Embed Size (px)
Citation preview

1
Running Title: A PPR protein aiding in translation and splicing 1 2 Corresponding author: Hakim Mireau 3 Institut Jean-Pierre Bourgin, Institut National de la Recherche Agronomique, 4 AgroParisTech, CNRS, Université Paris-Saclay, RD10, 78026 Versailles Cedex, 5 France 6 Tel : (+33) 130833070 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29
Plant Physiology Preview. Published on November 4, 2015, as DOI:10.1104/pp.15.01591
Copyright 2015 by the American Society of Plant Biologists
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

2
The MTL1 Pentatricopeptide Repeat Protein is Required for Both 30 Translation and Splicing of the mitochondrial NADH Dehydrogenase 31 Subunit 7 mRNA in Arabidopsis 32 33 Nawel Haïli, Noelya Planchard, Nadège Arnal1, Martine Quadrado, Nathalie 34 Vrielynck, Jennifer Dahan2, Catherine Colas des Francs Small, and Hakim Mireau* 35 36 Institut Jean-Pierre Bourgin, Institut National de la Recherche Agronomique, 37 AgroParisTech, CNRS, Université Paris-Saclay, RD10, 78026 Versailles Cedex, 38 France (N.H., N.P., N.A., M.Q., N.V., J.D., H.M.); Université Paris-Sud, Université 39 Paris-Saclay, 91405 Orsay Cedex, France (N.H., N.P.); Australian Research Council 40 Centre of Excellence in Plant Energy Biology, University of Western Australia, 41 Crawley WA 6009, Australia (C.CFS.) 42 43 44 *Corresponding Author 45 46 One sentence summary: A mitochondria-targeted Pentatricopeptide Repeat Protein 47 specifically facilitates the translation of a respiratory chain transcript without inducing 48 any cleavage in its upstream non-coding region. 49 50 Footnotes: 51 1 Present address: Institut National de la Recherche Agronomique, Centre National de 52 Ressources Génomiques Végétales, 31326 Castanet Tolosan, France. 53 2 Present address: Department of Plant, Soil, and Entomological Sciences, University 54 of Idaho, Moscow, ID 83844, USA. 55 56 Author to whom correspondence should be addressed: Email: 57 [email protected] 58 59 The author responsible for distribution of materials integral to the findings presented 60 in this article in accordance with the policy described in the Instructions for Authors 61 (www.plantphysiol.org) is: Hakim Mireau ([email protected]). 62 63
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

3
ABSTRACT 64 65 Mitochondrial translation involves a complex interplay of ancient bacterial-like 66 features and host-derived functionalities. Although the basic components of the 67 mitochondrial translation apparatus have been recognized, very few protein factors 68 aiding in recruiting ribosomes on mitochondria-encoded mRNAs have been identified 69 in higher plants. In this study, we describe the identification of the Arabidopsis 70 MITOCHONDRIAL TRANSLATION FACTOR 1 (MTL1) protein - a new member 71 of the PentatricoPeptide Repeat family - and show that it is essential for the 72 translation of the mitochondrial nad7 mRNA. We demonstrate that mtl1 mutant plants 73 fail to accumulate the Nad7 protein, even though the nad7 mature mRNA is produced 74 and bears the same 5’ and 3’ extremities as in wild-type plants. We next observed that 75 polysome association of nad7 mature mRNA is specifically disrupted in mtl1 mutants, 76 indicating that the absence of Nad7 results from a lack of translation of nad7 mRNA. 77 These findings illustrate that mitochondrial translation requires the intervention of 78 gene-specific nuclear-encoded PPR trans-factors and that their action does not 79 necessarily involve the 5’ processing of their target mRNA as previously observed. 80 Interestingly, a partial decrease in nad7 intron 2 splicing was also detected in mtl1 81 mutants suggesting that MTL1 is also involved in group II intron splicing. However, 82 this second function appears less essential for nad7 expression than its role in 83 translation. MTL1 will be instrumental to understand the multi-functionality of PPR 84 proteins and the mechanisms governing mRNA translation and intron splicing in plant 85 mitochondria. 86 87 88 89 90 91 92 93 94 95 96 97
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

4
INTRODUCTION 98 99 Translation is the fundamental process decoding the genetic message present on 100 mRNAs into proteins. In plant cells, mRNA translation occurs in the cytoplasm but 101 also in two organelles, mitochondria and plastids. Because of their prokaryotic origin, 102 the translation machineries operating in these two organelles share many 103 characteristics with the bacterial translation apparatus (Bonen, 2004). However, most 104 of these bacterial-like features have been modified throughout evolution and current 105 organellar translation systems cooperate with numerous nuclear-encoded eukaryotic 106 trans-factors. The divergence from bacteria is particularly obvious in plant 107 mitochondria notably because mitochondrial mRNAs lack typical Shine and Dalgarno 108 (SD) motif in their 5’ leaders, and alternative start codons other than AUG are often 109 used to initiate translation (Bonen, 2004). Proteomic and bioinformatic analyses 110 allowed the identification of most proteins and RNA factors forming the core of the 111 plant mitochondrial translation machinery including translation initiation and 112 elongation factors as well as ribosomal proteins (Bonen, 2004; Bonen and Calixte, 113 2005). However, the dynamics of this machinery remains largely obscure. In 114 particular, nothing is known about the recruitment of mitochondrial ribosomes on 5’ 115 untranslated regions (UTR) in the absence of SD motif and about the recognition of 116 the correct translation initiation codon by the small ribosomal subunit. The high 117 degree of sequence divergence among 5’ leaders of mitochondrial genes suggests a 118 ribosome recruitment mechanism involving gene-specific cis-sequences and 119 transfactors (Hazle and Bonen, 2007; Choi et al., 2012). Up to now, only two proteins 120 belonging to the Pentatricopeptide Repeat (PPR) family have been found to promote 121 mitochondrial translation in higher plants (Uyttewaal et al., 2008b; Manavski et al., 122 2012). How they facilitate translation is still unclear, like for the few characterized 123 PPR proteins shown to participate in plastid translation (Fisk et al., 1999; Schmitz-124 Linneweber et al., 2005; Cai et al., 2011; Zoschke et al., 2012a; Zoschke et al., 125 2012b). The plastid PENTATRICOPEPTIDE REPEAT PROTEIN 10 (PPR10) 126 protein of maize is the only one for which the function has been elucidated at the 127 molecular level. It was shown that upon binding PPR10 impedes the formation of a 128 stem loop structure in the 5’ leader of the ATP synthase subunit c (atpH) mRNA 129 permitting the recruitment of ribosomes through the liberation of an SD motif (Prikryl 130 et al., 2011). 131
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

5
PPR proteins represent a large family of RNA binding proteins that has massively 132 expanded in terrestrial plants (Barkan and Small, 2014). Most eukaryotes encode a 133 handful of these proteins, whereas plant nuclear genomes express over 400 PPR 134 proteins that are almost exclusively predicted to target mitochondria and/or plastids 135 (Lurin et al., 2004; O'Toole et al., 2008). This family of proteins is characterized by 136 the succession of tandem degenerate motifs of approximately 35 amino acids (Small 137 and Peeters, 2000; Lurin et al., 2004). Based on the length of these repeats, the PPR 138 family has been divided in two groups of roughly equal size in higher plants. P-type 139 PPR proteins contain only successions of canonical 35-amino-acid repeats (P), 140 whereas PLS PPR proteins are composed of sequential repeats of P, short (S) and long 141 (L) PPR motifs. P-type PPR proteins were shown to participate in various aspects of 142 organellar RNA processing whereas PLS PPR proteins have been almost exclusively 143 associated with C-to-U RNA editing (reviewed in Barkan and Small, 2014; Hammani 144 and Giegé, 2014). Recent crystal structures showed that PPR motifs adopt an 145 antiparallel helix-turn-helix fold whose repetition forms a solenoid-like structure 146 (Ringel et al., 2011; Howard et al., 2012; Ban et al., 2013; Yin et al., 2013; Coquille 147 et al., 2014; Gully et al., 2015). PPR tracks organize highly specific interaction 148 domains that were shown to associate with single-stranded RNAs (Schmitz-149 Linneweber et al., 2005; Beick et al., 2008; Williams-Carrier et al., 2008; Uyttewaal 150 et al., 2008a; Pfalz et al., 2009; Cai et al., 2011; Hammani et al., 2011; Prikryl et al., 151 2011; Zhelyazkova et al., 2011; Khrouchtchova et al., 2012; Manavski et al., 2012; 152 Ke et al., 2013; Yin et al., 2013). The mechanism of sequence-specific RNA 153 recognition by PPR proteins was recently uncovered and combinations involving 154 amino acid 6 of one motif and amino acid 1 of the subsequent motif correlate strongly 155 with the identity of the RNA base to be bound (Barkan et al., 2012; Takenaka et al., 156 2013; Yagi et al., 2013). 157 Besides those involved in RNA editing, few mitochondria-targeted PPR proteins have 158 been characterized to date. Thus, our knowledge of the mechanisms governing the 159 production and the expression of mitochondrial RNAs in higher plants is very limited. 160 In this analysis, we describe the function of a novel mitochondria-targeted PPR 161 protein of Arabidopsis, called MITOCHONDRIAL TRANSLATION FACTOR 1 162 (MTL1). Genetic and biochemical analyses indicate that MTL1 is essential for the 163 translation of the mitochondrial nad7 mRNA. Effectively, the Nad7 protein does not 164 accumulate to detectable levels in mtl1 mutants and this absence correlates with a lack 165
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

