Embed Size (px)
Citation preview

J. Mol. Biol. (1996) 260, 304–311
COMMUNICATION
RNP Export is Mediated by Structural Reorganizationof the Nuclear Pore Basket
Elena Kiseleva 1,2,3, Martin W. Goldberg 1, Bertil Daneholt 2 andTerence D. Allen 1*
Messenger RNA leaves the cell nucleus as ribonucleoprotein (RNP)1CRC Department ofparticles. The nucleocytoplasmic translocation of the particles takes placeStructural Cell Biologythrough the nuclear pore complex (NPC) and includes two steps: bindingPaterson Institute for Cancerto the NPC and transit through its central channel. The NPC basket is aResearch, Christie Hospital
National Health Service fishtrap-like component of NPC facing the nucleoplasm. Its position in theNPC strongly suggests that it has an important role in the initial stepsTrust, Manchester
M20 9BX, UK of macromolecular export from the nucleus. Here we report a cyclicrearrangement of the basket structure in relation to the translocation of a2Department of Cell and specific messenger RNP (mRNP) of exceptional size, the Balbiani ring RNP
Molecular Biology, Medical particles in the salivary gland cells in Chironomus. We used field emissionNobel Institute, Karolinska in-lens scanning electron microscopy (FEISEM), transmission electronInstitutet, S-171 77 microscopy (TEM), and immunocytochemistry to analyse the structuralStockholm, Sweden organization of the basket during the mRNP export. Our observations
reveal five configurations of the basket which are presented in a model of3Institute of Cytology andbasket reorganization related to the state of mRNP penetration into theGenetics, NovosibirskNPC. We suggest that the functional role of the basket is to anchor the630090, RussiamRNP particle to the NPC and position it in correct orientation at theentrance to the central channel of the NPC.
7 1996 Academic Press Limited
Keywords: nuclear pore; RNP export; nuclear pore basket; Chironomus;*Corresponding author scanning microscopy
To study the functional role of the NPC basketsin mRNP export we used salivary gland cellsof Chironomus, as they contain abundant(pre)messenger RNP particles of giant size, theBalbani ring (BR) RNP particles (Mehlin &Daneholt, 1993), the product of amplified transcrip-tion from polytene chromosomes. During transcrip-tion, the BR RNA molecules, 35 to 40 kb in size, arecomplexed with proteins and packed into largeRNP particles, 50 nm in diameter (Mehlin &Daneholt, 1993). The primary transcripts containfour introns (Wieslander, 1994), but as all of themare short (less than 1.5 kb), the RNP particles areonly marginally reduced in size during processing.Since the particles remain large, they can be easilyidentified, not only during assembly on the BRgenes (Andersson et al., 1980; Olins et al., 1980;Skoglund et al., 1983) but also in the nucleoplasm(Olins et al., 1980; Skoglund et al., 1983, 1986) and
