Embed Size (px)
Citation preview

477
BIOLOGY OF REPRODUCTION 62, 477–490 (2000)
Role of Transforming Growth Factor-a and the Epidermal Growth Factor Receptorin Embryonic Rat Testis Development1
Elena Levine,3 Andrea S. Cupp, Linda Miyashiro, and Michael K. Skinner2
Center for Reproductive Biology, Department of Genetics and Cell Biology, Washington State University, Pullman,Washington 99164-4231
ABSTRACT
Embryonic testis development requires the morphogenesis ofcords and growth of all cell populations to allow organ forma-tion. It is anticipated that coordination of the growth and dif-ferentiation of various cell types involves locally producedgrowth factors. The current study was an investigation of thehypothesis that transforming growth factor-a (TGF-a) is in-volved in regulating embryonic testis growth. TGF-a has previ-ously been shown to function in the postnatal testis. TGF-a andother members of the epidermal growth factor (EGF) family actthrough the epidermal growth factor receptor (EGFR) to stimu-late cell proliferation and tissue morphogenesis. To understandthe potential actions of TGF-a in the embryonic testis, generalcell proliferation was investigated. Characterization of cell pro-liferation in the rat testis throughout embryonic and postnataldevelopment indicated that each cell type has a distinct patternof proliferation. Germ cell growth was transiently suppressedaround birth. Interstitial cell growth was high embryonically anddecreased to low levels around birth. A low level of Sertoli cellproliferation was observed at the onset of testis cord formation.Sertoli cell proliferation in early embryonic development waslow; the levels were high later in embryonic development andremained high until the onset of puberty. Both TGF-a and theEGFR were shown to be expressed in the embryonic and post-natal rat and mouse testis. Perturbation of TGF-a function usingneutralizing antibodies to TGF-a on testis organ cultures dra-matically inhibited the growth of both embryonic and neonataltestis. TGF-a antibodies had no effect on cord formation. TheTGF-a antibody was found to be specific for TGF-a in Westernblots when compared to EGF and heregulin. Testis growth wasalso inhibited by perturbation of EGFR signaling using an EGFRkinase inhibitor. Therefore, TGF-a appears to influence embry-onic testis growth but not morphogenesis (i.e., cord formation).Treatment of embryonic testis organ cultures with exogenousTGF-a also perturbed development, leading to an increased pro-liferation of unorganized cells. Testis from EGFR and TGF-aknockout mice were analyzed for testis morphology. TGF-aknockout mice had no alterations in testis phenotype, whileEGFR knockout mice had a transient decrease in the relativeamount of interstitial cells before birth. Observations suggestthat there may be alternate or compensatory factors that allowtestis growth to occur in the apparent absence of TGF-a actionsin the mutant mice. In summary, the results obtained suggestthat TGF-a is an important factor in the regulation of embryonic
1The research was supported by NIH grants (HD20583, DK45889) toM.K.S. E.L. was supported by the Biochemistry Program of the GraduateProgram in Biological Sciences (PIBS) in the Department of Biochemistry,University of California at San Francisco.2Correspondence. FAX: 509-335-2176; e-mail: [email protected] address: Biological Sciences Department, California PolytechnicState University, San Louis Obispo, CA 93407.
Received: 15 April 1999.First decision: 13 May 1999.Accepted: 15 October 1999.Q 2000 by the Society for the Study of Reproduction, Inc.ISSN: 0006-3363. http://www.biolreprod.org
testis growth, but other factors will also be involved in the pro-cess.
INTRODUCTIONThe development of the testis requires coordination of
the growth and differentiation of several cell types. Thetestis is formed initially from an undifferentiated bipotentialgonad (i.e., embryonic Day [e]13 in the rat). At this stageit is indistinguishable from the ovary and possesses no mor-phological organization. The first sign of male developmentis the differentiation of precursor Sertoli cells, which thenaggregate with germ cells and undergo a transition frommesenchyme to epithelia to form cords (i.e., e13.5 to e14in the rat) [1, 2]. The testis cords remain solid until a lumenforms after the onset of puberty (i.e., postnatal Day 10 inthe rat) to convert them into tubules (i.e., postnatal Days14 to 18 in the rat). The seminiferous tubules are the siteof spermatogenesis, in which spermatogonia develop intospermatozoa in close interaction with the Sertoli cells thatmake up the walls of the tubule [3–5]. Surrounding theSertoli cells is a layer of peritubular myoid cells that areresponsible for contraction of the tubule [6–8) and regula-tion of differentiated function [9, 10]. In the interstitialspace between the tubules are Leydig cells, stromal fibro-blasts, vascular and lymphatic endothelial cells, and mac-rophages [9].
Dramatic growth occurs in the testis during embryonicand early postnatal development [11]. One of the earliestmale-specific characteristics of the testis after the formationof cords is rapid growth. This extensive growth does notoccur in the ovary [12]. The peak of Sertoli cell prolifer-ation occurs in the late embryo at e20–21 just before birth[13]. This proliferation in the embryonic and early postnatalperiod is crucial for establishing the mature adult size ofthe testis since no Sertoli cell proliferation occurs after ini-tiation of puberty [14, 15]. In addition, this proliferation isnecessary for reproductive function, since a sufficient pop-ulation of Sertoli cells is necessary for adequate sperm pro-duction in the adult [13, 16, 17]. The majority of informa-tion about testicular growth regulation has concerned post-natal growth or occasionally late embryonic growth [11,13–19]. Several growth factors have been identified in thepostnatal testis and are postulated to be involved in theregulation of puberty and spermatogenesis [9, 20, 21]. Thecurrent study was an investigation of growth in the embry-onic testis.
One family of growth factors that may play a role inembryonic testis growth is the epidermal growth factor(EGF) family. There are multiple related ligands in thisfamily, including EGF, transforming growth factor a (TGF-a), amphiregulin, heparin-binding EGF, crypto, betacellu-lin, and heregulin [22–24]. The receptors include the epi-dermal growth factor receptor (EGFR/HER1/erbB1),HER2/neu, HER3, and HER4 [25, 26]. The EGFR familyof receptors function by ligand-dependent dimerization and

478 LEVINE ET AL.
activation of the tyrosine kinase in the cytoplasmic domain[27, 28]. EGF and its related ligands are capable of induc-ing proliferation, differentiation, and migration in many dif-ferent cell types [29]. The EGF family, and particularly theheregulin receptors, have been associated with stimulationof proliferation and malignant transformation. In addition,these factors are known to stimulate growth and morpho-genesis in several developing systems including the kidney,lung, mammary gland, and microvascular endothelial cells[30–36].
EGF and the related ligand transforming growth factora (TGF-a) are expressed in the postnatal testis [37–42].TGF-a and EGF have various effects on the proliferation,differentiation, and migration of Sertoli, Leydig, and peri-tubular cells [37, 39, 43]. Although they have not been aswell characterized, other members of the EGF/TGF-a fam-ily are also found in the testis [26, 44, 45]. Since the TGF-a/EGF family of ligands have the ability to stimulate bothgrowth and morphogenesis, they have the potential to playa critical role in early testis development. The current ex-periments were designed to investigate the hypothesis thatthe TGF-a family of growth factors has a role in embryonictestis development.
MATERIALS AND METHODS
Dissections and Organ Cultures
Timed-pregnant Sprague-Dawley rats were obtainedfrom Charles River (Wilmington, MA). Plug date was con-sidered to be embryonic Day 0 (e0). For e13 dissections,gonads were dissected out with the mesonephros; for later-stage dissections, testes alone were dissected. The organswere cultured at 378C in 5% CO2 in drops of medium onMillicell CM filters (Millipore, Bedford MA) floating onthe surface of 0.5–1 ml of CMRL 1066 medium (GibcoBRL, Gaithersburg, MD) supplemented with penicillin-streptomycin, insulin (10 mg/ml), and transferrin (10 mg/ml). Antibodies and factors were added directly to the cul-ture medium. The medium was changed every 1 or 2 days.Cultures of e13 gonads 1 mesonephros were incubated for3 days, by which point cords were well developed; e14testis cultures were incubated for 4 days. For e18 and post-natal Day 0 (0d) organ cultures, the testes were cultured asfragments (halves of e18 testes and eighths of 0d testes),since entire testes were too large to culture intact withoutconsiderable necrosis in the centers. The tunica was re-moved from 0d testis before culturing. The testes collectedfor morphological analysis prior to cell culture were fixedand processed immediately after microdissection or wereobtained from whole embryos that had been fixed prior tomicrodissection. All procedures were approved by the ap-propriate University Animal Care committees.