6
of association of nad7 mature mRNA with mitochondrial polysomes. Interestingly, a 166 partial but significant decrease in nad7 intron 2 splicing was also detected in mtl1 167 mutants suggesting that the MTL1 protein is also involved in group II intron splicing. 168 Since the decrease in splicing was only partial, this second function of MTL1 appears 169 less essential for nad7 expression than its role in translation. 170 171 172
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

7
RESULTS 173 174 Arabidopsis mtl1 mutants display a slow growth phenotype 175 In an effort to better understand gene expression in higher plant mitochondria, a series 176 of Arabidopsis mutants bearing T-DNA insertions predicted to affect mitochondrially-177 targeted P-type PPR proteins was collected. The search for interesting mutants 178 revealed the mtl1-1 line for which homozygous mutant plants displayed significantly 179 retarded growth on soil compared to wild type (WT, Fig. 1A). The affected PPR gene 180 in this line corresponded to the At5g64320 gene and encoded an 82-kD protein 181 comprising 16 PPR repeats according to predictions (Fig. 1B) (Lurin et al., 2004). A 182 second T-DNA insertion line affecting the same gene was subsequently identified. 183 This second allelic mutant, named mtl1-2, displayed the same growth alterations as 184 mtl1-1, strongly suggesting that the developmental phenotype observed in these lines 185 was effectively associated with inactivation of the At5g64320 gene (Fig. 1A and 1B). 186 RT-PCR analysis indicated that no detectable full length mRNA derived from the 187 At5g64320 gene accumulates in both mtl1-1 and mtl1-2 mutant plants, supporting that 188 both identified mutant lines represented null mutants (Fig. 1C). 189 mtl1 mutant plants showed various developmental abnormalities compared to wild-190 type plants. Both mutant lines grew rather slowly compared to the wild type but 191 reached about 80% of the size of Col-0 plants when cultured on soil for two and half 192 months (Supplemental Fig. S1A). Additionally, mtl1 plants generally bear deformed 193 and dark green rosette leaves (Fig. 1A). They also needed nearly twice as much time 194 to flower compared to the wild type, but are fertile (Supplemental Fig. S1B). Seeds 195 produced by homozygous mtl1 mutant plants were darker than normal Arabidopsis 196 seeds but they germinate with around 80% efficiency on soil or in vitro (Supplemental 197 Fig. S1C). 198 199 The Arabidopsis MTL1 gene encodes a mitochondrion-targeted PPR protein. 200 The cellular distribution of the MTL1 protein was verified by expressing a GFP 201 translational fusion comprising the first 109 amino acids of MTL1 in transgenic 202 Arabidopsis plants. Roots of transformed plants were observed under a confocal 203 microscope and the GFP fluorescence appeared as a punctuated signal distributed 204 throughout the cytoplasm. The use of MitoTracker® Red indicated that these signals 205
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

8
corresponded to mitochondria (Fig. 2A). To confirm these results and to gain insight 206 into the submitochondrial distribution of MTL1, a C-terminal fusion comprising the 207 full-length MTL1 protein and ten copies of the MYC epitope tag was expressed in 208 Arabidopsis. This strategy was favored, as rabbit polyclonal antibodies produced 209 against fragments of MTL1 did not succeed in detecting the protein from Arabidopsis 210 protein extracts. Expression of the MTL1-MYC protein in the mtl1-1 mutant line 211 rescued the mutant phenotypes, strongly supporting the functionality of the fusion 212 protein (Supplemental Fig. S2A and S2B). The distribution of the tagged MTL1 was 213
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.
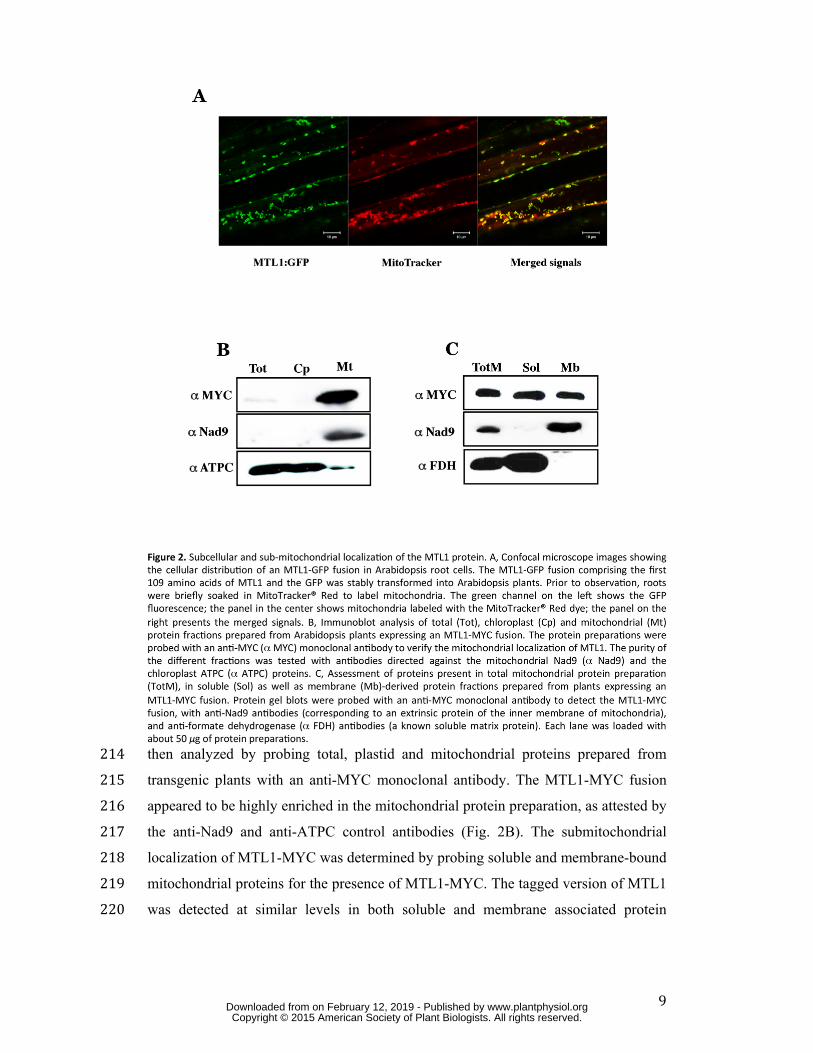
9
then analyzed by probing total, plastid and mitochondrial proteins prepared from 214 transgenic plants with an anti-MYC monoclonal antibody. The MTL1-MYC fusion 215 appeared to be highly enriched in the mitochondrial protein preparation, as attested by 216 the anti-Nad9 and anti-ATPC control antibodies (Fig. 2B). The submitochondrial 217 localization of MTL1-MYC was determined by probing soluble and membrane-bound 218 mitochondrial proteins for the presence of MTL1-MYC. The tagged version of MTL1 219 was detected at similar levels in both soluble and membrane associated protein 220
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

10
preparations (Fig. 2C). The use of anti-Nad9 and anti-FDH control antibodies 221 confirmed the proper separation of both mitochondrial protein fractions. 222 223 mtl1 mutants are impaired in the synthesis of respiratory complex I 224 The mitochondrial localization of MTL1 led us to consider that a defective respiratory 225 activity may be at the origin of mtl1 mutant phenotypes. To clarify the origin of a 226 potential respiratory perturbation in these plants, we analyzed the steady state levels 227 of the different respiratory chain complexes in comparison to the wild type. 228 Mitochondria prepared from mtl1 and wild-type plants were lysed in the presence of 229 digitonin and the solubilized respiratory complexes were separated on blue-native gel. 230 Coomassie blue staining showed that most respiratory complexes accumulated to 231 similar levels in wild-type and mutant plants, except for complex I and the super 232 complex I + III (Fig. 3A left panel). An in-gel NADH-dehydrogenase activity test 233 confirmed the lack of both forms of complex I in mtl1 mutants (Fig. 3A right panel). 234 To further analyze the impact of mtl1 mutations on Arabidopsis respiration, oxygen 235 uptake in the dark was determined on detached leaves in both mutant and wild-type 236 plants. This led us to show that mtl1 plants consumed nearly twice as much oxygen as 237 in Col-0 plants (Supplemental Fig. S3). Since this increase of oxygen consumption 238 could be consecutive of a strong induction of the alternative respiratory pathways in 239 the mutant plants, the expression levels of alternative NADH dehydrogenases (NDA 240 and NDB) and alternative oxidase (AOX) genes were measured. Quantitative RT-241 PCR indicated that steady state levels of AOX1A, NDA1 and NDB4 transcripts over 242 accumulate from 4 to 8 fold in mtl1 plants (Fig. 3B). The strong increase in AOX 243 expression was further estimated by immunoblot analysis of mitochondrial extracts 244 with an antibody to AOX. The results showed a highly significant increase in the 245 AOX signal in both mtl1 mutants (Fig. 3C). Our results indicated that mtl1 plants are 246 complex I respiratory mutants and that the alternative respiratory pathway is strongly 247 activated in these plants. 248 249 mtl1 mutants do not accumulate the mitochondrial Nad7 protein. 250 To better understand the lack of assembled complex I in mtl1 plants, we next analyzed 251 the expression of all mitochondria-encoded complex I subunits in the mutants. In a 252 first approach, steady-state levels and processing efficiency of mitochondria-encoded 253 complex I mRNAs were analyzed by RNA gel blot analysis in both mtl1 and wild-254
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

11
type plants. This approach showed that all nine mitochondrial mRNAs encoding 255 complex I subunits accumulate to detectable levels in mtl1 plants (Fig. 4A and 256 Supplemental Fig. S4). Most of the corresponding mature transcripts are produced in 257 amounts close to wild-type levels, with a slight increase notably for nad1, nad4, nad5 258 and nad6 mRNAs. The only exceptions are the nad7 and nad9 mature transcripts that 259 are much less abundant in mtl1 plants compared to Col-0 (Fig. 4A). In the case of 260 nad7, this decrease was associated with the over-accumulation of an unspliced mRNA 261 precursor, indicating that the loss of MTL1 caused, directly or indirectly, a splicing 262
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