during translocation through the NPC (Stevens &Swift, 1966; Mehlin et al., 1992, 1995).
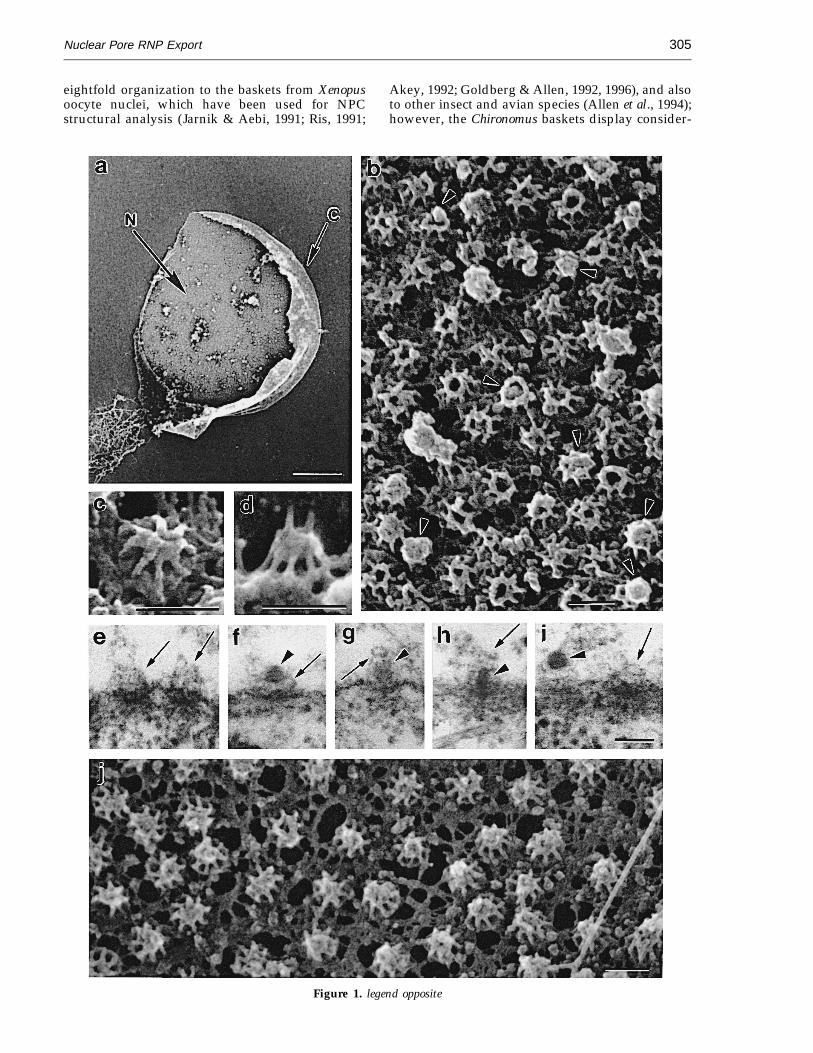
The basket structures in the Chironomus NPCswere studied with FEISEM after the nuclei wereisolated from salivary gland cells, fixed inglutaraldehyde and tannic acid, and processed asdescribed (Goldberg & Allen, 1992, 1996). Figure 1shows a partly opened cell nucleus (Figure 1a),displaying the nucleoplasmic side of the nuclearenvelope with many baskets (Figure 1b), andexamples of individual baskets (Figure 1c and d).NPC baskets extend into the nucleoplasm for 20 to40 nm and are composed of eight filaments, about10 nm in width and 20 to 40 nm in length (Fig-ure 1b to d). The filaments are anchored at theperiphery of the nuclear coaxial ring proximal tothe nuclear envelope and joined to each other bytheir distal ends, often forming a terminal ring(Figure 1b; cf. inset in Figures 3e and 4). In TEMsections of salivary gland cells, the baskets appearas fibrous structures extending into the nucleo-plasm (Figure 1e), and occasionally a terminal ringcan also be visualized (Figure 1g and i). It can beconcluded that Chironomus baskets have a similar
Abbreviations used: RNP, ribonucleoprotein; NPC,nuclear pore complex; FEISEM, field-emission in-lensscanning electron microscopy; TEM, transmissionelectron microscopy; BR, Balbiani ring.
0022–2836/96/280304–08 $18.00/0 7 1996 Academic Press Limited
Nuclear Pore RNP Export 305
eightfold organization to the baskets from Xenopusoocyte nuclei, which have been used for NPCstructural analysis (Jarnik & Aebi, 1991; Ris, 1991;
Akey, 1992; Goldberg & Allen, 1992, 1996), and alsoto other insect and avian species (Allen et al., 1994);however, the Chironomus baskets display consider-
Figure 1. legend opposite

Nuclear Pore RNP Export306
ably more variation in the morphology of theNPC-distal part (Figure 1b, arrowheads), whichmay be a consequence of the intense nucleocyto-plasmic export in salivary gland cells. In contrast,the nuclear envelope of Xenopus (Figure 1j) pre-pared in the same manner from Xenopus oocyte atstage 6, which is low in transcriptional activity(Davidson, 1986), displays a more or less uniformstructure of baskets.
