Embed Size (px)
Citation preview

17
Molecular and Cellular Biochemistry 203: 17–21, 2000.© 2000 Kluwer Academic Publishers. Printed in the Netherlands.
Sarco/endoplasmic reticulum Ca2+-pump isoformSERCA3a is more resistant to superoxide damagethan SERCA2b
Kelly A. Barnes, Sue E. Samson and Ashok K. GroverDepartment of Medicine, McMaster University, Hamilton, Ontario, Canada
Received 31 March 1999; accepted 8 July 1999
Abstract
Endo/sarcoplasmic reticulum (ER) Ca2+-pumps are important for cell survival and communication but they are inactivated byreactive oxygen species (ROS). We have previously reported that the Ca2+-pump isoform SERCA3a is more resistant than SERCA2bto damage by peroxide. Since peroxide and superoxide differ in their redox potentials, we now report the effects of superoxide onthe two Ca2+-pump isoforms. We isolated microsomes from HEK293 cells transiently transfected with SERCA2b or SERCA3acDNA. We exposed these microsomes to superoxide which was generated using xanthine plus xanthine oxidase and catalase toprevent accumulation of peroxide due to superoxide dismutation. Superoxide damaged the Ca2+- transport activity of both isoformsbut SERCA3a was damaged at higher concentrations of superoxide and upon longer periods of exposures than was SERCA2b.Thus the SERCA3a isoform is more resistant than SERCA2b to inactivation by both superoxide and peroxide. (Mol CellBiochem 203: 17–21, 2000)
Key words: Free radicals, Ca-Mg-ATPase, ischaemia, coronary artery, immune-response
Abbreviations: ER – endoplasmic reticulum; ROS – reactive oxygen species; XO – xanthine oxidase
Introduction
During cell activation, Ca2+-sequestered in the endoplasmicreticulum (ER) is released into the cytosol through IP
3-
dependent Ca2+-channels or Ca2+-induced Ca2+-releasechannels [8, 16, 21]. ER Ca2+-pumps remove cytosolic Ca2+ intothe lumen of ER. Thus, the ER Ca2+-pumps have a dualfunction: initiating cell deactivation by removing cytosolicCa2+ and sequestering this Ca2+ into the ER for releaseduring the next signal transduction cycle. Ca2+-entry intothe cell in response to Ca2+ release from the ER alsocontributes to both cell activation and refillingthe ER withCa2+. Three genes encode theER Ca2+ pumps: SERCA1, 2 and3 [4, 9, 15, 22]. All the SERCA proteins have a very highdegree of identity with each other. The fast twitch skeletalmuscle expresses SERCA1 and cardiac or slow twitch skeletal
muscles express a splice of SERCA2 (SERCA2a). Mosttissues, including vascular smooth muscle,express anothersplice of SERCA2 (SERCA2b). Reactive oxygen species(ROS, or oxygen free radicals) which are formed duringcardiac ischaemia-reperfusion or during immune responsefollowing infection can inactivate SERCA2 Ca2+-pumps [3,6, 18–20]. Tracheal epithelium, mast cells,lymphoid cells,platelets and aortic endothelium express a different Ca2+-pump gene, SERCA3 [1, 2 ,4 ,5, 17, 22, 23]. These tissues arealso frequently exposed to oxidative stress. For instance, theendothelium in the arteries is exposed to oxidative stressmore often because ROS are generated in the arterial lumenby macrophages or by catalytic actions of Fe2+ liberated fromdecomposition of haemoglobin. We surmise that an expressionof an ROS resistant Ca2+-pump protein would enable thesetissues to function during infection and in mild cases of
Address for offprints: A.K. Grover, Department of Medicine, HSC 4N41, McMaster University, 1200 Main Street West, Hamilton, Ontario L8N3Z5 Canada

18
ischaemia-reperfusion. Indeed, SERCA3a is more resistantto peroxide damage than SERCA2 and in coronary arterythe SERCA-pump in endothelium is more resistant thanin smooth muscle to inactivation by peroxide [12, 14]. Bothsuperoxide and peroxide can damage SERCA2b [7, 10, 11,13]. However,the two ROS differ in their redox potentialsuch that superoxide can act as a reducing or as anoxidizing agent while peroxide acts only as an oxidant.Therefore, we now compare the effects of superoxide onSERCA2b and SERCA3a. We chose these isoforms becausethese are the most widely expressed splices of these twogenes [5, 17].