12
defect of nad7 mRNA (Fig. 4A). Quantitative RT-PCR analysis measuring the 263 splicing efficiency of all intron-containing mitochondrial transcripts allowed us to 264 determine that this splicing defect concerns the second intron of nad7 (Supplemental 265 Fig. S5). A moderate 2-fold decrease of nad2 intron 1 and 2 splicing efficiency was 266 also detected in mtl1 mutants with this analysis. To analyze the processing of nad 267 transcripts in more details, the 5’ and 3’ extremities of all mitochondria-encoded nad 268 mRNAs were amplified by circular RT-PCR in both wild-type and mtl1 plants. No 269 obvious processing differences for any of these mRNAs could be identified in the 270
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

13
mutants. Cloning and sequencing of the obtained amplification products further 271 confirmed that all complex I mature transcripts in mtl1 plants bear 5’ and 3’ 272 extremities that are identical to the ones found in wild-type, including nad7 273 (Supplemental Fig. S6). Shorter cRT-PCR products for nad7 were more apparent in 274 mtl1 plants compared to the wild type. Cloning and sequencing of these fragments in 275 both Col-0 and mtl1 mutants indicated that they derived from 5’-truncated nad7 276 transcripts that appear accumulate to slightly higher levels in the mutants. Their 277 abundance is insufficient to be visible on RNA gel blot (Fig. 4A). We next reasoned 278 that partial destabilization of nad7 and nad9 mature transcripts could result from 279 difficulties in expressing the Nad7 and/or Nad9 proteins. Consequently, mitochondrial 280 protein extracts were prepared from both mutants and wild-type plants and probed 281 with anti-Nad7 and anti-Nad9 antibodies. Whereas the Nad9 protein was still 282 detectable although significantly reduced, no trace of Nad7 could be found in either 283 mtl1 mutant lines (Fig. 4B). This showed that mtl1 mutants did not accumulate 284 detectable levels of Nad7 protein and that the lack of this mitochondrial protein was 285 very likely responsible for the different molecular and physiological perturbations 286 observed in mtl1 plants. 287 288 The nad7 mRNA does not associate with translating mitochondrial polysomes in 289 mtl1 mutants. 290 We next wondered whether the lack of Nad7 protein in mtl1 plants could simply 291 result from insufficient accumulation of the nad7 mature mRNA. To answer this 292 question, we used the Arabidopsis bso-insensitive roots 6 (bir6) ppr mutant that was 293 also described to accumulate low levels of mature nad7 transcript in response to 294 difficulties to splice the first nad7 intron (Supplemental Fig. S7A, Koprivova et al., 295 2010). Comparative RNA gel blot analyses suggested that bir6 plants accumulate less 296 nad7 mature transcript compared to mtl1 mutants (Supplemental Fig. S7B). Indeed, 297 quantitative RT-PCR further showed that bir6 plants contain only 6% of wild-type 298 levels of mature nad7 transcripts whereas mtl1-1 about 20% (Supplemental Fig. S7C). 299 Probing of mitochondrial proteins with anti-Nad7 and anti-Nad9 antibodies showed 300 that bir6 plants accumulate slightly reduced levels of the Nad9 protein compared to 301 wild-type and, unlike mtl1 mutants, very low but still detectable levels of Nad7 302 (Supplemental Fig. S7D). Thereby, the comparison between mtl1 and bir6 mutants 303 supported that the loss of Nad7 in mtl1 plants was not directly linked to the low levels 304
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

14
of nad7 mature transcript, but likely from molecular events involving directly the 305 production of the Nad7 protein. However, the lack of antibody against the other 306 mitochondria-encoded complex I subunits could not allow us to rule out the 307 possibility that other Nad proteins were also missing in mtl1 mutants. 308 Therefore, the translation status of all mitochondria-encoded nad transcripts was 309 analyzed by polysome sedimentation analysis in both wild-type and mtl1-1 plants. 310 Polysomes were isolated from Arabidopsis inflorescences and fractionated on 311 continuous sucrose density gradient. Ten fractions were collected along the gradients 312 after centrifugation. Polysome integrity was shown by the distribution of ribosomal 313 RNAs along the gradients in the presence of MgCl2 (Supplemental Fig. S8A). The 314 disruption of polysomes by addition of EDTA in the gradients indicated that 315 polysomal RNA migrated towards the center and the bottom of the gradients, whereas 316 free mRNAs accumulate in the upper fractions (Supplemental Fig. S8A). RNA was 317 extracted from each fraction and subjected to RNA gel blot analysis using probes 318 specifically recognizing each mitochondria-encoded complex I transcripts (Fig. 5). 319 The hybridized membranes were then exposed to phosphorimager screen and the 320 relative distribution of each hybridization signal was determined for each mature 321 mRNA (Supplemental Fig. S9). Most mRNAs distributed similarly all along the 322 gradients in Col-0 and mtl1 extracts, indicating that free and polysome-associated 323 transcripts exist for all complex I transcripts. In contrast, the distribution of nad7 324 mRNA was drastically different between mtl1-1 and wild-type plants. In the former, 325 most of the nad7 signal was concentrated in the top 4 fractions, suggesting that no 326 ribosomes are loaded on nad7 mature transcripts in the absence of a functional copy 327 of MTL1. No such difference was found for the other complex I transcripts, 328 supporting a specific role of MTL1 in the translation of nad7 mRNA. We could 329 conclude from these results that nad7 mRNA is not properly translated in the absence 330 of MTL1 and that subsequently Nad7 is indeed not produced in mtl1 mutants. 331 To further support the role of MTL1 in mitochondrial translation, we analyzed the 332 distribution of the MTL1-MYC protein fusion on a polysome gradient prepared from 333 mtl1-complemented plants (Supplemental Fig. S2A). Total proteins were prepared 334 from the ten collected fractions and fractions 2 to 10 were subjected to immunoblot 335 analysis using anti-MYC antibodies. The result indicates that the MTL1-MYC fusion 336 is detected all along the gradient and does co-sediment with heavy polysome fractions 337 on sucrose density gradient (Supplemental Fig. S8B). 338
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

15
339 340
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

16
DISCUSSION 341 342 MTL1 is a P-type PPR protein essential for nad7 mature mRNA translation 343 PPR proteins have been implicated in multiple aspects of organelle gene expression, 344 but very few of them have been specifically associated with the translation of 345 mRNAs. In this study, we convincingly showed that the mitochondria-targeted MTL1 346 protein of Arabidopsis is required for the translation initiation of the nad7 mRNA. To 347 reach this conclusion, we observed that the Nad7 protein did not accumulate in mtl1 348 mutants and that this deficiency likely resulted from the lack of synthesis of Nad7 as 349 nad7 mature mRNA does not co-sediment with translating mitochondrial polysomes 350 on sucrose density gradient. We also showed that the absence of Nad7 synthesis does 351 not result from an incorrect processing of nad7 transcripts as nad7 mature mRNAs 352 bear the same 5’ and 3’ extremities in mtl1 and wild-type plants. Furthermore, 353 comparison with another Arabidopsis ppr mutant (bir6, Koprivova et al., 2010) 354 allowed us to show that the lack of Nad7 protein accumulation in mtl1 plants did not 355 result from an insufficient accumulation of nad7 mature mRNA. At last, MTL1 was 356 found to co-sediment with translating polysome fractions on sucrose density gradient. 357 We also observed a significant reduction of nad9 expression in mtl1 plants. A role of 358 MTL1 in nad9 translation is unlikely as nad9 transcripts still co-sediment with 359 mitochondrial polysomes in mtl1-1 plants. It is more likely that the effect observed on 360 nad9 expression is secondary to the loss of respiratory complex I as observed in many 361 other complex I deficient mutants (Gutierres et al., 1997; Keren et al., 2009; Kuhn et 362 al., 2011; Hsu et al., 2014; Hsieh et al., 2015). 363 Very little is known about the molecular mechanisms governing the expression of 364 mitochondrial genes in plants. Translation is by far the least understood RNA 365 processing step in plant mitochondria. Due to its origin, the mitochondrial translation 366 machinery exhibits certain bacterial-type characteristics (Bonen, 2004). However in 367 plants, the vast majority of mitochondrial mRNAs lacks a Shine-Dalgarno (SD) motif 368 upstream of their translation initiation codon, certainly because the mitochondrial 18S 369 rRNA does not contain the corresponding anti-SD sequence (Hazle and Bonen, 2007). 370 In bacteria, the ribosomal S1 protein was shown to promote ribosome recruitment for 371 mRNAs bearing leaders devoid of SD motif (Sørensen et al., 1998). Such a 372 mechanism is unlikely to permit ribosome association to mitochondrial mRNAs in 373
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

