Embed Size (px)
Citation preview

1
Aortic valve calcification is the leading cause of aortic valve stenosis, which is a progressive disease that results
in heart failure, angina, syncope, and death. The prevalence of aortic valve stenosis is ≈0.2% among adults between the ages of 50 and 59 years but increases to 2.8% in adults ≥75 years of age. Aortic valve stenosis does not cause symptoms for long periods of time, but the rate of death is >50% at 2 years when patients become symptomatic unless aortic valve replacement is performed.1,2 It is now recognized that aortic valve calcifica-tion is not a passive process of calcium deposition that occurs with advanced age, but, instead, it is an active process that involves valve, circulating, and bone marrow–derived cells along with biochemical and mechanical factors.3,4
Wnts are lipid-modified glycosylated proteins encoded by 19 genes. They bind to extracellular Frizzled receptors leading to activation of multiple signaling pathways that are diverted into the main branches: the classical or canonical
β-catenin–dependent pathway and the β-catenin–independent pathways, collectively known as the noncanonical path-ways.5–8 Wnt signaling pathways have multiple functions in heart valves in the regulation of endothelial-to-mesenchymal transition and cell proliferation during the initial stages of endocardial cushion and heart valve formation.9,10 There is strong evidence suggesting the involvement of canonical Wnt signaling cascade in valve calcification.11 Caira et al12 found that Lrp5 (low-density lipoprotein receptor–related protein 5)/Wnt3 signaling markers were highly expressed in human calcified aortic valves. Despite the fact that there is evidence implicating Wnt signaling in osteogenic differentiation and development, little is known about the role of noncanoni-cal Wnt ligands, such as WNT5a, WNT5b, and WNT11, in valvular calcification. The prototypical noncanonical ligand WNT5a has been implicated in the development of atheroscle-rotic plaques, such as in smooth muscle cells and macrophage
© 2016 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.116.308394
Objective—The mechanisms underlying the pathogenesis of aortic valve calcification remain unclear. With accumulating evidence demonstrating that valve calcification recapitulates bone development, the crucial roles of noncanonical Wnt ligands WNT5a, WNT5b, and WNT11 in osteogenesis make them critical targets in the study of aortic valve calcification.
Approach and Results—Using immunohistochemistry, RT-PCR, Western blotting, and tissue culture, we examined the tissue distribution of WNT5a, WNT5b, and WNT11 in noncalcified and calcified aortic valves and their effects on human aortic valve interstitial cells (HAVICs). Only focal strong immunostaining for WNT5a was seen in and around areas of calcification. Abundant immunostaining for WNT5b and WNT11 was seen in inflammatory cells, fibrosis, and activated myofibroblasts in areas of calcified foci. There was significant correlation between WNT5b and WNT11 overall staining and presence of calcification, lipid score, fibrosis, and microvessels (P<0.05). RT-PCR and Western blotting revealed abundant expression of both Wnts in stenotic aortic valves, particularly in bicuspid valves. Incubation of HAVICs from noncalcified valves with the 3 noncanonical Wnts significantly increased cell apoptosis and calcification (P<0.05). Treatment of HAVICs with the mitogen-activated protein kinase-38β and GSK3β inhibitors significantly reduced their mineralization (P<0.01). Raman spectroscopy identified the inorganic phosphate deposits as hydroxyapatite and showed a significant increase in hydroxyapatite deposition in HAVICs in response to WNT5a and WNT11 (P<0.05). Similar crystallinity was seen in the deposits found in HAVICs treated with Wnts and in calcified human aortic valves.
Conclusions—These findings suggest a potential role for noncanonical Wnt signaling in the pathogenesis of aortic valve calcification. (Arterioscler Thromb Vasc Biol. 2017;37:00-00. DOI: 10.1161/ATVBAHA.116.308394.)
Key Words: aortic valve ◼ bone marrow ◼ calcium ◼ human ◼ lipids
Received on: February 19, 2016; final version accepted on: November 28, 2016.From the Division of Cardiology and Division of Cardiac Surgery, McGill University Health Centre, Montreal, Quebec, Canada (I.A., B.Y., H.A.-K.,
B.B., L.O., M.A.-R., B.d.V., D.S.-T., A.S.); Department of Material Engineering, McGill University, Montreal, Quebec, Canada (M.C., O.G.); and Montreal Heart Institute, Université de Montréal, Montreal, Quebec, Canada (E.R., J.C.T.).
*These authors contributed equally to this article.The online-only Data Supplement is available with this article at http://atvb.ahajournals.org/lookup/suppl/doi:10.1161/ATVBAHA.116.308394/-/DC1.Correspondence to Adel Schwertani, PhD, DM, McGill University Health Centre, Glen EM12224, 1001 Blvd Decarie, Montreal, H4A 3J1, Quebec,
Canada. E-mail [email protected]
Role of Noncanonical Wnts Signaling Pathway in Human Aortic Valve Calcification
Isabella Albanese,* Bin Yu,* Hamood Al-Kindi, Bianca Barratt, Leah Ott, Mohammad Al-Refai, Benoit de Varennes, Dominique Shum-Tim, Marta Cerruti, Ophélie Gourgas, Eric Rhéaume,
Jean-Claude Tardif, Adel Schwertani
Original Research
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
by guest on M
arch 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from

