Embed Size (px)
Citation preview

BACTERIOLOGICAL REVIEWS, June 1974. p. 164-198 Vol. 38, No. 2Copyright © 1974 American Society for Microbiology Printed in U.S.A.
Saccharomyces cerevisiae Cell CycleLELAND H. HARTWELL
Department of Genetics, University of Washington, Seattle, Washington 98195
INTRODUCTION ......................................... 164SYNCHRONOUS CULTURES .................. ....................... 165Assay of Landmarks ......... ................................ 165Induction Synchrony .......... ............................... 166Selection Synchrony.167
CELL DIVISION CYCLE MUTANTS ......................................... 167Diagnostic Landmarks and Terminal Phenotypes ............... ................ 167First Cycle Arrest and Execution Points.168The Mutant Collection .......... ............................... 171Role of cdc Genes in the Life Cycle.171
COMPONENT PROCESSES OF THE CELL CYCLE .............. ................ 171Bud Emergence ......................................... 171Bud Growth ......................................... 173Cytokinesis and Cell Separation.......................................... 173Nuclear Structure ............................ 174Spindle Plaque Cycle ............................ 176Nuclear DNA Components ............................ 176Initiation of Nuclear DNA Synthesis ........ .................... 178Nuclear DNA Synthesis ............................ 179Mitochondrial DNA ............................ 180Two-Micron Circular DNA ............................ 181Macromolecule Synthesis.182
HOW ARE THE EVENTS OF THE CELL CYCLE COORDINATED? ...... ........ 183Dependent Pathways of Landmarks.............................................. 183Dependent Pathways of Gene Expression.184
HOW IS CELL DIVISION CONTROLLED? ...................................... 186Transition from Mitosis to Conjugation ......................................... 186Transition from Mitosis to Stationary Phase .................................... 188Transition from Mitosis to Meiosis ........................................... 188
HOW IS GROWTH INTEGRATED WITH DIVISION? ........... ................. 190THE CONCEPT OF "START"............................................ 191
INTRODUCTION
The yeast Saccharomyces cerevisiae is a sin-gle-celled organism capable of rapid division ondefined medium. Each cell reproduces by bud-ding and the bud grows in size throughout thecell cycle, thereby providing a morphologicalindicator of cycle progress. The organism pos-sesses an elegant and extensively exploitedgenetic system. Cells of ploidy from haploid tooctaploid undergo the same mitotic cycle sothat recessive mutations affecting cycle prog-ress can be detected in haploids and analyzedby complementation in diploids, and the effectsof gene dosage can be examined in polyploids.Furthermore, the basic elements of cell struc-ture, macromolecule synthesis, chromosomereplication, and chromosome segregation in S.cerevisiae are extensively homologous to thosein higher plant and animal cells. These factstaken together suggest that S. cerevisiae may bethe most tractable of experimental organisms incurrent use for a combined genetic, physiologi-
cal, and biochemical analysis of the mitotic celldivision cycle in eukaryotes. This review consti-tutes an attempt to relate the potential of thisorganism for studies on cell division and toreview our current meager understanding of thisprocess in S. cerevisiae. Readers seeking a lessparochial treatment of the cell cycle are referredto an excellent monograph that synthesizesobservations from many organisms includingprokaryotes and eukaryotes (114).
In considering the cell cycle as a whole one isimpressed with the diversity of biological proc-esses that must be coordinated to assure theultimate goal of cellular reproduction. Mitchi-son appropriately stated "My thesis ... is thatwe can find the same processes of morphogene-sis and periodic gene expression in the minia-ture system of the cell cycle and that differen-tiation can be found and investigated as prop-erly here as in the whole developing organism"(115). I find it useful to consider the problems ofmorphogenesis and differentiation in the cellcycle in terms of three questions which are
64
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
admittedly neither comprehensive nor mutuallyexclusive:How are the events of the cell cycle coordi-
nated? How does the cell assure that any twodiscontinuous cell cycle events such as deox-yribonucleic acid (DNA) synthesis and nucleardivision always occur in the proper order?How is cell division controlled? How does the
cell arrest the mitotic cell division cycle andcommence upon an alternative developmentalprogram such as sexual conjugation, meiosis, orstationary phase?How is growth integrated with division? How
does the cell accomplish the precise doubling ofall of its macromolecular components betweentwo consecutive cell cycles under conditions ofbalanced growth?The material selected for the present review
was chosen in large part because of its relevanceto one or more of these questions. Although itwill not be possible at the present time toprovide satisfying answers to any of thesequestions, it will be possible during the course ofthis review to rephrase them. Whether thataccomplishment represents an exercise in se-mantics or a measure of progress, the readermust decide.The discontinuous events that occur once
during each cell cycle constitute landmarks ofcell cycle progress by which we can assess acell's position. The landmarks of the S. cerevi-siae cell cycle will be discussed in greater detailbelow, but it may be useful to consider theirtemporal order at this time (Fig. 1). For reasonsthat will become apparent later it is appropriateto consider the cycle as commencing with anunbudded cell in the G1 interval of the cycle.The nucleus contains a single-spindle plaque, astructure embedded in the nuclear membranefrom which microtubules arise. Three events,whose precise temporal order has not beendetermined, mark the end of the G1 interval:spindle plaque duplication, the initiation ofDNA synthesis, and the emergence of the bud.The DNA synthetic interval or S period consti-tutes about 25% of the cycle. The spindleplaques separate to form the complete spindle,and the bud grows in size throughout theremainder of the cycle. The end of G2 is markedby the migration of the nucleus to the neck ofthe cell where it undergoes the first stage ofnuclear division, medial nuclear division, con-comitant with the elongation of the spindlemicrotubules. The second stage of nuclear divi-sion, late nuclear division, is followed by cyto-kinesis, or cell membrane separation, which isfollowed in turn by cell wall separation. Thisevent completes the cycle with the productionof two unbudded cells. A diploid cell growing atthe optimal temperature, 30 C, may complete a
INI
C Cs
GI3 iDS (
D ( M DS
\ G-ny/
PDBE
PS
mND
NM
FIG. 1. Landmarks of the S. cerevisiae cell divisioncycle. Abbreviations: PD, plaque duplication; BE,bud emergence; iDS, initiation of DNA synthesis;DS, DNA synthesis; PS, plaque separation; NM,nuclear migration; mND, medial nuclear division;SE, spindle elongation; IND, late nuclear division;CK, cytokinesis: CS, cell separation. Distance be-tween events does not necessarily reflect interval oftime between events.
cycle in about 100 min. Although the literatureon the S. cerevisiae cell cycle will be extensivelyreferenced below, it is appropriate to acknowl-edge at this point that much of our currentunderstanding of the yeast cell cycle is theresult of pioneering research carried out forseveral years in the laboratories of H. 0. Hal-vorson, C. F. Robinow, and D. H. Williamson.
SYNCHRONOUS CULTURES
Assay of LandmarksThe experimental analysis of the cell cycle
frequently requires synchronous cultures and anumber of methods employing either inductionor selection (114) have been devised to achievepartially synchronous division of S. cerevisiae.In such experiments, it is necessary to follow atleast one landmark as a measure of the degree ofsynchrony, and more than one is preferable. Abrief discussion of the synchrony techniquesavailable for yeast can be usefully integratedwith another methodological consideration, howeach of the landmarks are assayed.The most convenient landmark is bud emer-
gence because it can be monitored by directvisual examination and there is essentially nosubjective element involved in distinguishingan unbudded from a budded cell.DNA synthesis has also been used as a
measure of synchrony. However, because the Speriod occupies a significant but not easilydetermined portion of the cycle, this measure-ment is inherently less accurate than that of
165VOL. 38, 1974
T
0
1
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
bud emergence. DNA synthesis cannot be moni-tored in yeast by thymine or thymidine incorpo-ration, possibly because the organism lacksthymidine kinase (59); however, it can be mea-sured by incorporation of radioactive adenine(18, 187) or uracil (69) into alkali-resistant,acid-precipitable material. Recently mutantscapable of incorporating thymidine 5'-mono-phosphate have been described (16, 89, 90, 182),and these may greatly facilitate DNA studies inyeast.
Nuclear division has been used to followsynchrony and is in principle a good marker,because during the brief interval between nu-clear migration and medial nuclear division thenucleus is found in the neck of the cell and thisstage can be used as a "mitotic index." Visual-izing the position of the nucleus requires fixa-tion of the cells and staining, usually withGiemsa (145).
Cytokinesis can be monitored by fixing cellswith formaldehyde followed by the digestion ofthe wall with snail digestive juice (71). If theparent cell and bud come apart following thistreatment, they are presumed to have com-pleted cell membrane division or cytokinesis.The event measured empirically in this mannerfollows nuclear division and precedes cell sepa-ration, but there is no direct evidence that itcoincides with cytoplasmic membrane division.Most frequently cell division, measured ei-
ther by particle count or by visual examination,has been used to monitor synchrony. When aparticle-counting device is employed, it is usu-ally necessary to sonically treat samples prior tocounting in order to facilitate separation of cellsthat have completed physiological division, butthat may otherwise remain adhered for severalcell cycles (147). As an alternative to sonication,cells have been scored by visual examination ina hemocytometer by using the following conven-tion. A particle with a single bud is scored asone cell and a particle containing two or threebuds is scored as two cells (192). This conven-tion is based upon the observation that underconditions of balanced growth, cell separationin one cycle precedes bud emergence in the nextcycle (192). Of course, this method does notscore the cell division event itself, but ratherthe immediately succeeding bud emergenceevent. Finally, it should be pointed out thatsome authors have employed a different con-vention by counting a unit with a bud as onecell until the bud reaches a certain size, where-upon the unit is counted as two cells. Thislatter procedure is not recommended becauseit is both arbitrary and subjective.
Other parameters of cell cycle progress and
cell synchrony include particle volume distribu-tions (64, 150, 168), the ratio of bud size to par-ent cell size (155), and the stepwise appearanceof certain enzymes (65).Synchrony once attained frequently decays
rapidly. The parent cell always buds considera-bly before the daughter cell in haploid strains(19) as well as in diploids that are homozygousfor mating type, and the same is probably trueof diploid strains growing slowly on poor carbonsources. Hence, the longest maintenance ofsynchrony is to be expected with diploid orhigher ploidy cells growing rapidly. It is quite asimple matter to determine whether or not aparticular combination of strain and cultureconditions produces synchronous budding ofparent and daughter cell and is therefore condu-cive to the maintenance of synchrony. Becausecell association is frequently maintained aftermother and daughter have both budded, onecan merely examine asynchronous cultures mi-croscopically to see if such pairs have equal orunequal size buds.
Induction SynchronyAny method that induces an asynchronous
population to become synchronous is subject tothe criticism that the inducing treatment maycause profound changes in cellular physiologyand lead to aberrant behavior during the ensu-ing synchronous cycle. An advantage of induc-tion synchrony, however, is the large yield ofsynchronous cells that can be produced by thesemethods.One of the oldest and still most popular
methods of achieving synchronous yeast cul-tures is that devised by Williamson and Scopes(185, 195) which involves shifting stationary-phase cells between growth medium and starva-tion medium coupled with the selection of thelarger cells in the population. This techniquetakes advantage of the fact that stationarycultures of yeast produced by many types ofnutritional deprivation (see section on growthintegrated with division) are arrested synchro-nously with respect to cell cycle progress at apoint between cell separation and the initiationof DNA synthesis. However, stationary-phasecells are usually quite heterogeneous in size,and the degree of synchrony in starting new cellcycles upon subculturing in fresh medium ismarkedly improved by selection of the largercells in the population (184). The media shiftsmay induce some additional phasing of the cells(193). The method is capable of producingenormous quantities of highly synchronouscells, but suffers from the fact that two pro-cesses are occurring simultaneously, synchro-
166 HARTWELL
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
nous division and synchronous emergence fromstationary phase. The stepwise occurrence ofany particular event may be a consequence ofthe latter as well as the former.Any method that arrests cell division at a
unique stage is potentially capable of giving riseto a synchronous culture. X rays (163), hydrox-yurea (159), temperature-sensitive cdc-mutants(130), and the mating substances, a factor (18,73) and a factor (L. Wilkinson and J. Pringle,submitted for publication), have all been usedin this capacity with some degree of success.
Selection SynchronySelection synchrony involves selecting a
small portion of an asynchronous population ofcells on the basis of some physical attributewhich changes during the cell cycle. Themethod suffers from the disadvantage that onlya small percentage of the original population(usually less than 10%) can be used and, hence,has a much lower yield than induced synchronytechniques. One major advantage is that thetechnique does not require unbalanced growthand should, in theory, therefore cause fewerperturbations in the ensuing cycle. Further-more, selection techniques can be adapted toexamine events that occurred prior to the selec-tion procedure during asynchronous growth,thus avoiding the perturbation problem al-together.Two methods have been employed to achieve
selection synchrony. One is the method ofMitchison and Vincent (116) which involvesselecting the youngest cells by their slow rate ofsedimentation in a sucrose gradient. This tech-nique has been applied to S. cerevisiae byTauro, Halvorson, and Carter (64, 168) whoobtained a fraction of cells containing 80%unbudded cells which displayed partially syn-chronous growth upon subculturing. However,this method did cause physiological changes inthe cells because a 1-h lag in the commence-ment of division was observed in an asynchro-nous culture derived by recombining all of thefractions from such a sucrose gradient (168).Perhaps a substance of lower osmolarity thansucrose would not produce this detrimentaleffect.
Sebastian, Carter, and Halvorson have usedrate sedimentation on sucrose gradients in azonal rotor to fractionate an asynchronous pop-ulation of cells according to size (150) and,hence, according to age in the cell cycle. Theywere able to obtain synchronous cultures fromsamples removed from the top, the middle, orthe bottom of the gradient. Because the entirecycle is displayed on the gradient, this method
has two important advantages. First, all of thecells in the population can be used, and, second,subculturing of the cells is often unnecessary.Direct chemical measurements on the frac-tionated cells allowed an analysis of composi-tional changes in DNA and enzymes during thecell cycle, and pulse labeling with a radioactiveprecursor prior to fractionation would permit ananalysis of synthesis during the cycle. It wouldbe helpful for assessing the potential utility ofthis technique if additional parameters weremonitored in the gradients, such as the presenceor absence of buds, bud size, and the morpho-logical stage of the nucleus.A second method takes advantage of the
observations by Mitchison (112) and Scopesand Williamson (147) that mass and volume donot increase coordinately during the yeast cellcycle, thus producing a density fluctuation.Upon banding an asynchronous culture isopyc-nically in a density gradient of low osmolarity,the cells are fractionated so that the lightestcells are near the end of the cycle and thedensest cells are near the time ofDNA synthesis(69, 183). Partially synchronous cultures can beobtained from either fraction.This selection technique has also been
adapted to avoid the necessity of subculturingthe cells after fractionation. In one experimentan asynchronous culture of S. cerevisiae waspulse labeled with radioactive amino acids, andthen the population of cells was isopycnicallybanded (155). By determining the radioactivityincorporated into ribosomal proteins isolatedfrom the lightest or the densest cells, it waspossible to follow the rate of synthesis of theseproteins at two points in the cell cycle. Further-more, by interspersing a chase period betweenthe time of labeling and the time of isopycnicbanding, synthesis at any time in the cyclecould be examined.
CELL DIVISION CYCLE MUTANTS
Diagnostic Landmarks and TerminalPhenotypes
One of the strong advantages of S. cerevisiaeas an experimental organism is its geneticsystem (for a review, see reference 123). Thischaracteristic has been exploited for cell cycleanalysis in the isolation of cell division cycle(cdc) mutants (75). In theory, a cdc geneproduct is necessary for one and only one of thediscontinuous events of the cell cycle. Empiri-cally, a conditional mutation in a cdc gene isrecognized by the fact that, after a shift to therestrictive condition, all of the mutant cellsfrom an asynchronous clone first exhibit defec-
167VOL. 38, 1974
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
tive behavior at the same landmark in the cellcycle. This landmark will be defined as thediagnostic landmark (the term initial defect hasbeen applied to this parameter previously [74],but the word initial carries a misleading conno-tation). A consequence of the fact that all cellsfrom an asynchronous population first arrestnormal development at the same landmark isthat nearly all of the mutant cells assume thesame, albeit often aberrant, morphology (theterminal phenotype, previously termed the ter-mination point [751) after extended incubationat the restrictive temperature. It is this lattercharacteristic that has permitted the detectionof cdc mutants (75, 76) from among a largercollection of temperature-sensitive mutants(68).
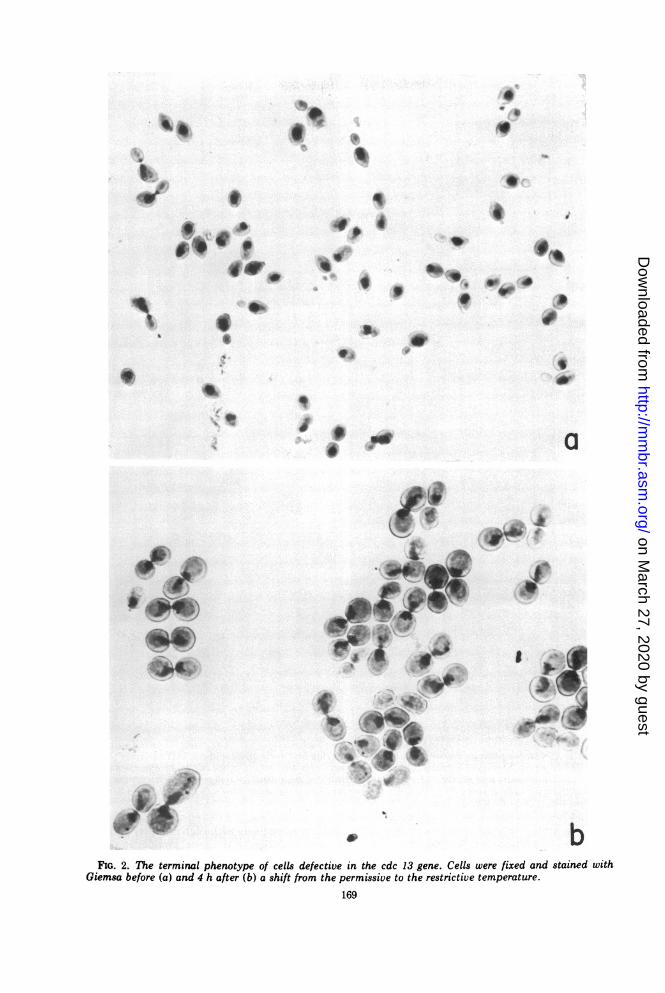
Cells carrying the cdc 13 mutation that werefixed and stained before and after incubation atthe restrictive temperature are displayed inFig. 2a and b, respectively. The diagnosticlandmark for this mutant is medial nucleardivision. The population of cells arrests afterhaving completed bud emergence, DNA synthe-sis, and nuclear migration, but before complet-ing medial nuclear division, late nuclear divi-sion, cytokinesis, or cell separation. Bud growthcontinues, but no new buds appear. Hence, theterminal phenotype is a cell with a large budand a nucleus located in the cell neck. Theidentical morphologies of these cells are strik-ingly different from the originally asynchronouscell population and from other non cdc temper-ature-sensitive mutants even when the nucleusis not revealed by staining. The terminal pheno-type for different cdc genes is often different,but is the same for all alleles of a particulargene, and this fact strongly suggests that each ofthe cdc genes does indeed perform an indispen-sible function in one and only one of thediscontinuous cell cycle events (76).
