Embed Size (px)
Citation preview

Insect Biochem., 1976, Vol. 6, pp. 571 to 578. Pergamon Press. Printed in Great Britain
RIBOSOMAL REPLACEMENT AND DEGRADATION DURING METAMORPHOSIS OF THE
BLOWFLY, CALLIPHORA VICINA
ALBERTO PROTZEL, S. SRIDHARA,* and LEO LEVENB(~K
Laboratory of Physical Biology, National Institute of Arthritis, Metabolism, and Digesive Diseases, National Institutes of Health, Bethesda,
Maryland 20014, U:S.A.
(Received 7 June, 1976)
Abstract--Larvae of the blowfly Calliphora vicina were either raised on a semi-defined diet containing labeled uridine and/or lysine, or injected with these isotopes. Labeled ribosomal RNA was then isolated and its specific activity determined to estimate ribosomal half-lives during development. The larger and smaller ribosomal sub-units turn over with statistically indistinguishable half-lives of about 8.3 days. The CMP component of rRNA turns over significantly more slowly (half-life: 10.4 days) than the corresponding UMP (8.5 days). However, by making some likely assumptions and including the specific activity of the precursor UTP pool in the computation, the probable half-life of the ribosomal pool during metamorphosis is calculated to be about 2.2 days. This value indicates that over 90~o of the relatively constant ribosomal pool is newly synthesized by pharate adult cells during metamor- phosis, with an equivalent amount being degraded by larval cells (excluding the fat body) during this period. Fat body ribosomes are destroyed in situ prior to pupariation to form a degraded, relatively insoluble material ('pellet RNA') which is metabolically inert.
INTRODUCTION
THE TOTAL ribosome content of mature 3rd instar lar- vae of the blowfly Calliphora vicina R. D. (= C. eryth- rocephala, Meig) decreases by about 459/o shortly before pupariation, with but little change in the number of ribosomes thereafter (SRIDI-IAgA and LEW~OOK, 1974). Such unexpected constancy of ribosomes during the profound morphological changes occurring during fly metamorphosis (P~gEZ, 1910; BARRIT and BraT, 1971) suggests either meta- bolic stability of the ribosomal population, or essen- tially equal rates of ribosomal breakdown and de novo synthesis. The former possibility is incompatible with the observations that isotopic precursors are incor- porated into rRNA (CROSSLEY, 1972; PETR1 et al., 1971), and pre-labeled RNA is broken down (ALONSO, 1973) during metamorphosis of Diptera. The present experiments were therefore designed to quantitatively determine the rates of ribosomal degradation, and by inference, the corresponding rates of biosynthesis dur- ing pharate adult development.
Under most circumstances our experiments would lead to an estimate of the rate of ribosomal turnover. However, in the case of metamorphosing insects, where larval tissues are being degraded, and adult organs are concurrently in process of development, true ribosomal turnover is complicated by what may
* Present Address: Established Investigator of the Amer- ican Heart Assn., The Department of Biological Sciences, Northwestern University, Evanston, Illinois 60201, U.S.A.
be termed ribosomal replacement, i.e. the replacement of larval ribosomes by their newly synthesized adult counterparts. Turnover and such replacement cannot easily be distinguished experimentally.
Our earlier study (SRIDI-t~.A and LEVENaOOK, 1974) also indicated that the observed loss in C. vicina ribo- somes before pupariation was attributable to destruc- tion of larval fat body ribosomes. Concomitant with the loss of these ribosomes was the appearance in the fat body of a low mol. wt (4-7S), relatively insol- uble RNA-like material termed 'pellet RNA'. We now present further evidence that 'pellet RNA' is derived from rRNA, and that, in contrast to the latter, it does not appear to turn over.
MATERIALS AND METHODS
Materials
[2-1*C]-uridine (52.7 mCi/rn-mole), L-[G-aH]-lysine (2.5Ci/m-mole), [5-3H]-uridine (26.4Ci/m-mole), Omni- fluor and Triton X-100 were purchased from New England Nuclear. Polygram Cel 300 PEI sheets for thin layer chro- matography were obtained from Brinkman Instruments, Inc. Acrylamide and N-N-Methylene-bis-acrylamide were purchased from Eastman, the latter being recrystal- lized from acetone. N,N,N',N',-tetramethylethylenediamine (TEMED) was purchased from Sigma. All other reagents were analytical grade.
