Embed Size (px)
Citation preview

R.E. Brummett.1 G.G. Teugels2
1 The WorldFish Centre (formerly ICLARM), BP 2008, Yaoundé, Cameroon
[email protected] (corresponding author)
2 Royal Museum for Central Africa, B-3080 Tervuren, Belgium [email protected]
RIVERS OF THE LOWER GUINEANRAINFOREST: BIOGEOGRAPHY AND
SUSTAINABLE EXPLOITATION
ABSTRACT
The Lower Guinean rainforest of Southern Cameroon, con-tinental Equatorial Guinea, Gabon and the People’s Republic of theCongo and its associated biodiversity is being destroyed at a rate of1 million ha per year by poorly regulated timber exploitation andslash and burn agriculture. An important component of the rainfor-est is the river that drains it. Although very little studied and poor-ly understood, these rivers drain over 500 000 km2 and have beenestimated to contain at least 500 fish species, of which a large per-centage may be endemic. In the process of deforestation, the fishare being destroyed along with the trees and other wildlife.
149
Keywords: rainforest rivers, Central Africa,fish biodiversity, sustainable management

The rivers and swamps of the Lower Guineanforest comprise the Lower Guinean ichthyologicalprovince and possess different species from those ofthe Sudano-Nilotic province to the north and theCongo province to the East and South. The icthyofau-na of these rivers is dominated by the Siluriformes (6families, 23 genera, 102 species), the Characiformes (2families, 20 genera, 62 species), the Cichlidae (17 gen-era, 54 species) the Cyprinidae (10 genera, 79 species)and the Mormyridae (14 genera, 49 species). Amongthese are a large number of ornamentals, many ofwhich are rare and unusual, fetching high prices inEurope and North America.
The 8 million people who live in the LowerGuinean forest depend heavily upon the integrity of theriver ecosystem for their livelihoods. Estimates fromCameroon put the productivity of capture fisheries inforest river basins at 1.1 tons km-2 yr-1. Extrapolated tothe entire Lower Guinean forest, this translates into acash value of over US$1.4 thousand million per year,more than twice the value of all other non-timber for-est products combined.
Increasing population and poverty, coupledwith false valuations of rainforest biodiversity, haveled to unregulated logging, habitat destruction andover-exploitation. In addition, fishing rainforest riversincreasingly involves the use of poisons that are high-ly destructive of the entire food web. New and diversenatural resource management and exploitation strate-gies are needed to add value to rainforest river ecosys-tems to justify their preservation and improve thelivelihoods of rainforest communities.
INTRODUCTION
The Lower Guinean ichthyological province(Figure 1) extends in an arc along the NE corner of theGulf of Guinea from the Cross River in the NW to justshort of the Congo in the SE and includes some 50major and minor rivers (Table 1). It is sandwiched in-between the Nilo-Sudan and Congo provinces. To thewest and north, the Cross and Sanaga Rivers form theboundary with the Nilo-Sudan fauna. To the east andsouth, lies the Congo basin, separated from the LowerGuinean by a series of highlands, terminating with theChaillu Mountains in the PR Congo. The river systemsof the Lower Guinean province drain over 500 000 km2
of tropical rainforest (Mahé and Olivry 1999), formingan integral part of the rainforest ecosystem.
150 Rivers of the lower Guinean rainforest:
Figure 1. Ichthyological provinces of Africa, based on Roberts(1975) as modified by Lévéque (1997) and redrawn accordingto new hydrological basin mapping published by FAO (2000). 1= Maghreb, 2 = Nilo-Sudan, 3 = Upper Guinea, 4 = LowerGuinea, 5 = Congo Basin, 6 = Quanza, 7 = Zambezi, 8 = EastCoast, 9 = Southern, 10 = Malagasy.
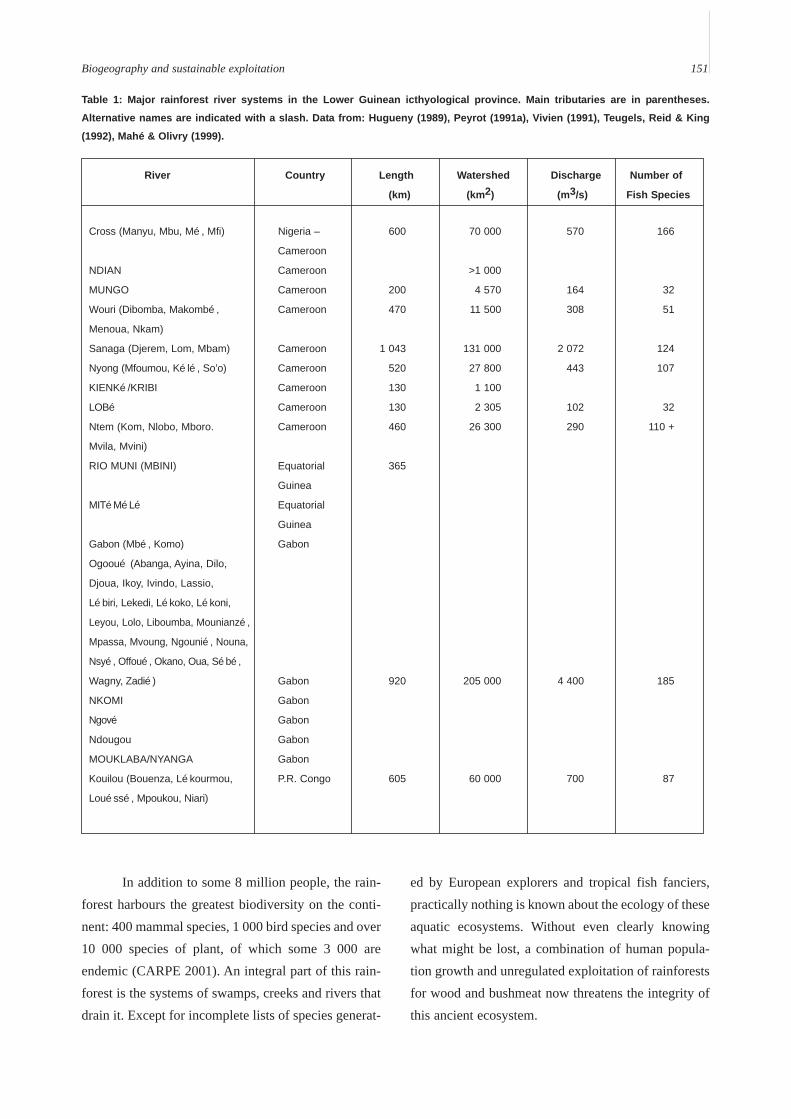
Table 1: Major rainforest river systems in the Lower Guinean icthyological province. Main tributaries are in parentheses.
Alternative names are indicated with a slash. Data from: Hugueny (1989), Peyrot (1991a), Vivien (1991), Teugels, Reid & King
(1992), Mahé & Olivry (1999).
River Country Length Watershed Discharge Number of
(km) (km2) (m3/s) Fish Species
Cross (Manyu, Mbu, Mé, Mfi) Nigeria – 600 70 000 570 166
Cameroon
NDIAN Cameroon >1 000
MUNGO Cameroon 200 4 570 164 32
Wouri (Dibomba, Makombé, Cameroon 470 11 500 308 51
Menoua, Nkam)
Sanaga (Djerem, Lom, Mbam) Cameroon 1 043 131 000 2 072 124
Nyong (Mfoumou, Kélé, So’o) Cameroon 520 27 800 443 107
KIENKé/KRIBI Cameroon 130 1 100
LOBé Cameroon 130 2 305 102 32
Ntem (Kom, Nlobo, Mboro. Cameroon 460 26 300 290 110 +
Mvila, Mvini)
RIO MUNI (MBINI) Equatorial 365
Guinea
MITéMéLé Equatorial
Guinea
Gabon (Mbé, Komo) Gabon
Ogooué (Abanga, Ayina, Dilo,
Djoua, Ikoy, Ivindo, Lassio,
Lébiri, Lekedi, Lékoko, Lékoni,
Leyou, Lolo, Liboumba, Mounianzé,
Mpassa, Mvoung, Ngounié, Nouna,
Nsyé, Offoué, Okano, Oua, Sébé,
Wagny, Zadié) Gabon 920 205 000 4 400 185
NKOMI Gabon
Ngové Gabon
Ndougou Gabon
MOUKLABA/NYANGA Gabon
Kouilou (Bouenza, Lékourmou, P.R. Congo 605 60 000 700 87
Louéssé, Mpoukou, Niari)
In addition to some 8 million people, the rain-forest harbours the greatest biodiversity on the conti-nent: 400 mammal species, 1 000 bird species and over10 000 species of plant, of which some 3 000 areendemic (CARPE 2001). An integral part of this rain-forest is the systems of swamps, creeks and rivers thatdrain it. Except for incomplete lists of species generat-
ed by European explorers and tropical fish fanciers,practically nothing is known about the ecology of theseaquatic ecosystems. Without even clearly knowingwhat might be lost, a combination of human popula-tion growth and unregulated exploitation of rainforestsfor wood and bushmeat now threatens the integrity ofthis ancient ecosystem.
Biogeography and sustainable exploitation 151

MATERIALS AND METHODS
Since September 2000, WorldFish Centre hasbeen working with rainforest communities in theLower Guinean ichthyological province of SouthernCameroon. In partnership with the Institut deRecherche Agricole pour le Développment (IRAD) anumber of biological studies have been carried out onbiodiversity, reproductive seasonality, sexual matura-tion and feeding habits of the ichthyofauna of theNyong River. With the collaboration of the Ministerede l’Elévage, des Pêches et des Industries Animales deCameroun (MINEPIA), additional work has been doneon exploitation strategies and a needs assessment offishing communities on the Ntem River. This latterparticularly focused on the role of women in aquaticresource exploitation.