Testis Suspension and Reaggregation
To generate a testicular suspension from 0d testis, thetunica was removed and the testes were digested with0.125% trypsin, 0.1% EDTA, and 0.02 mg/ml DNase inHanks’ Balanced Salt Solution (HBSS) for 15 min at 378C.The trypsin was inactivated with 10% calf serum. The sam-ples were triturated with a pipette tip and washed twice in1 ml HBSS by resuspending, spinning for 2 min, and re-moving the supernatant. The remaining pellet was resus-pended in a small volume (10 ml from 6–7 pooled testes)and contained a single-cell suspension according to visualinspection under a microscope. Drops of 2 ml of this sus-
pension were cultured and allowed to reaggregate on float-ing filters as for intact organs.
Genomic DNA Isolation and Polymerase Chain Reaction(PCR) for SRY
To determine the sex of e13 gonads, embryonic tailswere collected to make genomic DNA by standard proce-dures. Briefly, the tissue was homogenized through a 25-gauge needle in digestion buffer (100 mM NaCl, 10 mMTris, pH 8, 25 mM EDTA, 0.5% SDS) and digested withproteinase K (0.15 mg/ml) for at least 4 h at 608C. Thesamples were then extracted twice with an equal volume ofphenol:chloroform:isoamyl alcohol (25:24:1) and once withchloroform:isoamyl alcohol. The DNA was precipitated byadding 1/10 volume 7.5 M NH4Ac and 3 volumes coldethanol and incubating at 2808C for 1 h before centrifu-gation at 48C for 30 min. Pellets were dried and resus-pended in 10 ml H2O. PCR was performed by using 1 mlof genomic DNA with the consensus SRY primers 59CGGGATCCATGTCAAGCGCC CCATGAATGCATT-TATG 39 and 59 GCGGAATTCACTTTAGCCCTCCGAT-GAGGCTGA TAT 39, producing a PCR product of 240base pairs (bp). PCR was performed using an annealingtemperature of 558C for 30 cycles.
RNA Isolation and Reverse Transcription-PolymeraseChain Reaction (RT-PCR)
RNA for RT-PCR was obtained by freezing samples ondry ice and then using the Tri Reagent (Sigma ChemicalCo., St. Louis, MO) for RNA isolation. Reverse transcrip-tion was performed using murine Moloney leukemia virus-RT under standard conditions. RT-PCR was performed forEGFR at 608C annealing temperature for 30 cycles and forTGF-a at 658C for 5 cycles followed by 558C for 30 cycles.The EGFR primer sequences were 59TGCGpTCTCTTG-CCGGAATGTCAG39 and 59GCAGTGGGGGCCGTCA-ATGTAGT39, producing a PCR product of 205 bp. TheTGF-a primer sequences were 59ACCCTTTATCACACA-GTTTT39 and 59CCCTGGCTGTCCTC ATTATCACCTG39, producing a PCR product of 380 bp. The identity of thePCR products was confirmed by restriction digests.
Embedding, Histology, and Immunocytochemistry forProliferating Cell Nuclear Antigen (PCNA)
Tissues were fixed in Histochoice (Amresco, Solon, OH)and embedded in paraffin or in JB4 (Polysciences, War-rington, PA) according to standard procedures. Sectionswere stained with hematoxylin and eosin (paraffin sections)or with toluidine blue (JB4 sections) according to standardprocedures. Immunocytochemistry for PCNA was per-formed according to standard procedures. Briefly, 7-mmsections were deparaffinized and rehydrated, quenched in20% methanol/3% hydrogen peroxide, and blocked in 5%serum for several hours at room temperature before incu-bation with primary antibody monoclonal anti-PCNA clonePC10 (Sigma) at 1:500 dilution overnight at 48C. Second-ary antibody (biotinylated sheep anti-mouse biotin fromAmersham Pharmacia Biotech, Piscataway, NJ) was de-tected by using the Vectastain kit (Vector Laboratories, Bur-lingame, CA) and diaminobenzadine. Slides were counter-stained lightly with hematoxylin to visualize the tissue.
Labeling indices for the percentage of each cell typestained for PCNA were calculated by counting random re-gions on at least 3 different sections from 3 different ex-

479ROLE OF TGF-a/EGFR IN EMBRYONIC TESTIS GROWTH
periments. The total number of cells counted in specificregions ranged from 155 to 396 Sertoli cells, 80 to 396germ cells, 338 to 862 interstitial cells, and 139 to 306peritubular cells. Counts were performed by two differentpeople independently for comparison, and data were com-bined. Data are presented as the mean 6 SEM to assess thepercentage of stained cells. The data from the two individ-uals generally compared as shown by the reproducibility ofthe data.
DNA Assay
To assay the DNA content of organs, each organ wassonicated in 100 ml ethidium bromide buffer (EBB; 20 mMNaCl, 5 mM EDTA, 10 mM Tris, pH 7.5) and stored at2208C. DNA content then was determined fluorometricallywith ethidium bromide as previously described [37]. Brief-ly, 0.25 nM ethidium bromide and 100 units/ml heparin inEBB were added to each sample; samples were vortexedand incubated for 15 min at room temperature. Fluorescentemission was measured and quantified by using a standardcurve with calf thymus DNA from 0.5 mg to 6 mg DNA.
Imaging and Area Quantification
Images of whole organs were obtained by using a Lu-mina digital scanning camera (Leaf Systems, Southbor-ough, MA). Area was calculated by using Adobe Photoshop(Mountain View, CA). To determine the relative percentageof interstitium in testis, representative sections were imagedand analyzed using Adobe Photoshop. The outline of eachcord was traced and the area within each was calculated,as was the total area. The percentage of interstitium wascalculated as area cords/total area.
Western Blotting
Western blotting was performed according to standardprocedures. Protein samples were electrophoresed on a 8–25% Phast gradient gel (Phast System; Pharmacia, Ala-meda, CA) and transferred to nitrocellulose by using thePhastTransfer semidry electrophoretic transfer unit (Phar-macia). After blocking overnight in 5% dry milk/1% gly-cine/0.05% Nonidet P-40 (NP40) in 50 mM Tris-HCl (pH7.4)/0.15 M NaCl (TBS), the primary antibodies againstTGF-a were incubated with the blots for 4 h. After threewashes in TBS/NP40 and one in TBS, the blots were in-cubated with secondary antibody anti-sheep conjugated tohorseradish peroxidase (Sigma) at 1:3000 for 1 h. Afterthree washes in TBS/NP40, one in TBS, and three in water,detection was carried out using the ECL system (Amer-sham).
Reagents
The antibodies against TGF-a (S574 raised against ratTGF-a and S509 raised against human TGF-a) were ob-tained from East Acres Biological (Southbridge, MA).These TGF-a antibodies have been shown to be neutraliz-ing polyclonal antibodies and to cross-react with mouse,rat, human, and bovine TGF-a and not to cross-react withEGF, amphiregulin, TGF-b, or fibroblast growth factor(FGF) on Western blots (East Acres Biological). The EGFRinhibitor AG1478 was obtained from Calbiochem (La Jolla,CA). AG1478 is a highly selective competitive inhibitor ofATP binding to the EGFR. The in vitro IC50 for EGFR is3 nM, while the IC50 for HER2 and the platelet-derived
growth factor receptor are greater than 100 mM. TGF-a andrecombinant human heregulin-a were obtained from R&DSystems (Minneapolis, MN). TGF-a knockout mice wereprovided generously by Dr. Gerald Cunha (University ofCalifornia at San Francisco [UCSF], San Francisco, CA),who maintains a colony of mice previously developed [19].EGFR knockout mice were provided generously by ZenaWerb and Rik Derynk (UCSF) and by Terry Magnuson(Case Western University, Cleveland, OH).