2 Arterioscler Thromb Vasc Biol February 2017
infiltration.13–15 Other recent work supporting the involvement of WNT5a, coreceptor LRP6, and canonical Wnt signaling in vascular calcification merits the investigation of other non-canonical Wnt ligands and their roles in stenotic valve dis-ease.16–18 In this study, we hypothesized that the noncanonical Wnt ligands WNT5a, WNT5b, and WNT11 are upregulated in stenotic aortic valves and may contribute to the pathogen-esis of the disease. We, therefore, examined cellular and total protein expression of WNT5a, WNT5b, and WNT11 in aor-tic valves by immunohistochemistry and Western blotting. We also examined the relative mRNA expression of WNT11, WNT5a, and WNT5b in calcified aortic valves. We further isolated human aortic valve interstitial cells (HAVICs) from noncalcified and calcified aortic valves and assessed the effect of WNT5a, WNT5b, and WNT11 on cell mineralization using Alizarin Red S staining and Raman spectroscopy and deter-mined gene expression of osteogenic mediators.
Materials and MethodsMaterials and Methods are available in the online-only Data Supplement.
ResultsExpression of Noncanonical Wnt mRNAs and Protein in Human Aortic Valves and HAVICsThere were significantly higher expressions of WNT5a, WNT5b, and WNT11 mRNAs in calcified aortic valves com-pared with normal valves (Figure 1A; P<0.05).
The mRNA expression of WNT5b was significantly higher in calcified bicuspid aortic valves compared with calcified tri-cuspid aortic valves (Figure 1B; P<0.05). The mRNA expres-sion of WNT5a and WNT11 were higher in calcified bicuspid valves compared with calcified tricuspid aortic valves, but it did not reach statistical significance (Figure 1B).
Western blot results showed robust WNT5b expression in cultured HAVICs isolated from stenotic aortic valves and ste-notic aortic valve tissues and no detectable expression in normal dermal fibroblast cells. Although there is a WNT5b signal at the predicted size of 37 kDa, there is also strong signal at 75 kDa, suggesting the occurrence of post-translational modifications or potential dimerization of the Wnt ligands (Figure 1C). Cell lysate from HEK293 cells revealed a similar expression pattern to what was observed in human aortic valve tissue (data not shown). Both aortic valve tissues and cultured HAVICs show abundant WNT11 protein expression (Figure 1D).
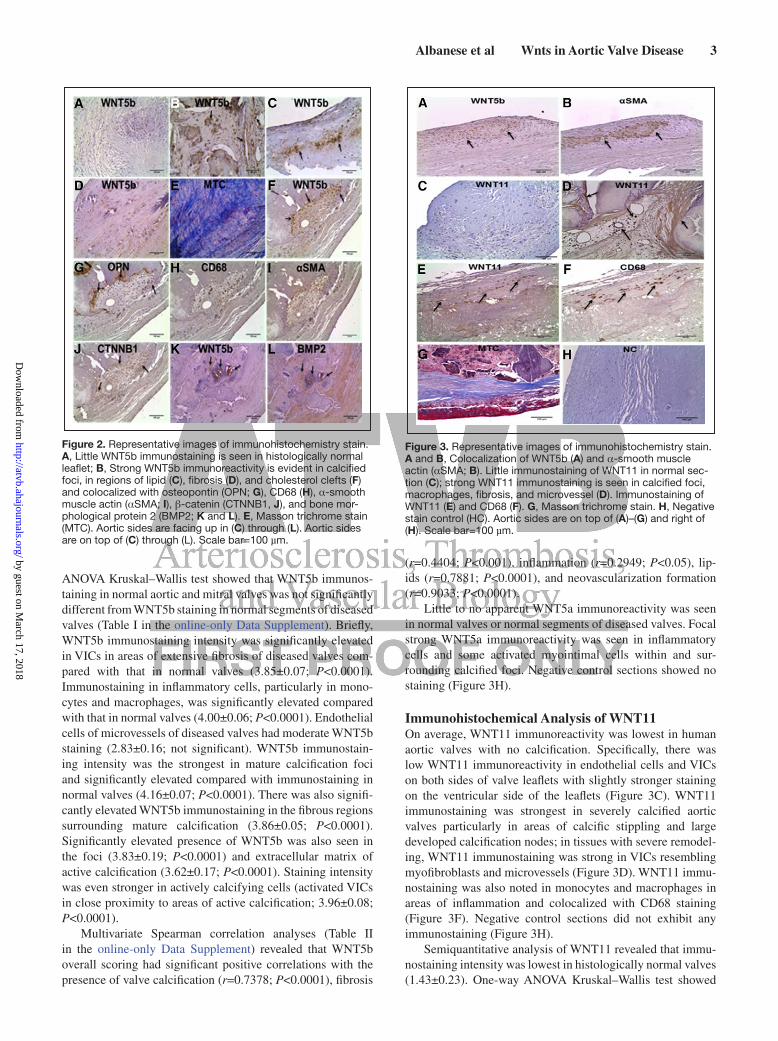
Immunohistochemical Analysis of WNT5a and WNT5bThere was little immunoreactivity for WNT5b in histo-logically normal valves. Specifically, low immunoreactivity
was evident in valvular interstitial cells (VICs) and endo-thelial cells on both aortic and ventricular sides of the leaf-let (Figure 2A). WNT5b immunostaining was strongest in severely calcified valves, with immunoreactivity in VICs on both the aortic and ventricular sides of valve leaflet and in areas of calcific stippling and large developed calcification nodes (Figure 2B). Strong WNT5b immunostaining was also evident in macrophages in areas of inflammation (Figure 2B) and in region of lipid (Figure 2C). WNT5b immunostaining was strong in severely remodeled valves. In these tissues, WNT5b immunoreactivity was abundantly seen in VICs in areas of extensive fibrosis (Figure 2D and 2E) and valve thick-ening (Figure 3A and 3B). Although WNT5b immunostaining was found in quiescent VICs, immunoreactivity was stronger in activated VICs and in osteoblasts (Figure 2F) and colocal-ized with osteopontin (Figure 2G), β-catenin (Figure 2J), bone morphological protein 2 (Figure 2 L), CD68 (Figure 2H), and α-smooth muscle actin (αSMA; Figures 2I and 3B).
Semiquantitative analyses of the immunohistochemi-cal data revealed that WNT5b immunostaining intensity was lowest in histologically normal valves (2.62±0.18). One-way
Nonstandard Abbreviations and Acronyms
ALP alkaline phosphatase
HAVICs human aortic valve interstitial cells
OSM osteogenic medium
VICs valve interstitial cells
Figure 1. A, mRNA expression of noncanonical Wnt ligands in human aortic valves. Wnt5a, Wnt5b, and Wnt11 mRNA expres-sion (normalized to GAPDH) are elevated in calcified aortic valves compared with normal aortic valves. Relative normalized expression level is presented as mean±SEM (*P<0.05). B, mRNA expression of Wnt ligands in human bicuspid and tricuspid aortic valves. mRNA expression of Wnt5b and Wnt11 (normalized to GAPDH) is elevated in bicuspid aortic valves compared with that in tricuspid aortic valves. Relative normalized expression level is presented as mean±SEM (*P<0.05). C, WNT5b protein detec-tion. Left: Human dermal fibroblast (DF) cells, human aortic valve interstitial cells (HAVICs), and calcified aortic valves (CAV). Right: CAV and noncalcified aortic valves (NCAVs). D, WNT11 protein detection. Left: human DF cells, HAVICs, and CAV. Right: CAV and NCAVs.
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
Albanese et al Wnts in Aortic Valve Disease 3
ANOVA Kruskal–Wallis test showed that WNT5b immunos-taining in normal aortic and mitral valves was not significantly different from WNT5b staining in normal segments of diseased valves (Table I in the online-only Data Supplement). Briefly, WNT5b immunostaining intensity was significantly elevated in VICs in areas of extensive fibrosis of diseased valves com-pared with that in normal valves (3.85±0.07; P<0.0001). Immunostaining in inflammatory cells, particularly in mono-cytes and macrophages, was significantly elevated compared with that in normal valves (4.00±0.06; P<0.0001). Endothelial cells of microvessels of diseased valves had moderate WNT5b staining (2.83±0.16; not significant). WNT5b immunostain-ing intensity was the strongest in mature calcification foci and significantly elevated compared with immunostaining in normal valves (4.16±0.07; P<0.0001). There was also signifi-cantly elevated WNT5b immunostaining in the fibrous regions surrounding mature calcification (3.86±0.05; P<0.0001). Significantly elevated presence of WNT5b was also seen in the foci (3.83±0.19; P<0.0001) and extracellular matrix of active calcification (3.62±0.17; P<0.0001). Staining intensity was even stronger in actively calcifying cells (activated VICs in close proximity to areas of active calcification; 3.96±0.08; P<0.0001).
Multivariate Spearman correlation analyses (Table II in the online-only Data Supplement) revealed that WNT5b overall scoring had significant positive correlations with the presence of valve calcification (r=0.7378; P<0.0001), fibrosis
(r=0.4404; P<0.001), inflammation (r=0.2949; P<0.05), lip-ids (r=0.7881; P<0.0001), and neovascularization formation (r=0.9033; P<0.0001).
Little to no apparent WNT5a immunoreactivity was seen in normal valves or normal segments of diseased valves. Focal strong WNT5a immunoreactivity was seen in inflammatory cells and some activated myointimal cells within and sur-rounding calcified foci. Negative control sections showed no staining (Figure 3H).
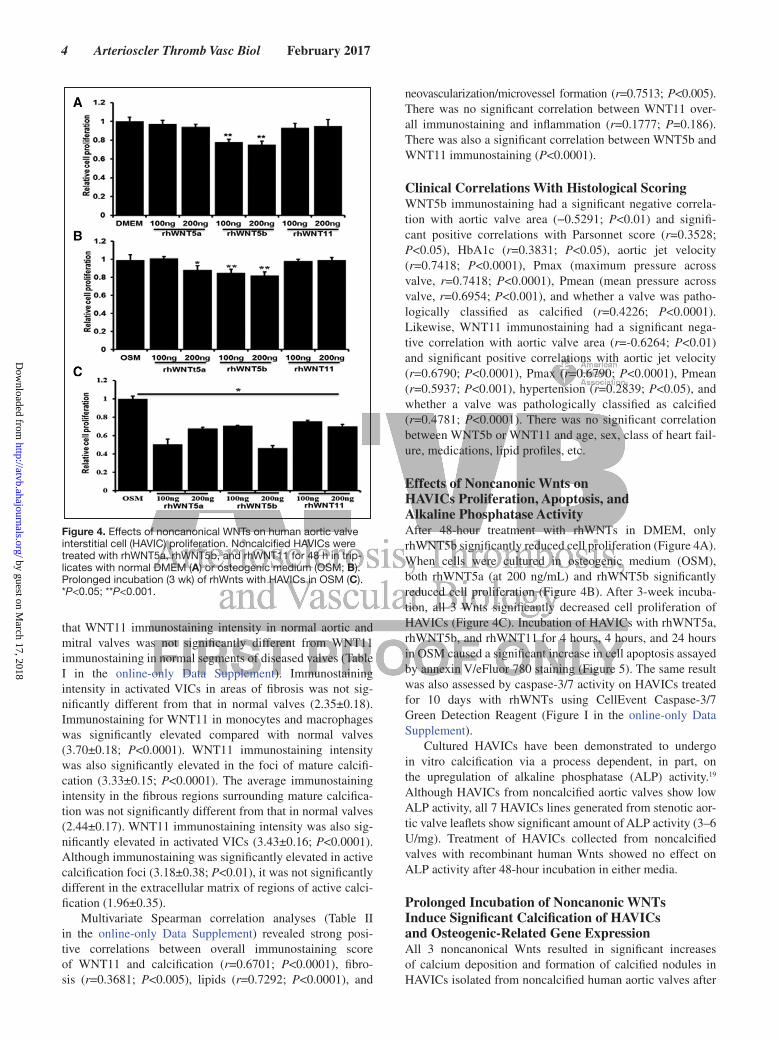
Immunohistochemical Analysis of WNT11On average, WNT11 immunoreactivity was lowest in human aortic valves with no calcification. Specifically, there was low WNT11 immunoreactivity in endothelial cells and VICs on both sides of valve leaflets with slightly stronger staining on the ventricular side of the leaflets (Figure 3C). WNT11 immunostaining was strongest in severely calcified aortic valves particularly in areas of calcific stippling and large developed calcification nodes; in tissues with severe remodel-ing, WNT11 immunostaining was strong in VICs resembling myofibroblasts and microvessels (Figure 3D). WNT11 immu-nostaining was also noted in monocytes and macrophages in areas of inflammation and colocalized with CD68 staining (Figure 3F). Negative control sections did not exhibit any immunostaining (Figure 3H).
Semiquantitative analysis of WNT11 revealed that immu-nostaining intensity was lowest in histologically normal valves (1.43±0.23). One-way ANOVA Kruskal–Wallis test showed
Figure 2. Representative images of immunohistochemistry stain. A, Little WNT5b immunostaining is seen in histologically normal leaflet; B, Strong WNT5b immunoreactivity is evident in calcified foci, in regions of lipid (C), fibrosis (D), and cholesterol clefts (F) and colocalized with osteopontin (OPN; G), CD68 (H), α-smooth muscle actin (αSMA; I), β-catenin (CTNNB1, J), and bone mor-phological protein 2 (BMP2; K and L). E, Masson trichrome stain (MTC). Aortic sides are facing up in (C) through (L). Aortic sides are on top of (C) through (L). Scale bar=100 µm.
Figure 3. Representative images of immunohistochemistry stain. A and B, Colocalization of WNT5b (A) and α-smooth muscle actin (αSMA; B). Little immunostaining of WNT11 in normal sec-tion (C); strong WNT11 immunostaining is seen in calcified foci, macrophages, fibrosis, and microvessel (D). Immunostaining of WNT11 (E) and CD68 (F). G, Masson trichrome stain. H, Negative stain control (HC). Aortic sides are on top of (A)–(G) and right of (H). Scale bar=100 µm.
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
4 Arterioscler Thromb Vasc Biol February 2017
that WNT11 immunostaining intensity in normal aortic and mitral valves was not significantly different from WNT11 immunostaining in normal segments of diseased valves (Table I in the online-only Data Supplement). Immunostaining intensity in activated VICs in areas of fibrosis was not sig-nificantly different from that in normal valves (2.35±0.18). Immunostaining for WNT11 in monocytes and macrophages was significantly elevated compared with normal valves (3.70±0.18; P<0.0001). WNT11 immunostaining intensity was also significantly elevated in the foci of mature calcifi-cation (3.33±0.15; P<0.0001). The average immunostaining intensity in the fibrous regions surrounding mature calcifica-tion was not significantly different from that in normal valves (2.44±0.17). WNT11 immunostaining intensity was also sig-nificantly elevated in activated VICs (3.43±0.16; P<0.0001). Although immunostaining was significantly elevated in active calcification foci (3.18±0.38; P<0.01), it was not significantly different in the extracellular matrix of regions of active calci-fication (1.96±0.35).
Multivariate Spearman correlation analyses (Table II in the online-only Data Supplement) revealed strong posi-tive correlations between overall immunostaining score of WNT11 and calcification (r=0.6701; P<0.0001), fibro-sis (r=0.3681; P<0.005), lipids (r=0.7292; P<0.0001), and
neovascularization/microvessel formation (r=0.7513; P<0.005). There was no significant correlation between WNT11 over-all immunostaining and inflammation (r=0.1777; P=0.186). There was also a significant correlation between WNT5b and WNT11 immunostaining (P<0.0001).
Clinical Correlations With Histological ScoringWNT5b immunostaining had a significant negative correla-tion with aortic valve area (−0.5291; P<0.01) and signifi-cant positive correlations with Parsonnet score (r=0.3528; P<0.05), HbA1c (r=0.3831; P<0.05), aortic jet velocity (r=0.7418; P<0.0001), Pmax (maximum pressure across valve, r=0.7418; P<0.0001), Pmean (mean pressure across valve, r=0.6954; P<0.001), and whether a valve was patho-logically classified as calcified (r=0.4226; P<0.0001). Likewise, WNT11 immunostaining had a significant nega-tive correlation with aortic valve area (r=-0.6264; P<0.01) and significant positive correlations with aortic jet velocity (r=0.6790; P<0.0001), Pmax (r=0.6790; P<0.0001), Pmean (r=0.5937; P<0.001), hypertension (r=0.2839; P<0.05), and whether a valve was pathologically classified as calcified (r=0.4781; P<0.0001). There was no significant correlation between WNT5b or WNT11 and age, sex, class of heart fail-ure, medications, lipid profiles, etc.
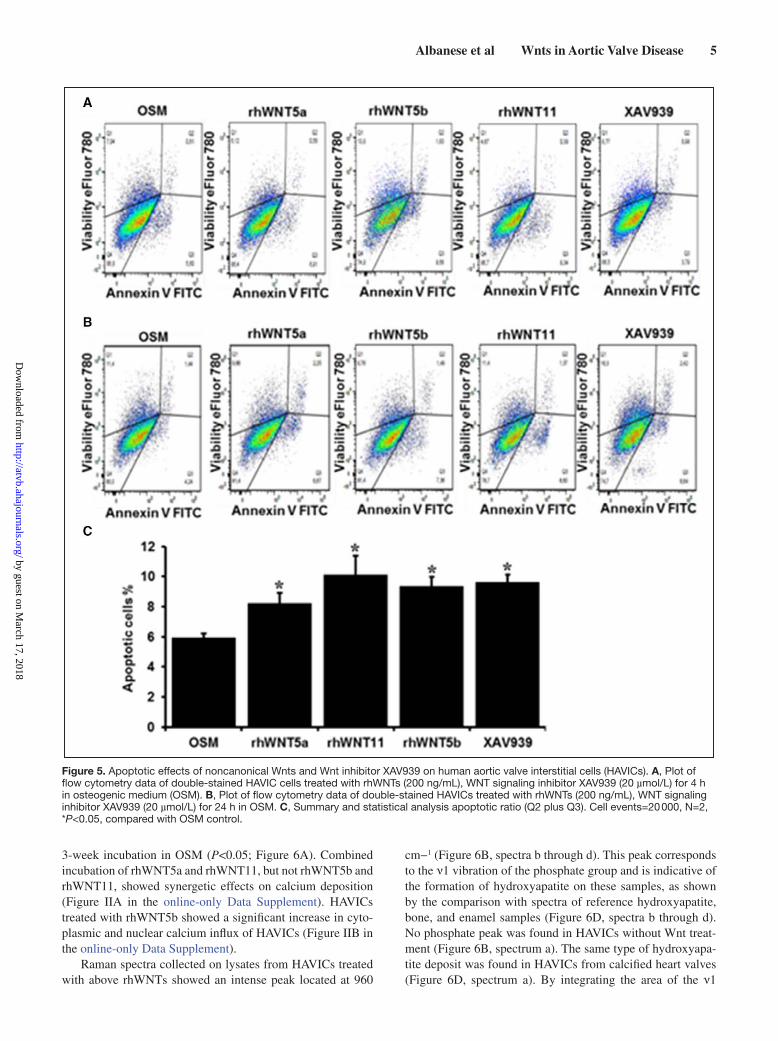
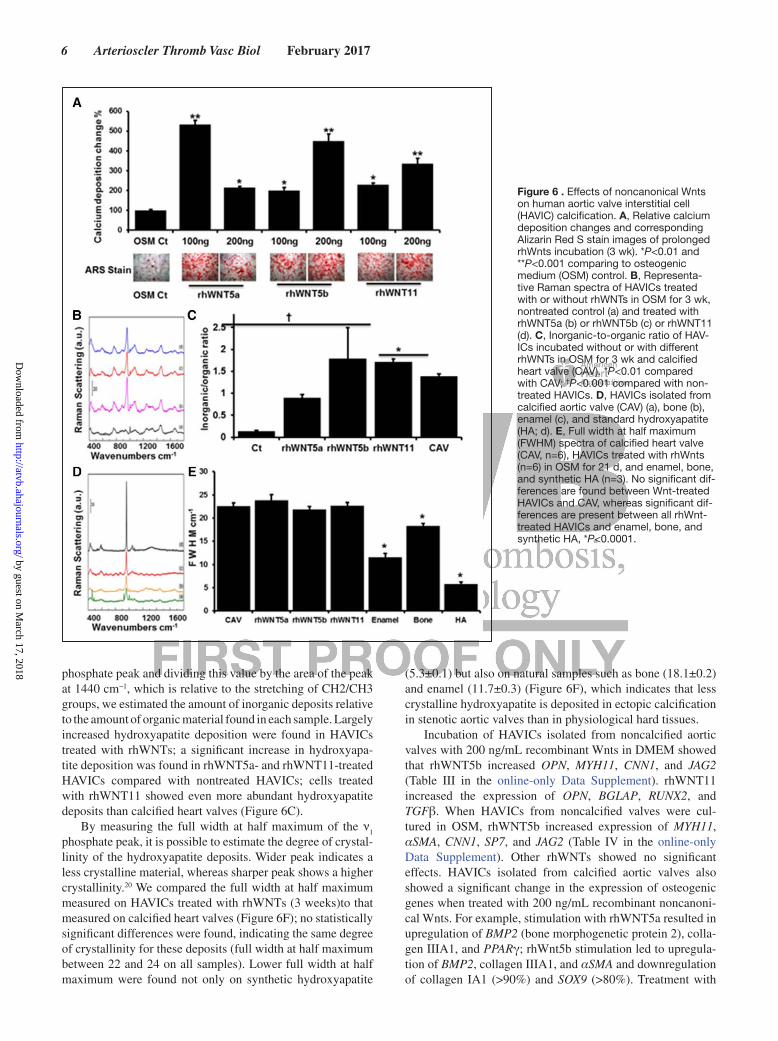
Effects of Noncanonic Wnts on HAVICs Proliferation, Apoptosis, and Alkaline Phosphatase ActivityAfter 48-hour treatment with rhWNTs in DMEM, only rhWNT5b significantly reduced cell proliferation (Figure 4A). When cells were cultured in osteogenic medium (OSM), both rhWNT5a (at 200 ng/mL) and rhWNT5b significantly reduced cell proliferation (Figure 4B). After 3-week incuba-tion, all 3 Wnts significantly decreased cell proliferation of HAVICs (Figure 4C). Incubation of HAVICs with rhWNT5a, rhWNT5b, and rhWNT11 for 4 hours, 4 hours, and 24 hours in OSM caused a significant increase in cell apoptosis assayed by annexin V/eFluor 780 staining (Figure 5). The same result was also assessed by caspase-3/7 activity on HAVICs treated for 10 days with rhWNTs using CellEvent Caspase-3/7 Green Detection Reagent (Figure I in the online-only Data Supplement).
Cultured HAVICs have been demonstrated to undergo in vitro calcification via a process dependent, in part, on the upregulation of alkaline phosphatase (ALP) activity.19 Although HAVICs from noncalcified aortic valves show low ALP activity, all 7 HAVICs lines generated from stenotic aor-tic valve leaflets show significant amount of ALP activity (3–6 U/mg). Treatment of HAVICs collected from noncalcified valves with recombinant human Wnts showed no effect on ALP activity after 48-hour incubation in either media.
Prolonged Incubation of Noncanonic WNTs Induce Significant Calcification of HAVICs and Osteogenic-Related Gene ExpressionAll 3 noncanonical Wnts resulted in significant increases of calcium deposition and formation of calcified nodules in HAVICs isolated from noncalcified human aortic valves after
Figure 4. Effects of noncanonical WNTs on human aortic valve interstitial cell (HAVIC) proliferation. Noncalcified HAVICs were treated with rhWNT5a, rhWNT5b, and rhWNT11 for 48 h in trip-licates with normal DMEM (A) or osteogenic medium (OSM; B). Prolonged incubation (3 wk) of rhWnts with HAVICs in OSM (C). *P<0.05; **P<0.001.
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
Albanese et al Wnts in Aortic Valve Disease 5
3-week incubation in OSM (P<0.05; Figure 6A). Combined incubation of rhWNT5a and rhWNT11, but not rhWNT5b and rhWNT11, showed synergetic effects on calcium deposition (Figure IIA in the online-only Data Supplement). HAVICs treated with rhWNT5b showed a significant increase in cyto-plasmic and nuclear calcium influx of HAVICs (Figure IIB in the online-only Data Supplement).
Raman spectra collected on lysates from HAVICs treated with above rhWNTs showed an intense peak located at 960
cm−1 (Figure 6B, spectra b through d). This peak corresponds to the ν1 vibration of the phosphate group and is indicative of the formation of hydroxyapatite on these samples, as shown by the comparison with spectra of reference hydroxyapatite, bone, and enamel samples (Figure 6D, spectra b through d). No phosphate peak was found in HAVICs without Wnt treat-ment (Figure 6B, spectrum a). The same type of hydroxyapa-tite deposit was found in HAVICs from calcified heart valves (Figure 6D, spectrum a). By integrating the area of the ν1
Figure 5. Apoptotic effects of noncanonical Wnts and Wnt inhibitor XAV939 on human aortic valve interstitial cells (HAVICs). A, Plot of flow cytometry data of double-stained HAVIC cells treated with rhWNTs (200 ng/mL), WNT signaling inhibitor XAV939 (20 µmol/L) for 4 h in osteogenic medium (OSM). B, Plot of flow cytometry data of double-stained HAVICs treated with rhWNTs (200 ng/mL), WNT signaling inhibitor XAV939 (20 µmol/L) for 24 h in OSM. C, Summary and statistical analysis apoptotic ratio (Q2 plus Q3). Cell events=20 000, N=2, *P<0.05, compared with OSM control.
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from
6 Arterioscler Thromb Vasc Biol February 2017
phosphate peak and dividing this value by the area of the peak at 1440 cm−1, which is relative to the stretching of CH2/CH3 groups, we estimated the amount of inorganic deposits relative to the amount of organic material found in each sample. Largely increased hydroxyapatite deposition were found in HAVICs treated with rhWNTs; a significant increase in hydroxyapa-tite deposition was found in rhWNT5a- and rhWNT11-treated HAVICs compared with nontreated HAVICs; cells treated with rhWNT11 showed even more abundant hydroxyapatite deposits than calcified heart valves (Figure 6C).
By measuring the full width at half maximum of the ν1
phosphate peak, it is possible to estimate the degree of crystal-linity of the hydroxyapatite deposits. Wider peak indicates a less crystalline material, whereas sharper peak shows a higher crystallinity.20 We compared the full width at half maximum measured on HAVICs treated with rhWNTs (3 weeks)to that measured on calcified heart valves (Figure 6F); no statistically significant differences were found, indicating the same degree of crystallinity for these deposits (full width at half maximum between 22 and 24 on all samples). Lower full width at half maximum were found not only on synthetic hydroxyapatite
(5.3±0.1) but also on natural samples such as bone (18.1±0.2) and enamel (11.7±0.3) (Figure 6F), which indicates that less crystalline hydroxyapatite is deposited in ectopic calcification in stenotic aortic valves than in physiological hard tissues.
Incubation of HAVICs isolated from noncalcified aortic valves with 200 ng/mL recombinant Wnts in DMEM showed that rhWNT5b increased OPN, MYH11, CNN1, and JAG2 (Table III in the online-only Data Supplement). rhWNT11 increased the expression of OPN, BGLAP, RUNX2, and TGFβ. When HAVICs from noncalcified valves were cul-tured in OSM, rhWNT5b increased expression of MYH11, αSMA, CNN1, SP7, and JAG2 (Table IV in the online-only Data Supplement). Other rhWNTs showed no significant effects. HAVICs isolated from calcified aortic valves also showed a significant change in the expression of osteogenic genes when treated with 200 ng/mL recombinant noncanoni-cal Wnts. For example, stimulation with rhWNT5a resulted in upregulation of BMP2 (bone morphogenetic protein 2), colla-gen IIIA1, and PPARγ; rhWnt5b stimulation led to upregula-tion of BMP2, collagen IIIA1, and αSMA and downregulation of collagen IA1 (>90%) and SOX9 (>80%). Treatment with
Figure 6 . Effects of noncanonical Wnts on human aortic valve interstitial cell (HAVIC) calcification. A, Relative calcium deposition changes and corresponding Alizarin Red S stain images of prolonged rhWnts incubation (3 wk). *P<0.01 and **P<0.001 comparing to osteogenic medium (OSM) control. B, Representa-tive Raman spectra of HAVICs treated with or without rhWNTs in OSM for 3 wk, nontreated control (a) and treated with rhWNT5a (b) or rhWNT5b (c) or rhWNT11 (d). C, Inorganic-to-organic ratio of HAV-ICs incubated without or with different rhWNTs in OSM for 3 wk and calcified heart valve (CAV). *P<0.01 compared with CAV; †P<0.001 compared with non-treated HAVICs. D, HAVICs isolated from calcified aortic valve (CAV) (a), bone (b), enamel (c), and standard hydroxyapatite (HA; d). E, Full width at half maximum (FWHM) spectra of calcified heart valve (CAV, n=6), HAVICs treated with rhWnts (n=6) in OSM for 21 d, and enamel, bone, and synthetic HA (n=3). No significant dif-ferences are found between Wnt-treated HAVICs and CAV, whereas significant dif-ferences are present between all rhWnt-treated HAVICs and enamel, bone, and synthetic HA, *P<0.0001.
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from

