Embed Size (px)
Citation preview

Molecular and Cellular Pathobiology
Role of Megakaryocytes in Breast CancerMetastasis to BoneWalter Jackson III1, Donna M. Sosnoski1, Sara E. Ohanessian2, Paige Chandler1,Adam Mobley1, Kacey D. Meisel1, and Andrea M. Mastro1
Abstract
Little is known about how megakaryocytes may affect metas-tasis beyond serving as a source of platelets. In this study, weexplored the functional implications of megakaryocyte accumu-lation in the femurs of mice after injection of metastatic or non-metastatic breast cancer cells in 4T1.2 BALB/cJ and MDA-MB-231nude mouse models. At bone metastatic sites, but not primarygrowth sites, tumor growth was associated with increased mega-karyopoiesis in both model systems. In the orthotopic BALB/cJmodel, extramedullary hematopoiesis occurred in the spleen,resulting in a four-fold increase in megakaryocytes. In supportof the hypothesis that reducing megakaryocytes may reduce
metastasis, we found that thrombopoietin-deficient mice exhib-ited a 90% relative decrease in megakaryocytes, yet they devel-oped more aggressive metastasis than wild-type hosts. In humanclinical specimens, we observed an increase in megakaryocytes inthe bone marrow of 6/8 patients with metastatic breast cancercompared with age- and gender-matched controls. Taken togeth-er, our results suggested that an increase in megakaryocytesoccurring in response tometastatic cells entering thebonemarrowconfers some measure of protection against metastasis, challeng-ing present views on the role of megakaryocytes in this setting.Cancer Res; 77(8); 1942–54. �2017 AACR.
IntroductionBone is a complex and sophisticated organ. Resident marrow
cells differentiate either from hematopoietic (HSC) or mesen-chymal stem cells (MSC). Chondrocytes, osteoblasts, fibro-blasts, adipocytes, endothelial cells, and myocytes originatefrom MSCs (1). Megakaryocytes differentiate from HSCs in theendosteal niche under the influence of thrombopoietin (TPO)secreted by osteoblasts (2). Mature megakaryocytes are direct-ed to the vascular niche by stromal-derived factor-1 (SDF-1)where they produce and release platelets into the bloodstream.In addition, megakaryocytes play a role in bone metabolism,for example, megakaryocytes inhibit osteoclast function andenhance osteoblast proliferation (3). Bone metastasis is theleading cause of death from breast cancer. Both osteoblasts andosteoclasts are fundamental in the process of metastasesgrowth in the bone. Because thrombocytosis is often seen inmany types of advanced cancer, megakaryocytes may assume amuch larger role in cancer progression and metastasis thanpreviously known.
"Megakaryocytes join the war on cancer" is the title of acommentary (4) regarding a novel mechanism of host sup-
pression of tumorigenesis via an increase in platelets resultingfrom an increase in megakaryocytes in the bone marrow oftumor-bearing mice (5). In that study, Lewis lung tumor cellsinoculated in the flanks of mice led to an increase in mega-karyocytes in the bone marrow and an increase in circulatingplatelets. The authors concluded that platelet-derived throm-bospondin-1 (TSP-1) plays a critical role in suppressing tumorangiogenesis (5). In a different study, Li and colleagues suggestthat megakaryocytes are the first line of defense against prostatecancer metastasis to bone, noting a reduction in bone metas-tasis of PC-3 cells in mice treated with megakaryocyte growthand development factor, TPO, prior to tumor inoculation (6).In contrast, others have suggested that megakaryocytes andplatelets act as "primers" of the metastatic niche encouragingcancer colonization (7, 8). Thus, when megakaryocytes join thewar, whose side are they on?
While carrying out immunohistochemistry of bone (9), wedetected increased numbers of megakaryocytes in the marrow ofathymic mice bearing metastatic breast cancer (MDA-MB-231).This observation led us to question whether the megakaryocytesincreased in numbers prior to the arrival of the cancer cells tocreate a hospitable niche or if the megakaryocyte increased inresponse to cancer cells in the bone? Using a mouse xenograftmetastatic model, we found that the increase in megakaryocyteswas seen after metastases were evident. However, in a model inwhich themammary gland tumor didnotmetastasize, no increasein the megakaryocytes was observed. In a syngeneic immuno-competent mouse mammary carcinoma metastasis model, 4T1.2(10), an increase in megakaryocytes was not found in the bonemarrow, but rather in the spleen. In Tpo knockout mice, mega-karyocytes and platelet counts were reduced by 90% comparedwith normal mice. When injected with 4T1.2 cancer cells, theTpo�/� mice developed metastases more rapidly than either wild-type or heterozygous cohorts.
1Department of Biochemistry and Molecular Biology, The Pennsylvania StateUniversity, University Park, Pennsylvania. 2Pathology and Laboratory Medicine,Penn State Hershey Medical Center, Hershey, Pennsylvania.
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
Corresponding Author: Andrea M. Mastro, Pennsylvania State University, 428 SFrear, University Park, PA 16802. Phone: 814-863-0152; Fax: 814-863-0724;E-mail: [email protected]
doi: 10.1158/0008-5472.CAN-16-1084
�2017 American Association for Cancer Research.
CancerResearch
Cancer Res; 77(8) April 15, 20171942
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084

In order to determine if an increase in megakaryocytesoccurred in human metastatic cancer patients, megakaryocyteswere counted in bone marrow autopsy samples from patientswho died from metastatic breast cancer and from age- andgender-matched subjects whose death was noncancer related.Increased numbers of megakaryocytes were found in six out ofeight of the cancer patient samples compared with matchedcontrols.
Taken together, the data suggest that megakaryocytes increaseas part of a protective response to metastatic cancer. The loss ofmegakaryocytes permits a more aggressive metastatic response.
Materials and MethodsCell culture
MDA-MB-231-1833-luc (11) is an aggressive bone seekingclone of the MDA-MB-231 human metastatic breast cancer cellline. The cells, a gift, 2010, from Dr. Joan Massagu�e (MemorialSloan-Kettering Cancer Center, New York, NY) were grown inDMEM (Corning Cellgro), 10% FBS (PAA Laboratories), and 100U/mL penicillin/100 mg/mL streptomycin (Corning Cellgro). Themouse mammary carcinoma lines 4T1.2, (bone metastatic) and67NR(nonmetastatic; ref. 10)were a gift fromDr.RobinAnderson(Peter MacCallum Cancer Centre, Melbourne, Victoria, Australia)in 2008. The nonimmunogenic luciferase tagged 4T1.2 cells (12)were obtained (2009) from Dr. Shoukat Dedhar (British Colum-bia Cancer Research Centre, Vancouver, British Columbia,Canada), and maintained in DMEM, 10% FBS, 1% nonessentialamino acids (Corning Cellgro) penicillin/streptomycin, and5 mg/mL puromycin; 67NR cells were maintained in DMEM,10% FBS, 1% NEAA, and penicillin/streptomycin. MEG-01, ahuman megakaryoblastic line, from Dr. Melissa Kacena (IndianaUniversity, Bloomington, IN) in 2009, were maintained inRPMI1640 supplemented with 10% FBS and penicillin/strepto-mycin. Murine preosteoblasts, MC3T3-E1, obtained in 2004 fromDr. Norman Karin (Pacific Northwest National Laboratory, Rich-land, WA) were grown as previously described (13). Primary,authenticated human osteoblasts, NHOst, were purchased fromLonza and grown as recommended. All cell lines were tested andwere free of mycoplasma. The MDA-MB-231 and MEG-01 cellsbothwere authenticatedby STRprofiling (Genetica)most recentlyin 2016.Nouniversal standards are available for authentication ofthe murine cell lines (14).
Mouse experimentsAll animal experiments were approved by the Penn State
University IACUC, protocol 40832. Animals were anesthetizedby inhalation of isoflurane and oxygen. For the xenograft model,female athymic mice, obtained from Charles River Laboratories,Frederick, MD, were inoculated with MDA-MB-231-1833 cellspassaged five times without antibiotics. For the experimentalmetastatic model, 1 � 105 cells were suspended in 100 mL ofsterile PBS and injected into the left cardiac ventricle (15, 16) of 8-week-oldmice; for thenonmetastaticmodel, 1�106 cells in 25mLof PBS were inoculated into the fourth mammary gland. Inocu-lation of PBS alone served as a control. The mice receivingintracardiac injections (six per group) were sacrificed 1, 4, 10,20, and 30 days postinjection. Mice receiving mammary glandinjections (six per group) were sacrificed 4, 14, 24, and 34 dayspostinjection. Upon sacrifice, blood was collected by cardiacpuncture; femurs were removed, fixed in 4% paraformaldehyde,
and decalcified in 500 mmol/L EDTA. Bones were paraffin-embedded using standard histologic protocols.
