and the RNA was extracted with acidicphenol–chloroform (25:24:1 vol/vol/vol phenol/chloroform/isoamyl alcohol)and then with chloroform/isoamyl alcohol (24:1 vol/vol). After precipitationwith ethanol, the RNA was dissolved in 20 µl PBS. For quality control, 1 µl ofeach sample was analyzed by gel electrophoresis. Equal amounts of sense andantisense RNA were mixed, heated to 95 °C for 5 min, and reannealed by slowcooling over several hours. Annealing and quality of the dsRNA were moni-tored by electrophoresis with nondenaturing conditions and after digestionwith 0.5 µg/ml RNase I.
Note: Supplementary information can be found on the Nature Biotechnologywebsite.
AcknowledgmentsWe thank Monika Mielich for excellent technical assistance. This work was carried out at the University of Basel, Institute of Zoology, and was supportedby the Swiss National Science Foundation, the Ott Foundation, and the Human Frontier Science Program Organization.
Figure 3. A cDNA fragment is sufficient for the analysis of gene functionin vivo by RNAi. In a subtractive hybridization screen we identified severalcandidate guidance cues for commissural axons in the form of cDNAfragments. To pre-select the functionally relevant candidates for furtheranalysis, we tested them with our newly developed in ovo RNAi approach.As an example, we show the analysis of commissural axon trajectories in three different embryos after injection and electroporation of dsRNAderived from one of our candidate genes that showed a strikingphenotype (A–C) compared to a control embryo (D). In contrast tocommissural axons in the control embryo, most commissural axons in theexperimental embryos stalled in the floor plate (arrows) and only very fewfibers managed to reach the contralateral floor plate border (arrowheads).Strikingly, most axons stalled after growing ∼ 60% of the width of the floorplate. The floor plate is indicated by dashed lines; the arrows mark thevirtual line within the floor plate that is reached by most axons. In contrast,in age-matched control-electroporated embryos (D), commissural axonshave left the floor plate and already extended a considerable distancealong the contralateral floor plate border. Bar, 50 µm.
A B
C D
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reb
iote
chn
olo
gy

TECHNICAL REPORT
nature biotechnology • VOLUME 21 • JANUARY 2003 • www.nature.com/naturebiotechnology96
Competing interests statementThe authors declare that they have no competing financial interests.
Received 14 August 2002; accepted 22 October 2002
1. Lo, D.C. Challenges for neuroscience in a post-genome world. Nat. Neurosci. 4,1153–1154 (2001).
2. Fire, A. et al. Potent and specific genetic interference by double-stranded RNA inCaenorhabditis elegans. Nature 391, 806–811 (1998).
3. Muramatsu, T., Mizutani, Y., Ohmori, Y. & Okumura, J. Comparison of three nonvi-ral transfection methods for foreign gene expression in early chicken embryos inovo. Biochem. Biophys. Res. Commun. 230, 376–380 (1997).
4. Itasaki, N., Bel-Vialar, S. & Krumlauf, R. ‘Shocking’ developments in chick embryol-ogy: electroporation and in ovo gene expression. Nat. Cell Biol. 1, E203–E207(1999).
5. Müller, U.Ten years of gene targeting: targeted mouse mutants, from vector designto phenotype analysis. Mech. Dev. 82, 3–21 (1999).
6. Tang, J., Rutishauser, U. & Landmesser, L. Polysialic acid regulates growth conebehavior during sorting of motor axons in the plexus region. Neuron 13, 405–414(1994).
7. Stoeckli, E.T. & Landmesser, L.T. Axonin-1, NrCAM, and NgCAM play differentroles in the in vivo guidance of chick commissural neurons. Neuron 14,1165–1179 (1995).
8. Burstyn-Cohen, T. et al. F-spondin is required for accurate pathfinding of commis-sural axons at the floor plate. Neuron 23, 233–246 (1999).
9. Perrin, F.E., Rathjen, F.G. & Stoeckli, E.T. Distinct subpopulations of sensory affer-ents require F11 or axonin-1 for growth to their target layers within the spinal cordof the chick. Neuron 30, 707–723 (2001).
10. Momose, T. et al. Efficient targeting of gene expression in chick embryos by micro-electroporation. Dev. Growth Differ. 41, 335–344 (1999).
11. Swartz, M., Eberhart, J., Mastick, G.S. & Krull, C.E. Sparking new frontiers: usingin vivo electroporation for genetic manipulations. Dev. Biol. 233, 13–21 (2001).
12. Li, Y.-X., Farrell, M.J., Liu, R., Mohanty, N. & Kirby, M.L. Double-stranded RNAinjection produces null phenotypes in zebrafish. Dev. Biol. 217, 394–405 (2000).
13. Wianny, F. & Zernicka-Goetz, M. Specific interference with gene function by double-stranded RNA in early mouse development. Nat. Cell Biol. 2, 70–75 (2000).
14. Zhao, Z., Cao, Y., Li, M. & Meng, A. Double-stranded RNA injection produces non-specific defects in zebrafish. Dev. Biol. 229, 215–223 (2001).
15. Stoeckli, E.T. & Landmesser, L.T. Axon guidance at choice points. Curr. Opin.Neurobiol. 8, 73–79 (1998).
16. Ikonomov, O.C., Kulesa, M.C., Shisheva, A.C. & Jacob, M.H. Innervation and tar-get tissue interactions induce Rab-GDP dissociation inhibitor (GDI) expressionduring peripheral synapse formation in developing chick ciliary ganglion neuronsin situ. J. Neurosci. 18, 6331–6339 (1998).
17. Stenmark, H. & Olkkonen, V.M. The Rab GTPase family. Genome Biol. 2, Reviews3007.1–3007.7 (2001).
18. Geyer, M. & Wittinghofer, A. GEFs, GAPs, GDIs and effectors: taking a closer (3D)look at the regulation of Ras-related GTP-binding proteins. Curr. Opin. Struct. Biol.7, 786–792 (1997).
19. Perrin, F.E. & Stoeckli E.T. Use of lipophilic dyes in studies of axonal pathfinding invivo. Microsc. Res. Tech. 48, 25–31 (2000).
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reb
iote
chn
olo
gy