Embed Size (px)
Citation preview

Role of Models in Population Biology
Understanding Predicting Implications for
empirical work
0
50
100
150
0 10 20 30 40 50
Days
Den
sity
Rotifers(after Halbach 1979)
0
50
100
150
0 10 20 30 40 50Days
Den
sity
15 oC
25 oC

Pheasants

Data collection Regional and statewide abundance
estimates Predicting quality of coming
pheasant season
2003: 1.36 pheasants per 100 miles2004: 1.75 pheasants per 100 miles 29% increase

Commission's upland game program manager, Scott Taylor
Nebraska's pheasant population is at its highest level since 1995, according to rural mail carrier surveys conducted in the spring and summer by the Nebraska Game and Parks Commission.
The surveys indicate the population is improved over last year and substantially improved over the previous five-year average.
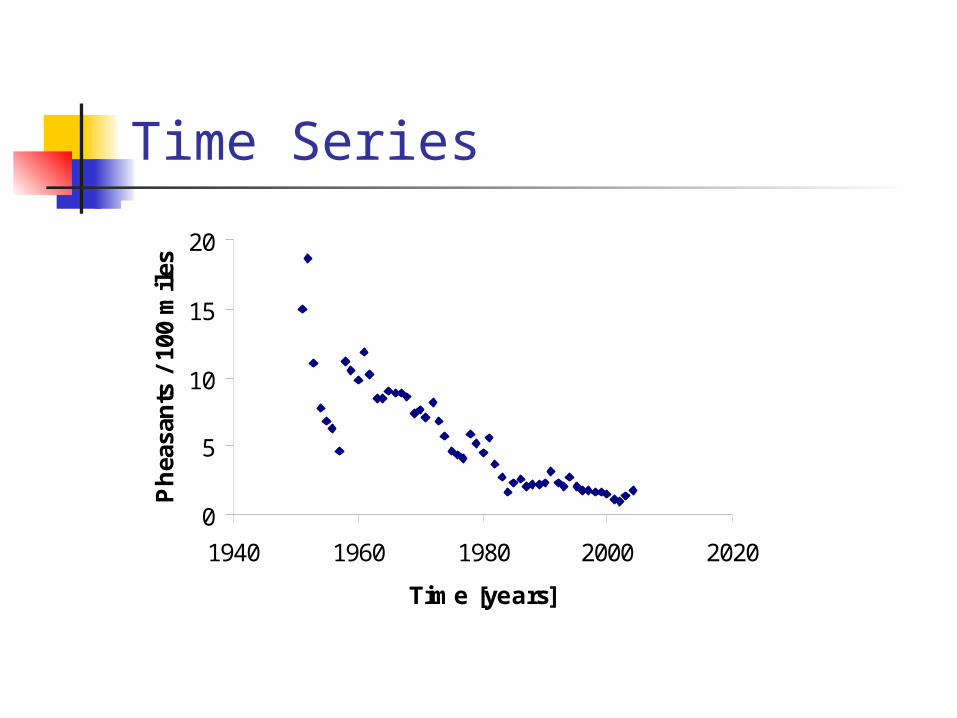
Time Series
0
5
10
15
20
1940 1960 1980 2000 2020
Time [years]
Ph
easa
nts
/ 1
00 m
iles
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
16.0
18.0
20.0
1940 1950 1960 1970 1980 1990 2000 2010
Time [years]
Ph
easa
nts
/ 1
00 m
iles
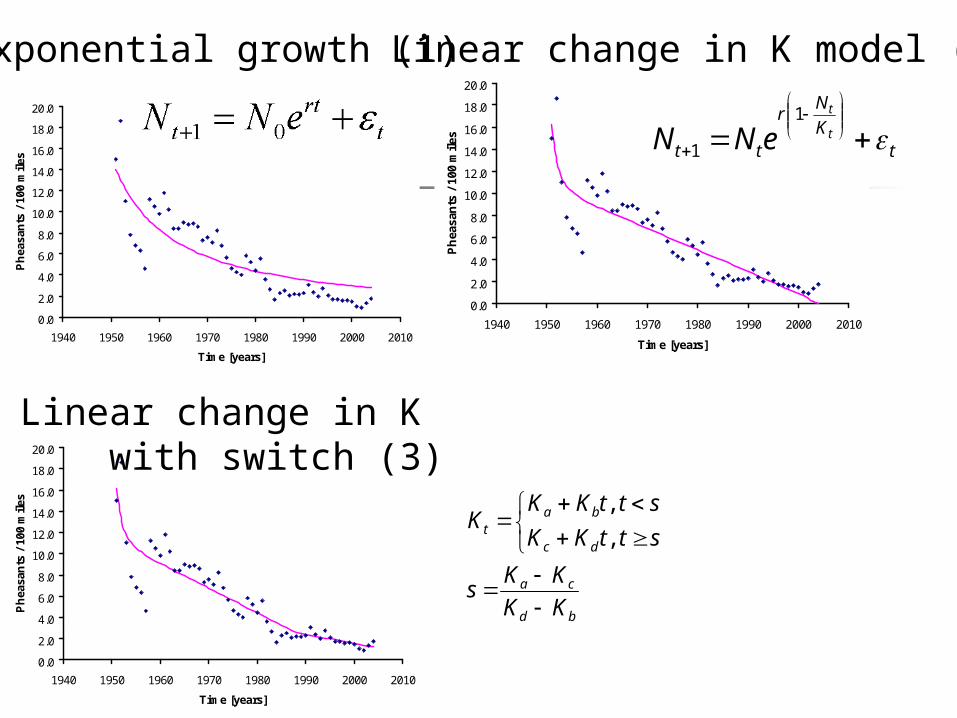
Exponential growth (1)
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
16.0
18.0
20.0
1940 1950 1960 1970 1980 1990 2000 2010
Time [years]
Ph
easa
nts
/ 1
00 m
iles
Linear change in K model (2)
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
16.0
18.0
20.0
1940 1950 1960 1970 1980 1990 2000 2010
Time [years]
Ph
easa
nts
/ 1
00 m
iles
Linear change in K with switch (3)
tK
Nr
ttt
t
eNN
1
1
bd
ca
dc
bat
KK
KKs
sttKK
sttKKK
,
,