Albanese et al Wnts in Aortic Valve Disease 7
rhWnt11 significantly increased expression of BMP2, colla-gen IA1, αSMA, and PPARγ (Table V in the online-only Data Supplement).
Effects of Noncanonical Wnts on Phospho-Mitogen-Activated Protein Kinases and Effects of Pathway Inhibitors on CalcificationWhen noncalcified HAVICs were treated with 100 ng/mL of rhWNTs, phosphorylation of c-Jun N-terminal kinase 1/2/3 (JNK1/2/3), serine–threonine protein kinase 1 (Akt1), and mitogen-activated protein kinase-38β (MAPK38) were sig-nificantly increased, and rhWNT5a showed the strongest increase (Figure III in the online-only Data Supplement). Stenotic aortic valve tissues showed significant increase of phosphor-p38 and pJNK (Figure IV in the online-only Data Supplement). Incubation of HAVICs with the MAPK38 inhib-itor SB203580 or the GSK3β inhibitor CHIR99021 signifi-cantly decreased calcium deposition. Conversely, incubation of HAVICs with the ERK1/2 inhibitor PD184352 significantly increased calcium deposition (Figure V in the online-only Data Supplement).
Incubation of HAVICs with the mTOR inhibitor SP600125 (20 ng/mL) or the JNK inhibitor rapamycin (9 nmol/L) in combination with the 3 different rhWNTs showed synergic effects on calcium deposition with SP600125 but not with rapamycin (Figure VI in the online-only Data Supplement).
DiscussionAortic valve calcification is an active process that involves mechanical and biochemical factors leading to valve degen-eration and stenosis. The critical features of aortic valve cal-cification that involve VIC activation to osteoblast and bone formation lead to the growing interest in exploring the role of bone metabolism pathways in the aortic valve calcifica-tion such as Wnt pathways.3,4,21 Involvement of canonical Wnt signaling in human valve calcification has been previously reported11,12,22; however, this study implicates noncanoni-cal Wnt ligands WNT5b and WNT11 in the pathogenesis of aortic valve calcification. The mRNA expression of WNT5a, WNT5b, and WNT11 was higher in calcified valves compared with normal valves. Interestingly, we found significantly higher mRNA expression of WNT5b in calcified bicuspid aor-tic valves compared with calcified tricuspid aortic valves. We also demonstrated that HAVICs from calcified aortic valves express abundant levels of WNT5b and WNT11 proteins via Western blotting, whereas dermal fibroblasts showed mini-mal to no expression of these ligands. Moreover, in noncal-cified HAVICs, all 3 noncanonical Wnts were found to have strong apoptotic effect and induced calcification of these cells. Taken together, these findings suggest a possible role for non-canonical Wnt pathway in the pathogenesis of aortic valve calcification.
The Wnt-Frizzled system, involving 19 Wnt ligands and 10 Frizzleds, plays key roles in the development and function of multiple organs including the cardiovascu-lar system.22 Wnt signaling is also thought to be required for different aspects of cardiac and vascular development,
including myocardial specification, cardiac morphogen-esis, cardiac valve formation, and endothelial and vascular smooth muscle cell proliferation.23 Furthermore, Wnt modu-lators were found to be involved in several cardiac diseases including myocardial infarction, arrhythmia, and heart fail-ure.24–26 Several in vitro and animal studies demonstrated the role of Wnt/β-catenin signaling in inducing osteogenic signaling and the development of aortic valve disease.11,27–29 Caira et al12 demonstrated an active endochondral bone for-mation mechanism in the development of heart valve disease associated with high expression of Wnt3a (important canoni-cal Wnt factors) and coreceptors LRP5/6 in human calcified aortic valves. Askevold et al30 found increased serum level and tissue expression of Wnt modulators (sFRP3, WIF-1, and DKK-1) in symptomatic aortic valve stenosis. In this study, we found high mRNA expression of WNT5a, WNT5b, and WNT11 in calcified aortic valves compared with nor-mal aortic valves. Our histological studies showed that both WNT5b and WNT11 have high immunostaining in areas of active calcification in calcified valves and in activated osteo-blast-like VICs. WNT5b immunostaining was also high in fibrous regions surrounding active calcification in diseased valves. The presence of these noncanonical Wnts in areas of increased matrix deposition, inflammation, and calcifica-tion suggests possible autocrine and paracrine roles in aortic valve disease. In particular, the strong immunostaining of Wnt5b and Wnt11 in macrophages in inflammatory regions of diseased valves suggests that inflammatory cells provide a potential mechanism for the model proposed by Aikawa et al where atherosclerotic plaque inflammation is the trigger for osteogenesis and calcification.
The aortic valve is an avascular structure that is made up of VICs arranged in 3 layers. VICs maintain the integrity of valve tissue by secreting extracellular matrix, including col-lagen, elastin, and glycosaminoglycans, that provide strength and elasticity to the valve.31 In situ transition of VICs to osteoblast-like cells is an important mechanism implicated in the pathogenesis of aortic valve calcification. The activated VICs are responsive to osteogenic mediators such as BMPs that stimulate osteoblasts to initiate skeletal bone forma-tion.32,33 In this study, we generated primary cultures of VICs from calcified human aortic valves. This allows for more tar-geted studies of the specific functional roles of these cells in heart disease. Our Western blot findings in isolated HAVICs provide further evidence for the presence of Wnt protein for-mation. Although there is evidence supporting the role of WNT5b as a regulator of osteoblastic lineage allocation,34,35 its exact role in this process is not fully understood. Here, we show profound osteogenic effect of Wnts on HAVICs, which was further confirmed by a spectroscopic analysis of the deposits. On incubation in OSM for 21 days, only HAVICs treated with Wnts showed the presence of hydroxyapatite deposits, whereas nontreated HAVICs did not significantly mineralize. The amount of hydroxyapatite deposits was lower than that found in calcified heart valves for cells treated with WNT5a and higher for those treated with WNT11. The crystallinity of the hydroxyapatite deposits found in Wnt-treated AVICs was comparable to that found in calcified heart valves. This suggests that the deposits formed in the presence
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from

8 Arterioscler Thromb Vasc Biol February 2017
of Wnt are actually similar to those found in calcified heart valves, thus confirming the importance of Wnts in heart valve mineralization.
Normal human aortic valves contain few macrophages, whereas calcified human aortic valves have significant infil-tration of macrophages and leukocytes. This suggests that aortic valve disease, similar to atherosclerosis, can be consid-ered as an active inflammatory process, and it has been sug-gested that macrophage and inflammatory cell infiltration is involved in the early development of aortic valve disease.36,37 Both WNT5b and WNT11 were found to have significantly elevated immunostaining in monocytes and macrophages in calcified valves. Several studies highlight that some inflam-matory cytokines may induce ectopic calcification of human adipose tissue–derived mesenchymal stem cells via activation of noncanonical Wnt pathways.38 In addition, inflammatory cytokines can promote the production of proapoptotic ligands such as transforming growth factor-β1 and tumor necro-sis factor–related apoptosis–inducing ligand that have been detected in human calcified aortic valves.39 Wnt proteins are important in organogenesis and tissue hemostasis, and several studies highlight their role in apoptosis.40,41 In this study, all 3 Wnts had strong apoptotic effect on noncalcified HAVICs that may suggest possible mechanism of valve degeneration and calcification. This is further supported by the WNT11-increased expression of transforming growth factor-β and by the findings that the ERK1/2 inhibitor PD184352, that is known to increase cell apoptosis, significantly increased HAVICs calcification. Interestingly, although all noncanoni-cal Wnts studied here significantly increased HAVICs cal-cification, none had significant effect on ALP activity. This suggests that noncanonical Wnts induce calcification inde-pendent of ALP activity via mechanisms similar to what has been reported in vascular calcification pathogenesis,42 a path-way that may involve apoptosis.
Bicuspid aortic valve is the most common congenital car-diac abnormality with an estimated prevalence of 1% to 2%. It is associated with incidence of aortic valve calcification, aortic aneurysm formation, and aortic dissection.43 The exact patho-genesis of the formation of BAV is not yet fully understood. It is thought there is a genetic component, especially given the association between BAV and other congenital abnormali-ties such as coarctation of the aorta.44 Mutations in NOTCH1 gene, a transmembrane receptor that has a role in determining cell outcome in organogenesis, were noted in 2 families with BAV and also in sporadic cases.45,46 It was found that Notch1 normally represses the gene encoding BMP2 in murine aor-tic valves in vivo and in aortic VICs in vitro.47 Several stud-ies highlight the counteractive interaction between NOTCH and Wnt signaling in heart development and diseases.48,49 In this study, we found a significant increase of Wnt5b in bicus-pid compared with tricuspid valves, and this may suggest its involvement in the development and pathogenesis of this dis-ease. Moreover, all noncanonical Wnts increased expression of BMP2 in calcified HAVICs. There was also increased phos-phorylation of GSK3β and MAPK38β, both of which have previously been shown to be involved in Wnt activation and osteogenic differentiation. These findings were further sup-ported by our observation of significant decrease in HAVICs
calcification in response to treatment with the GSK3β and MAPK38 inhibitors.
In addition to the evidence suggesting a role for WNT5b and WNT11 signaling in the pathogenesis of aortic valve cal-cification, both ligands demonstrated interesting significant correlations with various clinical parameters. It is a general rule that the pressure gradient across a valve is inversely related to the area of the orifice.50 Both WNT5b and WNT11 immunostaining demonstrated significant negative corre-lations with the area of the orifice and significant positive correlations with aortic jet velocity, Pmax, and Pmean. This suggests direct associations between WNT5b and WNT11 protein expression and aortic valve stenosis. Additionally, it has been previously demonstrated in osteoblasts that mechani-cal stimulation causes increased Wnt production, which then affects the functionality of those osteoblasts.51 This raises the possibility that it is actually the increased pressure across a valve that is stimulating increased Wnt production and that this increased Wnt signaling then mediates pathological cal-cium deposition through activation of VICs. In this study, we found that WNT5b has a positive correlation with Parsonnet score. This score was developed as a preoperative risk-assess-ment score for cardiac surgeries.52 By taking into account 37 factors, such as age, sex, congestive heart failure, hyperten-sion, diabetes mellitus and morbid obesity, among others, the Parsonnet score has been successfully used preoperatively to predict surgical outcomes. The fact that WNT5b positively correlates with Parsonnet score suggests that WNT5b may be used as a prognostic marker for valve replacement operations.
Given our findings that rhWNT5a, rhWNT5b, and rhWNT11 all induce expression of osteopontin, a key regu-lator of orthotopic and heterotropic mineralization, future studies warrant the investigation of the relationship between noncanonical Wnt signaling and osteopontin in vascular and valvular mineralization.
There is increasing evidence that valve extracellular matrix, 50% of which consists of collagen by dry weight, plays a role in aortic valve calcification, specifically via its interac-tion with VICs.53 Our findings demonstrate differential roles of Wnt signals on collagen IIIA1 and IA1, with rhWNT5a and rhWNT5b contributing to the upregulation of collagen IIIA1; meanwhile, rhWNT5b decreased the expression of collagen IA1, and rhWNT11 increased its expression. This may be explained in part by work by Eriksen et al54 demonstrating that turnover of collagen type I is significantly increased in aortic stenosis, particularly around calcified nodules. Their findings also demonstrate slight upregulation of type III collagen in calcified valves. Together, this suggests that Wnt signaling could be contributing to aortic valve calcification by mediat-ing interactions between VICs and extracellular matrix.
In summary, we demonstrate here increased expression of 3 important members of the Wnt family, WNT5a, WNT5b, and WNT11, in the valves of patients with aortic valve ste-nosis, and the association of this increase with clinical and pathological characteristics of the disease. Given that there is no previous work on the involvement of noncanonical Wnt pathway in aortic valve calcification, our findings make an important contribution to developing a better understand-ing of this pathological process. Because mTOR and JNK
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from

Albanese et al Wnts in Aortic Valve Disease 9
inhibition did not diminish calcium deposition, noncanonical Wnt signaling relays promoting VIC mineralization have yet to be fully elucidated. These findings also illustrate the impor-tance of future studies in determining functional roles of Wnt signaling molecules in the osteogenic differentiation of valve interstitial cells and aortic valve calcification.
PerspectivesEven in the absence of valvular stenosis, aortic valve calci-fication is associated with an increased risk of cardiovascu-lar events and mortality from cardiovascular causes, which is alarming given that there are no proven treatments to avoid or slow its development. Interfering with the Wnt pathway may provide a novel way to treat the disease.
Sources of FundingThis work was supported by the Canadian Institute of Health Research and the Heart and Stroke Foundation of Quebec.
DisclosuresNone.
References 1. Otto CM, Prendergast B. Aortic-valve stenosis–from patients at risk to
severe valve obstruction. N Engl J Med. 2014;371:744–756. doi: 10.1056/NEJMra1313875.
2. Thaden JJ, Nkomo VT, Enriquez-Sarano M. The global burden of aor-tic stenosis. Prog Cardiovasc Dis. 2014;56:565–571. doi: 10.1016/j.pcad.2014.02.006.
3. Li C, Xu S, Gotlieb AI. The progression of calcific aortic valve disease through injury, cell dysfunction, and disruptive biologic and physical force feedback loops. Cardiovasc Pathol. 2013;22:1–8. doi: 10.1016/j.carpath.2012.06.005.
4. Merryman WD, Schoen FJ. Mechanisms of calcification in aortic valve disease: role of mechanokinetics and mechanodynamics. Curr Cardiol Rep. 2013;15:355. doi: 10.1007/s11886-013-0355-5.
5. Rao TP, Kühl M. An updated overview on Wnt signaling pathways: a prelude for more. Circ Res. 2010;106:1798–1806. doi: 10.1161/CIRCRESAHA.110.219840.
6. Cha SW, Tadjuidje E, Tao Q, Wylie C, Heasman J. Wnt5a and Wnt11 interact in a maternal Dkk1-regulated fashion to activate both canonical and non-canonical signaling in Xenopus axis formation. Development. 2008;135:3719–3729. doi: 10.1242/dev.029025.
7. Cha SW, Tadjuidje E, White J, Wells J, Mayhew C, Wylie C, Heasman J. Wnt11/5a complex formation caused by tyrosine sulfation increases canonical signaling activity. Curr Biol. 2009;19:1573–1580. doi: 10.1016/j.cub.2009.07.062.
8. Christman MA 2nd, Goetz DJ, Dickerson E, McCall KD, Lewis CJ, Benencia F, Silver MJ, Kohn LD, Malgor R. Wnt5a is expressed in murine and human atherosclerotic lesions. Am J Physiol Heart Circ Physiol. 2008;294:H2864–H2870. doi: 10.1152/ajpheart.00982.2007.
9. Tian Y, Cohen ED, Morrisey EE. The importance of wnt signaling in cardiovascular development. Pediatric Cardiol. 2010;31:342–348. doi: 10.1007/s00246-009-9606-z.
10. Flaherty MP, Kamerzell TJ, Dawn B. Wnt signaling and cardiac differ-entiation. Prog Mol Biol Transl Sci. 2012;111:153–174. doi: 10.1016/B978-0-12-398459-3.00007-1.
11. Gu GJ, Chen T, Zhou HM, Sun KX, Li J. Role of Wnt/β-catenin signaling pathway in the mechanism of calcification of aortic valve. J Huazhong Univ Sci Technolog Med Sci. 2014;34:33–36. doi: 10.1007/s11596-014-1228-x.
12. Caira FC, Stock SR, Gleason TG, McGee EC, Huang J, Bonow RO, Spelsberg TC, McCarthy PM, Rahimtoola SH, Rajamannan NM. Human degenerative valve disease is associated with up-regulation of low-density lipoprotein receptor-related protein 5 receptor-mediated bone formation. J Am Coll Cardiol. 2006;47:1707–1712. doi: 10.1016/j.jacc.2006.02.040.
13. Xin H, Xin F, Zhou S, Guan S. The Wnt5a/Ror2 pathway is associated with determination of the differentiation fate of bone marrow mesenchymal
stem cells in vascular calcification. Int J Mol Med. 2013;31:583–588. doi: 10.3892/ijmm.2013.1242.
14. Guan S, Wang Z, Xin F, Xin H. Wnt5a is associated with the differentia-tion of bone marrow mesenchymal stem cells in vascular calcification by connecting with different receptors. Mol Med Rep. 2014;10:1985–1991. doi: 10.3892/mmr.2014.2449.
15. Briolay A, Lencel P, Bessueille L, Caverzasio J, Buchet R, Magne D. Autocrine stimulation of osteoblast activity by Wnt5a in response to TNF-α in human mesenchymal stem cells. Biochem Biophys Res Commun. 2013;430:1072–1077. doi: 10.1016/j.bbrc.2012.12.036.
16. Woldt E, Terrand J, Mlih M, et al. The nuclear hormone receptor PPARγ counteracts vascular calcification by inhibiting Wnt5a signalling in vas-cular smooth muscle cells. Nat Commun. 2012;3:1077. doi: 10.1038/ncomms2087.
17. Cheng SL, Ramachandran B, Behrmann A, Shao JS, Mead M, Smith C, Krchma K, Bello Arredondo Y, Kovacs A, Kapoor K, Brill LM, Perera R, Williams BO, Towler DA. Vascular smooth muscle LRP6 limits arteriosclerotic calcification in diabetic LDLR-/- mice by restraining noncanonical Wnt signals. Circ Res. 2015;117:142–156. doi: 10.1161/CIRCRESAHA.117.306712.
18. Shao JS, Cheng SL, Pingsterhaus JM, Charlton-Kachigian N, Loewy AP, Towler DA. Msx2 promotes cardiovascular calcification by activating paracrine Wnt signals. J Clin Invest. 2005;115:1210–1220. doi: 10.1172/JCI24140.
19. Vincent SD, Buckingham ME. How to make a heart: the origin and regu-lation of cardiac progenitor cells. Curr Top Dev Biol. 2010;90:1–41. doi: 10.1016/S0070-2153(10)90001-X.
20. Pucéat E, Reynard B, Lécuyer C. Can crystallinity be used to deter-mine the degree of chemical alteration of biogenic apatites? Chem Geol. 2004;205:83–97.
21. Chester AH, El-Hamamsy I, Butcher JT, Latif N, Bertazzo S, Yacoub MH. The living aortic valve: from molecules to function. Glob Cardiol Sci Pract. 2014;2014:52–77. doi: 10.5339/gcsp.2014.11.
22. Rajamannan NM. The role of Lrp5/6 in cardiac valve disease: LDL-density-pressure theory. J Cell Biochem. 2011;112:2222–2229. doi: 10.1002/jcb.23182.
23. Gessert S, Kühl M. The multiple phases and faces of wnt signaling dur-ing cardiac differentiation and development. Circ Res. 2010;107:186–199. doi: 10.1161/CIRCRESAHA.110.221531.
24. Dawson K, Aflaki M, Nattel S. Role of the Wnt-Frizzled system in car-diac pathophysiology: a rapidly developing, poorly understood area with enormous potential. J Physiol. 2013;591:1409–1432. doi: 10.1113/jphysiol.2012.235382.
25. Daskalopoulos EP, Hermans KC, Janssen BJ, Matthijs Blankesteijn W. Targeting the Wnt/frizzled signaling pathway after myocardial infarc-tion: a new tool in the therapeutic toolbox? Trends Cardiovasc Med. 2013;23:121–127. doi: 10.1016/j.tcm.2012.09.010.
26. Naito AT, Shiojima I, Komuro I. Wnt signaling and aging-related heart disorders. Circ Res. 2010;107:1295–1303. doi: 10.1161/CIRCRESAHA.110.223776.
27. Alfieri CM, Cheek J, Chakraborty S, Yutzey KE. Wnt signaling in heart valve development and osteogenic gene induction. Dev Biol. 2010;338:127–135. doi: 10.1016/j.ydbio.2009.11.030.
28. Chen JH, Chen WL, Sider KL, Yip CY, Simmons CA. β-Catenin mediates mechanically regulated, transforming growth factor-β1-induced myofibro-blast differentiation of aortic valve interstitial cells. Arterioscler Thromb Vasc Biol. 2011;31:590–597. doi: 10.1161/ATVBAHA.110.220061.
29. Rajamannan NM. Oxidative-mechanical stress signals stem cell niche mediated Lrp5 osteogenesis in eNOS(-/-) null mice. J Cell Biochem. 2012;113:1623–1634. doi: 10.1002/jcb.24031.
30. Askevold ET, Gullestad L, Aakhus S, Ranheim T, Tønnessen T, Solberg OG, Aukrust P, Ueland T. Secreted Wnt modulators in symptom-atic aortic stenosis. J Am Heart Assoc. 2012;1:e002261. doi: 10.1161/JAHA.112.002261.
31. Liu AC, Joag VR, Gotlieb AI. The emerging role of valve interstitial cell phenotypes in regulating heart valve pathobiology. Am J Pathol. 2007;171:1407–1418. doi: 10.2353/ajpath.2007.070251.
32. Mohler ER 3rd, Chawla MK, Chang AW, Vyavahare N, Levy RJ, Graham L, Gannon FH. Identification and characterization of calcifying valve cells from human and canine aortic valves. J Heart Valve Dis. 1999;8:254–260.
33. Mohler ER 3rd, Gannon F, Reynolds C, Zimmerman R, Keane MG, Kaplan FS. Bone formation and inflammation in cardiac valves. Circulation. 2001;103:1522–1528.
34. Hurson CJ, Butler JS, Keating DT, Murray DW, Sadlier DM, O’Byrne JM, Doran PP. Gene expression analysis in human osteoblasts exposed
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from

