Embed Size (px)
Citation preview

Biochimica et Biophysics Acta. 752 (1983) 178- 18 1 Elsevier
178
BBA Report
BBA 50042
ROUTE OF INCORPORATION OF ALVEOLAR PALMITATE AND CHOLINE INTO SURFACTANT PHOSPHATIDYLCHOLINE IN RABBITS
HARRIS JACOBS, ALAN JOBE, MACHIKO IKEGAMI, SALLY JONES and DEBORAH MILLER
Fetal- Maternal Research Laboratories, Department of Pediatrics, Harbor -UCLA Medical Center, Torrance, CA 90509 (U.S.A.)
(Received December 15 th, 1982)
Key worak Lung surjactani; Phosphatidylcholine; Precursor incorporation; (Rabbit)
Intratracheal injection of 3-day-old rabbits with radioactively labeled palmitic acid and choline results in an S-lo-fold increase in the efficiency of their incorporation into surfactant phosphatidylcholine when compared
to the intravenous injection of these precursors. Based on labeling patterns in microsomal, lamellar body and
alveolar wash fractions, the incorporation appears to be via normal surfactant synthetic pathways. In- tratracheal injection of phospholipid precursors is useful for producing relatively high specific activity natural
surfactant.
Surfactant is a complex mixture of lipids and protein which is synthesized by pulmonary type 11 pneumocytes and packaged into lamellar bodies
for secretion to the alveolar surface [ 1,2]. Most of
the phospholipid of surfactant is phosphati-
dylcholine (PC) [3]. One approach to the study of
surfactant metabolism has been to inject animals
intravenously with radioactively labeled precursors
of PC. The radioactively labeled PC can then be
followed through lung subcellular fractions to the
alveolar space by autoradiography [4,5] or by iso-
lating subcellular fractions and measuring PC specific activities [1,2,6-g]. Another approach has
been to administer a solution of radioactively
labeled natural or synthetic surfactant directly into
the airways and again use autoradiography, specific activities or total count recovery to study metabo- lism [9-131. The present experiment was designed to compare the pattern of incorporation of radio- actively labeled palmitate and choline into surfac- tant PC by intratracheal injection versus that after administration by intravenous injection.
Abbreviation: PC, phosphatidylcholine.
0005-2760/83/$03.00 0 1983 Elsevier Science Publishers B.V.
Three injection solutions were prepared with
[ “C]choline (50 Ci/mol) and 9,10-[3H]palmitic
acid 918 Ci/mmol) that was complexed to al-
bumin [ 1,6]. Solutions 1 and 2 contained 8.23 PCi
of [‘4C]choline/ml and 137 PCi of [3H]palmitic
acid/ml in a 1 : 1 mixture of Lactated Ringers Injection (Travanol Lab., Inc.) and distilled water.
Unlabeled natural rabbit surfactant was isolated
as previously described [ 131 and was added to
solution 1 to a final concentration of 0.75 pm01 of surfactant PC/ml of solution. No surfactant was
added to solution 2. Solution 3 was made with 0.9% saline and contained 7.5 PCi of [14C]-. choline/ml and 125 PCi of [ 3H]palmitic acid/ml.
44 3-day-old New Zealand White rabbits were
taken from their litters on the day of injection. 12 rabbits were injected intratracheally with solution 1 and 12 other rabbits were injected intratrache- ally with solution 2. The trachea of each rabbit was isolated and 0.35 ml/ 100 g body weight was injected with a 30 gauge needle [ 131. This provided 2.88 pCi of [‘4C]choline/100 g body weight and 48 PCi of [ 3H]palmitic acid/ 100 g body weight. Three rabbits from each group were killed at 1, 5, 15 and 25 h after injection by an intraperitoneal
injection of pentobarbital followed by exanguina-
tion. 20 rabbits were injected via an external jugu- lar vein with 0.4 ml/ 100 g body weight of solution
3, providing 3.0 PC1 of [‘4C]choline/ 100 g body
weight and 50 pCi of [3H]palmitate/100 g body
weight. These doses were approximately the same
as those received by rabbits given solution 1 or 2.
