Embed Size (px)
Citation preview

Current Biology Vol 22 No 10R400
6. Karlsson, K.A., Gall, A.J., Mohns, E.J.,Seelke, A.M., and Blumberg, M.S. (2005). Theneural substrates of infant sleep in rats. PLoSBiol. 3, e143.
7. Seelke, A.M., and Blumberg, M.S. (2008). Themicrostructure of active and quiet sleep ascortical delta activity emerges in infant rats.Sleep 31, 691–699.
8. Frank, M.G., and Heller, H.C. (2005).Unresolved issues in sleep ontogeny:a response to Blumberg et al. J. Sleep Res. 14,98–101.
9. Khazipov, R., Sirota, A., Leinekugel, X.,Holmes, G.L., Ben-Ari, Y., and Buzsaki, G.(2004). Early motor activity drives spindlebursts in the developing somatosensorycortex. Nature 432, 758–761.
10. Shaffery, J.P., Roffwarg, H.P., Speciale, S.G.,and Marks, G.A. (1999). Ponto-geniculo-occipital-wave suppression amplifieslateral geniculate nucleus cell-sizechanges in monocularly deprivedkittens. Brain Res. Dev. Brain Res. 114,109–119.
11. Aton, S.J., Seibt, J., Dumoulin, M., Jha, S.K.,Steinmetz, N., Coleman, T., Naidoo, N., andFrank, M.G. (2009). Mechanisms ofsleep-dependent consolidation of corticalplasticity. Neuron 61, 454–466.
12. Corner, M.A., Schade, J.P., Sedlacek, J.,Stoeckart, R., and Bot, A.P. (1967).Developmental patterns in the central nervoussystem of birds. I. Electrical activity in thecerebral hemisphere, optic lobe andcerebellum. Prog. Brain Res. 26, 145–192.
13. Speciale, S.G., Nowaczyk, T., and Jouvet, M.(1976). A longitudinal study of bioelectricactivity in the pre- and post-hatch chick. Dev.Psychobiol. 9, 539–547.
14. Kovach, J.K. (1970). Development andmechanisms of behavior in the chick embryoduring the last five days of incubation. J. Comp.Physiol. Psychol. 73, 392–406.
15. Dang-Vu, T.T., Schabus, M., Desseilles, M.,Sterpenich, V., Bonjean, M., and Maquet, P.(2010). Functional neuroimaging insights intothe physiology of human sleep. Sleep 33,1589–1603.
16. Morrison, A.R., and Pompeiano, O. (1970).Vestibular influences during sleep. VI.Vestibular control of autonomic functionsduring the rapid eye movements ofdesynchronized sleep. Arch. Ital. Biol. 108,154–180.
17. Lickliter, R. (2011). The integrated developmentof sensory organization. Clin. Perinatol. 38,591–603.
18. Graven, S.N. (2000). Sound and the developinginfant in the NICU: conclusions andrecommendations for care. J. Perinatol. 20,S88–93.
Avian Sleep Group, Max Planck Institute forOrnithology, Eberhard-Gwinner-Strasse,82319 Seewiesen, Germany.E-mail: [email protected]
DOI: 10.1016/j.cub.2012.03.036
RNA-Directed DNA Methylation:Getting a Grip on Mechanism
Small RNAs guide repressive chromatinmodifications to regions of the genomecontaining transposons and repeats. An Arabidopsis genetic screen revealsthat the guidancemachinery includes a novel ATPase complex that could act asa dynamic molecular gripper.