For the syngeneic immunocompetent model, mouse mamma-ry tumor cells, 4T1.2 and 67NR, were injected, 5� 104 in 25 mL ofsterile PBS, into the 4th mammary gland of 13-week-old femaleBALB/cJ (Jackson Laboratory, Bar Harbor, ME). Sacrifice occurredon days 7, 14, 32, and 40 days postinoculation.
Mice were weighed before cancer cell inoculation and againat sacrifice. Tumor growth was assessed morphometricallyusing an electronic caliper, and tumor volumes were calculatedaccording to the formula V (mm3) ¼ L (major axis) � W2
(minor axis) / 2 (17). On termination, blood, femurs andspleens were collected. Femurs and spleens were fixed andparaffin-embedded.
Image analysisThe MDA-MB-231-1833-luc and 4T1.2-luc cells were engi-
neered to express luciferase for real-time visualization with anIVIS50 Imaging System (Xenogen). The mice were injected with10 uL/g of body weight of luciferin (15 mg/mL in sterile PBS)10–15 minutes prior to imaging. Live animal imaging was per-formed weekly. Total tumor burden was determined by calculat-ing the total amountof photonflux from the luciferase tagged cellswithin the entiremouse. Living Image(R), version 2.60.1/Igor Pro4.09A software was used to analyze the IVIS images and tocalculate the tumor burden.
IHCSections of formalin-fixed, decalcified, and paraffin-embedded
femurs (10 mm) and fixed and paraffin-embedded spleens (5 mm)were mounted on glass slides, treated with 0.5% trypsin for 15minutes at 37�C, and stained overnight at 4�C with a primaryrabbit polyclonal antimouse von Willebrand (vWF; Abcam)1:500 in TBS, 1% donkey serum. Endogenous peroxidase wasblocked with 3% hydrogen peroxide for 45 minutes at roomtemperature and the sections were further stained with secondarybiotinylated donkey anti-rabbit (Abcam) 1:1,000 in TBS, 1%donkey serum for 2 hours at room temperature. Immunostainingwas completed using Vectastain Elite ABC reagent and ImmPACT3,3'-Diaminobenzidine Kit (Vector Laboratories, Burlingame,CA), and a hematoxylin counterstain. vWF-positive megakaryo-cytes were counted in 30 fields, �400 magnification per eachfemur and spleen with an 20� light microscope.
Human bone marrow samples were obtained from anautopsy set (Penn State Hershey Medical Center) that includedeight subjects who died with metastatic breast cancer, and age-and gender-matched individuals who died of noncancer-relat-ed causes. Formalin fixed paraffin-embedded sections werestained for vWF using a polyclonal rabbit anti-human vWF(Dako) diluted 1:800 in TBS, 1% donkey serum as the primaryantibody. A Keyence BZ-X700 microscope (Keyence Corp.) wasemployed to image, scan, and quantify the area of eachsample. Megakaryocytes were counted and reported as mega-karyocytes/mm2.
Blood collection and analysisBlood was obtained by cardiac puncture from anesthetized
mice. Whole blood (100–500 mL) was collected in EDTA forcomplete blood counts; the remaining blood was clotted at roomtemperature for 2 to 3 hours, centrifuged at 400 � g, 10 minutes,and the serum fraction collected and stored at �80�C.
Megakaryocytes and Bone Metastasis
www.aacrjournals.org Cancer Res; 77(8) April 15, 2017 1943
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084

Complete blood cell counts were performed on a Hemavet950S (Drew Scientific). For some experiments, platelet countswere performed by flow cytometry following the method ofAlugupalli and colleagues (18). Briefly, whole blood samplescollected in sodium citrate were fluorescently labeled for theplatelet markers CD 41 and CD 61. Counts were performed usingan Amnis FlowSight Imaging flow cytometer.
SDF-1 and TPO serum levels were determined by ELISA assays(R&D Systems) following the manufacturer's protocol.
BALB/cJ TPO knockout miceC57Bl/6 Tpo knockout mice were created and characterized
by de Sauvage and colleagues (19). Frozen embryos were a giftfrom Genentech. The embryos were re-derived by The JacksonLaboratory and backcrossed with BALB/cJ mice for 10 genera-tions to obtain Tpo heterozygous mice with a BALB/cJ geneticbackground of >98%. A breeding colony was established atPenn State University to generate Tpo�/� female mice. Geno-type determinations were made from tail snips of 3-week-oldpups (Supplementary Fig. S1).
In vitro megakaryocyte assaysMEG-01 cells were tested for proliferation, adherence, and
ploidy in the presence of conditioned media from cancer cellsand osteoblasts. Conditioned media were prepared by replacingthe cancer cell, MDA-MB-231 or 4T1.2, or osteoblast, murineMC3T3-E1 or human NHOst differentiation medium with serumfree RPMI1640, and incubating at 37�C for 24 hours. The mediawere collected and centrifuged (300� g, 10minutes) and stored at�80�C until used. Conditioned media were prepared from nearlyconfluent cultures of cancer cells, differentiated osteoblasts, orfrom osteoblasts incubated for 24 hours with 50% conditionedmedium from cancer cells. These conditioned media containedsecretions from the osteoblasts in response to the cancer cellsecretions, and are referred to as "double conditioned medium."As described in the figure legends, proliferation was based on anincrease in viable cell numbers (MTS Promega, CellTiter96 Aque-ous One). Adherence of MEG-01 was carried out as describedpreviously (20). The degree of ploidy was measured by flowcytometry following propidium iodide staining. Megakaryocyteprogenitors in primarymouse bonemarrowwere quantified usingtheMegaCult–Cassay systemas directed (StemCell Technologies).
Statistical analysisThe number of animals per group was determined by previous
experiments and discussionwith a statistician. Data analyses wereperformedwithGraphpad Prism software. One-way and two-wayANOVAwith Bonferroni or Tukeymultiple comparison tests wereused to compare platelet counts, red blood cell (RBC), whiteblood cell (WBC), WBC populations, megakaryocyte counts, andserum cytokine concentrations. Student t test was utilized toanalyze flow cytometry platelet counts and human bone marrowmegakaryocyte data.
ResultsAn increase in megakaryocytes followed the appearance ofmetastasis of cancer cells in the bone marrow of athymic mice
Wenoted an increase inmegakaryocytes in the femurs of tumor-bearingmice, approximately 1monthafter intracardiac inoculationof MDA-MB-231 cells. Using mouse models, we asked when the
increase in megakaryocytes occurred during the metastatic process.An increase in megakaryocytes prior to colonization of metastaseswould support the theory that megakaryocytes prime the premeta-static niche. However, an increase in megakaryocytes after theappearance of metastases would suggest that the cancer cells inthe bonemarrowaffectmegakaryopoiesis. In addition, comparisonof a metastatic model (intracardiac injection) to a nonmetastatic(mammary gland injection) would indicate whether primarytumors systemically elicited an increase in megakaryocytes.
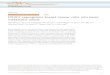
Mice were injected intracardially with 1833-luc or with PBS(control), and sacrificed, 1, 4, 10, 20, and30days postinoculation.Additionally, another group of mice were inoculated in themammary gland, and sacrificed 4, 14, 24, and 34 days postinoc-ulation. IVIS imaging was performed weekly. Metastases weredetected between 4 and 10 days in mice receiving intracardiacinjections (Fig. 1A). Mice injected orthotopically showed no signsof metastasis over the course of the study (Fig. 1E). The mega-karyocytes in both femurs of each mouse were visualized byimmunohistochemical staining of bone sections for von Will-ebrand factor (Fig. 1B). In the metastatic group, femurs of micefrom the 30-day cohort contained about twice the number ofmegakaryocytes compared with other cohorts (Fig. 1C). The micesacrificed on day 30 had on average 13.0 (� 2.5 SD) megakar-yocytes/field compared with an average of 7.0 (� 1SD) mega-karyocytes/field in all other groups. The number of megakaryo-cytes in the bonemarrow increasedwith time, and correlatedwiththe whole body tumor burden (Fig. 1D). In the nonmetastaticmodel, there was no difference in megakaryocyte numbers in theanimals with or without tumors (Fig. 1F). Although there was anonsignificant increase in megakaryocyte numbers in the femursof PBS and tumor injected mice over time, it may be attributed tothe aging of the mice.
