Embed Size (px)
Citation preview

Role of the Hsp90 protein chaperone, and of trxG chromatin regulators, as
capacitors of morphological evolution
Giacomo Cavalli, Institute of Human Genetics, CNRS. Montpellier, April 2004
a literature seminar held as part of a teaching course in Genetics, University of Montpellier 2, using as sources the articles:Rutherford, S. L., and Lindquist, S. (1998). Hsp90 as a capacitor for morphological evolution [see comments]. Nature 396, 336-342.And:Sollars, V., Lu, X., Xiao, L., Wang, X., Garfinkel, M. D., and Ruden, D. M. (2003). Evidence for an epigenetic mechanism by which Hsp90 acts as a capacitor for morphological evolution. Nat Genet 33, 70-74.


Introductory text to the subject (from Nature 396, 336-42)
Among the major heat-shock proteins, Hsp90 is unique in its functions. It is not required for the maturation or maintenance of most proteins in vivo. Most of its many identified cellular targets are signal transducers–cell-cycle and developmental regulators whose conformational instability is relevant to their roles as molecular switches. Through low affinity interactions characterized by repeated cycles of binding and release, Hsp90 keeps these unstable signalling proteins poised for activation until they are stabilized by conformational changes associated with signal transduction. Minor changes in amino-acid sequence can have substantial effects on a protein's conformational stability and Hsp90 recognizes structural features common to unstable proteins rather than specific sequence motifs. Thus, individual members of highly homologous protein families, such as steroid-hormone receptors or cyclin-dependent or Src-family kinases, can varygreatly in their dependence on Hsp90.