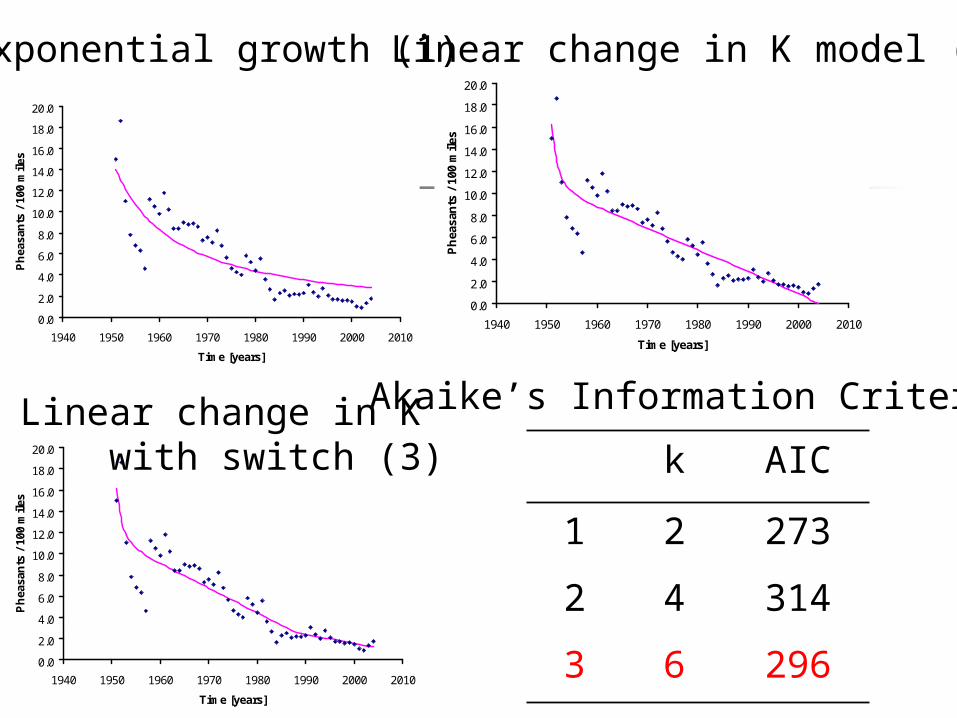
Akaike’s Information Criterion
AIC = -2L + 2k L = log likelihood ~ probability the
model is correct, given the data k = number of parameters in the
model Low AIC is better
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
16.0
18.0
20.0
1940 1950 1960 1970 1980 1990 2000 2010
Time [years]
Ph
easa
nts
/ 1
00 m
iles
Exponential growth (1)
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
16.0
18.0
20.0
1940 1950 1960 1970 1980 1990 2000 2010
Time [years]
Ph
easa
nts
/ 1
00 m
iles
Linear change in K model (2)
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
16.0
18.0
20.0
1940 1950 1960 1970 1980 1990 2000 2010
Time [years]
Ph
easa
nts
/ 1
00 m
iles
Linear change in K with switch (3)
Akaike’s Information Criterion
k AIC
1 2 273
2 4 314
3 6 296