First Cycle Arrest and Execution Pointscdc mutants can be classified into two cate-
gories depending upon the number of cell cyclesthat they complete after a shift to the restric-tive temperature. In the first category are cdcmutants that traverse several cell cycles after ashift to the restrictive temperature. In thesecond category are mutants that behave asfollows: those cells that are before a certainpoint in the cell cycle (the execution point) ar-rest development in the first cycle at the re-strictive temperature and those cells that areafter the execution point complete the firstcycle, and arrest in the second cell cycle (75).Mutants in this second category are said toexhibit first cycle arrest, because a cell that
commences the cycle at the restrictive tempera-ture will arrest in the first cycle.
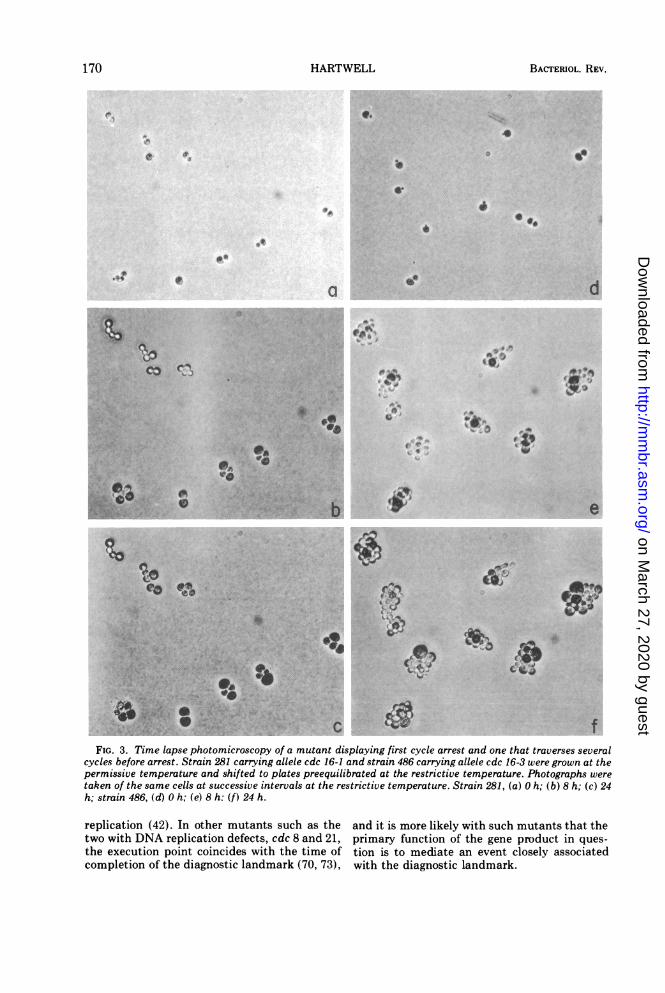
In several cdc genes there are representativesof alleles that do give first cycle arrest and otheralleles that do not give first cycle arrest (76).Mutants carrying different alleles of the cdc 16gene are displayed in Fig. 3 before and afterincubation at the restrictive temperature.These two mutants both arrest. with the sameterminal phenotype as that displayed in Fig. 2,a cell with one large bud, but one allele permitsseveral cycles of division before arrest at therestrictive temperature and the other allelecauses arrest in the first or second cycle depend-ing upon the position of the cell in the cycle atthe time of the shift. Although there are severalmolecular mechanisms consistent with thesetwo responses, the presence in a gene of allelesthat do not exhibit first cycle arrest suggeststhat this gene product is normally present inexcess. Hence, other alleles of this same genethat do exhibit first cycle arrest are probablythermolabile for the function and not for thesynthesis of the gene product in question.
For those mutant strains that do exhibitfirst-cycle arrest the execution point can bedetermined by examining many cells by time-lapse photomicroscopy (75, 76). A mutant de-fective in gene cdc 13 before and several hoursafter a shift to the restrictive temperature isdisplayed in Fig. 4. The terminal phenotype ofthis mutant is a cell with a large bud. Cells thatwere early in the cell cycle at the permissivetemperature, as evidenced by having no bud ora small bud, arrest at the restrictive tempera-ture as one cell with a large bud. Cells that werelater in the cell cycle, as evidenced by largerbuds, arrest development after forming twocells each with a large bud. Therefore, theexecution point for this mutant is passed duringgrowth at the permissive temperature at a pointin the cycle represented by the arrow in Fig. 4.From the fraction of cells before the executionpoint one can calculate the fraction of time inthe cell cycle at which execution occurs aftercorrecting for the age distribution of the popula-tion (76).The execution point in some mutants consid-
erably precedes the time of occurrence of thediagnostic landmark. The primary defect forsuch mutants must lie in some early, as yetundetected, process upon which the completionof the diagnostic landmark is dependent. Forexample, many of the nuclear division mutantsexhibit execution points during the DNA rep-lication period, and one surmises that pro-cesses which are necessary for chromosomesegregation normally occur during chromosome
168 HARTWELL
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from
9'
*4
.1f
S
'C
0aI-
SS
5.
o,0 9
i
ee_
I
.tOlv 0
0a
TO
-
U_I'
4.
v bFIG. 2. The terminal phenotype of cells defective in the cdc 13 gene. Cells were fixed and stained with
Giemsa before (a) and 4 h after (b) a shift from the permissive to the restrictive temperature.
169
'S
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from
BACTERIOL. REV.
FIG. 3. Time lapse photomicroscopy of a mutant displaying first cycle arrest and one that traverses severalcycles before arrest. Strain 281 carrying allele cdc 16-1 and strain 486 carrying allele cdc 16-3 were grown at thepermissive temperature and shifted to plates preequilibrated at the restrictive temperature. Photographs weretaken of the same cells at successive intervals at the restrictive temperature. Strain 281, (a) 0 h; (b) 8 h; (c) 24h; strain 486, (d) 0 h; (e) 8 h: (f) 24 h.
replication (42). In other mutants such as the and it is more likely with such mutants that thetwo with DNA replication defects, cdc 8 and 21, primary function of the gene product in ques-the execution point coincides with the time of tion is to mediate an event closely associatedcompletion of the diagnostic landmark (70, 73), with the diagnostic landmark.
170 HARTWELL
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
FIG. 4. Determination by time lapse photomicros-copy of the execution point in a cdc mutant. Cells ofstrain 428D1 with a temperature-sensitive mutationin gene cdc 13 were grown at 23 C and shifted ontoplates pre-equilibrated to 36 C. Cells were photo-graphed at the time of the shift, and individual cellswere cut from larger fields and arranged in order ofbud size at the time of the shift (inner circle). Thesame fields were photographed after 6 h incubation atthe restrictive temperature, and the cell(s) producedby each individual cell on the inner circle was cut outand arranged (outer circle). Each cell on the outercircle consists of a parent cell with a large budattached. Cells early in the cycle (before EX) arresteddevelopment in the first cell cycle at the restrictivetemperature and cells later in the cycle (after EX)finished the first cycle and arrested in the secondcycle.
The Mutant CollectionExamination of about 1,500 temperature-sensi-tive mutant clones by morphological criteriarevealed the presence of 148 cdc mutants (76).Complementation studies demonstrated thatthese mutations define 32 complementationgroups and that mutations in each of thesegroups are recessive. Genetic analysis suggestedthat 30 of these complementation groups are
defined by single mutations in nuclear genes,and 14 of these genes were mapped on the yeastgenetic map (76). (Recent results indicate thatthe single mutant representing cdc 22 carriestwo mutations, both of which are necessary fortemperature sensitivity [L. H. Hartwell, unpub-lished data], and the mutant representing cdc32 could not be analyzed because it producesinviable spores in crosses.) No evidence for theclose clustering of functionally related geneswas found in this study. A summary of thediagnostic landmarks, terminal phenotypes,and execution points of mutants with lesions inthe cdc genes is presented in references 74 and
76. Mutant strains harboring these mutationsmay be obtained from the Yeast Stock Center,Dr. R. K. Mortimer, Donner Laboratory, Uni-versity of California, Berkeley, Calif. 94720.
Role of cdc Genes in the Life CycleAlthough these mutations were isolated in
haploid cells of a mating type during vegetativegrowth, subsequent analysis has demonstratedthat most and perhaps all perform indispensiblefunctions for the mitotic cycle at every stage ofthe life cycle. Life cycle stages that are basicallydistinct either in the origin of the cells or in thecapabilities of the cells but which require amitotic cycle include: vegetative growth in cellsof a, a, and a/a mating type, cells emergingfrom stationary phase to vegetative growth,zygotes resulting from the fusion of two hap-loid cells, and spores germinating to vegetativegrowth (Fig. 5). Most of the cdc genes havebeen tested for their requirement in the mitoticcycle at each of these stages of the life cycleand with possibly only one exception all genestested are required at all stages (76; Pringle,Maddox, Reid, and Hartwell, unpublisheddata).Perhaps of greater interest is the question of
whether the genes that mediate mitosis alsoperform indispensible functions for meiosis.The same cdc mutations have been tested byGiora Simchen for their role in the meiotic cycle(157). Of 20 genes that were tested, 13 (cdc 2, 4,5, 7-9, 13, 14, 16, 17, 20, 23, and 28) appear to beessential for meiosis, because diploid cellshomozygous for these mutations were drasti-cally impaired in their ability to sporulate at therestrictive temperature and, in some cases, evenat the permissive temperature.
COMPONENT PROCESSES OF THECELL CYCLE
This section will review some of the currentinformation available on the cytological, geneti-cal, and biochemical processes that underlie thelandmarks of the cell cycle. The reader isreferred to the authoritative review by Matile etal. (108) for a more comprehensive discussion ofyeast cytology.
Bud EmergenceThe future daughter cell appears early in the
cell cycle as a discrete morphological entity, thebud. The site at which the bud arises becomes achannel (the neck) connecting parent anddaughter cell through which the nucleus andother organelles pass into the daughter cell, inwhich the nucleus divides and at which cyto-kinesis and cell wall separation take place.
Early in the cell cycle, prior to the emergence
171VOL. 38, 1974
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
nutrientreplenishment
0 0\mitosis meiosis ond
sporulotion
Zgote Ascusspore ---
germination
FIG. 5. The life cycle of S. cerevisiae emphasizingthe relationship of the mitotic cycle to the otherdevelopmental pathways; a and a refer to mating typealleles; unlabelled cells may be a. a, or aa. Mitotichaploid cells of mating type a or a may enterstationary phase under conditions of nutrient deple-tion or may fuse with haploid cells of opposite matingtype to create a diploid zygote which in turn producesaa diploid mitotic cells. Mitotic aa diploid cells mayenter stationary phase under conditions of nutrientdepletion or may undergo meiosis and sporulation in amedium containing acetate and lacking a nitrogensource. The resulting haploid a and a spores willgerminate to produce haploid mitotic cells whennutrients are replenished. Not shown on the figure are
other pathways of development: haploid stationarycells or spores may fuse to form a zygote withoutpassing through the mitotic cycle and diploid station-ary cells may undergo meiosis and sporulation with-out the intermediacy of a mitotic cycle.of the bud, a stainable circular rim which isprobably a precursor of the bud scar appears onthe parent cell (78). The scars which appear onmother and daughter cells after cell separationcontain chitin. Hayashibe and Katohda haveclaimed that the chitin is synthesized prior tothe budding event and hence is temporally andprobably spacially associated with the appear-ance of this circular rim (78). This interpreta-tion has been challenged by Cabib et al. (25),who suggest an inadequacy of the methodemployed by Hayashibe and Katohda to mea-
sure chitin and who support the view thatchitin is synthesized later in the cell cycle nearthe time of cell separation (see below). If thelatter interpretation is correct, then the chitinwould have to be added to the rim later in thecycle as it matured into the bud scar.
Small membrane-bound vesicles accumulateprior to bud emergence, and they are foundlocalized at the site of emergence of the youngbud (119; Fig. 8b, d). The plasmalemma of the
young bud contains small circular invaginationswhen observed by freeze-fracture electron mi-croscopy, and it has been suggested that theinvaginations arise by the fusion of these vesi-cles with the plasmalemma of the new bud(119).Within the cytoplasm, adjacent to the cyto-
plasmic membrane, a ring of fibrous materialhas recently been detected and been shown tobe present in the youngest bud detectable andto remain in the neck throughout the growth ofthe bud (B. Byers, personal communication).The ultimate fate of this structure at the time ofcell separation is unknown.The suggestion that the bud emerges explo-
sively as a naked protoplast due to the localizedrupture of the cell wall (131) appears to bewrong in detail because even the smallest budscontain a thick cell wall (107, 109, 154). Never-theless, the basic idea inherent in this proposalthat the force behind bud emergence may be thehydrostatic pressure of the cell acting on alocalized weakening of the wall is still a viablepossibility and may account for the observationthat the first few buds on a new parent celloccur in the region of maximal wall curvature(51).
It has been suggested that the double-plaqueconfiguration of the spindle plaque may play acausative role in the budding process since cellswith very small buds aways contain a double-spindle plaque (20). Furthermore, the doubleplaque is found closely associated with andoriented in the direction of the new bud with theextranuclear microtubules extending into thebud (Fig. 8d; 20). Observations on two cdcmutants support the contention that the doubleplaque has a role in bud emergence. Culturescontaining the cdc 1 mutation arrest with someof the cells containing tiny buds and these cellsalso have the double-spindle plaque configura-tion while other cells arrest without buds andthese cells have a single-spindle plaque (20).Cells carrying a mutation in cdc 4 arrest in thedouble-plaque configuration and although theydo not initiate DNA synthesis, they do undergoseveral cycles of bud emergence terminatingdevelopment with as many as five buds on asingle cell (20, 70).The phenotype of mutants defective in cdc 4
also suggests that some type of clock controlsthe bud emergence event because successivebudding events at the restrictive temperaturemaintain a periodicity of about one cell cycletime (70). This clock runs in the absence ofDNA synthesis, nuclear division, cytokinesis, orcell separation, events which do not occur inthis mutant at the restrictive temperature.The cdc 24 gene product is probably inti-
0Stotionorp
cell
nutrientdepletion
0
0conjugat ion
172 HARTWELL
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
mately involved in bud emergence becausemutations in this gene block budding, but notDNA synthesis or nuclear division, and becausethe execution point is precisely at the time ofbud emergence (74). In populations of cellscarrying the cdc 1 lesion some of the cells arrestunbudded and others form tiny buds (71, 75);although the cells synthesize DNA, they do notcomplete nuclear division, and macromoleculesynthesis is rapidly arrested (71). It is difficultto attribute to this gene product a role in oneknown cell cycle event, especially in view of theproperties of the cdc 24 mutants. Mutations incdc genes 19, 22, 25, 29, and 32 also prevent budemergence (76), but they have not been wellcharacterized either because the available al-leles do not show first-cycle arrest or becausethe genetic situation is complicated.
Bud GrowthThe bud grows in size throughout the cell
cycle (112), and this fact greatly facilitates cellcycle studies with budding yeast because therelative size of the bud and parent cell providesa marker by which one can order individualcells within the cycle (187). The new daughtercell may continue to grow between the end ofone cell cycle (cell separation) and the begin-ning of the next (bud emergence), but theextent of this growth is markedly dependentupon the culture conditions. Mitchison (112)measured individual cells during the course ofthe cell cycle and found that the mother cellmaintained a constant volume, whereas John-son (92), who measured a population of cellsand ordered individuals according to bud size,concluded that the mother cell may undergo asmuch as a 50% cyclic variation in volume overthe course of the cycle. The parent cell increasesslowly in volume over the course of manygenerations, and it has been suggested that thisincrease may be due to the accumulation of budscar material (102, 125).The yeast wall is composed of glucan, man-
nan, and protein with small amounts of chitin.The bud wall is not derived from the material ofthe mother cell wall but is newly synthesized(32, 174). The glucan and mannan componentsof the wall are synthesized continuouslythroughout the cell cycle (156). Three groupshave investigated the topology of wall growthduring the division cycle. The site of addition ofnew glucose residues to the glucan component ofthe bud wall was determined by autoradiogra-phy of cells pulse labeled in [3H glucose andextracted to remove all nonglucan material(94). New glucan was added at the tip of thebud distal to the parent cell. 2-Deoxyglucose
induces lysis of growing yeast cells at this samesite possibly by inhibiting the synthesis ofglucan without preventing glucan breakdown atthe growing point in the wall (93).The appearance of newly synthesized man-
nan on the cell wall was studied by labeling oldmannan with fluorescein-conjugated con-canavalin A, followed by growth for an intervalin the absence of concanavalin A conjugate(173, 174). The fluorescence was found to beuniformly distributed over the parent cell andthe region of the bud proximal to the parentcell. The portion of the bud distal to the parentcell was unlabeled, indicating that the newlysynthesized mannan was added at the tip of thebud distal to the parent cell.The simple picture of cell wall growth at the
tip of the bud is confused by a third investiga-tion using fluorescein-labeled antibody whicharrived at a different conclusion (32). Newlysynthesized wall antigen was found to be laiddown on the new bud in the region proximal tothe parent cell. It seems that there are only twoways to reconcile these observations. Eitherthere are strain differences in the mode of wallsynthesis or the unidentified antigen(s) investi-gated in the third study represents a thirdcomponent of the wall that is added to the newbud wall at a point 180° removed on the budsurface from the site of addition of the glucanand mannan components.The addition of an extracellular enzyme,
invertase, to the wall has also been studied byusing fluorescent antibody and found to bedeposited only in newly synthesized bud walls(175). The reported results were not of sufficientresolution to decide between addition at the tipof the growing bud distal to the parent cell or atthe neck region, proximal to the parent cell.