Statistical Analysis
All the data were obtained from a minimum of threedifferent experiments unless otherwise stated. Each datapoint was converted to a mean and SEM from multipleexperiments determined as indicated in the figure legends.Data were analyzed by a Student’s t-test or ANOVA asindicated in the figure legends. Different superscript lettersdenote a statistical difference.
RESULTS
Proliferation Pattern in Developing Testis
Investigation of the effects of a particular factor ongrowth in the developing testis requires an understandingof the overall pattern of growth of different cell types atvarious stages of development. The cell populations thatare proliferating provide potential targets for the locallyproduced growth factors (e.g., TGF-a). The pattern of cellproliferation in the early rat testis was determined by usingimmunocytochemistry for PCNA (Fig. 1). PCNA is an aux-iliary protein of DNA polymerase that is required for DNAreplication during S-phase and therefore is expressed onlyin proliferating cells [46–48]. The earliest time examinedwas e14, which is the earliest point at which the testis isdistinguishable morphologically from the ovary, just aftercords have formed (Fig. 1A). Additional time points wereexamined through late embryonic and early postnatal de-velopment up to the early pubertal period of 20 days post-natal (20d). The pattern of PCNA staining changed duringembryonic and early postnatal development (Fig. 1). At e14(Fig. 1A), many cells in both the cords and the interstitiumexpressed PCNA, although many of the Sertoli nuclei thatwere lined up at the edges of the cords were unstained. Bye16 (Fig. 1B), all cells in the cords were stained, as werethe peritubular cells surrounding the cords. The interstitiumcontained both PCNA-positive and PCNA-negative cells.The pattern at e18 (Fig. 1C) was similar except that moregerm cells in the centers of the cords did not stain. Thishigh percentage of PCNA-positive cells throughout the lateembryonic testis is consistent with the rapid growth of theorgan. At birth (0d; Fig. 1D) there was little staining ofinterstitial, peritubular, or germ cells, but almost all Sertolicells were stained. By 5d (Fig. 1E), the cells in the cordshave proliferated so that the cords occupy much more ofthe area of the testis than in the embryonic testis. The Ser-toli cells were still stained heavily for PCNA, and somegerm cells had begun expressing PCNA again. At 10d (Fig.1F), some Sertoli cells had left the cell cycle; at 20d, theSertoli cells did not stain (data not shown). Similar resultswere obtained with the adult 60d testis (data not shown).Various spermatogenic cells were stained, including sper-matogonia at the outside edges of the tubules and morecentrally located spermatocytes. In addition, meiotic figureswere visible. A high magnification of 10d testis is presented
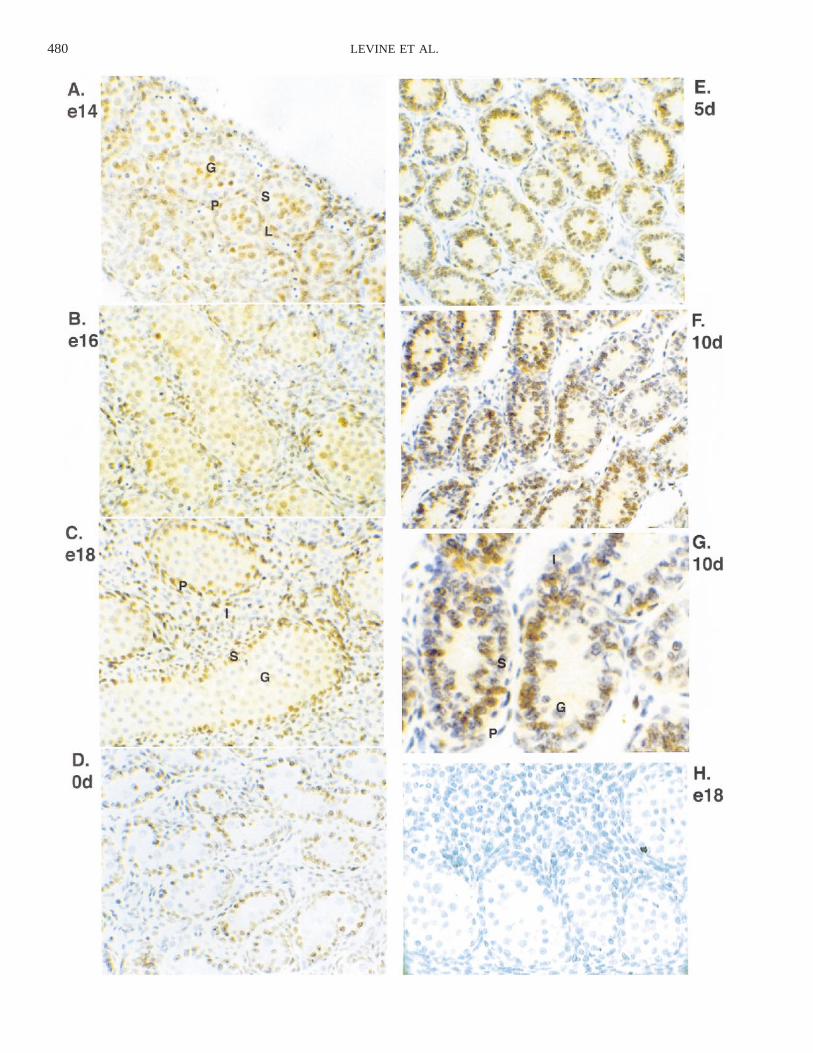
480 LEVINE ET AL.

481ROLE OF TGF-a/EGFR IN EMBRYONIC TESTIS GROWTH
FIG. 2. Labeling indices of the percentage of PCNA-positive cells foreach cell type at various stages of development. Labeling indices werecalculated by counting stained and unstained cells in random regions ofreplicate slides from at least 3 different sections in 3 different experiments.Data are presented as the mean 6 SEM.
FIG. 3. Expression of TGF-a and EGFR mRNA by RT-PCR in the devel-oping testis. Lanes 1–5, TGFa; lanes 6–9, EGFR. Individual lanes: 1, mo-lecular weight markers; 2, e14; 3, e18; 4, 0d; 5, no template; 6, e14; 7,e18; 8, 0d; 9, no template. The sizes of the PCR products were 380 bpfor TGF-a and 205 bp for EGFR. Representative of at least 3 differentexperiments.
b
FIG. 1. PCNA immunocytochemistry in the developing rat testis; (G)high magnification of F. PCNA-positive cells are brown while negativecells are purple from hematoxylin counterstain. H) A negative control e18hematoxylin stain. Representative cells: P, peritubular cell; S, Sertoli cell;G, germ cell; L or I, interstitial cell. G) Two different stages of germ cellare labeled (spermatogonia and spermatocyte). 3200 except for G, whichis 3400. Representative of at least three different experiments done inreplicate.
in Figure 1G to show specific cell populations. A controlbackground stain with nonimmune IgG is shown in Figure1H.
Labeling indices were calculated at ages e14, e18, 0d,5d, and 10d to extend the observations with a calculationof the nuclear PCNA staining pattern and ratio of cellsstained (Fig. 2). Proliferation decreased in germ cells bye18, was minimal at 0d, and returned to high levels by 5dpostnatal. The interstitium proliferated from the initiationof testis development through the late embryo and de-creased substantially in the early postnatal period. Sertolicells did not appear to be proliferating at the time of cordformation (e14) but proliferated extensively through the restof embryonic and early postnatal development until puber-ty. The decrease of Sertoli cell proliferation at the onset ofpuberty has been previously documented [14]. Peritubularproliferation was high until after birth and then decreaseddramatically. Observations suggest that all the testis cellpopulations provide potential targets for locally producedgrowth factors (e.g., TGF-a).
Expression of TGF-a and EGFR in the Embryonic Testis
Both TGF-a and EGFR are expressed in the postnataltestis. To investigate their presence in the embryonic rattestis, RNA was isolated from testes of various ages andRT-PCR was performed. RT-PCR for TGF-a and EGFR(Fig. 3) demonstrated that both genes were expressedthroughout embryonic testis development at e14, e18, and0d. Similar results were obtained from e14 and 0d mousetestis RNA (data not shown).