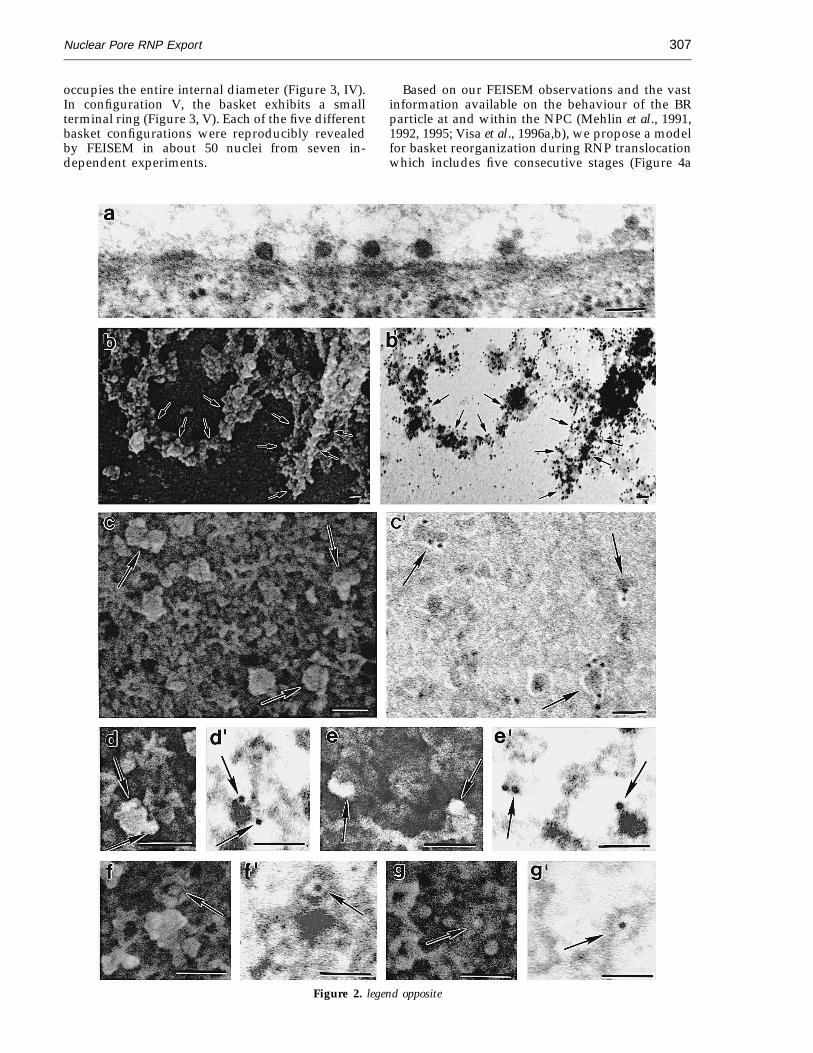
Numerous large particles were observed con-nected to Chironomus baskets in TEM (Figure 2a)and FEISEM specimens (Figure 1b, arrowheads). Todecide whether these globular particles, approxi-mately 50 nm in diameter, were BR RNP particles,we stained the specimen with an antibody againsta 45 kDa RNA-binding protein, known to be anabundant protein in the BR particles (Wurtz et al.,1996). As shown in Figure 2b to g', the 45 kDaprotein is present in nascent RNP particles at BRtranscription units (Figure 2b and b'), and as alsolabelled the majority of the globular particles at orwithin the NPC basket (Figure 2c to g'). Theseresults provide good evidence for the view thatmost of the large particles interacting with thenucleoplasmic periphery of the NPC are indeed BRRNP particles.
TEM analysis of sections cut perpendicular to thenuclear envelope reveals globular RNP particlesassociated with or ‘‘embraced’’ by the baskets(Figure 1f to h, arrowheads). When the particleshave penetrated into the central channel of theNPC, they appear rod-shaped (Figure 1h). Thisstriking conformational change of translocating BRRNP particles has been studied in detail both withconventional TEM (Stevens & Swift, 1966; Mehlinet al., 1991) and with electron tomography forthree-dimensional reconstruction (Mehlin et al.,1992, 1995). The tomographic analysis has shownthat a BR particle is best described as a ring-like structure, consisting of a bent RNP ribbon(Skoglund et al., 1986). The particle docks in frontof the entrance to the central channel of the NPC.Upon translocation, the ribbon extends and moves
through the channel or transporter region of theNPC, and the remaining portion of the ribbonrotates (Mehlin et al., 1995). Eventually, the wholeribbon has been straightened out and the leadingend has emerged into the cytoplasm (cf. Figure 4).This drastic conformational change of the BRparticle is required if the 50 nm particle is topass through the central channel of the NPC withits maximal diameter of 25 nm (Dworetzky &Feldherr, 1988).
Although the TEM approach has been suitablefor analysis of the BR RNP particle itself duringtranslocation, it has not been informative in studiesof the interaction between the NPC basket andthe particle. As shown above, FEISEM providesdirectly observed three-dimensional information onthe fine structure of a large number of baskets thatwould be impossible by thin section reconstructionin TEM (Figure 1b to d versus e to i), although itshould be kept in mind that some shrinkage of theNPC structures may occur during the critical pointdrying. Using the FEISEM technique, we have nowbeen able to demonstrate that the basket is modifiedin a defined manner in relation to mRNP export.