A simple (discrete) population model
Nt+1 = Nt + B - D + I – Epop next time period
pop. this time period
births deaths immigrants emigrants
Change in abundance
Nt+1 - Nt = B - D + I - E
N = B - D + I - E

Basic closed population model
Simplifying assumption: no movement between populations Abundance
Nt+1 = Nt + B – D
Change in abundance
N = B – D
if B > D then population increaseif B< D then population decreaseif B = D then stable population

Basic discrete model
Let number of births and deaths depend on population size:
i.e. B = bN D = dNHence
Nt+1 = Nt + (b – d) Nt
Let r = b – dNt+1 = Nt + r Nt = (1 + r) Nt = R Nt
R is the finite rate of increase (a.k.a. )

Varying
0500
1000
15002000250030003500
0 1 2 3 4 5 6 7 8 9 10
time (t)
po
pu
lati
on
siz
e N
(t)
1.0
1.2
0.8

Continuous growth
Recall that
N=(b-d)Nt
As t => 0
rNdt
dN

Population growth vs abundance
rteNtN
rNdt
dN
0
Where N0 is the initial population size
Nt is the population size at time t
r is the per capita growth rate

Different Names for r
Instantaneous rate of increase Intrinsic rate of increase Malthusian parameter Per capita rate of population
increase over short time interval
r = b – d
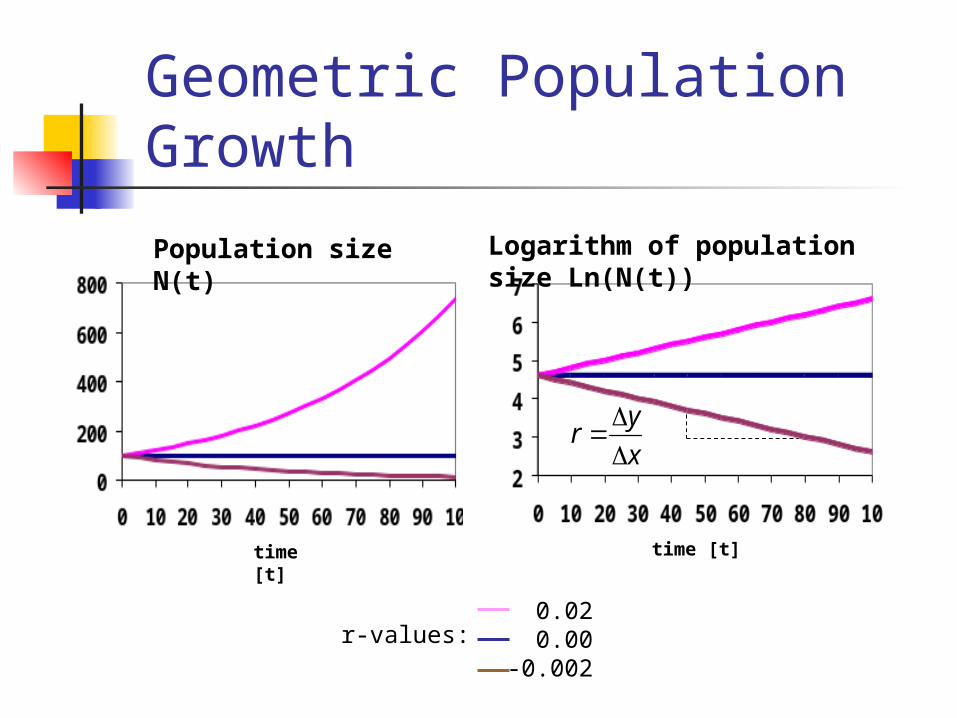
Geometric Population Growth
time [t]
Population size N(t)
0.020.00
-0.002
r-values:
time [t]
Logarithm of population size Ln(N(t))
yr
x