Studies of yeast illustrate the specificity of Hsp90: at normal temperatures, reductions in Hsp90 levels that have no apparent effects on cell growth or metabolism can completely abolish signalling through Hsp90-dependent pathways. Conditions that cause general protein damage can divert Hsp90 from its normal targets to other partially denatured proteins. Because of its dual involvement with inherently unstable signal transducers on the one hand, and with the cellular response to stress on the other, Hsp90 may link developmental programs to environmental contingency.
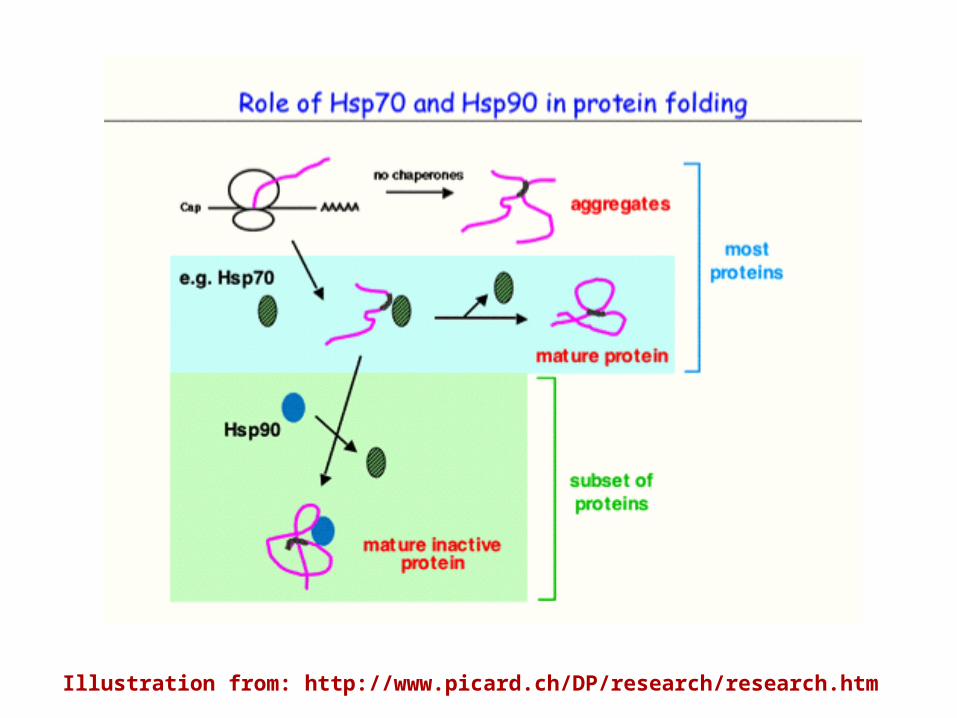
Illustration from: http://www.picard.ch/DP/research/research.htm
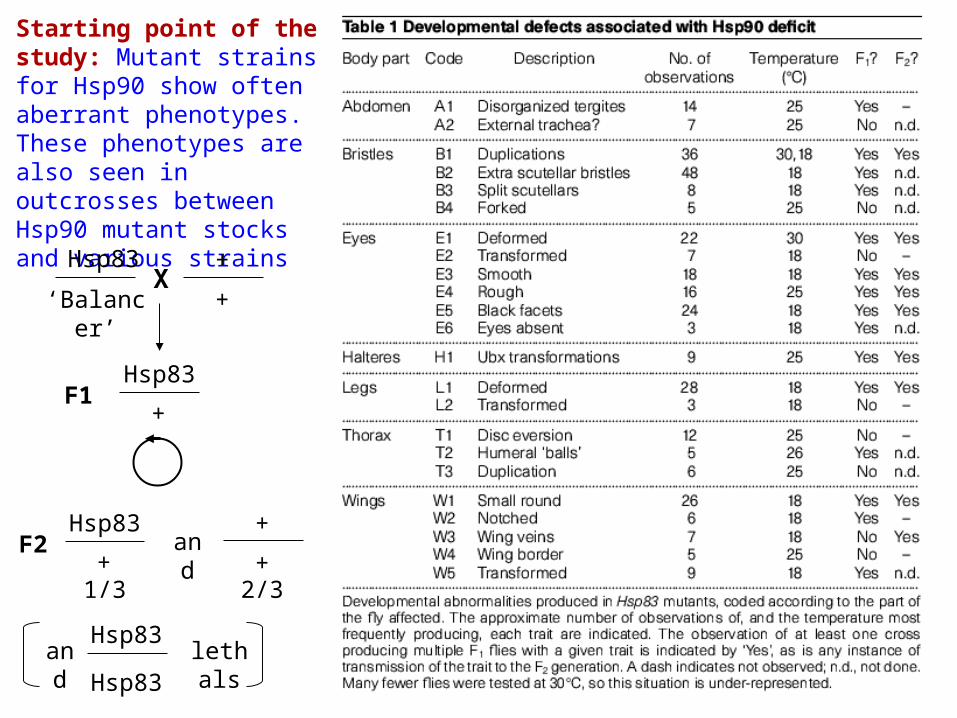
Starting point of the study: Mutant strains for Hsp90 show often aberrant phenotypes. These phenotypes are also seen in outcrosses between Hsp90 mutant stocks and various strains
Hsp83
‘Balancer’X
+
+
Hsp83
+
Hsp83
+ +
+
F1
F2 and
1/3 2/3
andHsp83
Hsp83lethals

Examples of phenotypes
Mutant Hsp83 stocks
Crosses between Hsp83 mutants and laboratory stocks (phenotypes are often worse than in established Hsp83 stocks)
Effect of the Hsp90 inhibitor geldanamycin
« Bad » phenotypes observed in double Hsp90 mutants (heteroallelic viable combinations)
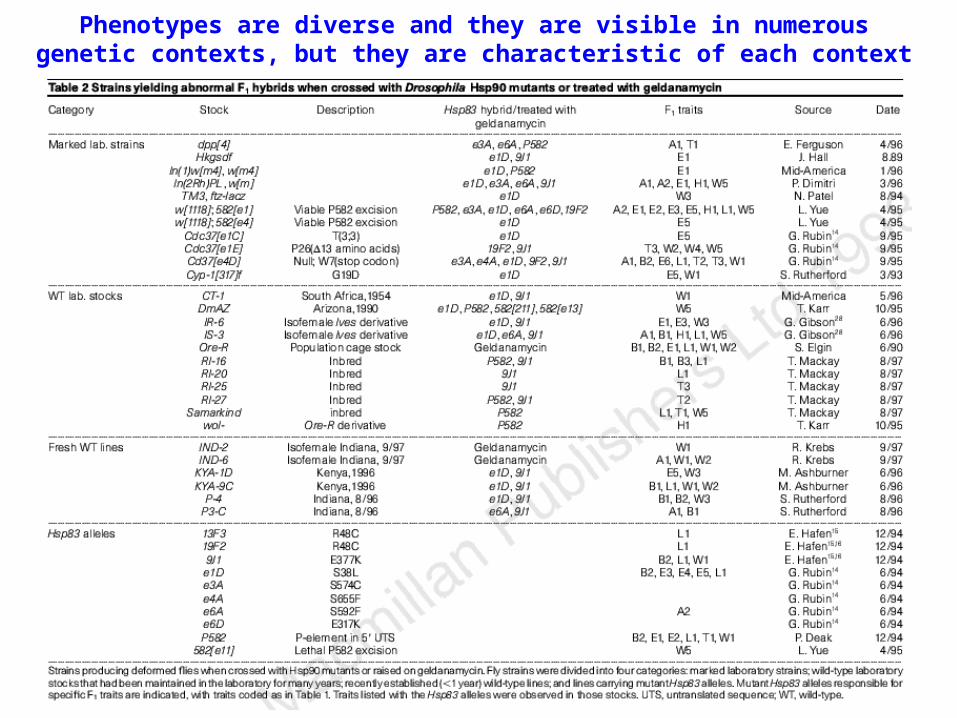
Phenotypes are diverse and they are visible in numerous genetic contexts, but they are characteristic of each context

Origin of the phenotypes?