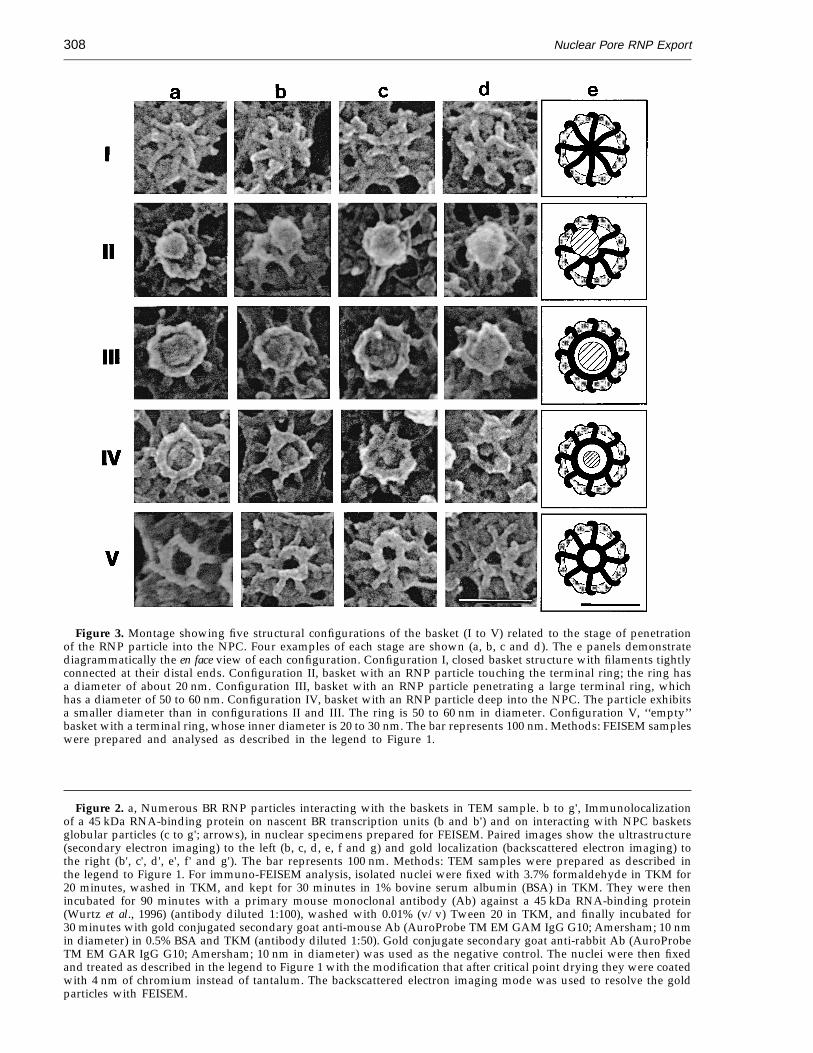
As shown in Figure 3, five different basketconfigurations were observed, two apparentlyempty (I and V) and three with associated RNPparticles (II to IV). In configuration I, the basketis closed, with the distal ends of the filamentsinterwoven (Figure 3, I). In configuration II, an RNPparticle is attached to the distal ends of thefilaments and a small terminal ring, 20 to 30 nm indiameter, can be seen (Figure 3, II). At this stage theRNP particle may be attached to the edge ofthe ring, prior to a more centralized location andthe ring may be incomplete (Figure 3, IIa). Inconfiguration III, an RNP particle fills the internalring space of the basket and the ring exhibits anincreased inner diameter of 50 to 60 nm (Figure 3,III). In configuration IV, the particle is deeper intothe basket and appears smaller in diameter; thebasket terminal ring has slightly decreased in sizeand the translocating RNP particle no longer
Figure 1. Nucleus from a Chironomus salivary gland cell and NPC baskets visualized by FEISEM and TEM. a, Isolatedand partly opened nucleus demonstrating cytoplasmic (C) and nucleoplasmic (N) sides of the nuclear envelope. b, Thenucleoplasmic side of the nuclear envelope with numerous basket structures (low magnification); arrowheads indicatethe RNP particles attached to the basket. c and d, Morphology of basket structure at high magnification (c, top view;d, side view). e and i, ‘‘Empty’’ basket structures (arrows) on sections of salivary gland cells. f to h, TEM thin sectionsof BR RNP particles (arrowheads) interacting with or passing through the baskets. j, The nucleoplasmic side of simi-larly prepared Xenopus oocyte nuclear envelope with numerous uniform baskets. Bars represent 10 mm for a and100 nm for b to j. Methods: for FEISEM, four to six salivary glands from fourth instar larvae of Chironomus thummiwere treated at 0°C for 30 seconds in 2% (v/v) Nonidet P-40 (NP-40) in TKM (100 mM KCl, 1 mM MgCl2, 10 mMtriethanolamine-HCl (pH 7.0)) and then transferred to 0.025% NP-40 in TKM. The nuclei were released from the glandsby pipetting in TKM and further transferred onto a silicon chip, fixed for ten minutes in 2% (v/v) glutaraldehyde and0.2% (w/v) tannic acid in TKM, rinsed with TKM, and fixed for ten minutes in OsO4 in TKM. Further processing andFEISEM analysis were performed as described in studies of NPCs from Xenopus oocytes (Goldberg & Allen, 1992, 1996).For TEM, salivary glands were isolated from larvae and fixed in 2% glutaraldehyde for two hours at 4°C and 1% (w/v)osmium tetroxide for 30 minutes at 4°C, dehydrated, and embedded in Epon. Prior to isolation of salivary glands, thelarvae were kept in cultivation medium provided with pilocarpine (0.1 mg/ml) for three to five hours at 22°C tostimulate BR gene transcription. Xenopus oocyte samples were prepared as described (Goldberg & Allen, 1993, 1996)including maintenance of basket structure after brief detergent extraction.
Nuclear Pore RNP Export 307