17
plants as the S1 counterpart lacks the necessary RNA-binding domain (Hazle and 374 Bonen, 2007). To compensate for the absence of a general ribosome recruitment 375 system on mitochondrial mRNAs in plants, translation initiation likely relies on gene 376 specific trans-factors to allow for ribosome anchoring and guidance towards the 377 correct AUG start codon. Besides MTL1, very few others mitochondrial translation 378 factors were identified in higher plants so far. The first one was the Arabidopsis 379 PENTATRICOPEPTIDE REPEAT PROTEIN 336 protein that co-purifies with 380 mitochondrial polysomes but whose action on mitochondrial translation is currently 381 not understood (Uyttewaal et al., 2008b). More recently, the maize 382 PENTATRICOPEPTIDE REPEAT PROTEIN 6 (MPPR6) protein was demonstrated 383 to be essential for the translation of the ribosomal protein s3 (rps3) mitochondrial 384 mRNA (Manavski et al., 2012). However, mppr6 mutants accumulate 5’-extended 385 rps3 transcripts suggesting that the primary function of MPPR6 resides in the 5’ 386 processing of rps3 transcripts to yield translatable mature mRNA. Finally, two 387 fertility-restorer PPR proteins were shown to directly or indirectly inhibit the 388 translation of their cognate sterility-inducing mitochondrial gene. This concerns the 389 rice RF1A protein that upon RNA processing produces a non-translatable orf79 390 transcript and the radish PPR-B protein that is suspected to specifically impede the 391 loading or the progression of mitochondrial ribosomes on the orf138 mRNA (Kazama 392 et al., 2008; Uyttewaal et al., 2008a). Several PPR proteins have also been found to 393 participate in plastid translation and PPR10 is the only one for which the molecular 394 action has been elucidated (Fisk et al., 1999; Schmitz-Linneweber et al., 2005; Cai et 395 al., 2011; Prikryl et al., 2011; Zoschke et al., 2012a; Zoschke et al., 2012b). PPR10 396 function involves binding to the atpH 5’ mRNA leader to prevent the formation of a 397 stem loop structure which otherwise would sequestrate an SD sequence and thereby 398 impede ribosome entry on the atpH mRNA (Prikryl et al., 2011). The example of 399 PPR10 indicates that altering the structure of RNA sequences could represent a major 400 action of PPR proteins to disclose RNA segments and allow the action of other RNA 401 processing enzymes, notably the ones enhancing mRNA translation. In the case of 402 MTL1, we showed that the 5’-end processing of nad7 is not perturbed in mtl1 mutants 403 indicating that MTL1 primary function is not in nad7 5’ processing. Thereby, this 404 strongly supports a direct influence of MTL1 on nad7 mature mRNA translation 405 through a mechanism that still needs to be understood. According to the PPR code 406 (Barkan et al., 2012), a rather good putative binding site (GAATCATCTCTTT) for 407
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

18
MTL1 is present 35 nucleotides upstream of the nad7 AUG. However, we did not 408 observe obvious binding of recombinant MTL1 to this sequence by gel shift assay. 409 The 5’ UTR of nad7 transcript is 375-nucleotide long and as in most mitochondrial 410 mRNAs lacks any obvious Shine and Dalgarno sequence. MTL1 may be necessary to 411 recruit or guide the mitochondrial ribosomes toward the correct start codon of nad7. 412 To this end, MTL1 may influence short or long-range RNA interactions within nad7 413 5’ leader to prevent the formation of a stem-loop structure as proposed for the plastid 414 PPR10 protein. Alternatively, MTL1 may interact with the mitochondrial ribosomes 415 or may be necessary to recruit other co-factors essential for nad7 translation. Further 416 molecular investigations on the mode of action of MTL1 are necessary to find which 417 one of these hypotheses is correct. 418 419 The MTL1 protein is also essential for optimal splicing of nad7 intron 2. 420 Our results showed that mtl1 plants were also affected in the splicing of several 421 mitochondrial group II introns. This concerned introns present in nad2 and nad7 pre-422 mRNAs (Supplemental Fig. S5). The decrease measured for nad2 intron splicing was 423 very modest and induced no reduction in the accumulation of the corresponding 424 mature transcript (Supplemental Fig. S4). These very mild alterations could simply 425 represent pleiotropic effects of the loss of complex I as observed in several other 426 complex I mutants (Haïli et al., 2013; Colas des Francs-Small et al., 2014; Hsieh et 427 al., 2015) and do not support a direct role of MTL1 in the splicing of these introns. 428 Oppositely, the 16-fold decrease of nad7 intron 2 splicing efficiency resulted in a 429 measured 4-time reduction in the accumulation level of nad7 mature mRNA (Fig. 4 430 and Supplemental Fig. S5). This effect is sufficiently significant to support a role of 431 MTL1 in the splicing of this intron. However, the comparison made with the bir6 432 mutant clearly showed that the reduction of nad7 splicing in mtl1 mutants is 433 insufficient to explain the lack of complex I in these plants. This conclusion is further 434 supported by the recent study of the slow growth 3 (slo3) mutant which displays a 435 similar reduction in nad7 intron 2 splicing but still accumulates detectable respiratory 436 complex I (Hsieh et al., 2015). A comparison of published Arabidopsis mitochondrial 437 splicing mutants indicates that splicing reductions of at least 500 times compared to 438 the wild type seems necessary to result in a near complete loss of complex I on BN 439 gels (Koprivova et al., 2010; Kühn et al., 2011; Cohen et al., 2014; Colas des Francs-440 Small et al., 2014; Hsieh et al., 2015). The decrease in splicing efficiency can be 441
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

19
much milder though when several complex I introns are concomitantly affected in a 442 same mutant (Keren et al., 2012; Zmudjak et al., 2013). The manner by which PPR 443 proteins assist group II intron splicing is currently unknown at the molecular level. 444 Mitochondrial, but also plastid, group II introns have lost their ability to self-splice 445 and require the participation of numerous nuclear-encoded general and specific 446 factors to achieve efficient splicing in vivo (Bonen, 2008; Ostersetzer-Biran, 2014). 447 Although poorly conserved at the sequence level, group II introns arrange themselves 448 into a phylogenetically conserved secondary structure comprising 6 helical domains 449 that radiate from a central hub (Pyle et al., 2007). It has been speculated, based on 450 their activity that is likely limited to a passive binding to RNA, that P-type PPR 451 proteins may facilitate intron splicing by stabilizing the structure of group II introns 452 into their catalytically active forms. Several nuclear-encoded splicing factors were 453 previously reported to assist the removal of nad7 intron 2, namely the two general 454 splicing factors nMAT2 and mCSF1 as well as the SLO3 PPR protein, which is 455 specific for this intron (Keren et al., 2009; Zmudjak et al., 2013; Hsieh et al., 2015). It 456 is interesting to note that none of the corresponding single mutants showed a complete 457 loss of nad7 intron 2 splicing. This suggests that the loss of one of these factors at a 458 time may be partially compensated by the others and that in the absence of MTL1 the 459 nad7 intron 2 conserves a poorly active structure resulting in a sub-optimal splicing 460 reaction. In the future, it will be interesting to explore the inter-functionality of these 461 4 splicing factors and understand how they may functionally promote the removal of 462 nad7 intron 2 in Arabidopsis. 463 464 MTL1 is one of the first mitochondrial bi-functional PPR-P proteins identified in 465 plants 466 Our results revealed therefore that the Arabidopsis MTL1 protein carries two 467 independent activities, which are both required for the proper expression of the nad7 468 transcript. It is currently difficult to tell with certainty which of these two functions is 469 the most essential with respect to nad7 expression. However as indicated, we obtained 470 good indications supporting that the defect in nad7 intron 2 splicing is insufficient to 471 explain the lack of complex I in mtl1 mutants, which would favor the translation as 472 being the most essential role for MTL1. This hypothesis is hard to validate since we 473 have no way to verify that nad7 mRNA translation would still not occur if only fully-474 spliced nad7 transcripts accumulated in mtl1 plants. Effectively, an increase in the 475
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

20
production of mature nad7 mRNA may result in a certain degree of translation of 476 nad7 transcript in the absence of MTL1. It is not uncommon to observe multiple and 477 apparently unrelated effects of ppr mutations on RNA metabolism in plant organelles 478 (reviewed in Barkan and Small, 2013). Some of these effects can result from 479 secondary consequences of ppr mutations, whereas other ones can be directly 480 imputable to the inactivation of multi-functional PPR proteins. For instance, PPR 481 proteins involved in C-to-U RNA editing are often considered multi-functional in the 482 sense that they frequently edit multiple RNA targets, notably in plant mitochondria 483 (Hammani and Giegé, 2014). PPR proteins supporting completely different RNA 484 processing activities have been found much less frequently and interestingly known 485 examples concern PPR proteins having a role in mRNA translation. The plastid PGR3 486 and PPR10 proteins were shown to be involved in both stabilization and translation of 487 their cognate mRNA targets (Yamazaki et al., 2004; Cai et al., 2011; Prikryl et al., 488 2011). The analysis of PPR10 in maize provided a molecular framework to explain 489 how a PPR protein can achieve both of these functions. As already mentioned, PPR10 490 facilitates translation of the atpH mRNA by uncovering a ribosome binding site and 491 concomitantly through the same binding site stabilizes the atpH transcript by blocking 492 the progression of degrading ribonucleases (Prikryl et al., 2011). PPR proteins 493 involved in organelle translation like CRP1 and MPPR6 or the restorer of fertility 494 RF1A of rice have also been associated with RNA cleavage events (Fisk et al., 1999; 495 Kazama et al., 2008; Manavski et al., 2012). Others like PGR3, ATP4, SVR7 but also 496 CRP1 are required for the translation of distinct mRNAs (Barkan et al., 1994; 497 Yamazaki et al., 2004; Cai et al., 2011; Zoschke et al., 2012a; Zoschke et al., 2012b). 498 Oppositely, PPR proteins engaged in intron splicing are rarely found involved in other 499 mRNA processing events, and MTL1 stands as one of the first counterexample in 500 plants. A precedent was found though in plastids with the maize PPR5 protein which 501 was shown to facilitate intron splicing of trnG-UCC pre-mRNA and to concomitantly 502 stabilize the unspliced precursor by masking a ribonuclease-sensitive site (Williams-503 Carrier et al., 2008). Another example concerns the AEF1/MRP25 that is essential for 504 the splicing of atpF in plastids but which also promotes RNA editing of nad5 and 505 atpF in mitochondria and plastids respectively (Yap et al., 2015). The multi-506 functionality of PPR proteins is facilitated by the fact that this class of RNA binding 507 proteins show a relaxed recognition specificity to some extend which allows them to 508 recognize multiple RNA targets sharing limited sequence homology (Barkan et al., 509
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