They depend on Hsp90, since they can be observed in several independent mutant alleles, they get worse in general in the presence of two mutant alleles (heteroallelism), and since they are also induced by the Hsp90 inhibitor geldanamycin
In Hsp90 mutant context, development might become more sensitive to environmental noise
Alternatively, Hsp90 might induce an increase in the global mutation rate
Finally, Hsp90 might buffer against phenotypic variation dependent on the accumulation of genetic polymorphism. In case of a decrease of Hsp90 levels, this cryptic variation might become exposed and thus originate visible phenotypes. The previous data seem to suggest this third hypothesis, since the observed phenotypes depend on genetic background of the tested strains. This seems to exclude a global increase in mutation or an increased sensitivity to environmentally induced stress, since both conditions should result in a more random distribution of phenotypes
Mechanism?
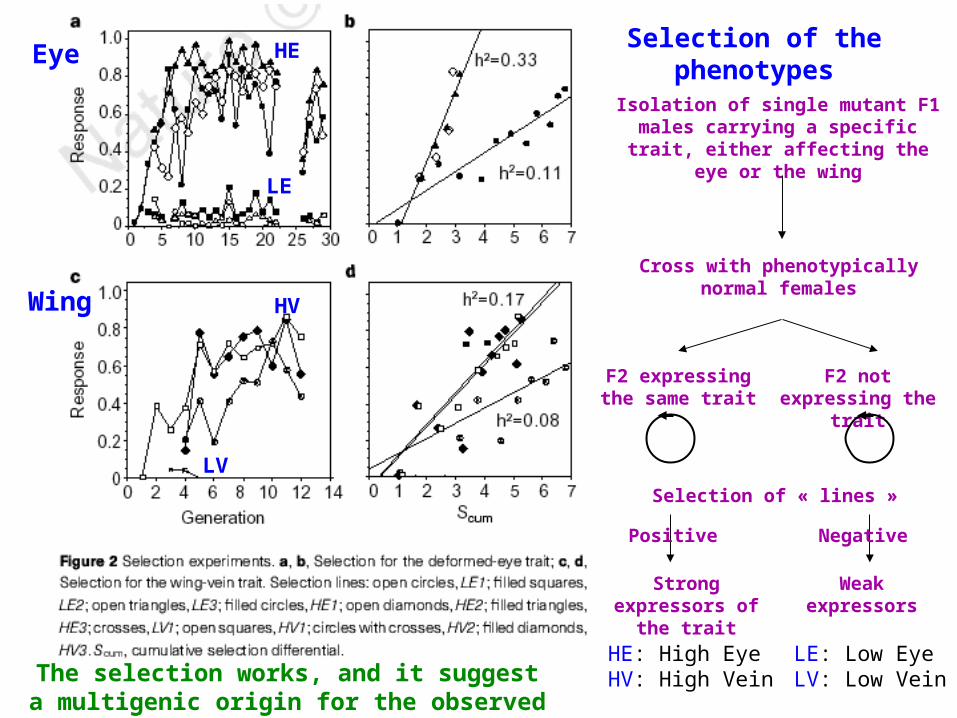
Selection of the phenotypes
Isolation of single mutant F1 males carrying a specific trait, either affecting the eye or the wing
Cross with phenotypically normal females
F2 expressing the same trait
F2 not expressing the trait
Selection of « lines »
Strong expressors of the trait
Weak expressors
HE: High Eye HV: High Vein
LE: Low EyeLV: Low Vein
Eye
Wing
HE
LE
Positive Negative
HV
LV
The selection works, and it suggest a multigenic origin for the observed traits!