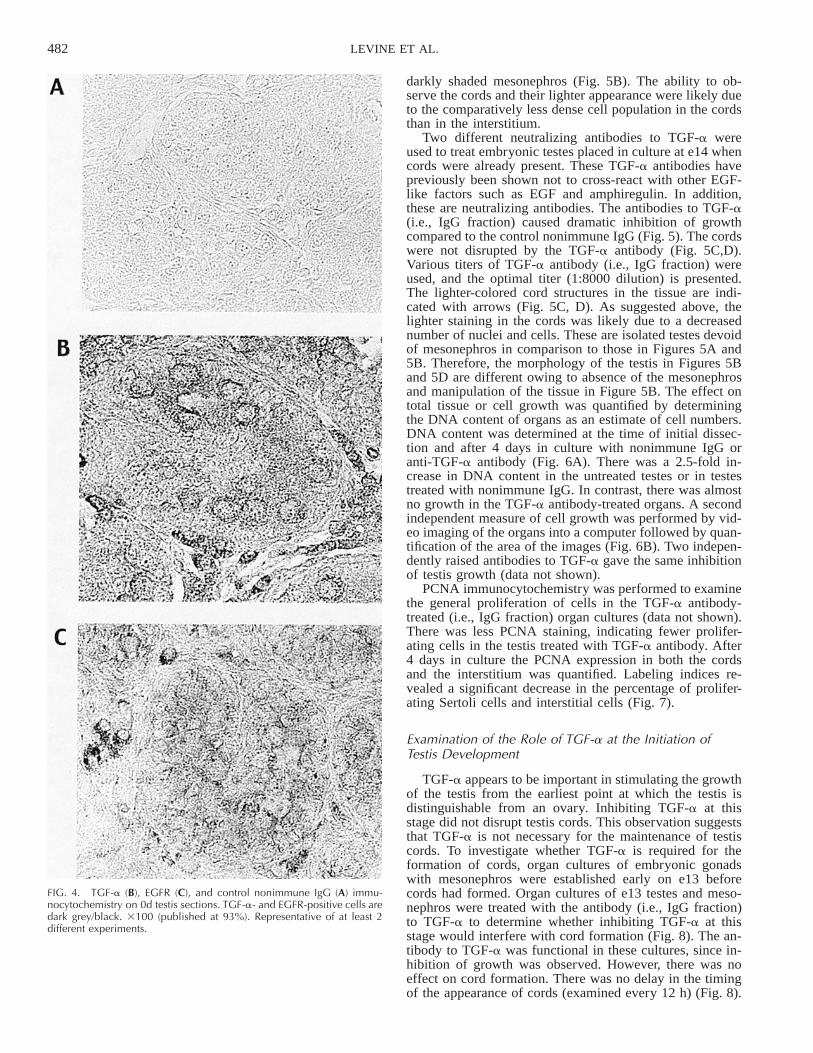
Immunocytochemistry was performed to confirm the ex-pression of the TGF-a and EGFR proteins. TGF-a andEGFR immunocytochemistry utilized sections from 0d rattestis. The TGF-a and EGFR were primarily localized tothe interstitial cells with lower levels of staining in thecords (Fig. 4). Preabsorbtion of the TGF-a antibody withexcess TGF-a eliminated the staining detected in the im-munocytochemistry (data not shown). Observations sug-gested that interstitial cells and to a lesser extent the cordsin the 0d testis express the TGF-a and EGFR proteins andsupported the mRNA data presented in Figure 3. In addi-tion, the specificity of the TGF-a antibody was indicatedby the immunocytochemistry control.
Functional Perturbation of Embryonic Testis Developmentwith TGF-a-Neutralizing Antibody
To investigate whether TGF-a is important for stimulat-ing the growth of the embryonic testis, organ cultures ofthe embryonic testis were established. Growing the testesin organ culture allowed direct perturbation of their devel-opment. Testes from e14 embryos grew during 4 days inculture and maintained their cord structure. Testes from e13embryos cultured with the mesonephros developed cordsover 3 days in culture (Fig. 5A). The top part of the mi-crograph (marked ‘‘T’’) is the testis connected to the lower
482 LEVINE ET AL.
FIG. 4. TGF-a (B), EGFR (C), and control nonimmune IgG (A) immu-nocytochemistry on 0d testis sections. TGF-a- and EGFR-positive cells aredark grey/black. 3100 (published at 93%). Representative of at least 2different experiments.
darkly shaded mesonephros (Fig. 5B). The ability to ob-serve the cords and their lighter appearance were likely dueto the comparatively less dense cell population in the cordsthan in the interstitium.
Two different neutralizing antibodies to TGF-a wereused to treat embryonic testes placed in culture at e14 whencords were already present. These TGF-a antibodies havepreviously been shown not to cross-react with other EGF-like factors such as EGF and amphiregulin. In addition,these are neutralizing antibodies. The antibodies to TGF-a(i.e., IgG fraction) caused dramatic inhibition of growthcompared to the control nonimmune IgG (Fig. 5). The cordswere not disrupted by the TGF-a antibody (Fig. 5C,D).Various titers of TGF-a antibody (i.e., IgG fraction) wereused, and the optimal titer (1:8000 dilution) is presented.The lighter-colored cord structures in the tissue are indi-cated with arrows (Fig. 5C, D). As suggested above, thelighter staining in the cords was likely due to a decreasednumber of nuclei and cells. These are isolated testes devoidof mesonephros in comparison to those in Figures 5A and5B. Therefore, the morphology of the testis in Figures 5Band 5D are different owing to absence of the mesonephrosand manipulation of the tissue in Figure 5B. The effect ontotal tissue or cell growth was quantified by determiningthe DNA content of organs as an estimate of cell numbers.DNA content was determined at the time of initial dissec-tion and after 4 days in culture with nonimmune IgG oranti-TGF-a antibody (Fig. 6A). There was a 2.5-fold in-crease in DNA content in the untreated testes or in testestreated with nonimmune IgG. In contrast, there was almostno growth in the TGF-a antibody-treated organs. A secondindependent measure of cell growth was performed by vid-eo imaging of the organs into a computer followed by quan-tification of the area of the images (Fig. 6B). Two indepen-dently raised antibodies to TGF-a gave the same inhibitionof testis growth (data not shown).
PCNA immunocytochemistry was performed to examinethe general proliferation of cells in the TGF-a antibody-treated (i.e., IgG fraction) organ cultures (data not shown).There was less PCNA staining, indicating fewer prolifer-ating cells in the testis treated with TGF-a antibody. After4 days in culture the PCNA expression in both the cordsand the interstitium was quantified. Labeling indices re-vealed a significant decrease in the percentage of prolifer-ating Sertoli cells and interstitial cells (Fig. 7).
Examination of the Role of TGF-a at the Initiation ofTestis Development
TGF-a appears to be important in stimulating the growthof the testis from the earliest point at which the testis isdistinguishable from an ovary. Inhibiting TGF-a at thisstage did not disrupt testis cords. This observation suggeststhat TGF-a is not necessary for the maintenance of testiscords. To investigate whether TGF-a is required for theformation of cords, organ cultures of embryonic gonadswith mesonephros were established early on e13 beforecords had formed. Organ cultures of e13 testes and meso-nephros were treated with the antibody (i.e., IgG fraction)to TGF-a to determine whether inhibiting TGF-a at thisstage would interfere with cord formation (Fig. 8). The an-tibody to TGF-a was functional in these cultures, since in-hibition of growth was observed. However, there was noeffect on cord formation. There was no delay in the timingof the appearance of cords (examined every 12 h) (Fig. 8).

483ROLE OF TGF-a/EGFR IN EMBRYONIC TESTIS GROWTH
FIG. 5. E13 testis 1 mesonephros organcultures after 1 day (A) and after 3 days(B) in culture. The upper structure (T) isthe testis attached to the lower, dark-col-ored mesonephros (M). Representative ofmore than 10 different experiments. Thediameter of the organ was approximately1000 mm. E14 testis organ cultures treatedwith control nonimmune IgG (C) or withanti-TGF-a IgG antibody S574 (D) after 4days in culture. Representative cords aremarked by arrows placed on the lighter-colored structures and are shown in C andD. This is a microdissected testis devoid ofmesonephros with some manipulation incomparison to that shown in A and B.Representative of 10 experiments.