21
2012). However, the apparent restriction of PPR proteins engaged in splicing to a 510 single function could reflect the necessity for these proteins to fulfill their action with 511 very high specificity, preventing them from having several RNA binding targets. The 512 dual activity of MTL1 in both nad7 mRNA translation and splicing represents 513 therefore an interesting new situation for PPR proteins that very likely involves 514 distinct RNA binding sites. Additional investigations are required to understand how 515 MTL1 can control these two apparently disconnected RNA processing events. MTL1 516 stands therefore as a valuable model to understand the multi-functionality of PPR 517 proteins and its molecular deciphering will provide new insight on the mechanisms 518 governing mRNA expression in plant mitochondria. 519 520 MATERIAL AND METHODS 521 522 Plant Material, Growth Conditions and Complementation Analysis 523 The Arabidopsis Col-0 plants were obtained from the Arabidopsis Stock Centre of 524 Institut National de la Recherche Agronomique in Versailles 525 (http://dbsgap.versailles.inra.fr/portail/). The N539066 (mtl1-1), N567427 (mtl1-2) 526 and bir6 (N500310) Arabidopsis T-DNA insertion lines were recovered from the 527 European Arabidopsis Stock Centre (http://arabidopsis.info/). Plants homozygous for 528 the insertions were identified by PCR genotyping. mtl1-1 and mtl1-2 mutants were 529 genotyped using the MTL1-3 and MTL1-5 primers combined with the LB-SALK2 530 primer. Plants were grown on soil under long-day conditions (16 h of light and 8 h of 531 dark). For complementation test with a MYC tagged version of MTL1, the complete 532 MTL1 gene was amplified by PCR from Arabidopsis total DNA using GWMTL1-1 533 and GWMTL1-11 primers, cloned into the pDONR207 vector by Gateway® BP 534 reaction (Invitrogen), and subsequently transferred into the pGWB20 expression 535 vector (Nakagawa et al., 2007) by LR reaction (Invitrogen). The resulting vector was 536 transformed into Agrobacterium tumefaciens C58C51 and used for floral dip 537 transformation of mtl1-1/MTL1 and mtl1-2/MTL1 heterozygous plants. Transformed 538 plants were selected on hygromycin and transgenic homozygous mutants were 539 identified by PCR analysis using the GWMTL1-2 and MTL1-8 primers to find 540 homozygous mutant plants, the LBSALK2 and GWMTL1-2 primers to check for the 541
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

22
presence of the T-DNA mutations, and the P35SU and GWMTL1-2 primers to check 542 for the presence of the complementing transgene (MTL1-MYC). 543 544 Primers 545 The primers used in this work are listed in Supplemental Table S1. 546 547 Analyses of Targeting via GFP Fusion 548 The DNA region encoding the first 109 amino acids of MTL1 was PCR amplified 549 with GWMTL1-1 and GWMTL1-6 primers. The AttB1 and AttB2 recombination sites 550 were then completed in a second PCR amplification using the GW5 and GW3 551 oligonucleotides. The PCR product was cloned into pDONR207 vector using the 552 Gateway® BP clonase enzyme mix, and sequenced to check PCR accuracy. The 553 obtained entry clone was then transferred into pMDC83 expression vector (Curtis and 554 Grossniklaus, 2003) by Gateway® LR reaction to create a translational fusion 555 between the MTL1 targeting sequence and the GFP coding sequence. The resulting 556 vector was transformed into Agrobacterium tumefaciens C58C51 and used for floral 557 dip transformation of Arabidopsis Col-0 plants. Transgenic plants were selected on 558 hygromycin and GFP fluorescence was visualized in root cells by confocal 559 microscopy. Prior to observation, root cell mitochondria were labeled by soaking the 560 samples for 5 minutes in a solution containing 0.1 µM MitoTracker® Red (Molecular 561 probes). 562 563 Protein extraction and immunoblot analysis 564 Mitochondria and chloroplasts were isolated from Arabidopsis flower buds and young 565 leaves respectively using protocols described in (Uyttewaal et al., 2008a). Total and 566 organelle protein samples were extracted in buffer containing 30 mM Hepes/KOH, 567 pH 7.7, 10 mM Mg-acetate, 150 mM K-acetate, 10% [v/v] glycerol, 0.5% [w/v] 568 CHAPS and separated by SDS-PAGE. After electrophoresis, proteins on gels were 569 transferred onto a polyvinylidene difluoride (PVDF) membrane (Perkin-Elmer) and 570 incubated with specific polyclonal antibodies recognizing either Nad9 (diluted 571 1:100,000 (Lamattina et al., 1993)), FDH (diluted 1:2,000 (Colas des Francs-Small et 572 al., 1993), ATPC (diluted 1:5,000; a gift of Jorg Meurer, Ludwig-Maximilians-573 Universität, Germany), Nad7 (diluted 1:2,000 (Pineau et al., 2008)), AOX (diluted 574 1:50; (Elthon et al., 1989)) and PORIN (diluted 1:10,000; a gift of D. Day, University 575
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

23
of Western Australia, Australia). Following hybridization of peroxidase-conjugated 576 secondary antibodies, signals were detected using Western Lightning ECL plus 577 reagents (Perking Elmer) and visualized by a LAS4000 chemiluminescence analyzer 578 (Fuji Film). The apparent molecular masses of the proteins were estimated with pre-579 stained molecular mass markers (Fermentas). Analysis of the sub-mitochondrial 580 distribution of MTL1 protein was performed as described in (Uyttewaal et al., 2008a). 581 582 BN-PAGE and complex I activity assay 583 The equivalent of 500 µg of total mitochondrial proteins were solubilized in buffer 584 containing 50 mM bis-Tris/HCl pH 7, 750 mM 6-aminohexanoic acid, 0.5 mM 585 EDTA, 5 g/g of mitochondrial protein of digitonin. Samples were electrophoresed in a 586 4 to 13% [w/v] Nupage Bis/Tris native gel (Life Technologies) according to the 587 manufacturer’s instructions. Following electrophoresis, the gels were either stained 588 with Coomassie Blue or incubated in a buffer containing 0.1 M Tris-HCl pH 7.4, 0.2 589 mM NADH, 0.2 % [w/v] nitroblue tetrazolium to reveal the NADH dehydrogenase 590 activity of complex I. A fixing solution containing 30% [v/v] methanol, 10% [v/v] 591 acetic acid was then used to stop the reaction. 592 593 RNA Extraction and Analysis 594 Total RNA was isolated using TRizol reagent (Life Technologies) following the 595 manufacturer’s recommendations and treated with RNase-free DNAse I (RNeasy 596 Mini Kit, Qiagen). For RT-PCR experiment, approximately 500 ng of total RNA were 597 reverse transcribed using random hexamers (Eurofins) and 400 units of reverse 598 transcriptase (Thermo-Scientific) in a final volume of 20 µL. For PCR amplifications, 599 2 µL of a 50-fold dilution of the cDNA solution were used as template. Amplification 600 of the MTL1 gene transcript was performed using GWMTL1-1 and GWMTL1-6 601 primers. PCR on the BIO2 gene transcript was used as a control using the BIO2-6 and 602 BIO2-8 primers. Quantitative RT-PCR analyses of mitochondrial transcripts 603 abundance and splicing efficiency were performed as in described in Haïli et al., 604 2013. For Northern-blot analysis, 15 µg of RNA was loaded on formaldehyde agarose 605 gels and transferred onto a nylon membrane (Genescreen). RNA integrity, loading 606 and transfer were verified by staining the membrane with methylene blue. 607 Hybridizations with phosporus-32 radiolabeled gene-specific probes were performed 608 under high-stringency conditions overnight at 65°C in Church Buffer (7% [w/v] SDS, 609
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

24
0.25 M Na2HPO4, pH 7.4, 2 mM EDTA, 200 µg mL-1 heparin). Blots were washed 610 and hybridized transcripts were detected and analyzed using a phosphorimager screen 611 (Fla5000-Fuji). Primers used to generate gene-specific probes are listed in 612 Supplementary Table S1. 613 Circular RT-PCR analysis was performed as in Haïli et al., 2013. Primer pairs used to 614 perform the different cRT-PCR on complex I genes are listed in Supplementary Table 615 S1. 616 617 Polysome association analysis 618 Polysomes were prepared as previously described (Barkan, 1993) except that they 619 were isolated from flowers buds and not from leaf tissue. Ten fractions of 900 µL 620 starting from the top of the gradients were collected. RNAs from each fraction were 621 extracted using TRizol reagent (Life Technologies) following the manufacturer’s 622 recommendations, precipitated with ethanol and resuspended in 30 µL of RNAse-free 623 water. 10 µL of each RNA fraction were fractionated on 1.5% [w/v] formaldehyde 624 agarose gels, transferred onto a nylon membrane (Genescreen) and hybridized with 625 gene-specific probes as described above. 626 627 Gas exchange measurements 628 Respiratory O2 consumption was measured as in Haïli et al., 2013. 629 630 SUPPLEMENTAL MATERIAL 631 632 The following materials are available in the online version of this article. 633 634 Supplemental Figure S1. Phenotypes of mtl1 mutant plants in comparison to wild 635 type. 636 Supplemental Figure S2. An MTL1-MYC translational fusion can functionally 637 complement mtl1 mutants. 638 Supplemental Figure S3. Respiratory activity measurements in wild-type (Col-0) 639 and mtl1 plants. 640 Supplemental Figure S4. RNA gel blot analysis determining the steady state levels 641 of mitochondria-encoded complex I mRNAs in the wild type and mtl1 mutants. 642 Supplemental Figure S5. Quantitative analysis of intron splicing and mature mRNA 643 accumulation in mtl1 mutants. 644
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