Platelet counts, performed with an automated counter, indi-cated variation from mouse to mouse (Fig. 1G). However, onaverage, the mice injected with 1833 intracardially showed anincrease in platelets on day 30, the same time that the megakar-yocytes were elevated (Fig. 1G). The mice with mammary glandinjections did not have a significantly elevated platelet count (Fig.1G). These data suggested that tumor cells, once they colonizedthe marrow, elicited an increase in megakaryocytes. The increasedid not appear to be due to a systemic factor released from theprimary tumor but rather a response to the presence of the cancercells in themarrow. Nonetheless, it cannot be ruled out that if theprimary tumor had been permitted to grow for a longer time, itmight eventually have led to increased megakaryocyte numbers.
BALB/cJ mice injected with 4T1.2 showed an increase insplenic megakaryocytes
In order to determine if the increase in megakaryocytesoccurred in mice with an intact immune system, we used asyngeneicmodel of immunocompetent BALB/cJmice andmurinemammary tumor cells. The megakaryocyte response to nonmeta-static 67NR cells (primary tumors) was compared with that ofmice inoculatedwith4T1.2 cells (primary andmetastatic tumors).Over the course of 6 weeks, six mice per group including a PBSgroupwere sacrificed at 7, 14, 32, and 40 days postinoculation. Tomonitormetastasis, IVIS imagingwas performedweekly (Fig. 2A).Mice were weighed before inoculation and again before sacrifice;tumors were measured immediately prior to sacrifice. In allgroups, body weights increased over the course of the study. Onday 32, 4T1.2 tumors averaged 154.05 (�43.84 SD) mm3
Jackson et al.
Cancer Res; 77(8) April 15, 2017 Cancer Research1944
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084

1 4 10 20 30Days
15
10
5
01 4 10 20 30
MK
s/Fi
eld
MK
s/Fi
eld
Days
PBS1833
IC
Photon flux
20
15
10
5
01×106 1×107 1×108 1×109 1×1010
4 14 24 34Days
PBS1833
MK
s/Fi
eld
10.0
7.5
5.0
2.5
0.04 14 24 34
Days
MG
A
C
E
G
F
D
BP
late
lets
(×10
3 /µL
) 1,500
1,250
1,000
750
500
250
0
Days Days Days Days1 4 10 20 30 1 4 10 20 30 4 14 24 34 4 14 24 34
IC PBS IC 1833 MG PBS MG 1833
10
20
30
Figure 1.
Increase in megakaryocyte numbers in the bone marrow of mice with metastatic but not nonmetastatic breast cancer in a xenograft model. Six athymic mice perendpoint were inoculated into the heart (intracardiac) or into the mammary gland (MG) with MDA-MB-231-LUC (clone 1833) or with PBS. A, Representative IVISimages of nudemice inoculated intracardially with 1833-LUC. At 1, 4, 10, 20, and 30 days following intracardiac inoculation, the mice were sacrificed, and the left andright femurs processed as described in Materials and Methods and stained for von Willebrand factor (vWF). Megakaryocytes (MK) were counted using lightmicroscopy (�400). B, Representative paraffin section stained for vWF to identify megakaryocytes (arrow). Scale bar, 20 mm C,Megakaryocytes were counted in atotal of two femurs permouse inoculated intracardially (IC). Shown is a graph of the average number of megakaryocytes per field (30 fields at�400magnification).Themice sacrificedat day30hadonaverage 13.0� 2.5 SDmegakaryocytes/field comparedwith anaverageof 7.0� 1.0 SDmegakaryocytes/field formice sacrificed 1day postinoculation. Metastatic cancer caused a significant increase (��� , P < 0.001) in the number of megakaryocytes in the femur at 30 days postinoculation.D, Tumor burden from 1833 inoculation measured from the IVIS images correlated with megakaryocyte numbers per field (Pearson correlation theory). Asthe tumor burden increased, the number of megakaryocytes in the bone marrow of the femurs also increased. Each square represents a cohort of mice for days 10,20, and 30. Each dot represents an individualmouse (n¼ 5 or 6 per time). Samples are grouped by time of sacrifice following tumor cell inoculation.E,RepresentativeIVIS images of nude mice inoculated in the 4th mammary gland with 1833-LUC. At 4, 14, 24, and 34 days following inoculation, mice were sacrificed andfemurs treated as inA. F,Megakaryocyteswere counted in a total of 12 femurs, two permouse, as described inC. Shown are the averagemegakaryocytes/field� SD.Therewas no significant difference between the tumor-bearingmice and those inoculatedwith PBS at any of the times tested.G, Platelet counts from blood samplesof all cohorts were analyzed by a HEMAVET 950FS. Shown are the counts per sample and the mean for each time. One-way ANOVA with repeated measureswas used to analyze each intervention across time and when appropriate a Fisher LSD post hoc test was used to compare pairwise differences. Independentt tests with alpha level correctionswere used to compare the different treatment groups at corresponding time points. Only themice with intracardiac inoculation of1833 showed a significant (#, P < 0.05) increase in platelets with time.
Megakaryocytes and Bone Metastasis
www.aacrjournals.org Cancer Res; 77(8) April 15, 2017 1945
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084

comparedwith 67NR tumors that averaged 649.08 (�165.15 SD)mm3. By day 40, 4T1.2 tumors averaged 211.81 (�23.82 SD)mm3whereas 67NR tumors averaged 833.58 (�177.72 SD)mm3.Although the 67NR tumors grew to nearly four times the volumeof the 4T1.2, they didnotmetastasize, and therewas no increase inmegakaryocytes.
Because of high levels of G-CSF secreted by 4T1.2 cells, micewith 4T1.2 tumors exhibited splenomegaly (Fig. 2B) and extra-medullary hematopoiesis (17). The spleens of mice injected with4T1.2 cells weighed 0.60 g (�0.5 SD) andmeasured 2.6 cm (�0.4SD) in length. By comparison, spleen weights and lengths forPBS and 67NR were (0.09� 0.01 g, 1.63� 0.05 cm) and (0.09�0.01 g, 1.38 � 0.08 cm), respectively.
There was no significant increase in megakaryocyte numbersin the marrow of tumor-bearing mice (4T1.2 or 67NR) com-pared with those inoculated with PBS at any time (Fig. 2C). Theaverage number of megakaryocytes observed across all cohortswas 8.3 (�1.2 SD) megakaryocytes/field. However, there werefewer megakaryocytes in the 4T1.2 injected mice at day 32 (Fig.2C). This decrease might reflect the large increase in extrame-dullary megakaryopoiesis in the spleen. In contrast, there was asignificant increase in megakaryocytes (Fig. 2D) in the spleensof the 4T1.2 tumor-bearing mice compared with the PBS- and67NR-inoculated mice. The average numbers of megakaryo-cytes/field �SD in the spleens of the 14 day, cancer cellinoculated cohort was 3.5 � 1.2 compared with 1.2 � 0.5 inthe PBS cohort, and 0.93 � 0.42 in the 67NR cohort. In thespleens of the 32 days, 4T1.2 cell inoculated cohort the averagewas 6.7 � 1.2 compared with 1.6 � 0.4 in the PBS-injected and1.15 � 0.65 in the 67NR inoculated mice. By 40 days aftercancer inoculation, the average number of megakaryocytes/field in the spleens of the 4T1.2 mice was 5.1 � 1.6 comparedwith 3.0 � 1 megakaryocytes/field in the PBS controls and 2.31� 0.5 in the 67NR-inoculated mice.
There alsowas a significant increase in platelet counts over timein mice injected with 4T1.2 but not with 67NR or PBS (Fig. 2E)reflecting the increase inmegakaryocytes. The increase in plateletsin the 4T1.2 inoculated mice was confirmed by a flow cytometricmeasure of CD41þ, CD61þmarkers. At 40 days postinjection, the4T1.2-bearing mice had on average 1,196 � 182.3 (SD) platelets� 103/mL; PBS-injected mice had 384.3 � 27.0 (SD) platelets �103/mL (P < 0.01).
Analysis of the complete blood counts indicated severalchanges (Fig. 3). The RBCs were significantly decreased in themice inoculated with 4T1.2 compared with the PBS-injectedmice. In contrast, the mice with 67NR had increased RBCcounts early on. The decrease with 4T1.2 erythropoiesis wasrecently reported in another study (21). The 4T1.2 cells alsoinduced a leukemoid reaction (Fig. 3B), resulting in a markedincrease in total WBC populations in the 4T1.2 mice comparedwith control and 67NR inoculated mice (Fig. 3B; ref. 17).There was also a corresponding increase in individual WBCpopulations (Fig. 3C). The populations of WBCs shifted sig-nificantly from leukocytes to granulocytes (mostly neutro-phils) in both 4T1.2 and 67NR inoculated mice comparedwith PBS-treated mice.