In addition to these academic studies, effortsare underway to organize fishing communities on theSo’o, Mungo (Moliwe) and Ntem Rivers in an effort toimprove the efficiency and sustainability of riverexploitation and management. Groups have beenformed by the villages themselves and these have actedas the interface between WorldFish Centre, theGovernment of Cameroon and the local population.These groups have identified ecotourism and exportingornamental fishes as high priority activities.
The ultimate goal of this work is to establishfunctional village-based monitoring and managementprograms that would ensure the sustainability of newand diversified natural resource exploitation. As back-ground to this effort, WorldFish Centre undertook anextensive survey of existing knowledge on the rainfor-est river ecosystem, its current uses and threats to itsintegrity. This paper reports the outcomes of thisresearch and uses the documented perceptions of cur-rent resource users within the province to identifyneeds and indicate the direction for further work.
RESULTS AND DISCUSSION
Unlike the uplifting and rifting that affectedeastern and southern Africa during the Miocene, theriver courses in central Africa are extremely ancient,having not been substantially disrupted since thePrecambrian (Beadle1981; Peyrot 1991a). Evidencereviewed by Lévêque (1997) seems to indicate that, atsome time prior to the Miocene, there was a muchgreater overlap in African fish distribution than is cur-rently the case. The Lower Guinean ichthyologicalprovince corresponds closely with the extent of humidforest refugia during the last dry phase of the conti-nent, 20 000 – 15 000 years bp (Maley 1987; Schwartz1991) and is similar to the distribution pattern of aquat-ic molluscs in the region (van Damme 1984). It seemslikely that a more broadly distributed group of archaictaxa related to the modern species in the Lower andUpper Guinean provinces were repeatedly and/or pro-gressively isolated during the several dry phases thatreduced the extent of rainforest between 70 000 yearsbp and the present (Lévêque 1997).
In general, the aquatic resources of the LowerGuinean province are “blackwater” rivers, with a meanpH between 5 and 6 and electrical conductivitybetween 20 and 30 µS/cm. Water temperature isalways between 20 and 30° C. The water in these riversis clear and tea-coloured as a result of the low dis-solved nutrient concentration, low light (due to nar-rowness of valleys, canopy cover and often cloudyskies) and the large amount of allochthonous vegeta-tive matter that falls or flushes into the water from thesurrounding forest (Welcomme and de Merona 1988).
In terms of hydrology, there are within theprovince two general types of river: 1) the tropicalsavannah type to the north (Cross, Mungo, Wouri,Sanaga), which have uni-modal discharge patterns(Figure 2a) and, 2) the 13 rivers that flow out of thepresent rainforest, which have a bimodal dischargepattern (Figure 2b). In general, the magnitude offluctuation is greater in the north (up to 8 m on the
152 Rivers of the lower Guinean rainforest:
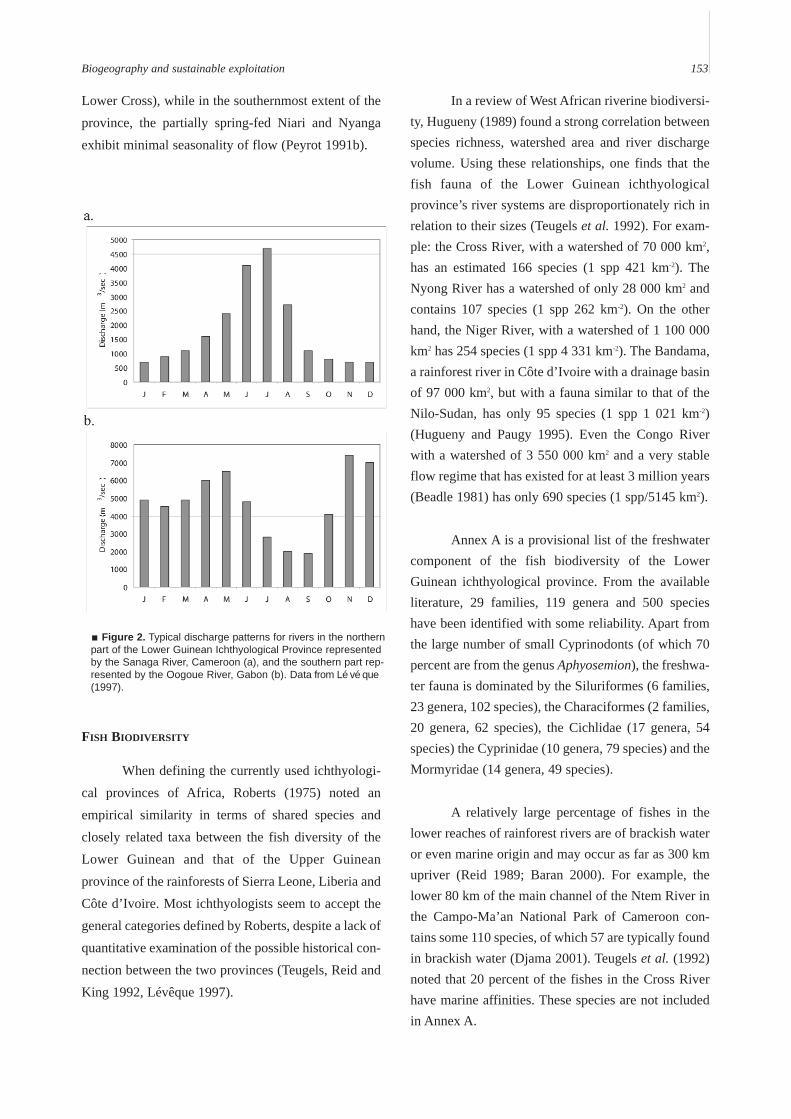
Lower Cross), while in the southernmost extent of theprovince, the partially spring-fed Niari and Nyangaexhibit minimal seasonality of flow (Peyrot 1991b).
FISH BIODIVERSITY
When defining the currently used ichthyologi-cal provinces of Africa, Roberts (1975) noted anempirical similarity in terms of shared species andclosely related taxa between the fish diversity of theLower Guinean and that of the Upper Guineanprovince of the rainforests of Sierra Leone, Liberia andCôte d’Ivoire. Most ichthyologists seem to accept thegeneral categories defined by Roberts, despite a lack ofquantitative examination of the possible historical con-nection between the two provinces (Teugels, Reid andKing 1992, Lévêque 1997).
In a review of West African riverine biodiversi-ty, Hugueny (1989) found a strong correlation betweenspecies richness, watershed area and river dischargevolume. Using these relationships, one finds that thefish fauna of the Lower Guinean ichthyologicalprovince’s river systems are disproportionately rich inrelation to their sizes (Teugels et al. 1992). For exam-ple: the Cross River, with a watershed of 70 000 km2,has an estimated 166 species (1 spp 421 km-2). TheNyong River has a watershed of only 28 000 km2 andcontains 107 species (1 spp 262 km-2). On the otherhand, the Niger River, with a watershed of 1 100 000km2 has 254 species (1 spp 4 331 km-2). The Bandama,a rainforest river in Côte d’Ivoire with a drainage basinof 97 000 km2, but with a fauna similar to that of theNilo-Sudan, has only 95 species (1 spp 1 021 km-2)(Hugueny and Paugy 1995). Even the Congo Riverwith a watershed of 3 550 000 km2 and a very stableflow regime that has existed for at least 3 million years(Beadle 1981) has only 690 species (1 spp/5145 km2).
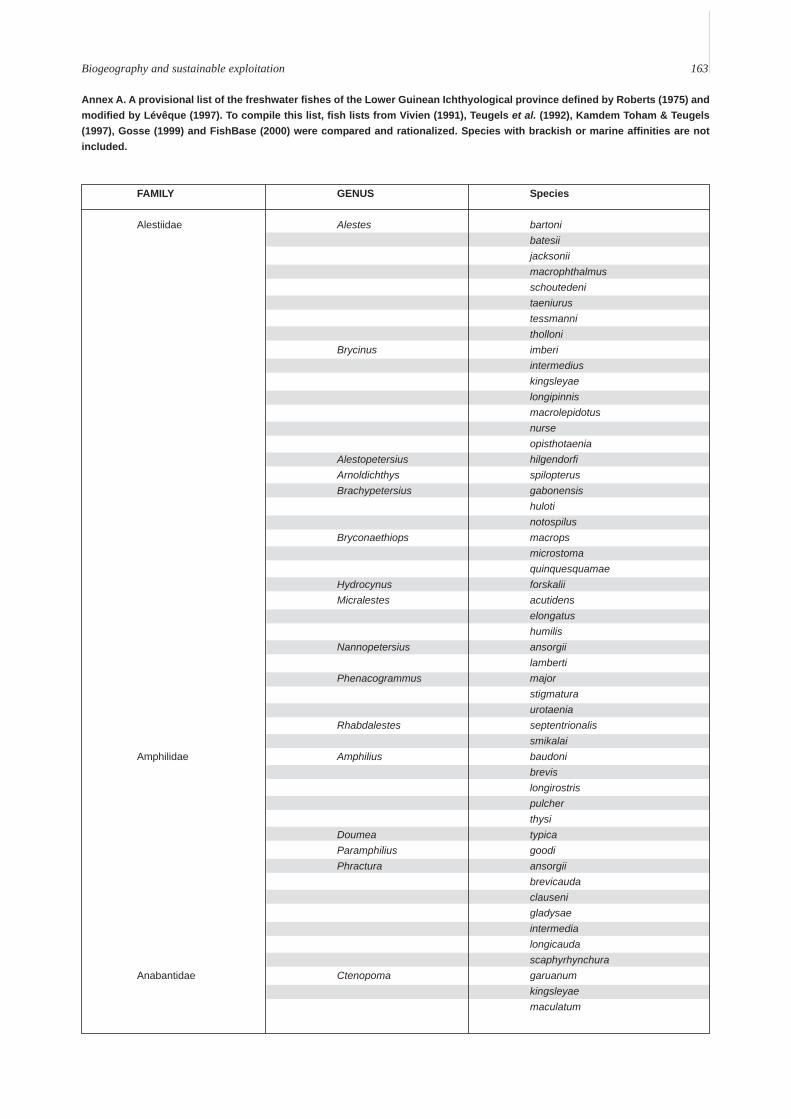
Annex A is a provisional list of the freshwatercomponent of the fish biodiversity of the LowerGuinean ichthyological province. From the availableliterature, 29 families, 119 genera and 500 specieshave been identified with some reliability. Apart fromthe large number of small Cyprinodonts (of which 70percent are from the genus Aphyosemion), the freshwa-ter fauna is dominated by the Siluriformes (6 families,23 genera, 102 species), the Characiformes (2 families,20 genera, 62 species), the Cichlidae (17 genera, 54species) the Cyprinidae (10 genera, 79 species) and theMormyridae (14 genera, 49 species).
