Embed Size (px)
Citation preview

Scanning Electron Microscopy of Tilia PollenAuthor(s): T. C. Chambers and H. GodwinSource: New Phytologist, Vol. 70, No. 4 (Jul., 1971), pp. 687-692Published by: Wiley on behalf of the New Phytologist TrustStable URL: http://www.jstor.org/stable/2434743 .
Accessed: 14/06/2014 13:13
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Wiley and New Phytologist Trust are collaborating with JSTOR to digitize, preserve and extend access to NewPhytologist.
http://www.jstor.org
This content downloaded from 195.78.108.60 on Sat, 14 Jun 2014 13:13:39 PMAll use subject to JSTOR Terms and Conditions

New Phytol. (I97I) 70, 687-692.
SCANNING ELECTRON MICROSCOPY OF TILIA POLLEN
BY T. C. CHAMBERS AND H. GODWIN
Botany School, University of Melbourne, and Botany School, Cambridge
(Received 20 January I971)
SUMMARY
A method is described for studying simultaneously the intact and the sectional view of pollen walls. Scanning electron micrographs of such fractured pollen grains of Tilia platyphyllos Scop., T. cordata Mill. and the putative hybrid T. x vulgaris Hayne assist in the interpretation of a number of observations made by light microscopy on the differences between these taxa and confirm and amplify our interpretation of T. platyphyllos made from early transmission electron microscopy of embedded and sectioned grains. The new information makes imperative the distinction between two types of exine reticulation, one bacula-bordered and the other bacula- centric (the Tilia type).
INTRODUCTION
In the decade since we completed our earlier account of the wall structure of Tilia platyphyllos using transmission electron microscopy (Chambers and Godwin, I96I) there have been a number of important advances in techniques yielding new information on the fine structure of pollen. Improved embedding resins, aldehyde fixatives and general refinement of techniques have greatly improved our understanding of the morphogenesis and internal structure of pollen. For the pollen morphologist primarily interested in interpretation and identification of pollen, the most significant advance has been the commercial production of the scanning electron microscope: instruments are now avail- able giving a routine resolution of better than I50 A. Scanning electron microscopy has given us views of intact pollen grains of a quality undreamed of a decade ago but results have hitherto contributed little to our understanding of the arrangement of the sub- surface layers of the wall structure and indeed serious investigators of pollen structure have used the more laborious methods of embedding in hard resins and sectioning for transmission microscopy along with their scanning electron micrographs (e.g. Ferguson and Webb, I970).
A recent paper by Cerceau et al. (I970) elegantly demonstrates the value of disruption of pollen grains by use of ultrasonics prior to scanning electron microscopy. Our attempts with ultrasonics have not been as successful. Further, it must be realized that ultrasonic devices are hazardous to a degree making protection of personnel necessary if pollen were to be broken routinely at the frequency and with the length of exposure that has been recommended. In the present paper a simpler alternative method of fracturing the grains is described which even in its present crude form gives results comparable to those described from the use of ultrasonics.
In our I96I paper we were primarily concerned with the structure of the wall of one species, T. platyphyllos. Light microscopists (e.g. Beug, I970; Andrew, I97I) have
687
This content downloaded from 195.78.108.60 on Sat, 14 Jun 2014 13:13:39 PMAll use subject to JSTOR Terms and Conditions

688 T. C. CHAMBERS AND H. GODWIN
continued and indeed refined their methods of differentiating T. platyphyllos from T. cordata and each of these from the probable hybrid T. x vulgaris (referred to as T. x europea in I96I). It became urgent not only to establish the nature of the structures which account for the optically distinctive behaviour of the two species, but to elucidate a type of exine organization hitherto not properly understood.
METHODS
Pollen samples were taken from herbarium material housed in the Herbarium, Botany School, Cambridge, the identification of which had been verified by an independent taxonomist on characters other than pollen structure. These specimens were selected from the list of those studied by Andrew (I97I, Table i). Mature anthers, some partially dehisced, were selected and for most specimens these were acetolysed by the double method described by Andrew (I970). A distilled water suspensionl of pollen was applied to a slightly sticky aluminium specimen-stub surface prepared by supporting it, well cleaned, in an upright position and applying to it a single small drop of acetone glue extract prepared by placing about 5 cm2 double-sided sticky tape (self-adhesive vinyl tape ioi8; Arabol-Edwardson) in IO ml of acetone. It may be significant that the acetone extracted only some of the adhesive component, the tape remaining sticky after extraction. Immediately on evaporation of the acetone (i.e. about i minute) a drop of the pollen in distilled water was added and after a further minute, when most of the grains had settled to the adhesive surface, the water was carefully blotted simultaneously from opposite edges with pieces of tissue and this left the pollen evenly spread. Stubs so prepared were left overnight to dry.