Both TPO and SDF-1 are important cytokines for megakaryo-poiesis. Decreases in serum TPO and SDF-1 concentrations werenoted at 32 and 40 days postinoculation for 4T1.2mice (Fig. 3D).The 67NR, on the other hand, expressed increased levels. Thus,lower serum TPO levels were associated with metastatic cancer.
SDF-1 levels dropped in both the 4T1.2 and 67NRmice (Fig. 3E).The decrease in SDF-1 may be inversely correlated with theincrease seen in G-CSF concentrations (22).
Tpo�/� mice developed metastasis more rapidlythan Tpoþ/þ and Tpoþ/� mice
In order to further investigate a role for megakaryocytes incancer metastasis, we utilized Tpo knockout mice, which producereduced numbers of megakaryocytes (19). Phenotypically, theBALBc/J Tpo knockout strain presented negligible levels of TPO,>90% reduction in megakaryocytes in the bone marrow andspleen, and a nearly 10-fold reduction in platelets compared withwild-typemice. Heterozygousmice showed intermediate levels ofthese four phenotypic parameters (Table 1).
Tpoþ/þ (wild-type), Tpoþ/� (heterozygotes), and Tpo�/�
(knockout) BALB/cJ mice were inoculated with 4T1.2, andmaintained for 4 to 5 weeks to allow for metastasis to occur.We predicted that the Tpo�/� mice would have fewer metastasesthan wild type or heterozygotes if the megakaryocytes werepositively involved with the metastatic colonization ofthe bone. Contrary to our prediction, the Tpo�/� mice devel-oped more aggressive metastases than either heterozygous orwild-type mice (Fig. 4A–C). The IVIS images revealed that theTpo�/� mice developed widespread metastases (Fig. 4A and B)by about 33 days. The heterozygotes showed extensive metas-tases by approximately day 35, whereas the wild-type mice didnot show strong evidence of widespread metastases until aboutday 40. The days of sacrifice were significantly earlier for theTpo�/� mice (Fig. 4C) compared with the Tpoþ/� and WT mice.The reduction of the RBCs was greater in the knockout mice atthe time of sacrifice compared with the wild-type and hetero-zygote mice (Fig. 4D). All groups exhibited splenomegalyfollowing injection of 4T1.2 cells.
A count of the megakaryocytes in the femurs and spleens (Fig.4E and F) revealed a significant increase only in the megakaryo-cytes in the femurs of Tpoþ/� mice inoculated with the 4T1.2 butnot in the other cohorts. In contrast, the increase in megakaryo-cytes in the spleens was greatest in the wild-type mice. Thisincrease was also seen in the heterozygotes. There were too fewmegakaryocytes in the knockout mice to be significant. WBCincreased in mice injected with wild-type, heterozygotes, andknockoutmice (Fig. 4G). However, an analysis of the populationsof WBC (Fig. 4H) revealed a shift in lymphocytes to granulocytesin the wild type and in the heterozygotes but not in the knockoutmice. The percentage of monocytes remained unchanged amongthe groups.
The combination of metastatic cancer cells andosteoblasts produce factors that stimulate aspects ofmegakaryopoiesis in vitro
The in vivo data suggested that cancer cells that metastasized tothe bone produced factors that enhanced megakaryopoiesis. Inprevious work, we found that osteoblasts in the presence ofmetastatic cells undergo an inflammatory response and secreteseveral cytokines, some of which are known to take part in themegakaryopoiesis (13). Thus, we asked if factors produced by thecombination of cultures of osteoblasts and cancer cells enhancedsteps in megakaryopoieis. We carried out a series of in vitro assaysusing a human megakaryocyte line, MEG-01, and conditionedmedia fromMDA-MB-231 cancer cells, or fromMC3T3-E1 osteo-blasts or conditioned medium from MC3T3-E1 incubated with
Jackson et al.
Cancer Res; 77(8) April 15, 2017 Cancer Research1946
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084

cancer cell conditioned medium (double conditioned medium).We also tested a model of 4T1.2 murine cells and normal humanosteoblasts (NHOsts). We asked if the various conditioned
media stimulatedMEG-01proliferation/differentiation includingadherence and ploidy development. We also assayed for mega-karyocyte colony–forming units (CFU-MK).
7 14 32 40Days
4.0×107
3.0×107
2.0×107
1.0×107
07 14 32 40
Days
Pho
ton
flux
0.8
0.6
0.4
0.2
0.0PBS 4T1.2
Wei
ght (
g)
PBS PBS4T1.2 4T1.267NR 67NR
Femur
121110
9876543210
7 14 32 40
7 14 32 40 7 14 32 40 7 14 32 40
7.5
5.0
2.5
0.07 14 32 40
MK
s/Fi
eld
MK
s/Fi
eld
Pla
tele
ts (×
103 /
µL)
DaysDays
Days Days Days
Spleen
PBS 4T1.2 67NR1,500
1,250
1,000
750
500
250
0
1,500
1,250
1,000
750
500
250
0
1,500
1,250
1,000
750
500
250
0
A
B
C
E
D
Figure 2.
Splenicmegakaryocytes increased inmice injectedwithmurinemetastatic 4T1.2, but notwith nonmetastatic 67NR breast cancer cells. Six BALB/cJmice per timewereinjected in the fourth mammary gland with either 4T1.2 or 67NR cells or with PBS. Mice were sacrificed 7, 14, 32, and 40 days postinjection. A, RepresentativeIVIS images of mice inoculated with 4T1.2 (left) and graphs of average photon flux values (right). B, Image comparison of spleens from PBS (right)- and 4T1.2 (left)-injected mice. The spleens of mice injected with 4T1.2 weighed on average 0.60 g � 0.5 SD compared with spleens from PBS-injected mice, 0.09 g � 0.01 SD.C,Shown is abar graphof the averagenumberofmegakaryocytes (MK)perfield�SD(�400) in the femurs frommice injectedwith4T1.2, 67NR,orPBS.Atday32, therewere fewer megakaryocyte in the femurs of the 4T1.2 mice than in the femurs of the 67NR inoculated and less than in the PBS-treatedmice.D, Shown is a bar graph ofthe average number of megakaryocytes per field � SD in the spleens of the same groups of mice. There was a significant increase over time in the numbers ofmegakaryocyte in the spleen in the 4T1.2 bearing mice, but not in the 67NR- or PBS-injected mice. E, Platelet counts were analyzed by Hemavet 950FSfor eachgroupofmice. Shownare the individual counts and themean.Platelet countswere comparedwithin each treatment. Therewere significant increases in plateletcounts over time for the mice injected with 4T1.2. The PBS-injected mice showed an increase in the number of platelets after day 7 compared with the other times,but there was no increase thereafter. Platelet counts for 67NR injected mice did not change over time. � , P < 0.05; �� , P < 0.01; ��� , P < 0.001; #, P < 0.05.
Megakaryocytes and Bone Metastasis
www.aacrjournals.org Cancer Res; 77(8) April 15, 2017 1947
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084

Proliferation (Fig. 5A) of MEG-01 carried out with 10% FBS or1% FBSwas significantly increased with 10% double conditionedmedium, i.e. medium fromMC3T3-E1 incubated withMDA-MB-231 conditioned medium. The increase was approximately two-to three-fold above the numbers of the untreated cells. Stimula-tion of proliferation to a similar extent was also seen with 5%conditionedmedium. In contrast the conditionedmedia from the4T1.2, NHOst, and combinations did not significantly increaseproliferation of the MEG-01 cells (Fig. 5B).
An important stage of megakaryopoiesis is the adherenceof megakaryocytes to bone marrow endothelial cells (23).
The conditioned media from osteoblasts or the double condi-tion media caused adherence of the MEG-01 cells in thecultures (Fig. 5C). Conditioned medium from the cancer cellsalone did not.
Another aspect of the megakaryocyte differentiation is devel-opment of polyploidy through endoreplication. Incubating thecells with conditioned media from osteoblasts, cancer cells ordouble conditionedmedia did not lead to a significant increase inploidy (Supplementary Fig. S3).