These rabbits were killed in groups of four at 1, 5,
13, 17 and 25 h after injection. From each of the 44 rabbits we isolated the
alveolar wash, a microsomal and a lamellar body fraction. Immediately after killing, the chest of
each animal was opened, the trachea cannulated
and the lungs were thoroughly washed with saline
to yield an alveolar wash fraction [ 1,131. The lungs
of each rabbit then were homogenized in 8 ml of
0.32 m sucrose, 0.01 M Tris-HCl, 0.15 M NaCl,
0.001 M CaCl,, 0.001 M MgSO, and 0.0001 M EDTA at pH 7.4 [ 1,131, and 0.5 ml of the homo-
genate was saved. From the rest of the homo-
genate we isolated lamellar bodies and a micro- somal fraction by differential and sucrose density
gradient centrifugation [ 1,6,7,13]. The lamellar
body fraction was that fraction recovered between
0.45 and 0.55 M sucrose buffers while the micro-
somal fraction was recovered as the pellet which
sedimented through 1 M sucrose buffer at 100000
x g. For each rabbit, lipids in each fraction were
extracted [14] and PC was isolated in duplicate from each lipid extract by one-dimensional thin-
layer chromatography [1,6,13]. One spot was as-
sayed for phosphate according to the method of Bartlett [ 151 and its duplicate for radioactivity in
Aquasol- scintillation fluid (New England
Nuclear Corporation). All 3 H and I4 C counts/ min
were corrected for cross-channel contamination,
and the specific activity of PC was determined on
each fraction and expressed as counts/min per pm01 PC.
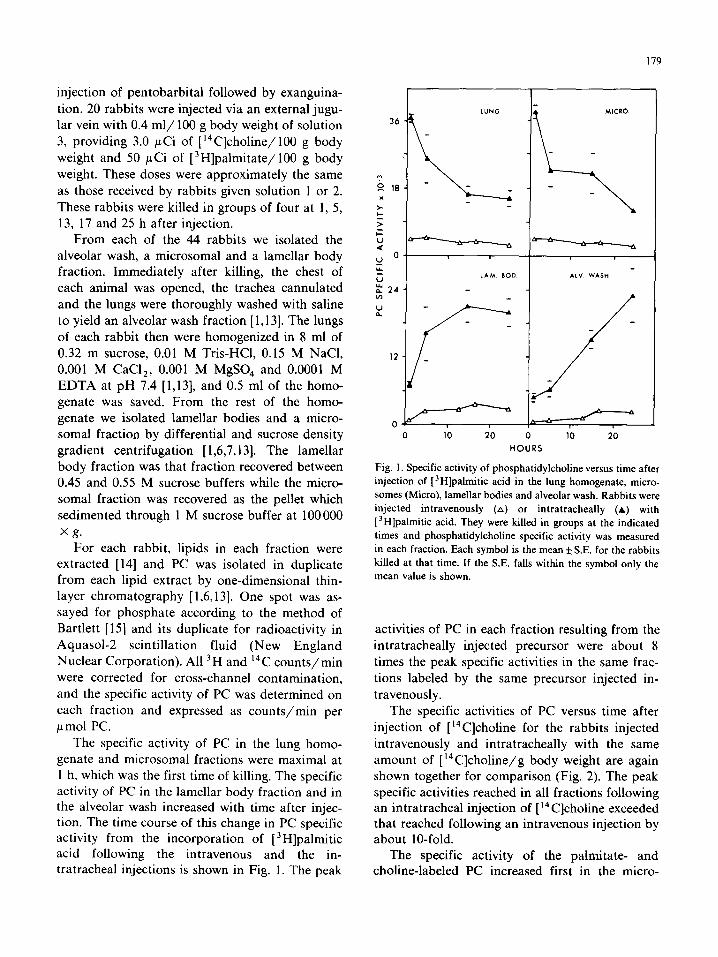
The specific activity of PC in the lung homo-
genate and microsomal fractions were maximal at 1 h, which was the first time of killing. The specific activity of PC in the lamellar body fraction and in the alveolar wash increased with time after injec- tion. The time course of this change in PC specific activity from the incorporation of [3H]palmitic acid following the intravenous and the in- tratracheal injections is shown in Fig. 1. The peak
0 0 10 20 0 10 20
HOURS
Fig. 1. Specific activity of phosphatidylcholine versus time after
injection of [ 3H]palmitic acid in the lung homogenate, micro-
somes (Micro), lamellar bodies and alveolar wash. Rabbits were
injected intravenously (A) or intratracheally (A) with
[ ‘Hlpalmitic acid. They were killed in groups at the indicated
times and phosphatidylcholine specific activity was measured
in each fraction. Each symbol is the mean f SE. for the rabbits
killed at that time. If the S.E. falls within the symbol only the
mean value is shown.
activities of PC in each fraction resulting from the
intratracheally injected precursor were about 8
times the peak specific activities in the same frac-
tions labeled by the same precursor injected in-
travenously.