Samples required for study of wall details in sectional view were then fractured by taking a clean stub and pushing it against the stub with pollen grains attached and, while applying considerable pressure, twisting the clean stub slightly and then lifting it clear. The importance of an extremely thin layer of adhesive becomes obvious at this stage as it is essential to the success of this method that the fragments of grains are not deeply embedded in the glue.
Obviously with specimens fractured in this way useful broken faces with wall details will require rotation and variable tilting in the microscope in order to scan the best areas in an optically ideal way. For this reason the operation is simpler and quicker if the area of fractured grains is exactly in the centre of the specimen stub.
All specimens except one illustrated in Plate 3, No. I4 were coated in a double rotating device at 450 to the source (Edwards High Vacuum Company prototype) with a thin layer (300-400 A) of gold: palladium 6o: 40 and examined at 30 kV on either a Stereoscan IIA or S4 (Cambridge Scientific Instrument Company). The specimen illustrated in Plate 3, No. 14 is carbon coated and photographed at 5 kV.
WALL STRUCTURE
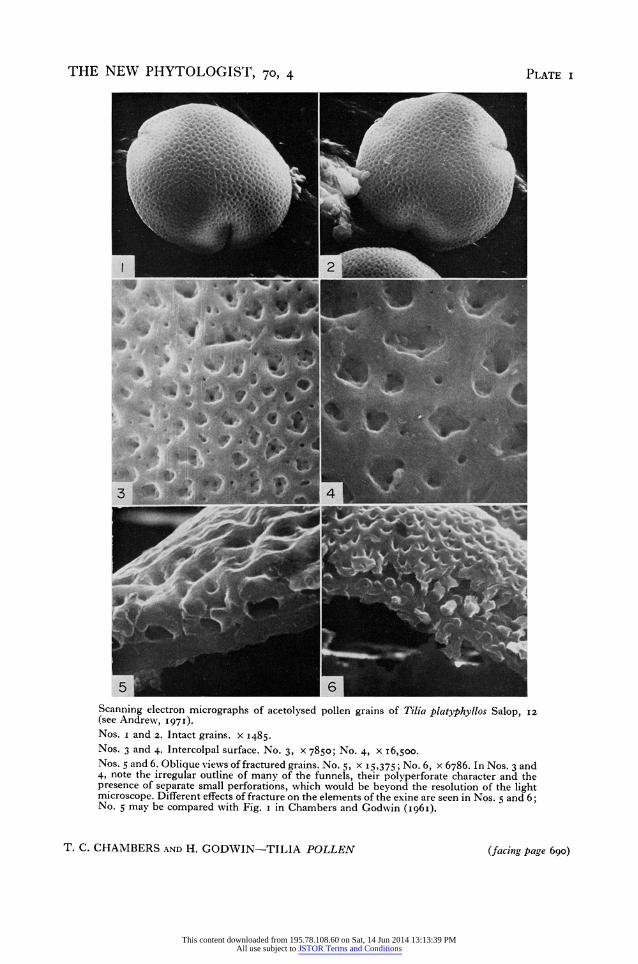
Tilia platyphyllos Scop. (Plate i, Nos. i-6, Plate 3, Nos. II-4) As we reported in i96I from transmission electron microscopy, the exine is a little
over I gm thick, the outer, the inner and the cavity layers each between 0.3 ,um and 0.4 gm in thickness. The outer layer of the exine is made up of the extended walls of numerous funnel shaped hollow columns that separate the surface layer from the basal one. We now see that these are sufficiently irregular in shape (Plate i, Nos. 3 and 4)
This content downloaded from 195.78.108.60 on Sat, 14 Jun 2014 13:13:39 PMAll use subject to JSTOR Terms and Conditions

Tilia pollen 689 (a view which corresponds with recent observations by means of light microscopy) that wewereinerror in illustrating them (Fig. I, p. 395, I96I paper) as circular. In view of their irregular shape it is best to describe their distribution in terms of a-verage centre to centre distances and this further justifies the use of the frequency of funnels (of a size sufficient to be resolved by light microscopy) along a iO gm line on a polar view of a grain as one of the most useful diagnostic characters in differentiating this species from T. cordata (see Andrew, I97I). For typical T. platyphyllos this frequency ranges from eight to twelve but occasionally in extreme examples large irregular funnels are as few as six or seven/Io gm.