Cytokinesis and Cell SeparationWhen the mature bud separates from the
parent cell at the conclusion of the cell cycle(cell separation), a permanent birth scar re-mains on the surface of the bud and a perma-nent bud scar is left on the parent cell (1, 5, 6).The bud and birth scars can be observed byfluorescence microscopy following staining withprimulin (Fig. 6; 165, 166) or brightener (78),and the number of generations that a particularcell has undergone can be determined by count-ing the number of bud scars (9). The bud scarhas the appearance of a crater on the surface ofthe parent cell with a raised, circular rim that iscomposed in part of chitin (3, 21).An elegant biochemical and cytological anal-
ysis of the cell separation or septation processhas been undertaken by Cabib and co-workers
VOL. 38, 1974 173
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from
HARTWELL
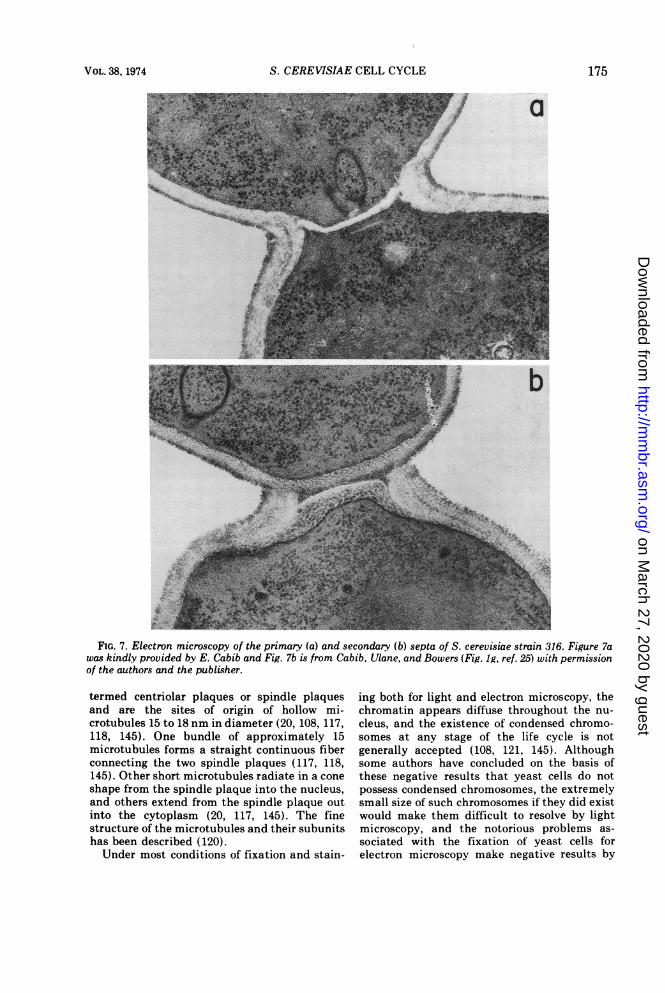
(for a review see reference 25). Cell separationhas been divided into two cytologically distin-guishable stages: the formation of the primaryseptum, which is composed of chitin, and theaddition tothis of a secondary septum which iscomposed of glucan (Fig. 7; 25). The chitin issynthesized during a discrete interval of the cellcycle (22), sometime between bud emergence
and cell separation, although the precise timingof this synthesis relative to other cell cycleevents is not obvious from the published data.The suggestion that the chitin is actually syn-
thesized prior to bud emergence (78) is disputedby Cabib et al. on technical grounds (25) as
discussed previously.A particulate enzyme preparation capable of
chitin synthesis has been described by Cabiband co-workers, and the chitin synthetase isfound in cell lysates as an inactive zymogen thatcan be converted to active enzyme by incuba-tion with trypsin or a vesicle-bound activatorfound in yeast cells (22, 23, 96). The activator,which is a proteolytic enzyme (77), is in turn
subject to inhibition by a heat-stable protein(22). A model invoking these three elements hasbeen proposed to account for the localizeddeposition of chitin in the bud scar rim (22, 24,25).
Four gene products that are necessary for cell
FIG. 6. The bud scars of a diploid strain of S.cerevisiae. Note the presence of a stained rim on theneck of the budded cell. This photograph was kindlyprovided by Streiblova and Beran. It appeared previ-ously in Beran (9), and is reproduced with permissionof the authors and publisher.
separation (cdc 3, cdc 10, cdc 11, and cdc 12)have been identified by mutation (71). Thesemutants do not complete the process of cyto-kinesis or cell membrane separation (71) basedupon an empirical test; however, the relation-ship between the point of arrest in these mu-tants and the two stages of septation defined byCabib et al. (25) is at present unknown. First-cycle arrest is displayed by some of the mutantsin three of these genes, and the execution pointsfor all three are early in the cell cycle considera-bly before cytokinesis itself. In fact, two exhibitexecution points near the time of bud emer-gence. It would be interesting to examine thesemutants for defects in formation of the circularrim that precedes bud emergence or in chitinsynthesis.The claim that "protein disulfide reductase"
is a cellular division enzyme in yeasts (132)deserves comment. This conclusion stems fromthe observation that a morphologic variant ofCandida albicans isolated in 1940 (106) waslater found to have a lower activity of proteindisulfide reductase than its parent strain (132).The comparison of two strains separately cul-tured for 16 years is of questionable validity but,even granting that the biochemical differencearose at the time of the morphological differ-ence, the two changes may both be pleiotropicmanifestations of some other primary lesion.Until it can be demonstrated that a singlegenetic alteration in the structural gene for theprotein, disulfide reductase, results in filamen-tous growth, the conclusion that this enzymeplays a role in division is unwarranted.
Nuclear Structure
The nucleus of the yeast cell is surrounded bya pair of double-unit membranes, the nuclearenvelope, which remains intact throughout themitotic cycle (108). The nuclear envelope isstudded with circular pores approximately 80 to90 nm in diameter at a density of 1 to 1.5 persquare micrometer (121) that are filled withgranules smaller than ribosomes (145).A portion of the nucleus stains differentially,
has a crescent shape during part of the mitoticcycle, and is believed to be the nucleolus (145).Evidence has been presented that the crescentregion is the site of synthesis of ribosomalribonucleic acid (RNA) (162).
During part of the mitotic cycle two disks arepresent in the nuclear membrane. The crosssection of the disks appear by electron micros-copy as bands of material forming a structureabout 150 nm in length and several hundrednanometers wide. These structures have been
BACTERIOL. REV.174
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from
S. CEREVISIAE CELL CYCLE
FIG. 7. Electron microscopy of the primary (a) and secondary (b) septa of S. cerevisiae strain 316. Figure 7awas kindly provided by E. Cabib and Fig. 7b is from Cabib, Ulane, and Bowers (Fig. 1g, ref. 25) with permissionof the authors and the publisher.
termed centriolar plaques or spindle plaquesand are the sites of origin of hollow mi-crotubules 15 to 18 nm in diameter (20, 108, 117,118, 145). One bundle of approximately 15microtubules forms a straight continuous fiberconnecting the two spindle plaques (117, 118,145). Other short microtubules radiate in a coneshape from the spindle plaque into the nucleus,and others extend from the spindle plaque outinto the cytoplasm (20, 117, 145). The finestructure of the microtubules and their subunitshas been described (120).Under most conditions of fixation and stain-
ing both for light and electron microscopy, thechromatin appears diffuse throughout the nu-cleus, and the existence of condensed chromo-somes at any stage of the life cycle is notgenerally accepted (108, 121, 145). Althoughsome authors have concluded on the basis ofthese negative results that yeast cells do notpossess condensed chromosomes, the extremelysmall size of such chromosomes if they did existwould make them difficult to resolve by lightmicroscopy, and the notorious problems as-sociated with the fixation of yeast cells forelectron microscopy make negative results by
VOL. 38, 1974 175
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
this technique equally unconvincing. Indeed,Williamson, by using permanganate fixation,has described structures which he suggested tobe condensed chromosomes (188), and Tamaki(167) and Fischer et al. (50) have reportedseeing condensed chromosomes visible by lightmicroscopy, although these reports are not con-sidered to be definitive (108). Chromatin fibersfrom S. cerevisiae have been reported to besimilar in structure to those from higher eu-karyotes in that they have a diameter of about17.5 nm (58). Proteins have been extracted fromyeast chromatin and have been termed histonesbecause they are basic and because they appearto influence the accessibility of the DNA fortranscription and the structure of DNA asmeasured by thermal denaturation (176, 179).However, the relationship of these proteins tothe histones of higher eukaryotes and their rolein vivo are unknown.
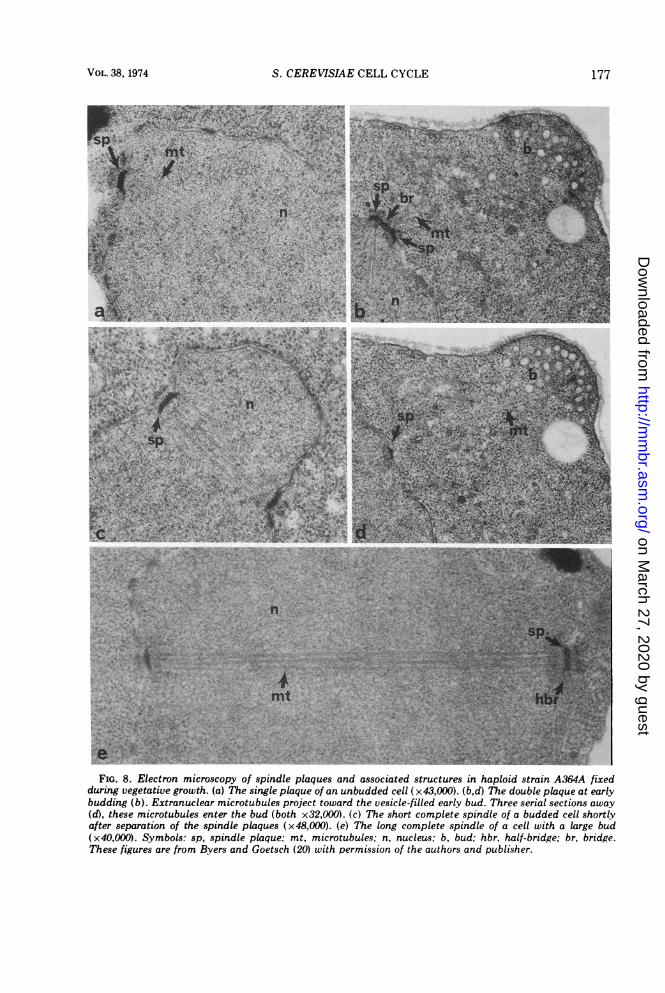
Spindle plaque cycleThe cytology of the spindle plaque has been
studied during the mitotic cycle by Moens andRapport (117) and in cdc mutants by Byers andGoetsch (20). An unbudded cell early in the cellcycle contains a single, dense spindle plaquelying in the nuclear envelope (Fig. 8a). Bothintra- and extranuclear microtubules originatefrom it (20, 117, 145). At about the time of budemergence the spindle plaque duplicates, re-sulting in two plaques lying side by side andconnected by a short bridge (Fig. 8b; 20, 117).This double-plaque configuration is usuallyfound in close proximity to the site of budding.The extranuclear microtubules emanating fromthe outer surface extend into the new bud and,as discussed earlier, it has been suggested thatthe double-plaque configuration plays a causa-tive role in the budding process (20). The cdc28 mediated step is the earliest known gene-controlled event in the cell cycle (see section onevents of cell cycle coordination), and muta-tions in this gene specifically prevent plaqueduplication (20).
Next, the two spindle plaques rapidly migrateto opposite sides of the nucleus while remainingwithin the nuclear membrane, and a continuousstraight fiber of microtubules of length about0.8 ,um connects them (Fig. 8c). This structurehas been termed the complete spindle (20, 117).The cdc 4-mediated step is the second knownstep in the pathway of gene-controlled events,and the expression of this gene is necessary forthe separation of the two spindle plaques (20).-Some time after DNA replication the nucleus
migrates into the neck of the bud, and through-
out this process the two plaques maintain arelatively constant distance of separation ofabout 0.8 gum (20, 117). It has been emphasizedthat the force for nuclear migration and for theinitial stage of nuclear elongation is probablynot supplied by the complete spindle becauseits orientation is random with respect to thelong axis of the cell and because the spindleoften does not extend the length of the nucleus(20, 145).
After nuclear migration and prior to thecompletion of nuclear division the completespindle abruptly elongates in the long axis ofthe cell to about 6 to 8 Atm as does the entirenucleus (Fig. 8e; 20, 117, 145). The two spindleplaques are now found at opposite ends of theelongated nucleus, and it is not unlikely thatthe growth of the spindle provides the force forthis final stage of nuclear elongation. It istempting to imagine that chromosome segrega-tion occurs during the spindle elongation, butthere is no evidence to support this idea. Thefinal spindle elongation is blocked in mutantsdefective in the initiation ofDNA synthesis (cdc7), DNA synthesis (cdc 8 and 21), and medialnuclear division (cdc 2, 6, 9, 13, 16, 17, 20, 21,and 23; reference 20). Some of the latter groupof mutants might well be defective in processesdirectly involved in chromosome segregationand nuclear division such as the spindle elonga-tion step. However all of the mutants have anapparently normal complete spindle and,hence, there is no evidence that any are defec-tive in microtubule assembly (B. Byers and L.Goetsch, personal communication).
Vesicles of endoplasmic reticulum accumu-late in the bud neck as the nuclear membraneand cytoplasmic membranes pinch in two (145).Mutants defective in late nuclear division (cdc5, 14, and 15) arrest at the elongated spindlestage and could be defective in one of theseterminal steps of nuclear division (20).
Nuclear DNA ComponentsThe total amount of DNA in a haploid yeast
cell has been reported by various workers to bebetween 1.2 and 1.4 x 10'0 daltons (33, 48, 133,148, 194). About 90% of this DNA bands inCsCl2 gradients at a density of about 1.699g/cm3 (127), and most of this main band DNA iscontained in the nucleus because it is retainedin isolated nuclei (12, 40).
Renaturation studies with whole-cell DNAindicate that the bulk of the cellular DNA has akinetic complexity equivalent to a genome sizeof 9.2 x 109 daltons (13), a value which is inreasonably good agreement with the total DNA
176 HARTWELL
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from
S. CEREVISIAE CELL CYCLE
FIG. 8. Electron microscopy of spindle plaques and associated structures in haploid strain A364A fixedduring vegetative growth. (a) The single plaque of an unbudded cell (x43,000). (b,d) The double plaque at earlybudding (b). Extranuclear microtubules project toward the vesicle-filled early bud. Three serial sections away(d), these microtubules enter the bud (both x32,000). (c) The short complete spindle of a budded cell shortlyafter separation of the spindle plaques (x48,000). (e) The long complete spindle of a cell with a large bud(x40,OOO). Symbols: sp, spindle plaque: mt, microtubules; n, nucleus; b, bud; hbr, half-bridge; br, bridge.These figures are from Byers and Goetsch (20) with permission of the authors and publisher.
VOL. 38, 1974 177
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
content of the haploid cell. Although no evi-dence of a repetitive DNA fraction was seen inthis study, other data suggested the presence of11% repetitive DNA with a kinetic complexityof about 2 x 107 daltons in nuclear preparationsfrom Saccharomyces carlsbergenesis (31). Therepetitive DNA might be due in part to riboso-mal DNA and/or 2-gm circular DNA (see be-low).
Genetic studies have identified 17 independ-ently segregating groups of centromere-associated genes in diploid cells, and a statisti-cal assessment of this data suggested that thisnumber may represent the complete chromo-some complement (124). If a haploid cell con-tains 17 chromosomes the average chromosomewould have a molecular weight of about 5 to 6 x108 of DNA, an amount only about one-fifththat of the Escherichia coli chromosome (26).This small size probably accounts for the inabil-ity of cytologists to reproducibly visualize indi-vidual chromosomes in the yeast nucleus but,on the other hand, their small size has facili-tated physical studies of intact yeast nuclearDNA.
Sedimentation studies reveal that the bulk ofthe cellular DNA sediments as large moleculeswith an average molecular weight number ofabout 6 x 108 (14, 138). In one study thedistribution of molecular weights was found tobe broad, ranging from 5 x 107 to 1.4 x 108(138), whereas in another it was narrow, rangingfrom 4 x 108 to 6 x 108 (14). The average molec-ular weight number of DNA in these investiga-tions is in striking agreement with the expectedaverage molecular weight of DNA per chromo-some and suggests that each chromosome maycontain a single DNA molecule. However, sincethe equations that relate sedimentation velocityof DNA molecules to molecular weights havenot been validated with molecules this large,the sedimentation data alone does not provethis important point.
Direct visualization of yeast DNA moleculesbefore and after velocity sedimentation revealedthat the molecules are linear and range incontour length from 50 to 355 qm (excludingmitochondrial DNA and 2-gm circular DNA)with an average value of 165 jim (137, 140). Theaverage length corresponds to a molecularweight of 3.8 x 108. Because any artifactualbreakage of these long molecules would lead toan underestimate of their true size these dataare interpreted to demonstrate that the yeastchromosomes each contain a single, linear DNAmolecule that is continuous through the cen-tromere (137, 140).The availability of yeast strains that are
monosomic or disomic for one or a few chromo-somes provides a method for associating thegenetic linkage groups with specific-size DNAmolecules by the cosedimentation of differen-tially labeled aneuploid and euploid DNA prep-arations (14, 49, 55, 140). Data obtained by thisapproach suggest that the linkage group desig-nated as chromosome I is contained on a DNAmolecule of about 4 x 108 daltons (49) and thatchromosome VIII is associated with a moleculeof about 4 to 5 x 108 daltons (14, 140). Thesecorrelations of changes in sedimentation profilewith alterations in ploidy for certain nucleargenes demonstrate that the large DNA mole-cules are indeed the repository of the nucleargenetic information.A satellite DNA band of a density of 1.705
g/cm3 (y band) is produced from nuclearDNA when the large molecules are sheared tomolecular weights of 3 x 107 or less, but is notobserved if the nuclear DNA molecules aregreater than 8 x 107 daltons (40); Gamma-bandDNA accounts for 11% of the total nuclear DNAand hybridizes with 18 and 25S ribosomal RNA(40, 43) and contains the 5S ribosomal RNAcistrons (146) as well. Because there are about140 ribosomal RNA cistrons in the haploidgenome of S. cerevisiae (43, 52, 144, 149) thatare each transcribed into a molecule of 2.5 x 106daltons (178), it can be calculated that aboutone-half of the My band DNA is accounted for bythe ribosomal RNA cistrons (40). Furthermore,the shear data suggests that these are clusteredin units of 10 to 32 cistrons (40) and that theindividual units are separated from one anotherby DNA of main-band density (40, 44). Themajority of the ribosomal cistrons appear to belocated on chromosome I (95, 135), based uponthe fact that strains monosomic for chromosomeI have a lower percentage of rDNA.