A relatively large percentage of fishes in thelower reaches of rainforest rivers are of brackish wateror even marine origin and may occur as far as 300 kmupriver (Reid 1989; Baran 2000). For example, thelower 80 km of the main channel of the Ntem River inthe Campo-Ma’an National Park of Cameroon con-tains some 110 species, of which 57 are typically foundin brackish water (Djama 2001). Teugels et al. (1992)noted that 20 percent of the fishes in the Cross Riverhave marine affinities. These species are not includedin Annex A.
Biogeography and sustainable exploitation 153
Figure 2. Typical discharge patterns for rivers in the northernpart of the Lower Guinean Ichthyological Province representedby the Sanaga River, Cameroon (a), and the southern part rep-resented by the Oogoue River, Gabon (b). Data from Lévéque(1997).

Endemism in rainforest fishes seems to be rel-atively high (Teugels and Guégan 1994), although it isvery difficult from the scanty documentation to deter-mine exactly how many of the single reports for aspecies are due to endemism, lack of adequate distribu-tion data or simple misidentification (Stiassny 1996).In particular, the Cyprinodontiformes are prone toendemism, with some species occupying only a fewhundred square meters of bog, or an isolated creek(Welcomme and de Merona 1988). These small fishes,of which there are at least 100 species in the province,account for a substantial portion of the overall speciesrichness.
In addition, a number of fishes move up anddown the river according to their reproductive season-ality. Cyprinids and Citharinids, in particular membersof the genera Labeo and Distichodus, are reported byfishing communities in the Upper Cross and NtemRivers of Cameroon to undertake spawning runs dur-ing the latter part of the long rainy season (October-December) when rivers are swollen and marginalforests are flooded, providing cover and food for lar-vae and fry (Lowe-McConnell 1975; du Feu, 2001).The result of this is that species diversity measuredover the year changes substantially according to whichfishes are moving up or down stream at any particularpoint in time (Lowe-McConnell 1977).
The high fish species richness in the LowerGuinean province is probably the result of three mainfactors: 1) the relative stability of the hydrologicalregime in these rivers since the Eocene (compared tothe Nilo-Sudan province), 2) the highly sculpted natureof the watershed (compared to both the Congo andNilo-Sudan provinces) and, 3) the large number ofmicrohabitats created in rainforest rivers by the forestitself.
ECOLOGICAL ASSOCIATIONS
The forested nature of the watershed is themajor determinant of productivity and fish communitystructure in rainforest rivers. Stream width, depth, cur-rent velocity and substrate type have been identified ascritical in determining the spatial distribution of mostspecies (Lowe-McConnell 1975; Kamdem-Toham andTeugels 1997). These are all in one way or another,determined by the degree of canopy closure over theriver from the surrounding forest. The low primaryproductivity in rainforest water means that food websare mostly based on allochthonous plant materialsfrom the forest. The hydrological regime and watertemperature are directly influenced by the presence ofthe forest. The large amounts of dead wood influencedepth and current velocity and provide shelter frompredation, thus partitioning the stream and creating alarge number of microhabitats (Figure 3).
154 Rivers of the lower Guinean rainforest:
Figure 3. The So’o river, Cameroon in mid-April 2002 show-ing the large amounts of allochthonous wood that creates bothstructure and food producing surface in rainforest river ecosys-tems. Photo by the author.

Both species diversity and richness increase asone moves downstream from swamp, to first-order for-est stream (of which there are a particularly large num-ber in African rainforests) to medium-sized tributary tothe main channel, primarily through the addition ofspecies rather than through replacement (Géry 1965;Welcomme and de Merona 1988; Kamdem-Toham andTeugels 1998). Flooded swamp forest, either perma-nent or annual, is a typical feature of rainforest riverheadwaters. These contain very low dissolved oxygenand very high carbon dioxide concentrations (pH is inthe range of 4-5), but large quantities of allochthonousmaterials on a substrate of organic mud that generatesabundant food for those species with accessory breath-ing organs or resistance to very low oxygen concentra-tions. A large number of larval fishes that survive bybreathing from the air-water interface use the floodedforest as a nursery making this biotope particularlyimportant to overall ecosystem integrity.
First order rainforest streams are typically <5 mwide, <50 cm deep and are characterized by longstretches of shallow riffle interrupted by deeper, lower-velocity pools into which fish shelter during periodicdry spells when streams stop flowing. Relief in rain-forests tends to be low, so current velocity seldomexceeds 0.5 m/sec. Canopy closure ranges between 25and nearly 100 percent. Substrate is typically com-posed of leaf-covered sand or gravel.
Medium-sized streams are transitional zones(Lévêque 1997). As one proceeds downstream, theyfeature decreasing canopy closure, current velocity,allochthonous material and electrical conductivity andincreasing depth, fine sediment, large boulders, dis-solved oxygen and pH.
The main channel of rainforest rivers is themost stable biotope and offers the greatest range ofmicrohabitats (Welcomme and de Merona 1988).Citing Gosse’s (1963) work on the Yangambi portionof the Zaire River, which is broadly similar to Lower
Guinean ecosystems and shares a certain percentage oftheir biodiversity, Lowe-McConnell (1975) noted that,within the main channel, fish species richness andabundance are higher in shallow marginal waters alongbanks and islands than in mid-river. Gosse developed a“bank coefficient” that relates the length of water-bankcontact (including bays, islands, etc.) to species rich-ness. In these areas, sheltered from the main current,abundant aquatic vegetation representing a number ofgenera (Anubias, Crinum, Commelina, Limnophyton,Nympahea, et al.) creates habitats for a wide variety ofspecies and their offspring (Kamdem-Toham andTeugels 1998).
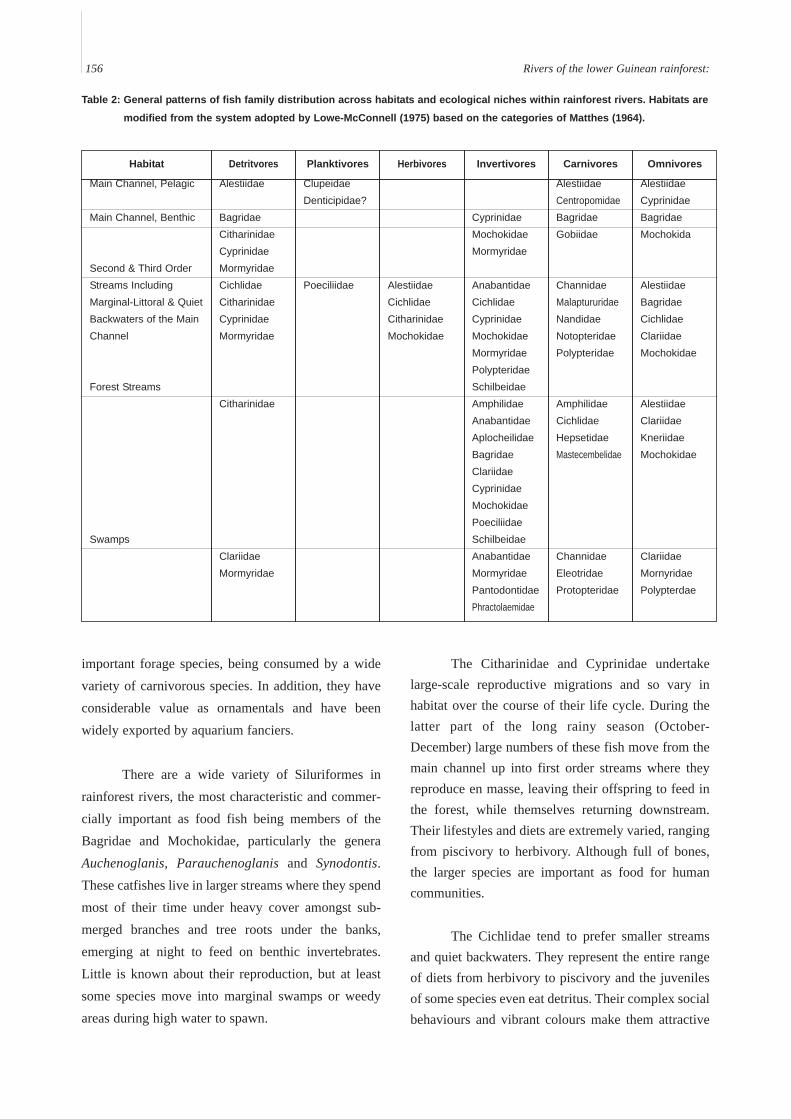
The nature of forest river food webs means thatmost species rely on carnivory or detritivory of onetype or another for survival and growth, planktivorybeing especially rare. Invertivores are the largest feed-ing guild in swamps and forest streams, whileomnivory and herbivory are more common as one pro-ceeds downstream. In general, fishes with highly spe-cialized diets are more common downstream due to thelarger number of specialized feeding niches (micro-habitats). Although the high degree of evolutionaryadaptation by fishes to the variety of rainforest riverhabitats means that for every family of fishes thereseem to be a species or life-history stage for everyhabitat, some general trends among family preferencesare evident (Table 2).