FIG. 7. Labeling index for the percentage of each cell type positive forPCNA on e14 testis organ cultures treated with control nonimmune oranti-TGF-a antibody for 4 days. Labeling indices were calculated bycounting stained and unstained cells in random regions of at least 3 dif-ferent sections from 3 different experiments. ** Statistically significant dif-ference from control (P , 0.001); * statistically significant difference fromcontrol (P , 0.02) as determined by a t-test.
FIG. 6. Quantitation of e14 testis organ cultures treated with control non-immune IgG or with anti-TGF-a IgG antibody S574 for 4 days in culture.(A) DNA content of the organ cultures; (B) area of the organ cultures. (A)DNA content (mg) was measured by ethidium bromide fluorometry onextracts of testis organ cultures. Data are presented as the mean 6 SEMfor 3 separate experiments with n 5 9 (Initial), n 5 21 (Control Ab), andn 5 22 (TGF-a Ab). (B) Area (mm2) was measured using NIH Image after1 day and after 4 days in culture. Data are presented as the mean 6 SEMfor 3 separate experiments with n 5 13 (Initial 5 1 day), n 5 23 (ControlAb), and n 5 24 (TGF-a Ab). Different superscripts designate statisticallysignificant difference (P , 0.01) as determined by ANOVA.
Investigation of the Role of TGF-a Later in Embryonicand Perinatal Development
In order to determine whether TGF-a is required for tes-tis growth in perinatal development, testis organ cultures atlater embryonic ages were treated with an antibody to TGF-a and assayed for DNA content as a measure of cell num-ber. There was still dramatic inhibition of testis growth ate18 by the antibody to TGF-a (Fig. 9A). The control organstreated with nonimmune IgG exhibited only about a 1.5-fold increase in DNA content after 6 days in culture insteadof the 2.5-fold increase seen with e14 testis. The antibodyto TGF-a abolished all growth. The smaller increase in sizeof the control organs may reflect a decreased growth and/or a limitation on growth in the organ culture system. The0d testes grew less, and growth was inhibited partially by

484 LEVINE ET AL.
FIG. 8. E13 testis 1 mesonephros organ cultures treated with controlnonimmune IgG or anti-TGF-a IgG antibody S574 for 3 days. (A) Controlnonimmune IgG; (B) anti-TGF-a IgG antibody. T, testis; M, mesonephros.A and B are the same magnification. Representative of 4 different exper-iments.
c
FIG. 9. Effects of control nonimmune IgG or anti-TGF-a antibody S574on the growth of e18 (A) and 0d (B) testis. DNA content (mg) was deter-mined at the time of dissection (initial) or after 4 days in culture. A) Mean6 SEM for 2 separate experiments with n 5 22 for all conditions. (B)Mean 6 SEM for 3 separate experiments with n 5 28 (initial) and n 529 (control Ab and anti-TGF-a antibody) using 1/8 of testis. (C) Effects ofEGFR tyrosine phosphorylation inhibitor AG1478 (160 nM) or control sol-vent dimethyl sulfoxide (DMSO) on the growth of e14 testis cultured for4 days; mean 6 SEM for 3 separate experiments with n 5 13 (initial), n5 14 (DMSO), and n 5 15 (AG1478). Different superscripts designatestatistically significant differences (P , 0.01) as determined by ANOVA.
the antibody to TGF-a (Fig. 9B). Observations suggest thatTGF-a is important for testis growth throughout embryonicdevelopment and in early postnatal development.
Perturbation of the EGFR by a Tyrphostin Inhibitor
To confirm the importance of TGF-a in testis growth, anindependent approach of perturbing the EGFR signaling

485ROLE OF TGF-a/EGFR IN EMBRYONIC TESTIS GROWTH
FIG. 10. Effects of exogeneous TGF-a (50ng/ml) on testis cultures. A,B) E14 testiscultured for 4 days in the absence (A) orpresence (B) of TGF-a (40 ng/ml). Repre-sentative of 5 different experiments. (C,D)Dissociation-reaggregation cultures of 0dtestis in the absence (C) or presence (D) ofTGF-a (100 ng/ml). Representative of 2different experiments. Representative cordsare marked by arrows. The ring of unorga-nized tissue around the edge in B and Dis marked by arrowheads.
pathway was used. The action of the EGFR was blockedwith use of a highly selective tyrosine kinase inhibitor ofthe tyrphostin family known as AG1478 [49, 50]. Treat-ment of e14 testes in organ culture with 160 nM AG1478resulted in a striking inhibition of growth (Fig. 9C). Treat-ment with 80 nM AG1478 was found to be less effective,although it resulted in some inhibition of growth (data notshown). Similar growth inhibition was apparent when 0dtestis cultures were treated with AG1478 (data not shown).Observations suggest that the TGF-a receptor EGFR is re-quired for optimal embryonic testis growth.
Effect of Exogeneous TGF-a on Testis Organ Cultures
If TGF-a is important in promoting the proliferation ofcells in the embryonic testis, exogeneous TGF-a may in-fluence testis growth. Testis organ cultures at e14 weretreated with 50 ng/ml of TGF-a and then stained for PCNAto determine whether there was an increase in the numberof proliferating cells. A 25-ng/ml and a 100-ng/ml dose ofTGF-a gave results similar to those with the 50-ng/ml dose(data not shown). Addition of TGF-a to the culture mediumcaused a ring of unorganized tissue around the edge of theorgan (Fig. 10A, B). Immunocytochemistry for PCNAdemonstrated that almost all the cells in the outer ring wereproliferating (data not shown). This suggests that cells inthe embryonic testis are capable of responding to exoge-
neous TGF-a by inducing additional proliferation. Thesame effect was seen with reaggregated cultures of a cellsuspension from a 0d testis. When a 0d testis was disso-ciated to single cells and the cells were allowed to reag-gregate on a floating filter, cords reformed in the aggregate(Fig. 10C). When these cultures were treated with exoge-neous TGF-a, a ring of unorganized cells developed aroundthe edge just as in the intact testis organ cultures (Fig. 10C,D).
Examination of Testis from EGFR and TGF-aKnockout Mice
The data with the neutralizing antibodies suggested thatTGF-a is critical for growth of the early embryonic testis.Since TGF-a functions through the EGFR, it was possiblethat EGFR knockout mice might show a defect in testisgrowth. Testis from EGFR knockout mice were examinedat several ages starting with e13 (the equivalent of e15 inthe rat). Despite dramatic effects on other organs and onthe overall size of the EGFR knockout mice [51–53], thetestes were grossly normal (Fig. 11). There was apparentlyno decrease in size in the early embryonic testis comparedto that in wild-type littermates. Later in development therewas some decrease in size, but it was less than the overalldecrease in size of the animals (data not shown). It appears

486 LEVINE ET AL.
FIG. 11. A) E18 wild-type (WT) and B) e18 knockout (KO) mice testes.JB4 sections were stained with toluidine blue. 3200 (published at 80%).Data are representative of more than 10 sections from different testes.
FIG. 12. Quantitation of percentage interstitium in EGFR knockout andwild-type testes from e18 and 0d mice. Computer analysis of the area ofthe interstitium was performed by subtracting the combined areas of thecords from the total area of each testis section. Data are presented as themean 6 SEM; numbers of different testes analyzed were n 5 11 (e18WT), n 5 16 (e18 KO), n 5 9 (0d WT), and n 5 8 (0d KO). The e18 micewere from two independently produced knockout mouse lines. Differentsuperscripts designate statistically significant difference (P , 0.01) as de-termined by ANOVA.
that the testis is able to compensate for the lack of theEGFR.
When sections through the testes were examined, thereappeared to be a subtle effect on the growth of the inter-stitium in the EGFR knockout mice. Less interstitium waspresent in the knockout mice than in the wild-type litter-mates (Fig. 11). Quantitation of this effect revealed that ate18 there was approximately a 20% decrease in the per-centage of interstitium in the EGFR knockout testis (Fig.12). The difference in the area of the interstitium observedwas found to be due to a decrease in cell number throughquantitation of cell numbers in selected sections (data notshown). In the 0d testis, the difference between the area ofthe interstitium in the wild-type and knockout testis wasnot significant (data not shown). Therefore, the EGFRknockout testis phenotype is a transient decrease in inter-stitial cell growth late in embryonic development.