Initiation of Nuclear DNA SynthesisIn synchronous yeast cultures the majority of
the cellular DNA replicates early in the cellcycle at about the time of bud emergence (134,186, 192). However, because results with syn-chronous cultures are always suspect, a moredefinitive investigation is that of Williamsonwho showed by an autoradiographic approachthat in unperturbed, asynchronous populationsmost of the cellular DNA (and therefore thenuclear DNA) replicates during an intervalcomprising approximately one-quarter of thecell cycle beginning at about the time of budemergence (187).
Events that are required for the initiation ofDNA synthesis (that is, events that constitute aprecondition for the onset of synthesis), but are
178 HARTWELL
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
not required for DNA replication (elongation)itself, should satisfy the following criterion. Allcells that have entered the DNA syntheticperiod will have completed the event in ques-tion and, hence, they will be able to completethe full round of DNA synthesis after inhibitionof the initiation event is imposed. Because theDNA synthetic period occupies only about 25%of the cell cycle, the imposition of an initiationblock in an asynchronous culture would permita DNA increase of only about 15% (80). In asynchronous culture if the inhibition of initia-tion is imposed just after completion of thesensitive event then nearly a complete round ofDNA synthesis should occur.
Evidence of this type has been presentedsuggesting that a factor (18, 81) and a factor (L.Wilkinson and J. Pringle, submitted for publi-cation), the mating pheromones (see below),cycloheximide and anisomycin (inhibitors ofprotein synthesis; 80, 160, 190), and lesions inthe cdc 28, cdc 4, and cdc 7 genes (73, 80)prevent the initiation but not the completion ofDNA synthesis. However, this type of datum isnot sufficient to distinguish a true initiationblock from a leaky replication block, a replica-tion block with a delayed onset of expression, ora specific block in the early portion of DNAsynthesis.Such a distinction can, however, be made by
a direct examination of DNA molecules forreplication structures. As will be pointed outbelow, DNA molecules in the process of replica-tion contain "bubbles" and "forks" that can bevisualized by electron microscopy (130, 140).When the DNA obtained from a replicationmutant (cdc 8) that had been incubated at therestrictive temperature for an interval sufficientto accumulate all of the cells at the block wasexamined, a large fraction of the moleculescontained multiple small bubbles (T. D. Petesand C. S. Newlon, personal communication).This result is interpreted as being due to theslow, "leaky" replication of DNA in essentiallyall of the cells. However, when the DNA fromthe purported initiation of DNA synthesis mu-tant (cdc 7) was examined, very few moleculeshad bubbles. Nevertheless, nuclear DNA syn-thesis in the cdc 7 mutant at the restrictivetemperature was even more leaky than that inthe cdc 8 mutant. This result is interpreted tomean that most of the cells with the cdc 7mutation are arrested prior to the actual begin-ning of chromosome replication. Leakiness ofDNA synthesis in this mutant must be due toan occasional cell that leaks through the initia-tion block and then rapidly replicates its entiregenome so that only a small fraction of the
molecules from the population contain bubblesat any one time. Because the other purportedinitiation blocks, a factor, a factor, protein syn-thesis inhibitors, and mutations in cdc 28 andcdc 4, have been shown to be steps that precedethe cdc 7-mediated step in the cell cycle (81),these results provide strong confirmation of thesuggestion that all of these agents act on pre-conditions for DNA replication.The finding that inhibitors of protein synthe-
sis impose blocks specifically in the initiation ofDNA synthesis suggests that S. cerevisiae un-like most eukaryotes is capable of replicating itsgenome in the absence of protein synthesis.DNA synthesis in the absence of protein synthe-sis apparently leads to the complete duplicationof all chromosomes since cells that have rep-licated their DNA under these conditions cansubsequently complete nuclear division (160)and cell division (80) without further DNAsynthesis. Hence, all of the proteins neededboth for the initiation and the replication of the200 replicons (140) in the yeast genome arepresent at the onset of the S period. It is notclear whether these proteins synchronously ini-tiate all replicons at the beginning of S orwhether they merely accumulate at the begin-ning of S and are later used to mediate theasynchronous initiation of replicons throughoutthe S period.
Nuclear DNA SynthesisThe structure of replicating DNA molecules
from S. cerevisiae has been examined (130,140). DNA molecules which contained terminalforks and/or internal bubbles were obtainedfrom synchronous cultures in the S phase withhigh frequency but not in those obtained fromG1 cells (Fig. 9). Furthermore, in DNA obtainedfrom cells following a shift from isotopicallydense to light medium, molecules with thesestructures were highly enriched among thosethat were intermediate in density between thefully dense and fully hybrid peaks. Both of thesecorrelations strongly suggest that forked andbubbled structures are replication intermedi-ates. The terminal forks are assumed to arise byinternal initiation of replication that had pro-gressed by replication to the molecular terminusor by terminal initiation sites that had pro-gressed internally. The bubbles are presumed toarise by internal initiation of replication. A highproportion of these molecules contained multi-ple initiation sites and the center-to-centerdistances between these sites ranged from 3 to86 ,um with clusters at 15 to 20 and 30 to 35um. In this respect, the chromosomes of S.cerevisiae closely resemble the chromosomes of
179VOL. 38, 1974
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

HARTWELL
FIG. 9. An electron micrograph of a replicatingDNA molecule displaying a "bubble." The arrowsindicate the limits of the bubble; kindly provided byC. Newlon and W. Fangman.
animal somatic cells which also have multipleinternal sites of replication initiation spaced atintervals of 15 to 60 gAm (27, 85) in contrast tothe chromosome of E. coli which contains asingle initiation site that serves a 1,400-,umreplicon (26).
In synchronous cultures the proportion ofrDNA to total DNA remains constant through-out the S period (54). This result suggests thatthe replication of the rDNA is not restricted to asmall portion of the S period. However, thisconclusion is only valid if the length of the Speriod in the individual cell was nearly as longas the length of the S period in the synchronizedculture, an assumption which is reasonable inthis experiment, but not proven.Two known gene functions, cdc 8 and cdc 21,
are required for DNA replication (70, 73). Bothhave execution points considerably after budemergence and are probably required thereforethroughout the DNA synthetic period. Indeed,cells carrying these mutations cease ongoingDNA synthesis abruptly following a shift frompermissive to restrictive temperature (70, 73).One of these mutants, cdc 8, has been found tobe temperature-sensitive for DNA synthesis in apermeabilized cell system that incorporatesdeoxynucleotide triphosphates into DNA (79)
and, hence, this mutant is probably not defec-tive for precursor synthesis. The other mutanthas not been tested.Mutants with a diagnostic defect in medial
nuclear division arrest with the same terminalphenotype as those with diagnostic defects inDNA replication. However, the former synthe-size DNA before arrest at medial nuclear divi-sion, whereas the latter do not. Furthermore,many of the medial nuclear division mutantshave execution points around the time of DNAsynthesis. These facts suggest the possibilitythat some or all of the medial nuclear divisionmutants might be defective in proteins that arerequired for the completion of chromosomeduplication, but not for the polymerization ofdeoxyribonucleotides. The DNA ligase enzymeof E. coli is an example of such a protein (136).
Other specific inhibitors of DNA synthesis inS. cerevisiae include hydroxyurea (159), Treni-mon (88), and fluorodeoxyuridine (48). Cellsthat are arrested by these inhibitors assume thesame terminal phenotype as do cells carryingdefects in gene cdc 8 or cdc 21.
Repair and recombination of DNA moleculesmay be either restricted to the DNA syntheticperiod or induced during the S period as evi-denced by two observations. The capacity toundergo X ray-induced mitotic recombinationis periodic in synchronous yeast cultures with apeak at about the time of DNA synthesisinitiation (48), and survival to high doses of Xrays is much greater in budding yeast than inunbudded cells (7).
Mitochondrial DNA
Reviews on mitochondrial DNA and mito-chondrial genetics have appeared recently (15,103), and these areas will be only briefly andselectively reviewed here.
In addition to nuclear DNA, the yeast cellcontains mitochondrial DNA that constitutes 5to 20% of the total cellular DNA and differs indensity (1.683) and base composition (17%G+C) from nuclear DNA (10, 110). Althoughthe size and topology of the mitochondrial DNAmolecule is still a subject of controversy (seereference 15), recent gentle lysis and handlingprocedures have permitted the isolation of amajority of the mitochondrial DNA moleculesfrom sucrose gradients as linear structures witha length of about 21 lsm (56, 137). Directvisualization of lysed mitochondria revealedsome closed, circular molecules of about 25 gmin length, and it has been suggested that thisstructure represents the native molecule (83).
180 BACTrERIOL. REV.
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
The renaturation kinetics of mitochondrialDNA is reported to be compatible with aminimum genome equivalent to 25 ;m of DNA(83), but an anomalous behavior of mitochon-drial DNA in renaturation experiments suggestscaution in interpreting these results (31).
Irreversible changes in the mitochondrialDNA occur spontaneously with high frequencyresulting in respiratory deficient (petite) yeastand can be induced by many agents in close to100% of the cells in a population. The mitochon-drial DNA from petites frequently displays analtered density and base composition (10 11110, 126) and may be entirely missing in somepetites (36, 56, 127-129).The ratio of mitochondrial to nuclear DNA is
fairly constant under a variety of growth condi-tions (53, 189), and some control mechanismprobably exists to coordinate their relative ratesof synthesis. Although the replication of mito-chondrial DNA has been reported to occurwithin a brief portion of the cell cycle, immedi-ately following nuclear DNA replication in syn-chronous cultures of S. lactis (161), similarstudies with S. cerevisiae suggest a continuousreplication of mitochondria DNA throughoutthe cell cycle (191).One possible model for achieving a constant
relationship between the two DNA componentswould be for the duplication of one to bedependent upon the duplication of the other.This simple model appears to be ruled out,however, since conditions are known that per-mit the synthesis of either nuclear or mitochon-drial DNA while preventing that of the other.Thus, nuclear DNA replication occurs in theabsence of mitochondrial DNA synthesis, be-cause yeast strains lacking mitochondrial DNAare viable (56, 128). Conversely, mitochondrialDNA synthesis continues under conditions thatprevent nuclear DNA synthesis. When cytoplas-mic protein synthesis is prevented by cyclohexi-mide, the net synthesis of mitochondrial DNAcontinues in the absence of nuclear DNA syn-thesis for about one generation time, approxi-mately doubling in amount (60). Even morestriking are results obtained in the presence of afactor where net accmulation of mitochondrialDNA continued at a normal rate in the absenceof nuclear DNA synthesis for at least twogenerations undergoing a fourfold increase inamount (139). In certain mutants that aredefective in the initiation of nuclear DNAreplication, mitochondrial DNA synthesis pro-ceeds uninhibited at the restrictive temperature(38, C. S. Newlon and W. L. Fangman, personalcommunication).
Two gene products that are required fornuclear DNA replication (cdc 8 and cdc 21) arealso essential for mitochondrial DNA synthesis(C. S. Newlon and W. L. Fangman, personalcommunication). However, the three genes thatare required for the initiation of nuclear DNAsynthesis are not necessary for mitochondrialDNA synthesis (38, C. S. Newlon and W. L.Fangman, personal communication). Perhapsother, as yet unidentified, gene products arespecifically involved in initiation of mitochon-drial DNA synthesis.
In view of the dependence of the initiation ofreplicons in yeast (80, 160, 190), in animal cells(84), and in bacterial cells (100) upon proteinsynthesis, the apparent insensitivity of mito-chondrial DNA synthesis to the inhibition ofeither cytoplasmic or mitochondrial proteinsynthesis is quite interesting. In the presence of200 Mg of chloramphenicol per ml, a specificinhibitor of mitochondrial protein synthesis,yeast cells continued mitochondrial DNA syn-thesis at a normal rate for at least two genera-tions (60). Although the level of chloram-phenicol used in these experiments was suffi-cient to inhibit respiratory activity, the degreeof inhibition of mitochondrial protein synthesiswas not measured. However, a lack of depend-ence of mitochondrial DNA replication uponmitochondrial protein synthesis can also bereasoned from the fact that petites with grosslyaltered mitochondrial DNA are able to replicatethis DNA, although they must not be capable ofsynthesizing an active mitochondrial proteinsynthetic system (103).
Two-Micron Circular DNA
About 1 to 5% of the total yeast cell DNA ispresent as 2-am closed, circular DNA moleculeswith a density similar to that of the main bandnuclear DNA (2, 61, 158). The renaturationkinetics of isolated 2-Mim circles are consistentwith their size, and this result suggests that allof the molecules are identical in base sequence(4). The cellular location of this DNA fraction isunclear at present, but the isolation of 2-amcircles free of both nuclear and mitochondrialDNA in association with a membrane-contain-ing component suggests the possibility of adistinct organelle location (34). Mechanismsmust exist to coordinate the synthesis of 2-,umDNA with that of nuclear and mitochondrialDNA, but this question has not yet been ex-amined. Because of the ease with which thesemolecules can be isolated and characterized,they should be very useful in investigations of
181VOL. 38, 1974
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
the molecular details of DNA replication inyeast.
Macromolecule synthesisTotal cell mass increases continuously
throughout the cell cycle (19, 147, 168). Al-though some experiments suggested a periodicincrease in total cell RNA (184, 187), it is nowgenerally accepted that both protein and RNAaccumulate continuously throughout the cell cy-cle (37, 57, 69, 150, 183, 184). Synthesis of thetwo stable ribosomal RNAs and total transferRNA have each been examined and also foundto occur throughout the cell cycle (170). Theresolution in these experiments was not suffi-cient, however, to rule out the possibility of abrief cessation of RNA and/or protein synthesisduring mitosis and, in view of the documenta-tion of this phenomenon in animal cells (142), itwould be of interest to examine this possibilityby an autoradiographic approach.Kuenzi and Fiechter have investigated the
carbohydrate composition of the yeast cell dur-ing the cell cycle (98, 99). In cells synchronizedby glucose starvation and refeeding, the glucanand mannan contents per milligram of dryweight remained relatively constant, but theglycogen and trehalose contents decreasedabruptly at the time of budding (98). In cellssynchronized by selection from a growing popu-lation of unbudded cells by sedimentation in adensity gradient the contents of all four car-bohydrates were found to remain relativelyconstant throughout the cell cycle (99). Kuenziand Fiechter suggest that glycogen and treha-lose may provide a source of energy for thebudding cycle under the special conditions ofglucose limitation (99).The phenomenon of periodic or discontinuous
enzyme synthesis during the cell cycle has beendocumented in many organisms and is poten-tially of great significance to our understandingof the cell division cycle (reviewed in references65, 113). For a large number of enzymes thathave been examined, total activity was ob-served to double abruptly (step synthesis) at aspecific time in synchronous cultures, whereasfor other enzymes an abrupt increase followedshortly by a decrease (pulse synthesis) wasfound. The former pattern is thought to signifythe restricted synthesis of a stable enzymeduring a discrete interval of the cell cycle,whereas the latter is explained by the samehypothesis for an unstable enzyme. Periodicenzyme synthesis has been extensively studiedin S. cerevisiae by Halvorson and his colleagues(65).
Step synthesis is apparently quite general inS. cerevisiae, having been documented for 33different enzymes (65). Five different permeaseactivities also exhibit a periodic doubling (29).No example of continuous enzyme synthesisduring the cell cycle of Saccharomyces has beenreported, although such examples have beenreported in other organisms including the yeastSchizosaccharomyces pombe (113). However,the synthesis of the ribosomal proteins appearto occur continuously during the cell cycle of S.cerevisiae (155). It has been suggested thatsome examples of continuous enzyme synthesismay in reality be the summation of severalpatterns of discontinuous synthesis by each ofseveral non-allelic genes that code for proteinswith the same function or activity (168).A number of observations limit the possible
interpretations of this phenomenon. Althoughalmost all of the experiments have been donewith cells synchronized by one of two ratherharsh procedures, either by induced synchronyusing shifts from starvation to growth mediumas in the procedure of Williamson and Scopes(195) or selection synchrony utilizing sucrosegradients to select the small cells in the popula-tion (168), the hypothesis that the observationsare artifacts of the synchronization techniquesappears to be ruled out on several counts. First,periodic synthesis has been observed for manydifferent enzymes. Second, although the twosynchronization techniques are harsh, they arequite different and yet the step time of aparticular enzyme is nearly the same regardlessof the synchronization regime (168). Third, theresult is reproducible in successive cycles in thesame experiment (39, 57, 169). Finally, thephenomenon has been reproduced for threeenzymes by the technique of Sebastian et al.(150), which does not require presynchroniza-tion of the culture.Any model that invokes the activity of the
enzyme itself in the production of repressor ormetabolism of inducer to control the step inenzyme activity appears to be incompatiblewith observations for certain enzymes in yeast.Thus a-glucosidase, f3-galactosidase, and f-glucosidase display step synthesis at the sametime in the cell cycle in synchronous culturessynthesizing the enzyme at the basal rate and ata highly induced rate (30, 63). The enzymeuridine 5'-diphosphate galactose-4-epimerase is.reported to undergo step synthesis at the sametime in the cell cycle in cultures that areinduced as well as in cultures producing theenzyme constitutively (65). Finally, any one ofseveral non-allelic genes (124) endow the cell
182 HARTWELL
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
carrying the dominant allele with the capacityto produce the same enzyme, a-glucosidase(67). The patterns of a-glucosidase synthesis incells carrying more than one of these genes hasbeen interpreted to be the result of severaldistinct steps of enzyme synthesis occurring atdifferent times in the cell cycle, with each stepbeing the result of synthesis from one of thenon-allelic genes (168). However, it is not clearthat these non-allelic genes are the structuralgenes for the enzyme(s) and, in fact, at leastone, MAL6, is apparently not structural butregulatory (171).
Halvorson and his colleagues have discussedseveral models (65) to explain these phenom-ena, but favor a sequential chromosome tran-scription model. "In its most explicit form thismodel assumes that the order of genes on thechromosome determines the program for tran-scription and, thus, subsequent translation dur-ing the cell cycle. Thus, there is a linearrelationship between the time of ordered en-zyme appearance and the position of genesalong the chromosome" (65). As pointed out bythe authors, in order for this model to accountfor the observations it is also necessary that themessenger RNA of yeast be metabolically un-stable, and this appears to be the case (86, 177).In support of this model is the finding thatenzymes from closely linked genes, f3-galactosi-dase and f3-glucosidase in Saccharomyces lactis(66), and aspartokinase, phosphoribosyl-adeno-sine 5'-triphosphate-pyrophosphorylase, andthreonine deaminase on chromosome V of S.cerevisiae (169), exhibit steps at about the sametime in the cell cycle, whereas enzymes fromgenes that are unlinked exhibit steps as early as0.1 and as late as 0.8 in the cell cycle (169).Although the sequential transcription modelhas strong appeal, it is important to note thatthere is no evidence that the steps in enzymeactivity are correlated with the time of tran-scription of the relevant gene or the translationof this transcript as would be required by themodel.