25
Supplemental Figure S6. Circular RT-PCR analysis of mitochondrial complex I 645 genes (nad) in the wild type (Col-0) and mtl1 mutants. 646 Supplemental Figure S7. Comparative analysis of mtl1 and bir6 mutants. 647 Supplemental Figure S8. The MTL1 protein co-sediments with mitochondrial 648 polysomes on sucrose density gradient. 649 Supplemental Figure S9. Quantitation of polysome sedimentation hybridization 650 results. 651 Supplemental Table S1. DNA primers used in this analysis. 652 653 654 LEGENDS TO FIGURES 655 656 Figure 1. Arabidopsis mtl1 mutants are delayed in their development. A, Comparative 657 vegetative phenotypes of mtl1 and Col-0 plants. Homozygous mtl1 mutants grow 658 much slower than wild-type plants on soil, and produce plants with twisted rosette 659 leaves. The photography was taken after 10 weeks of culture in long day conditions. 660 B, Schematic diagram showing the predicted domain structure of the Arabidopsis 661 MTL1 protein. Thick arrows along the protein indicate PPR-P repeats and the black 662 rectangle at the N-terminus depicts the mitochondrial targeting sequence. Thin arrows 663 above and below the protein indicate locations of T-DNA insertions harbored in mtl1-664 1 and mtl1-2 mutants. C, Reverse-Transcription PCR analysis of MTL1 transcripts in 665 Col-0 and mtl1 mutants. Five hundred ng of total RNA from plants of the indicated 666 genotypes were reverse transcribed with random primers. Resulting cDNAs were 667 PCR amplified with oligonucleotides bracketing T-DNA insertion sites and the 668 resulting amplification products were size-fractionated by agarose gel electrophoresis. 669 The BIO2 cDNA was separately amplified to control the efficiency of the reverse 670 transcription reaction. Reverse transcriptase was either added (RT+) or omitted (RT-) 671 from the initial reverse transcription reaction. 672 673 Figure 2. Subcellular and sub-mitochondrial localization of the MTL1 protein. A, 674 Confocal microscope images showing the cellular distribution of an MTL1-GFP 675 fusion in Arabidopsis root cells. The MTL1-GFP fusion comprising the first 109 676 amino acids of MTL1 and the GFP was stably transformed into Arabidopsis plants. 677 Prior to observation, roots were briefly soaked in MitoTracker® Red to label 678 mitochondria. The green channel on the left shows the GFP fluorescence; the panel in 679
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

26
the center shows mitochondria labeled with the MitoTracker® Red dye; the panel on 680 the right presents the merged signals. B, Immunoblot analysis of total (Tot), 681 chloroplast (Cp) and mitochondrial (Mt) protein fractions prepared from Arabidopsis 682 plants expressing an MTL1-MYC fusion. The protein preparations were probed with 683 an anti-MYC (α MYC) monoclonal antibody to verify the mitochondrial localization 684 of MTL1. The purity of the different fractions was tested with antibodies directed 685 against the mitochondrial Nad9 (α Nad9) and the chloroplast ATPC (α ATPC) 686 proteins. C, Assessment of proteins present in total mitochondrial protein preparation 687 (TotM), in soluble (Sol) as well as membrane (Mb)-derived protein fractions prepared 688 from plants expressing an MTL1-MYC fusion. Protein gel blots were probed with an 689 anti-MYC monoclonal antibody to detect the MTL1-MYC fusion, with anti-Nad9 690 antibodies (corresponding to an extrinsic protein of the inner membrane of 691 mitochondria), and anti-formate dehydrogenase (α FDH) antibodies (a known soluble 692 matrix protein). Each lane was loaded with about 50 µg of protein preparations. 693 694 Figure 3. mtl1 mutants are complex I respiratory mutants. A, Blue native gel analysis 695 of mitochondrial respiratory complexes extracted from wild-type and mtl1 mutant 696 plants. The gel presented on the left was stained with Coomassie blue whereas the gel 697 on the right was stained to reveal the NADH dehydrogenase activity of complex I. 698 About 200 µg of protein treated with digitonin were loaded per lane. I, III, V indicate 699 the positions of the respective respiratory chain complexes. I+III corresponds to the 700 I+III super-complex. Location of native molecular markers is indicated. B, 701 Quantitative RT-PCR measuring the relative steady-state level of alternative NADH 702 dehydrogenase (NDA and NDB) and alternative oxidase (AOX) transcripts in mtl1-1 703 and mtl1-2 mutants. C, Analysis of the accumulation level of the mitochondrial 704 alternative oxidase (AOX) in both wild-type and mtl1 plants. Total mitochondrial 705 protein preparations obtained from plants of the indicated genotypes were probed with 706 anti-AOX antibodies (α AOX). Fifty µg of proteins were loaded in each lane. 707 Coomassie blue staining of part of the gel is shown below the blots to attest for 708 equivalent loading of the different samples. 709 710 Figure 4. The Nad7 mitochondrial protein does not accumulate in mtl1 mutants. A, 711 Analysis of nad7 and nad9 mRNA abundance in wild-type and mtl1 plants. Fifteen 712
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

27
micrograms of total RNA were size fractionated on denaturing agarose gels, blotted 713 and analyzed by hybridization to radiolabeled DNA probes corresponding to nad7 and 714 nad9 mRNA. Signals corresponding to the expected mature transcripts are indicated 715 by (M) and precursors by (P). (*) points to unexplained hybridization signals that may 716 not be directly related to the nad9 mitochondrial locus. Ethidium bromide staining of 717 rRNAs is shown below the blots and serves as loading control. B, Detection of both 718 Nad7 and Nad9 proteins in mitochondrial extracts of mtl1 mutants. Mitochondrial 719 proteins prepared from plants of the indicated genotypes were analyzed by 720 immunoblot assay. The blots were probed with antisera to Nad7 (α Nad7) and Nad9 721 (α Nad9) mitochondrial complex I subunits and to PORIN (α PORIN) used as 722 loading control. 723 724 Figure 5. The nad7 mature mRNA does not sediment with mitochondrial polysomes 725 in mtl1-1 mutant. Flower bud RNA extracts prepared from Col-0 and mtl1-1 plants 726 were fractionated in 15-55% sucrose density gradients by ultracentrifugation and 727 under conditions maintaining polysome integrity. Nine fractions of equal volume 728 were collected (2 to 10 as fraction 1, devoid of RNA, was discarded). Equal aliquots 729 of each fraction were analyzed by RNA gel blot analysis and hybridized to DNA 730 probes detecting each one of the mitochondria-encoded complex I genes (nad1 to 731 nad9). A representative blot is shown for each mRNA and each genotype. (M) 732 indicates the band corresponding to the mature transcript size for each gene. 733 734 Table S1. DNA primers used in this analysis. 735 736 Figure S1. Phenotypes of mtl1 mutant plants in comparison to the wild type. A, 737 Maximum height reached by Col-0 and mtl1 adult plants. B, Number of days after 738 sowing that is necessary to see the emergence of the first flower. C, Germination 739 efficiency measured in vitro after stratification. Means±SE (n=3). 740 741 Figure S2. An MTL1-MYC translational fusion can functionally complement the mtl1 742 mutants. A, Comparative photography showing a wild-type plant (Col-0), an mtl1-1 743 mutant and an mtl1-1 mutant expressing an MTL1-MYC translational fusion. The 744 wild-type phenotype exhibited by the mutant expressing the fusion attest for the 745 functionality of the MTL1-MYC construct. Shown plants were cultivated for 8 weeks 746
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

28
in long day conditions. B, Mitochondrial protein preparations were obtained from the 747 plants of the indicated genotypes and probed with an anti-Nad7 antibody. The 748 recovery of Nad7 protein synthesis in the mtl1-1 plant transformed with the MTL1-749 MYC construct attests for the function of the MTL1-MYC translational fusion. 750 751 Figure S3. Respiratory activity measurements in wild-type (Col-0) and mtl1 plants. 752 Oxygen consumptions by rosette leaf fragments detached from plants of the indicated 753 genotypes were measured using a Clark electrode in the dark. Means±SE (n=3). FW, 754 fresh weight. 755 756 Figure S4. RNA gel blot analysis determining the steady state levels of mitochondria-757 encoded complex I mRNAs in the wild type and mtl1 mutants. Fifteen µg of total 758 RNA extracted from wild type (Col-0) and both mtl1 mutants were separated on 759 agarose gel and blotted. Membranes were hybridized with DNA probes corresponding 760 to the complex I genes indicated on the figure (nad genes). (M) Mature mRNA; (P) 761 Precursor mRNA. 762 763 Figure S5. Quantitative analysis of intron splicing and mature mRNA accumulation in 764 mtl1 mutants. A, Quantitative RT-PCR results measuring the splicing efficiency of 765 each mitochondrial intron in mtl1 mutants. The histogram shows log2 ratios of spliced 766 to unspliced forms in the mutants as compared with the Col-0 control. B, Quantitative 767 RT-PCR results measuring the accumulation levels of mature mRNAs in mtl1 768 mutants. Only transcripts that undergo splicing were considered. Log2 ratios between 769 mtl1 mutants and wild-type are shown (after normalization to 18S nuclear rRNA). 770 Three technical replicates and three biological replicates were used per genotype; 771 standard errors are indicated. 772 773 Figure S6. Circular RT-PCR analysis of mitochondrial complex I genes (nad) in the 774 wild type (Col-0) and mtl1 mutants. Complementary DNAs was produced after 775 circularization of RNAs extracted from the plants of the indicated genotype and 776 subsequently amplified with divergent primers located near the extremities of the 777 mentioned nad genes. Amplification products were run on agarose gel. The major 778 bands for each gene was cloned and sequenced from Col-0. In the case of nad7, the 779 major PCR product was cloned and sequenced from both Col-0 and mtl1 mutants. The 780
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