While we can confirm our earlier report that perforations in the walls of the funnels open into the voids at their bases (Plate i, No. 5) we now see that they are also to be found at all levels and frequently just below the lip of the funnel (Plate i, Nos. 3 and 4). In addition fine openings penetrate the outer exine between the major funnels. These are the openings to small funnels, some having a central pore and one or more very fine lateral channels similar to those of the larger funnels. These minute funnels are beyond the range of the light microscope but grains where these are numerous may have a slightly distinctive texture when viewed in light optics.
In I96I we described 'smaller columns' which appeared to be solid and we interpreted these as 'possibly ... not complete pillars but ... stalactites or stalagmites'. The Stereoscan pictures indicate that both are present, the stalactites being considerably more frequent, but we cannot say whether they are all hollow. Most of these stalactites we interpret as baculoid funnels (Plate i, Nos. 5 and 6).
A very noticeable feature of the fractured grains is the manner in which these baculoid funnels have often broken out of the inner exine (Plate 3, No. I3) like teeth from their sockets, with however a flat transverse fracture and a clear circular socket (Plate i, No. 5). This is a feature that might well have an explanation in terms of the ontogeny of the bacula (see Angold, I968 for Endymion). In Tilia we frequently see that transverse breakages occur across the stem of the funnels (Plate No. i, 6).
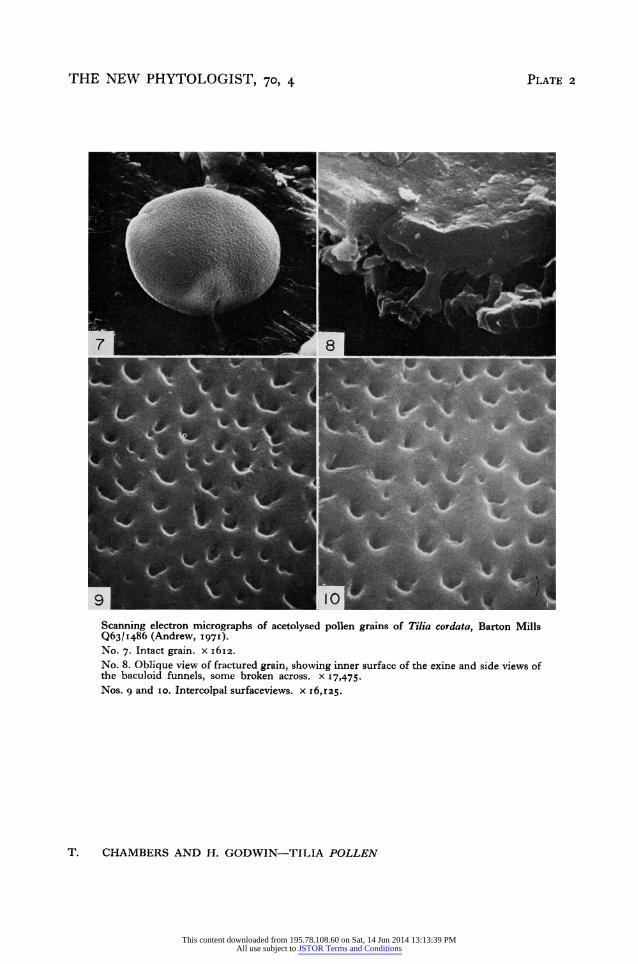
T. cordata Mill. (Plate 2, Nos. 7-IO) The much finer 'mesh' reported by light microscopists and used routinely in the
Quaternary sub-department at Cambridge for diagnosis of the pollen of this species is seen in scanning electron micrographs (Plate 2, Nos. 6-io) to consist of funnels basically similar to but consistently smaller, much closer together and more regular than those of T. platyphyllos. The major funnels of T. cordata of which there are frequently as many as twenty along a io urm line are sufficiently small to be very difficult to study satis- factorily by scanning electron microscopy (Plate 2, Nos. 9 and io). Minute funnels perforating the outer exine between the main funnels also occur in T. cordata.
The cross-sectional structure of the wall of T. cordata (Plate 2, No. 8) is essentially similar to that of T. platyphyllos but the overall dimensions on the examples we have studied here suggest that the wall is a little thicker I.I-I.3 ,um.