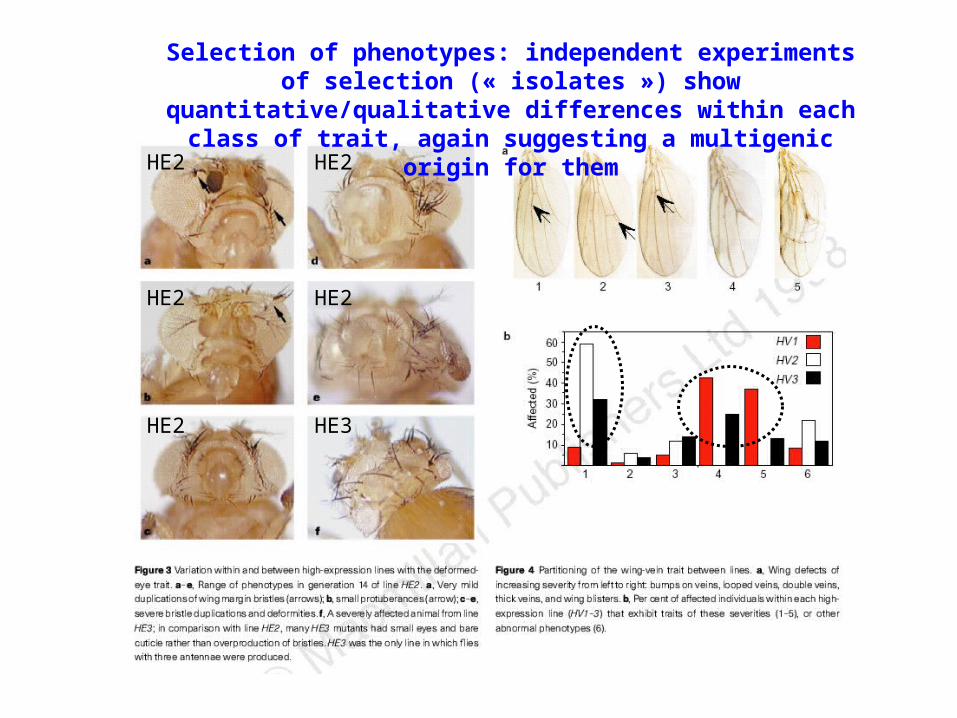
HE2
HE2
HE2
HE2
HE2
HE3
Selection of phenotypes: independent experiments of selection (« isolates ») show quantitative/qualitative differences within each class
of trait, again suggesting a multigenic origin for them
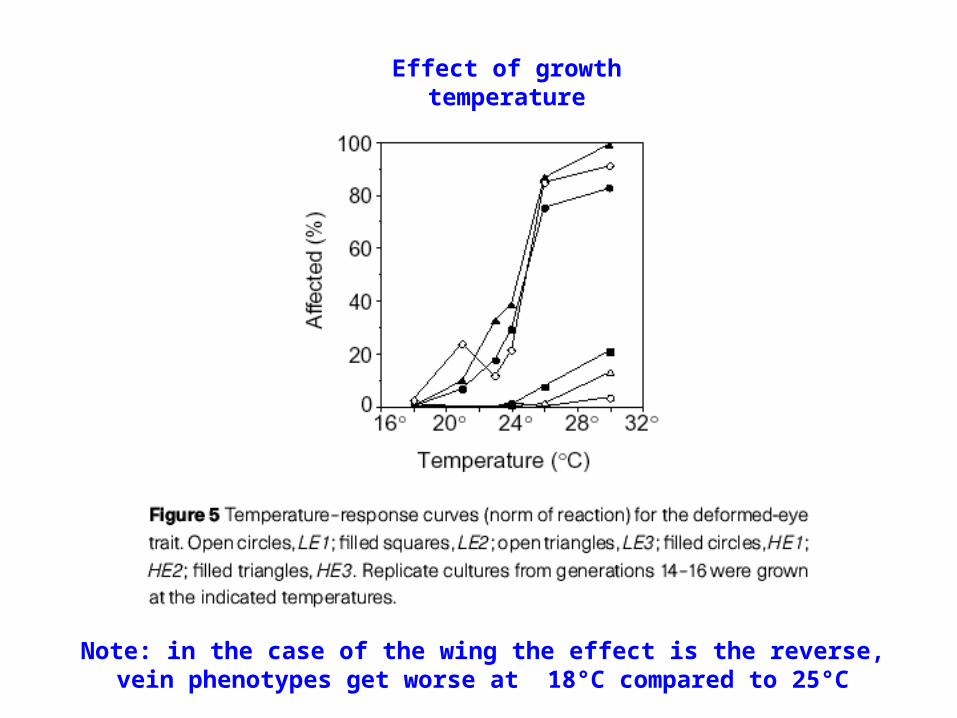
Effect of growth temperature
Note: in the case of the wing the effect is the reverse, vein phenotypes get worse at 18°C compared to 25°C

Genetic determinants of the observed effectsIt is possible to design crosses allowing to isolate fly populations carrying any given chromosome (in one or two copies) from a specific genetic background, either the background corresponding to a HE “stock” or a control non-selected background. This allows comparing the percentage of
phenotypes obtained in each genetic background to the control (HE) frequency
The analysis shows that the phenotypes have at least two determinants on chromosome 2, and at least one on chromosome 3

Genotyping the phenotypically affected flies: does maintenance of the phenotypes requires continuous impairment of Hsp90?