10 Arterioscler Thromb Vasc Biol February 2017
to dexamethasone identifies altered developmental pathways as puta-tive drivers of osteoporosis. BMC Musculoskelet Disord. 2007;8:12. doi: 10.1186/1471-2474-8-12.
35. van Tienen FH, Laeremans H, van der Kallen CJ, Smeets HJ. Wnt5b stimu-lates adipogenesis by activating PPARgamma, and inhibiting the beta-catenin dependent Wnt signaling pathway together with Wnt5a. Biochem Biophys Res Commun. 2009;387:207–211. doi: 10.1016/j.bbrc.2009.07.004.
36. Coté N, Mahmut A, Bosse Y, Couture C, Pagé S, Trahan S, Boulanger MC, Fournier D, Pibarot P, Mathieu P. Inflammation is associated with the remodeling of calcific aortic valve disease. Inflammation. 2013;36:573–581. doi: 10.1007/s10753-012-9579-6.
37. New SE, Aikawa E. Cardiovascular calcification: an inflammatory disease. Circ J. 2011;75:1305–1313.
38. Fukuyo S, Yamaoka K, Sonomoto K, Oshita K, Okada Y, Saito K, Yoshida Y, Kanazawa T, Minami Y, Tanaka Y. IL-6-accelerated calcification by induction of ROR2 in human adipose tissue-derived mesenchymal stem cells is STAT3 dependent. Rheumatology (Oxford). 2014;53:1282–1290. doi: 10.1093/rheumatology/ket496.
39. Galeone A, Brunetti G, Oranger A, Greco G, Di Benedetto A, Mori G, Colucci S, Zallone A, Paparella D, Grano M. Aortic valvular interstitial cells apoptosis and calcification are mediated by TNF-related apoptosis-inducing ligand. Int J Cardiol. 2013;169:296–304. doi: 10.1016/j.ijcard.2013.09.012.
40. Pećina-Slaus N. Wnt signal transduction pathway and apoptosis: a review. Cancer Cell Int. 2010;10:22. doi: 10.1186/1475-2867-10-22.
41. Kahn M. Can we safely target the WNT pathway? Nat Rev Drug Discov. 2014;13:513–532. doi: 10.1038/nrd4233.
42. Kwun IS, Alcantara EH, Shin M-Y, Beattie JH. Zn deficiency promotes calcification in vascular smooth muscle cells independent of alp action and in part by pit1 up-regulation. FASEB J. 2013;27:
43. Losenno KL, Chu MW. Bicuspid aortic valve disease. CMAJ. 2013;185:1599. doi: 10.1503/cmaj.121875.
44. Abdulkareem N, Smelt J, Jahangiri M. Bicuspid aortic valve aortopa-thy: genetics, pathophysiology and medical therapy. Interact Cardiovasc Thorac Surg. 2013;17:554–559. doi: 10.1093/icvts/ivt196.
45. Mohamed SA, Aherrahrou Z, Liptau H, Erasmi AW, Hagemann C, Wrobel S, Borzym K, Schunkert H, Sievers HH, Erdmann J. Novel missense mutations (p.T596M and p.P1797H) in NOTCH1 in patients with bicuspid aortic valve. Biochem Biophys Res Commun. 2006;345:1460–1465. doi: 10.1016/j.bbrc.2006.05.046.
46. Foffa I, Ait Alì L, Panesi P, Mariani M, Festa P, Botto N, Vecoli C, Andreassi MG. Sequencing of NOTCH1, GATA5, TGFBR1 and TGFBR2 genes in familial cases of bicuspid aortic valve. BMC Med Genet. 2013;14:44. doi: 10.1186/1471-2350-14-44.
47. Nigam V, Srivastava D. Notch1 represses osteogenic pathways in aor-tic valve cells. J Mol Cell Cardiol. 2009;47:828–834. doi: 10.1016/j.yjmcc.2009.08.008.
48. Wang Y, Wu B, Chamberlain AA, Lui W, Koirala P, Susztak K, Klein D, Taylor V, Zhou B. Endocardial to myocardial notch-wnt-bmp axis regulates early heart valve development. PLoS One. 2013;8:e60244. doi: 10.1371/journal.pone.0060244.
49. Rusanescu G, Weissleder R, Aikawa E. Notch signaling in cardiovas-cular disease and calcification. Curr Cardiol Rev. 2008;4:148–156. doi: 10.2174/157340308785160552.
50. Gorlin R, Gorlin SG. Hydraulic formula for calculation of the area of the stenotic mitral valve, other cardiac valves, and central circulatory shunts. I. Am Heart J. 1951;41:1–29.
51. Novo G, Fazio G, Visconti C, Carità P, Maira E, Fattouch K, Novo S. Atherosclerosis, degenerative aortic stenosis and statins. Curr Drug Targets. 2011;12:115–121.
52. Bernstein AD, Parsonnet V. Bedside estimation of risk as an aid for deci-sion-making in cardiac surgery. Ann Thorac Surg. 2000;69:823–828.
53. Rodriguez KJ, Piechura LM, Porras AM, Masters KS. Manipulation of valve composition to elucidate the role of collagen in aortic valve calcification. BMC Cardiovasc Disord. 2014;14:29. doi: 10.1186/1471-2261-14-29.
54. Eriksen HA, Satta J, Risteli J, Veijola M, Väre P, Soini Y. Type I and type III collagen synthesis and composition in the valve matrix in aor-tic valve stenosis. Atherosclerosis. 2006;189:91–98. doi: 10.1016/j.atherosclerosis.2005.11.034.
Highlights• Noncanonical WNTs were upregulated in human stenotic aortic valves and in primary cultured human aortic valve interstitial cells isolated from
stenotic aortic valves.• Noncanonical WNTs induced human aortic valve interstitial cell calcification in vitro and showed similar hydroxyapatite formation to that in
bone.• The effect of noncanonical WNTs on human aortic valve interstitial cells was mediated by mitogen-activated protein kinase signaling pathway.
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from

Jean-Claude Tardif and Adel SchwertaniBenoit de Varennes, Dominique Shum-Tim, Marta Cerruti, Ophélie Gourgas, Eric Rhéaume,
Isabella Albanese, Bin Yu, Hamood Al-Kindi, Bianca Barratt, Leah Ott, Mohammad Al-Refai,Role of Noncanonical Wnts Signaling Pathway in Human Aortic Valve Calcification
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2016 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
published online December 8, 2016;Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/early/2016/12/08/ATVBAHA.116.308394World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org/content/suppl/2017/12/05/ATVBAHA.116.308394.DC2 http://atvb.ahajournals.org/content/suppl/2016/12/16/ATVBAHA.116.308394.DC1
Data Supplement (unedited) at:
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on March 17, 2018
http://atvb.ahajournals.org/D
ownloaded from

Supplemental Materials
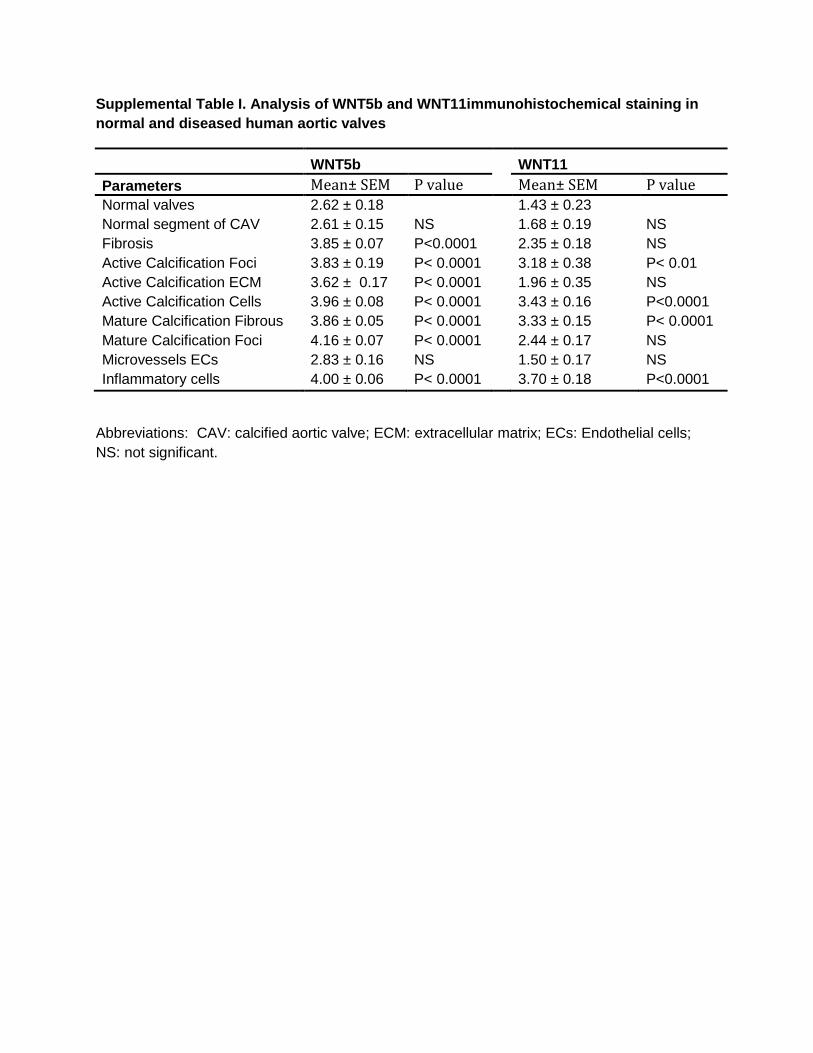
Supplemental Table I. Analysis of WNT5b and WNT11immunohistochemical staining in
normal and diseased human aortic valves
WNT5b
WNT11
Parameters Mean± SEM P value
Mean± SEM P value Normal valves 2.62 ± 0.18
1.43 ± 0.23
Normal segment of CAV 2.61 ± 0.15 NS
1.68 ± 0.19 NS
Fibrosis 3.85 ± 0.07 P<0.0001
2.35 ± 0.18 NS
Active Calcification Foci 3.83 ± 0.19 P< 0.0001
3.18 ± 0.38 P< 0.01
Active Calcification ECM 3.62 ± 0.17 P< 0.0001
1.96 ± 0.35 NS
Active Calcification Cells 3.96 ± 0.08 P< 0.0001 3.43 ± 0.16 P<0.0001
Mature Calcification Fibrous 3.86 ± 0.05 P< 0.0001 3.33 ± 0.15 P< 0.0001
Mature Calcification Foci 4.16 ± 0.07 P< 0.0001 2.44 ± 0.17 NS
Microvessels ECs 2.83 ± 0.16 NS 1.50 ± 0.17 NS
Inflammatory cells 4.00 ± 0.06 P< 0.0001
3.70 ± 0.18 P<0.0001
Abbreviations: CAV: calcified aortic valve; ECM: extracellular matrix; ECs: Endothelial cells;
NS: not significant.
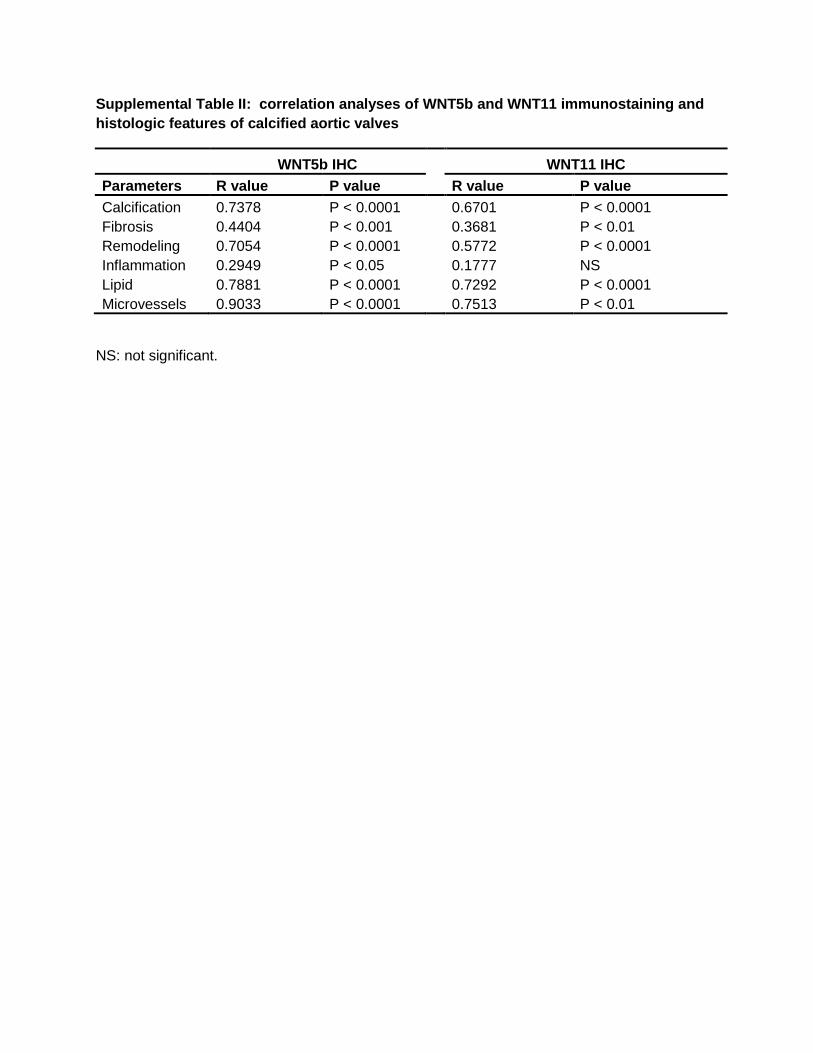
Supplemental Table II: correlation analyses of WNT5b and WNT11 immunostaining and
histologic features of calcified aortic valves
WNT5b IHC
WNT11 IHC
Parameters R value P value
R value P value
Calcification 0.7378 P < 0.0001
0.6701 P < 0.0001
Fibrosis 0.4404 P < 0.001
0.3681 P < 0.01
Remodeling 0.7054 P < 0.0001
0.5772 P < 0.0001
Inflammation 0.2949 P < 0.05
0.1777 NS
Lipid 0.7881 P < 0.0001
0.7292 P < 0.0001
Microvessels 0.9033 P < 0.0001
0.7513 P < 0.01
NS: not significant.
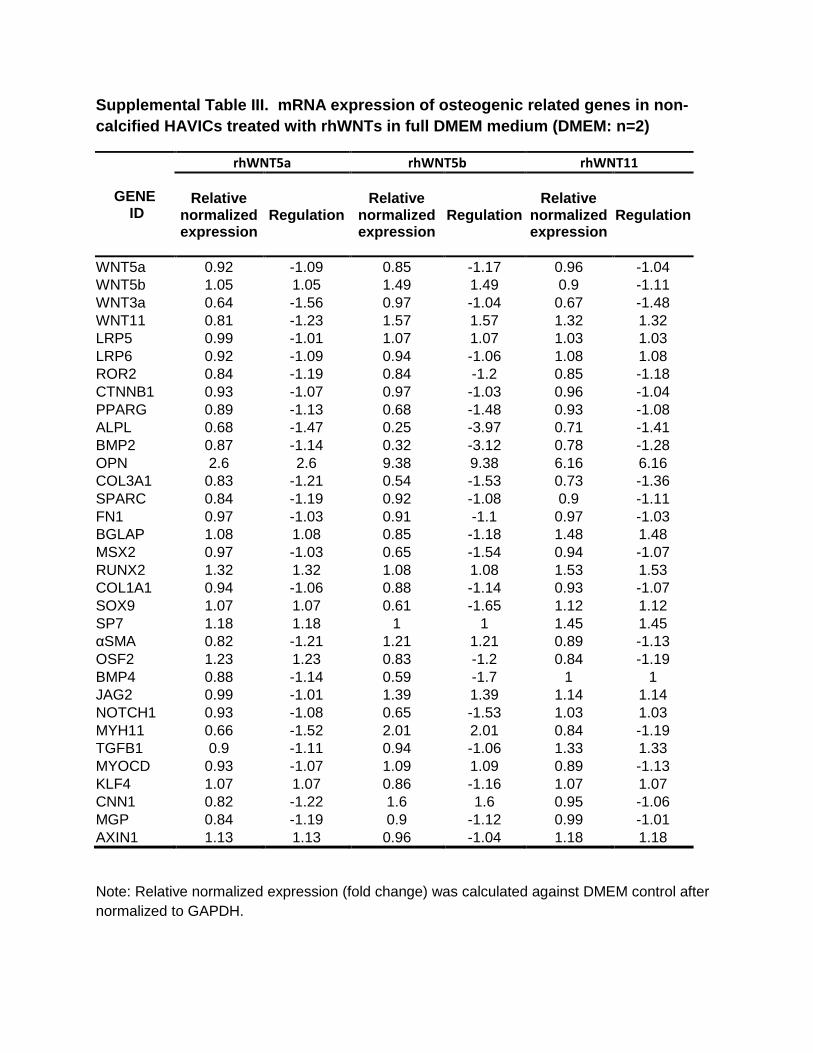
Supplemental Table III. mRNA expression of osteogenic related genes in non-
calcified HAVICs treated with rhWNTs in full DMEM medium (DMEM: n=2)
GENE
ID
rhWNT5a rhWNT5b rhWNT11
Relative normalized
expression Regulation
Relative normalized
expression Regulation
Relative normalized
expression Regulation
WNT5a 0.92 -1.09 0.85 -1.17 0.96 -1.04 WNT5b 1.05 1.05 1.49 1.49 0.9 -1.11 WNT3a 0.64 -1.56 0.97 -1.04 0.67 -1.48 WNT11 0.81 -1.23 1.57 1.57 1.32 1.32
LRP5 0.99 -1.01 1.07 1.07 1.03 1.03 LRP6 0.92 -1.09 0.94 -1.06 1.08 1.08 ROR2 0.84 -1.19 0.84 -1.2 0.85 -1.18
CTNNB1 0.93 -1.07 0.97 -1.03 0.96 -1.04 PPARG 0.89 -1.13 0.68 -1.48 0.93 -1.08
ALPL 0.68 -1.47 0.25 -3.97 0.71 -1.41 BMP2 0.87 -1.14 0.32 -3.12 0.78 -1.28
OPN 2.6 2.6 9.38 9.38 6.16 6.16 COL3A1 0.83 -1.21 0.54 -1.53 0.73 -1.36 SPARC 0.84 -1.19 0.92 -1.08 0.9 -1.11
FN1 0.97 -1.03 0.91 -1.1 0.97 -1.03 BGLAP 1.08 1.08 0.85 -1.18 1.48 1.48
MSX2 0.97 -1.03 0.65 -1.54 0.94 -1.07 RUNX2 1.32 1.32 1.08 1.08 1.53 1.53 COL1A1 0.94 -1.06 0.88 -1.14 0.93 -1.07
SOX9 1.07 1.07 0.61 -1.65 1.12 1.12 SP7 1.18 1.18 1 1 1.45 1.45 αSMA 0.82 -1.21 1.21 1.21 0.89 -1.13 OSF2 1.23 1.23 0.83 -1.2 0.84 -1.19 BMP4 0.88 -1.14 0.59 -1.7 1 1 JAG2 0.99 -1.01 1.39 1.39 1.14 1.14
NOTCH1 0.93 -1.08 0.65 -1.53 1.03 1.03 MYH11 0.66 -1.52 2.01 2.01 0.84 -1.19 TGFB1 0.9 -1.11 0.94 -1.06 1.33 1.33 MYOCD 0.93 -1.07 1.09 1.09 0.89 -1.13
KLF4 1.07 1.07 0.86 -1.16 1.07 1.07 CNN1 0.82 -1.22 1.6 1.6 0.95 -1.06 MGP 0.84 -1.19 0.9 -1.12 0.99 -1.01 AXIN1 1.13 1.13 0.96 -1.04 1.18 1.18
Note: Relative normalized expression (fold change) was calculated against DMEM control after
normalized to GAPDH.