29
5’ and 3’ termini of each transcript are indicated relatively to the translation initiation 781 and termination codon respectively. (H2O) water control, (M) DNA size marker. 782 783 Figure S7. Comparative analysis of mtl1 and bir6 mutants. A, Comparative 784 photography showing the phenotypes of Col-0, mtl1-1, mtl1-2 and bir6 plants after 12 785 weeks on soil. B, RNA gel blot analysis determining the steady state levels of nad7 786 mature mRNA in mtl1 and bir6 mutants in comparison to the wild type. (M) Mature 787 mRNA; (P) Precursor mRNA. C, Quantitative RT-PCR measuring the accumulation 788 levels of spliced and unspliced forms of nad7 in Col-0, mtl1-1 and bir6 plants. 789 Quantitative data normalized to the 18S nuclear rRNA are shown. D, Comparative 790 detection of both Nad7 and Nad9 proteins in mitochondrial extracts of mtl1 and bir6 791 mutants. Mitochondrial proteins prepared from plants of the indicated genotypes were 792 analyzed by immunoblot assay. The blots were probed with antisera to Nad7 and 793 Nad9 mitochondrial complex I subunits and to PORIN used as loading control. 794 795 Figure S8. The MTL1 protein co-sediments with mitochondrial polysomes on sucrose 796 density gradient. A, Agarose gels stained with ethidium bromide showing RNA 797 fractions recovered from a representative polysome fragmentation experiments. 798 Flower bud RNA extracts were fractionated in continuous sucrose density gradients 799 by ultracentrifugation under conditions either maintaining (MgCl2) or disrupting 800 (EDTA) polysome integrity. Only aliquots of fractions 2 to 10 are shown. The gels 801 reveal that polysomal RNAs migrate towards the center and the bottom of the 802 gradients, whereas free mRNAs accumulate in the upper fractions. B, Immunoblot 803 analysis detecting the MTL1-MYC protein in polysome fractions prepared from the 804 mtl1-1 complemented mutant in the presence of MgCl2. The hybridized membrane 805 was subsequently stained with Coomassie blue to monitor the protein repartition 806 along the gradient (Coomassie). 807 808 Figure S9. Quantitation of polysome sedimentation hybridization results. 809 Hybridization signals corresponding to mature mRNAs in each fraction (see Fig. 5) 810 for the indicated gene were determined using the ImageQuant software (GE 811 Healthcare Life Sciences) in both Col-0 and mtl1-1 mutants. For each gene and 812 genotype, the hybridization signals quantified in each fractions were cumulated and 813
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

30
the percentage of this total signal to which contributes each fraction was calculated 814 and is shown on the presented graphs. 815 816 ACKNOWLEDGMENTS 817 818 We are thankful to Guillaume Tcherkez for technical assistance with respiration 819 measurements and to Françoise Budar for critical reading of the manuscript. The 820 Institut Jean-Pierre Bourgin benefits from the support of the LabEx Saclay Plant 821 Sciences-SPS (ANR-10-LABX-0040-SPS). 822 823 824
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

Parsed CitationsBan T, Ke J, Chen R, Gu X, Tan MHE, Zhou XE, Kang Y, Melcher K, Zhu JK, Xu HE (2013) Structure of a PLS-classPentatricopeptide Repeat Protein Provides Insights into Mechanism of RNA Recognition. J Biol Chem 288: 31540-31548
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Barkan A (1993) Nuclear Mutants of Maize with Defects in Chloroplast Polysome Assembly Have Altered Chloroplast RNAMetabolism. Plant Cell 5: 389-402
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Barkan A (2011) Expression of plastid genes: organelle-specific elaborations on a prokaryotic scaffold. Plant Physiol 155: 1520-1532Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Barkan A, Rojas M, Fujii S, Yap A, Chong YS, Bond CS, Small I (2012) A combinatorial amino Acid code for RNA recognition bypentatricopeptide repeat proteins. PLoS Genet 8: e1002910
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Barkan A, Small I (2013) Pentatricopeptide Repeat Proteins in Plants. Annu Rev Plant Biol 65: 415-442Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Barkan A, Walker M, Nolasco M, Johnson D (1994) A nuclear mutation in maize blocks the processing and translation of severalchloroplast mRNAs and provides evidence for the differential translation of alternative mRNA forms. EMBO J 13: 3170-3181
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Beick S, Schmitz-Linneweber C, Williams-Carrier R, Jensen B, Barkan A (2008) The pentatricopeptide repeat protein PPR5stabilizes a specific tRNA precursor in maize chloroplasts. Mol Cell Biol 28: 5337-5347
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bonen L (2004) Translational machinery in plant organelles. Pp. 323-345 in H. Daniell and C. Chase, eds. Molecular biol- ogy andbiotechnology of plant organelles. Springer, the Netherlands.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bonen L (2008) Cis- and trans-splicing of group II introns in plant mitochondria. Mitochondrion 8: 26-34Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bonen L, Calixte S (2005) Comparative Analysis of Bacterial-Origin Genes for Plant Mitochondrial Ribosomal Proteins. Mol BiolEvol 23: 701-712
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Cai W, Okuda K, Peng L, Shikanai T (2011) PROTON GRADIENT REGULATION 3 recognizes multiple targets with limited similarityand mediates translation and RNA stabilization in plastids. Plant J 67: 318-327
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Choi B, Acero MM, Bonen L (2012) Mapping of wheat mitochondrial mRNA termini and comparison with breakpoints in DNAhomology among plants. Plant Mol Biol 80: 539-552
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Cohen S, Zmudjak M, Colas des Francs-Small C, Malik S, Shaya F, Keren I, Belausov E, Many Y, Brown GG, Small I, et al (2014)nMAT4, a maturase factor required for nad1pre-mRNA processing and maturation, is essential for holocomplex I biogenesis inArabidopsis mitochondria. Plant J 78: 253-268
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Colas des Francs-Small C, Falcon de Longevialle A, Li Y, Lowe E, Tanz SK, Smith C, Bevan MW, Small I (2014) ThePentatricopeptide Repeat Proteins TANG2 and ORGANELLE TRANSCRIPT PROCESSING439 Are Involved in the Splicing of the www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from
Copyright © 2015 American Society of Plant Biologists. All rights reserved.

Multipartite nad5 Transcript Encoding a Subunit of Mitochondrial Complex I. Plant Physiol 165: 1409-1416Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Colas des Francs-Small, C., Ambard-Bretteville, F., Small, I.D. and Rémy, R. (1993) Identification of a major soluble protein inmitochondria from nonphotosynthetic tissues as NAD-dependent formate dehydrogenase. Plant Physiol 102, 1171-1177.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Coquille S, Filipovska A, Chia T, Rajappa L, Lingford JP, Razif MFM, Thore SEP, Rackham O (2014) An artificial PPR scaffold forprogrammable RNA recognition. Nature Commun 5: 1-9
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Curtis MD and Grossniklaus U (2003) A Gateway Cloning Vector Set for High-Throughput Functional Analysis of Genes in Planta.Plant Physiol 133: 462-469
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Elthon TE, Nickels RL, McIntosh L (1989) Monoclonal antibodies to the alternative oxidase of higher plant mitochondria. PlantPhysiol 89: 1311-1317
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fisk DG, Walker MB, Barkan A (1999) Molecular cloning of the maize gene crp1 reveals similarity between regulators ofmitochondrial and chloroplast gene expression. EMBO J 18: 2621-2630
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Colas des Francs-Small C, Ambard-Bretteville F, Darpas A, Sallantin M, Huet JC, Pernollet JC, Rémy R (1993) Variation of thepolypeptide composition of mitochondria isolated from different potato tissues. Plant Physiol 98: 273-278
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Giegé P, Brennicke A (2001) From gene to protein in higher plant mitochondria. C R Acad Sci III, Sci Vie 324: 209-217Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gully BS, Shah KR, Lee M, Shearston K, Smith NM, Sadowska A, Blythe AJ, Bernath-Levin K, Stanley WA, Small ID, et al (2015) Thedesign and structural characterization of a synthetic pentatricopeptide repeat protein Acta Crystallogr D Biol Crystallogr 71:196-208
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gutierres S, Sabar M, Lelandais C, Chetrit P, Diolez P, Degand H, Boutry M, Vedel F, de Kouchkovsky Y, De Paepe R (1997) Lackof mitochondrial and nuclear-encoded subunits of complex I and alteration of the respiratory chain in Nicotiana sylvestrismitochondrial deletion mutants. Proc Natl Acad Sci USA 94: 3436-3441
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Haïli N, Arnal N, Quadrado M, Amiar S, Tcherkez G, Dahan J, Briozzo P, Colas des Francs-Small C, Vrielynck N, Mireau H (2013)The pentatricopeptide repeat MTSF1 protein stabilizes the nad4 mRNA in Arabidopsis mitochondria. Nucleic Acids Res 41: 6650-6663
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hammani K, Colas des Francs-Small C, Takenaka M, Tanz SK, Okuda K, Shikanai T, Brennicke A, Small I (2011) ThePentatricopeptide Repeat Protein OTP87 Is Essential for RNA Editing of nad7 and atp1 Transcripts in Arabidopsis Mitochondria. JBiol Chem 286: 21361-21371
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hammani K, Giegé P (2014) RNA metabolism in plant mitochondria. Trends Plant Sci 19: 380-389Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hazle T, Bonen L (2007) Comparative analysis of sequences preceding protein-coding mitochondrial genes in flowering plants. www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from
Copyright © 2015 American Society of Plant Biologists. All rights reserved.