Finally we asked if the conditioned medium increased thenumber of CFU-MK in primary bone marrow (Supplementary
9876543210
7 14 32 40 7 14 32 40Days Days
RB
Cs
(M/µ
L)
Tota
l WB
Cs
(×10
3 /m
L) 50
40
30
20
10
0
PBS4T1.267NR
PBS4T1.267NR
PBS 4T1.2 67NR
GranulocytesLymphocytesMonocytes
A
C
D E
B
3,500
3,000
2,500
2,000
1,500
1,000
500
0
3,500
3,000
2,500
2,000
1,500
1,000
500
07 14 32 40 7 14 32 40
Days Days
TPO
(pg/
mL)
SD
F-1
(pg/
mL)
PBS4T1.267NR
PBS4T1.267NR
Figure 3.
Changes in the populations of blood cells and serum levels of TPO and SDF-1 in mice injected with 4T1.2 and 67NR cancer cells. Mice were injected with 4T1.2, 67NR,or PBS as described in the Fig. 2 legend. Complete blood counts were determined from Hemavet analysis at the times indicated following tumor cell injection.A, RBC counts were reduced in mice injected with 4T1.2. Shown is the average value � SD (n ¼ 6 per group). Mice injected with 4T1.2 had fewer RBCs thanthose injected with PBS at days 4, 32, and 40. The mice bearing 67NR had more RBC at day 7 and day 40 than the PBS-injected mice. B, The numberof WBCs increased three- to five-fold at days 32 and 40 only in the mice injected with 4T1.2 cells. C, The distribution of the WBC subtypes at day 40 followingtumor cell injections. The percentages of granulocytes increased significantly, with a corresponding decrease in the percentage of lymphocytes in the miceinjected with the 4T1.2 or 67NR. Greater than 95% of the granulocyte population was due to an increase in neutrophils. The percentage of monocytes did notchange among the groups. D, TPO concentrations in the sera were measured by ELISA. Mice injected with 67NR showed increased levels compared withPBS and to 4T1.2 mice at all times measured. The sera from the 4T1.2 mice had a significantly reduced concentration at day 40. E, SDF-1 serum concentrations weremeasured by ELISA. Concentrations dropped by day 32 in the 4T1.2- and 67NR-injected mice, and remained significantly lower in the 4T1.2 mice. Significantdifferences are indicated. � , P < 0.05; �� , P < 0.01; ��� , P < 0.001.
Jackson et al.
Cancer Res; 77(8) April 15, 2017 Cancer Research1948
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084

Fig. S4). None of the conditioned media affected CFU-MK. Onlythe combination of TPO, IL3, IL6, and IL11 significantly increasedCFU-MK.
An increase in megakaryocytes was observed in humans withmetastatic breast cancer
Is the increase in megakaryocytes seen in mice with metastaticbreast cancer also present in the humanmetastatic patients? It wasvery difficult to obtain useable clinical bone marrow samplesfrom patients with metastatic bone cancer. However in samplesobtained from a matched autopsy set, six out of eight matchedpairs indicated an increase inmegakaryocytes in the bonemarrowof the patient who died ofmetastatic breast cancer comparedwithan age and gendermatchedpatientwhodied of noncancer-relatedcauses (Fig. 6 and Supplementary Table S1). The increase rangedfrom about 1.4- to 7.5-fold.
DiscussionVery little research has focused on the interaction of cancer
cells and megakaryocytes. Because megakaryocytes are largeand reside in the marrow, they are difficult to study. However,megakaryocytes are the sole source of platelets; and plateletshave been recognized to be associated with cancer for morethan 100 years (see ref. 24). Recent publications concerningthe role of megakaryocytes in metastasis offer conflictingtheories. In one case, Li and colleagues suggest that mega-karyocytes inhibit carcinoma growth in vitro and in vivo (6).When they injected nude mice with TPO to increase mega-karyocytes before inoculation of metastatic prostate carcinomaPC-3 cells, a decrease in hind limb tumor burden was seen.They reasoned that because megakaryocytes line the sinusoidsof the marrow, they are positioned to be one of the first cellsto interact with disseminated tumor cells. An inhibition inmetastases suggested that megakaryocytes suppressed PC-3cells via cell-to-cell contact (6). Zaslavsky and colleaguesindicated that platelets and megakaryocyte-derived TSP-1plays a critical role in suppressing tumor angiogenesis, whichlimits metastasis in the earliest stages of tumor growth.
In contrast to these protective mechanisms, Psaila and col-leagues (7) suggest that megakaryocytes promote metastasis.They imply that the vascular niche is a site for preferentialhoming and engraftment of malignant cells due in part to theincreased localization of megakaryocytes. They point out thatmegakaryocytes secrete an array of cytokines (7) that stimulateosteoblast proliferation and inhibit osteoclast differentiation.Megakaryocytes also secrete pro- and antiangiogenic cytokinessuch as VEGF and endostatin (25). In addition, following totalbody irradiation, megakaryocytes orchestrate regeneration ofthe marrow compartments (7). The megakaryocytes stimulateosteoblasts to proliferate; thus, they also may stimulate cancercell growth.
The lack of extensive research in the role of megakaryocytes inmetastatic cancer is partly due to thedifficulty inobtaininghumanbone samples that have not been compromised by destructivemetastasis. In addition there exists the technical challenge ofisolating intact megakaryocytes (7). In this study both a xenograftand a syngeneic model were used to determine the relationshipbetween megakaryocytes and tumor metastasis. We found anincrease in megakaryocytes with metastases but not with primarytumors. Because megakaryocytes produce a variety of cytokinesand chemokines that affect many cell types and processes, wefurther asked whether our data supported the idea that mega-karyocytes prepare a niche for the cancer cells as suggested byPsaila (7); or acted to protect against cancer cell colonization assuggested by Li (6) and Zaslavsky (5).
In this study, the xenograft intracardiac model provided evi-dence thatmetastases colonized themarrowprior to an increase inmegakaryocytes. The presence of primarymammary gland tumorsfor up to 34 days was not sufficient to increase megakaryocytes.We have previously reported that several cytokines including IL6increase in the bone marrow as part of the osteoblast inflamma-tory response to cancer cells (16). In addition, IL6 has been foundto increase the proliferation of premegakaryocytes (26). Thus, themegakaryocyte poolmay expand in response to an increase in IL6and other cytokines released by the stromal cells in the bone. Invitro experiments carried out with amegakaryocyte line supportedthe idea that osteoblasts in the presence of cancer cells producefactors that increase megakaryocyte proliferation and adherence,two important steps in megakaryopoiesis. Likely candidatesincluded TPO, IL6, and IL3. The conditioned media did notcontain TPO or IL3 as determined by ELISA. IL6 seemed unlikelybecause murine IL6 does not cross react with human cells.Moreover, addition of neutralizing antibody to IL6 to the culturesdid not change proliferation. AlsoMEG-01 cells secrete IL6, whichis likely autoreactive (27).
Using the syngeneic model of spontaneous metastasis fromthe mammary gland to the bone marrow, we found similarresults. In this case, the 67NR tumors were significantly largerthan the primary 4T1.2 tumors; nonetheless, there was noincreased megakaryopoiesis. Although there was no increasein megakaryocytes in the femurs of mice injected with 4T1.2compared with those inoculated with 67NR, or PBS, there was a4-fold increase of megakaryocytes in the spleens, first observed14 days postinoculation. The spleens also were significantlylarger than the control mice by day 14. The in vivo experimentsand the in vitro data taken together suggest that megakaryopoi-esis increased in response to the presence of metastatic cancercells in the bone marrow.
We used a Tpo knockout mouse to reduce megakaryocytes(andplatelets) speculating that theywould have fewermetastases.Contrary to our expectation, TPO knockout BALB/cJmice injectedorthotopically with 4T1.2 cells displayed more aggressive metas-tasis and a decreased survival compared with wild-type mice
Table 1. Phenotype of Tpoþ/þ, Tpoþ/�, and Tpo�/� mice
GenotypeTpo(pg/mL)
Megakaryocytes/field(spleen)
Megakaryocytes/field(femur)
Platelets(103/mL)
Tpoþ/þ 2,868.6 (� 271.7) 5.1 (� 1.6) 7.5 (� 1.8) 384.3 (� 46.7)Tpoþ/� 1,468.4 (� 246.9) 2.1 (� 1.8) 3.4 (� 1.2) 235.7 (� 12.2)Tpo�/� ND 0.2 (� 0.04) 0.3 (� 0.2) 42.0 (� 28.5)
NOTE: n ¼ 8; X � SEM.Abbreviation: ND, not detected.