Supplemental Table IV. mRNA expression of osteogenic related genes in non-calcified HAVICs treated with rhWNTs in osteogenic medium (OSM: n=2)
GENE
ID
rhWNT5a rhWNT5b rhWNT11
Relative normalized
expression Regulation
Relative normalized
expression Regulation
Relative normalized
expression Regulation
WNT5a 4.36 4.36 4.00 4.00 4.49 4.49
WNT5b 0.85 -1.17 1.63 1.63 1.01 1.01
WNT3a 1.17 1.17 1.22 1.22 1.04 1.04
WNT11 0.94 -1.06 1.05 1.05 0.77 -1.30
LRP5 0.88 -1.14 0.92 -1.09 0.90 -1.12
LRP6 0.84 -1.19 0.94 -1.07 0.91 -1.10
ROR2 2.65 2.65 2.45 2.45 2.40 2.40
CTNNB1 0.96 -1.04 1.04 1.04 0.92 -1.08
PPARG 0.88 -1.13 0.86 -1.17 0.95 -1.05
ALPL 0.92 -1.09 0.42 -2.41 1.21 1.21
BMP2 0.89 -1.13 0.40 -2.53 1.25 1.25
OPN 0.16 -6.40 0.64 -1.56 0.14 -7.29
COL3A1 4.99 4.99 4.19 4.19 5.07 5.07
SPARC 0.15 -6.48 1.18 1.18 0.93 -1.07
FN1 0.96 -1.04 1.12 1.12 0.90 -1.11
BGLAP 0.81 -1.24 0.69 -1.44 0.86 -1.16
MSX2 0.99 -1.01 0.64 -1.55 1.06 1.06
RUNX2 0.92 -1.09 0.74 -1.36 0.79 -1.26
COL1A1 5.53 5.53 5.19 5.19 5.47 5.47
SOX9 0.91 -1.10 0.71 -1.41 1.21 1.21
SP7 0.78 -1.29 1.25 1.25 0.83 -1.21
αSMA 1.02 1.02 1.52 1.52 0.79 -1.27
OSF2 1.20 1.20 1.26 1.26 1.01 1.01
BMP4 0.85 -1.18 0.64 -1.57 1.03 1.03
JAG2 1.84 1.84 2.92 2.92 2.02 2.02
NOTCH1 0.87 -1.14 0.67 -1.49 1.07 1.07
MYH11 0.84 -1.19 3.82 3.82 0.55 -1.80
TGFB1 0.90 -1.11 1.02 1.02 1.07 1.07
MYOCD 0.92 -1.09 1.21 1.21 0.85 -1.18
KLF4 0.87 -1.15 0.95 -1.05 0.94 -1.07
CNN1 3.37 3.37 8.11 8.11 3.45 3.45
MGP 0.91 -1.10 0.99 -1.01 0.87 -1.16
AXIN1 0.91 -1.10 0.82 -1.21 0.96 -1.04
Note: Relative normalized expression (fold change) was calculated against OSM control after normalized to GAPDH.
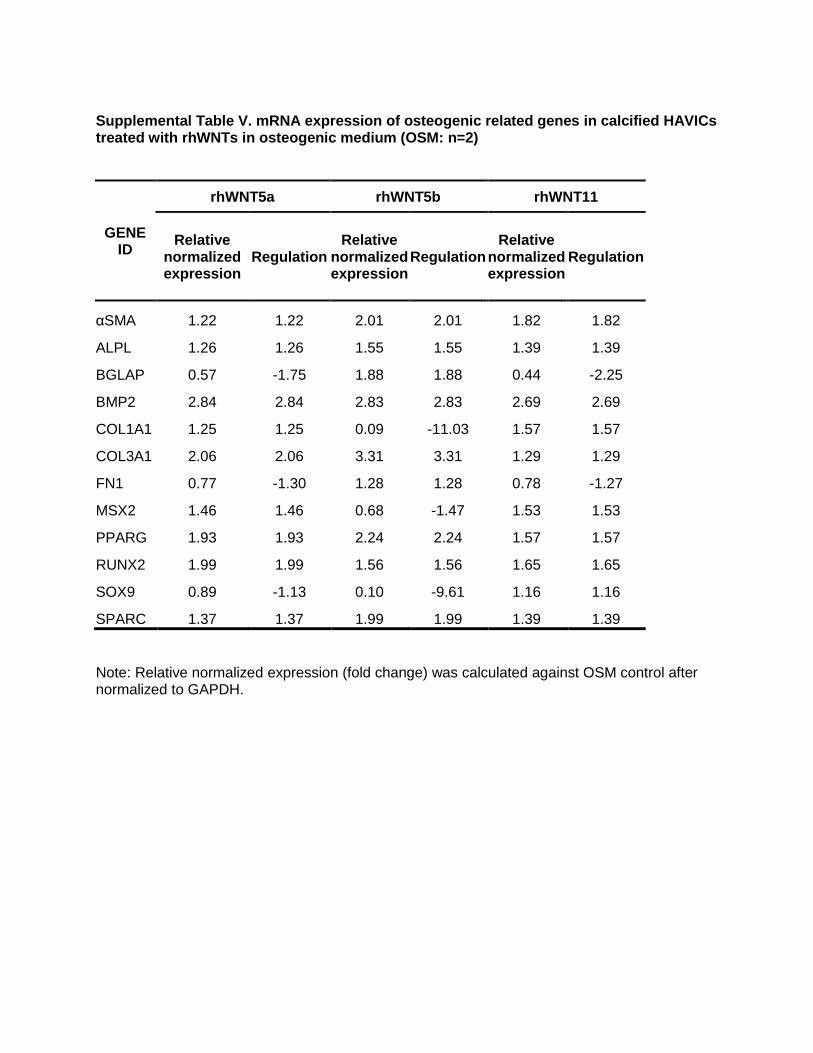
Supplemental Table V. mRNA expression of osteogenic related genes in calcified HAVICs treated with rhWNTs in osteogenic medium (OSM: n=2)
GENE
ID
rhWNT5a rhWNT5b rhWNT11
Relative normalized
expression Regulation
Relative normalized
expression Regulation
Relative normalized
expression Regulation
αSMA 1.22 1.22 2.01 2.01 1.82 1.82 ALPL 1.26 1.26 1.55 1.55 1.39 1.39 BGLAP 0.57 -1.75 1.88 1.88 0.44 -2.25
BMP2 2.84 2.84 2.83 2.83 2.69 2.69 COL1A1 1.25 1.25 0.09 -11.03 1.57 1.57 COL3A1 2.06 2.06 3.31 3.31 1.29 1.29
FN1 0.77 -1.30 1.28 1.28 0.78 -1.27 MSX2 1.46 1.46 0.68 -1.47 1.53 1.53 PPARG 1.93 1.93 2.24 2.24 1.57 1.57 RUNX2 1.99 1.99 1.56 1.56 1.65 1.65
SOX9 0.89 -1.13 0.10 -9.61 1.16 1.16 SPARC 1.37 1.37 1.99 1.99 1.39 1.39
Note: Relative normalized expression (fold change) was calculated against OSM control after normalized to GAPDH.
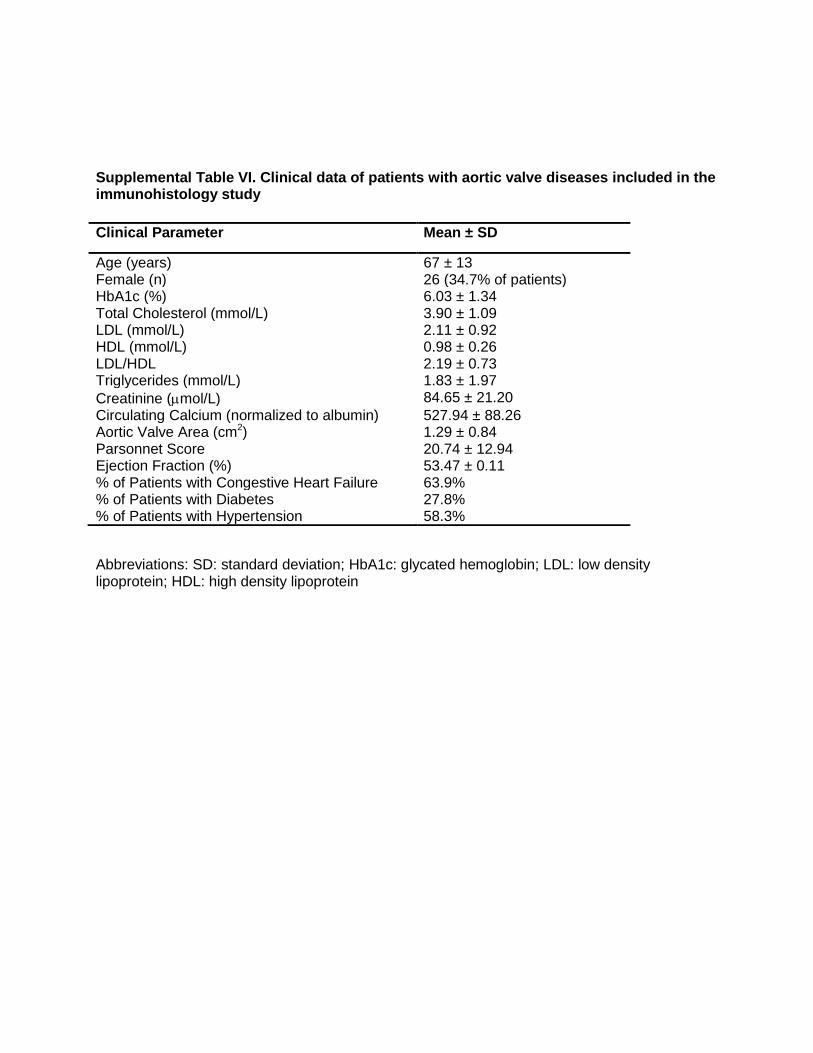
Supplemental Table VI. Clinical data of patients with aortic valve diseases included in the immunohistology study
Abbreviations: SD: standard deviation; HbA1c: glycated hemoglobin; LDL: low density lipoprotein; HDL: high density lipoprotein
Clinical Parameter Mean ± SD
Age (years) 67 ± 13 Female (n) 26 (34.7% of patients) HbA1c (%) 6.03 ± 1.34 Total Cholesterol (mmol/L) 3.90 ± 1.09 LDL (mmol/L) 2.11 ± 0.92 HDL (mmol/L) 0.98 ± 0.26 LDL/HDL 2.19 ± 0.73 Triglycerides (mmol/L) 1.83 ± 1.97
Creatinine (mol/L) 84.65 ± 21.20
Circulating Calcium (normalized to albumin) 527.94 ± 88.26 Aortic Valve Area (cm2) 1.29 ± 0.84 Parsonnet Score 20.74 ± 12.94 Ejection Fraction (%) 53.47 ± 0.11 % of Patients with Congestive Heart Failure 63.9% % of Patients with Diabetes 27.8% % of Patients with Hypertension 58.3%
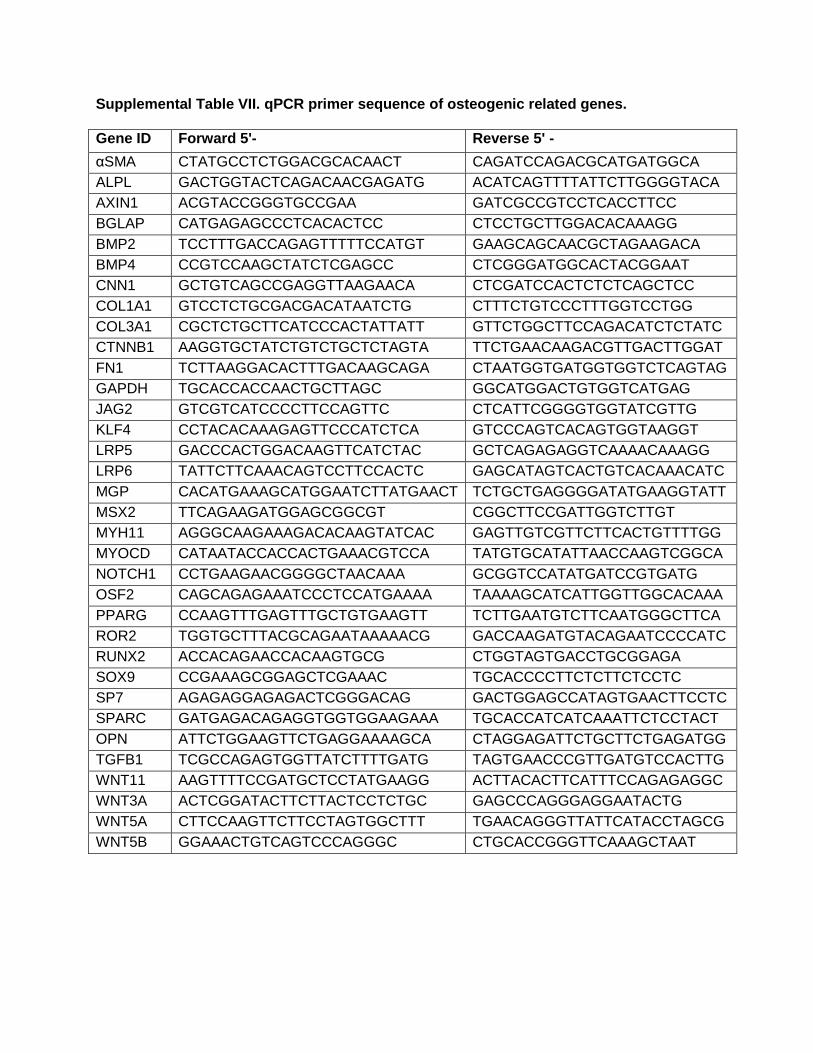
Supplemental Table VII. qPCR primer sequence of osteogenic related genes.
Gene ID Forward 5'- Reverse 5' -
αSMA CTATGCCTCTGGACGCACAACT CAGATCCAGACGCATGATGGCA
ALPL GACTGGTACTCAGACAACGAGATG ACATCAGTTTTATTCTTGGGGTACA
AXIN1 ACGTACCGGGTGCCGAA GATCGCCGTCCTCACCTTCC
BGLAP CATGAGAGCCCTCACACTCC CTCCTGCTTGGACACAAAGG
BMP2 TCCTTTGACCAGAGTTTTTCCATGT GAAGCAGCAACGCTAGAAGACA
BMP4 CCGTCCAAGCTATCTCGAGCC CTCGGGATGGCACTACGGAAT
CNN1 GCTGTCAGCCGAGGTTAAGAACA CTCGATCCACTCTCTCAGCTCC
COL1A1 GTCCTCTGCGACGACATAATCTG CTTTCTGTCCCTTTGGTCCTGG
COL3A1 CGCTCTGCTTCATCCCACTATTATT GTTCTGGCTTCCAGACATCTCTATC
CTNNB1 AAGGTGCTATCTGTCTGCTCTAGTA TTCTGAACAAGACGTTGACTTGGAT
FN1 TCTTAAGGACACTTTGACAAGCAGA CTAATGGTGATGGTGGTCTCAGTAG
GAPDH TGCACCACCAACTGCTTAGC GGCATGGACTGTGGTCATGAG
JAG2 GTCGTCATCCCCTTCCAGTTC CTCATTCGGGGTGGTATCGTTG
KLF4 CCTACACAAAGAGTTCCCATCTCA GTCCCAGTCACAGTGGTAAGGT
LRP5 GACCCACTGGACAAGTTCATCTAC GCTCAGAGAGGTCAAAACAAAGG
LRP6 TATTCTTCAAACAGTCCTTCCACTC GAGCATAGTCACTGTCACAAACATC
MGP CACATGAAAGCATGGAATCTTATGAACT TCTGCTGAGGGGATATGAAGGTATT
MSX2 TTCAGAAGATGGAGCGGCGT CGGCTTCCGATTGGTCTTGT
MYH11 AGGGCAAGAAAGACACAAGTATCAC GAGTTGTCGTTCTTCACTGTTTTGG
MYOCD CATAATACCACCACTGAAACGTCCA TATGTGCATATTAACCAAGTCGGCA
NOTCH1 CCTGAAGAACGGGGCTAACAAA GCGGTCCATATGATCCGTGATG
OSF2 CAGCAGAGAAATCCCTCCATGAAAA TAAAAGCATCATTGGTTGGCACAAA
PPARG CCAAGTTTGAGTTTGCTGTGAAGTT TCTTGAATGTCTTCAATGGGCTTCA
ROR2 TGGTGCTTTACGCAGAATAAAAACG GACCAAGATGTACAGAATCCCCATC
RUNX2 ACCACAGAACCACAAGTGCG CTGGTAGTGACCTGCGGAGA
SOX9 CCGAAAGCGGAGCTCGAAAC TGCACCCCTTCTCTTCTCCTC
SP7 AGAGAGGAGAGACTCGGGACAG GACTGGAGCCATAGTGAACTTCCTC
SPARC GATGAGACAGAGGTGGTGGAAGAAA TGCACCATCATCAAATTCTCCTACT
OPN ATTCTGGAAGTTCTGAGGAAAAGCA CTAGGAGATTCTGCTTCTGAGATGG
TGFB1 TCGCCAGAGTGGTTATCTTTTGATG TAGTGAACCCGTTGATGTCCACTTG
WNT11 AAGTTTTCCGATGCTCCTATGAAGG ACTTACACTTCATTTCCAGAGAGGC
WNT3A ACTCGGATACTTCTTACTCCTCTGC GAGCCCAGGGAGGAATACTG
WNT5A CTTCCAAGTTCTTCCTAGTGGCTTT TGAACAGGGTTATTCATACCTAGCG
WNT5B GGAAACTGTCAGTCCCAGGGC CTGCACCGGGTTCAAAGCTAAT