Materials and methods
Cell transfection and microsome preparation
SERCA2b and SERCA3a full length proteins were over-expressed in the cell line HEK293 using the plasmids PMT2-SERCA2b and PMT2-SERCA3a as described previously [14].Briefly, HEK293 cells were cultured to confluence in DMEMsupplemented with 10% fetal calf serum, 2 mM glutamine,50 µg/ml gentamicin and 0.5mM HEPES and split 1:2 24 h later.The cells were transfected by the calcium phosphate pre-cipitation method and 48 h later microsomes were prepared fromthem as described earlier [14]. The resulting microsomes weresuspended (protein concentration of 0.3–0.7 mg/ml) in 8%sucrose and 5 mM Na-azide and preincubated with superoxide.Superoxide was generated using xanthine plus xanthine oxidase(XO) as described earlier. Typically, microsomes were pre-incubated at 37°C in 10 mM imidazole pH 6.8 buffer containing0.3 mM xanthine, specified concentrations of XO and excess(120 units/ml) of catalase. XO was purified from bovine milkand assayed following the conversion of xanthine to uric acid.An enzyme unit is defined as the amount of enzyme whichconverts one µmole of xanthine to uric acid in one min.
Ca2+-transport
The microsomes, which had been preincubated in differentconcentrations of superoxide for 30 min at 37°C, were diluted6-fold for 45Ca2+-uptake in a medium containing: 30 mMimidazole-HCl pH 6.8 (pH at 37°C), 100 mM KCl, 5 mM MgCl
2,
5 mM Na-azide, 1mM EGTA, 0.85 mM CaCl2 (plus trace
amounts of 45CaCl2), 5 mM ATP, 5 mM oxalate and 15–30 µg
of microsomal protein [14]. This resulted in 5 µM [Ca2+] in theassay mixture. In an initial experiment it was determined thatthe uptake was linear for up to 5 min. Routinely, the sampleswere incubated for 2 min at 37°C and then filtered through0.45 µm nitrocellulose filters under suction.
Data analysis
Curve fitting was carried out using the software package Fig P.Null hypotheses were tested using a Student’s t-test and the pvalues < 0.05 were considered to be statistically significant.
Results
Time course of Ca2+-uptake
Transfecting cells with SERCA-pump cDNAs produces anincrease in the amount of the corresponding proteins andhence it would affect the initial rate of the 45Ca2+-uptake bythe microsomes but it may not increase the amount of endo-plasmic reticulum and hence the final uptake may not change.Figure 1 shows the time-course of the 45Ca2+-uptake bymicrosomes from HEK293 cells overexpressing SERCA2b andSERCA3a proteins and the untransfected HEK293 cells. TheATP-dependent uptake was examined in the presence of Na+-azide to inhibitany uptake by mitochondria. The 45Ca2+-uptakeinto the microsomes from cells with SERCA2b or SERCA3awas linear for up to 5 min. The uptake in 5 min was 6–20fold higher in the microsomes from the transfected cells thanin those from the untransfected cells. In order to examine theeffect of superoxide on the initial velocity of the pump wedetermined the uptake in 2 min in all the subsequentexperiments.
Fig. 1. Time course of 45Ca2+-uptake. Microsomes prepared fromtransfected and control cells were incubated with the 45Ca2+-uptakesolution for the specified lengths of time and then filtered as describedin the Materials and methods. Each data point represents mean ±S.E.M. of 6 replicates. This experiment was conducted three times toconfirm the observation that the uptake was linear for up to 5 min.