Megakaryocytes and Bone Metastasis
www.aacrjournals.org Cancer Res; 77(8) April 15, 2017 1949
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084

TPO+/+
TPO+/-
TPO-/-
14 21 33Days
Day
s of
max
pho
ton
flux
Day
s of
sur
viva
lR
BC
s (M
/µL)
WB
C (×
103 /
µL)
MK
s/fie
ld
MK
s/fie
ld
% W
BC
100
80
60
40
20
04T1.2
TPO
– + + +– –
+/+ +/– –/–
GranulocytesLymphocytesMonocytes
76543210
PBS4T1.2
PBS4T1.2
TPO+/+TPO+/+ TPO+/-TPO+/- TPO-/-TPO-/-
TPO+/+ TPO+/- TPO-/-
TPO+/+ TPO+/- TPO-/-
TPO+/+ TPO+/- TPO-/-
TPO+/+ TPO+/- TPO-/-
Genotypes
Genotypes
Genotypes
Genotypes
Genotypes
Genotypes
SpleenFemur
PBS4T1.2
PBS4T1.2
109876543210
60
40
20
0
50
40
30
20
10
0
50
40
30
20
10
0
14 32 40
14 28 35
9876543210
A B
C
D
E
G H
F
300
250
200
150
100
50
300
250
200
150
100
50
300
250
200
150
100
50
Figure 4.
Tpo�/� mice had fewer megakaryocytes (MK) but developed metastasis more rapidly than the Tpoþ/þ and Tpoþ/� mice. Mice were inoculated in the mammaryglandswith4T1.2orPBSasdescribed inMaterials andMethods.A,Representativewhole-animal luciferase imageof Tpo�/�, Tpoþ/þ, andTpoþ/�over timeuntil the dateof sacrifice. B, Maximum photon flux for each mouse with the average � SEM for each genotype (n ¼ 6–17/genotype). The tumors grew more rapidlyand the metastatic spread was greater in the Tpo�/� mice. C, The Tpo�/� mice (n ¼ 17) developed metastasis more rapidly than the Tpoþ/þ (n ¼ 11) and Tpoþ/�
mice (n ¼ 15). The average survival time was 39.2 (�1.9 SD) days for Tpoþ/þ mice, 37.3 (�3.9 SD) days for Tpoþ/� mice, and 33.1 (�6.1) days for Tpo�/� mice.D, RBC counts were lower in tumor-bearing groups compared with PBS-treated mice. In this cohort, statistical significance (P < 0.05) was reached at about30 to 40 days, the last time of sacrificewith Tpo�/�; this decreasewas also seen at earlier times inwild-type Tpomice (see Fig. 3A). E,Megakaryocyteswere counted inthe femurs (E) and spleens (F). As determined previously (Fig. 2), there was not a significant increase in the femurs of the wild-typemice injectedwith 4T1.2; however,there was an increase in megakaryocytes in the heterozygotes even though there were fewer total megakaryocytes than in the wild-type mice. The Tpo�/� mice hadvery few megakaryocytes in the femur and their numbers did not increase following injection of 4T1.2. The spleens of all three groups of mice showed increasedmegakaryocytes in the spleens although the Tpo�/� mice had so few megakaryocytes that the increase was not statistically significant. G, WBC populations of allthree genotypes of mice increased significantly compared with mice injected with PBS. The numbers were not significantly different among the three genotypes.H, Although the total number of WBC increased in all three groups, the Tpoþ/þ mice showed a shift in the population to >60% granulocytes compared withlymphocytes (see Fig. 3), the heterozygousmice, Tpoþ/�, showed about 50% granulocytes and 50% lymphocytes, but theTpo�/�mice did not showa change from thenontumor-bearing mice (�20% granulocytes, 75% lymphocytes). Shown are mean � SEM. Significance is indicated by � , P < 0.05; �� , P < 0.01; ��� , P < 0.001.
Jackson et al.
Cancer Res; 77(8) April 15, 2017 Cancer Research1950
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084

injected with 4T1.2. Was this effect due to the reduction ofmegakaryocytes or to the lack of platelets? There is considerableevidence that platelets aid inmetastatic spread through a variety ofprocesses (24). Activation of platelets may allow tumor cells toenter the circulation, and protect tumor cells from immuneelimination (24). It is also known that increased platelet levelsare a poor prognostic factor for breast cancer metastasis (28), andthromboembolism is a common cause of death in cancer patients
(29). There are various reports that platelets protect circulatingtumor cells (30).Mezouar and colleagues (31) observed that in anectopicmodel of colon cancer, treatment ofmice with clopidogrel(Plavix), an antiplatelet drug, decreased the size of the tumors andrestored hemostasis. In a syngeneic orthotopic model of pancre-atic cancer, clopidogrel significantly inhibited the development ofmetastases (31). However, the role of megakaryocytes was notinvestigated.
400
300
200
100
0
150
100
50
0
150
100
50
0
Viab
le c
ells
(per
cent
)
Per
cent
adh
eren
t cel
ls
Viab
le c
ells
(per
cent
)
10% Conditioned medium
10% Conditioned medium
10% Conditioned medium
1% FBS10% FBS
10% FBS
A
C
B
NoneNone
None
MC3T3-E
1
MC3T3-E
1
MC3T3-E
1
MDA-MB-23
1
MDA-MB-23
1
MDA-MB-23
1
MDA-MB-23
1/MC3T
3-E1
MDA-MB-23
1/MC3T
3-E1
MDA-MB-23
1/MC3T
3-E1
MDA-MB-23
1/NHOst
NHOst
4T1.2
4T1.2
/MC3T
3-E1
4T1.2
/NHOst
None
MDA-MB-23
1/NHOst
NHOst
4T1.2
4T1.2
/MC3T
3-E1
4T1.2
/NHOst
Figure 5.
Conditioned medium from MC3T3-E1 osteoblasts treated with MDA-MB-231 conditioned medium enhanced megakaryocyte proliferation and differentiation in vitro.A, To determine whether cancer cell/osteoblast conditioned medium affected megakaryocyte proliferation, MEG-01 cells were plated in quadruplicate in 96-wellplates, 2.5� 103/well inRPMI1640mediumwith 1%or 10%FBSandwith5%or 10%conditionedmediumor left inRPMI1640untreated(none).Theconditionedmediumwasprepared as described in Materials and Methods from MDA-MB-231 cancer cells, MC3T3-E1 osteoblasts, or with MC3T3-E1 treated with MDA-MB-231 conditionedmedium to produce double conditionedmedium, as indicated. The cultures were incubated for 4 days at 37�C. On days 3 and 4, wells were incubatedwith MTS reagent(Promega, CellTiter96AqueousOne) and readwith a TECAN (InfiniteM1000 Pro) reader to detect viable cells. Shown are the average values� SEMonday 4 for the 10%condition medium, normalized to the no treatment control. Results with 5% conditioned medium were similar. The double conditioned medium from MDA-MB-231/MC3T3-E1 significantly increased the proliferation of MEG-01 cells. The conditionedmedium from the cancer cells alone or osteoblasts alone slightly but not significantlyenhanced proliferation. �� , P < 0.02, ��� , P < 0.0004. B, Similar experiments were carried out with condition medium from 4T1.2 cells, NHOst, or 4T1.2/NHOst doubleconditioned medium. Shown are the average values �SEM of the assays in 10% FBS. The conditioned medium did not result in significantly more cell numbers.C, Adherence assays were carried out as described for megakaryocytes (20). MEG-01, 1 � 104 per well, 24-well plates were plated in duplicate or quadruplicate andincubated in RPMI1640, 1% FBS with condition medium as described in A. After an overnight incubation at 37� , the numbers of adherent MEG-01 were counted infour fields perwell at�200magnification. Shown are the average values�SEM from two experiments totaling six replicates. Conditionedmedium fromosteoblasts andosteoblast/cancer cell combinations significantly increased MEG-01 adherence compared with cancer cell conditioned medium alone. ��� , P < 0.0001, �� , P < 0.03.This effect on adherence was seen in as little as 1% conditioned medium. Conditioned medium alone from either MDA-MB-231 or 4T1.2 did not affect adherence.
Megakaryocytes and Bone Metastasis
www.aacrjournals.org Cancer Res; 77(8) April 15, 2017 1951
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084

The results of our study support the idea that circulatingplatelets are not sufficient to explain an aggressive metastaticphenotype. The finding that Tpo knockout mice with greatlyreduced platelet numbers had more aggressive metastasis thanthewild type, suggests that other changes in themegakaryocytes orbone marrow were responsible.