In contrast, testes from TGF-a knockout mice appearednormal at the developmental stages examined in compari-son to normal littermates (data not shown). Although theEGFR knockout phenotype supports a role for EGF familyligands in the developing testis, combined data suggest thatalternate and/or compensating TGF-a-related ligands and/or EGFR-related receptors appear to be present.
TGF-a Antibody Specificity
The presence of TGF-a antibody resulted in completeinhibition of testis growth. This neutralizing TGF-a anti-body has previously been shown to not cross-react withEGF, amphiregulin, TGF-b, or FGF. The inability of the
antibody to react with other EGF-like ligands such as EGFor amphiregulin suggests a high degree of specificity; how-ever, reactivity with another EGF-like ligand, heregulin, hasnot been shown. To further examine whether the antibodiesto TGF-a are able to recognize related ligands, a Westernblot was performed. The antibodies to TGF-a were foundto react only with TGF-a and not with either EGF or her-egulin-a (Fig. 13). A Western blot of Day 0 testis showedthe 25-kDa precursor and 6-kDa mature forms of the TGF-a protein (data not shown). Therefore, the TGF-a antibod-ies appear specific with the factors tested thus far.
DISCUSSION
The results provide information about the control ofgrowth during embryonic testis development. Immunocy-tochemistry for the proliferation marker PCNA confirmedthe dramatic growth of the embryonic testis. Each cell typehad its own temporal pattern of proliferation. During lateembryonic development, almost all somatic cells were pro-liferating both in the cords and in the interstitium. The num-ber of proliferating cells decreased first in the interstitium.A high percentage of proliferating cells continued to befound in the Sertoli cell population through early postnataldevelopment, decreasing by 10 days at the onset of pubertaltestis development. Germ cell proliferation was extensiveat e14 and e16, decreased by e18, and then stopped untilthe early postnatal period, at which point it increasedthrough 20d development. Therefore, all the testis cell typesprovide targets for locally produced growth factors such asTGF-a.
These observations based on PCNA as a marker of pro-liferating cells confirmed earlier experiments in which mi-totic figures were quantified [14] and the incorporation oftritiated thymidine was examined [13, 15]. The only differ-ence from the previous data is that the experiments dem-onstrated mitotic arrest of the germ cells by e18 [13, 54].In the previous thymidine incorporation experiments, e18

487ROLE OF TGF-a/EGFR IN EMBRYONIC TESTIS GROWTH
FIG. 13. Western blot with anti-TGF-a antibodies S574 and S509 ofrecombinant TGF-a, EGF, and heregulin-a (300 ng each). A) SDS gelstained with Coomassie Blue. B) Western blot with anti-TGF-a antibodyS509. C) Western blot with anti-TGF-a antibody S574. A,B,C) Lanes: 1,molecular weight markers; 2, heregulin-a; 3, EGF; 4, TGF-a. TGF-a andEGF are 6 kDa and heregulin-a is 7 kDa.
germ cells did not stain positive, whereas in the currentstudy, e18 germ cells stained positive for PCNA. A limi-tation to the use of PCNA is that the protein is retained bythe cell for a period after cell proliferation. Therefore,PCNA is a marker of general cell growth, and some PCNA
likely remains after mitotic arrest in male germ cells. Asimilar observation has been made for female germ cells,since there is PCNA staining in the oocyte while it is inmeiotic arrest [55]. PCNA staining has been shown to cor-relate extremely well with cell proliferation, but it is alsoinvolved in DNA repair [47, 55–58]. Increased levels ofDNA repair in germ cells could also explain the persistenceof PCNA at e18. Therefore, the PCNA data presented con-firm the previous studies, and the difference at e18 is likelya technical limitation to the use of PCNA. The extensivecell proliferation suggests a need for stimulators of growthin the embryonic testis. This dramatic growth is sex-spe-cific, since the ovary grows very little during late embry-onic development [12]. In addition, the differences in thetiming of proliferation of the various cell types suggests theneed for the regulation of growth through local growth fac-tors and receptors such as TGF-a and EGFR.
TGF-a was found to be important in stimulating thegrowth of the embryonic testis. The growth of e14 testis inorgan culture was inhibited by either of two neutralizingantibodies to TGF-a. Growth was measured by changes inDNA content of the organs and was confirmed by measur-ing the area of the organ. Quantitation by area is less ac-curate since it is a two-dimensional measurement of a three-dimensional tissue. However, it was still apparent that theantibodies to TGF-a effectively inhibited the growth of e14testis in organ culture. The neutralizing antibodies to TGF-a and the EGFR inhibitor were still able to inhibit thegrowth of late embryonic (e18) and perinatal (0d) testis.This suggests that TGF-a ligand is important for growththroughout early testis development. Immunocytochemistryfor PCNA in the organ cultures demonstrated that growthof both Sertoli and interstitial cells is inhibited by the an-tibodies to TGF-a. These observations suggest that TGF-ais important for promoting growth of both cell types. How-ever, the current experiments do not rule out possible in-direct effects through various cell-cell interactions. The ex-act location of TGF-a expression and action remain to bedetermined.
Testis growth was also inhibited by the tyrphostinAG1478, which is a specific inhibitor of the EGFR tyrosinekinase. AG1478 is selective for the EGFR and does notinhibit other growth factor receptor tyrosine kinases includ-ing the related HER2 kinase at the concentrations used inthese experiments [49]. Inhibition of growth of the embry-onic testis by AG1478, therefore, provides further evidencefor the importance of TGF-a-induced signaling in stimu-lating testis growth.
Despite dramatic effects on embryonic testis growth,neutralizing antibodies to TGF-a did not disrupt cord for-mation in e13 testis. TGF-a and EGF have been implicatedin morphogenetic events in other systems involving tubeformation and branching such as the kidney, lung, mam-mary gland, and endothelial cells [30–36]. Organ culturesof early e13 testis and mesonephros are susceptible to theinhibition of cord formation by serum [1, 59]. This dem-onstrates the possibility of perturbing cord formation in tes-tis organ cultures. The lack of an effect of the TGF-a an-tibodies on the development of cords in the testis suggeststhat TGF-a is not necessary for testis cord formation. IfTGF-a does play a role in testis cord formation, then otherfactors must be able to compensate in the organ cultures.
Exogenous TGF-a was found to affect testis organ cul-tures, implying that the cells are capable of responding toTGF-a. There was not an overall increase in the size orDNA content of the organ but an increase in disorganized

488 LEVINE ET AL.
cells around the outside of the testis. One reason could bethat TGF-a stimulates proliferation to such an extent thatthe cells can no longer maintain the cord structure; anothercould be an overstimulation of growth of the interstitiumsuch that it grows out beyond the cords. PCNA immuno-cytochemistry of the organs treated with TGF-a revealedan increase in the number of proliferating cells around theoutside of the testis, such that almost all the cells in thisregion expressed PCNA. These observations support theinterpretation that TGF-a treatment preferentially increasedthe proliferation of the more accessible cells on the outsideof the organs. Maintenance of the cord structure is clearlycompatible with some proliferation, since there is extensivePCNA staining in the cords throughout late embryonic andearly postnatal development. However, it is possible that anincreased rate of proliferation causes a disruption of thestructure.
In contrast to the extensive inhibition of testis growth byneutralizing TGF-a antibodies or inhibiting signalingthrough the EGFR, the lack of EGFR in knockout mice didnot cause a significant inhibition of testis growth. TheEGFR knockout mice did show a dramatic inhibition ofgrowth of other organs and of the whole animal [51–53].There was a decrease in the percentage of the interstitiumin late embryonic (e18) testis, suggesting that the growthof the interstitium was slightly inhibited or delayed. Thiseffect was observed in two different EGFR knockoutstrains. Interstitial volume and cell numbers were transient-ly decreased. Observations suggest that the interstitial cellsmay be more dependent on the presence of the EGFR forgrowth and less able to compensate for its absence thanSertoli cells.
There has been no evidence that TGF-a is capable ofinteracting directly with any other related receptors. Pre-viously, the TGF-a antibodies were shown to not cross-react with EGF, amphiregulin, TGF-b, or FGF. The inabilityto react with EGF suggests a high degree of specificity, butreactivity to heregulin had not been examined. Westernblotting of both antibodies against recombinant human EGFand heregulin-a failed to detect any cross reactivity. Al-though the antibodies used appear specific, further investi-gation is required to determine whether unidentified TGF-a-related ligands may be recognized by the antibodies andfunction normally in the stimulation of testis growth.