19
Effect of superoxide on Ca2+-uptake
Superoxide was generated by using xanthine + XO and anyperoxide formed due to dismutation of superoxide wasdegraded using excess catalase. Figure 2 shows the timecourse of the superoxide-dependent and superoxide-in-dependent inactivation of the 45Ca2+-uptake by microsomesprepared from the SERCA2b and SERCA3a transfected cells.The formation of superoxide monitored as disppearance ofxanthine or as reduction of cytochrome c was linear with timeduring this period (data not shown). Preincubating themicrosomes at 37°C by itself resulted in a time dependent lossof the 45Ca2+-uptake activity (Figs. 2A and 2B). The spon-taneous loss of activity was greater for SERCA2 than forSERCA3a. The loss of activity due to superoxide was ex-pressed compared to the loss of activity without the super-oxide in the same time period. Inactivation of 45Ca2+-uptakeby superoxide (XO) was less for SERCA3a than for SERCA2at 0, 30, 60 or 90 min.
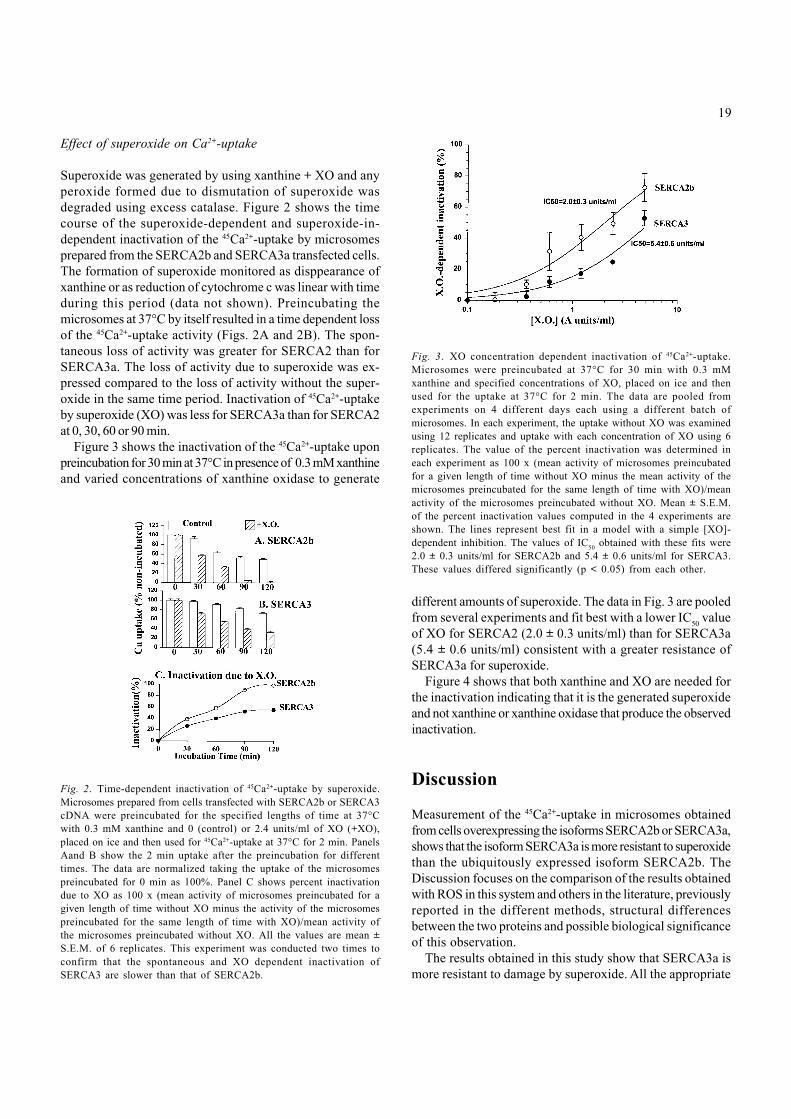
Figure 3 shows the inactivation of the 45Ca2+-uptake uponpreincubation for 30 min at 37°C in presence of 0.3 mM xanthineand varied concentrations of xanthine oxidase to generate
different amounts of superoxide. The data in Fig. 3 are pooledfrom several experiments and fit best with a lower IC
50 value
of XO for SERCA2 (2.0 ± 0.3 units/ml) than for SERCA3a(5.4 ± 0.6 units/ml) consistent with a greater resistance ofSERCA3a for superoxide.