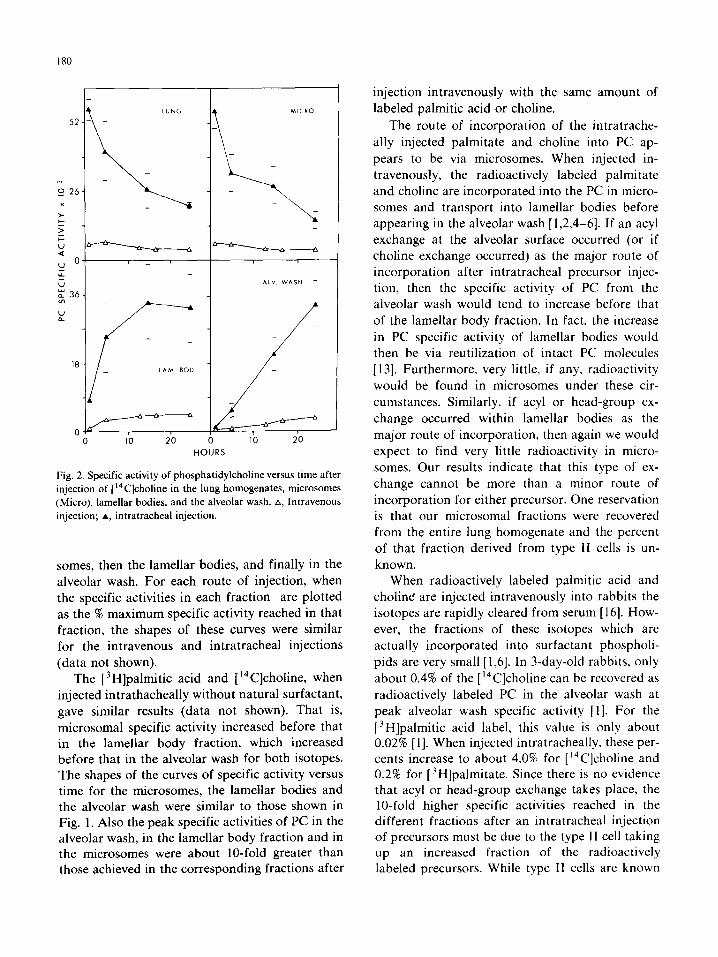
The specific activities of PC versus time after
injection of [‘4C]choline for the rabbits injected intravenously and intratracheally with the same
amount of [ “C]choline/g body weight are again shown together for comparison (Fig. 2). The peak
specific activities reached in all fractions following an intratracheal injection of [ “C]choline exceeded that reached following an intravenous injection by about IO-fold.
The specific activity of the palmitate- and choline-labeled PC increased first in the micro-
180
Ojd I& s
0 IO 20 0 10 20 HOURS
Fig. 2. Specific activity of phosphatidylcholine versus time after
injection of [ “C]choline in the lung homogenates, microsomes
(Micro), lamellar bodies. and the alveolar wash. A, Intravenous
injection; A, intratracheal injection.
somes, then the lamellar bodies, and finally in the
alveolar wash. For each route of injection, when the specific activities in each fraction are plotted
as the % maximum specific activity reached in that
fraction, the shapes of these curves were similar for the intravenous and intratracheal injections
(data not shown).
The [3H]palmitic acid and [‘4C]choline, when
injected intrathacheally without natural surfactant,
gave similar results (data not shown). That is,
microsomal specific activity increased before that in the lamellar body fraction, which increased before that in the alveolar wash for both isotopes. The shapes of the curves of specific activity versus time for the microsomes, the lamellar bodies and the alveolar wash were similar to those shown in Fig. 1. Also the peak specific activities of PC in the alveolar wash, in the lamellar body fraction and in the microsomes were about IO-fold greater than those achieved in the corresponding fractions after
injection intravenously with the same amount of
labeled palmitic acid or choline.