The observation that TGF-a is important for stimulatingthe growth of the embryonic testis is significant because ofthe importance of embryonic cell proliferation for adult tes-tis size and fertility. FSH is the most active agent found topromote testis growth and particularly Sertoli cell prolif-eration. FSH in vitro can stimulate Sertoli cell proliferation,and removal of FSH in vivo causes a dramatic decrease inSertoli proliferation in e18 testis [16, 17]. However, the ear-liest detectable FSH in the pituitary and the earliest de-tectable FSH receptors in the testis are in e17 rats [11].Other factors must be responsible for promoting the growthof the earlier testis development. These factors presumablyare produced locally, since testis growth is maintained inorgan culture. The current data imply that TGF-a is capableof performing this function in the early embryonic testis.In addition, it is not known what local testicular factorsmight be involved in the response to FSH in the late em-bryonic testis. b-Endorphin produced by Leydig cells hasbeen implicated in modulating the response to FSH, sinceblocking its action increases the stimulation of e20 Sertolicell proliferation by FSH [17]. It is possible that TGF-a isinvolved in the local mediation of FSH action, since the
TGF-a family appears to be important for testis growth inthe late embryo.
The involvement of other proteins such as basic FGFand activin has been suggested in testis growth at particulartimes in development. Basic FGF is expressed embryoni-cally by both germ cells and Leydig cells beginning arounde16, so it is a potential mediator of local testis cell inter-actions in the late embryo [60]. FGF increases the survivalof Sertoli cells in culture, but there are conflicting data onwhether it increases proliferation [61, 62]. Activin can en-hance the proliferation of postnatal Sertoli cells, and activinreceptor knockouts show decreased testis size and a delayin fertility [18, 63]. However, activin actually inhibits thegrowth of e14 testis in organ culture [64]. The data suggestthat any promotion of growth by activin occurs postnatally,but it may be a negative regulator of embryonic testisgrowth. TGF-a is therefore one of the first factors impli-cated thus far in the local promotion of early embryonictestis growth.
ACKNOWLEDGMENTS
We thank Urvashi Patel, Joel Brody, and Anne Donjacour for technicalassistance. We thank Paivi Miettinen, Zena Werb, Rik Derynk, GeraldCunha, and Terry Magnuson for providing knockout animals. We thankAnne Donjacour for critical reading of the manuscript and Julia Barfield,Susan Cobb, and Jaideep Chaudhary for assistance in preparation of themanuscript.
REFERENCES
1. Jost A, Magre S, Agelopoulou R. Early stages of testicular differen-tiation in the rat. Hum Genet 1981; 58:59–63.
2. Magre S, Jost A. The initial phases of testicular organogenesis in therat. An electron microscopy study. Arch Anat Microsc Morphol Exp1980; 69:297–318.
3. Pelletier RM and Byers SW. The blood-testis barrier and Sertoli celljunctions: structural considerations. Microsc Res Tech 1992; 20:3–33.
4. Russell LD, Bartke, A, Goh JC. Postnatal development of the Sertolicell barrier, tubular lumen, and cytoskeleton of Sertoli and myoid cellsin the rat, and their relationship to tubular fluid secretion and flow.Am J Anat 1989; 184:179–189.
5. Sertoli E. On the existence of special branched cells in the seminif-erous tubules of the human testis. Morgagni 1865; 7:31–39.
6. Bressler RS, Ross MH. Differentiation of peritubular myoid cells ofthe testis: effects of intratesticular implantation of newborn mousetestes into normal and hypophysectomized adults. Biol Reprod 1972;6:148–159.
7. Clermont Y. Contractile elements in the limiting membrane of theseminiferous tubules of the rat. Experimental Cell Research 1958; 15:438–440.
8. Ross MH. The fine structure and development of the peritubular con-tractile cell component in the seminiferous tubules of the mouse. AmJ Anat 1967; 121:523–557.
9. Skinner MK. Cell-cell interactions in the testis. Endocr Rev 1991; 12:45–77.
10. Skinner MK, Fetterolf PM, Anthony CT. Purification of a paracrinefactor, PModS, produced by testiscular peritubular cells that modulatesSertoli cell function. J Biol Chem 1988; 263:2884–2890.
11. Warren DW, Huhtaniemi IT, Tapanainen J, Dufau ML, Catt KJ. On-togeny of gonadotropin receptors in the fetal and neonatal rat testis.Endocrinology 1984; 114:470–476.
12. Mittwoch U, Delhanty JD, Beck F. Growth of differentiating testesand ovaries. Nature 1969; 224:1323–1325.
13. Orth JM. Proliferation of Sertoli cells in fetal and postnatal rats: aquantitative autoradiographic study. Anat Rec 1982; 203:485–492.
14. Clermont YPB. Quantitative study of the cell population of the sem-iniferous tubules in immature rats. Am J Anat 1957; 100:241–267.
15. Steinberger A, Steinberger E. Replication pattern of Sertoli cells inmaturing rat testis in vivo and in organ culture. Biol Reprod 1971; 4:84–87.
16. Orth JM. The role of follicle-stimulating hormone in controlling Ser-toli cell proliferation in testes of fetal rats. Endocrinology 1984; 115:1248–1255.

489ROLE OF TGF-a/EGFR IN EMBRYONIC TESTIS GROWTH
17. Orth JM. FSH-induced Sertoli cell proliferation in the developing ratis modified by beta-endorphin produced in the testis. Endocrinology1986; 119:1876–1878.
18. Boitani C, Stefanini M, Fragale A, Morena AR. Activin stimulatesSertoli cell proliferation in a defined period of rat testis development.Endocrinology 1995; 136:5438–5444.
19. Luetteke NC, Qiu TH, Peiffer RL, Oliver P, Smithies O, Lee DC.TGF alpha deficiency results in hair follicle and eye abnormalities intargeted and waved-1 mice. Cell 1993; 73:263–278.
20. Mullaney BP, Skinner MK. Basic fibroblast growth factor (bFGF)gene expression and protein production during pubertal developmentof the seminiferous tubule: follicle-stimulating hormone-induced Ser-toli cell bFGF expression. Endocrinology 1992; 13:2928–2934.
21. Mullaney BP, Skinner MK. Transforming growth factor-beta (beta 1,beta 2, and beta 3) gene expression and action during pubertal devel-opment of the seminiferous tubule: potential role at the onset of sper-matogenesis. Mol Endocrinol 1993; 7:67–76.
22. Marquardt H, Hunkapiller MW, Hood LE, Todaro GJ. Rat transform-ing growth factor type 1: structure and relation to epidermal growthfactor. Science 1984; 223:1079–1082.
23. Massague J, Pandiella A. Membrane-anchored growth factors. AnnuRev Biochem 1993; 62:515–541.
24. Peles E, Yarden Y. Neu and its ligands: from an oncogene to neuralfactors. Bioessays 1993; 15:815–824.
25. Carraway KLR, Sliwkowski MX, Akita R, Platko J V, Guy PM, Nui-jens A, Diamonti AJ, Vandlen RL, Cantley LC, Cerione RA. TheerbB3 gene product is a receptor for heregulin. J Biol Chem 1994;269;14303–14306.
26. Holmes WE, Sliwkowski MX, Akita RW, Henzel WJ, Lee J, Park,JW, Yansura D, Abadi N, Raab H, Lewis GD, Shephard H, Kuang W,Wood W, Goeddel D, Vandlen R. Identification of heregulin, a specificactivator of p185erbB2. Science 1992; 256:1205–1210.
27. Boonstra J, Rijken P, Humbel B, Cremers F, Verkleij A, van Bergenen Henegouwen P. The epidermal growth factor. Cell Biol Int 1995;19:413–430.
28. Carpenter, Cohen S. Epidermal growth factor. J Biol Chem 1990; 265:7709–7712.
29. Prigent SA, Lemoine NR. The type 1 (EGFR-related) family ofgrowth factor receptors and their ligands. Prog Growth Factor Res1992; 4:1–24.