occupies the entire internal diameter (Figure 3, IV).In configuration V, the basket exhibits a smallterminal ring (Figure 3, V). Each of the five differentbasket configurations were reproducibly revealedby FEISEM in about 50 nuclei from seven in-dependent experiments.
Based on our FEISEM observations and the vastinformation available on the behaviour of the BRparticle at and within the NPC (Mehlin et al., 1991,1992, 1995; Visa et al., 1996a,b), we propose a modelfor basket reorganization during RNP translocationwhich includes five consecutive stages (Figure 4a
Figure 2. legend opposite
Nuclear Pore RNP Export308
Figure 3. Montage showing five structural configurations of the basket (I to V) related to the stage of penetrationof the RNP particle into the NPC. Four examples of each stage are shown (a, b, c and d). The e panels demonstratediagrammatically the en face view of each configuration. Configuration I, closed basket structure with filaments tightlyconnected at their distal ends. Configuration II, basket with an RNP particle touching the terminal ring; the ring hasa diameter of about 20 nm. Configuration III, basket with an RNP particle penetrating a large terminal ring, whichhas a diameter of 50 to 60 nm. Configuration IV, basket with an RNP particle deep into the NPC. The particle exhibitsa smaller diameter than in configurations II and III. The ring is 50 to 60 nm in diameter. Configuration V, ‘‘empty’’basket with a terminal ring, whose inner diameter is 20 to 30 nm. The bar represents 100 nm. Methods: FEISEM sampleswere prepared and analysed as described in the legend to Figure 1.
Figure 2. a, Numerous BR RNP particles interacting with the baskets in TEM sample. b to g', Immunolocalizationof a 45 kDa RNA-binding protein on nascent BR transcription units (b and b') and on interacting with NPC basketsglobular particles (c to g'; arrows), in nuclear specimens prepared for FEISEM. Paired images show the ultrastructure(secondary electron imaging) to the left (b, c, d, e, f and g) and gold localization (backscattered electron imaging) tothe right (b', c', d', e', f' and g'). The bar represents 100 nm. Methods: TEM samples were prepared as described inthe legend to Figure 1. For immuno-FEISEM analysis, isolated nuclei were fixed with 3.7% formaldehyde in TKM for20 minutes, washed in TKM, and kept for 30 minutes in 1% bovine serum albumin (BSA) in TKM. They were thenincubated for 90 minutes with a primary mouse monoclonal antibody (Ab) against a 45 kDa RNA-binding protein(Wurtz et al., 1996) (antibody diluted 1:100), washed with 0.01% (v/v) Tween 20 in TKM, and finally incubated for30 minutes with gold conjugated secondary goat anti-mouse Ab (AuroProbe TM EM GAM IgG G10; Amersham; 10 nmin diameter) in 0.5% BSA and TKM (antibody diluted 1:50). Gold conjugate secondary goat anti-rabbit Ab (AuroProbeTM EM GAR IgG G10; Amersham; 10 nm in diameter) was used as the negative control. The nuclei were then fixedand treated as described in the legend to Figure 1 with the modification that after critical point drying they were coatedwith 4 nm of chromium instead of tantalum. The backscattered electron imaging mode was used to resolve the goldparticles with FEISEM.