Since selection was only based on phenotypes, but not on the
maintenance of the Hsp90 mutant chromosome, an obvious question was whether Hsp90 was always mutated in
the individuals carrying the traits
Note that out of the F1 cross, only 2/3 des mouches should carry the
mutation in the F2 and progressively less in the subsequent generations, while selection obtains up to 80% of
flies carrying the trait
For this reason they genotyped HE and LE flies
An example of how this is done is shown here for the starting strains
All tested flies (i.e. ten to fourty in different conditions) had lost the mutation !!!
Inactivation of Hsp90 required to initiate, but not to maintain, development of new traits!

Proposed model to explain the action of Hsp90 (extracted from Ref. 7, see the last slide of this document). Hsp90 might favour the accumulation of genetic variation that would normally remain cryptic thanks to the Hsp90-dependent buffering ability. In situations
impairing Hsp90 function (dependent on environmental stress or on mutation of the Hsp90 coding gene), this variation could become exposed and, if not deleterious, might be
subjected to selective pressure. This may ultimately lead to rapid fixation of phenotypic and genotypic variation that might result in the emergence of novel species

…but there are other data to be considered…
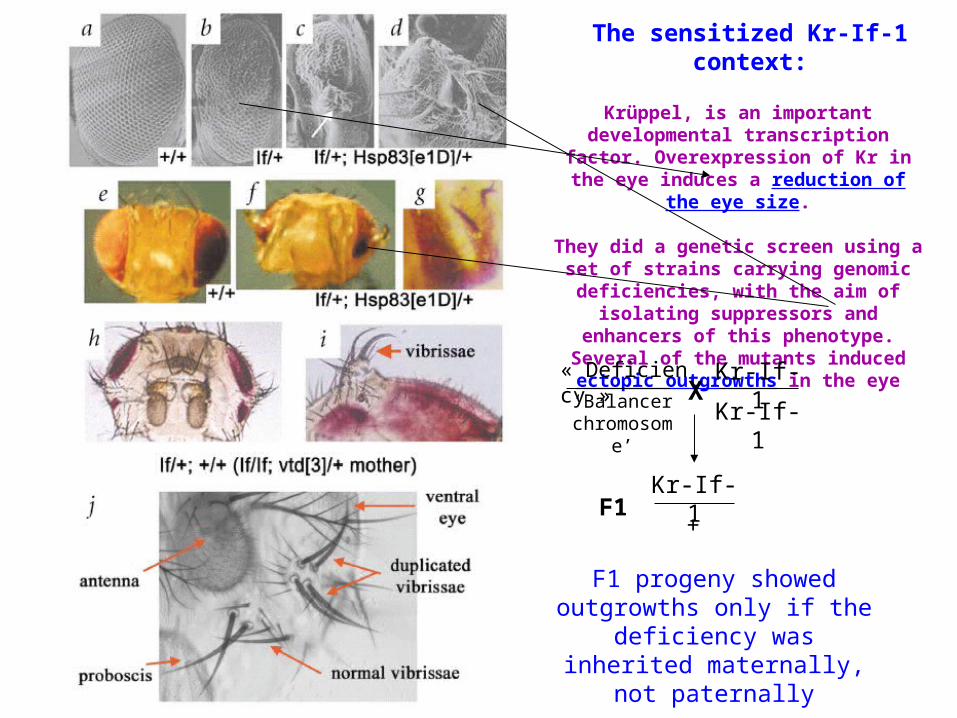
The sensitized Kr-If-1 context:
Krüppel, is an important developmental transcription factor. Overexpression of Kr in the eye induces a reduction of the eye
size.