Figure 4 shows that both xanthine and XO are needed forthe inactivation indicating that it is the generated superoxideand not xanthine or xanthine oxidase that produce the observedinactivation.
Discussion
Measurement of the 45Ca2+-uptake in microsomes obtainedfrom cells overexpressing the isoforms SERCA2b or SERCA3a,shows that the isoform SERCA3a is more resistant to superoxidethan the ubiquitously expressed isoform SERCA2b. TheDiscussion focuses on the comparison of the results obtainedwith ROS in this system and others in the literature, previouslyreported in the different methods, structural differencesbetween the two proteins and possible biological significanceof this observation.
The results obtained in this study show that SERCA3a ismore resistant to damage by superoxide. All the appropriate
Fig. 2. Time-dependent inactivation of 45Ca2+-uptake by superoxide.Microsomes prepared from cells transfected with SERCA2b or SERCA3cDNA were preincubated for the specified lengths of time at 37°Cwith 0.3 mM xanthine and 0 (control) or 2.4 units/ml of XO (+XO),placed on ice and then used for 45Ca2+-uptake at 37°C for 2 min. PanelsAand B show the 2 min uptake after the preincubation for differenttimes. The data are normalized taking the uptake of the microsomespreincubated for 0 min as 100%. Panel C shows percent inactivationdue to XO as 100 x (mean activity of microsomes preincubated for agiven length of time without XO minus the activity of the microsomespreincubated for the same length of time with XO)/mean activity ofthe microsomes preincubated without XO. All the values are mean ±S.E.M. of 6 replicates. This experiment was conducted two times toconfirm that the spontaneous and XO dependent inactivation ofSERCA3 are slower than that of SERCA2b.
Fig. 3. XO concentration dependent inactivation of 45Ca2+-uptake.Microsomes were preincubated at 37°C for 30 min with 0.3 mMxanthine and specified concentrations of XO, placed on ice and thenused for the uptake at 37°C for 2 min. The data are pooled fromexperiments on 4 different days each using a different batch ofmicrosomes. In each experiment, the uptake without XO was examinedusing 12 replicates and uptake with each concentration of XO using 6replicates. The value of the percent inactivation was determined ineach experiment as 100 x (mean activity of microsomes preincubatedfor a given length of time without XO minus the mean activity of themicrosomes preincubated for the same length of time with XO)/meanactivity of the microsomes preincubated without XO. Mean ± S.E.M.of the percent inactivation values computed in the 4 experiments areshown. The lines represent best fit in a model with a simple [XO]-dependent inhibition. The values of IC50 obtained with these fits were2.0 ± 0.3 units/ml for SERCA2b and 5.4 ± 0.6 units/ml for SERCA3.These values differed significantly (p < 0.05) from each other.

20
controls indicate that the observed damage is due to superoxideand not xanthine or xanthine oxidase because: (a) eitheragent alone is ineffective and both the agents are needed toproduce the inactivation, and (b) the damage depends notonly upon the concentration of XO used but also with thetime for which the microsomes are exposed to xanthine + XO,thereby correlating it with the exposure to the amount ofsuperoxide generated. The observed damage is due to super-oxide and not due to peroxide generated upon dismutation ofsuperoxide since a very high concentration of catalase wasincluded in the superoxide generating system to instant-aneously decompose the peroxide. The IC
50 value for XO for
SERCA2b in these experiments is similar to those usingpermeabilized coronary artery smooth muscle cells reportedpreviously but higher than those we had obtained usingmicrosomes from the artery [7, 8, 11]. However, catalase wasnot included in the previous study using the microsomes andthis may be the source of the discrepancy. One can not ruleout the possibility that part of the damage observed here isdue to superoxide increasing permeability of the microsomes.However, the cell type used for SERCA2b and SERCA3a wasthe same and hence any damage produced by superoxidewould be similar. Therefore, we conclude that the SERCA3apump is more resistant to superoxide than SERCA2b. Thisresult is in accordance with the previous observation that theSERCA3a pump is more resistant to peroxide than SERCA2b.Based on differences in cysteines between SERCA2 and 3and the observation that SERCA2 sensitivity to oxidativedamage increases with pH, we previously proposed that thecysteine 887 in SERCA2b is more likely to be the cause of its
greater susceptibility to peroxide [14]. Whether superoxideis involved in the same oxidative pathway as peroxide oracts via a different chain of reactions remains to be determined.