Nuclear Pore RNP Export 309
Figure 4. A tentative model for the dynamic changesof the NPC basket when a BR mRNP particle entersand passes through the basket. a, Closed ‘‘inactive’’basket. Basket filaments appear tightly connected toeach other by their distal ends. b, mRNP binding. RNPparticle touches the filaments in the distal region ofthe basket and initiates the formation of a terminalring. c, mRNP entering the basket. The terminal ringincreases in diameter concomitant with the penetrationof the particle into the basket. The length of basketfilaments decreases, probably due to their involvementin ring formation. The particle starts unfolding at theentrance of the central channel. d, Unfolding mRNPpassing deeper into the NPC. The mRNP particlebecomes translocated to the inner part of the NPC andgradually disappears from the basket ring. e, Open‘‘empty’’ basket. The basket ring gradually regresseswhile the individual basket filaments increase in length,approaching the configuration in a. Inset at top left:the basket in relation to the nuclear envelope (n,nucleoplasmic; c, cytoplasmic sides) and the three mainlayers of the NPC: the central spoke assemblysurrounded by the coaxial nuclear (above) and cyto-plasmic (below) rings (for recent reviews on NPC, seePante et al., 1994; Akey, 1995; Goldberg & Allen, 1995;Pante & Aebi, 1995).
to e). To allow the RNP particle to access the centralchannel of the NPC, the distal end of the basketgradually begins to open and a new structure isformed, the basket ring (Figure 4a to b). The ringexpands when the particle passes through the ringand docks at the entrance to the central channel,and the bent ribbon of the particle is graduallystraightened out and penetrates the central channel(Figure 4c). The basket ring retains its largediameter while translocation proceeds and theunfolded particle as a whole enters into the cen-tral channel (Figure 4d). The apparent decreasein diameter of the particle most likely reflects aconformational change of the particle: from aring-like to a rod-like structure. While we are awarethat the RNP particles from other sources (BR3 orother genes) on polytene chromosomes are smallerthan those from BR1 or BR2, the vast majority of allBR particles interacting with NPC baskets areapproximately 50 nm in diameter (Figure 2a andFigure 3). Smaller particles, centrally positioned,deep within the basket, most likely indicate laterstages of BR1 and BR2 granules exiting from thebasket ring as shown in the Figure 3, IV. When theparticle has left the basket, the basket ring closes(Figure 4e), and the initial state is reformed (Fig-ure 4a).
Although the model is well supported by theavailable data, a few uncertainties should bepointed out. Firstly, it is not yet possible to statewhether the basket ring is formed prior to, orsubsequent to, the attachment of the RNP particle.Secondly, it is not clear whether the ring doesdisassemble between each round of the transloca-tion cycle. However, as the ring can be seen to beincomplete when the RNP is attached, we favourthe view that the binding of the RNP particle elicitsring formation, and that the cycle comprises all thestages indicated in Figure 4.
Electron tomographic analysis of the transloca-tion of the RNP particle has shown that the 5' endof the RNP particle is in the lead through the NPC(Mehlin et al., 1992). This result implies that theglobular particle is positioned in a specific way infront of the central channel of the NPC prior to theentry of the unfolding ribbon into the channel. It isreasonable to assume that the basket structure isinvolved in this process. One possibility is that theproper positioning is a direct consequence of aspecific binding of the particle to the basket.Another possibility would be that the binding ofthe particle to the basket is less specific, but theparticle is held by the basket in front of the entranceto the central channel and a specific recognition canthen take place between the 5' end of the particle(perhaps cap structure (Hamm & Mattaj, 1990;Jarmolowski et al., 1994) and/or cap-bindingproteins (Izaurrald et al., 1995; Visa et al., 1996b)and one or more NPC components at the entranceto the channel).