One obvious candidate is TPO. The role for TPO in cancer isunknown; however, its role in maintaining HSC quiescence hasbeen established (32). The release of TPO in the endosteal orosteoblastic niche regulates the proliferation of resident HSCs. InMpl (the receptor for TPO) knockout mice, there is a reduction intheHSCpopulation. The hematopoietic niche (32) is a postulatedhaven formetastatic cells in the bone (33). Activation of the nicheand proliferation of HSC in a Tpo knockout mouse may permit amore fertile environment for metastases. This idea is furthersupported by the finding that megakaryocytes regulate HSCquiescence. Loss of megakaryocytes activate the HSC niche(34). Another piece of evidence that the HSC niche of the TPOknockout mice was modified, was the lack of a shift from leuko-cytes to granulocytes following injection of 4T1.2 (Fig. 4G)despite the increase in total WBC (Fig. 4F). We also have apreliminary observation that the bone volume and density inthe knockout mice is less than in the wild type.
TPO, together with SCF, GM-CSF, IL1, IL3, IL11, and IL6 areimportant for megakaryopoiesis (10). Additional chemokinessuch as LIX, MIP-1, and KC are thought to play a role in earlymegakaryocyte progenitor proliferation (35). Interestingly,these are produced by osteoblasts in response to cancer (13,36, 37). An increase in any one of those cytokines in wild-typemice may be due to host cells or be produced by the metastaticcells and could lead to more megakaryocytes. For example, 4T1tumor cells and MDA-MB-231 constitutively express genes forthe myeloid colony-stimulation factors G-CSF, GM-CSF, andM-CSF (17). The secretion of these growth factors contribute tothe extramedullary hematopoiesis, splenomegaly, and myelo-poiesis noted in the 4T1.2 injected mice. As has been reportedin a rat model of metastatic mammary carcinoma, a GM-CSF/
IL3-like cytokine, released from the cancer cells led to anincrease of megakaryocytes in the femur (7). In addition, ahost immune response to the cancer cells took place as severalcytokines including IL6 was released into the marrow fromresident osteoblasts (16, 38). The increase in IL6 could, in turn,promote megakaryopoiesis. The in vitro studies indicated thatIL6 probably did not act alone.
We also observed a significant decrease in SDF-1 (CXCL-12)concentrations in the bloodofwild-type BALB/c Jmice sacrificed atday 32 and day 40 post 4T1.2 inoculation. SDF-1 and its receptorCXCR4 are known to be important in various human cancer cellsand models of metastasis (39, 40). Likewise, CXCR4 is found onTregs and other bone marrow cells and appears to promote cellmobilization. The disruption of the CXCR4–SDF-1 axis mayfacilitate mobilization of Treg cells and other CXCR4þ cells intoperipheral blood. This disruptionmayalsoplay a role in cancer cellmobilization, as a decrease in SDF-1 levels late in the experimentaltime line seemed to coincide with more advanced metastasis.
In humans, G-CSF is often prescribed for patients who undergochemotherapy; and it causes extramedullary hematopoiesis (41).The resulting megakaryocytes are sometimes mistaken for cancercells and misdiagnosis occurs (42). From our data it appears thatmegakaryocytes increase in response to the presence of cancer cellsin the bone microenvironment; patients who died from meta-static breast cancer had increased numbers of megakaryocytes inthe bone marrow (Fig. 6). Winklemann and colleagues, reportedthat megakaryocyte ploidy, related to platelet production, washigher in patients who died of metastatic cancer compared withpatients who did not (43). Thus, this increase in platelets andmegakaryocytes correlated with more metastasis.
Paradoxically, high levels of anti-angiogenic TSP-1 secretedby platelets and megakaryocytes (5) and the cancer cell inhib-itory qualities that the megakaryocytes (6) possess, may help toslow down bone metastasis. TPO is used to help replenishplatelets in patients undergoing chemotherapy. There is littleresearch that focuses on TPO as a therapeutic. Our finding, thata decrease in megakaryocytes was associated with an increase inmetastasis, suggested that megakaryocytes may have a protec-tive role in metastatic cancer to bone. It is possible that in thenormal mouse they compete with the tumor cells for theendosteal niche or directly inhibit cancer growth as suggestedby Li and colleagues (6).
In conclusion, our data support the hypothesis that in the waron cancer,megakaryocytes battle againstmetastasis. However, thefindings create more questions. Clearly further studies with theTpo�/� mice may offer valuable insights into the metastaticprocess. One preliminary observation that is being pursued isthat the bones of the Tpo�/�mice are less dense than the bones ofthe wild type. Although metastases were not restricted to bone,but indeed may develop from cancer cells that find haven there.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: W. Jackson III, D.M. Sosnoski, A.M. MastroDevelopment of methodology: W. Jackson III, D.M. Sosnoski, A.M. MastroAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): W. Jackson III, D.M. Sosnoski, S.E. Ohanessian,A. Mobley, K.D. Meisel, A.M. Mastro
80
60
40
20
0
MK
s/m
m2
Metastatic cancer Noncancer relatedCause of death
Figure 6.
Increase in bone marrow megakaryocytes (MK) in patients with metastaticbreast cancer. Autopsy samples of bone from women who died of metastaticbreast cancer (n ¼ 8) and age-matched samples from women who died ofnoncancer-related deaths (n ¼ 8) were stained for megakaryocytes. Thesamples were sectioned, stained for vWF, and counted as described in MaterialsandMethods. In six of the eight paired cases, there was an increase in the ratio ofmegakaryocytes in the boneof thewomenwhodied ofmetastatic breast cancer.The average increase (3.23� 2.25 SD) approached significance (P<0.057) in thepatients with metastatic cancer (unpaired t test).
Cancer Res; 77(8) April 15, 2017 Cancer Research1952
Jackson et al.
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084

Analysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): W. Jackson III, D.M. Sosnoski, K.D. Meisel,A.M. MastroWriting, review, and/or revision of the manuscript: W. Jackson III, D.M.Sosnoski, S.E. Ohanessian, P. Chandler, A. Mobley, K.D. Meisel, A.M. MastroAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): W. Jackson III, D.M. Sosnoski, A.M. MastroStudy supervision: A.M. Mastro
AcknowledgmentsThe authors thank Ruth Nissley and John Cantolina of the Huck Institute
Microscopy and Cytometry Facility, Penn State University; Yu-Chi Chen,Nick Kendsersky, Johnathan Vicenty, Richa Pursnani, Emily Rutan, WilliamTurbitt, Ashaki Nehisi, Shuleika Lopez Ortiz, Karen Bussard, and VenkateshKrishnan for technical assistance; Dr. Connie J. Rogers, William Kraemer(Ohio State) and the Penn State Statistical Consulting Center for help with
data analysis; and Dr Jake Werner for his help with platelet analysis. Wegratefully acknowledge Genentech's generous gift of C57Blk6 Tpo -/- mouseembryos.
Grant SupportU.S. ArmyMedical andMateriel Command Breast Cancer Research Program,
W81XWH-10-1-0253 to A.M. Mastro, Alfred P. Sloan Foundation GraduateAward to W. Jackson.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received April 21, 2016; revised December 8, 2016; accepted December 26,2016; published OnlineFirst February 15, 2017.
References1. Wilson A, Trumpp A. Bone-marrow haematopoietic-stem-cell niches. Nat
Rev Immunol 2006;6:93–106.2. Papadantonakis N, Ravid K. Development of megakaryocytes. In:
Wicrema A, Kee B, editors. Molecular basis of hematopoiesis. NewYork: Springer; 2009. p95–126.
3. KacenaMA,Nelson T, CloughME, Lee SK, Lorenzo JA, Gundberg CM, et al.Megakaryocyte-mediated inhibition of osteoclast development. Bone2006;39:991–9.
4. Lawler J.Megakaryocytes join the war on cancer. Blood 2010;115:4326–7.
5. Zaslavsky A, Baek KH, Lynch RC, Short S, Grillo J, Folkman J,et al. Platelet-derived thrombospondin-1 is a critical negativeregulator and potential biomarker of angiogenesis. Blood 2010;115:4605–13.
6. Li X, Koh AJ, Wang Z, Soki FN, Park SI, Pienta KJ, et al. Inhibitory effects ofmegakaryocytic cells in prostate cancer skeletalmetastasis. J BoneMiner Res2011;26:125–34.
7. Psaila B, Lyden D, Roberts I. Megakaryocytes, malignancy and bonemarrow vascular niches. J Thromb Haemost 2012;10:177–88.
8. Psaila B, Kaplan RN, Port ER, Lyden D. Priming the 'soil' forbreast cancer metastasis: the pre-metastatic niche. Breast Dis 2006;26:65–74.