Supplemental Table VIII. Immunohistochemistry staining score
Score Distribution Intensity
0 No stain No stain
1 < 5% of ECM or cells Faint
2 < 25% of ECM or cells Light Brown
3 < 50% of ECM or cells Medium Brown
4 < 75% of ECM or cells Deep Brown
5 > 75% of ECM or cells Dark Brown

Supplemental Table IX. Aortic valve histopathology score
Feature Score: 0 Score: 1 Score: 2 Score: 3 Score: 4
Calcification None Calcium deposition in < 25% of tissue
Calcium deposition in 25-50% of tissue
Calcium deposition in >50% of tissue
N/A
Fibrosis (Mahmut et al.)
None Section surface < 25% fibrosis
Section surface 25-50% fibrosis
Section surface > 50% fibrosis
N/A
Remodeling (Warren et al.)
No fibrosis. Mild fibrous thickening. Cusp structural integrity maintained.
Moderate valve thickening. Some calcification.
Extensive thickening with a lot of calcification.
Extensive thickening and calcification. Destruction of most structural components.
Inflammation None Occasional cells or 1 group with > 50 cells
2-5 groups of > 50 cells
> 5 groups with > 50 cells or 1 group with > 500 cells
N/A
Lipids No lipid core Small lipid core (< 25% tissue surface)
Core is 25-50% lipid
Core is > 50% lipid
N/A
Microvessels None < 10 microvessels per section
10-20 microvessels per section
> 20 microvessels per section
N/A

Supplemental Table X. Raman Spectra Peak assignment
Raman shift (cm-1) Assignment
425 2 PO4 590 4 PO4 621 C-C Phenylalanine 758 Tryptophan
796 U, A, C (DNA)
960 1 PO4 1001 Phenylalanine
1031 C-H Phenylalanine 1078 3 PO4 1155 C-C, ν C-N in proteins 1182 C-H Tyrosine, Phenylalanine 1197 - 1200 C-C6H6 stretching Phe, Tyr, Trp 1329 C-H proteins 1450 C-H deformation proteins
1583 A, G (DNA)
1602 C=C Phe, Tyr
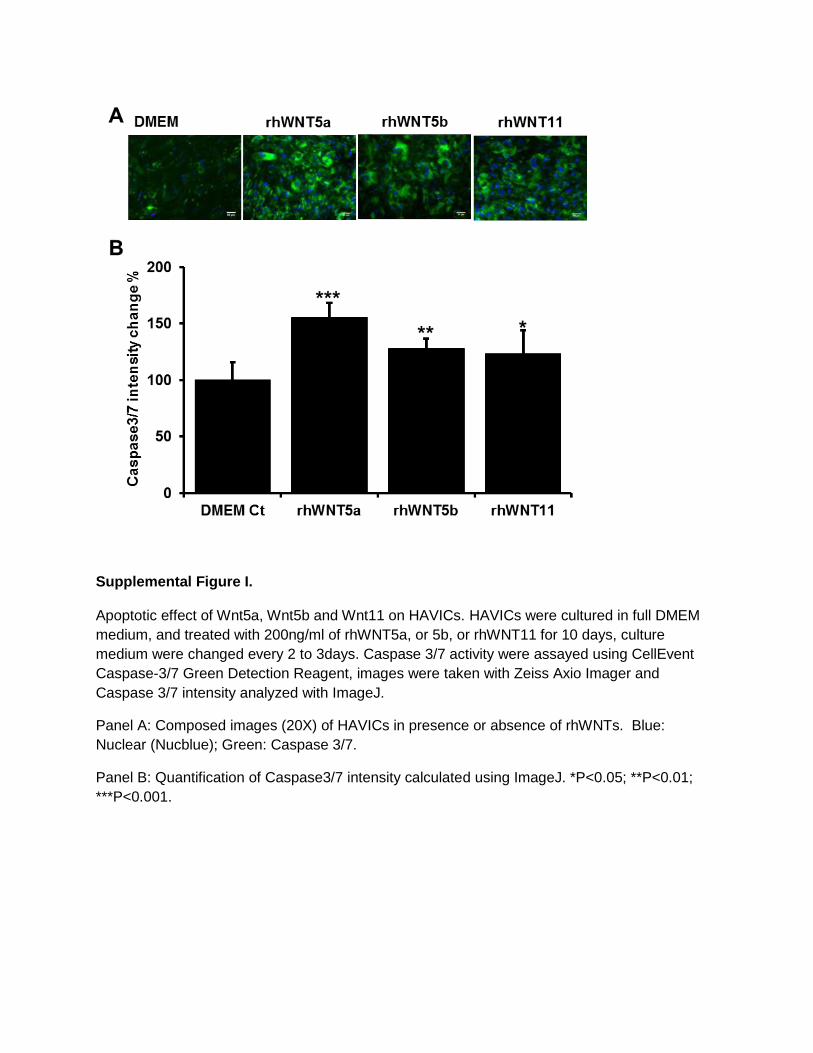
Supplemental Figure I.
Apoptotic effect of Wnt5a, Wnt5b and Wnt11 on HAVICs. HAVICs were cultured in full DMEM
medium, and treated with 200ng/ml of rhWNT5a, or 5b, or rhWNT11 for 10 days, culture
medium were changed every 2 to 3days. Caspase 3/7 activity were assayed using CellEvent
Caspase-3/7 Green Detection Reagent, images were taken with Zeiss Axio Imager and
Caspase 3/7 intensity analyzed with ImageJ.
Panel A: Composed images (20X) of HAVICs in presence or absence of rhWNTs. Blue:
Nuclear (Nucblue); Green: Caspase 3/7.
Panel B: Quantification of Caspase3/7 intensity calculated using ImageJ. *P<0.05; **P<0.01;
***P<0.001.
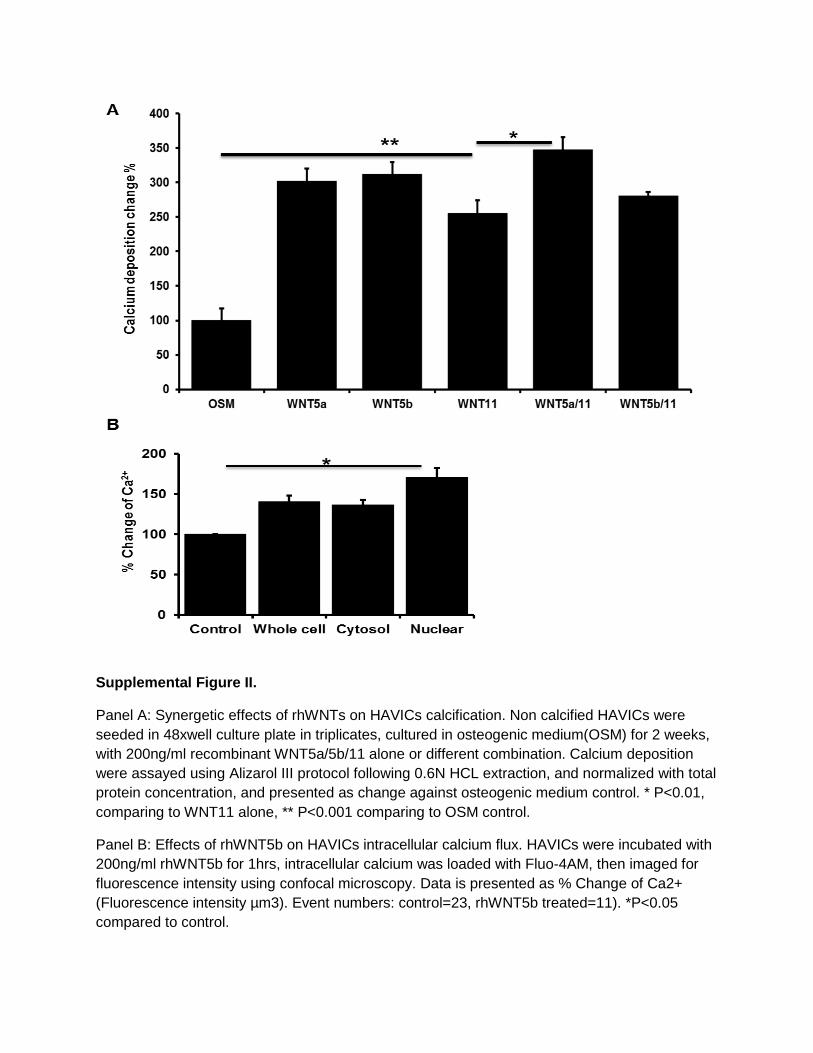
Supplemental Figure II.
Panel A: Synergetic effects of rhWNTs on HAVICs calcification. Non calcified HAVICs were
seeded in 48xwell culture plate in triplicates, cultured in osteogenic medium(OSM) for 2 weeks,
with 200ng/ml recombinant WNT5a/5b/11 alone or different combination. Calcium deposition
were assayed using Alizarol III protocol following 0.6N HCL extraction, and normalized with total
protein concentration, and presented as change against osteogenic medium control. * P<0.01,
comparing to WNT11 alone, ** P<0.001 comparing to OSM control.
Panel B: Effects of rhWNT5b on HAVICs intracellular calcium flux. HAVICs were incubated with
200ng/ml rhWNT5b for 1hrs, intracellular calcium was loaded with Fluo-4AM, then imaged for
fluorescence intensity using confocal microscopy. Data is presented as % Change of Ca2+
(Fluorescence intensity µm3). Event numbers: control=23, rhWNT5b treated=11). *P<0.05
compared to control.
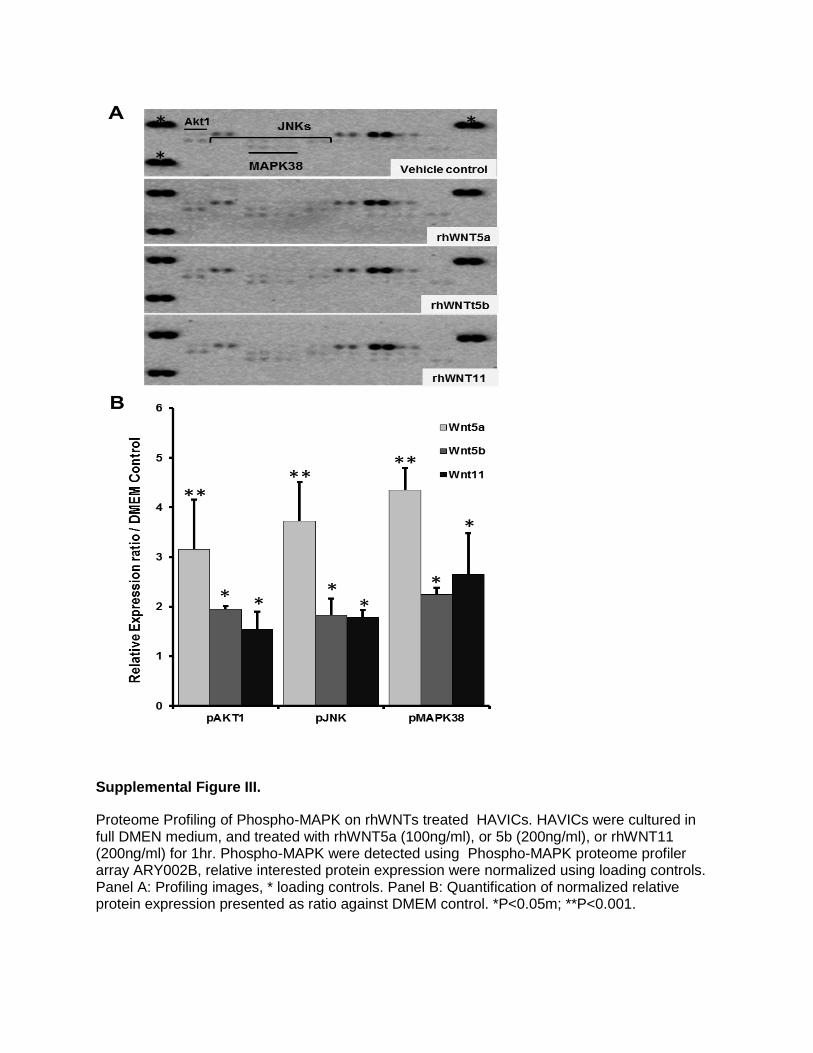
Supplemental Figure III.
Proteome Profiling of Phospho-MAPK on rhWNTs treated HAVICs. HAVICs were cultured in full DMEN medium, and treated with rhWNT5a (100ng/ml), or 5b (200ng/ml), or rhWNT11 (200ng/ml) for 1hr. Phospho-MAPK were detected using Phospho-MAPK proteome profiler array ARY002B, relative interested protein expression were normalized using loading controls. Panel A: Profiling images, * loading controls. Panel B: Quantification of normalized relative protein expression presented as ratio against DMEM control. *P<0.05m; **P<0.001.
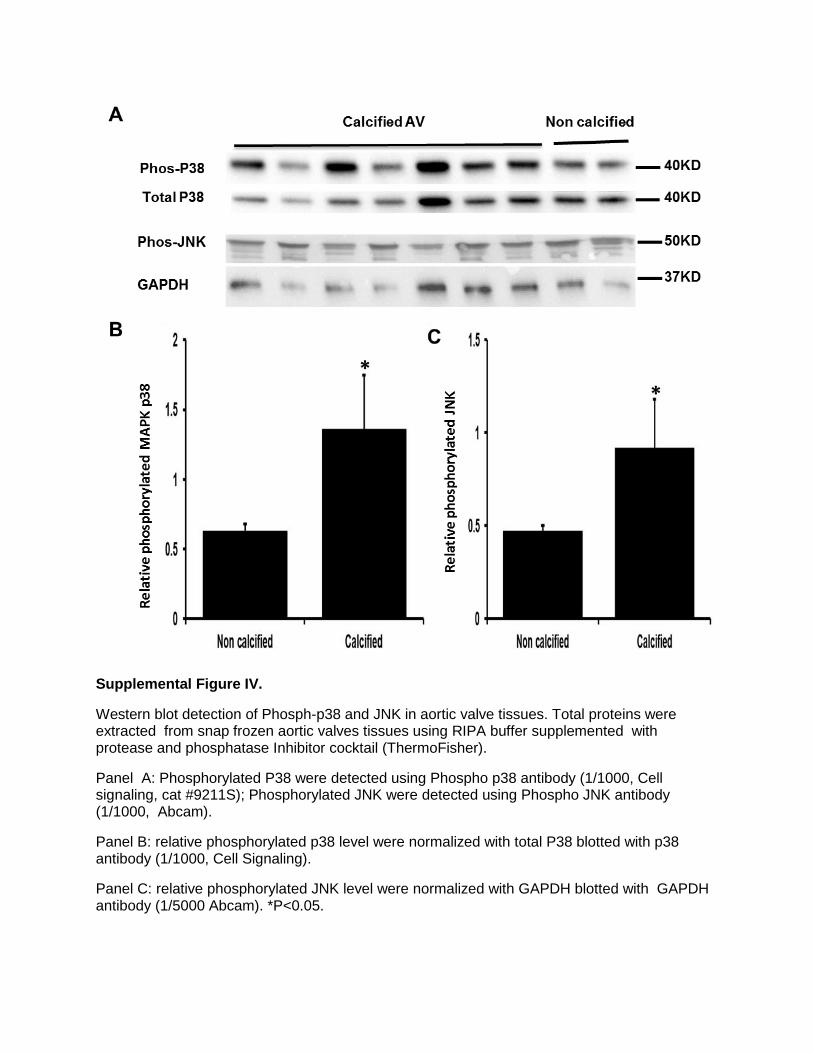
Supplemental Figure IV.
Western blot detection of Phosph-p38 and JNK in aortic valve tissues. Total proteins were extracted from snap frozen aortic valves tissues using RIPA buffer supplemented with protease and phosphatase Inhibitor cocktail (ThermoFisher).
Panel A: Phosphorylated P38 were detected using Phospho p38 antibody (1/1000, Cell signaling, cat #9211S); Phosphorylated JNK were detected using Phospho JNK antibody (1/1000, Abcam).
Panel B: relative phosphorylated p38 level were normalized with total P38 blotted with p38 antibody (1/1000, Cell Signaling).
Panel C: relative phosphorylated JNK level were normalized with GAPDH blotted with GAPDH antibody (1/5000 Abcam). *P<0.05.
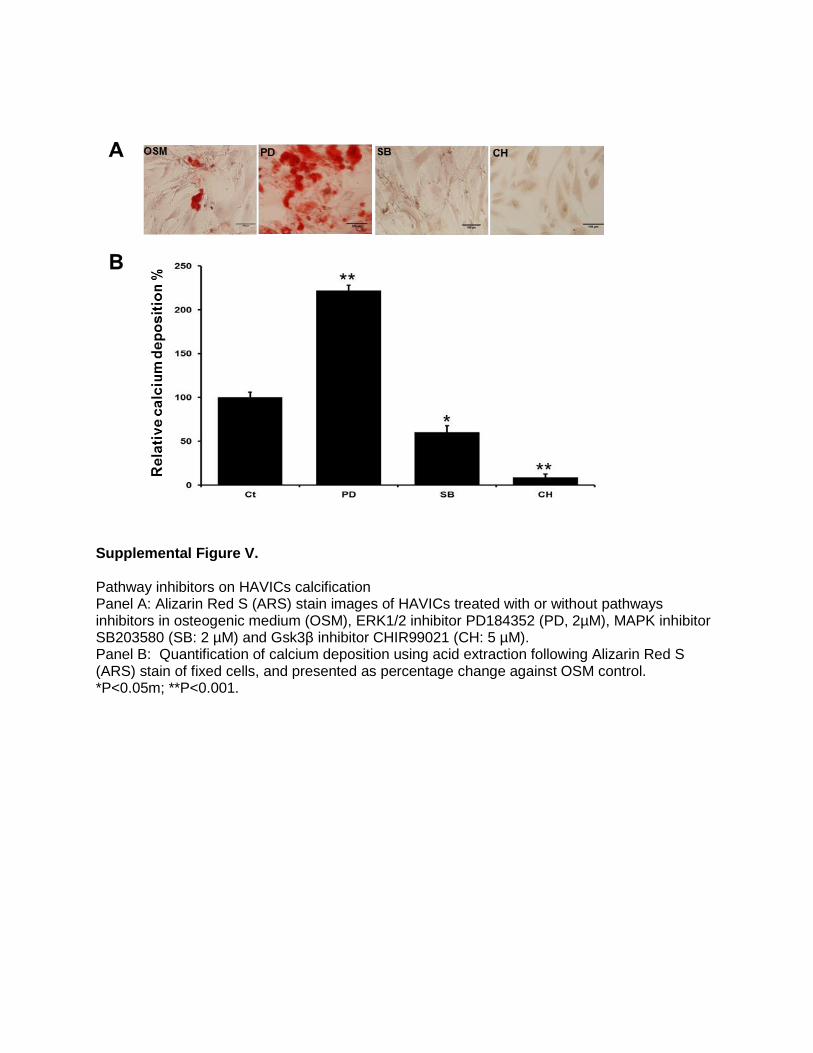
Supplemental Figure V.
Pathway inhibitors on HAVICs calcification
Panel A: Alizarin Red S (ARS) stain images of HAVICs treated with or without pathways inhibitors in osteogenic medium (OSM), ERK1/2 inhibitor PD184352 (PD, 2µM), MAPK inhibitor SB203580 (SB: 2 µM) and Gsk3β inhibitor CHIR99021 (CH: 5 µM). Panel B: Quantification of calcium deposition using acid extraction following Alizarin Red S (ARS) stain of fixed cells, and presented as percentage change against OSM control. *P<0.05m; **P<0.001.
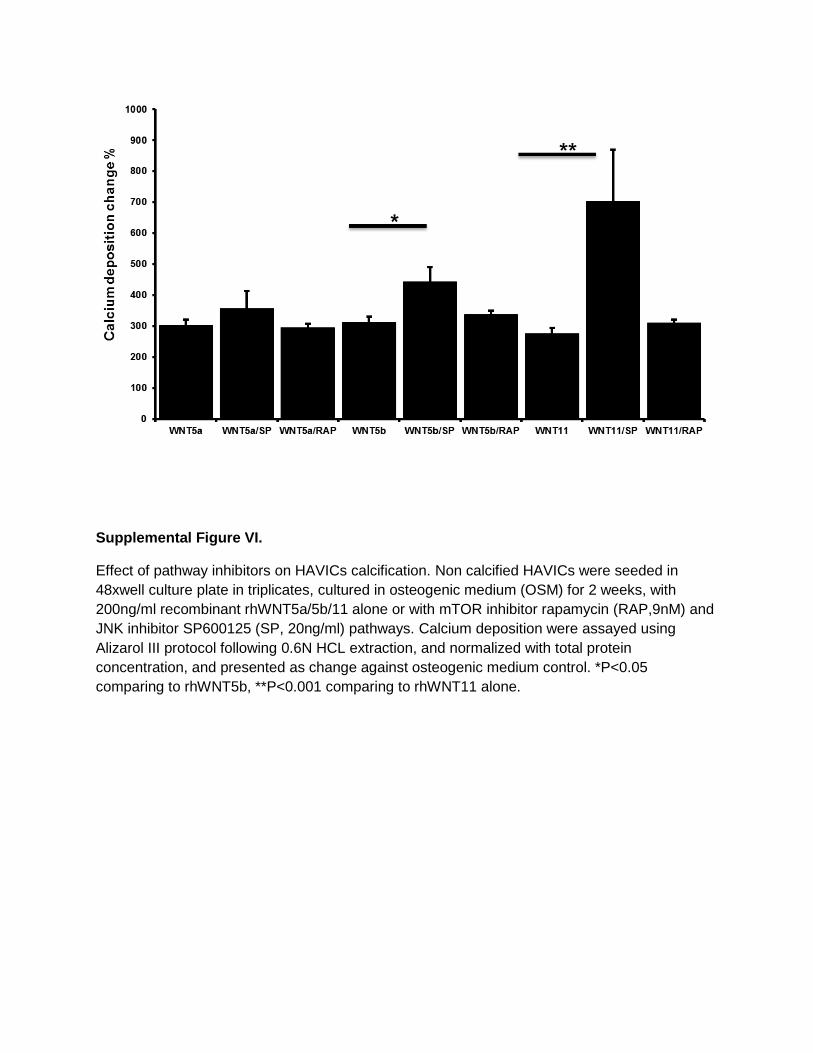
Supplemental Figure VI.
Effect of pathway inhibitors on HAVICs calcification. Non calcified HAVICs were seeded in
48xwell culture plate in triplicates, cultured in osteogenic medium (OSM) for 2 weeks, with
200ng/ml recombinant rhWNT5a/5b/11 alone or with mTOR inhibitor rapamycin (RAP,9nM) and
JNK inhibitor SP600125 (SP, 20ng/ml) pathways. Calcium deposition were assayed using
Alizarol III protocol following 0.6N HCL extraction, and normalized with total protein
concentration, and presented as change against osteogenic medium control. *P<0.05
comparing to rhWNT5b, **P<0.001 comparing to rhWNT11 alone.