Apparently strong confirmation of the se-quential transcription model was provided bythe observation that a change in the mapposition of two genes was correlated with achange in step times of the products of thesegenes (39). The genes gal, and lys2 are report-edly closely linked on the same side of thecentromere of chromosome II in the S.cerevisiae strain S288C (122), whereas they arereportedly far apart on opposite sides of thecentromere in strain 61009 (87). In synchronouscultures with strain S288C, Cox and Gilbert
(39) found that the step times for the enzymeproducts of these genes, galactokinase and a-aminoadipic acid reductase occurred at about0.5 and 0.3, respectively, in the cell cycle,whereas in strain 61009 they occurred at about0.7 and 0.2, respectively. Thus the greaterseparation of the two structural genes appearedto be correlated with a greater separation in steptimes for the synthesis of their gene products.Unfortunately, Cox and Gilbert did not attemptto confirm the different mapping results previ-ously reported from two different labs. In orderto generate the purported map differences twoinversions would be required. It is conceivablethat the reported difference is due to a mappingerror in one of the laboratories, and beforeaccepting the correlated change in time ofenzyme synthesis as support for the lineartranscription model, it is imperative that themap difference be confirmed in a single labora-tory.
If the sequential transcription model is cor-rect it appears that some genes may be excep-tions to the rule of discontinuous transcription.The synthesis of some, if not all, of the riboso-mal proteins occurs continuously during the cellcycle (155). However, one could accommodatethis observation with the sequential transcrip-tion model in one of two ways. First, eachribosomal protein may be encoded by severalnon-allelic genes with different times of expres-sion. This hypothesis seems unlikely because atleast one ribosomal protein, that conferringcycloheximide resistance to the 60S ribosomalsubunit, is encoded by a single gene, cyr2 (35).Second, the messenger RNAs for the ribosomalproteins may be exceptionally stable.The recent finding that ornithine transami-
nase in S. cerevisiae and fl-galactosidase in S.lactis are inducible at all times in the cell cycle(151) is incompatible with any model thatrestricts transcription potential within the cellcycle, but is not necessarily incompatible withordered transcription of the genome under fullyinduced or fully repressed conditions.
HOW ARE THE EVENTS OF THE CELLCYCLE COORDINATED?
Dependent Pathways of LandmarksMost of the cell cycle landmarks must occur
in the proper order to ensure the production oftwo viable daughter cells. How is this fixedorder achieved? A partial answer to this ques-tion comes from the observation that manylandmarks in the cell cycle are dependent fortheir occurrence upon the prior completion of
VOL. 38, 1974 183
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

HARTWELL
earlier landmarks. Such cases of dependencedictate a fixed sequence of events because nolate landmark can occur until all early land-marks upon which it is dependent have beencompleted.The sequences of dependent events can be
mapped with the use of specific cell cycle blocks(74). When the occurrence of one landmark isspecifically prevented by an inhibitor or tem-perature-sensitive mutation, some of the land-marks that normally follow are also prevented,whereas others are not. Those that do not occurare dependent upon the completion of theinhibited landmark, whereas those that dooccur are not dependent upon such completion.Of course, the assumption inherent in thisargument is that the specific inhibitor or muta-tion directly interferes with only one landmark.The extensive genetic and physiological charac-terization of the cdc mutants (76) lends supportto the validity of this assumption for thesetemperature-sensitive mutants, and it is a rea-sonable assumption for some inhibitors as well.The data currently available with a variety ofcell cycle inhibitors and cdc mutants was re-viewed recently (74). With this data one canconstruct the pathways of dependent eventsshown in Fig. 10. Evidence for some of therelationships shown in Fig. 10 is stronger thanfor others, and it is not unlikely that alterationsin this model will be made as more data becomeavailable. The model is presented more as aninducement to further experimentation ratherthan as the final word, and the interested readeris advised to consult Hartwell et al. (74) andByers and Goetsch (20) for the detailed argu-ments that support each dependent step.The landmarks of the cycle are apparently
ordered into two dependent pathways. Onepathway is comprised of the following land-marks: plaque duplication, plaque separation,initiation of DNA synthesis, DNA synthesis,medial nuclear division, late nuclear divisionand spindle elongation, cytokinesis, and cellseparation. The other pathway is composed ofthe following landmarks: bud emergence, nu-clear migration, cytokinesis, and cell separa-tion. Only the inner pathway of events must becompleted in one cycle in order for the cell tocommence upon a second cycle. If this model iscorrect, then the order of events in each of thetwo separate pathways is assured. Furthermore,the integration of the two pathways would beaccomplished by the facts that both pathwaysdiverge from a common landmark, plaque du-plication, and that both converge upon a com-mon landmark, cytokinesis.
PS
iDS
DS621
SE /
I- NM
FIG. 10. Dependent pathways of events in the cellcycle. A bbreviations are the same as those used in Fig.1. Small numbers refer to cdc genes that are requiredfor the particular step in question and small lettersdesignate inhibitors that block the step. MF, matingfactor; HU, hydroxyurea; TR, Trenimon.
Events on different pathways are independ-ent of one another. For example, DNA synthesisand nuclear division occur in the absence of budemergence in mutants defective in gene cdc 24at the restrictive temperature and, conversely,bud emergence and nuclear migration occur inthe absence of DNA synthesis in mutants defec-tive in cdc and 21 at the restrictive temperature(74). It would not be surprising, therefore, iflandmarks that belong to different pathwaysexhibited a reversal of order under two differentsets of conditions. Indeed, the order of theevents nuclear migration and nuclear divisiondoes appear to be variable under some condi-tions (143). In this context it is interesting tonote that the landmarks are so distributedbetween the two pathways that such an occur-rence would not result in the production ofinviable daughter cells, providing that bothpathways were eventually completed.
Dependent Pathways of Gene ExpressionThe demonstration that order is achieved in
the cell cycle by a dependence of late eventsupon early events raises another question. Whatis the basis of this dependence? At the next levelof analysis this question can be approached byinquiring whether the steps mediated by the cdcgene products are organized into dependentpathways. In theory, two-gene-mediated stepsmay be related to one another in one of fourpossible ways as shown in Fig. 11. They may berelated in one of two dependent sequences, with
184 BACTERIOL. REV.
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
step A being dependent upon the prior comple-tion of step B or vice versa. They may beindependent with either step A or step B beingcapable of completion in the absence of thecompletion of the other step, or they may beinterdependent so that neither step can occurwithout the concomitant occurrence of theother.Two rationales have been used to distinguish
between these four possible relationships (81).One is applicable to two steps for which reversi-ble conditional blocks are available that can beindependently imposed. This method involvesshifting cells from one restrictive condition tothe other and it is capable of distinguishing allfour possibilities (81, 91). A second method isapplicable to two blocks that need not bereversible and that cannot be independentlyimposed. For two-gene-mediated steps it re-quires a comparison of the phenotypes of thesingle-gene mutants with that of a mutantstrain defective in both genes. The doublemutant approach is capable of resolving three ofthe four possible relationships. For a discussionof these methods and their limitations thereader is referred to Hereford and Hartwell (81).By using this approach the sequence of gene-
mediated steps diagrammed in Fig. 12 wasdetermined. The steps are' organized into adependent sequence. Although the data are notextensive, they do suggest that the reason forthe dependent pathways of landmarks (Fig. 11)is because the expression of the gene productsthat mediate these landmarks is also organizedinto dependent sequences. For example, nucleardivision cannot take place until after the rep-lication of DNA because the gene productsthat mediate nuclear division cannot expressthemselves until after the gene products thatmediate nuclear DNA synthesis. More work ofthis type with additional inhibitors and mu-
dependent
2 dependent
3 independent
A B
B A
AB
A R4 interdependent
M-
FIG. 11. Four possible relationships between twogene-mediated steps. Two steps may be related in adependent sequence (1 or 2), in which case the firststep must occur before the second. They may beindependent (3), in which case either step can occur inthe absence of the other, or they may be interdepend-ent (4), where both steps must occur concomitantly.
SF-Ilab-2
cell cycle landmarks PD PS iDS DS mNN B SE
gee mediated steps lF.287, | 1|_7 1 2.8.21.HU | (9,13.16.20)_
FIG. 12. Dependent pathway of gene-mediatedsteps in the cell cycle. Succeeding steps are related ina dependent sequence (as in Fig. 11) of given order.Numbers refer to cdc genes and letters to inhibitors(HU, hydorxyurea; MF, mating factor) or to land-marks (as in Fig. 1). Designations over the same arroware interdependent (as in Fig. 11) unless they appearin parentheses, in which case order has not beendetermined.
tants will permit the construction of a detailedmap of gene expression and a more comprehen-sive test of this conclusion.We can now rephrase the question that
open this section as follows: what is themolecular basis of the dependence of geneexpression in the cell cycle? Although we do nothave an answer to this question, it may beuseful to consider two broad categories of pos-sibilities. The dependence might exist at thelevel of the synthesis of a gene product or at thelevel of its functional expression. For example,the fact that cdc 7 must express itself before cdc8 (Fig. 12) might be due to the fact that cdc 7product is required for the synthesis of cdc 8product (i.e., cdc 7 product is a positive regula-tor of the synthesis of cdc 8 product) or becausethe cdc 7 product is required for the function ofcdc 8 product (perhaps they are sequential stepsin a pathway so that the product produced bythe action of cdc 7 product is the substrate forcdc 8 product). A resolution between these twopossibilities is hampered by the fact that we donot know the molecular basis of the tempera-ture-sensitive lesion in either gene. If we candetermine whether the mutant gene productsare thermolabile for their synthesis or for theirfunction, then it may be possible to distinguishthese possibilities.The extensive data suggesting that many, if
not all, proteins are synthesized periodicallyduring the cell cycle (65) offers a plausible andtestable explanation for the dependent path-ways of landmarks and gene expression. Takentogether the data suggest a cascading system ofpositive regulation where the function of earlygene products turns on the synthesis of latergene products whose function in turn inducesthe synthesis of still other gene products, etc. Ifthis model is valid, then the arrest of the cellcycle with an inhibitor or with a cdc mutationmight arrest the program of sequential protein
185VOL. 38, 1974
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
synthesis or prevent the synthesis of cdc geneproducts that function later in the cell cycle.
Indeed, cells arrested in DNA synthesis byhydroxyurea do not acquire the capacity tocomplete nuclear division, cell division, orsubsequent rounds of budding in the absence offurther protein synthesis (159, 160), suggestingthat some of the proteins required for each ofthese processes cannot be synthesized untilDNA replication is completed. However, cellsthat are arrested in cell cycle progress by X-raytreatment continue to exhibit pulse synthesis ofDNA polymerase (47), and cells arrested atDNA synthesis by Trenimon continue to syn-thesize dihydrofolate reductase although thenormal step pattern of synthesis is altered (88).These results demonstrate that experimentalrationales exist for advancing an answer to thequestion, but much more data is needed beforegeneral conclusions are warranted.
HOW IS CELL DIVISIONCONTROLLED?
In the transition from mitotic cell division tothe three alternative stages of the yeast lifecycle-stationary phase, sexual conjugation,and meiosis-the mitotic cell division cycle isarrested and an alternative developmental pro-gram is undertaken (see Fig. 5). Do yeast cellsarrest the cell cycle at a specific stage beforecommencing on one of the alternatives, and if sois such arrest obligatory? A definitive answer tothese questions is not yet available for all threesituations, but all evidence points to a restric-tion of developmental potential to the G1 inter-val of the cell cycle and suggests that once a cellhas passed a critical point in G1, it is committedto completion of the mitotic cell cycle before itcan commence on one of the three alternativeprograms. The most complete data availableconcern the transition from the mitotic cell cy-cle to sexual conjugation.
Transition from Mitosis to ConjugationThe relationship between the mitotic cycle of
haploids and the sexual cycle may be relativelysimple. BUcking-Throm et al. (18) proposedthat the two haploid cells synchronize their cellcycles prior to cell fusion and zygote formation.Furthermore, they proposed that this synchron-ization is achieved in part through the agency ofthe constitutively produced a factor by cells of amating type. In this model, the a factor specifi-cally arrests a cells as unbudded, mononucleatecells prior to the initiation of DNA synthesis(i.e., in the G1 interval of the cell cycle). Somereciprocal mechanism for arresting the a cell at
the same point in the cycle was foreseen, andevidence is now at hand that an analogous afactor exists. Considerable evidence supportsthis model, but some points remain unproven.
First, let us consider the proposition that aand a cells are at the same point in the cell cycleat the time of cell fusion. Because the resultingzygote undergoes nuclear fusion followed by adiploid mitotic cycle, it is clear that at somepoint before or after fusion of the two haploidcells their mitotic cell cycles must becomesynchronized. It has been known for some timethat cell fusion can occur between unbuddedyeast spores (196) or stationary-phase yeastcells (17) without the intervention of a buddingcycle. Unbudded cells isolated from log culturesalso undergo cell fusion without prior budding(152). These results demonstrate that cells inthe unbudded portion of the cell cycle canundergo fusion, but they do not permit anyconclusion about cells in the rest of the cycle. Amore critical test of the proposition was pro-vided by a kinetic study of zygote morphologyafter the mixing of two asynchronous cultures ofhaploid cells (72). The first zygotes that wereformed were unbudded and binucleate and,hence, resulted from the fusion of two unbud-ded, mononucleate, haploid cells. This resultsuggests that cell fusion preferentially occursamong cells in the unbudded portion of the cellcycle, but does not rule out the possibility ofslower or less frequent conjugation betweenbudded cells.The most convincing evidence on this point
comes from an ultrastructural study of conjuga-tion by Byers and Goetsch (20). In a massmating induced by mixing two asynchronouspopulations they found that each of the twohaploid nuclei contained a single nuclear plaquein all mating pairs that had not yet completednuclear fusion. This result demonstrates thatthe two haploid nuclei are synchronized at thesingle-plaque stage at the time of cell fusion.This conclusion is supported by the finding ofSena et al. (153) that newly formed zygotesundergo DNA replication during the first bud-ding cycle. Because plaque duplication is aprerequisite for DNA replicaton and bud emer-gence in the cell cycle, all of the results collec-tively support the proposition that cell fusionnormally occurs at the unbudded portion of thecell cycle prior to plaque duplication and theinitiation of DNA synthesis.
Studies by Reid (personal communication)suggest that this portion of the cell cycle is notonly the preferred period for cell fusion, but thatconjugation is restricted to this interval. He
186 HARTWELL
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
arrested haploid a or a cells at various positionsin the cell cycle by shifting temperature-sensi-tive cdc mutants to the restrictive temperatureand then challenged them for mating with cellsof opposite mating type. Cells arrested at thecdc 28 block retained the ability to mate, andthis result is consistent with those reportedabove, for this mutant arrests as an unbuddedcell with a single nuclear plaque at a stage priorto the initiation of DNA synthesis (20, 73).Mutants arrested at all subsequent stages of thecell cycle exhibited drastically reduced matingpropensity, suggesting that conjugation is notpossible at other positions in the cell cycle.Mutants with a block in cytokinesis exhibited alow but significant ability to mate, and it isinteresting that these mutants continue to tra-verse the nuclear mitotic cycle at the restrictivetemperature (71). Two arguments suggestedthat the inability of cells blocked at most stagesof the cell cycle to mate was not an artifact oftheir being "sick." First, if the cells were shiftedback to the permissive temperature after incu-bation at the restrictive temperature they rap-idly regained the ability to mate (presumablydue to traverse of the cell cycle and arrival atGi). Second, synchronous cultures of a mutantwith a very late block in the cycle lost theability to mate at the restrictive temperature atthe time of bud emergence, long before reachingthe point of cell cycle arrest.Much circumstantial evidence suggests that
a factor and the recently described a factor arethe agents responsible for the synchronization ofthe haploid cell cycles prior to cell fusion, butproof is lacking. The existence of mating type-specific diffusible substances was first evi-denced by the finding of Levi (101) that a cellswhich were near but not physically touching acells underwent morphological changes. The acells elongated and enlarged, a process de-scribed by Levi as putting out "copulatoryprocesses." Cells with these processes are com-monly referred to as schmoos (104). The sub-stance responsible for this phenomenon (a fac-tor) is produced constitutively only by cells ofa mating type (a or aa), and only cells of a mat-ing type respond to it (a or aa) (45). Cells thatcarry at least one allele of both mating types(aa, aaa, aaa) neither produce nor respond to afactor (45). The a factor has been extensivelypurified, has a molecular weight of about 1,400,and contains a number of common amino acidsas well as cupric ions (46).When a factor is added to cells of a mating
type, cell division and nuclear DNA synthesisare arrested, but protein and RNA synthesis
continue (172); consequently the cells enlargeand elongate. The cell population becomessynchronously arrested as unbudded mononu-cleate cells that have not initiated DNA synthe-sis (18, 81), and each cell has a single nuclearplaque (20). Furthermore, the site of arrest hasbeen specifically mapped within the sequence ofthree gene functions that lead to the initiationof DNA synthesis, and the a factor sensitivestep is the same as that mediated by the cdc 28gene product, the first step in the sequence(Fig. 12; 81).
Recently a factor produced constitutively bycells of a mating type, a factor, has beendiscovered by the fact that it occasionallycauses shape changes in cells of a mating type(104). This substance specifically arrests a cellsas unbudded, mononucleate cells prior to theinitiation of DNA synthesis (L. Wilkinson andJ. Pringle, submitted for publication). The afactor arrested cells have a single nuclearplaque (B. Byers, personal communication)and, again, the site of arrest is at the cdc 28mediated step (L. Wilkinson and J. Pringle,submitted for publication). a factor has no effectupon cells of a mating type or upon a'/a diploidcells (L. Wilkinson and J. Pringle, submitted forpublication). Nothing is known of the chemistryof a factor. The relationship, if any, of these twofactors to substances that are reported to causemating-type-specific cell expansion (197) is un-known.The observations reported above strongly im-
plicate a and a factors in the synchronization ofthe haploid cell cycles prior to cell fusion. Thefactors have the appropriate specificity of pro-duction and response with respect to the matingtype alleles. The factors arrest cells at preciselythe point in the cell cycle at which conjugatingcells are blocked. Finally, a high proportion ofsterile mutants either fail to produce factor orrespond to factor, or both (104, 105).