We previously hypothesized that SERCA3a is part of acoping mechanism against oxidative stress in the tissuesexpressing it [14]. This hypothesis was based the resistanceto peroxide being greater for SERCA3a than for SERCA2b.The greater resistance of SERCA3a to superoxide obtainedin this study further supports this hypothesis.
Acknowledgements
We thank Drs. G. Shull (University of Cincinnati, USA), S.Asano (Toyama Medical & Pharmaceutical University, Japan)and J. Lytton (Calgary University, Canada) for the gift of theplasmids pMT2-SERCA2b and pMT2-SERCA3a. This workwas carried out in part as an undergraduate thesis project ofKB. The work was supported by Heart & Stroke Foundationof Ontario as a Grant-in-Aid and a Career Award to AKG.
References
1 . Anger M, Samuel JL, Marotte F, Wuytack F, Rappaport L, LompreAM: The sarco(endo)plasmic reticulum Ca2+-ATPase messengerRNA isoform, SERCA-3, is expressed in endothelial and epithelialcells in various organs. FEBS Lett 334: 45–48, 1993
2 . Anger M, Samuel JL, Marotte F, Wuytack F, Rappaport L,Lompre AM: In situ mRNA distribution of sarco(endo)plasmicreticulum Ca2+- ATPase isoforms during ontogeny in the rat. JMol Cell Cardiol 26: 539–550, 1994
3 . Araki N, Greenberg JH, Uematsu D, Sladky JT, Reivich M: Effectof superoxide dismutase on intracellular calcium in stroke. J CerebBlood Flow Metab 12: 43–52, 1992
4 . Burk SE, Lytton J, MacLennan DH, Shull GE: cDNA cloning,functional expression, and mRNA tissue distribution of a thirdorganellar Ca2+ pump. J Bio Chem 264: 18561–18568, 1989
5 . Dode L, Wuytack F, Kools PF, Baba-Aissa F, Raeymaekers L, Brike F,van d, V, Casteels R, Brik F: cDNA cloning, expression andchromosomal localization of the human sarco/endoplasmic reticulumCa(2+)-ATPase 3 gene [published erratum appears in Biochem J 1996,Nov 1; 319 (Pt 3): 1008]. Biochem J 318 (Pt 2): 689–699, 1996
6 . Downey JM: Free radicals and their involvement during long-term myocardial ischemia and reperfusion. Annu Rev Physiol52: (review) 487–504, 1990
7 . Elmoselhi A, Samson S, Grover AK: Heterogeneity in the effectsof reactive oxygen on coronary artery Ca2+ pumps loading IP3sensitive and insensitive SR Ca2+ pools. Am J Physiol: 1996
8. Gill DL, Ghosh TK, Bian J, Short AD, Waldron RT, Rybak SL:Function and organization of the inositol 1,4,5-trisphosphate-sensitive calcium pool. Adv Second Mess Phosphopro Res. 26:(review) 265–308, 1992
9 . Grover AK, Khan I: Calcium pump isoforms: Diversity, selectivityand plasticity. Cell Calcium13: 9–17,(review) 1992
10. Grover A, Samson SE: Impurities in commercial xanthine oxidaseinhibit Ca pump and interfere in contractility of pig coronaryartery. Biochem Biophys Res Commun 143: 575–581, 1987
Fig. 4. Control experiments to show that the inactivation of 45Ca2+-uptake is due to superoxide. Microsomes were preincubated at 37°Cfor 60 min with 0 or 0.3 mM xanthine and 0 or 2.4 units/ml of XO,placed on ice and then used for the uptake at at 37°C for 2 min. Thevalues shown are percent inactivation values computed as 100 x (meanactivity of microsomes preincubated for a given length of time withoutxanthine and XO minus the activity of the microsomes preincubatedfor the same length of time with xanthine or XO)/mean activity ofthe microsomes preincubated without xanthine and XO. The valuesare mean ± S.E.M. of 6 replicates. A significant (p < 0.05) inactivationwas observed only when both xanthine and XO were present in thepreincubation mixture. The inactivation observed with XO + xanthinewas significantly lower (p < 0.05) for SERCA3 than for SERCA2b.The experiment was conducted three times to confirm this conclusion.