Tilia x vulgaris Samples from a number of specimens judged on characters other than pollen struc-
ture as intermediate forms at present best referred to as T. x vulgaris possessed pollen of a variable funnel size and spacing but intermediate between that of T. platyphyllos and T. cordata with occasional grains with a structure (possibly only of one polar face) more closely approaching that of one of the putative parental species. These observations
This content downloaded from 195.78.108.60 on Sat, 14 Jun 2014 13:13:39 PMAll use subject to JSTOR Terms and Conditions

690 T. C. CHAMBERS AND H. GODWIN
confirm but add little to the conclusions of Andrew (I97I) and Beug (1970) and have not been further figured in this report.
DISCUSSION
An important outcome of the I96I paper was that it demonstrated that the pollen grain wall of Tilia platyphyllos was made up on 'an inner and an outer layer of exine joined by a system of hollow funnels and pillars crossing a cavity or void occupied by cyto- plasm'. This interpretation has since been adopted by palynologists employing high resolution optical microscopy in describing the pollen morphology of the genus and the differences between pollen grains of component species (Faegri and Iversen, I964; Beug, I97I). The results of scanning electron microscopy now reported confirm the primary conclusion that the wall morphologically resembles the crypt of a mediaeval church supported by massive columns most of which are conical with wide openings on the outer surface of the grain. Whereas, however, the wall section electron micro- graphs were interpreted as having only a few openings through the funnel walls to the voids, and these low down the columns, the Stereoscan pictures make it apparent that the funnel walls have many lateral openings through them especially evident in the upper, wider part of the funnel cone. The dimensions and frequency of these perforations are immediately evident in Plate I, No. 4 for T. platyphyllos. In the smaller funnels of T. cordata (Plate 2, No. 9 and io) they are smaller, fewer and less apparent but none the less similar.
These results give us the assurance that pollen grain surface reticulation, as observed by light microscopy, may be derived from two basically different kinds of exine anatomy. The commoner, seen a merveille in the Stereoscan pictures of Lilium (Heslop-Harrison, I968), is one in which the walls of the mesh are composed of aligned and more or less confluent bacula: it is what we may call a solid-wall mesh (a bacula-bordered mesh). The less common type, to which Tilia belongs, is quite other. The centre of each mesh is the central cavity of a funnel that may be regarded as a massive hollow baculum or possibly a bundle of fused bacula. The walls of contiguous craters (funnels) slope outwards to meet one another in a system of ridges that, at the highest optical plane, constitute a reticulum or mesh. This reticulation differs from that first described in that there are only cavities below these apparent strands of the net: it is a hollow-ridge mesh or a bacula-centred mesh.
This structural distinction is already almost explicit in Beug's descriptions of Tilia grains, and of course underlies the distinctive luxus-obscurus patterns pointed by Erdtman (1952). As a consequence of recognizing the exine structure of Tilia for what it is, we must abandon the use for bacula-centric reticulate grains, of terms appropriate only to grains of bacula-bordered mesh. We cannot recognize 'murus', 'lumen' or 'brochus' nor can we measure the width of the 'muri'. We can, however, characterize the reticulation by measuring the mean frequency of the perforate funnels, or (which comes to the same thing) their mean distance apart, and we can describe their shape, and, so far as technique allows, the number, shape and size of their perforations.