They did a genetic screen using a set of strains carrying genomic deficiencies,
with the aim of isolating suppressors and enhancers of this phenotype. Several of the mutants induced ectopic outgrowths
in the eye
« Deficiency »
‘Balancer chromosome’
XKr-If-1
+F1
Kr-If-1
Kr-If-1
F1 progeny showed outgrowths only if the deficiency was inherited
maternally, not paternally
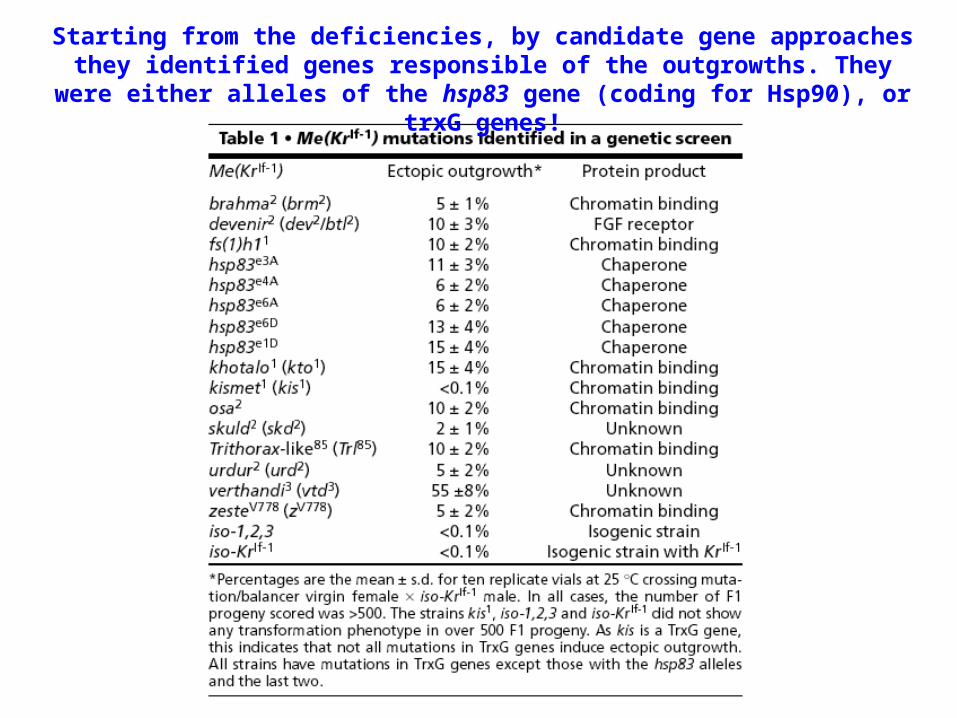
Starting from the deficiencies, by candidate gene approaches they identified genes responsible of the outgrowths. They were either alleles of the hsp83 gene (coding for
Hsp90), or trxG genes!
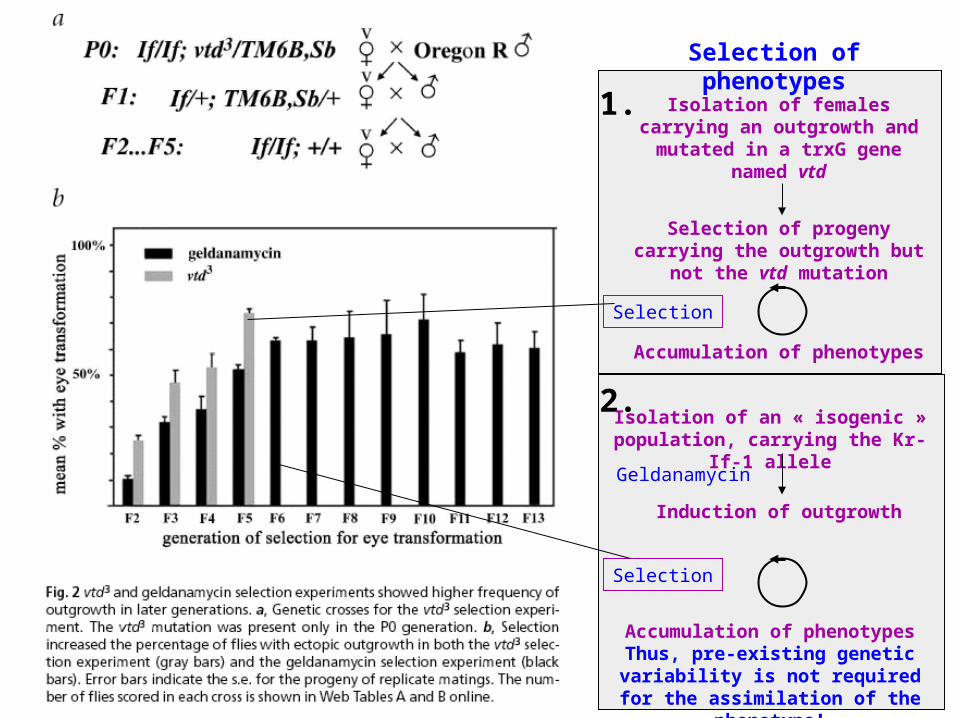
Selection of phenotypes
Isolation of females carrying an outgrowth and mutated in a trxG
gene named vtd
Selection of progeny carrying the outgrowth but not the vtd
mutation
Accumulation of phenotypes
1.