30. Barros EJ, Santos OF, Matsumoto K, Nakamura T, Nigam SK. Dif-ferential tubulogenic and branching morphogenetic activities ofgrowth factors: implications for epithelial tissue development. ProcNatl Acad Sci USA 1995; 92:4412–4416.
31. Ono M, Okamura K, Nakayama Y, Tomita M, Sato Y, Komatsu Y,Kuwano M. Induction of human microvascular endothelial tubularmorphogenesis by human keratinocytes: involvement of transforminggrowth factor-alpha. Biochem Biophys Res Commun 1992; 189:601–609.
32. Rogers SA, Ryan G, Hammerman MR. Metanephric transforminggrowth factor-alpha is required for renal organogenesis in vitro. AmJ Physiol 1992; 262:F533–539.
33. Seth R, Shum L, Wu F, Wuenschell C, Hall FL, Slavkin HC, War-burton D. Role of epidermal growth factor expression in early mouseembryo lung branching morphogenesis in culture: antisense oligo-deoxynucleotide inhibitory strategy. Dev Biol 1993; 158:555–559.
34. Snedeker SM, Brown CF, DiAugustine RP. Expression and functionalproperties of transforming growth factor alpha and epidermal growthfactor during mouse mammary gland ductal morphogenesis. Proc NatlAcad Sci USA 1991; 88:276–280.
35. Taub M, Wang Y, Szczesny TM, Kleinman HK. Epidermal growthfactor or transforming growth factor alpha is required for kidney tub-ulogenesis in matrigel cultures in serum-free medium. Proc Natl AcadSci USA 1990; 87:4002–4006.
36. Warburton D, Seth R, Shum L, Horcher PG, Hall FL, Werb Z, SlavkinHC. Epigenetic role of epidermal growth factor expression and sig-nalling in embryonic mouse lung morphogenesis. Dev Biol 1992; 149:123–133.
37. Mullaney BP, Skinner MK. Transforming growth factor-alpha and epi-dermal growth factor receptor gene expression and action during pu-bertal development of the seminiferous tubule. Mol Endocrinol 1992;6:2103–2113.
38. Radhakrishnan B, Oke BO, Papadopoulos V, DiAugustine RP, Suarez-Quian CA. Characterization of epidermal growth factor in mouse tes-tis. Endocrinology 1992; 131:3091–3099.
39. Skinner MK, Takacs K, Coffey RJ. Transforming growth factor-alpha
gene expression and action in the seminiferous tubule: peritubular cell-Sertoli cell interactions. Endocrinology 1989; 124:845–854.
40. Suarez-Quian CA, Dai MZ, Onoda M, Kriss RM, Dym M. Epidermalgrowth factor receptor localization in the rat and monkey testes. BiolReprod 1989; 41:921–932.
41. Suarez-Quian CA, Niklinski W. Immunocytochemical localization ofthe epidermal growth factor receptor in mouse testis. Biol Reprod1990; 43:1087–1097.
42. Teerds KJ, Rommerts FF, Dorrington JH. Immunohistochemical de-tection of transforming growth factor-alpha in Leydig cells during thedevelopment of the rat testis. Mol Cell Endocrinol 1990; 69:R1–6.
43. Tsutsumi O, Kurachi H, Oka T. A physiological role of epidermalgrowth factor in male reproductive function. Science 1986; 233:975–977.
44. Plowman GD, Green JM, McDonald VL, Neubauer MG, DistecheCM, Todaro GJ, Shoyab M. The amphiregulin gene encodes a novelepidermal growth factor-related protein with tumor-inhibitory activity.Mol Cell Biol 1990; 10:1969–1981.
45. Orr-Urtreger A, Trakhtenbrot L, Ben-Levy R, Wen D, Rechavi G,Lonai P, Yarden Y. Neural expression and chromosomal mapping ofNeu differentiation factor to 8p12-p21. Proc Natl Acad Sci USA 1993;90:1867–1871.
46. Bravo R, Frank R, Blundell PA, Macdonald-Bravo H. Cyclin/PCNAis the auxiliary protein of DNA polymerase-delta. Nature 1987; 326:515–517.
47. Hall PA, Levison DA, Woods AL, Yu CC, Kellock DB, Watkins JA,Barnes DM, Gillett CE, Camplejohn R, Dover R, Waseem N, LaneD. Proliferating cell nuclear antigen (PCNA) immunolocalization inparaffin sections: an index of cell proliferation with evidence of de-regulated expression in some neoplasms. J Pathol 1990; 162:285–294.
48. Prelich G, Tan CK, Kostura M, Mathews MB, So AG, Downey KM,Stillman B. Functional identity of proliferating cell nuclear antigenand a DNA polymerase-delta auxiliary protein. Nature 1987; 326:517–520.
49. Levitzki A, Gazit A. Tyrosine kinase inhibition: an approach to drugdevelopment. Science 1995; 267:1782–1788.
50. Lyall RM, Zilberstein A, Gazit A, Gilon C, Levitzki A, SchlessingerJ. Tyrphostins inhibit epidermal growth factor (EGF)-receptor tyrosinekinase activity in living cells and EGF-stimulated cell proliferation. JBiol Chem 1989; 264:14503–14509.
51. Miettinen PJ, Berger JE, Meneses J, Phung Y, Pedersen RA, WerbZ, Derynck R. Epithelial immaturity and multiorgan failure in micelacking epidermal growth factor receptor. Nature 1995; 376:337–341.
52. Sibilia M, Wagner EF. Strain-dependent epithelial defects in micelacking the EGF receptor [published erratum appears in Science 1995;269(5226):909]. Science 1995; 269:234–238.
53. Threadgill DW, Dlugosz AA, Hansen LA, Tennenbaum T, Lichti U,Yee D, LaMantia C, Mourton T, Herrup K, Harris RC, Barnard J,Yuspa S, Coffey R, Magnuson T. Targeted disruption of mouse EGFreceptor: effect of genetic background on mutant phenotype. Science1995; 269:230–234.
54. Hilscher B, Hilscher W, Bulthoff-Ohnolz B, Kramer U, Birke A, Pel-zer H, Gauss G. Kinetics of gametogenesis. I. Comparative histolog-ical and autoradiographic studies of oocytes and transitional prosper-matogonia during oogenesis and prespermatogenesis. Cell Tissue Res1974; 154:443–470.
55. Oktay K, Schenken RS, Nelson JF. Proliferating cell nuclear antigenmarks the initiation of follicular growth in the rat. Biol Reprod 1995;53:295–301.
56. Liu YC, Marraccino RL, Keng PC, Bambara RA, Lord EM, ChouWG, Zain SB. Requirement for proliferating cell nuclear antigen ex-pression during stages of the Chinese hamster ovary cell cycle. Bio-chemistry 1989; 28:2967–2974.
57. Shivji KK, Kenny MK, Wood RD. Proliferating cell nuclear antigenis required for DNA excision repair. Cell 1992; 69:367–374.
58. Xiong Y, Zhang H, Beach D. D type cyclins associate with multipleprotein kinases and the DNA replication and repair factor PCNA. Cell1992; 71:505–514.
59. Magre S, Jost A. Dissociation between testicular organogenesis andendocrine cytodifferentiation of Sertoli cells. Proc Natl Acad Sci USA1984; 81:7831–7834.
60. Koike S, Noumura T. Cell- and stage-specific expression of basic FGFin the developing rat gonads. Growth Regul 1994; 4:77–81.
61. Jaillard C, Chatelain PG, Saez JM. In vitro regulation of pig Sertolicell growth and function: effects of fibroblast growth factor and so-matomedin-C. Biol Reprod 1987; 37:665–674.

490 LEVINE ET AL.
62. Van Dissel-Emiliani FM, De Boer-Brouwer M, De Rooij DG. Effectof fibroblast growth factor-2 on Sertoli cells and gonocytes in co-culture during the perinatal period. Endocrinology 1996; 137:647–654.
63. Matzuk MM, Kumar TR, Bradley A. Different phenotypes for mice
deficient in either activins or activin receptor type II. Nature 1995;374:356–360.
64. Kaipia A, Toppari J, Huhtaniemi I, Paranko J. Sex difference in theaction of activin-A on cell proliferation of differentiating rat gonad.Endocrinology 1994; 134:2165–2170.





