Methods and Materials
Tissue Collection Archived heart valve tissue blocks (fixed in formalin and embedded in paraffin) were obtained in accordance with the McGill University Health Centre guidelines. These aortic and mitral valves were originally obtained from cardiac valve replacement surgeries between 2009 and 2012 at the McGill University Health Centre. A total of 17 non-calcified aortic valves, 55 calcified aortic valves and 18 non-calcified mitral valves) were collected for examination along with detailed clinical history (Supplemental Table VI). List of medications, lipid profile, the jet velocity through the aortic valve in cm/s (jet), the maximum pressure gradient across the valve (Pmax), the mean pressure gradient across the valve (Pmean) and whether or not the valve was identified in pathology as being calcified were all noted as well. All patients had severe heart failure (NYHA class III-IV). Additional aortic valves with focal calcification (n=5) and severely calcified (advanced) aortic valves (n=16) were also collected freshly at surgery for valve replacement. Five normal valves that had previously been collected from explanted hearts taken from patients receiving cardiac transplantation (mean age 53±19; 3 males) and stored at -80°C were obtained from the tissue bank at the Montreal Heart Institute. Small segments of 100-200mg of the valves were snap frozen in liquid nitrogen and stored at -80°C, segments adjacent to calcification nodule were used for subsequent protein and RNA extraction respectively. The study was approved by the McGill University Health Centre and the Montreal Heart Institute ethics committees; we have obtained informed consent from all participants involved in this study. Tissue expression of Wnts mRNA
Total RNAs were extracted from snap frozen aortic valve tissues using TRIzol (Invitrogen)/RNeasy Mini Kit (Qiagen) combining protocol. Total RNAs (1μg) were used for first strand cDNAs synthesis using SuperScript III First-Strand Synthesis System (Invitrogen). qPCRs were performed using QuantiFast® SYBR Green PCR kit (Qiagen) on LightCycler 1.5 (Roche). The qPCR data were analysed using PrimePCR Analysis (Bio-RAD), relative mRNA expressions of interested genes were normalized using housekeeping gene GAPDH. Sequences of gene-specific primers were list in Supplemental Table VII. Immunohistochemistry The paraffin-embedded tissue blocks were cut into 4μm sections using a microtome, six sections (first one of each five continued sections) were examined per lesion (for each antiserum) at ~20 μm intervals, and a total of 6 images were analyzed per section for histological scoring. The sections were incubated for 1 hour in 10% normal goat serum/PBS solution, then incubated overnight with primary antibodies in 0.1% BSA/PBS solution in humid chambers at 4°C. Primary antibodies used were: mouse anti human Wnt5a (Sigma Aldrich, cat#:SAB1402393 1/100), α-smooth muscle actin (Cedarlane, Cat#:CLT9000, 1/500); rabbit anti-human Wnt5b (Abcam ab115563, 1/500), Wnt11 (Thermo Scientific PA5-21712, 1/500), CD68 (Santa Cruz, sc9139 at 1:2000), ALPL (Sigma, HPA 007105 at 1/50), osteopontin (Bioworld, BS1264 at 1:100), CTNNB1(Sigma, HPA029159 at 1:200) and BMP2 (Novus, NBP1-19751 at 1:200). Secondary biotinylated goat anti-mouse IgG or goat anti-rabbit IgG (Vector Labs, BA1000, BA9200 at 1:200) were applied respectively, then followed by using Vectastain ABC Elite Kit (Vector Laboratories, Burlingame, CA) according to manufacturer protocols. Immunostaining was visualized by 1xDAB/H2O2 solution, subsequently counterstained with hematoxylin, and mounted with Permount (Sigma). Immunostaining without primary antibody

were done as negative control. Alizarin Red S and Masson’s Trichrome staining protocols were used for Calcium and collagen detection, respectively. Aortic valves were semi-quantitatively scored for overall immunostaining (intensity and distribution) in segments of fibrosis, calcified foci and surrounding extracellular matrix (ECM), microvessels and inflammatory cells (Supplemental Table VIII). Aortic valves were also assessed for overall level of calcification, fibrosis, remodeling,1 inflammation, lipid, and microvessel formation (Supplemental Table IX).2-4 Isolation and culture of Human Aortic Valve Interstitial Cells (HAVICs) Primary HAVICs lines were generated from fresh surgically removed human valve leaflets, and cultured in DMEM high glucose medium containing 10% FBS and 1x streptomycin/penicillin solution (Invitrogen).5 Briefly, aortic valve leaflets were washed with 1xHBSS buffer (Invitrogen), , cut into small pieces and incubated in DMEM media with Collagenase Type II (Sigma, 100U/ml) for 3 hours at 37°C in water bath with occasional vortex. The fully digested mixture were centrifuged at 500g at 4°C for 10 minutes; the supernatant was transferred into a new 50ml centrifuge tube and centrifuged at 1000g at 4°C for 10 minutes. Cell pellets were suspended in complete DMEM supplemented with 10% FBS, 1x Antibiotic Antimycotic solution (Invitrogen), and seeded in 75 cm2 culture flasks. HAVICs cells at passages 3 to 5 were used for experiments. HAVICs morphology is similar to myofibroblasts which are generally spindle shaped and spread evenly throughout the flask, and stained positive for anti-alpha smooth muscle actin (data not shown). For calcification inducing, HAVICs were cultured in osteogenic medium (OSM) which is of full DMEM medium supplemented with 2mM phosphate buffer (pH7.4). Western blotting Cultured HAVICs isolated from both calcified and non-calcified aortic valves, and human dermal fibroblast cells (ThermoFisher, C0135C) were re-suspended in RIPA buffer supplemented with cOmplete™, Mini, EDTA-free Protease Inhibitor Cocktail (Roche, 11836170001) for protein extraction. Samples were then centrifuged at 15000rpm at 4°C for 10 minutes and supernatants were collected, protein concentrations were measured by Protein DC protocol (Bio-Rad) and equal amount of total protein were loaded for 10% SDS-PAGE gel separation. The protein was transferred onto PVDF membrane at 300mA for 1 hour at RT, blocked for 1 hour at RT with 5% milk (1xTBST buffer), then incubated with primary antibodies against Wnt5a, Wnt5b or Wnt11 for 16hrs at 4°C; then followed by incubation with corresponding secondary antibodies: bovine anti-mouse, or anti-rabbit IgG-HRP (Santa Cruz Biotechnology, cat#sc-2371, sc-2370 at 1/5000), For signal development, Lumi-Light Western Blotting Substrate (Roche) was used according to manufacturer protocols. Measurement of Alkaline Phosphatase (ALP) activity Calcified and non-calcified HAVICs at 100% confluence were treated with rhWnt5a, rhWnt5b, rhWnt11(R & D Systems) at 100ng/ml, 200ng/ml and 300ng/ml for 48hrs. Medium and cellular ALP activities were assessed using Alkaline Phosphatase Activity Colorimetric Assay Kit (BioVision) according to manufacturer protocols. Briefly, culture medium was harvested and spun for 10mins at 12,000g; supernatant was used for ALP activity assay. Cells were washed twice with PBS, then incubated with shaking for 10 minutes at RT in cell lysis buffer (20mM Tris-