2.Isolation of an « isogenic »
population, carrying the Kr-If-1 allele
Induction of outgrowth
Accumulation of phenotypesThus, pre-existing genetic variability is not required for the assimilation of
the phenotype!
Geldanamycin
Selection
Selection
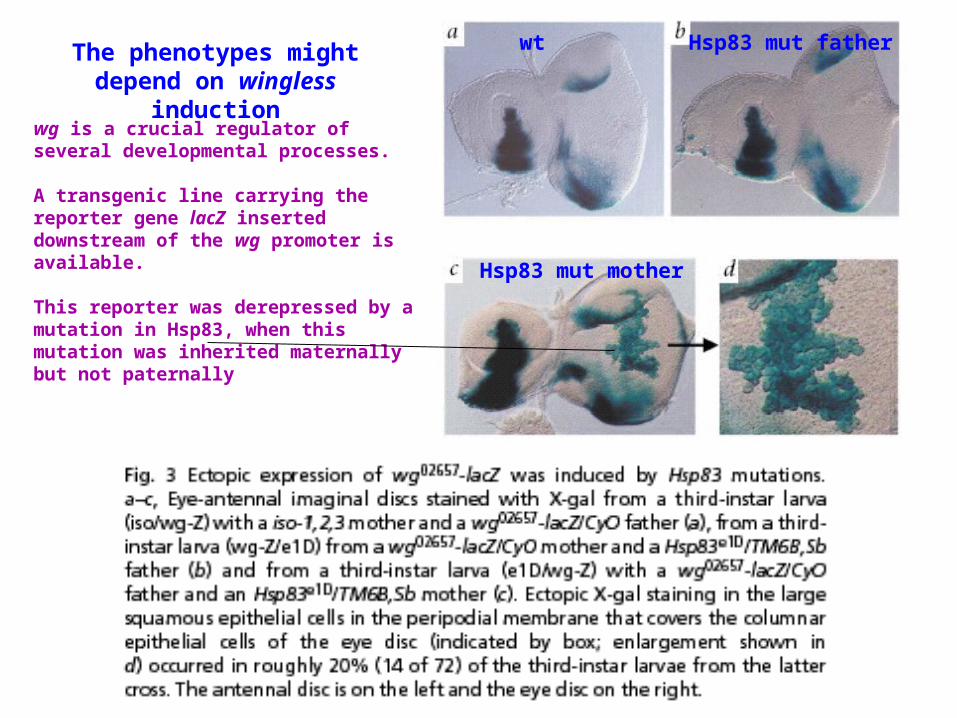
The phenotypes might depend on wingless induction
wg is a crucial regulator of several developmental processes.
A transgenic line carrying the reporter gene lacZ inserted downstream of the wg promoter is available.
This reporter was derepressed by a mutation in Hsp83, when this mutation was inherited maternally but not paternally
wt Hsp83 mut father
Hsp83 mut mother
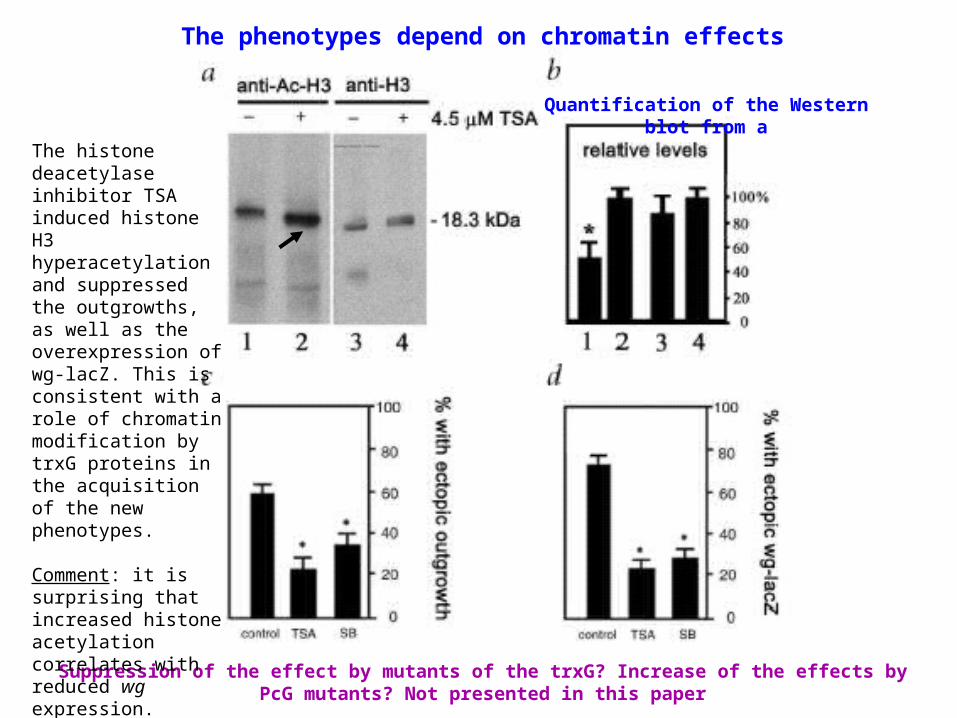
The phenotypes depend on chromatin effects
Suppression of the effect by mutants of the trxG? Increase of the effects by PcG mutants? Not presented in this paper
Quantification of the Western blot from a
The histone deacetylase inhibitor TSA induced histone H3 hyperacetylation and suppressed the outgrowths, as well as the overexpression of wg-lacZ. This is consistent with a role of chromatin modification by trxG proteins in the acquisition of the new phenotypes.
Comment: it is surprising that increased histone acetylation correlates with reduced wg expression.

Mother Nature’s « Tools » contributing to buffering of environmental variation and, at the same time, to genomic and phenotypic plasticity, might
play an important role for evolution of new species
Chromatin inheritance: the same DNA sequence can adopt different functional states, that are heritable during cell division and through mitosis (from yeast to mouse, maybe human, see refs 1, 2 and 4).
Stabilisation of homeostasis. Buffering of environmental changes. Since cells are capable of transmitting the memory of their identity to their progeny via chromatin inheritance in addition gene regulatory mechanisms based on the relative concentration of regulatory factors, this redundancy provides robustness to gene regulation and make it relatively insensitive to environmental changes.