Rearing of insects
C. vicina larvae were raised either on chopped horsemeat or aseptically on a semi-defined diet containing radioactive
571
572 ALBERTO PROTZEL, S. SRIDHARA, AND LEO LEVENBOOK
precursors (CHEN and LEVENBOOK, 1966; SRIDHARA and LEVENBOOK, 1974). Two hundred and fifty eggs were added to 75 ml of medium containing two to three #Ci of [5-3H] - uridine per ml in a 250 ml Erlenmeyer flask. Additional radioisotopes were added as required. All larvae were maintained at 25°C, at about 75~o relative humidity and 18 hr of illumination per day. Description of the blowfly life cycle raised under these conditions has been presented earlier (SRIDHARA and LEVENBOOK, 1974).
Injection of isotopes
Ribosomes and RNA from horsemeat-fed larvae were labeled by injection of radioactive isotope. Larvae at the desired stage of development were cooled on ice, anesthe- tized with carbon dioxide (dry ice) and individually in- jected laterally in the 2nd pre-anal segment with 1/~1 of radioisotope employing a fine capillary needle attached with polyethylene tubing to an Agla (Burroughs Wellcome Co.) syringe.
Isolation of Ribosomes, RNA and 'pellet RNA'
Procedures for the isolation and characterization of ribosomes and 'pellet RNA' have been described (SRID- HARA and LEVENBOOK, 1974). Ribosomes thus obtained had an A260/A2s0 ratio of at least 1.8 and consisted of about 55~o RNA and 45~o protein. Ribosomal protein was iso- lated by the procedure of SPITNIK-ELSON (1965) and its concentration measured by a modified Lowry procedure (Low et al., 1969).
To determine the rates of rRNA decay, total RNA from [5-3H]-uridine fed larvae was extracted according to GREEN~ERG (1969), except that the concentration of NaC1 in the extraction medium was 0.15 M, and 5 vol of extrac- tion medium per insect was employed. After three phenol
2 0 -
1 5 -
10
x
"O
~" 20 .=_
15
26S A"
18S
26S B
18S
5
1 2 3 4 5 CMS
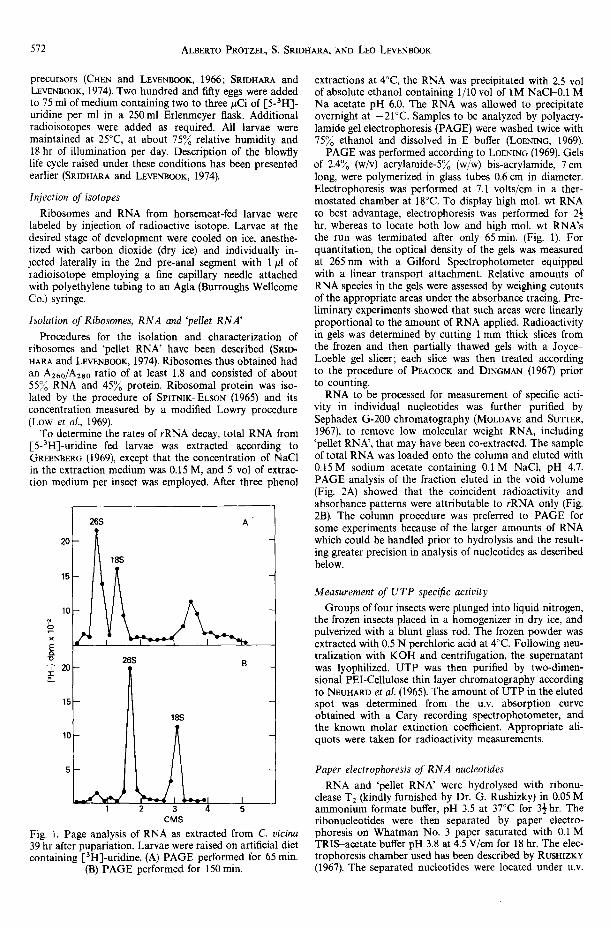
Fig. 1. Page analysis of RNA as extracted from C. vicina 39 hr after pupariation. Larvae were raised on artificial diet containing I-3H]-uridine. (A) PAGE performed for 65 min.
(B) PAGE performed for 150 min.
extractions at 4°C, the RNA was precipitated with 2.5 vol of absolute ethanol containing 1/10 vol of 1M NaCI-0.1 M Na acetate pH 6.0. The RNA was allowed to precipitate overnight at -21°C. Samples to be analyzed by polyacry- lamide gel electrophoresis (PAGE) were washed twice with 75~o ethanol and dissolved in E buffer (LOEN~NG, 1969).