21
11. Grover AK, Samson SE: Effect of superoxide radical on Ca2+
pumps of coronary artery. Am J Physiol 255: C297–C303, 198812. Grover AK, Samson SE: Sarco/endoplasmic reticulum Ca2+ pump
is more resistant to peroxide in endothelium than in smoothmuscle: Implications to coronary artery function. Am J Physiol273: 1250–1258, 1997
13. Grover AK, Samson SE, Fomin VP: Peroxide inactivates calciumpumps in pig coronary artery. Am J Physiol 263: H537–H543,1992
14. Grover AK, Samson SE, Misquitta CM: Sarco/endoplasmicreticulum Ca pump isoform SERCA3 is more resistant to peroxidethan SERCA2b. Am J Physiol 273: C420–C425, 1997
15. Lytton J, Westlin M, Burk SE, Shull GE, MacLennan DH:Functional comparisons between isoforms of the sarcoplasmic orendoplasmic reticulum family of calcium pumps. J Biol Chem267: 14483–14489, 1992
16. Missiaen L, Wuytack F, Raeymaekers L, de Smedt H, Droogmans G,Casteels R: Ca2+ extrusion across plasma membrane and Ca2+ uptakeby intracellular stores. Pharmacol Ther 50: 191–232,(review) 1991
17. Poch E, Leach S, Snape S, Cacic T, MacLennan DH, Lytton J:Functional characterization of alternatively spliced humanSERCA3 transcripts. Am J Physiol 275: C1449–C1458, 1998
18. Singal PK, Dhalla AK, Hill M, Thomas TP: Endogenousantioxidant changes in the myocardium in response to acuteand chronic stress conditions. Mol Cell Biochem 129: 179–186, 1993
19. Suzuki YJ, Ford GD: Inhibition of Ca(2+)-ATPase of vascularsmooth muscle sarcoplasmic reticulum by reactive oxygenintermediates. Am J Physiol 261: H568–H574, 1991
20. Todoki K, Okabe E, Kiyose T, Sekishita T, Ito H: Oxygen freeradical-mediated selective endothelial dysfunction in isolatedcoronary artery. Am J Physiol 262: H806–H812, 1992
21. Vanbreemen C, Chen Q, Laher I: Superficial buffer barrier functionof smooth muscle sarcoplasmic reticulum. Trends Pharmacol Sci16: 98–105, 1995
22 . Wu KD, Lee WS, Wey J, Bungard D, Lytton J: Localizationand quantification of endoplasmic reticulum Ca2+-ATPaseisoform transcripts. Am J Physiol – Cell Physiol 38: C775–C784, 1995
23. Wuytack F, Papp B, Verboomen H, Raeymaekers L, Dode L,Bobe R, Enouf J, Bokkala S, Authi KS, Casteels R: A Sarco/endoplasmic reticulum Ca2+-ATPase 3-Type Ca2+ pump is expressedin platelets, in lymphoid cells, and in mast cells. J Biol Chem 269:1410–1416, 1994





