The Cyprinodontiformes of the genusAphyosemion are typical species of small foreststreams, often with very restricted distributions. Inrainforest rivers, they are associated with shallower rif-fles through woody debris, moderate velocity and aclosed canopy, abundant leaves on a gravel substrateand dense aquatic vegetation. They have two basicreproductive strategies, either laying eggs directly onor into the substrate, or laying adhesive eggs that stickto aquatic plants and individuals exhibit a certainamount of flexibility between the two (Sterba 1966).They eat mostly insect larvae. Cyprinodonts are
Biogeography and sustainable exploitation 155
156 Rivers of the lower Guinean rainforest:
important forage species, being consumed by a widevariety of carnivorous species. In addition, they haveconsiderable value as ornamentals and have beenwidely exported by aquarium fanciers.
There are a wide variety of Siluriformes inrainforest rivers, the most characteristic and commer-cially important as food fish being members of theBagridae and Mochokidae, particularly the generaAuchenoglanis, Parauchenoglanis and Synodontis.These catfishes live in larger streams where they spendmost of their time under heavy cover amongst sub-merged branches and tree roots under the banks,emerging at night to feed on benthic invertebrates.Little is known about their reproduction, but at leastsome species move into marginal swamps or weedyareas during high water to spawn.
The Citharinidae and Cyprinidae undertakelarge-scale reproductive migrations and so vary inhabitat over the course of their life cycle. During thelatter part of the long rainy season (October-December) large numbers of these fish move from themain channel up into first order streams where theyreproduce en masse, leaving their offspring to feed inthe forest, while themselves returning downstream.Their lifestyles and diets are extremely varied, rangingfrom piscivory to herbivory. Although full of bones,the larger species are important as food for humancommunities.
The Cichlidae tend to prefer smaller streamsand quiet backwaters. They represent the entire rangeof diets from herbivory to piscivory and the juvenilesof some species even eat detritus. Their complex socialbehaviours and vibrant colours make them attractive
Table 2: General patterns of fish family distribution across habitats and ecological niches within rainforest rivers. Habitats are
modified from the system adopted by Lowe-McConnell (1975) based on the categories of Matthes (1964).
Habitat
Main Channel, Pelagic
Main Channel, Benthic
Second & Third OrderStreams IncludingMarginal-Littoral & QuietBackwaters of the MainChannel
Forest Streams
Swamps
Detritvores
Alestiidae
BagridaeCitharinidaeCyprinidaeMormyridaeCichlidaeCitharinidaeCyprinidaeMormyridae
Citharinidae
ClariidaeMormyridae
ClupeidaeDenticipidae?
Poeciliidae AlestiidaeCichlidaeCitharinidaeMochokidae
CyprinidaeMochokidaeMormyridae
AnabantidaeCichlidaeCyprinidaeMochokidaeMormyridaePolypteridaeSchilbeidaeAmphilidaeAnabantidaeAplocheilidaeBagridaeClariidaeCyprinidaeMochokidaePoeciliidaeSchilbeidaeAnabantidaeMormyridaePantodontidaePhractolaemidae
AlestiidaeCentropomidaeBagridaeGobiidae
ChannidaeMalaptururidaeNandidaeNotopteridaePolypteridae
AmphilidaeCichlidaeHepsetidaeMastecembelidae
ChannidaeEleotridaeProtopteridae
AlestiidaeCyprinidaeBagridaeMochokida
AlestiidaeBagridaeCichlidaeClariidaeMochokidae
AlestiidaeClariidaeKneriidaeMochokidae
ClariidaeMornyridaePolypterdae
Planktivores Herbivores Invertivores Carnivores Omnivores

aquarium fish, but most species are small and they arenot common in local fish markets. Cichlids breed yearround in shallow marginal areas where the majorityprovide substantial levels of parental care.
The Mormyridae are well adapted to the varietyof habitats available in forest rivers and exist in mostof them, but the majority are found in second orderstreams of moderate depth and current. While someMormyrids are diurnal shoal feeders, most are noctur-nal insectivores probing about in the sediments for lar-val forms and using their electrical generating abilityto navigate and identify conspecifics in the dark. Theyreproduce during high water periods when floodedswamps are available as nurseries. Mormyrids con-tribute substantially to the commercial and subsistencefisheries of the forest.
As in other river systems on other continents,juveniles and adults of many rainforest river speciesoccupy different habitats in order to avoid competitionand/or cannibalism. In general, adults tend to dominateareas with good foraging opportunities leaving small-er/younger individuals to shallower and swifter habi-tats where larger individuals cannot reach. Smallerspecies in general tend to be more specific in theirhabitat preference than are larger species (Kamdem-Toham and Teugels 1997).
CURRENT EXPLOITATION SYSTEMS
Welcomme (1976) estimated the total numberof first order rainforest streams in Africa at over 4 mil-lion with a combined total length equal to half of allwatercourses, making these the largest single riverineecosystem on the continent. Of the 8 million peoplewho live in the Lower Guinean rainforests, nearly 20percent are more or less fulltime fishers. Estimatesfrom Cameroon put the productivity of capture fish-eries in forest rivers basins at 1.1 tonnes km-2 yr-1 (duFeu 2001). Extrapolated to the entire Lower Guineanforest, this translates into a cash value of approximate-ly US$1.4 thousand million per year, more than twicethe value of all other non-timber forest products com-bined. Average fish consumption in Cameroonian rain-
forests is around 47 kg person-1 year-1, compared to 10kg for the general population (Obam 1992).
Fishing in rainforest rivers is severely con-strained by the large quantities of wood that accrue inthe streambed. By far the most common types of gearare passive set nets, traps and hook-lines of whichthere are a great variety in accordance with the diver-sity of the fish fauna. Also common, is a hook-and-linefishery dominated by small children and mainly target-ing immature cichlids.
Seasonal spawning migrations are reported fora number of species, (“most” according to Lowe-McConnell, 1975). Fishing communities have learnedto take advantage of these runs by constructing meshbarriers constructed of tree trunks and branches, boundtogether by vines and held in place by large stones(Figure 4). At the height of the rains, these structuresare submerged and gravid adults pass easily over them.After spawning and spending several months upstreamin flooded forests to forage, the adults once again headback downstream. However, by this time the water lev-els have declined and the fish find themselves trappedwhen they try to avoid the barrier. Juveniles apparent-ly pass through the mesh without problem.
Biogeography and sustainable exploitation 157
Figure 4. A traditional fish dam constructed with locally avail-able materials on the main channel of the So’o River, a tributaryof the Nyong in South-Central Cameroon. Fish migratingupstream at high water to feed in flooded forests and spawn canswim over the top of the dam. As water levels decline, returningadults are captured while juveniles pass through the mesh.

In Cameroon, a special type of reproductiveevent, known as the “dok” takes place during the longrains in October-November. Doks involving Labeobatesii and Distichodus spp. have been documented inthe Upper Cross and the Ntem, respectively. They typ-ically last no more than a few hours or days. Accordingto du Feu (2001), who interviewed fishers on theUpper Cross River in Cameroon, the village is alertedto the imminence of the spawning event by theupstream movement of fish. Two hours after the fishhave passed, the water turns white with milt, at whichtime the villagers set nets to block the return ofspawned out adults on their return downstream. Mendo the fishing with cast nets or even clubs, whilewomen clean and smoke the catch. Eggs are normallynot taken to ensure the continuation of the runs forfuture generations.
There are at least two traditional fisheries thatare allocated entirely to women. One involves the con-struction of small earthen dams across first order foreststreams during the dry season to capture smallChannids, Clariids and Mastecembelids. As water lev-els decline, the dams prevent fish from migratingdownstream. When the water gets low enough, thewomen wade in and bucket out the remainder, catchingthe fish by hand or with the help of baskets. This prac-tice is widespread in both the Lower Guinean andCongo ichthyological provinces and adds substantiallyto the protein intake of forest communities. Anotherfishery that is the exclusive domain of women is theuse of woven basket traps to catch the freshwaterprawn, Macrobrachium vollenhovenii (Figure 5).
Village leaders normally regulate access to afishery. Such management techniques as prohibitingthe collection of fish eggs during spawning runs andthe prohibition of certain gears are traditionallyenforced through the use of magic charms or “ju-ju”.Villagers are free to fish as long at they follow thebasic regulations. Visiting fishers, of which there areconsiderable numbers (an estimated 80 percent of fish-ers on Cameroonian rivers are of Malian or Nigerianorigin) must first seek permission of the village leader-ship and then pay a token fee, normally in the form ofpalm wine or a percentage of the catch.
Despite these traditional management systems,over-fishing has become an increasing problem as thehuman population grows and puts increasing pressureon resources. In addition, the use of fish poisons hasbecome increasingly frequent. Some of these are fromlocal plants and cause only temporary harm, but mostpoisoners now use Lindane or Gammelin 20, anorganochlorine insecticide used in cocoa productionand highly destructive of the entire food web. Humandeaths have been reported as a result of eating poi-soned fish. A recent survey conducted on the Ntem
158 Rivers of the lower Guinean rainforest:
Figure 5. Fishing for freshwater prawns (Macrobrachiumvollenhovenii) using woven basket traps is a traditional activityreserved exclusively for women in the Ntem River Basin.

River just upstream from the Campo-Ma’an ForestReserve in Southern Cameroon found that insecticidefishing had completely disrupted local aquatic ecosys-tems and had permitted the extension of the range ofthe electric catfish, Malapterurus electricus into thesmall rivers where they were previously not found.Because of the powerful shocks emitted by this fish,women have been forced to abandon their traditionaldam fishing in the area.