How could the cyclic rearrangement of the basketstructure be accomplished? It was noted that theappearance of the terminal ring is accompanied by

Nuclear Pore RNP Export310
a shortening of the individual basket filaments(Figure 3, II and III; see also Figure 4b and c). Thiscould imply that the distal ends of the eightfilaments are rebuilt to form the terminal ring.These filaments ends may possibly contain aprotein called NUP 153 (Cordes et al., 1993;Sukegawe & Blobel, 1993; Pante et al., 1994), whichcould be one component in such a reorganization.It has been demonstrated in vitro that the basket canbe ‘‘opened’’ by depleting divalent cations (Jarnik& Aebi, 1991). Perhaps the remarkable dynamics ofthe basket structure is somehow mediated bydivalent ions in vivo but both the regulation andmechanics of the process remain enigmatic.
In conclusion, the observations presented herestrongly suggest that the NPC basket is involved inmediating BR mRNP export from the nucleus. Thebasket may well be a multifunctional structure, butits primary role is likely one of selecting mRNPs forexport and in docking the RNP particles in thecorrect orientation properly at the gate of thechannel to accomplish a directed translocationthrough the channel. It will be important toestablish whether a similar configurational changeof the basket will be noted, maybe with a reducedor minimal distortion of the basket ring, when thetranslocation of smaller mRNPs, the small nuclearRNPs and the ribosomal subunits are studied.
AcknowledgementsThe authors thank C. Akey for critical discussion of the
manuscript, S. Rutherford for printing the photographs,G. R. Bennion for assistance, and P. Chantry andE. Magnusson for the artwork in Figures 3 and 4. Theresearch was supported by the Wellcome Trust Foun-dation, the Cancer Research Campaign and the SwedishNatural Science Research Council, the Kjell and MartaBeijer Foundation, and the Knut and Alice WallenbergFoundation.
ReferencesAkey, C. W. (1992). The nuclear pore complex: a
macromolecular transporter. In Nuclear Trafficking(Feldherr, C. M., ed.), pp. 3–14, Academic Press, SanDiego.
Akey, C. W. (1995). Structural plasticity of the nuclearpore complex. J. Mol. Biol. 248, 273–293.
Allen, T. D., Goldberg, M. W., Pelling, C. & Solovei, I.(1994). Evolutionary conservation of basket structurein nuclear pore complexes, and their possible role inmRNA export. In Proc. MSA (Bailey, G. &Garrat-Reed, A. J., eds), pp. 12–13, San FranciscoPress, San Fransisco.
Andersson, K., Bjorkroth, B. & Daneholt, B. (1980). Thein situ structure of the active 75S RNA genes inBalbiani rings of Chironomus tentans. Exp. Cell Res.130, 313–326.
Cordes, V. C., Reidenbach, C. S., Kohler, A., Stuurman,N., Vandriel, R. & Franke, W. W. (1993). Intramolec-ular filaments containing a nuclear pore complexprotein. J. Cell Biol. 123, 1333–1344.
Davidson, D. H. (1986). Gene Activity in Early Develop-ment, 3rd edit., Academic Press, New York.
Dworetzky, S. I. & Feldherr, C. M. (1988). Translocationof RNA-coated gold particles through the nuclearpores of oocytes. J. Cell Biol. 106, 575–584.
Goldberg, M. W. & Allen, T. D. (1992). High resolu-tion scanning electron microscopy of thenuclear envelope: demonstration of a new regular,fibrous lattice attached to the baskets of thenucleoplasmic face of the nuclear pores. J. Cell Biol.119, 1429–1440.
Goldberg, M. W. & Allen, T. D. (1993). The nuclearpore complex: three dimensional surface struc-ture revealed by field emission, in-lens scanningelectron microscopy with underlying structurebeing incovered by proteolysis. J. Cell Sci. 106,261–274.
Goldberg, M. W. & Allen, T. D. (1995). Structural andfunctional organization of the nuclear envelope.Curr. Opin. Cell Biol. 7, 301–309.
Goldberg, M. W. & Allen, T. D. (1996). The nuclear porecomplex and lamina: three dimensional structuresand interactions determined by field emission in-lensscanning electron microscopy. J. Mol. Biol. 257,848–865.
Hamm, J. & Mattaj, I. W. (1990). Monoethylated capstructures facilitate RNA export from the nucleus.Cell, 63, 109–118.