HCl pH=8.0, 150 mM NaCl, 0.2% NP40). Cell lysates were harvested and spun at 12000g for 5
minutes at 4C, supernatant was harvested for protein and ALP activity assay. Non-calcified and calcified HAVICs were also treated with rhWnts for 21 days in OSM or DMEM to determine effect of prolong incubation of rhWnts.
Cell proliferation assay Cell proliferation assay were performed using MTT (Thiazolyl Blue Tetrazolium Blue, Sigma) protocol. Briefly, HAVICs were incubated in the presence or absence of 200ng/ml rhWnt5a, rhWnt5b or rhWnt11 for 2 or 20 days in full DMEM medium or OSM. MTT solution (5mg/ml in DMEM) was added to cultured cells in 24xwell plates, and incubated for 2-3hrs at 37°C, then purple precipitation were extracted using MTT solvent (isopropanol with 4mM HCl, 0.1% Nondet P-40), and measured at OD570nm with 96xwell reading plate on Spectra Photometer (BioTek). Assessment of HAVICs apoptosis Apoptosis were assayed using Alexa Fluor 488 annexin V/Dead cell apoptosis kit for Flow Cytometry(ThermoFisher), the kit supplied prodidium iodide(PI) was replaced by FVD efluor 780 (Affymetrix eBioscience) because of the high background staining with HAVICs. Briefly, non-calcified HAVICs were treated with 200ng/ml of rhWNT5a, rhWNT5b or rhWNT11, and Wnt signalling inhibitor XAV939 (20µM) for 4hrs or 24hrs in OSM, cells were harvested by trypsin, and stained for Alexa Fluor 488 annexin V and FVD efluor 780 following manufacture’s instruction manual. Flow Cytometry data were collected using BD FACSCanto II, and analysed using FLowJo at the service of Immunophenotyping Platform at RI-MUHC. Prolonged incubation of rhWNTs on HAVICs apoptosis effects were also assessed using CellEvent Caspase-3/7 Green Detection Reagent (Invitrogen). Non-calcified HAVICs were seeded in slide chamber at 100% confluence for overnight, then changed with fresh medium, and treated with 200ng/ml rhWnt5a, rhWnt5b or rhWnt11 in complete DMEM for 10 days. Apoptosis were); briefly, cells were live stained with NucBlue (Molecular Probe) and Caspases 3/7 Green live staining kits. Caspase-3/7 staining was reviewed under fluorescence microscope (Zeiss Axio Imager), and Caspase-3/7 stain intensity were analysed using ImageJ with 9 random views from each group. Measurement of calcium mineralization Alizarin Red S (ARS) staining was used to visualize the calcium deposition and calcification nodule formation. Briefly, HAVICs treated with or without rhWnt5a, rhWnt5b or rhWnt11 at 100 and 200ng/ml in OSM for 3 weeks were fixed for 30min using 4% buffered formaldehyde at room temperature. Cells were then washed 4 times with excessive ddH2O, incubated with Alizarin Red S stain solution (40mM, pH4.2) for 20 minutes at room temperature, and again washed 4 times with excessive ddH2O. Calcification nodules were observed under microscopy, and phase contrast images were taken with inverted microscopy. After removing the ddH2O, calcium bound ARS were extracted by incubating with 10% Cetylpyridinium chloride (Sigma) solution at room temperature for 60 minutes. Extracts were transferred into new 1.5ml centrifuge tube, spun for 5 minutes at 12,000g and supernatant was aliquot in 96xwell reading plate for OD 405nm reading on Spectra Photometer (BioTek). Cells were further washed twice with ddH2O, and scraped into 1.5 ml centrifuge tube in cell lysis buffer (20mM Tris HCl pH7.4 with 2%SDS, 0.2M glycine), and heat treated at 80°C for 60 minutes with occasional vortex. Cell lysate were

centrifuged at 12,000g for 10 min at 4°C, total protein assayed as described above, and used for ARS normalization. For comparison, we also measured calcium deposition using Arsenazo III protocol following acid calcium extraction (0.6N HCL),6 and observed similar data to that of ARS protocol. For pathway inhibitors, non-calcified HAVICs were treated with the MAPKP38 inhibitor
SB203580 (2μM), GSK3 inhibitor CHIR99021 (5µM) and the ERK1/2 inhibitor PD184352 (2μM) for 3 weeks, calcification nodule and calcium deposition were assayed as above. Non-calcified HAVICs were also incubated with different combinations of rhWNTs (200ng/ml) and inhibitors of mTOR (SP600125, 20ng/ml) or JNK (Rapamycin, 9nM) pathways for 2 weeks in osteogenic medium, calcium deposition were assayed using Alizarol III protocol following 0.6N HCL extraction, and normalized with total protein concentration, and presented as change against osteogenic medium control. Raman spectroscopy Raman spectra were collected on dried pellets from cell lysates of HAVICs cultured in OSM in the presence or absence of 200ng/ml rhWnt5a, rhWnt5b or rhWnt11 for 21 days. To prepare these pellets, the lysates were centrifuged at 6,000 rpm for 20 min; then, the supernatants were removed and the pellets dried in vacuum overnight. The dried pellets were analyzed using a Raman spectrophotometer (Senterra, Bruker, Germany) equipped with a 785 nm diode laser of 100 mW power coupled with an optical microscope. Raman spectra were acquired using a 40x objective, in a spectral range from 400 to 1800 cm-1 at a resolution of 3.5 cm-1. Three different spots were scanned for each sample with an integration time of 60s and 4 co-additions. This peak corresponds to the ν1 vibration of the phosphate group, and is indicative of the formation of hydroxyapatite (HA) on these samples, all peaks were assigned accordingly (Supplemental
Table X). 7-12
Assessment of phosphorylated MAPKs Non-calcified HAVICs were treated with 100ng/ml rhWNT5a, 200ng/ml rhWNT5b or 200ng/ml rhWNT11 for 1hr in OSM and Phosphorlyated MAPKs were assayed using Proteomic ProfilerTM of Human Phospho-MAPK Array (R&D system) according to manufacturer’s instruction. Gene expression profiling of osteogenic mediators Non-calcified HAVICs were treated with rhWnt5a, rhWnt5b, rhWnt11 at 200ng/ml for 48hrs in complete DMEM medium or OSM. Total RNAs were isolated using RNeasy Mini kit (Qiagen), Total RNAs (1µg) was used for the first strand cDNA synthesis using Supertriscript III system (Lifetechnologies), qPCR were performed on Lightcycler 1.5 using SYBR fast qPCR kit (Qiagen). qPCR data were analysed using PrimerPCR software (Bio-RAD). Briefly, interested gene expression were first normalized with GAPDH gene expression using ΔΔCT protocol, then
relative normalized gene expression were calculated against relative control group, presented as relative normalized gene expression (fold) and regulation changes. More than 30 osteogenic related genes including WNTs, receptors, upstream and downstream regulatory genes are included (Supplemental Table VII). Gene expression of some of those osteogenic related genes were also analyzed on rhWNTs treated HAVICs isolated from calcified aortic valve. Confocal microscopy to analyze cytosolic and nuclear calcium influx

Cultured non-calcified HAVICs were treated with 200ng/ml rhWNT5b for 1 hr., and then loaded with 13 µmol/L of Fluo-4/AM (Molecular Probes). This analysis was done at the facility of Dr. Bkaily G (University of Sherbrooke, Quebec, Canada) following their well-established protocol.13,
14 Briefly, loaded HAVICs were examined with a Molecular Dynamics (Sunnyvale, California, USA) MultiProbe 2001 confocal system. Total16 sections with a step size of 0.1µm for each cell were obtained to constitute the real 3-D image using Molecular Dynamics' ImageSpace analysis and Volume software. The nucleus was stained with SYTO-11(Molecular Probes). Statistical Analysis Multivariant Spearman correlation analysis was used to test the association between the different Wnt immunohistochemical staining with their tissue analysis, mRNA expression and clinical parameters. One-way ANOVA Kruskal-Wallis tests were used to assess Wnt5b and Wnt11 immunostaining in normal aortic and mitral valves and normal segments of diseased valves compared to intensity of staining in segments of fibrosis, actively calcifying foci and extracellular matrix, areas of mature calcification, microvessels and inflammatory cells. It was also used to compare the effect of Wnts on ALP activity and calcification. Unpaired t tests were used to assess statistically significant differences in mRNA expression, Western blot densitometries measurements. Statistical analysis was performed using SPSS program Version 15.

References
1. Warren BA, Yong JL. Calcification of the aortic valve: Its progression and grading. Pathology. 1997;29:360-368
2. Mahmut A, Boulanger MC, Fournier D, Couture C, Trahan S, Page S, Arsenault B, Despres JP, Pibarot P, Mathieu P. Lipoprotein lipase in aortic valve stenosis is associated with lipid retention and remodelling. European journal of clinical investigation. 2013;43:570-578
3. Mohler ER, 3rd, Chawla MK, Chang AW, Vyavahare N, Levy RJ, Graham L, Gannon FH. Identification and characterization of calcifying valve cells from human and canine aortic valves. The Journal of heart valve disease. 1999;8:254-260
4. Friedman MS, Oyserman SM, Hankenson KD. Wnt11 promotes osteoblast maturation and mineralization through r-spondin 2. The Journal of biological chemistry. 2009;284:14117-14125
5. Gould RA, Butcher JT. Isolation of valvular endothelial cells. J Vis Exp. 2010:e2158 6. Attin T, Becker K, Hannig C, Buchalla W, Hilgers R. Method to detect minimal
amounts of calcium dissolved in acidic solutions. Caries Res. 2005;39:432-436 7. Penel G, Leroy G, Rey C, Bres E. Microraman spectral study of the po4 and co3
vibrational modes in synthetic and biological apatites. Calcified tissue international.63:475-481
8. Karampas IA, Kontoyannis CG. Characterization of calcium phosphates mixtures. Vibrational Spectroscopy. 2013;64:126-133
9. Baraga JJ, Feld MS, Rava RP. In situ optical histochemistry of human artery using near infrared fourier transform raman spectroscopy. Proceedings of the National Academy of Sciences. 1992;89:3473-3477
10. Salenius JP, Brennan JF, Miller A, Wang Y, Aretz T, Sacks B, Dasari RR, Feld MS. Biochemical composition of human peripheral arteries examined with nearinfrared raman spectroscopy. Journal of Vascular Surgery.27:710-719
11. Rygula A, Majzner K, Marzec KM, Kaczor A, Pilarczyk M, Baranska M. Raman spectroscopy of proteins: A review. . Journal of Raman Spectroscopy. 2013;44:1061-1076
12. Ramser K. Raman spectroscopy of single cells for biomedical applications. 2012;5:106-147
13. Bkaily G, Pothier P, D'Orleans-Juste P, Simaan M, Jacques D, Jaalouk D, Belzile F, Hassan G, Boutin C, Haddad G, Neugebauer W. The use of confocal microscopy in the investigation of cell structure and function in the heart, vascular endothelium and smooth muscle cells. Molecular and cellular biochemistry. 1997;172:171-194
14. Bkaily G, Avedanian L, Al-Khoury J, Chamoun M, Semaan R, Jubinville-Leblanc C, D'Orleans-Juste P, Jacques D. Nuclear membrane r-type calcium channels mediate cytosolic et-1-induced increase of nuclear calcium in human vascular smooth muscle cells. Canadian journal of physiology and pharmacology. 2015;93:291-297

膜的发展与 rDNA 转录活动的增强关系密切。CX-5461 主要通过诱导没有明显凋亡迹象增
殖的平滑肌细胞在 G2/M 细胞周期停滞,从而发挥调节作用。CX-5461 不能诱导 p53 的稳
定,但能够增加 p53 磷酸化和乙酰化并且能够激活共济失调毛细血管扩张突变/共济失调毛
细血管扩张和 Rad3相关的(Rad3-related, ATR)信号通路。抑制 ATR 但没有抑制共济失调
的毛细血管扩张突变基因消除了 CX-5461 的细胞抑制效应和 p53 的磷酸化作用。此外,针
对 GADD45抑制 p53或者沉默 p53 与抑制 ATR 时的效应相似。体内实验发现在用 CX-5461
治疗的血管中,p53 的磷酸化水平和乙酰化水平及共济失调毛细血管突变/ATR 通路的活动
均增强。
图 CX-5461的治疗对新生内膜增殖的影响
因此,Pol I可作为治疗靶点来抑制新生内膜的生长,这也侧面说明 Pol I是一个预防动
脉再狭窄的新型生物靶点。从机制上来说,Pol I 抑制剂通过激活 ATR-p53 轴导致平滑肌细
胞在 G2/M细胞周期停滞。
(Arterioscler Thromb Vasc Biol. 2017; 37:476-484.)
非经典Wnt信号通路在人类主动脉瓣钙化中的作用
主动脉瓣钙化是引起主动脉狭窄的主要原因,主动脉狭窄的进行性发展可导致心衰、心
绞痛、晕厥甚至死亡。主动脉狭窄在很长一段时间内没有症状,但一旦出现症状,如果不进
行主动脉瓣置换手术,其两年的死亡率大于 50%。主动脉钙化的病理机制尚不清楚。越来越
多的证据表明瓣膜钙化概括了骨骼的形成。非经典 WNT 配体 WNT5a、WNT5b 和 WNT11
在骨形成中发挥了重要作用,使得其成为主动脉瓣钙化研究中一个重要的靶点。
研究人员运用免疫组化、实时荧光定量 PCR,蛋白免疫印迹及组织培养等技术检测了钙
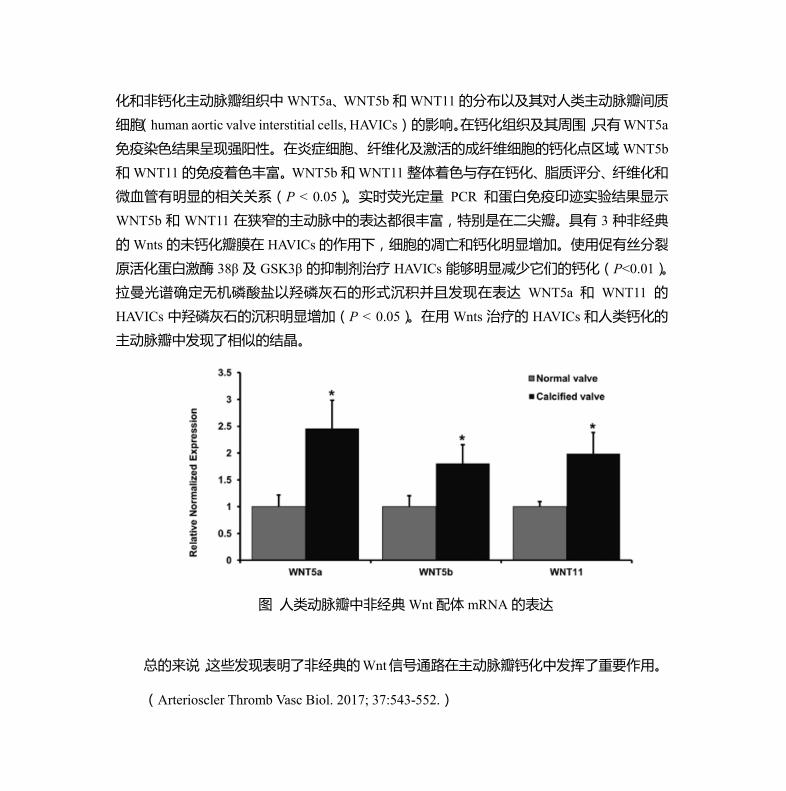
化和非钙化主动脉瓣组织中 WNT5a、WNT5b 和WNT11的分布以及其对人类主动脉瓣间质
细胞(human aortic valve interstitial cells, HAVICs) 的影响。在钙化组织及其周围,只有WNT5a
免疫染色结果呈现强阳性。在炎症细胞、纤维化及激活的成纤维细胞的钙化点区域 WNT5b
和 WNT11的免疫着色丰富。WNT5b 和WNT11 整体着色与存在钙化、脂质评分、纤维化和
微血管有明显的相关关系(P < 0.05)。实时荧光定量 PCR 和蛋白免疫印迹实验结果显示
WNT5b 和 WNT11 在狭窄的主动脉中的表达都很丰富,特别是在二尖瓣。具有 3 种非经典
的 Wnts 的未钙化瓣膜在 HAVICs 的作用下,细胞的凋亡和钙化明显增加。使用促有丝分裂
原活化蛋白激酶 38β 及 GSK3β 的抑制剂治疗 HAVICs 能够明显减少它们的钙化(P<0.01)。
拉曼光谱确定无机磷酸盐以羟磷灰石的形式沉积并且发现在表达 WNT5a 和 WNT11 的
HAVICs 中羟磷灰石的沉积明显增加(P < 0.05)。在用 Wnts 治疗的 HAVICs 和人类钙化的
主动脉瓣中发现了相似的结晶。
图 人类动脉瓣中非经典Wnt 配体 mRNA 的表达
总的来说,这些发现表明了非经典的Wnt信号通路在主动脉瓣钙化中发挥了重要作用。
(Arterioscler Thromb Vasc Biol. 2017; 37:543-552.)
在动脉粥样硬化形成过程中 P2Y12通过丝切蛋白促进了血管平滑肌细胞的迁移
动脉粥样硬化主要发生在大动脉,是引起中风、心脏疾病的重要原因,也是在全世界范
围内致死、致残的主要原因。动脉粥样硬化的形成涉及一系列的细胞和分子层面的改变,其
发生和发展的病理机制仍不明确。P2Y12是一个公认的在血小板上表达的受体,并且是噻吩
吡啶类抗血小板药物的靶点。然而,有证据表明血管壁上 P2Y12的表达在动脉粥样硬化的形
成中发挥了重要作用,但其具体的机制仍不清楚。近日,有研究者对血管壁上的 P2Y12如何
调节血管平滑肌细胞(vascular smooth muscle cells, VSMCs)迁移及促进动脉粥样硬化的形
成过程的分子机制进行了深入研究。





