Discrete vs continuous growth

Discrete vs continuous growth
Discrete growth across a time interval; birth and death discrete
realistic; hard to generalize results
Continuous instantaneous
growth;
birth and death continuous process
abstraction; easier to generalize
results
Ln() = r
= er
r > 0 > 1r = 0 = 1r < 0 0 < < 1

Model Assumptions

Model Assumptions Closed population Constant b and d Population grows exponentially indefinitely No genetic structure No age or size structure Species exist as single panmictic population
unstructured (random-mating) populations

Pheasants of Protection Island(Data from Lack 1967)
1
10
100
1,000
10,000
1937 1938 1939 1940 1941 1942 1943Year
Po
pu
lati
on
siz
e (
N)
1937 8 birds1938 30 birds
R = 30/8 = 3.75r = ln(3.75) = 1.3217
0
2,000
4,000
6,000
8,000
10,000
12,000
1937 1938 1939 1940 1941 1942 1943Year
Po
pu
lati
on
siz
e (
N)

Density dependence Population growth is affected
by limited resources food breeding sites territories
Density dependent factors influence birth and death rates b and d are not constant
Logistic population growth
Population size (N)
Population size (N)
b
b
Allee effect

Effects of crowding: decreased reproduction
Effect of density on fecundity in the Great Tit Parus major (Lack 1966)
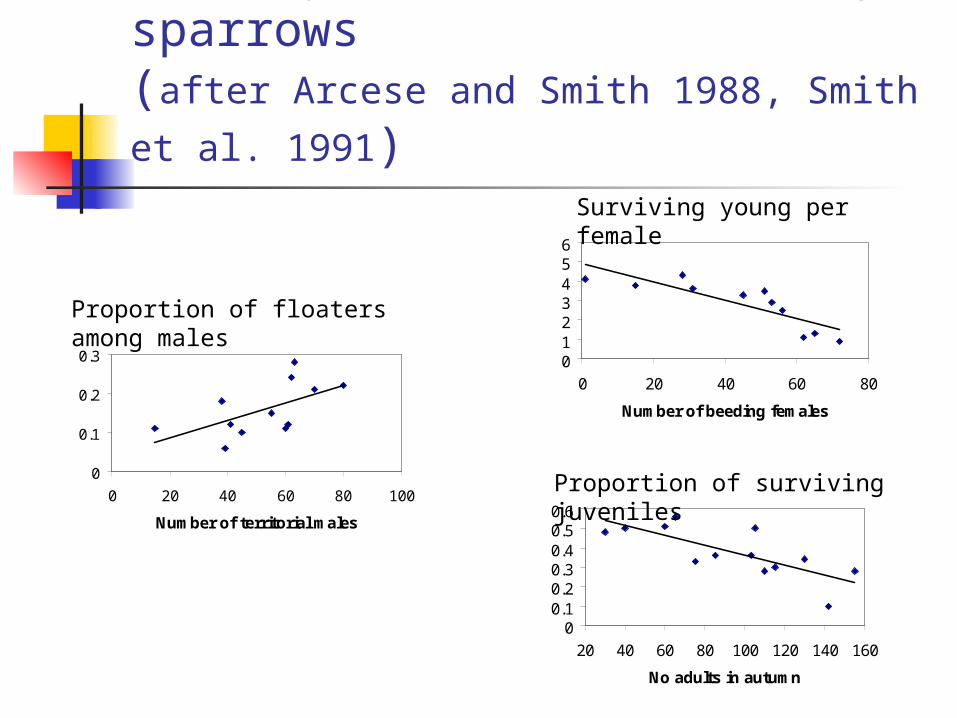
Density dependence in song sparrows (after Arcese and Smith 1988, Smith et al.
1991)
0123456
0 20 40 60 80
Number of beeding females
Surviving young per female
0
0.1
0.2
0.3
0 20 40 60 80 100
Number of territorial males
Proportion of floaters among males
00.10.20.30.40.50.6
20 40 60 80 100 120 140 160
No adults in autumn
Proportion of surviving juveniles