Nevertheless, caution has been suggested inaccepting the proposed roles for these factors inthe mating response (152) because the additionof a factor to mass matings can actually delayzygote formation (T. R. Manney, personal com-munication; 152). This observation is perhapsnot too surprising because an excess of what issurely an impure preparation of only one of thefactors is being added to cells that alreadycontain both factors in their appropriate biolog-ical concentrations. However, under one set ofrather artificial conditions a factor actuallypromotes mating. The mating of several tem-perature-sensitive cdc mutants at the restric-tive temperature is dramatically enhanced by
187VOL. 38, 1974
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
arresting the cells with a factor, presumablybecause the factor arrests the cell at the appro-priate position in the cell cycle and prevents thecell from blocking at an inappropriate position(B. Reid, personal communication). Neverthe-less, the proposed role for these factors can onlybe accepted when sterile mutants are found thatdo not produce the appropriate factor andwhose ability to mate can be restored by addingpurified factor.
Transition from Mitosis to Stationary PhaseA unified picture of how yeast cells control
the cell division cycle at the onset of nutritionaldeprivation is beginning to emerge. Proto-trophic yeast cells upon encountering nutri-tional conditions that are unsatisfactory forgrowth appear to arrest cell division at its onsetin the G1 portion of the cell division cycle.Beam et al. (7). noted that yeast cells placed
in starvation medium containing only glucoseand phosphate buffer accumulated as unbud-ded cells. Williamson and Scopes (185, 195)exploited the same fact in devising their syn-chronization scheme and demonstrated thatstationary-phase cells that have exhausted arich medium were not only unbudded, but werearrested prior to DNA synthesis in the cellcycle.A systematic investigation of the site of cell
cycle arrest after various defined states of nutri-tional deprivation was carried out by Pringleand Maddox (personal communication). Theyshowed that if a prototrophic yeast strain isstarved of carbon and energy source includingglucose, glycerol, acetate, lactate, or ethanol thepopulation of cells arrested with 97% or more ofthe cells unbudded. The cultures were lightlysonicated before scoring for budded and unbud-ded cells to disrupt clumps of cells. Starvationfor ammonia, sulfate, phosphate, biotin, orpotassium also elicited arrest within the unbud-ded portion of the cell cycle. The only obligatenutritional requirement of the prototrophicstrain that failed to produce uniform arrest atthe unbudded stage was magnesium.The question of strain specificity in this
response was examined by testing 14 unrelatedhaploid strains and 12 unrelated diploid strainsafter stationary-phase arrest in rich medium.All strains displayed a high percentage of un-budded cells in stationary phase, the mean forthe haploid strains being 95% and the mean forthe diploid strains being 97%.The site of cell cycle arrest after nutritional
deprivation was investigated with respect to thesequence of gene-mediated steps shown in Fig.
12. Cells carrying a cdc mutation were arrestedby nutritional deprivation at the permissivetemperature and then placed back into com-plete medium at the restrictive temperature.The emergence of the cells was followed by timelapse photomicroscopy. If a cell completed theparticular cdc gene-mediated step before nutri-tional arrest that cell should be capable ofcompleting one cell cycle at the restrictivetemperature and would arrest in the second cellcycle. If the cell had not completed the step, itshould have arrested in the first cell cycle. Theinhibitors, a factor and hydroxyurea, weretested in an analogous manner. The conclusionfrom these experiments was that the cells werearrested at or prior to the step that is sensitiveto a factor in cells of a mating type and ismediated by the cdc 28 gene product.These observations must mean that the cdc
28-mediated step or some prior step in the cellcycle is uniquely sensitive to nutritional depri-vation. At some time in the growth of theculture, cells that are before the cdc 28 step areunable to complete it, whereas cells that areafter this step continue through the entire cellcycle to complete cell separation. The responseof yeast cells to nutritional deprivation byarresting cell cycle progress in G1 may be acontrol mechanism that has evolved to copewith times of nutritional inadequacy. Perhapscells in G1 can retain viability under restingconditions for longer periods than cells arrestedin other parts of the cycle, but this idea has notbeen tested experimentally.G1 arrest is not, however, a necessary conse-
quence of all types of growth limitation, becausemost ways of artificially interfering with yeastgrowth do not lead to G1 arrest. Starvation ofauxotrophs for required amino acids or purinesor pyrimidines did not in general lead to G1arrest (J. Pringle and R. Maddox, personal com-munication). Furthermore, of 2,000 tempera-ture-sensitive mutants which are blocked in apresumably random sample of diverse, albeitunknown, metabolic processes, no more than 6showed G1 arrest (76).
Transition from Mitosis to MeiosisThe landmarks of the meiotic cycle in yeast
include DNA replication (41), genetic recombi-nation, first and second nuclear divisions, andspore wall formation (117). Although recombi-nation does occur during the mitotic cell cycle,its frequency is orders of magnitude higher inthe meiotic cycle. Bud emergence does notoccur during meiosis, and the first meioticnuclear segregation differs from mitosis in that
188 HARTWELL
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
homologous chromosome pairs are segregated.The second nuclear segregation has the samegenetic consequence as mitosis, and both nu-clear divisions involve the duplication and seg-regation of spindle plaques, although the mei-otic plaque is cytologically distinct from themitotic plaque (117). The first and secondchromosome segregations occur within a singlemembrane-bound nucleus, which at the com-pletion of the second division pinches off fournuclei (117). Cytokinesis and cell wall separa-tion are quite distinct in meiosis, being accom-plished by the growth of the spore wall aroundthe nuclear membrane (117).Despite some basic differences, the molecular
events that underlie many of the landmarks inmeiosis and mitosis must be fundamentallysimilar, because the same gene products medi-ate these processes in both the meiotic andmitotic cycles. Thirteen out of twenty cdc genesthat were tested were found to be essential formeiosis and sporulation (157). These essentialgenes include mutants with diagnostic defectsin plaque duplication, plaque separation, initia-tion of DNA synthesis, DNA synthesis, medialnuclear division, and late nuclear division of themitotic cell cycle. The stage of arrest of thesemutants in the meiotic cycle has not yet beeninvestigated..Of the seven genes that did not appear to be
essential for meiosis, because diploid cells ho-mozygous for mutations in these genes wereable to sporulate at the restrictive temperature,the diagnostic landmark for one (cdc 24) is budemergence and for three (cdc 3, 10, and 11) iscytokinesis. Because sporulation does not re-quire bud emergence and because cytokinesisappears to be accomplished during meiosis byan entirely different process than during mito-sis, this result is not unexpected.With this introduction we can pose the ques-
tion of whether there is a unique point in themitotic cell cycle from which a cell enters themeiotic program. It is clear that cells can beginthe meiotic process from the G1 interval of themitotic cycle because stationary-phase cellsundergo meiosis without the intervention of amitosis (41). However, can cells which havepassed the G1 interval of the mitotic cell cyclecommence upon a meiotic cycle without com-pleting the mitotic cycle first? The answer tothis question must be no for cells that have pro-gressed to the point of chromosome segregation(perhaps late nuclear division), because the firstmeiotic nuclear division is reductional and themitotic nuclear division is not. However, it isconceivable that cells between G1 and nuclear
division in the mitotic cycle would be able tochange their program to bring about a reduc-tional division, especially in view of the factthat DNA synthesis and nuclear division are, inpart, mediated by the same gene products.Two studies have been reported in which
synchronous mitotic cells were removed fromculture and challenged to undertake meiosis bya shift of medium at various times during themitotic cell cycle. Haber and Halvorson (62)found that cells in the budded portion of themitotic cycle could sporulate with high fre-quency, whereas unbudded cells sporulated withlower frequency. Milne found that the parentcell was able to sporulate with high efficiency atall stages of the mitotic cell cycle, but that thebud produced spores only if it was relativelylarge at the time of the medium shift (111).Both studies suggest that cells can undertakemeiosis from most stages of the mitotic cycle,and the different behavior of the unbudded cellsin the two experiments can probably be at-tributed to differences in strain or presporula-tion culture conditions, or both.However, in the context of the present discus-
sion, the important question is not answeredmerely by the observation that cells from moststages of the mitotic cycle eventually sporu-lated, but we must ask whether or not theycompleted the mitotic cycle before undergoingmeiosis. Milne (111) showed that some of theunbudded cells at the time of the medium shiftproceeded to form small buds, and the propor-tion of such cells was that expected from thosecells which were within 10 min of budding in themitotic cycle. All of the cells that acquired orhad small or large buds at the time of themedium shift proceeded to complete mitosisand cytokinesis despite the fact that the buddid not grow appreciably in the meiotic me-dium. All of the unbudded cells did not appearto pass through a mitotic nuclear division.These results suggest that under the conditionsof this experiment those cells which were within10 min of the bud emergence event completedthe mitotic cycle before undertaking the meioticcycle, whereas those unbudded cells which werebefore this point in the mitotic cycle proceededdirectly to meiosis. Although confirmation ofthis observation with other strains and cultureconditions is highly desirable, the results dosuggest that the transition from mitosis tomeiosis is controlled within the G1 interval ofthe cell cycle. Perhaps further experiments withcdc mutants will permit a more rigorous test ofthis idea and a greater resolution of the point ofcontrol.
VOL. 38, 1974 189
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

HARTWELL
HOW IS GROWTH INTEGRATED WITHDIVISION?
Although some heterogeneity in the size ofcells that are at the same stage of the cell cycledoes exist (92), on the average all of the cell'smacromolecular components are precisely dou-bled once each cycle under conditions of bal-anced growth. Very little hard information isavailable to suggest how this coordinationmight be achieved, and even the qualitativedata that is at hand relates only to the grossestof parameters such as size or mass. Neverthe-less, a few observations suggest the outline of a
model for the integration of growth and divisionthat may at least serve to stimulate furtherexperimentation.Growth and division might be coordinated in
one of two fundamentally different ways. First,it might be that one or more successive incre-ment of growth require the completion of one or
more successive cell cycle landmark. In thismodel cell growth beyond a certain size couldnot occur until a specific landmark had beenpassed. This mechanism could serve to main-tain cell size only if progress past the landmarkswere rate limiting and not growth. Second, itmight be that the passage of one or more
landmarks requires growth beyond some spe-cific size or sizes. This mechanism could serveto maintain cell size only if growth were ratelimiting and not the passage of landmarks. Ofcourse, a combination of the two models is alsopossible. Let us consider a few observations on
the growth of yeast cells with respect to celldivision in the context of these possibilities.
It does not appear that growth is dependentupon the completion of any particular landmarkin the cell cycle. Although careful quantitationis lacking, it is quite evident from visual inspec-tion that cells arrested at all of the landmarks inthe cell cycle identified by the cdc mutantscontinue to grow in size after the time of arrestand that the arrested cells attain sizes many-fold in excess of the normal size range afterseveral cycle times at the restrictive tempera-ture (42). The increase in size is accompaniedby accumulation of macromolecules becausethe cdc mutants continue to synthesize stableRNA species at an uninhibited rate for at leastone-or two cycles after arrest (42, 70, 71, 73).Furthermore, yeast cells arrested prior toplaque duplication by the a or a mating factors(172; L. Wilkinson and J. Pringle, submitted forpublication), at DNA synthesis by hydroxyurea(159) or Trenimon (88), and at nuclear divisionby X-ray treatment (82, 164) continue to in-crease dry mass and accumulate protein and
RNA, eventually becoming several times largerthan untreated cells. Although quantitation ofmacromolecule synthesis and size increase inthe cdc mutants is essential before firm conclu-sions can be drawn, it appears that growth is notdependent upon the completion of plaque du-plication, plaque separation, bud emergence,initiation of DNA synthesis, DNA replication,medial or late nuclear division, spindle elonga-tion, cytokinesis, or cell separation.
Conversely, most of the cell cycle events canbe completed without extensive growth. Al-though stationary-phase yeast cells are uni-formly arrested in the G1 interval of the cellcycle, they are very heterogeneous in size. Somecells are of normal size, some are extremelysmall (193), and the average cell contains lessmass, protein, and RNA than the averagegrowing cell (141). Most of the decrease inaverage size occurs during the last cell cycle ortwo during the approach to stationary phase(141). We may conclude that as the mediumbecomes deficient for growth, the cells are ableto complete the last division or two without thenormal amount of growth. This same phenome-non has been documented for cells that ceasegrowth due to ammonia, sulfate, or phosphatelimitation (J. Pringle and R. Maddox, personalcommunication). The origin of the small cellsunder conditions of nutrient limitation was welldocumented for one particular situation (111).Synchronous cultures of cells growing in glu-cose-containing, rich medium were shifted tomedium containing only acetate. Most unbud-ded cells did not bud after the shift. Buddedcells, even those with the smallest buds, com-pleted both nuclear division and cytokinesis.However, the buds did not grow perceptibly insize so that cells with small buds produced onenormal-size cell (the parent cell) and one verytiny cell (the bud), 'and it is not unlikely thatthe size heterogeneity in most stationary-phasepopulations arises in the same way. Indeed thishypothesis could explain the bimodal size dis-tribution of cells observed in stationary cultures(194). This result suggests that all stages of thecell cycle after bud emergence can be completedin the absence of the normal amount of growth.Furthermore, if we combine the observationthat the last cycle upon approach to stationaryphase is not accompanied by much growth withthe observation that stationary-phase cells arearrested at or prior to the cdc 28-mediated step,it is reasonable to conclude that all steps afterthe cdc 28 step can be completed in the absenceof a normal amount of growth.There is evidence that growth is required for
some early event in the cell cycle. Williamson
190 BACTERIOL. REV.
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
Presence of nutrientsCarbon and energy sourceNitrogenSulfurPhosphorousPotassiumBiotin
Growth to a critical sizeCompletion of IND in previous cycleAbsence of opposite mating type factor.Absence of conditions that promote meiosis
Iis START 0
Integrate -. Execute cdc 28 PD
FIG. 13. Requirements for "start" in the cell cycle.
and Scopes (192) noted that the small cells instationary phase populations took longer to budin fresh medium than the larger cells. A quan-titative study of this phenomenon by time lapsephotomicroscopy (J. Pringle, personal commu-nication) demonstrated that an inverse rela-tionship exists between the size of a cell and thelength of time before the onset of budding infresh medium. Because the small cells growduring the interval before budding this observa-tion can best be explained by the hypothesisthat some early event in the cycle prior to budemergence requires the attainment of a certaincell size for completion. The deficit of thesmall cells was made up by the time of budding,for the interval between first and second bud-ding cycles was found to be the same for theoriginally small and originally large cells.Another observation that is consistent with a
unique requirement for growth at some earlystep in the cell cycle is provided by studies ofvon Meyenburg (181) and Beck and von Meyen-burg (8). They found that when the growth rateand doubling time of S. cerevisiae cultures werevaried by limiting the supply of glucose in achemostat, almost all of the increased cell cycletime at slower growth rates was accounted forby an increase in the length of time spent in theunbudded portion of the cell cycle, whereas thelength of time spent in the budded portion wasalmost unaffected by the growth rate.
THE CONCEPT OF "START"The results reviewed in the previous sections
suggest that the control of cell division may beachieved in the G1 portion of the yeast cellcycle, in particular at those steps that precedeand include the step mediated by the cdc 28gene product. Haploid cells appear to beable to mate only at this point in the cycle, andthe same restriction may hold for diploid cellsembarking upon the meiotic program. Cellsthat are nutritionally deprived of carbon andenergy, ammonia, sulfate, phosphate, potas-
sium, or biotin arrest here, and there appears tobe a unique requirement for growth in this earlyportion of the cycle.We have coined the term "start" for those
events in early G1 that culminate in the execu-tion of the cdc 28-mediated step (75). Beforethis step, we view the yeast cell as undifferen-tiated in the sense that it is capable of under-taking several alternative developmental pro-grams, but after this step it appears to becommitted to the mitotic cell cycle. We reasonthat many inputs are integrated by the cell atthis unique point of control (Fig. 13) and thatonly if all are sufficient does the cell commititself to a new cell cycle.This concept obviously requires further ex-
perimental testing, but it does provide an ap-pealing unity to some of the diverse questions ofregulation posed at the outset of this review. Ifthese speculations are substantiated we will beable to rephrase these questions in the followingways: how do the mating factors, the pathwaysof carbon, nitrogen, sulfur, phosphorous, potas-sium, and biotin assimilation, and the processesof macromolecule synthesis communicate withthe cdc 28 gene product, and how are all of thesediverse inputs integrated at the "start" event?Recalling that the regulation of cellular prolifer-ation (28), the integration of growth and divi-sion (97), and the control of cellular differentia-tion (180) occur within the G1 interval of themammalian cell cycle, it would not be -unrea-sonable to expect that answers to these ques-tions would have import beyond the boundariesof mycology.
ACKNOWLEDGMENTSMy work with yeast has been encouraged by the
enthusiasm of my colleague, Calvin McLaughlin, andmy interest in cell cycle is the consequence of stimu-lating collaboration with Joe Culotti, Lynna Hereford,Charles Milne, John Pringle, Brian Reid, Dick Shul-man, and Linda Wilkinson. I would like to thankBreck Byers, Walt Fangman, Jim Haber, HarlynHalvorson, and Martin Slater for reading and criticiz-
VOL. 38, 1974 191
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
ing this manuscript.Unpublished work from my laboratory was sup-
ported by Public Health Service grant GM-17709from the National Institute of General MedicalSciences.
LITERATURE CITED1. Agar, H. D., and H. C. Douglas. 1955. Studies of
budding and cell wall structure of yeast. J.Bacteriol. 70:427-434.
2. Avers, C. J., F. E. Billheimer, H. P. Hoffmann,and R. M. Pauli. 1968. Circularity of yeastmitochondrial DNA. Proc. Nat. Acad. Sci.U.S.A. 61:90-97.
3. Bacon, J. S. D., E. Davidson, D. Jones, and I. F.Taylor. 1966. The location of chitin in theyeast cell wall. Biochem. J. 101:36C-38C.
4. Bak, A. L., C. Christiansen, and G. Christian-sen. 1972. Circular, repetitive DNA in yeast.Biochem. Biophys. Acta 269:527-530.
5. Bartholomew, J. W., and T. Mittwer. 1953.Demonstration of yeast bud scars with theelectron microscope. J. Bacteriol. 65:272-275.
6. Barton, A. A. 1950. Some aspects of cell divisionin S. cerevisiae. J. Gen. Microbiol. 4:84-86.
7. Beam, C. A., R. K. Mortimer, R. G. Wolfe, andC. A. Tobias. 1954. The relation of radioresist-ance to budding in Saccharomyces cerevisiae.Arch. Biochem. Biophys. 49:110-122.