9. Bussard KM, Okita N, Sharkey N, Neuberger T, Webb A, MastroAM. Localization of osteoblast inflammatory cytokines MCP-1 andVEGF to the matrix of the trabecula of the femur, a target area formetastatic breast cancer cell colonization. Clin Exp Metastasis 2010;27:331–40.
10. Eckhardt BL, Parker BS, van Laar RK, Restall CM, Natoli AL, Tavaria MD,et al. Genomic analysis of a spontaneous model of breast cancermetastasis to bone reveals a role for the extracellular matrix. Mol CancerRes 2005;3:1–13.
11. Cailleau R, Young R, Olive M, Reeves WJJr. Breast tumor cell lines frompleural effusions. J Natl Cancer Inst 1974;53:661–74.
12. Chen YC, Prabhu KS, Das A, Mastro AM. Dietary selenium supplementa-tion modifies breast tumor growth and metastasis. Int J Cancer 2013;133:2054–64.
13. Bussard KM, Venzon DJ, Mastro AM. Osteoblasts are a major source ofinflammatory cytokines in the tumor microenvironment of bone meta-static breast cancer. J Cell Biochem 2010;111:1138–48.
14. Cao Y, Slaney CY, Bidwell BN, Parker BS, Johnstone CN, Rautela J, et al.BMP4 inhibits breast cancer metastasis by blocking myeloid-derivedsuppressor cell activity. Cancer Res 2014;74:5091–102.
15. Phadke PA, Mercer RR, Harms JF,. Jia Y, Kappes JC, Frost AR, et al. Kineticsof metastatic breast cancer cell trafficking in bone. Clin Cancer Res2006;12:1431–40.
16. BussardKM,MastroAM. Ex-vivo analysis of the bonemicroenvironment inbone metastatic breast cancer. J Mammary Gland Biol Neoplasia 2009;14:387–95.
17. DuPre SA, Hunter KWJr.Murine mammary carcinoma 4T1 induces aleukemoid reaction with splenomegaly: association with tumor-derivedgrowth factors. Exp Mol Pathol 2007;82:12–24.
18. Alugupalli KR, Michelson AD, Barnard MR, Leong JM. Serial determina-tions of platelet counts in mice by flow cytometry. Thromb Haemost2001;86:668–71.
19. Murone M, Carpenter DA, de Sauvage FJ. Hematopoietic deficiencies in c-mpl and TPO knockout mice. Stem Cells 1998;16:1–6.
20. Levine RF, Eldor A, HyAm E, Gamliel H, Fuks Z, Vlodavsky I. Megakaryo-cyte interaction with subendothelial extracellular matrix is associated withadhesion, platelet-like shape change, and thromboxane A2 production.Blood 1985;66:570–6.
21. Liu M, Jin X, He X, Pan L, Zhang X, Zhao Y. Macrophages support splenicerythropoiesis in 4T1 tumor-bearing mice. PLoS ONE 2015;10:e0121921.
22. Petit I, Szyper-KravitzM, Nagler A, LahavM, Peled A, Habler L, et al. G-CSFinduces stem cell mobilization by decreasing bone marrow SDF-1 and up-regulating CXCR4. Nat Immunol 2002;3:687–94.
23. Avecilla ST, Hattori K, Heissig B, Tejada R, Liao F, Shido K, et al. Chemo-kine-mediated interaction of hematopoietic progenitors with the bonemarrow vascular niche is required for thrombopoiesis. Nat Med 2004;10:64–71.
24. Gay LJ, Felding-Habermann B. Contribution of platelets to tumour metas-tasis. Nat Rev Cancer 2011;11:123–34.
25. MundyG.Metastasis to bone: causes, consequences and therapeutic oppor-tunities. Nat Rev Cancer 2002;2:584–93.
26. Berl T, Schwertschlag U. Preclinical pharmacologic basis for clinicaluse of rhIL-11 as an effective platelet-support agent. Oncology2000;14:12–20.
27. Navarro S, Mitjavila MT, Katz A, Doly J, Vainchenker W. Expression ofinterleukin 6 and its specific receptor by untreated and PMA-stimulatedhuman erythroid and megakaryocytic cell lines. Exp Hematol 1991;19:11–7.
28. Taucher S, Salat A, Gnant M, Kwasny W, Mlineritsch B, Menzel RC, et al.Impact of pretreatment thrombocytosis on survival in primary breastcancer. Thromb Haemost 2003;89:1098–106.
29. TrikhaM,NakadaMT. Platelets and cancer: implications for antiangiogenictherapy. Semin Thromb Hemost 2002;28:39–44.
30. Varon D, Shai E. Platelets and their microparticles as keyplayers in pathophysiological responses. J Thromb Haemost 2015;13:S40–6.
31. Mezouar S, Darbousset R, Dignat-George F, Panicot-Dubois L, Dubois C.Inhibition of platelet activation prevents the P-selectin and integrin-dependent accumulation of cancer cell microparticles and reduces tumorgrowth and metastasis in vivo. Int J Cancer 2015;136:462–75.
32. YoshiharaH,Arai F,HosokawaK,Hagiwara T, TakuboK,NakamuraY, et al.Thrombopoietin/MPL signaling regulates hematopoietic stem cell quies-cence and interaction with the osteoblastic niche. Cell Stem Cell 2007;1:685–97.
33. Shiozawa Y, Taichman RS. Cancer stem cells and the bone marrowmicroenvironment. Bonekey Rep 2012;2012:pii: 48.
34. Bruns I, Lucas D, Pinho S, Ahmed J, Lambert MP, Kunisaki Y, et al.Megakaryocytes regulate hematopoietic stem cell quiescence throughCXCL4 secretion. Nat Med 2014;20:1315–20.
www.aacrjournals.org Cancer Res; 77(8) April 15, 2017 1953
Megakaryocytes and Bone Metastasis
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084

35. Choong ML, Yong YP, Tan AC, Luo B, Lodish HF. LIX: a chemokine with arole in hematopoietic stem cells maintenance. Cytokine 2004;25:239–45.
36. KinderM,ChislockE, BussardKM, ShumanL,MastroAM.Metastatic breastcancer induces an osteoblast inflammatory response. Exp Cell Res 2008;314:173–83.
37. Sosnoski DM, Krishnan V, Kraemer WJ, Dunn-Lewis C, Mastro AM.Changes in cytokines of the bone microenvironment during breast cancermetastasis. Int J Breast Cancer 2012;2012:160265.
38. Mercer R, Miyasaka C, Mastro AM. Metastatic breast cancer cells sup-press osteoblast adhesion and differentiation. Clin Exp Metastasis2004;21:427–35.
39. Muller A, Homey B, Soto H, Ge N, Catron D, Buchanan ME, et al.Involvement of chemokine receptors in breast cancer metastasis. Nature2001;410:50–6.
40. Williams SA, Harata-Lee Y, Comerford I, Anderson RL, Smyth MJ, McCollSR. Multiple functions of CXCL12 in a syngeneic model of breast cancer.Mol Cancer 2010;9:250.
41. Redmond J3rd, Kantor RS, Auerbach HE, Spiritos MD, MooreJT. Extramedullary hematopoiesis during therapy with granulo-cyte colony-stimulating factor. Arch Pathol Lab Med 1994;118:1014–5.
42. Hoda SA, Resetkova E, Yusuf Y, Cahan A, Rosen PP. Megakaryocytesmimicking metastatic breast carcinoma. Arch Pathol Lab Med2002;126:618–20.
43. Winkelmann M, Stockler J, Grassmuck J, Pfitzer P, Schneider W. Ploidypattern of megakaryocytes in patients with metastatic tumors with andwithout paraneoplastic thrombosis and in controls. Haemostasis1984;14:501–7.
Cancer Res; 77(8) April 15, 2017 Cancer Research1954
Jackson et al.
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084

2017;77:1942-1954. Published OnlineFirst February 15, 2017.Cancer Res Walter Jackson III, Donna M. Sosnoski, Sara E. Ohanessian, et al. Role of Megakaryocytes in Breast Cancer Metastasis to Bone
Updated version
10.1158/0008-5472.CAN-16-1084doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2017/02/15/0008-5472.CAN-16-1084.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/77/8/1942.full#ref-list-1
This article cites 42 articles, 6 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/77/8/1942.full#related-urls
This article has been cited by 2 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/77/8/1942To request permission to re-use all or part of this article, use this link
on March 23, 2020. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 15, 2017; DOI: 10.1158/0008-5472.CAN-16-1084