Density Dependence in Death Rate
Increased crowding increased death rate
d = d0 + c N
d0 : death rate under uncrowded conditions
c : measures strength of density dependence
N : current population size

Density Dependence in Birth Rate
Increased crowding decreased birth rate
b = b0 – a N
b0 : birth rate under uncrowded conditions
a : measures strength of density dependence
N : current population size
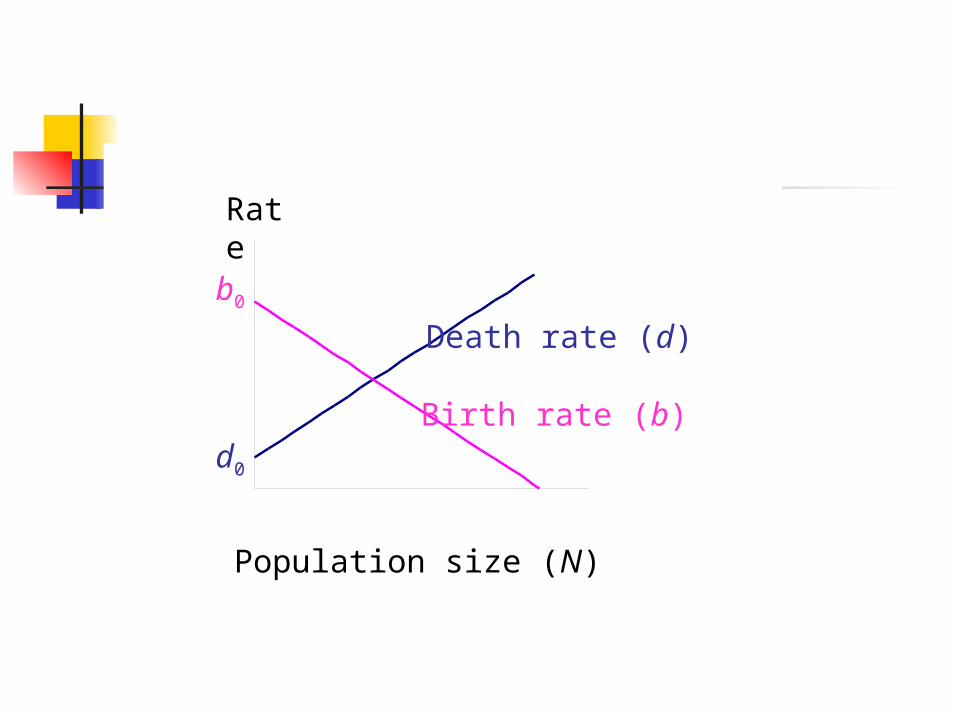
Population size (N)
Rate
b0
d0
Death rate (d)
Birth rate (b)

Interpretation of Carrying Capacity K Defined as the level of abundance above
which the population tends to decline It is population specific: depends on
available resources and space (quality and quantity), abundance of predators and competitors
It is an abstraction: crude summary of interactions of a population with environment
Units are in number of individuals supported by environment

Stable Equilibrium
N = K b = dN > K
population declines towards K
N < K population
increasestowards K
Population size (N)
Rate
b0
d0
K
Death rate (d)
Birth rate (b)

Carrying capacity, K
1dN N
rNdt K
Unused proportion of carrying capacity
14% of area is white or “unused”

K=100
1 – N/K Nt+1
N = 7 1 - 7/100= 0.93
rN (0.93) population growths fast
unused proportion of carrying capacity

K=100
1 – N/K Nt+1
N = 7 1 - 7/100= 0.93
rN (0.93) population growths fast
N = 98
1 - 98/100= 0.02
rN (0.02) population growths slowly
unused proportion of carrying capacity

K=100
1 – Nt/K Nt+1
N = 7 1 - 7/100= 0.93
rN (0.93) population growths fast
N = 98
1 - 98/100= 0.02
rN (0.02) population growths slowly
N = 105
1 - 105/100= -0.05
rN (-0.05) population declines
unused proportion of carrying capacity