8. Beck, C., and H. K. von Meyenburg. 1968.Enzyme pattern and aerobic growth of Sac-charomyces cerevisiae under various degreesof glucose limitation. J. Bacteriol. 96:479-486.
9. Beran, K. 1968. Budding of yeast cells, theirscars and ageing. Advan. Microb. Physiol.2:143-171.
10. Bernardi, G., F. Carnevali, A. Nicolaieff, G.Piperno, and G. Tecce. 1968. Separation andcharacterization of a satellite DNA from ayeast cytoplasmic "petite" mutant. J. Mol.Biol. 37:493-505.
11. Bernardi, G., M. Faures, G. Piperno, and P. P.Slonimski. 1970. Mitochondrial DNA's fromrespiratory-sufficient and cytoplasmic respira-tory-deficient mutant yeast. J. Mol. Biol.48:23-42.
12. Bhargava, M. M., and H. 0. Halvorson. 1971.Isolation of nuclei from yeast. J. Cell Biol.49:423-429.
13. Bicknell, J. N., and H. C. Douglas. 1970. Nucleicacid homologies among species ofSaccharomyces. J. Bacteriol. 101:505-512.
14. Blamire, J., D. R. Cryer, D. B. Finkelstein, andJ. Marmur. 1972. Sedimentation properties ofyeast nuclear and mitochondrial DNA. J. Mol.Biol. 67:11-24.
15. Borst, P. 1972. Mitochondrial nucleic acids.Annu. Rev. Biochem. 41:333-376.
16. Brendel, M., and R. H. Haynes. 1972. Kineticsand genetic control of the incorporation ofthymidine monophosphate in yeast DNA.Mol. Gen. Genet. 117:39-44.
17. Brock, T. D. 1961. Physiology of the conjugationprocess in the yeast, Hansenula wingei. J.
Gen. Microbiol. 26:487-497.18. Bucking-Throm, E., W. Duntze, L. H. Hartwell,
and T. R. Manney. 1973. Reversible arrest ofhaploid yeast cells at the initiation of DNAsynthesis by a diffusible sex factor. Exp. CellRes. 76:99-110.
19. Burns, V. W. 1956. Temporal studies of celldivision. I. The influence of ploidy and tem-perature on cell division in-S. cerevisiae. J.Cell Comp. Physiol. 47:357-375.
20. Byers, B., and L. Goetsch. 1973. Duplication ofspindle plaques and integration of the yeastcell cycle. Cold Spring Harbor Symp. Quant.Biol. 30 (in press).
21. Cabib, E., and B. Bowers. 1971. Chitin and yeastbudding. Localization of chitin in yeast budscars. J. Biol. Chem. 246:152-159.
22. Cabib, E., and V. Farkas. 1971. The control ofmorphogenesis: an enzymatic mechanism forthe initiation of septum formation in yeast.Proc. Nat. Acad. Sci. U.S.A. 68:2052-2056.
23. Cabib, E., and F. A. Keller. 1971. Chitin andyeast budding. Allosteric inhibition of chitinsynthetase by a heat-stable protein fromyeast. J. Biol. Chem. 246:167-173.
24. Cabib, E., and R. Ulane. 1973. Yeast chitinsynthetase. Separation of the zymogen fromits activating factor and recovery of the latterin the vacuole fraction. J. Biol. Chem.248: 1451-1458.
25. Cabib, E., R. Ulane, and B. Bowers. 1974. Amolecular model for morphogenesis: the pri-mary septum of yeast. In B. L. Horecker andE. R. Stadtman. Curr. Top. Cell. Reg. (inpress).
26. Cairns, J. 1963. The bacterial chromosome andits manner of replication as seen by autoradi-ography. J. Mol. Biol. 6:208-213.
27. Callan, H. G. 1972. Replication of DNA in thechromosomes of eukaryotes. Proc. Roy. Soc.Ser. B 181:19-41.
28. Cameron, I. L. 1971. Cell proliferation andrenewal in the mammalian body, p. 45-85. InI. L. Cameron and J. D. Thrasher (ed.),Cellular and molecular renewal in the mam-malian body. Academic Press Inc., New York.
29. Carter, B. L. A., and H. 0. Halvorson. 1973.Periodic changes in rate of amino acid uptakeduring the yeast cell cycle. J. Cell Biol.58:401-409.
30. Carter, B. L. A., and H. 0. Halvorson. 1973. Anevaluation of the oscillatory repression modelof periodic enzyme synthesis in yeast. Exp.Cell Res. 76:152-158.
31. Christiansen, C., A. Leth Bak, A. Stenderup,and G. Christiansen. 1971. Repetitive DNA inyeasts. Nature N. Biol. 231:176-177.
32. Chung, K. L., R. Z. Hawirko, and P. K. Isaac.1965. Cell wall replication in Saccharomycescerevisiae. Can. J. Microbiol. 11:953-957.
33. Ciferri, O., S. Sora, and 0. Tiboni. 1969. Effectof gene dosage on tryptophan synthetase ac-tivity in Saccharomyces cerevisiae. Genetics61:567-576.
192 HARTWELL
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
34. Clark-Walker, G. D. 1972. Isolation of circularDNA from a mitochondrial fraction fromyeast. Proc. Nat. Acad. Sci. U.S.A.69:388-392.
35. Cooper, D., D. V. Banthorpe, and D. Wilkie.1967. Modified ribosomes conferring resist-ance to cycloheximide in mutants of Sac-charomyces cerevisiae. J. Mol. Biol.26:347-350.
36. Corneo, G., C. Moore, D. R. Sanadi, L. I.Grossman, and J. Marmur. 1966. Mitochon-drial DNA in yeast and some mammalianspecies. Science 151:687-689.
37. Cottrell, S. F., and C. J. Avers. 1970. Evidence ofmitochondrial synchrony in synchronous cellcultures of yeast. Biochem. Biophys. Res.Commun. 38:973-980.
38. Cottrell, S., M. Rabinowitz, and G. S. Getz.1973. Mitochondrial deoxyribonucleic acidsynthesis in a temperature-sensitive mutant ofdeoxyribonucleic acid replication of Sac-charomyces cerevisiae. Biochemistry 12:4374-4378.
39. Cox, C. G., and J. B. Gilbert. 1970. Nonidenticaltimes of gene expression in two strains ofSaccharomyces cerevisae with mapping dif-ferences. Biochem. Biophys. Res. Commun.38:750-757.
40. Cramer, J. H., M. M. Bhargava, and H. 0.Halvorson. 1972. Isolation and characteriza-tion of -y DNA of Saccharomyces cerevisiae. J.Mol. Biol. 71:11-20.
41. Croes, A. F. 1966. Duplication of DNA duringmeiosis in Baker's yeast. Exp. Cell Res.41:452-454.
42. Culotti, J., and L. H. Hartwell. 1971. Geneticcontrol of the cell division cycle in yeast. III.Seven genes controlling nuclear division. Exp.Cell Res. 67:389-401.
43. de Kloet, S. R. 1970. The formation of ribonu-cleic acid in yeast: hybridization of highmolecular weight RNA species to yeast DNA.Arch. Biochem. Biophys. 136:402-412.
44. de Kloet, S. R. 1973. Distribution of ribosomalribonucleic acid cistrons among yeast chromo-somes. J. Bacteriol. 114:1034-1039.
45. Duntze, W., V. MacKay, and T. R. Manney.1970. Saccharomyces cerevisiae: a diffusiblesex factor. Science 168:1472-1473.
46. Duntze, W., D. Stotzler, E. Bucking-Throm, andS. Kalbitzer. 1973. Purification and partialcharacterization of a-factor, a mating typespecific inhibitor of cell reproduction fromSaccharomyces cerevisiae. Eur. J. Biochem.35:357-365.
47. Eckstein, H., V. Paduch, and H. Hilz. 1967.Synchronized yeast cells. HI. DNA synthesisand DNA polymerase after inhibition of celldivision by X rays. Eur. J. Biochem. 3:224-231.
48. Esposito, R. 1968. Genetic recombination insynchronized cultures of Saccharomycescerevisiae. Genetics 59:191-210.
49. Finkelstein, D. B., J. Blamire, and J. Marmur.
1972. Location of ribosomal RNA cistrons inyeast. Nature N. Biol. 240:279-281.
50. Fischer, P., B. Weingartner, and U. Winters-berger. 1973. Visualization of chromosome-like structures in protoplasts of the yeast. Exp.Cell Res. 79:452-456.
51. Freifelder, D. 1960. Bud position in Sac-charomyces cerevisiae. J. Bacteriol.80:567-568.
52. Fukuhara, H. 1967. Informational role of mito-chondrial DNA studied by hybridization withdifferent classes of RNA in yeast. Proc. Nat.Acad. Sci. U.S.A. 58:1065-1072.
53. Fukuhara, H. 1969. Relative proportions of mito-chondrial and nuclear DNA in yeast undervarious conditions of growth. Eur. J. Biochem.11:135-139.
54. Gimmler, G. M., and E. Schweizer. 1972. rDNAreplication in a synchronized culture of Sac-charomyces cerevisiae. Biochem. Biophys.Res. Commun. 46:143-149.
55. Goldberg, S., T. 0yen, J. M. Idriss, and H. 0.Halvorson. 1972. Use of disomic strains tostudy the arrangement of ribosomal cistrons inSaccharomyces cerevisiae. Mol. Gen. Genet.116:139-157.
56. Goldring, E. S., L. I. Grossman, D. Krupnick, D.R. Cryer, and J. Marmur. 1970. The petitemutation in yeast. Loss of mitochondrial deox-yribonucleic acid during induction of petiteswith ethidium bromide. J. Mol. Biol.52:323-335.
57. Gorman, J., P. Tauro, M. LaBerge, and H.Halvorson. 1964. Timing of enzyme synthesisduring synchronous division in yeast. Bio-chem. Biophys. Res. Commun. 15:43-49.
58. Gray, R. H., J. B. Peterson, and H. Ris. 1973.The organization of yeast nucleohistone fibers.J. Cell Biol. 58:244-247.
59. Grivell, A. R., and J. F. Jackson. 1968. Thymi-dine kinase: evidence for its absence fromNeurospora crassa and some other microor-ganisms, and the relevance of this to thespecific labellin'g of deoxyribonucleic acid. J.Gen. Microbiol. 54:307-317.
60. Grossman, L. I., E. S. Goldring, and J. Marmur.1969. Preferential synthesis of yeast mitochon-drial DNA in the absence of protein synthesis.J. Mol. Biol. 46:367-376.
61. Guerineau, M., C. Granchamp, C. Paoletti, andP. Slonimski. 1971. Characterization of a newclass of circular DNA molecules in yeast.Biochem. Biophys. Res. Commun.42:550-557.
62. Haber, J. E., and H. 0. Halvorson. 1972. Cellcycle dependency of sporulation in Sac-charomyces cerevisiae. J. Bacteriol.109:1027-1033.
63. Halvorson, H. O., R. M. Bock, P. Tauro, R.Epstein, and M. LaBerge. 1966. Periodic en-zyme synthesis in synchronous cultures ofyeast, p. 102-116. In I. L. Cameron and G. M.Padilla (ed.), Cell synchrony. Academic PressInc., New York.
VoL. 38, 1974 193
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
64. Halvorson, H. O., B. L. A. Carter, and P. Tauro.1971. Use of synchronous cultures of yeast to
study gene position, p. 462-470. In L. Gross-man and K. Moldave (ed.), Methods in en-
zymology, vol. XXI. Academic Press Inc.,New York.
65. Halvorson, H. O., B. L. A. Carter, and P. Tauro.1971. Synthesis of enzymes during the cellcycle. Advan. Microbial Physiol. 6:47-106.
66. Halvorson, H., J. Gorman, P. Tauro, R. Epstein,and M. LaBerge. 1964. Control of enzyme
synthesis in synchronous cultures of yeast.Fed. Proc. 23:1002-1008.
67. Halvorson, H. O., S. Winderman, and J. Gor-man. 1963. Comparison of the a-glucosidasesof Saccharomyces produced in response to fivenon-allelic maltose genes. Biochim. Biophys.Acta 67:42-53.
68. Hartwell, L. H. 1967. Macromolecule synthesisin temperature-sensitive mutants of yeast. J.Bacteriol. 93:1662-1670.
69. Hartwell, L. H. 1970. Periodic density fluctua-tion during the yeast cell cycle and the selec-tion of synchronous cultures. J. Bacteriol.104:1280-1285.
70. Hartwell, L. H. 1971. Genetic control of the celldivision cycle in yeast. II. Genes controllingDNA replication and its initiation. J. Mol.Biol. 59:183-194.
71. Hartwell, L. H. 1971. Genetic control of the celldivision cycle in yeast. IV. Genes controllingbud emergence and cytokinesis. Exp. Cell Res.69:265-276.
72. Hartwell, L. H. 1973. Synchronization of haploidyeast cell cycles, a prelude to conjugation.Exp. Cell Res. 76:111-117.
73. Hartwell, L. H. 1973. Three additional genes
required for DNA synthesis in Saccharomycescerevisiae. J. Bacteriol. 115:966-974.
74. Hartwell, L. H., J. Culotti, J. R. Pringle, and B.J. Reid. 1974. Genetic control of the celldivision cycle in yeast: a model. Science183:46-51.
75. Hartwell, L. H., J. Culotti, and B. Reid. 1970.Genetic control of the cell division cycle inyeast. I. Detection of mutants. Proc. Nat.Acad. Sci. U.S.A. 66:352-359.
76. Hartwell, L. H., R. K. Mortimer, J. Culotti, andM. Culotti. 1973. Genetic control of the celldivision cycle in yeast. V. Genetic analysis ofcdc mutants. Genetics 74:267-286.
77. Hasilik, A., and H. Holzer. 1973. Participation ofthe tryptophan synthetase inactivating sys-
tem from yeast in the activation of chitinsynthetase. Biochem. Biophys. Res. Commun.53:552-559.
78. Hayashibe, M., and S. Katohda. 1973. Initiationof budding and chitinring. J. Gen. Appl.Microbiol. 19:23-39.
79. Hereford, L. M., and L. H. Hartwell. 1971.
Defective DNA synthesis in permeabilizedyeast mutants. Nature N. Biol. 234:171-172.
80. Hereford, L. M., and L. H. Hartwell. 1973. Role
of protein synthesis in the replication of yeastDNA. Nature N. Biol. 244:129-131.
81. Hereford, L. M., and L. H. Hartwell. 1974.Sequential gene function in the initiation of S.cerevisiae DNA synthesis. J. Mol. Biol. 84:.445-461.
82. Hilz, H., and H. Eckstein. 1964. Teilungssyn-chronisierte hefezellen. I. UnterschiedlicheWirkungen von Rbntgenstrahlen und Cyto-statischen Verbindungen auf Stoffwechsel undZellteilung. Biochem. Z. 340:351-382.
83. Hollenberg, C. P., P. Borst, and E. F. J. VanBruggen. 1970. Mitochondrial DNA. V. A 25 A
closed circular duplex DNA molecule in wild-type yeast mitochondria. Structure and ge-netic complexity. Biochem. Biophys. Acta209:1-15.
84. Hori, T., and K. G. Lark. 1973. Effect of pu-romycin on DNA replication in Chinese ham-ster cells. J. Mol. Biol. 77:391-404.
85. Huberman, J. A., and A. D. Riggs. 1968. On themechanism ofDNA replication in mammalianchromosomes. J. Mol. Biol. 32:327-341.
86. Hutchison, H. T., L. H. Hartwell, and C. S.McLaughlin. 1969. A temperature-sensitiveyeast mutant defective in RNA production. J.Bacteriol. 99:807-814.
87. Hwang, Y. L., G. Lindegren, and C. C. Linde-gren. 1966. Genetic study of lysine biosyn-thesis in yeast. Can. J. Genet. Cytol. 8:471-480.
88. Jaenicke, L., K. Scholz, and M. Donike. 1970.Synthese der Dihydrofolat-Reduktase in Syn-chronisierter Hefe. Eur. J. Biochem.13:137-141.
89. Jannsen, S., E. R. Lochmann, and R. Megnet.1970. Specific incorporation of exogenous thy-midine monophosphate into DNA in Sac-charomyces cerevisiae. FEBS Lett. 8:113-115.
90. Jannsen, S., I. Witte, and R. Megnet. 1973.Mutants for the specific labelling of DNA inSaccharomyces cerevisiae. Biochem. Biophys.Acta 299:681-685.
91. Jarvick, J., and D. Botstein. 1973. A geneticmethod for determining the order of events ina biological pathway. Proc. Nat. Acad. Sci.U.S.A. 70:2046-2050.
92. Johnson, B. F. 1965. Morphometric analysis ofyeast cells. Exp. Cell Res. 39:577-583.
93. Johnson, Byron F. 1968. Lysis of yeast cell wallsinduced by 2-deoxyglucose at their sites ofglucan synthesis. J. Bacteriol. 95:1169-1172.
94. Johnson, B. F., and E. Jean Gibson. 1966.Autoradiographic analysis of regional cell wallgrowth of yeasts. III. Saccharomycescerevisiae. Exp. Cell Res. 41:580-591.
95. Kaback, D. B., M. M. Bhargava, and H. 0.Halvorson. 1973. Location and arrangement ofgenes coding for ribosomal RNA in Sac-charomyces cerevisiae. J. Mol. Biol.79:735-739.
96. Keller, F. A., and E. Cabib. 1971. Chitin andyeast budding. Properties of chitin synthetase
HARTWELL194
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
from Saccharomyces carlsbergensis. J. Biol.Chem. 246:160-166.
97. Killander, D., and A. Zetterberg. 1965. Quan-titative cytochemical studies on interphasegrowth. I. Determination of DNA, RNA, andmass content of age determined mouse fibro-blasts in vitro and of intercellular variation ingeneration time. Exp. Cell Res. 38:272-284.
98. Kiienzi, M. T., and A. Fiechter. 1969. Changesin carbohydrate composition and trehalase-activity during the budding cycle of Sac-charomyces cerevisiae. Arch. Mikrobiol.64:396-407.
99. Kiienzi, M. T., and A. Fiechter. 1972. Regulationof carbohydrate composition of Sac-charomyces cerevisiae under growth limita-tion. Arch. Mikrobiol. 84:254-265.
100. Lark, K. G. 1969. Initiation and control of DNAsynthesis. Annu. Rev. Biochem. 38:569-604.
101. Levi, J. D. 1956. Mating reaction in yeast.Nature (London) 177:753-754.
102. LieblovA, J., K. Beran, and E. StreibloA. 1964.Fractionation of a population of Sac-charomyces cerevisiae yeasts by centrifuga-tion in a dextran gradient. Folia Microbiol.Prague 9:205-213.