INTEGRATED WATERSHED MANAGEMENT
The productivity of rainforest river ecosystemsdepends upon maintaining the integrity of the entireseries of biotopes of which it consists. Without the for-est, there would be no material inputs that feed the fishin the lower reaches. Without the first and second orderrainforest streams, there would be no reproductivemigrations and, consequently, the number and diversi-ty of fish would be drastically reduced. Without themain channel system, the overall productivity of theriver would be seriously diminished.
Unfortunately, increasing population andpoverty, coupled with false valuations of rainforestbiodiversity have led to habitat destruction and over-exploitation (Stiassny 1996). The Congo Basin hasalready lost an estimated 46 percent of its rainforest tologging and conversion to agriculture and continues tolose forested watershed at an average rate of 7 percentper year (Revenga et al. 1998). In addition, theseforests are being harvested in a largely irresponsiblemanner that not only takes out the valuable timber, butalso crushes the under story, alters stream courses andincreases runoff and siltation. Roads, saw mills andother infrastructure associated with logging attractspeople into the forest, resulting in wholesale transfor-mation of the ecosystem (Burns 1972, Garman andMoring 1993).
Kamdem-Toham and Teugels (1999) describedthe changes that occur in and around the rainforestrivers in the Ntem River basin as a result of poorlymanaged logging operations:
Absence of forest canopy above streamsHeavy siltationAbundant primary production (algae)Uniform watercourse; absence of riffles;pools dominant habitat typeNo cover/shelter for fish
In terms of water quality, these changes in habi-tat resulted in large decreases in water clarity and dis-solved oxygen and large increases in temperature andconductivity. In undisturbed sites, water was clearbrown with a mean temperature of 23.5°, dissolvedoxygen between 2.5 and 4.2 mg l
-1(measured at noon)
and electrical conductivity between 20 and 30 υS cm-1.
In sites affected by logging, the water was cloudy witha mean temperature of 34°, dissolved oxygen of <1.0mg l
-1and average electrical conductivity of 48 υS cm
-1.
Changes of this magnitude can wreak havoc on aquat-ic life and may last for many years (Chutter 1969;Growns and Davis 1991).
Forestry management practices exist that couldsubstantially reduce the negative impacts of logging(Davies and Nelson 1994; Smith, Brown and Pope2001). An economic and social re-evaluation of rain-forest river fisheries in relation to timber exploitationmight encourage changes in current forest manage-ment policy (CARPE 2001). However, substantialwork needs to be done if the vested interests of politi-cians and logging companies are to be thwarted.
A first step is the generation of expert systemsthat can be used to monitor the status of aquaticecosystems as changes take place in the watershed.Several attempts have been made at the generation of aworkable Index of Biotic Integrity (IBI) such as thatused to track changes in temperate zone streams, butparameterisation has been a problem. The best effort todate in Central Africa is that of Kamdem-Toham andTeugels (1999), but gaps still exist. Existing datasetson aquatic biodiversity and ecology in Central Africaare weak, at best and this makes it very difficult todevelop quantitative tools (Lévêque 1997).
Biogeography and sustainable exploitation 159

160 Rivers of the lower Guinean rainforest:
Coupled with this valuation exercise should bethe development of improved management andexploitation strategies that could actually increase thevalue of aquatic ecosystems and justify their preserva-tion, while at the same time improving rural liveli-hoods. Forest river ecosystems are currently unman-aged and unregulated in any formal sense. TheDepartment of Fisheries in Cameroon does not evenhave a policy or planned program of work on riverineecosystems outside of a number of small dams (M.O.Baba, Director of Fisheries, personal communication,Yaoundé, April 2002). The most widely promotedmethod of increasing the productivity of aquaticecosystems in Central Africa is to increase fishingpressure through the introduction of subsidies onmotors and other fishing equipment and this withoutany clear idea as to the size of the resource or level ofcurrent exploitation.
While some increased pressure might be war-ranted in some areas, the upper limit for this strategy isprobably already in sight for most places. Careful reg-ulation of fishing gear and seasons based on scientificdata might be a more widely applicable strategy for
increasing catches of certain species in some rivers. Inaddition, integrated aquaculture in rainforest water-sheds could take advantage of abundant water suppliesand organic matter and might even be used in stock-enhancement or ranching where feasible or necessary.This might not be exclusively limited to the traditionalfood fishes. Species with value (both locally and inter-nationally) as ornamental aquarium fishes are unusual-ly abundant in rainforest rivers and fetch much higherprices per kilogram than food fish (Tlusty 2002).
Working with communities to both develop thetools and manage ecosystems might be worth trying(CARPE 2001). WorldFish Centre community basedmanagement of aquatic ecosystems program inBangladesh has produced positive outcomes. Otheragencies working in the rainforest, most notably theCentre for International Forestry Research (CIFOR)and the World Wildlife Fund (WWF) have had somesuccess in community forestry management and suchefforts need to be strengthened and broadened toinclude the most valuable non-timber forest product ofthem all: rainforest fishes.

REFERENCES
Baran E. 2000. Biodiversity of estuarine fish faunas inWest Africa. Naga, The WorldFish CentreQuarterly, 234: 4-9.
Beadle L.C. 1981. The inland waters of tropical Africa,second edition. London, Longman. 475 pp.
Burns J.W. 1972. Some effects of logging and associatedroad construction on northern Californiastreams. Transactions of the AmericanFisheries Society, 1011: 1-17.
CARPE. 2001. Congo Basin information series. CentralAfrican Regional Program for theEnvironment. Gland, Switzerland, WorldWildlife Fund.
Chutter F.M. 1969. The effects of silt and sand on theinvertebrate fauna of streams and rivers.Hydrobiologia, 341: 57-76.
Davies P.E. & M. Nelson. 1994. Relationships betweenriparian buffer widths and the effects of log-ging on stream habitat, invertebrate communi-ty composition and fish abundance. AustralianJournal of Marine and Freshwater Resources,45: 1289-1305.
Djama T. 2001. Inventaire qualitatif de poisons dansl’UTO Campo-Ma’an. Document 11. Projectd’Aménagement et de Conservation de laBiodiversité de Campo-Ma’an, Kribi,Cameroon.
Du Feu T.A. 2001. Fish and fisheries in the southern zoneof the Takamanda Forest Reserve, SouthwestCameroon. Consultant’s report to theCameroonian-German project: Protection ofForests around Akwaya PROFA, GTZ,Yaoundé, Cameroun.
FishBase. 2000. Electronic database on fishes.International Center for Living AquaticResources Management, Penang, Malaysia.
Garman G.C. & Moring J.R. 1993. Diet and annual pro-duction of two boreal river fishes followingclearcut logging. Environmental Biology ofFishes, 36: 301-311.
Géry J. 1965. Poissons du Basin d l’Invindo. BiologicaGabonica, 1: 375-393.
Gosse J.P. 1999. Document for the study of the zoogeog-raphy of African freshwater fishes. (http://www.naturalsciences.be/science/collec-tions/africanfish). Royal Brussels, BelgianInstitute of Natural Sciences.
Growns I.O. & Davis J.A. 1991. Comparison of themacroinvertebrate communities in streams inlogged and undisturbed catchments 8 yearsafter harvesting. Australian Journal of Marineand Freshwater Resources, 42: 689-706.
Hugueny B. 1989. West African rivers as biogeographicislands: Species richness of fish communities.Oecologia, 79: 236-243.
Hugueny B. & Paugy D. 1995. Unsaturated fish commu-nities in African rivers. American Naturalist,146: 162-169.
Kamdem-Toham A. &. Teugels G.G. 1997. Patterns ofmicrohabitat use among fourteen abundantfishes of the lower Ntem River BasinCameroon. Aquatic Living Resources, 10:289-298.
Kamdem-Toham A. & Teugels G.G. 1998. Diversity pat-terns of fish assemblages in the Lower NtemRiver Basin Cameroon, with notes on potentialeffects of deforestation. Archives ofHydrobiology, 1414: 421-446.
Kamdem-Toham A. & Teugels G.G. 1999. First data on anindex of biotic integrity (IBI) based on fishassemblages for the assessment of the impactof deforestation in a tropical West African riversystem. Hydrobiologia, 397: 29-38.
Lévêque C. 1997. Biodiversity dynamics and conserva-tion: The freshwater fishes of tropical Africa.Cambridge, UK, Cambridge University Press.
Lowe-McConnell R.H. 1975. Fish communities in tropi-cal freshwaters. London, Longman.
Lowe-McConnell R.H. 1977. Ecology of fishes in tropi-cal waters. London, Edward Arnold.
Biogeography and sustainable exploitation 161

Mahé G. & Olivry J.C. 1999. Les apports en eau douce àl’Atlantique depuis les côtes de l’Afriqueintertropicale. In Sciences de la terre et desplanets. C.R. Academie de Sciences de Paris,328: 621-626.
Maley J. 1987. Fragmentation de la forêt dense humide etextension des biotopes Montagnards au quater-naire récent. Palaeoecology of Africa, 18: 207-334.
Matthes H. 1964. Les poissons du Lac Tumba et de larégion d’Ikela. Etude systématique etécologique. Annales, Série 8, SciencesZoologiques 126. Terverun, Belgium, MuséeRoyal de l’Afrique Centrale.
Obam A. 1992. Conservation et mise en valeur de forêtsau Cameroun. Yaounde, Cameroun,Imprimerie National.
Peyrot R. 1991a. La géologie de l’Afrique centrale. In R.Lanfranchi & B. Clist, Aux Origines deL’Afrique Centrale. Centre International desCivilizations Bantu, Libreville, Gabon, CentreCulturels Français d’Agrique Centrale.
Peyrot R. 1991b. Hydrologie de l’Afrique centrale. In R.Lanfranchi & B. Clist, Aux Origines deL’Afrique Centrale, Centre International desCivilizations Bantu, Libreville. Gabon, CentreCulturels Français d’Agrique Centrale.