Judith Bender
In addition to genes and sequencesneeded to replicate chromosomes,eukaryotic genomes are riddled withpotentially destructive transposonsand transposon-derived repeats. Thetransposon agenda is to integratecopies of itself into new genomic sites,creating insertional mutations thatcan damage host cell genes. Eventransposon repeats that are notcompetent for movement can expressaberrant RNAs and proteins that saphost cell resources. A fundamentalmechanism by which eukaryotic cellsfight back against transposons is totarget them for chromatin-basedtranscriptional silencing. Abreakthrough in understanding howthe eukaryotic cell tells genomic ‘us’from ‘them’ came from studies in plantsand fission yeast showing that smallRNAs produced from transposon andrepeat sequences guide chromatinmodifications back to matchingsequences in the genome [1]. Morerecently, analogous pathways havebeen described for germline-generatedtransposon small RNAs in Drosophila
and mammals [2–4]. However, manyquestions remain about smallRNA-guided chromatin pathways,including how small RNAs areproduced, how the small RNAs areharnessed to detect matchingDNA sequences, and howchromatin-modifying enzymes arerecruited to the sites of detection.A new study in this issue of CurrentBiology [5], using a sensitive andcomprehensive genetic screeningsystem in Arabidopsis, has identifiedkey components of the guidancemachinery for small RNA-directed DNAmethylation of transposons andrepeats in the plant genome.
Factors previously recoveredfrom this genetic screening systeminclude RNA polymerase subunits,RNA-processing and -binding factors,a chromatin-remodeling protein, a DNAmethyltransferase, and an intriguingprotein called DMS3 [6–10]. DMS3contains a hinge dimerization domainsimilar to the hinge domains foundin structural maintenance ofchromosome (SMC) proteins thatcontrol chromosome organization, but,unlike other SMC proteins, DMS3 lacks
an ATPase domain [6]. In this newstudy, Lorkovic and colleagues presentthe most recent discovery from thegenetic screen, an ATPase DMS11 [5].They show that DMS11 interacts withDMS3 to constitute a complex withthe potential for driving a dynamiccomponent of the RNA-directed DNAmethylation machinery.Based on a combination of genetic
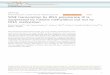
and biochemical approaches, thecurrent view is that RNA-directedDNA methylation involves tethering ofsmall RNA–protein complexes at thetarget region on the DNA by nascentnon-coding RNA transcripts that readthrough the target region (Figure 1) [11].Small RNA complexes couldbase-pair with nascent transcripts in asimilar interaction to the small RNAcomplex–messenger RNA interactionthat occurs during RNA interference, orthey could base-pair with unwoundsingle-stranded DNA in the transcribedregion. Small RNA complexes thenserve as platforms to recruit DNAmethyltransferases and otherchromatin-modifying factors to thetarget region. Plants have evolved anRNApolymerase variant—Pol V— thatis specifically dedicated to making thetethering transcripts for RNA-directedDNA methylation. How Pol V differsfrom RNA polymerase II to facilitatenon-coding transcription in DNAmethylated regions of the genomeis a key question in this field.For genetic dissection of the
small RNA-directed DNA methylationpathway, including Pol V function,

Pol V**
Current Biology
??
11ATPase
11ATPase
Hinge3
Hinge3
Figure 1. A mechanistic model for RNA-dependent DNA methylation.
RNA-dependent DNA methylation requirestranscription through the target region, medi-ated in plants by Pol V. DMS3, DMS11, andperhaps a partner protein (?) could form aring that promotes Pol V processivity by grip-ping together theRNA transcript and templateDNA. Asterisksmark potential targets for basepairing with a small RNA protein complex.
DispatchR401
Lorkovic and colleagues used atarget-plus-silencer double transgenesystem [5,6]. The target transgeneexpresses green fluorescent protein(GFP) in the growing shoot of the plant,and the silencer transgene expressessmall RNAs corresponding to thetarget promoter sequences. Whenthe two transgenes are combined, thetarget promoter region becomesDNA-methylated and transcriptionallysilenced, shutting off GFP. Throughmutations that reactivate GFPexpression, the double transgenescreening system has allowedidentification of eleven factors affectingevery step of the silencing pathway [5].Mutations in the DMS3 hinge domainprotein or the DMS11 ATPase reducepromoter DNA methylation on thetarget transgene, but do not affect thelevels of primary small RNAs producedby the silencer transgene [5,6].Furthermore, dms3 and dms11mutations prevent production of PolV-dependent transcripts atendogenous DNA methylation targetswhere such transcripts accumulate tohigh enough levels to be detectable[5,6,12]. Together, these findingsindicate that the DMS3–DMS11complex is required to promote Pol Vactivity. The dms11 mutation causesweak reactivation of GFP expressionrelative to dms3 and polVmutations [5].A potential explanation for the weakdms11 phenotype is that Arabidopsisencodes six DMS11-related ATPases,
at least some of which could act ina partially redundant manner withDMS11.