103. Linnane, A. W., J. M. Haslam, H. B. Lukins,and P. Nagley. 1972. The biogenesis of mito-chondria in microorganisms. Annu. Rev. Mi-crobiol. 26:163-198.
104. MacKay, V., and T. R. Manney. 1974. Muta-tions affecting sexual conjugation and relatedprocesses in Saccharomyces cerevisiae. I. Iso-lation and phenotypic characterization of non-mating mutants. Genetics 76:255-271.
105. MacKay, V., and T. R. Manney. 1974. Muta-tions affecting sexual conjugation and relatedprocesses in Saccharomyces cerevisiae. II. Ge-netic analysis of nonmating mutants. Genetics76:273-288.
106. MacKinnon, J. E. 1940. Dissociation in Candidaalbicans. J. Infect. Dis. 66:59-77.
107. Marchant, R., and D. G. Smith. 1968. Budformation in Saccharomyces cerevisiae and acomparison with the mechanism of cell divi-sion in other yeasts. J. Gen. Microbiol.53:163-169.
108. Matile, Ph., H. Moor, and C. F. Robinow. 1969.Yeast cytology, p. 219-302. In A. H. Rose andJ. S. Harrison (ed.), The yeast, vol. I. Aca-demic Press Inc., New York.
109. McClary, D. O., and W. D. Bowers, Jr. 1965. Theintegrity of the cell wall during bud formationin yeasts. Can. J. Microbiol. 11:447-452.
110. Mehrotra, B. D., and H. R. Mahler. 1968.Characterization of some unusual DNAs fromthe mitochondria from certain "petite" strainsof Saccharomyces cerevisiae. Arch. Biochem.Biophys. 128:685-703.
111. Milne, C. P. 1972. Investigation of the relation-ship between mitosis and meiosis in Sac-charomyces cerevisiae. Masters thesis, Uni-versity of Washington, Seattle.
112. Mitchison, J. M. 1958. The growth of single cells.II. Saccharomyces cerevisiae. Exp. Cell Res.15:214-221.
113. Mitchison, J. M. 1969. Enzyme synthesis insynchronous culture. Science 165:657-663.
114. Mitchison, J. M. 1971. The biology of the cellcycle. Cambridge University Press, London.
115. Mitchison, J. M. 1973. Differentiation in the cellcycle, p. 1-11. In M. Balls and F. S. Billett(ed.), The cell cycle in development anddifferentiation. Cambridge University Press,London.
116. Mitchison, J. M., and W. S. Vincent. 1965.Preparation of synchronous cell cultures bysedimentation. Nature (London) 205:987-989.
117. Moens, P. B., and E. Rapport. 1971. Spindles,spindle plaques and meiosis in the yeastSaccharomyces cerevisiae (Hansen). J. CellBiol. 50:344-361.
118. Moor, H. 1966. Ultrastrukturen im Zellkern derBackerhefer. J. Cell Biol. 29:153-155.
119. Moor, H. 1967. Endoplasmic reticulum as theinitiator of bud formation in yeast (S.cerevisiae). Arch. Mikrobiol. 57:135-146.
120. Moor, H. 1967. Der Feinbau der Mikrotubuli inHefe nach Gefrieratzung. Protoplasma64:89-103.
121. Moor, H., and K. Miuhlethaler. 1963. Fine struc-ture in frozen-etched yeast cells. J. Cell Biol.17:609-628.
122. Mortimer, R. K., and D. C. Hawthorne. 1966.Genetic mapping in Saccharomyces. Genetics53:165-173.
123. Mortimer, R. K., and D. C. Hawthorne. 1969.Yeast genetics, p. 385-460. In A. H. Rose andJ. S. Harrison (ed.), The yeast, vol. I. Aca-demic Press Inc., New York.
124. Mortimer, R. K., and D. C. Hawthorne. 1973.Genetic mapping in Saccharomyces. IV. Map-ping of temperature-sensitive genes and use ofdisomic strains in localizing genes. Genetics74:33-54.
125. Mortimer, R. K., and J. R. Johnston. 1959. Lifespan of individual yeast cells. Nature (Lon-don) 183:1751-1752.
126. Mounolou, J. C., H. Jakob, and P. P. Slonimski.1966. Mitochondrial DNA from yeast 'petite"mutants: specific changes of buoyant densitycorresponding to different cytoplasmic muta-tions. Biochem. Biophys. Res. Commun.24:218-224.
127. Moustacchi, E., and D. H. Williamson. 1966.Physiological variations in satellite compo-nents of yeast DNA detected by density gradi-ent centrifugation. Biochem. Biophys. Res.Commun. 23:56-61.
128. Nagley, P., and A. W. Linnane. 1970. Mitochon-drial DNA deficient petite mutants of yeast.Biochem. Biophys. Res. Commun.39:989-996.
129. Nagley, P., and A. W. Linnane. 1972. Biogenesisof mitochondria. XXI. Studies on the natureof the mitochondrial genome in yeast: the
VOL. 38, 1974 195
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

BACTERIOL. REV.
degenerative effects of ethidium bromide onmitochondrial genetic information in a re-spiratory competent strain. J. Mol. Biol.66:181-193.
130. Newlon, C. S., T. D. Petes, L. M. Hereford,and W. L. Fangman. 1974. Replication ofyeast chromosomal DNA. Nature (London)247:32-35.
131. Nickerson, W. J. 1963. Molecular basis of form inyeasts. Bacteriol. Rev. 27:305-324.
132. Nickerson, W. J., and G. Falcone. 1956. Identifi-cation of protein disulfide reductase as acellular division enzyme in yeasts. Science124:722-723.
133. Ogur, M., S. Minckler, G. Lindegren, and C. C.Lindegren. 1952. The nucleic acids in a poly-ploid series of Saccharomyces. Arch. Biochem.Biophys. 40:175-184.
134. Ogur, M., S. Minckler, and D. 0. McClary. 1953.Desoxyribonucleic acid and the budding cyclein the yeasts. J. Bacteriol. 66:642-645.
135. Oyen, T. B. 1973. Chromosome I as a possiblesite for some rRNA cistrons in Saccharomycescerevisiae. FEBS Lett. 30:53-56.
136. Pauling, C., and L. Hamm. 1969. Properties of aradiation-sensitive mutant of Escherichia coli.II. DNA replication. Proc. Nat. Acad. Sci.U.S.A. 64:1195-1202.
137. Petes, T. D., B. Byers, and W. L. Fangman.1973. Size and structure of yeast chromosomalDNA. Proc. Nat. Acad. Sci. U.S.A.70:3072-3076.
138. Petes, T. D., and W. L. Fangman. 1972. Sedi-mentation properties of yeast chromosomalDNA. Proc. Nat. Acad. Sci. U.S.A.69:1188-1191.
139. Petes, T., and W. L. Fangman. 1973. Preferen-tial synthesis of yeast mitochondrial DNA ina factor-arrested cells. Biochem. Biophys.Res. Commun. 55:603-609.
140. Petes, T., C. S. Newlon, B. Byers, and W. L.Fangman. 1973. Yeast chromosomal DNA:size, structure, and replication. Cold SpringHarbor Symp. Quant. Biol. (in press).
141. Polakis, E. S., and W. Bartley. 1966. Changes indry weight, protein, deoxyribonucleic acid,ribonucleic acid and reserve and structuralcarbohydrate during the aerobic growth cycleof yeast. Biochem. J. 98:883-887.
142. Prescott, D. M., and M. A. Bender. 1962. Syn-thesis of RNA and protein during mitosis inmammalian tissue culture cells. Exp. CellRes. 26:260-268.
143. Ramirez, C., and J. J. Miller. 1962. Observationson vegetative nuclear division in Sac-charomyces cerevisiae. Can. J. Microbiol.8:603-608.
144. Retel, J., and R. J. Planta. 1968. The investiga-tion of the ribosomal RNA sites in yeast DNAby the hybridization technique. Biochim. Bio-phys. Acta 169:416-429.
145. Robinow, C. F., and J. Marak. 1966. A fiberapparatus in the nucleus of the yeast cell. J.Cell Biol. 29:129-151.
146. Rubin, G. M., and J. E. Sulston. 1973. Physical
linkage of the 5S cistrons to the 18S and 28Sribosomal RNA cistrons in Saccharomycescerevisiae. J. Mol. Biol. 79:521-530.
147. Scopes, A. W., and D. H. Williamson. 1964. Thegrowth and oxygen uptake of synchronouslydividing cultures of Saccharomyces cerevisiae.Exp. Cell Res. 35:361-371.
148. Schweizer, E., and H. 0. Halvorson. 1969. Onthe regulation of ribosomal RNA synthesis inyeast. Exp. Cell Res. 56:239-244.
149. Schweizer, E., C. MacKechnie, and H. 0. Hal-vorson. 1969. The redundancy of ribosomaland transfer RNA genes in Saccharomycescerevisiae. J. Mol. Biol. 40:261-277.
150. Sebastian, J., B. L. A. Carter, and H. 0.Halvorson. 1971. Use of yeast populationsfractionated by zonal centrifugation to studythe cell cycle. J. Bacteriol. 108:1045-1050.
151. Sebastian, J., B. L. A. Carter, and H. 0.Halvorson. 1973. Induction capacity of en-zyme synthesis during the cell cycle of Sac-charomyces cerevisiae. Eur. J. Biochem.37:516-522.
152. Sena, E. P., D. N. Radin, and S. Fogel. 1973.Synchronous mating in yeast. Proc. Nat.Acad. Sci. U.S.A. 70:1373-1377.
153. Sena, E., J. Welch, D. Radin, and S. Fogel. 1973.DNA replication during mating in yeast. Ge-netics 74:S248-249.
154. Sentandreu, R., and D. H. Northcote. 1969. Theformation of buds in yeast. J. Gen. Microbiol.55:393-398.
155. Shulman, R. W., Hartwell, L. H., and Warner, J.R. 1973. Synthesis of ribosomal proteins dur-ing the yeast cell cycle. J. Mol. Biol.73:513-525.
156. Sierra, J. M., R. Sentandreu, and J. R. Vil-lanueva. 1973. Regulation of wall synthesisduring Saccharomyces cerevisiae cell cycle.FEBS Lett. 34:285-290.
157. Simchen, G. 1974. Are mitotic functions indis-pensible in meiosis? Genetics 76:745-753.
158. Sinclair, J. H., B. J. Stevens, P. Sanghavi, andM. Rabinowitz. 1967. Mitochondrial satelliteand circulat DNA filaments in yeast. Science156:1234-1237.
159. Slater, M. L. 1973. Effect of reversible inhibitionof deoxyribonucleic acid synthesis on the yeastcell cycle. J. Bacteriol. 113:263-270.
160. Slater, M. L. 1974. Recovery of yeast fromtransient inhibition of DNA synthesis. Nature(London) 247:275-276.
161. Smith, D., P. Tauro, E. Schweizer, and H. 0.Halvorson. 1968. The replication of mitochon-drial DNA during the cell cycle in Sac-charomyces lactis. Proc. Nat. Acad. Sci.U.S.A. 60:936-942.
162. Smitt, W. W. S., J. M. Vlak, I. Molenaar, andTh. H. Rozijn. 1973. Nucleolar function of thedense crescent in the yeast nucleus. Exp. CellRes. 80:313-321.
163. Spoerl, E., and D. Looney. 1959. Synchronizedbudding of yeast cells following X-irradiation.Exp. Cell Res. 17:320-327.
164. Spoerl, E., L. E. Loveless, T. H. Weisman, and
196 HARTWELL
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

S. CEREVISIAE CELL CYCLE
R. J. Balske. 1954. Studies on cell division. II.X-radiation as a division inhibiting agent. J.Bacteriol. 67:394-401.
165. Streiblova, E., and K. Beran. 1963. Types ofmultiplication scars in yeasts, demonstratedby fluorescence microscopy. Folia Microbiol.8:221-227.
166. Streiblova, E., and K. Beran. 1963. Demonstra-tion of yeast scars by fluorescence microscopy.Exp. Cell Res. 30:603-605.
167. Tamaki, H. 1965. Chromosome behavior at mei-osis in Saccharomyces cerevisiae. J. Gen.Microbiol. 41:93-98.
168. Tauro, P., and H. 0. Halvorson. 1966. Effect ofgene position on the timing of enzyme synthe-sis in synchronous cultures of yeast. J. Bacter-iol. 92:652-661.
169. Tauro, P., H. 0. Halvorson, and R. L. Epstein.1968. Time of gene expression in relation tocentromere distance during the cell cycle ofSaccharomyces cerevisiae. Proc. Nat. Acad.Sci. U.S.A. 59:277-284.
170. Tauro, P., E. Schweizer, R. Epstein, and H. 0.Halvorson. 1969. Synthesis of macromoleculesduring the cell cycle in yeast, p. 101-118. In G.M. Padilla, G. L. Whitson, and I. L. Cameron(ed.), The cell cycle. Gene-enzyme inter-actions. Academic Press Inc., New York.
171. ten Berge, A. M. A., G. Zoutewelle, and K. W.van de Poll. 1973. Regulation of maltosefermentation in Saccharomyces carlsbergene-sis. I. The function of the gene MAL., asrecognized by mal, mutants. Mol. Gen.Genet. 123:233-246.
172. Throm, E., and W. Duntze. 1970. Mating-type-dependent inhibition of deoxyribonucleic acidsynthesis in Saccharomyces cerevisiae. J. Bac-teriol. 104:1388-1390.
173. Tkacz, J. S., E. B. Cybulska, and J. 0. Lampen.1971. Specific staining of wall mannan inyeast cells with fluorescein-conjugated con-canavalin A. J. Bacteriol. 105:1-5.
174. Tkacz, J. S., and J. 0. Lampen. 1972. Wallreplication in Saccharomyces species: use offluorescein-conjugated concanavalin A to re-veal the site of mannan insertion. J. Gen.Microbiol. 72:243-247.
175. Tkacz, J. S., and J. 0. Lampen. 1973. Surfacedistribution of invertase on growing Saccharo-myces cells. J. Bacteriol. 113:1073-1075.
176. Tonino, G. J. M., and T. H. Rozijn. 1966. On theoccurrence of histones in yeast. Biochem.Biophys. Acta 124:427-429.
177. TOnnesen, T., and J. D. Friesen. 1973. Inhibitorsof ribonucleic acid synthesis in Sac-charomyces cerevisiae: decay rate of messen-ger ribonucleic acid. J. Bacteriol. 115:889-896.
178. Udem, S. A., and J. R. Warner. 1972. RibosomalRNA synthesis in Saccharomyces cerevisiae.J. Mol. Biol. 65:227-242.
179. Van der Vliet, P. C., G. J. M. Tonino, and T. H.Rozijn. 1969. Studies on the yeast nucleus. HI.Properties of a deoxyribonucleoprotein complex derived from yeast. Biochem. Biophys.Acta 195:473-483.
180. Vendrely, R., and C. Vendrely. 1956. The resultsof cytophotometry in the study of the deox-yribonucleic acid (DNA) content of the nu-cleus. Int. Rev. Cytol. 5:171-197.
181. von Meyenburg, H. K. 1968. Der Sprossungszyk-lus von Saccharomyces cerevisiae. Pathol.Microbiol. 31:117-127.
182. Wickner, R. B. 1974. Mutants of Saccharo-myces cerevisiae that incorporate deoxy-thymidine-5W-monophosphate into deox-yribonucleic acid in vivo. J. Bacteriol.117:252-260.
183. Wiemken, A., P. Matile, and H. Moor. 1970.Vacuolar dynamics in synchronously buddingyeast. Arch. Mikrobiol. 70:89-103.
184. Williamson, D. H. 1964. Division synchrony inyeasts, p. 351-379. In E. Zeuthen (ed.), Syn-chrony in cell division and growth. John Wileyand Sons, Inc., New York.
185. Williamson, D. H. 1964. Techniques for synchro-nizing yeast cells, p. 589-591. In E. Zeuthen(ed.), Synchrony in cell division and growth.John Wiley & Sons, Inc., New York.
186. Williamson, D. H. 1964. The timing of deox-yribonucleic acid synthesis in the cell cycle ofSaccharomyces cerevisiae. Biochem. J.90:25-26P.
187. Williamson, D. H. 1965. The timing of deox-yribonucleic synthesis in the cell cycle ofSaccharomyces cerevisiae. J. Cell Biol.25:517-528.
188. Williamson, D. H. 1966. Nuclear events in syn-chronously dividing yeast cultures, p. 81-101.In I. L. Cameron and G. M. Padilla (ed.), Cellsynchrony. Academic Press Inc., New York.
189. Williamson, D. H. 1970. The effect of environ-ment and genetic factors on the replication ofmitochondrial DNA in yeast, p. 247-276. In P.L. Miller (ed.), Control of organelle develop-ment. XXIV Symposium of the Society forExperimental Biology. Academic Press Inc.,New York.
190. Williamson, D. H. 1973. Replication of thenuclear genome in yeast does not requireconcomitant protein synthesis. Biochem. Bio-phys. Res. Commun. 52:731-740.
191. Williamson, D. H., and E. Moustacchi. 1971.The synthesis of mitochondrial DNA duringthe cell cycle in the yeast Saccharomycescerevisiae. Biochem. Biophys. Res. Commun.42:195-201.
192. Williamson, D. H., and A. W. Scopes. 1960. Thebehavior of nucleic acids in synchronouslydividing cultures of Saccharomyces cerevisae.Exp. Cell Res. 20:338-349.
193. Williamson, D. H., and A. W. Scopes. 1961.Synchronization of division in cultures ofSaccharomyces cerevisiae by control of theenvironment, p. 217-242. In G. G. Meynelland H. Gooder (ed.), Microbial reaction to theenvironment. Eleventh Symposium of the So-ciety for General Microbiology. CambridgeUniversity Press, New York.
194. Williamson, D. H., and A. W. Scopes. 1961. Thedistribution of nucleic acids and protein be-
VOL. 38, 1974 197
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from

198 HARTWELL BACrERIOL. REV.
tween different sized yeast cells. Exp. Cell 196. Winge, O., and 0. Laustsen. 1939. Saccharo-Res. 24:151-153. myces Ludwig II Hansen, a balanced hetero-
195. Williamson, D. H., and A. W. Scopes. 1962. zygote. C. R. Trav. Lab. Carlsberg Ser.A rapid method for synchronizing division in Physiol. 22:357-370.the yeast, Saccharomyces cerevisiae. Nature 197. Yanigashima, N. 1969. Sexual hormones in(London) 193:256-257. yeast. Planta 87:110-118.
on March 27, 2020 by guest
http://mm
br.asm.org/
Dow
nloaded from





