Mol Biol Evol 24: 1101-1112Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Howard MJ, Lim WH, Fierke CA, Koutmos M (2012) Mitochondrial ribonuclease P structure provides insight into the evolution ofcatalytic strategies for precursor-tRNA 5' processing. Proc Natl Acad Sci USA 109: 16149-16154
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hsieh W-Y, Liao J-C, Chang C-Y, Harrison T, Boucher C, Hsieh M-H (2015) The SLOW GROWTH3 Pentatricopeptide RepeatProtein Is Required for the Splicing of Mitochondrial NADH Dehydrogenase Subunit7Intron 2 in Arabidopsis. Plant Physiol 168:490-501
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hsu Y-W, Wang H-J, Hsieh M-H, Hsieh H-L, Jauh G-Y (2014) Arabidopsis mTERF15 Is Required for Mitochondrial nad2 Intron 3Splicing and Functional Complex I Activity. PLoS ONE 9: e112360
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kazama T, Nakamura T, Watanabe M, Sugita M, Toriyama K (2008) Suppression mechanism of mitochondrial ORF79 accumulationby Rf1 protein in BT-type cytoplasmic male sterile rice. Plant J 55: 619-628
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ke J, Chen R-Z, Ban T, Zhou XE, Gu X, Tan MHE, Chen C, Kang Y, Brunzelle JS, Zhu J-K, et al (2013) Structural basis for RNArecognition by a dimeric PPR-protein complex. Nat Struct Mol Biol 20: 1377-1382
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Keren I, Bezawork-Geleta A, Kolton M, Maayan I, Belausov E, Levy M, Mett A, Gidoni D, Shaya F, Ostersetzer-Biran O (2009)AtnMat2, a nuclear-encoded maturase required for splicing of group-II introns in Arabidopsis mitochondria. RNA 15: 2299-2311
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Keren I, Tal L, Colas des Francs-Small C, Araújo WL, Shevtsov S, Shaya F, Fernie AR, Small I, Ostersetzer-Biran O (2012) nMAT1, anuclear-encoded maturase involved in the trans-splicing of nad1 intron 1, is essential for mitochondrial complex I assembly andfunction. Plant J 71: 413-426
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Khrouchtchova A, Monde RA, Barkan A (2012) A short PPR protein required for the splicing of specific group II introns inangiosperm chloroplasts. RNA 18: 1197-1209
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Koprivova A, Francs-Small des CC, Calder G, Mugford ST, Tanz S, Lee B-R, Zechmann B, Small I, Kopriva S (2010) Identification ofa pentatricopeptide repeat protein implicated in splicing of intron 1 of mitochondrial nad7 transcripts. J Biol Chem 285: 32192-32199
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kühn K, Carrie C, Giraud E, Wang Y, Meyer EH, Narsai R, Francs-Small des CC, Zhang B, Murcha MW, Whelan J (2011) The RCC1family protein RUG3 is required for splicing of nad2 and complex I biogenesis in mitochondria of Arabidopsis thaliana. Plant J 67:1067-80.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lamattina L, Gonzalez D, Gualberto J, Grienenberger JM (1993) Higher plant mitochondria encode an homologue of the nuclear-encoded 30-kDa subunit of bovine mitochondrial complex I. Eur J Biochem 217: 831-838
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lurin C, Andrés C, Aubourg S, Bellaoui M, Bitton F, Bruyère C, Caboche M, Debast C, Gualberto J, Hoffmann B, et al (2004)Genome-wide analysis of Arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. PlantCell 16: 2089-2103
Pubmed: Author and TitleCrossRef: Author and Title www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from
Copyright © 2015 American Society of Plant Biologists. All rights reserved.

Google Scholar: Author Only Title Only Author and Title
Manavski N, Guyon V, Meurer J, Wienand U, Brettschneider R (2012) An Essential Pentatricopeptide Repeat Protein Facilitates 5'Maturation and Translation Initiation of rps3 mRNA in Maize Mitochondria. Plant Cell 24: 3087-3105
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nakagawa T, Kurose T, Hino T, Tanaka K, Kawamukai M, Niwa Y, Toyooka K, Matsuoka K, Jinbo T, Kimura T (2007) Development ofseries of gateway binary vectors, pGWBs, for realizing efficient construction of fusion genes for plant transformation. J BiosciBioeng 104: 34-41
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
O'Toole N, Hattori M, Andrés C, Iida K, Lurin C, Schmitz-Linneweber C, Sugita M, Small I (2008) On the expansion of thepentatricopeptide repeat gene family in plants. Mol Biol Evol 25: 1120-1128
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ostersetzer-Biran O (2014) Group II intron splicing factors in plant mitochondria. Front Plant Sci 5:35Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pfalz J, Bayraktar OA, Prikryl J, Barkan A (2009) Site-specific binding of a PPR protein defines and stabilizes 5' and 3' mRNA terminiin chloroplasts. EMBO J 28:2042-52
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pineau B, Layoune O, Danon A, De Paepe R (2008) L-galactono-1,4-lactone dehydrogenase is required for the accumulation ofplant respiratory complex I. J Biol Chem 283: 32500-32505
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Prikryl J, Rojas M, Schuster G, Barkan A (2011) Mechanism of RNA stabilization and translational activation by a pentatricopeptiderepeat protein. Proc Natl Acad Sci USA 108: 415-420
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pyle AM, Fedorova O, Waldsich C (2007) Folding of group II introns: a model system for large, multidomain RNAs? Trends BiochemSci 32: 138-145
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ringel R, Sologub M, Morozov YI, Litonin D, Cramer P, Temiakov D (2011) Structure of human mitochondrial RNA polymerase.Nature 478: 269-273
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Schmitz-Linneweber C, Williams-Carrier R, Barkan A (2005) RNA immunoprecipitation and microarray analysis show a chloroplastPentatricopeptide repeat protein to be associated with the 5' region of mRNAs whose translation it activates. Plant Cell 17: 2791-2804
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Small ID, Peeters N (2000) The PPR motif - a TPR-related motif prevalent in plant organellar proteins. Trends Biochem Sci 25: 46-47
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sørensen MA, Fricke J, Pedersen S (1998) Ribosomal protein S1 is required for translation of most, if not all, natural mRNAs inEscherichia coli in vivo. J Mol Biol 280: 561-569
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Takenaka M, Zehrmann A, Brennicke A, Graichen K (2013) Improved Computational Target Site Prediction for PentatricopeptideRepeat RNA Editing Factors. PLoS ONE 8: e65343
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

Uyttewaal M, Arnal N, Quadrado M, Martin-Canadell A, Vrielynck N, Hiard S, Gherbi H, Bendahmane A, Budar F, Mireau H (2008a)Characterization of Raphanus sativus Pentatricopeptide Repeat Proteins Encoded by the Fertility Restorer Locus for OguraCytoplasmic Male Sterility. Plant Cell 20: 3331-3345
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Uyttewaal M, Mireau H, Rurek M, Hammani K, Arnal N, Quadrado M, Giegé P (2008b) PPR336 is Associated with Polysomes in PlantMitochondria. J Mol Biol 375: 626-636
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Williams-Carrier R, Kroeger T, Barkan A (2008) Sequence-specific binding of a chloroplast pentatricopeptide repeat protein to itsnative group II intron ligand. RNA 14: 1930-1941
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yagi Y, Hayashi S, Kobayashi K, Hirayama T, Nakamura T (2013) Elucidation of the RNA Recognition Code for PentatricopeptideRepeat Proteins Involved in Organelle RNA Editing in Plants. PLoS ONE 8: e57286
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yap A, Kindgren P, Colas des Francs-Small C, Kazama T, Tanz SK, Toriyama K and Small I (2015) AEF1/MPR25 is implicated in RNAediting of plastid atpF and mitochondrial nad5, and also promotes atpF splicing in Arabidopsis and rice. Plant J, 81: 661-669
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yamazaki H, Tasaka M, Shikanai T (2004) PPR motifs of the nucleus-encoded factor, PGR3, function in the selective and distinctsteps of chloroplast gene expression in Arabidopsis. Plant J 38: 152-163
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yin P, Li Q, Yan C, Liu Y, Liu J, Yu F, Wang Z, Long J, He J, Wang H-W, et al (2013) Structural basis for the modular recognition ofsingle-stranded RNA by PPR proteins. Nature 5: 168-171
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhelyazkova P, Hammani K, Rojas M, Voelker R, Vargas-Suarez M, Borner T, Barkan A (2011) Protein-mediated protection as thepredominant mechanism for defining processed mRNA termini in land plant chloroplasts. Nucleic Acids Res 40: 3092-3105
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zmudjak M, Colas des Francs-Small C, Keren I, Shaya F, Belausov E, Small I, Ostersetzer-Biran O (2013) mCSF1, a nucleus-encoded CRM protein required for the processing of many mitochondrial introns, is involved in the biogenesis of respiratorycomplexes I and IV in Arabidopsis. New Phytol 199: 379-394
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zoschke R, Kroeger T, Belcher S, Schöttler MA, Barkan A, Schmitz-Linneweber C (2012a) The pentatricopeptide repeat-SMRprotein ATP4 promotes translation of the chloroplast atpB/E mRNA. Plant J 72: 547-558
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zoschke R, Qu Y, Zubo YO, Börner T, Schmitz-Linneweber C (2012b) Mutation of the pentatricopeptide repeat-SMR protein SVR7impairs accumulation and translation of chloroplast ATP synthase subunits in Arabidopsis thaliana. J Plant Res. 126:403-14
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon February 12, 2019 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.





