Izaurralde, E., Lewis, J., Gamberi, Ch., Jarmolowski, A.,McGuigan, C. & Mattaj, I. W. (1995). A cap-bindingprotein complex mediating U snRNA export. Nature,376, 709–712.
Jarmolowski, A., Boelens, E., Izaurralde, E. & Mattaj,I. W. (1994). Nuclear export of different classes ofRNA is mediated by specific factors. J. Cell Biol. 124,627–635.
Jarnik, M. & Aebi, U. J. (1991). Toward a more complete3-D structure of the nuclear pore complex. Struct.Biol. 107, 291–308.
Mehlin, H. & Daneholt, B. (1993). The Balbiani ringparticle: a model for the assebly and export from thenucleus. Trends Cell Biol. 3, 443–447.
Mehlin, H., Skoglund, U. & Daneholt, B. (1991). Transportof Balbiani ring granules through nuclear pores inChironomus tentans. Exp. Cell Res. 193, 72–77.
Mehlin, H., Daneholt, B. & Skoglund, U. (1992).Translocation of a specific premessenger ribonucleo-protein particle through the nuclear pore studiedwith electron microscope tomography. Cell, 69,605–613.
Mehlin, H., Daneholt, B. & Skoglund, U. (1995).Structural interaction between the nuclear porecomplex and a specific translocating RNP particle.J. Cell Biol. 129, 1205–1216.
Olins, A. L., Olins, D. E. & Franke, W. W. (1980).Stereo-electron microscopy of the nucleoli, Balbianiring and endoplasmic reticulum in Chironomussalivary gland cells. Eur. J. Cell Biol. 22, 714–723.
Pante, N. & Aebi, U. (1995). Exploring nuclear porecomplex structure and function in molecular detail.J. Cell Sci. Suppl. 19, 1–11.
Pante, N., Basts, R., McMorrow, B. & Aebi, U. (1994).Interaction and three-dimensional localization of agroup of nuclear pore complex protein. J. Cell Biol.126, 603–617.
Ris, H. (1991). The 3D-structure of the nuclear porecomplex as seen by high voltage electron microscopyand high resolution low voltage scanning electronmicroscopy. EMSA Bull. 21, 54–56.

Nuclear Pore RNP Export 311
Skoglund, U., Andersson, K., Bjorkroth, B., Lamb, M. M.& Daneholt, B. (1983). Visualization of the formationand transport of a specific hnRNP particle. Cell, 34,847–855.
Skoglund, U., Andersson, K., Strandberg, B. & Daneholt,B. (1986). Three-dimensional structure of a specificpre-messenger RNP particle established by electronmicroscope tomography. Nature, 319, 560–564.
Stevens, B. J. & Swift, H. (1966). RNA transport fromnucleus to cytoplasm in Chironomus salivary glands.J. Cell Biol. 31, 55–77.
Sukegawe, J. & Bobel, G. (1993). A nuclear pore complexprotein that contains zinc, finger motifs binds DNA,and faces the nucleoplasm. Cell, 72, 29–38.
Visa, N., Alzhanova-Ericsson, A., Sun, X., Kiseleva, E.,Bjorkroth, B., Wurtz, T. & Daneholt, B. (1996a). Apre-mRNA binding protein accompanies the RNA
from the gene through the nuclear pores and into thepolysomes. Cell, 84, 253–264.
Visa, N., Izaurralde, E., Ferreira, J., Daneholt, B. & Mattaj,I. W. (1996b). A nuclear cap-binding complex bindsBalbiani ring pre-mRNA cotranscriptionally andaccompanies the ribonucleoprotein particle duringnuclear export. J. Cell Biol. 133, 5–14.
Wieslander, L. (1994). The Balbiany ring multigenefamily: coding repetitive sequences and evolution ofa tissue specific cell function. Prog. Nucl. Acids Res.Mol. Biol. 48, 275–313.
Wurtz, T., Kiseleva, E., Nacheva, G., Alzhanova-Ericsson,A. T., Rosen, A. & Daneholt, B. (1996). Identificationof two RNA-binding proteins in Balbiany ringpremessenger RNP granules and the presence ofthese proteins in specific subsets of hnRNP, particles.Mol. Cell. Biol. 16, 1425–1435.
Edited by J. Karn
(Received 20 March 1996; received in revised form 1 May 1996; accepted 15 May 1996)





