Reid G.M. 1989. The living waters of Korup rainforest: Ahydrobiological survey report and recommen-dations with emphasis on fish and fisheries.WWF Report 3206/A8:1. Gland, Switzerland,World Wildlife Fund.
Revenga C., Murray S., Abramovitz J. & Hammond A.,1998. Watersheds of the world: Ecologicalvalue and vulnerability. Washington, DC,World Resources Institute.
Roberts T.R., 1975. Geographical distribution of Africanfreshwater fishes. Zoological Journal of theLinnaean Society, 57: 249-319.
Schwartz D. 1991. Les paysages de l’Afrique centralependant le quaternaire. In R. Lanfranchi & B.Clist, Aux Origines de L’Afrique Centrale,Centre International des Civilizations Bantu,Libreville, Gabon, Centre Culturels Françaisd’Agrique Centrale.
Smith F., Brown A.V. & Pope M. 2001. Meiofauna inintermittent streams differ among watershedssubjected to five methods of timber harvest.Hydrobiologia, 464: 1-8.
Sterba G. 1966. Freshwater fishes of the world. London,Studio Vista Ltd.
Stiassny M.L.J. 1996. An overview of freshwater biodi-versity with some lessons from African fishes.Fisheries, 219: 7-13.
Teugels G.G. & Guégan J.F. 1994. Diversité biologiquedes poisons d’eaux douces de la Basse-Guinéeet de l’Afrique Centrale. In G.G. Teugels, J.-F.Guégan & J.-J. Albaret, eds. Diversitébiologique des poissons des eaux douces etsaumâtres d’Afrique. Annales SciencesZoologiques. Tervuren, Belgium, Musée Royalde L’Afrique Centrale. 275 pp.
Teugels G.G., Reid G. & King R.P. 1992. Fishes of theCross River Basin: Taxonomy, zoogeography,ecology and conservation. Annales SciencesZoologiques, Terverun, Belgium, MuséeRoyale de l’Afrique Centrale. 266 pp.
Tlusty M. 2002. The benefits and risks of aquaculturalproduction for the aquarium trade.Aquaculture, 205: 203-219.
Van Damme D. 1984. The freshwater mollusca of north-ern Africa. Developments in Hydrobiologia25. Dordrecht, The Netherlands, W. JunkPublishers.
Vivien J. 1991. Faune du Cameroun: Guide des mam-mifères et poissons. Yaoundé, Cameroun,GICAM et Ministère de la Coopération et duDéveloppement.
Welcomme R.L. 1976. Some general and theoretical con-siderations on the fish yield of African rivers.Journal of Fish Biology, 8: 351-364.
Welcomme R.L. & de Merona B. 1988. Fish communitiesin rivers. In C. Leveque, M.N. Bruton & G.W.Sentongo, eds. Biology and Ecology ofAfrican Freshwater Fishes, Collection Travauxet Documents. Paris, Editions de l’ORSTOM.216 pp.
162 Rivers of the lower Guinean rainforest:
Biogeography and sustainable exploitation 163
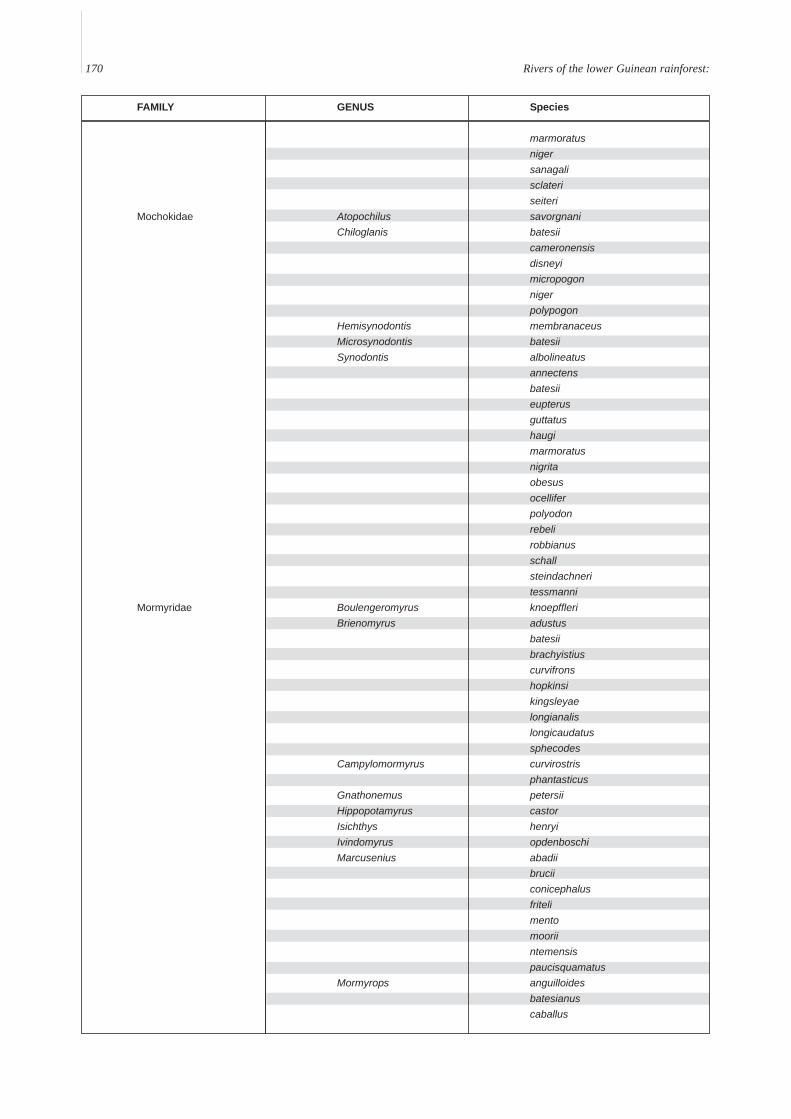
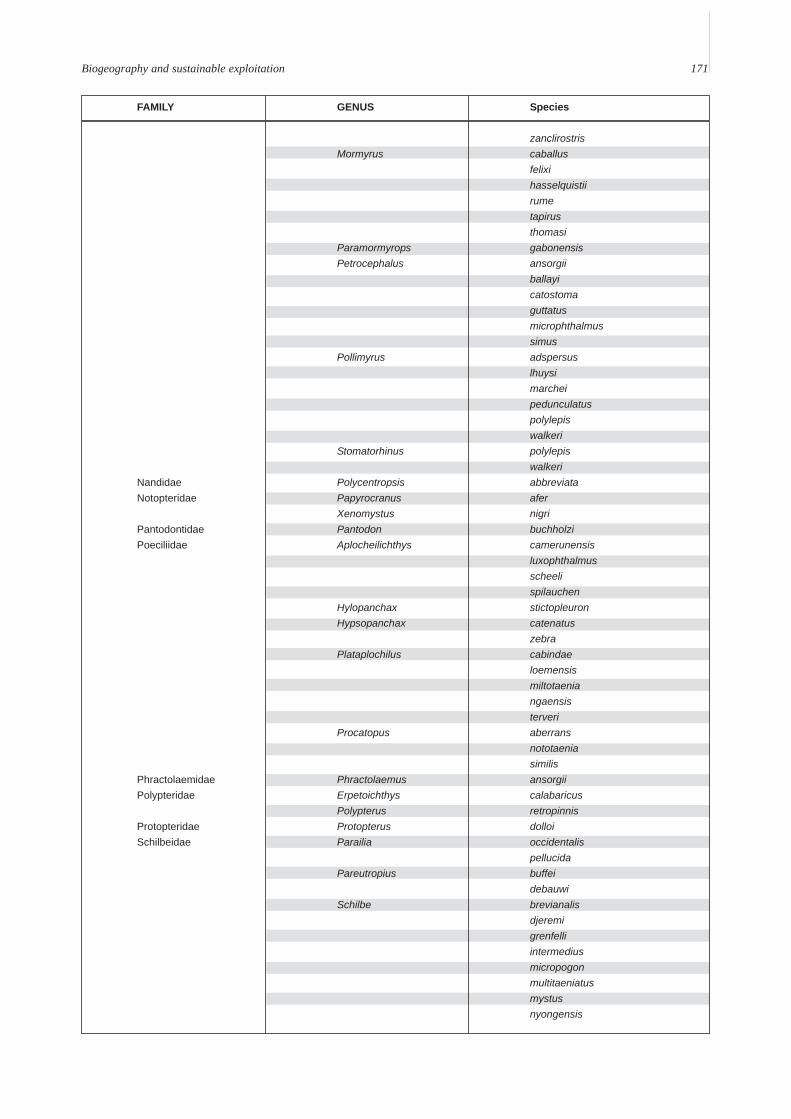
Annex A. A provisional list of the freshwater fishes of the Lower Guinean Ichthyological province defined by Roberts (1975) andmodified by Lévêque (1997). To compile this list, fish lists from Vivien (1991), Teugels et al. (1992), Kamdem Toham & Teugels(1997), Gosse (1999) and FishBase (2000) were compared and rationalized. Species with brackish or marine affinities are notincluded.