The DMS11 ATPase is in a distinctstructural class from the ATPasesfound in SMC hinge domain proteins[5]. However, the functionalorganization of the DMS11–DMS3complex with an ATPase at one endand a hinge domain at the other endis still analogous to the SMC hingedomain protein organization. SMChinge domain proteins form V-shapedheterodimers with the ATPase domainsat the ends of the V and the hinge–hingeinteraction at the point of the V [13,14].The ATPase ends of the SMC hingeheterodimers bind partner proteins toform ring structures that can encircleDNA strands. ATP hydrolysis providesa means to regulate the opening andthe closing of the ring. Pairs ofDMS11–DMS3 complexes could formanalogous dynamic ring structures thathold together DNA and/or RNA strandsduring Pol V transcription. For example,DMS11–DMS3 rings could holdnascent Pol V transcripts in associationwith the DNA template to promote Pol Vprocessivity, and to create a structurethat allows access of small RNAcomplexes to either unpaired RNAor DNA in the transcribed region(Figure 1). Whether DMS11–DMS3rings would include partner proteinsremains to be determined.
Lending support to the modelthat DMS11–DMS3 acts as a gripperfor RNA-directed DNA methylation,the structurally related mammalianATPase–hinge domain proteinSmcHD1 is required for maintainingDNA methylation on the silencedX-chromosome in femalemice — another process that involvesRNA–chromatin interactions [15].Compared with SmcHD1 and the SMChinge domain proteins, the separationof the ATPase and the hinge domainsinto two interacting proteins in theDMS11–DMS3 complex presents a newvariation on the theme of hinge domainprotein organization. This separationcould allow shuffling of DMS11-relatedATPase subunits for functionaldiversification. The discovery ofDMS11 and its interaction with DMS3in RNA-directed DNA methylationbroadens the spectrum of potentialATPase–hinge protein functions toinclude retention of nascentnon-coding transcripts on templateDNA at genomic regions where thesetranscripts serve as molecular tethers.
References1. Moazed, D. (2009). Small RNAs in
transcriptional gene silencing and genomedefence. Nature 457, 413–420.
2. Aravin, A.A., Sachidanandam, R., Girard, A.,Fejes-Toth, K., and Hannon, G.J. (2007).Developmentally regulated piRNA clustersimplicate MILI in transposon control. Science316, 744–747.
3. Kuramochi-Miyagawa, S., Watanabe, T.,Gotoh, K., Totoki, Y., Toyoda, A., Ikawa, M.,Asada, N., Kojima, K., Yamaguchi, Y., Ijiri, T.W.,et al. (2008). DNA methylation ofretrotransposon genes is regulated by Piwifamily members MILI and MIWI2 in murine fetaltestes. Genes Dev. 22, 908–917.
4. Wang, S.H., and Elgin, S.C. (2011). DrosophilaPiwi functions downstream of piRNAproduction mediating a chromatin-basedtransposon silencing mechanism in femalegerm line. Proc. Natl. Acad. Sci. USA 108,21164–21169.
5. Lorkovic, Z.J., Naumann, U., Matzke, A.J.M., andMatzke,M. (2012). InvolvementofaGHKLATPasein RNA-directed DNAmethylation in Arabidopsisthaliana. Curr. Biol. 22, 933–938.