Hsp90. This chaperone protein allows to key regulatory proteins to remain functional even upon mild mutations, since it favours their correct folding. Thus, mild mutations are tolerated provided Hsp90 is functional.
Buffers development from genetic or environmental sources of variability
Might induce a strong morphological variability when its activity is reduced by mutations or by strong stresses that divert this protein from its regular targets (ref 3, 5).
If among the new phenotypes there were favourable ones, natural selection might fix them in natural populations
Cooperation between Hsp90 and chromatin factors (ref 6) may contribute to rapid fixation of new traits in a population

Relevant bibliography:
1) Grewal, S. I. S., and Klar, A. J. S. (1996). Chromosomal inheritance of epigenetic states in fission yeast during mitosis and meiosis. Cell 86, 95-101.
2) Cavalli, G., and Paro, R. (1998). The Drosophila Fab-7 chromosomal element conveys epigenetic inheritance during mitosis and meiosis. Cell 93, 505-518.
3) Rutherford, S. L., and Lindquist, S. (1998). Hsp90 as a capacitor for morphological evolution [see comments]. Nature 396, 336-342.
4) Morgan, H. D., Sutherland, H. G., Martin, D. I., and Whitelaw, E. (1999). Epigenetic inheritance at the agouti locus in the mouse. Nat Genet 23, 314-318.
5) Queitsch C, Sangster TA, Lindquist S (2002) Hsp90 as a capacitor of phenotypic variation. Nature 417, 618-24. Epub 2002 May 12
6) Sollars, V., Lu, X., Xiao, L., Wang, X., Garfinkel, M. D., and Ruden, D. M. (2003). Evidence for an epigenetic mechanism by which Hsp90 acts as a capacitor for morphological evolution. Nat Genet 33, 70-74.
7) Sangster TA, Lindquist S, Queitsch C (2004). Under cover: causes, effects and implications of Hsp90-mediated genetic capacitance. Bioessays. 26, 348-62





![Hsp90-Targeted Library - Chemdiv · HtpG (high-temperature protein G), whereas Archaebacteria lack a Hsp90 representative [24]. All eukaryotes possess cytosolic members, called Hsp90](https://img.pdfslide.us/doc/110x75/5c687a8609d3f2f5638b9b2b/hsp90-targeted-library-htpg-high-temperature-protein-g-whereas-archaebacteria.jpg)






