Logistic Growth Curve
0
20
40
60
80
100
120
140
160
180
200
0 20 40 60 80 100
Time (t)
Popu
latio
n si
ze (N
)
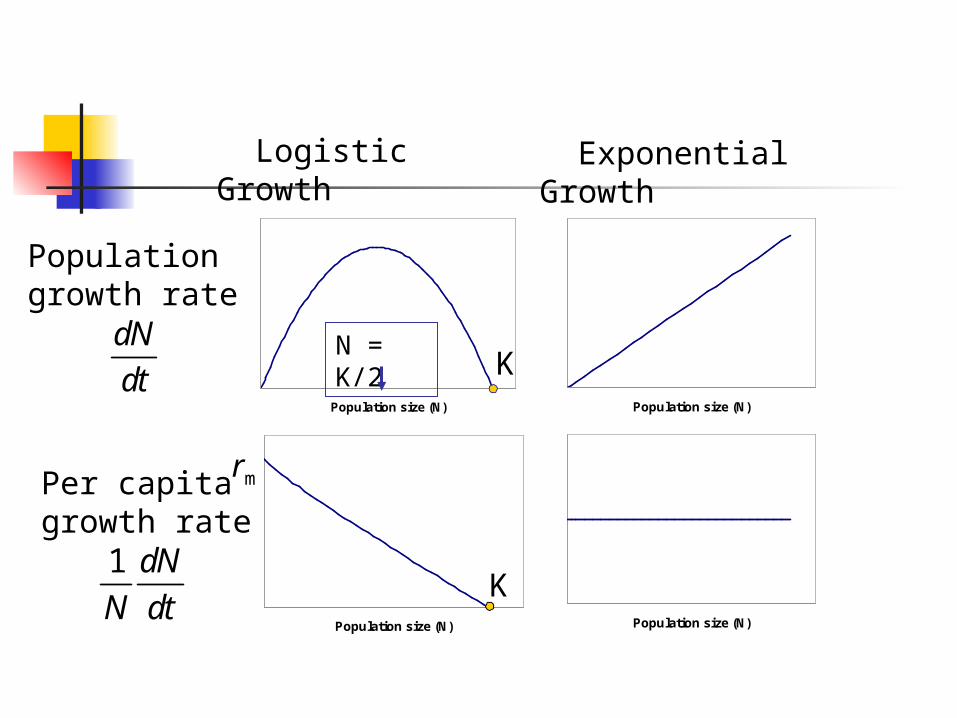
Population size (N)
Population growth rate
Logistic Growth
KdN
dtN = K/2
Population size (N)
Exponential Growth
Population size (N)
Per capita growth rate
K1 dN
N dt
rm
Population size (N)

Time Lag (t) Sometimes individuals do not
immediately adjust their growth and reproduction when resources change, and these delays can affect population dynamics
Causes Seasonal availability of resources Growth responses of prey populations Age and size structure of consumer
populations
1 tdN N
rNdt K

Time Lag Delay differential equation Important parameters:
Length of time lag, “Response time” = 1/r
r controls population growths 0 < r < 0.368
population increases smoothly to K 0.368 < r < 1.570
damped oscillations r > 1.570
stable limit cycle

Time (t)
Po
pu
lati
on
siz
e (
N)
“medium” r damped oscillations
Time (t)
Po
pu
lati
on
siz
e (
N)
“small” r
Time (t)
Po
pu
lati
on
siz
e (
N)
Period ( 4
Amplitude(increases with r)
“large” r stable limit cycle