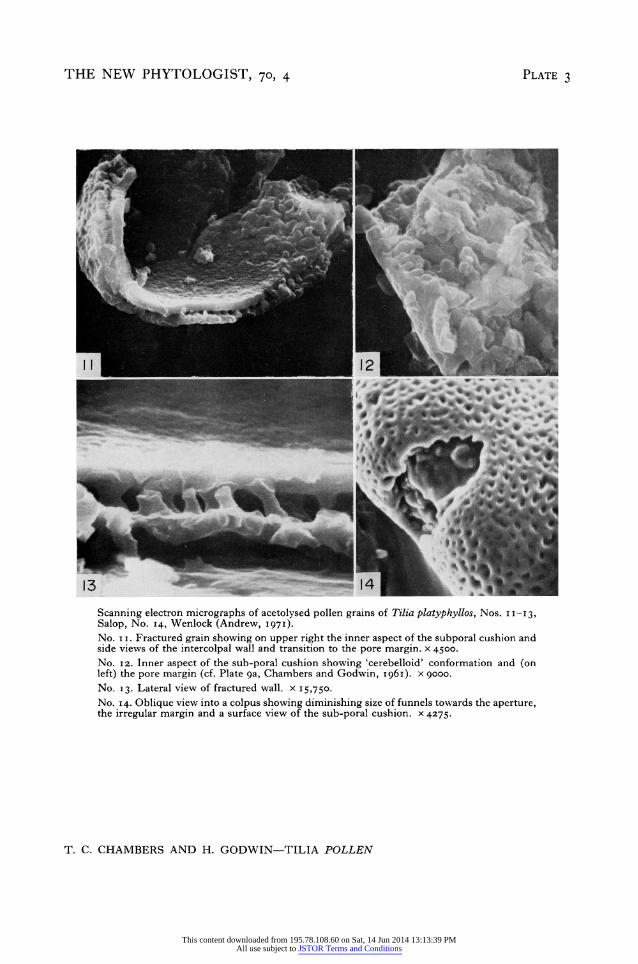
A conspicuous feature of the Tilia pollen grain is the ovoid mass of thickening that lies inside and around each pore. It was shown in electron microscopic section (Chambers and Godwin, I96I) to have a 'cerebelloid' configuration, and sub-poral or circumporal cushions of this kind have been seen subsequently in many types of pollen grain. In Helleborusfoetidus their ontogeny was followed from an early stage (Echlin and Godwin,
This content downloaded from 195.78.108.60 on Sat, 14 Jun 2014 13:13:39 PMAll use subject to JSTOR Terms and Conditions
THE NEW PHYTOLOGIST, 70, 4 PLATE I
2
5 6 _ Scanning electron micrographs of acetolysed pollen grains of Tilia platyphyllos Salop, I2 (see Andrew, I971). Nos. i and 2. Intact grains. x 1485. Nos. 3 and 4. Intercolpal surface. No. 3, x 7850; No. 4, x I6,500. Nos. 5 and 6. Oblique views of fractured grains. No. 5, x 15,375; No. 6, x 6786. In Nos. 3 and 4, note the irregular outline of many of the funnels, their polyperforate character and the presence of separate small perforations, which would be beyond the resolution of the light microscope. Different effects of fracture on the elements of the exine are seen in Nos. 5 and 6; No. 5 may be compared with Fig. i in Chambers and Godwin (I96I).
T. C. CHAMBERS AND H. GODWIN-TILIA POLLEN (facing page 690)
This content downloaded from 195.78.108.60 on Sat, 14 Jun 2014 13:13:39 PMAll use subject to JSTOR Terms and Conditions
THE NEW PHYTOLOGIST, 70, 4 PLATE 2
_' - ~~~~~~~~~~~~~~~~~~~~~~~~~4
"A
9 80
Scanning electron micrographs of acetolysed pollen grains of Tilia cordata, Barton Mills Q63/1|486 (Andrew, '97'). No. 7. Intact grain. x i6Iz. No. 8. Oblique view of fractured grain, showing inner surface of the exine and side views of the baculoid funnels, some broken across. x 17,475. Nos. g and io. Intercolpal surfaceviews. x I6,z25.
T. CHAMBERS AND H. GODWIN-TILIA POLLEN
This content downloaded from 195.78.108.60 on Sat, 14 Jun 2014 13:13:39 PMAll use subject to JSTOR Terms and Conditions
THE NEW PHYTOLOGIST, 70, 4 PLATE 3
jest
04o- N, t
f* *
13 ;9* 14 Scanning electron micrographs of acetolysed pollen grains of Tilia platyphyllos, Nos. II-I3,
Salop, No. I4, Wenlock (Andrew, I 97I). No. I i. Fractured grain showing on upper right the inner aspect of the subporal cushion and side views of the intercolpal wall and transition to the pore margin. x 4500. No. i2. Inner aspect of the sub-poral cushion showing 'cerebelloid' conformation and (on left) the pore margin (cf. Plate ga, Chambers and Godwin, I96I). x gooo. No. I3. Lateral view of fractured wall. x 15,750. No. 14. Oblique view into a colpus showing diminishing size of funnels towards the aperture, the irregular margin and a surface view of the sub-poral cushion. x 4275.
T. C. CHAMBERS AND H. GODWIN-TILIA POLLEN
This content downloaded from 195.78.108.60 on Sat, 14 Jun 2014 13:13:39 PMAll use subject to JSTOR Terms and Conditions

Tilia pollen 69i I969) when they appeared as a secondary exine of sporopollenin laid down upon meander- ing thin 'tapes' or 'lamellae'. With age these curving strands of sporopollenin thicken and condense but between them there persists a matrix of the original cytoplasmic material. It seems probable that the sub-poral cushions of Ti/ia are one expression of a general phenomenon of the stranded or lamellate secondary exine (Godwin, i968). This stranded organization is demonstrated less equivocally by the Stereoscan pictures (Plate 3, Nos. i i and I2) than in reconstructions from transmission electron micrographs. It has probably been emphasized by the removal of any cellulose originally accompanying the sporopollenin by the acetolysis pre-treatment. It is apparent that the elements are irregularly cylindric rather than lamellate.