FAMILY GENUS Species
Alestiidae Alestes bartonibatesiijacksoniimacrophthalmusschoutedenitaeniurustessmannitholloni
Brycinus imberiintermediuskingsleyaelongipinnismacrolepidotusnurseopisthotaenia
Alestopetersius hilgendorfiArnoldichthys spilopterusBrachypetersius gabonensis
hulotinotospilus
Bryconaethiops macropsmicrostomaquinquesquamae
Hydrocynus forskaliiMicralestes acutidens
elongatushumilis
Nannopetersius ansorgiilamberti
Phenacogrammus majorstigmaturaurotaenia
Rhabdalestes septentrionalissmikalai
Amphilidae Amphilius baudonibrevislongirostrispulcherthysi
Doumea typicaParamphilius goodiPhractura ansorgii
brevicaudaclausenigladysaeintermedialongicaudascaphyrhynchura
Anabantidae Ctenopoma garuanumkingsleyaemaculatum

164 Rivers of the lower Guinean rainforest:
FAMILY GENUS Species
nebulosumnigropannosumpetherici
Microctenopoma nanumAplocheilidae Aphyosemion abacinum
ahliamietiamoenumarnoldiaureumaustraleavichangbamilekorumbatesiibitaeniatumbivittatumbualanumbuytaerticalliurumcameronenseceliaecinnamomeumcitreinpinniscoelestecyanostictumdargeiedeanumelbertiescherichiexigoideumexiguumfallaxfranzwernerifulgensgabunensegardnerigeorgiaehanneloreaeheinemanniheraherzogihofmannijorgenscheelikekemensekouamenselambertiloennbergiilouessenselugensmaculatummarmoratummimodonmirabilendianumocellatumoeseriogoense

Biogeography and sustainable exploitation 165
FAMILY GENUS Species
ottogartneripaschenipassaroiprimigeniumpuerzlipunctatumpyrophoreraddairectogoenseriggenbachirobertsonirubrolabialeschluppiseegersisjoestedtisplendidumsplendopleurestriatumthysitirbakivolcanumwachtersizygaima
Diapteron abacinumcyanostictumfulgensgeorgiae
Epiplatys berkenkampicallipteronesekanusgrahamihuberiinfrafasciatusneumannisangmelinensissexfasciatussinga
Bagridae Anaspidoglanis ansorgiiboutchangaimacrostoma
Auchenoglanis ahliballayiguiralilongicepsmonkeipantherinuspietschmanni
Chrysichthys aluuensisauratusdagetifilamentosusfurcatusnigrodigitatusogooensispersimilislongidorsalisnigrodigitatus
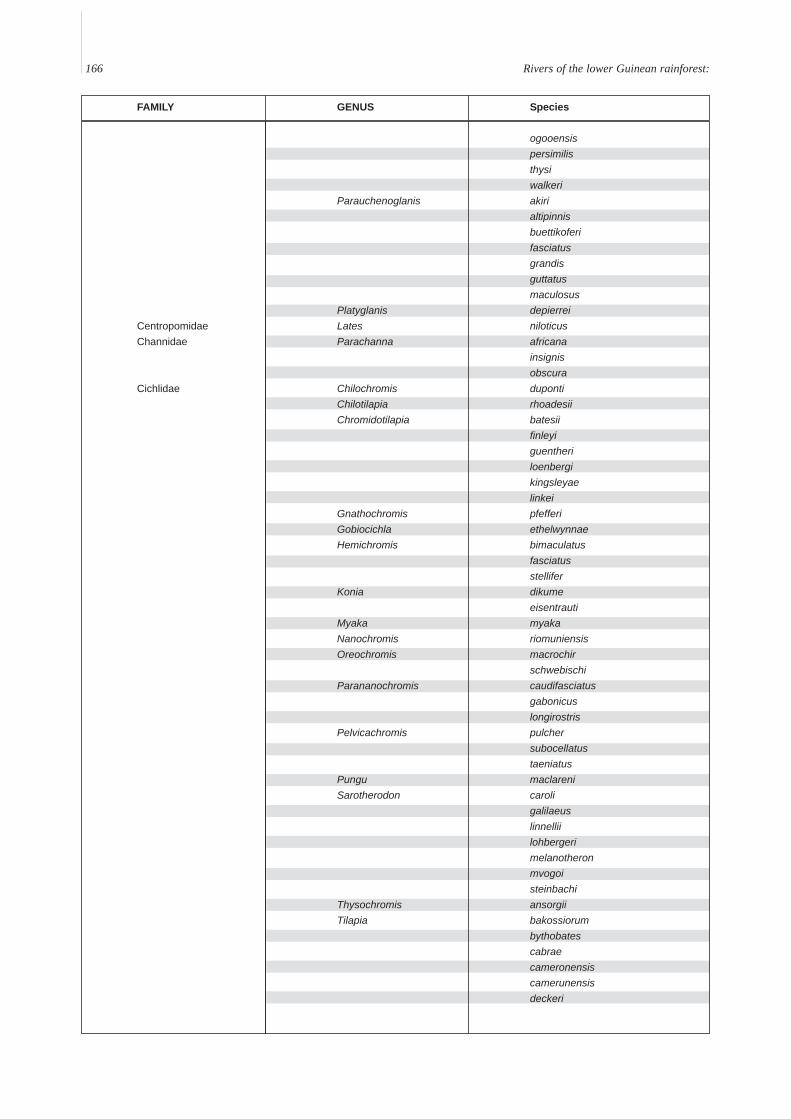
166 Rivers of the lower Guinean rainforest:
FAMILY GENUS Species
ogooensispersimilisthysiwalkeri
Parauchenoglanis akirialtipinnisbuettikoferifasciatusgrandisguttatusmaculosus
Platyglanis depierreiCentropomidae Lates niloticusChannidae Parachanna africana
insignisobscura
Cichlidae Chilochromis dupontiChilotilapia rhoadesiiChromidotilapia batesii
finleyiguentheriloenbergikingsleyaelinkei
Gnathochromis pfefferiGobiocichla ethelwynnaeHemichromis bimaculatus
fasciatusstellifer
Konia dikumeeisentrauti
Myaka myakaNanochromis riomuniensisOreochromis macrochir
schwebischiParananochromis caudifasciatus
gabonicuslongirostris
Pelvicachromis pulchersubocellatustaeniatus
Pungu maclareniSarotherodon caroli
galilaeuslinnelliilohbergerimelanotheronmvogoisteinbachi
Thysochromis ansorgiiTilapia bakossiorum
bythobatescabraecameronensiscamerunensisdeckeri
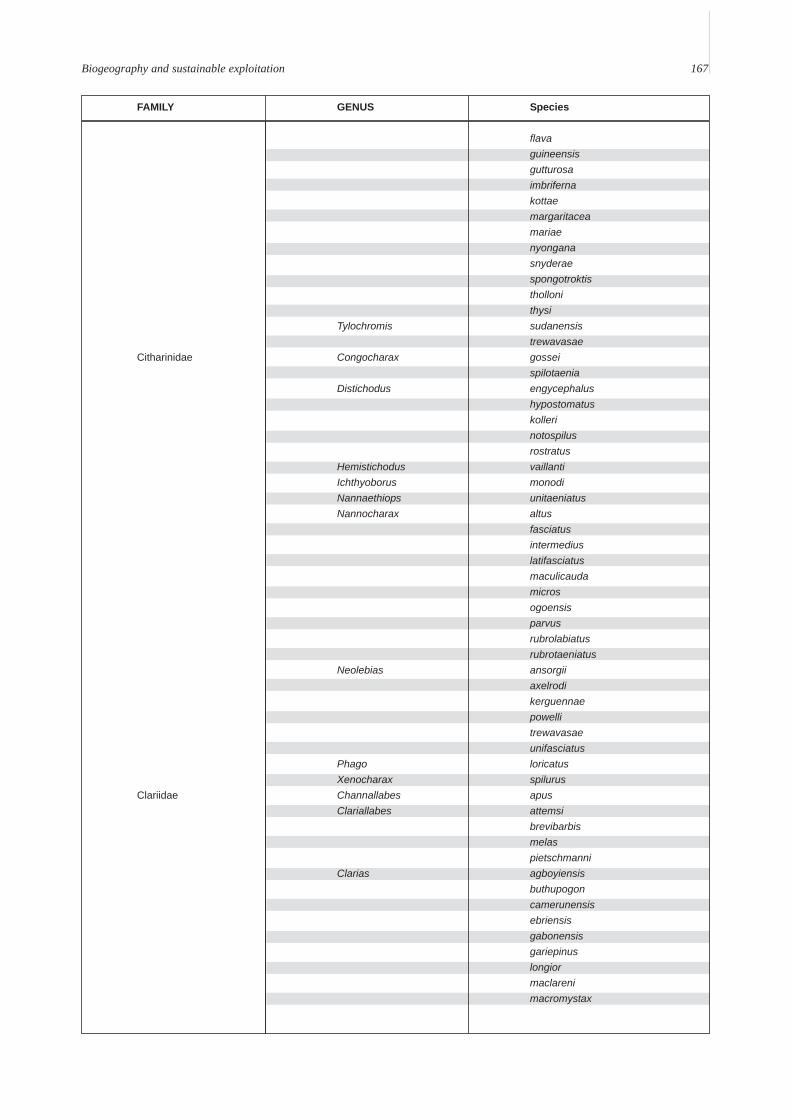
Biogeography and sustainable exploitation 167
FAMILY GENUS Species
flavaguineensisgutturosaimbrifernakottaemargaritaceamariaenyonganasnyderaespongotroktisthollonithysi
Tylochromis sudanensistrewavasae
Citharinidae Congocharax gosseispilotaenia
Distichodus engycephalushypostomatuskollerinotospilusrostratus
Hemistichodus vaillantiIchthyoborus monodiNannaethiops unitaeniatusNannocharax altus
fasciatusintermediuslatifasciatusmaculicaudamicrosogoensisparvusrubrolabiatusrubrotaeniatus
Neolebias ansorgiiaxelrodikerguennaepowellitrewavasaeunifasciatus
Phago loricatusXenocharax spilurus
Clariidae Channallabes apusClariallabes attemsi