Discrete time density-dependent models Discrete population growth model has
built-in time lag of length 1.0 complex behavior
Different ways to incorporate density dependence
1.
2.
3. Ricker model:
1 1 tt t t
NN N rN
K
1
1
tNr
Kt tN N e
1 1t t tN N N
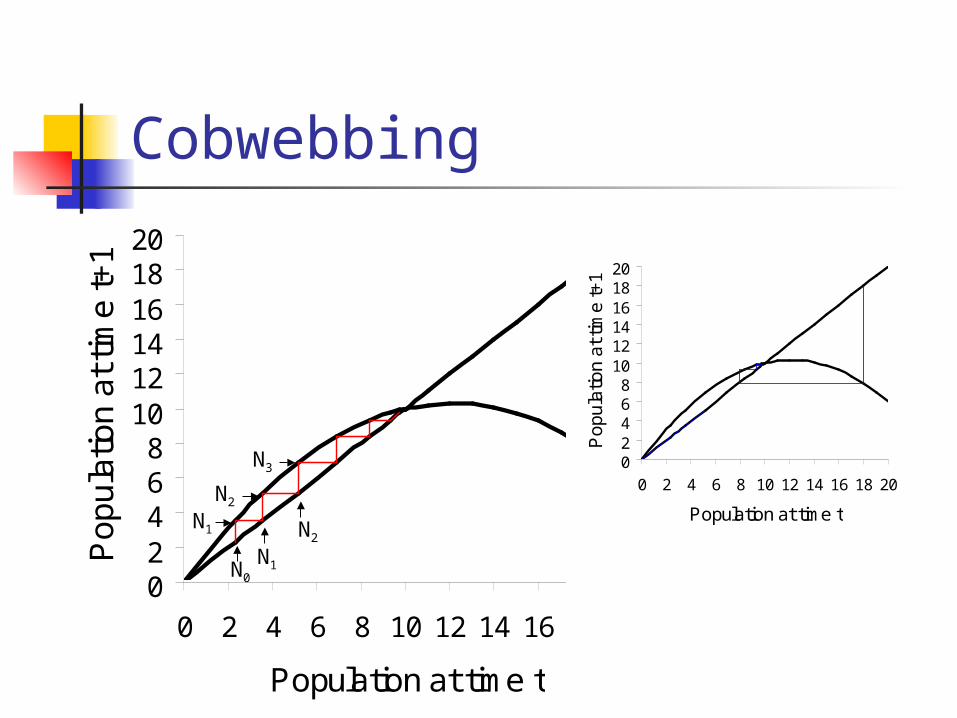
Cobwebbing
02468
101214161820
0 2 4 6 8 10 12 14 16 18 20
Population at time t
Po
pu
latio
n a
t tim
e t+
1
02468
101214161820
0 2 4 6 8 10 12 14 16 18 20
Population at time t
Po
pu
latio
n a
t tim
e t+
1N0
N1
N1 N2
N2
N3

Discrete time density-dependent models
In (1) and (3) dynamics depend solely on r
r < 2.0 population approaches K
2.0 < r < 2.449 2-point limit cycle2.449 < r < 2.57 4-point limit cycler > 2.57 chaos
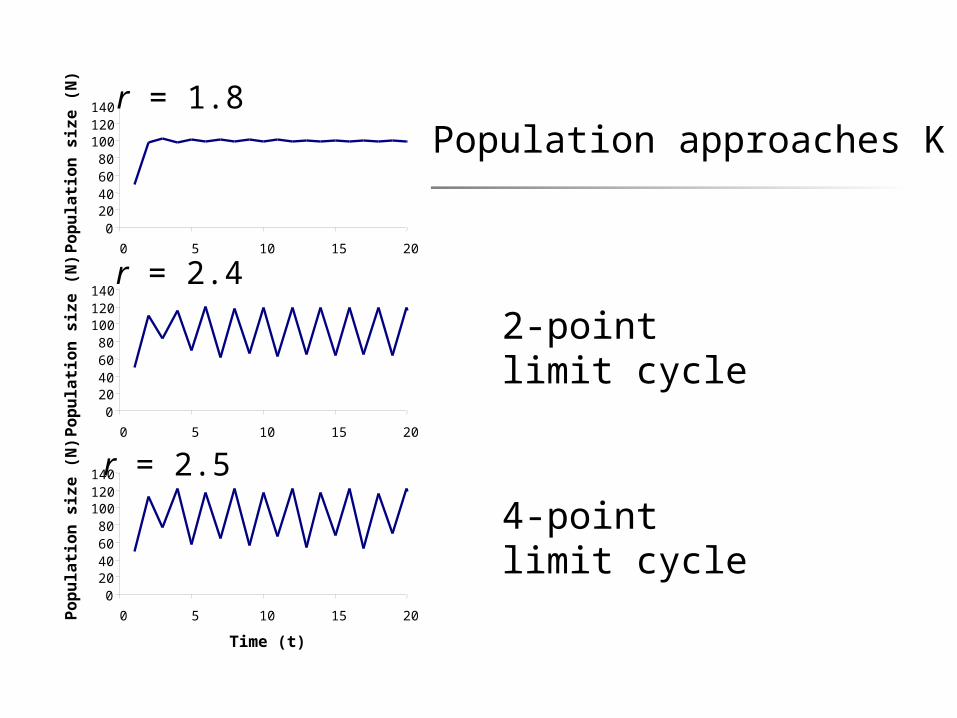
020406080
100120140
0 5 10 15 20
Po
pu
lati
on
siz
e (
N) r = 1.8
Population approaches K
020406080
100120140
0 5 10 15 20
r = 2.4
Po
pu
lati
on
siz
e (
N)
2-point limit cycle
020406080
100120140
0 5 10 15 20
Time (t)
r = 2.5
Po
pu
lati
on
siz
e (
N)
4-point limit cycle