A third feature clearly displayed by the Stereoscan pictures and otherwise hard to discern is the character of the pore aperture, now seen as an unthickened ovoid opening with an irregular outline determined only by contiguous funnels (Plate 3, Nos. ii and I3), and internally floored by the strands of the sub-poral cushion. There is room for investigation of the inter-relationship of intine and exine in this poral region in relation to the stimulating suggestions of Knox and Heslop-Harrison (1970) of the localization of pollen-wall proteins in poral regions and enzymic activity associated with them.
In the fourth place serial sections in our I96I report have a new significance. Many features overlooked in the original paper are now easily comprehensible. Thus in Plates 7 and ga (I96I) the smaller pores in the upper exine layer, and also near the rims of the funnels, are clearly present in the sections, just as they can be seen in the three dimen- sional Stereoscan image (Plate i, Nos. 3 and 4, Plate 2, Nos. 9 and io). In the same way re-examination of Plate ga (I96I) poses the question whether the voids in the exine wall may extend down between the strands of the sub-poral cushion? If so, the cytoplasm of the interbacula cavities may also continue down through this 'cerebelloid' structure.
ACKNOWLEDGMENTS
We are indebted to Professor P. W. Brian, F.R.S., Botany School, Cambridge for labora- tory facilities and to Miss Robin Andrew and Dr Patrick Echlin for their helpful advice, and to the Cambridge Scientific Instrument Company for their generosity in making available time on a Stereoscan electron microscope. One of us (T.C.C.) is indebted to the President and Fellows of Clare Hall for a Visiting Fellowship, the Royal Society for a travel grant and colleagues in the University of Melbourne who made study leave possible.
REFERENCES
ANDREW, R. (I970). The Cambridge pollen reference collection. Studies in the Vegetational History of the British Isles (Ed. by D. Walker and R. G. West). University Press, Cambridge.
ANDREW, R. (I97i). Exine pattern in the pollen of British species of Tilia. New Phytol., 70, 683. ANGOLD, R. E. (I968). The ontogeny and fine structure of the pollen grain of Endymion nonz-scriptus. Rev.
Palaeobot. Palynol., 3, 205. BEUG, H. J. (I97). Leitfaden der Pollenbestimmuniig far Mitteleuropa und anigrenzende Gebiete. Teile
II/III. Fischer-Verlag, Stuttgart. CERCEAU, M.-T., HIDEUX, M., MARCEAU, L. & ROLAND, F. (I970). Cassure du pollen par les ultrasons pour
l'etude structurale de l'exine au microscope electronique a balayage. C.R. hebd. S4anc. Acad. Sci., Paris, 270, 66.
CHAMBERS, T. C. & GODWIN, H. (I96I). The fine structure of the pollen wall of Tilia platyphyllos. New Phytol., 6o, 393.
ECHLIN, P. & GODWIN, H. (I969). The ultrastructure and ontogeny of pollen in Helleboruitsfoetiduts L. III. The formation of the pollen grain wall. Y. Cell Sci., 5, 459.
ERDTMAN, G. (I952). Pollen Morphology and Plant Taxonomy, Vol. I. Almquist & Wiksell, Stockholm. FAEGRI, K. & IVERSEN, J. (I964). Textbook of Pollen Analysis. Munksgaard, Copenhagen.
This content downloaded from 195.78.108.60 on Sat, 14 Jun 2014 13:13:39 PMAll use subject to JSTOR Terms and Conditions

692 T. C. CHAMBERS AND H. GODWIN FERGUSON, I. K. & WEBB, D. (1970). Pollen morphology in the genus Saxifraga and its taxonomic signifi-
cance. Bot. J7. Linn. Soc., 63, 295. GODWIN, H. (I968). The origin of the exine. Nezv Phytol., 67, 667. HESLOP-HARRISON, J. (I968). Pollen wall development. Science, N. Y., x6x, 230. KNOX, R. B. & HESLOP-HARRISON, J. (1970). Pollen-wall proteins: localization and enzymic activity. Y.
Cell Sci., 6, I.
This content downloaded from 195.78.108.60 on Sat, 14 Jun 2014 13:13:39 PMAll use subject to JSTOR Terms and Conditions





