The route of incorporation of the intratrache-
ally injected palmitate and choline into PC ap-
pears to be via microsomes. When injected in-
travenously, the radioactively labeled palmitate and choline are incorporated into the PC in micro-
somes and transport into lamellar bodies before
appearing in the alveolar wash [ 1,2,4-61. If an acyl exchange at the alveolar surface occurred (or if
choline exchange occurred) as the major route of
incorporation after intratracheal precursor injec- tion, then the specific activity of PC from the alveolar wash would tend to increase before that
of the lamellar body fraction. In fact, the increase
in PC specific activity of lamellar bodies would
then be via reutilization of intact PC molecules [13]. Furthermore, very little, if any, radioactivity
would be found in microsomes under these cir-
cumstances. Similarly, if acyl or head-group ex-
change occurred within lamellar bodies as the
major route of incorporation, then again we would expect to find very little radioactivity in micro-
somes. Our results indicate that this type of ex-
change cannot be more than a minor route of incorporation for either precursor. One reservation
is that our microsomal fractions were recovered
from the entire lung homogenate and the percent of that fraction derived from type II cells is un-
known. When radioactively labeled palmitic acid and
choline are injected intravenously into rabbits the isotopes are rapidly cleared from serum [ 161. How-
ever, the fractions of these isotopes which are
actually incorporated into surfactant phospholi-
pids are very small [ 1,6]. In 3-day-old rabbits, only
about 0.4% of the [ “C]choline can be recovered as
radioactively labeled PC in the alveolar wash at
peak alveolar wash specific activity [l]. For the
[‘Hlpalmitic acid label, this value is only about 0.02% [ 11. When injected intratracheally, these per- cents increase to about 4.0% for [ “C]choline and 0.2% for [jH]palmitate. Since there is no evidence that acyl or head-group exchange takes place, the IO-fold higher specific activities reached in the different fractions after an intratracheal injection of precursors must be due to the type II cell taking up an increased fraction of the radioactively labeled precursors. While type II cells are known

to take up surfactant phospholipids from the al-
veolar surface [9,11- 131, phagocytosis is not gener-
ally considered to be a major property of these
cells. The surface of the cell adjacent to the base-
ment membrane is nearest to the capillary bed and
is the surface which has been assumed to transport
precursors used for surfactant synthesis. The type
II cells in this experiment were exposed to the
precursors on their alveolar surface. Nevertheless,
they were capable of internalizing both precursors and utilizing them quite effectively for surfactant
synthesis. This occurred independent of whether the precursors were mixed with surfactant prior to injection.
This lo-fold increase in alveolar wash PC
specific activity after an intratracheal injection
versus an intravenous injection is useful for pro-
duction of a naturally labeled surfactant with a high specific activity. A radioactively labeled
surfactant which has a high specific activity allows for the use of only a small quantity in in- tratracheal labeling experiments. The advantage of
this is that one can be certain of minimal perturba-
tion of endogenous pools while still administering sufficient radioactivity for the required measure- ments.
Acknowledgements
This work was supported by NIH Grant HD- 11932 from Child Health and Development, De-
181
partment of Health and Human Services, by Re-
search Career Development Award HD-HL-00252
to A.J. and NIH Research Service Award HL-
06544 to H.J.
References
1 Jacobs, H.C., Jobe, A.H., Ikegami, M. and Jones, S. (1981)
J. Biol. Chem. 257, 1805-1810
2 Young, S.L., Kremers, S.A., Apple, J. Crapo, J.D. and
Brumley, G.W. (1981) J. Appl. Physiol. 51, 248-253
3 King, R.J. (1974) Fed. Proc. 33, 2238
4 Chevalier, G. and Collet, A.J. (1972) Anat. Rec. 174,
289-310
5 Askin, F.B. and Kuhn, C. (1971) Lab. Invest. 25, 260-268
6 Jobe, A.H. (1977) Biochim. Biophys. Acta 489, 440-453
7 Jobe, A.H., Kirkpatrick, E. and Gluck, L. (1978) J. Biol.
Chem. 253, 3810-3816
8 Baritussio, A.G., Magoon, M.W., Goerke, J. and Clements,
J.A. (1981) B&him. Biophys. Acta 666, 382-393
9 Geiger, K., Gallagher, M.L. and Hedley-Whyte, J. (1975) J.
Appl. Physiol. 39 759-766
10 Scarpelh, E.M., Condorelli, S., Colacicco, G. and Cosmi,
11
12
E.V. (1975) Pediat. Res. 9, 195-201
Hallman, M., Epstein, B.L. and Gluck, L. (1981) J. Clin.
Invest. 68, 742-75 I Glatz, T.H., Ikegami, M. and Jobe, A.H. (1982) Pediat. Res.
16, 711-715
13
14
Jacobs, H.C., Jobe, A.H., Ikegami, M. and Conaway, D.J.
and Jones, S.J. (1983) J. Biol. Chem. 258, 4159-4165
Bligh, E.G. and Dyer, W.J. (1959) Can. J. B&hem. Physiol.
37,911-917
1s Bartlett, G.R. (1959) J. Biol. Chem. 234, 466-468
16 Jobe, A.H. (1979) Biochim. Biophys. Acta 574, 268-279





