r=1.8
N(t)
N(t
+1)
0.0 0.2 0.4 0.6 0.8 1.0
0.0
0.2
0.4
0.6
0.8
1.0
N(t)
N(t
+1)
N0
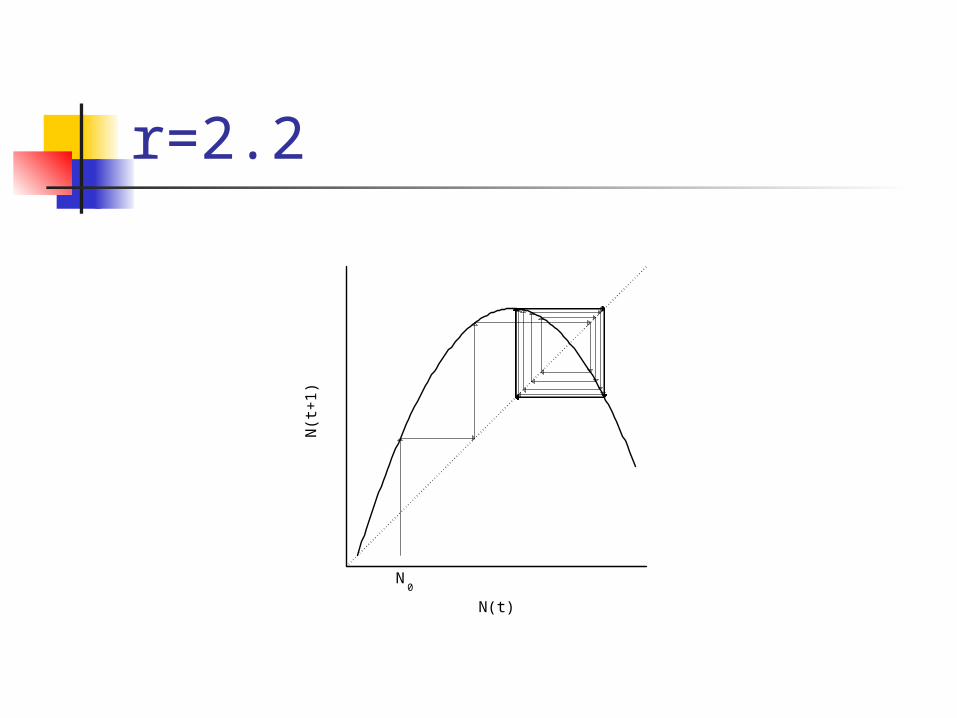
r=2.2
N(t)
N(t
+1)
N0

Chaotic behaviorr = 2.8
020406080
100120140
0 5 10 15 20
Time (t)
Po
pu
lati
on
siz
e (
N)
N0 = 50
N0 = 51

Predicting the future
0
100
200
300
400
0 50 100 150 200
Nt
Nt+1
Ricker model with K = 100 and r = 3.5
0
100
200
300
400
0 50 100 150 200
Nt
Nt+10

Model assumptions

Model assumptions
Increased density causes a (linear) decline in population growth rate
No variability in carrying capacity All other assumptions same as for
exponential growth (assume single, panmictic population, no age/stage structure)

Discrete Logistic Substitute b and d in
Nt+1 = RNt = (1+b - d) Nt
Nt+1 = Nt +[b0-aNt - d0-cNt]Nt
Rearrange and replace K = b0-d0 / (a+c)
1 1 tt t t
NN N rN
K





