brevibarbismelaspietschmanni
Clarias agboyiensisbuthupogoncamerunensisebriensisgabonensisgariepinuslongiormaclarenimacromystax
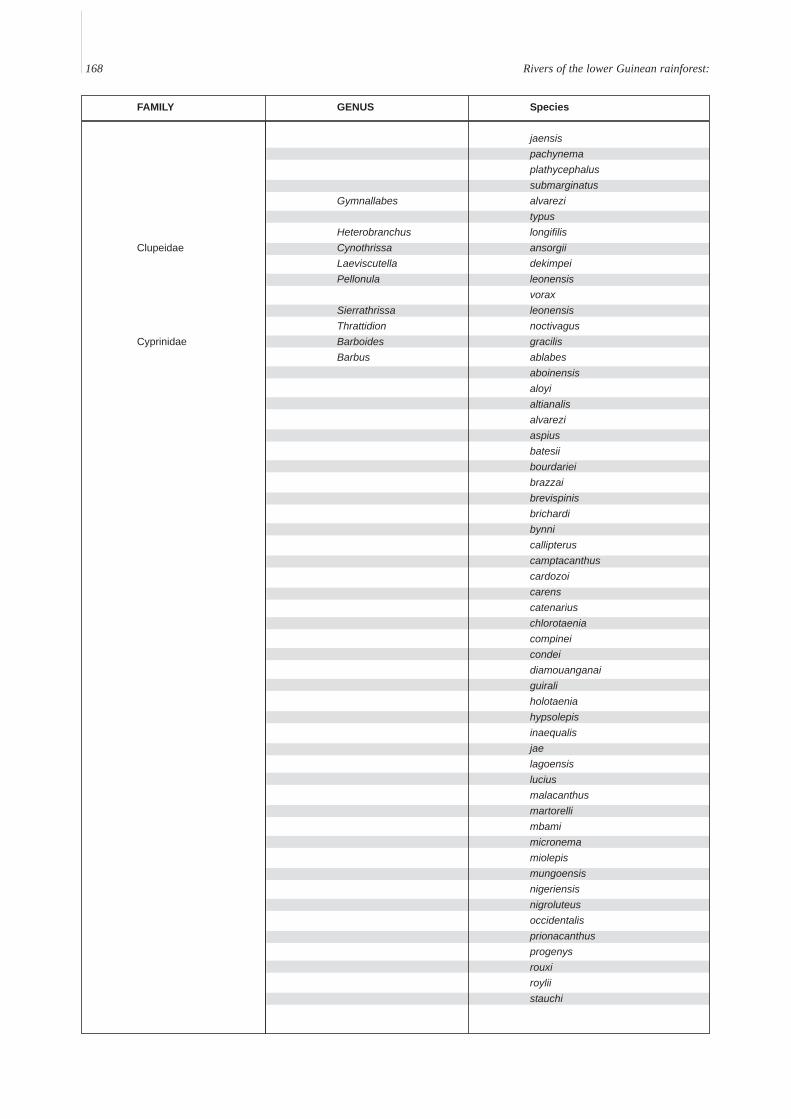
168 Rivers of the lower Guinean rainforest:
FAMILY GENUS Species
jaensispachynemaplathycephalussubmarginatus
Gymnallabes alvarezitypus
Heterobranchus longifilisClupeidae Cynothrissa ansorgii
Laeviscutella dekimpeiPellonula leonensis
voraxSierrathrissa leonensisThrattidion noctivagus
Cyprinidae Barboides gracilisBarbus ablabes
aboinensisaloyialtianalisalvareziaspiusbatesiibourdarieibrazzaibrevispinisbrichardibynnicallipteruscamptacanthuscardozoicarenscatenariuschlorotaeniacompineicondeidiamouanganaiguiraliholotaeniahypsolepisinaequalisjaelagoensisluciusmalacanthusmartorellimbamimicronemamiolepismungoensisnigeriensisnigroluteusoccidentalisprionacanthusprogenysrouxiroyliistauchi
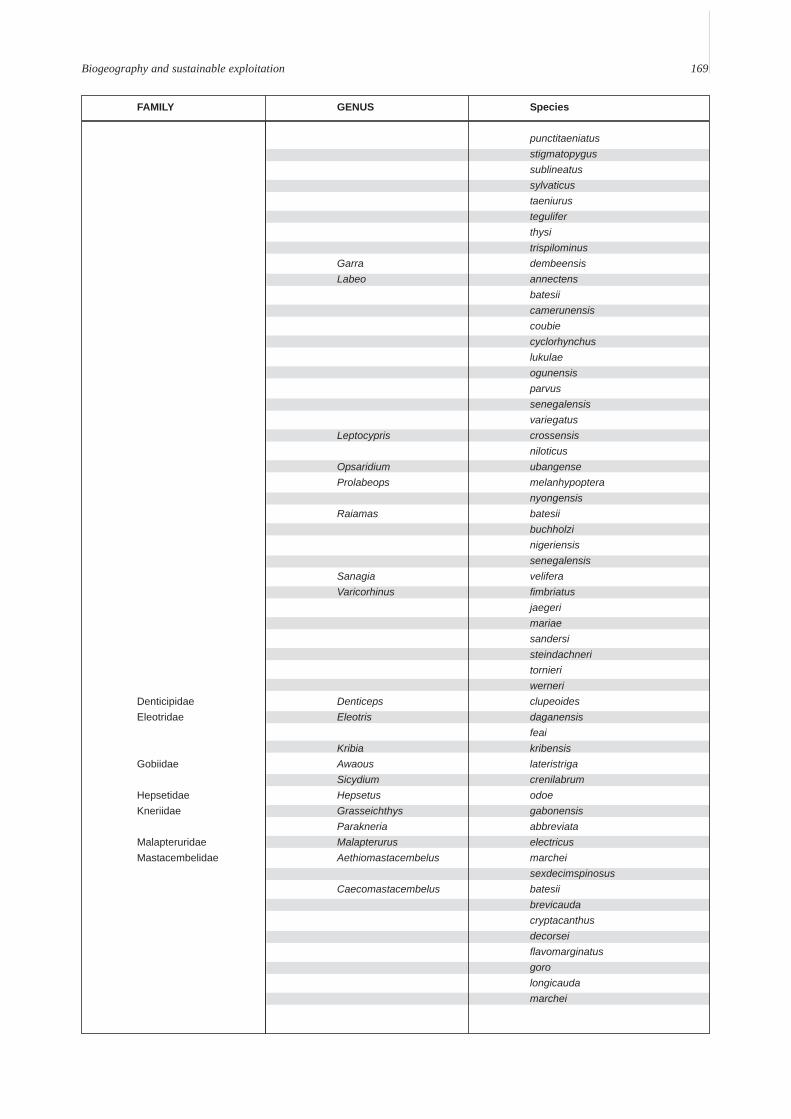
Biogeography and sustainable exploitation 169
FAMILY GENUS Species
punctitaeniatusstigmatopygussublineatussylvaticustaeniurusteguliferthysitrispilominus
Garra dembeensisLabeo annectens
batesiicamerunensiscoubiecyclorhynchuslukulaeogunensisparvussenegalensisvariegatus
Leptocypris crossensisniloticus
Opsaridium ubangenseProlabeops melanhypoptera
nyongensisRaiamas batesii
buchholzinigeriensissenegalensis
Sanagia veliferaVaricorhinus fimbriatus
jaegerimariaesandersisteindachneritornieriwerneri
Denticipidae Denticeps clupeoidesEleotridae Eleotris daganensis
feaiKribia kribensis
Gobiidae Awaous lateristrigaSicydium crenilabrum
Hepsetidae Hepsetus odoeKneriidae Grasseichthys gabonensis
Parakneria abbreviataMalapteruridae Malapterurus electricusMastacembelidae Aethiomastacembelus marchei
sexdecimspinosusCaecomastacembelus batesii
brevicaudacryptacanthusdecorseiflavomarginatusgorolongicaudamarchei
170 Rivers of the lower Guinean rainforest:
FAMILY GENUS Species
marmoratusnigersanagalisclateriseiteri
Mochokidae Atopochilus savorgnaniChiloglanis batesii
cameronensisdisneyimicropogonnigerpolypogon
Hemisynodontis membranaceusMicrosynodontis batesiiSynodontis albolineatus
annectensbatesiieupterusguttatushaugimarmoratusnigritaobesusocelliferpolyodonrebelirobbianusschallsteindachneritessmanni
Mormyridae Boulengeromyrus knoepffleriBrienomyrus adustus
batesiibrachyistiuscurvifronshopkinsikingsleyaelongianalislongicaudatussphecodes
Campylomormyrus curvirostrisphantasticus
Gnathonemus petersiiHippopotamyrus castorIsichthys henryiIvindomyrus opdenboschiMarcusenius abadii
bruciiconicephalusfritelimentomooriintemensispaucisquamatus
Mormyrops anguilloidesbatesianuscaballus
Biogeography and sustainable exploitation 171
FAMILY GENUS Species
zanclirostrisMormyrus caballus
felixihasselquistiirumetapirusthomasi
Paramormyrops gabonensisPetrocephalus ansorgii
ballayicatostomaguttatusmicrophthalmussimus
Pollimyrus adspersuslhuysimarcheipedunculatuspolylepiswalkeri
Stomatorhinus polylepiswalkeri
Nandidae Polycentropsis abbreviataNotopteridae Papyrocranus afer
Xenomystus nigriPantodontidae Pantodon buchholziPoeciliidae Aplocheilichthys camerunensis
luxophthalmusscheelispilauchen
Hylopanchax stictopleuronHypsopanchax catenatus
zebraPlataplochilus cabindae
loemensismiltotaeniangaensisterveri
Procatopus aberransnototaeniasimilis
Phractolaemidae Phractolaemus ansorgiiPolypteridae Erpetoichthys calabaricus
Polypterus retropinnisProtopteridae Protopterus dolloiSchilbeidae Parailia occidentalis
pellucidaPareutropius buffei
debauwiSchilbe brevianalis
djeremigrenfelliintermediusmicropogonmultitaeniatusmystusnyongensis





