6. Kanno, T., Bucher, E., Daxinger, L., Huettel, B.,Bohmdorfer, G., Gregor, W., Kreil, D.P.,Matzke, M., and Matzke, A.J. (2008). Astructural-maintenance-of-chromosomes hingedomain-containing protein is required forRNA-directed DNA methylation. Nat. Genet. 40,670–675.
7. Daxinger, L., Kanno, T., Bucher, E., van derWinden, J., Naumann, U., Matzke, A.J., andMatzke, M. (2009). A stepwise pathway forbiogenesis of 24-nt secondary siRNAs andspreading of DNA methylation. EMBO J. 28,48–57.
8. Gao, Z., Liu, H.L., Daxinger, L., Pontes, O.,He, X., Qian, W., Lin, H., Xie, M., Lorkovic, Z.J.,Zhang, S., et al. (2010). An RNA polymeraseII- and AGO4-associated protein acts inRNA-directed DNA methylation. Nature 465,106–109.
9. Eun, C., Lorkovic, Z.J., Naumann, U., Long, Q.,Havecker, E.R., Simon, S.A., Meyers, B.C.,Matzke, A.J., and Matzke, M. (2011). AGO6functions in RNA-mediated transcriptionalgene silencing in shoot and root meristems inArabidopsis thaliana. PLoS One 6, e25730.
10. Naumann, U., Daxinger, L., Kanno, T., Eun, C.,Long, Q., Lorkovic, Z.J., Matzke, M., andMatzke, A.J. (2011). Genetic evidence that DNAmethyltransferase DRM2 has a direct catalyticrole in RNA-directed DNA methylation inArabidopsis thaliana. Genetics 187, 977–979.
11. Haag, J.R., and Pikaard, C.S. (2011).Multisubunit RNA polymerases IV and V:purveyors of non-coding RNA for plantgene silencing. Nat. Rev. Mol. Cell Biol. 12,483–492.
12. Wierzbicki, A.T., Ream, T.S., Haag, J.R., andPikaard, C.S. (2009). RNA polymerase Vtranscription guides ARGONAUTE4 tochromatin. Nat. Genet. 41, 630–634.
13. Nasmyth, K., and Haering, C.H. (2009). Cohesin:its roles and mechanisms. Annu. Rev. Genet.43, 525–558.
14. Wood, A.J., Severson, A.F., and Meyer, B.J.(2010). Condensin and cohesin complexity: theexpanding repertoire of functions. Nat. Rev.Genet. 11, 391–404.
15. Blewitt, M.E., Gendrel, A.V., Pang, Z.,Sparrow, D.B., Whitelaw, N., Craig, J.M.,Apedaile, A., Hilton, D.J., Dunwoodie, S.L.,Brockdorff, N., et al. (2008). SmcHD1,containing a structural-maintenance-of-chromosomes hinge domain, has a critical rolein X inactivation. Nat. Genet. 40, 663–669.
Brown University, MCB Department, 185Meeting St, Providence, RI 02912, USA.E-mail: [email protected]
DOI: 10.1016/j.cub.2012.04.010










![The Fascinating World of RNA Interference · 2009. 1. 21. · dopsis, AGO4 plays a crucial role in generating rasi-RNAs that participate in RNA-directed DNA methylation [37], AGO6](https://img.pdfslide.us/doc/110x75/60c5beb91a11df3b4f5d8c3c/the-fascinating-world-of-rna-interference-2009-1-21-dopsis-ago4-plays-a-crucial.jpg)

![RNA N6-methyladenosine reader IGF2BP3 regulates cell ......RNA methylation modification information and partici-pate in the translation and degradation of downstream RNA [4]. Colon](https://img.pdfslide.us/doc/110x75/60e95ed35f0c0378245d0e57/rna-n6-methyladenosine-reader-igf2bp3-regulates-cell-rna-methylation-modification.jpg)