PAGE was performed according to LOENING (1969). Gels of 2.4~o (w/v) acrylamide-5~o (w/w) bis-acrylamide, 7 cm long, were polymerized in glass tubes 0.6 cm in diameter. Electrophoresis was performed at 7.1 volts/cm in a ther- mostated chamber at 18°C. To display high mol. wt RNA to best advantage, electrophoresis was performed for 2½ hr, whereas to locate both low and high mol. wt RNA's the run was terminated after only 65 min. (Fig. 1). For quantitation, the optical density of the gels was measured at 265 nm with a Gilford Spectrophotometer equipped with a linear transport attachment. Relative amounts of RNA species in the gels were assessed by weighing cutouts of the appropriate areas under the absorbance tracing. Pre- liminary experiments showed that such areas were linearly proportional to the amount of RNA applied. Radioactivity in gels was determined by cutting 1 mm thick slices from the frozen and then partially thawed gels with a Joyce- Loeble gel slicer; each slice was then treated according to the procedure of PEACOCK and DINGMAN (1967) prior to counting.
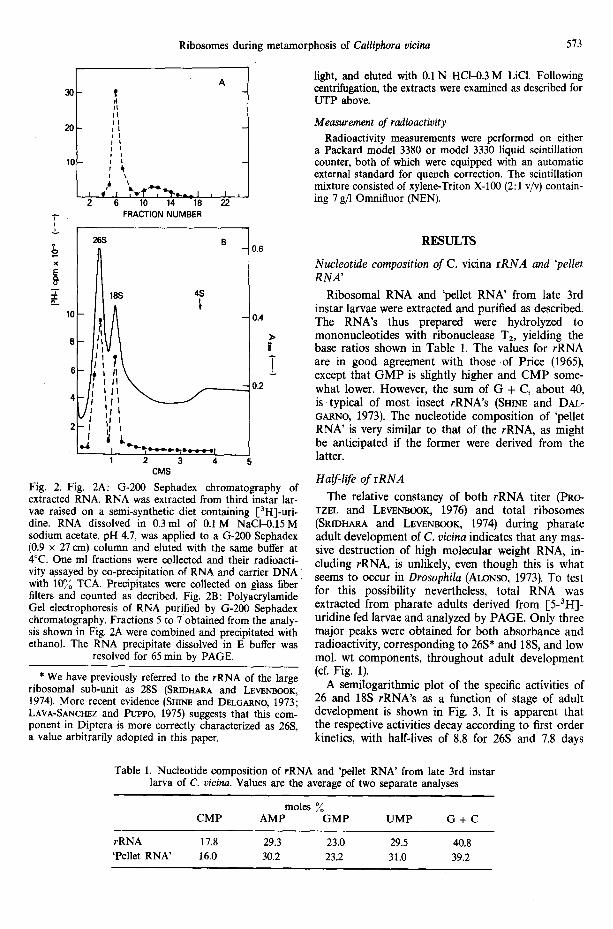
RNA to be processed for measurement of specific acti- vity in individual nucleotides was further purified by Sephadex G-200 chromatography (MOLDAVE and SUTTER, 1967), tO remove low molecular weight RNA, including 'pellet RNA', that may have been co-extracted. The sample of total RNA was loaded onto the column and eluted with 0.15M sodium acetate containing 0.1 M NaC1, pH 4.7. PAGE analysis of the fraction eluted in the void volume (Fig. 2A) showed that the coincident radioactivity and absorbance patterns were attributable to rRNA only (Fig. 2B). The column procedure was preferred to PAGE for some experiments because of the larger amounts of RNA which could be handled prior to hydrolysis and the result- ing greater precision in analysis of nucleotides as described below.
Measurement of UTP specific activity Groups of four insects were plunged into liquid nitrogen,
the frozen insects placed in a homogenizer in dry ice, and pulverized with a blunt glass rod. The frozen powder was extracted with 0.5 N perchloric acid at 4°C. Following neu- tralization with KOH and centrifugation, the supernatant was lyophilized. UTP was then purified by two-dimen- sional PEI-Cellulose thin layer chromatography according to NEUHARD et al. (1965). The amount of UTP in the eluted spot was determined from the u.v. absorption curve obtained with a Cary recording spectrophotometer, and the known molar extinction coefficient. Appropriate ali- quots were taken for radioactivity measurements.
Paper electrophoresis of RNA nucleotides
RNA and 'pellet RNA' were hydrolysed with ribonu- clease T 2 (kindly furnished by Dr. G. Rushizky) in 0.05 M ammonium formate buffer, pH 3.5 at 37°C for 3½ hr. The ribonucleotides were then separated by paper electro- phoresis on Whatman No. 3 paper saturated with 0.1 M TRIS-aeetate buffer pH 3.8 at 4.5 V/cm for 18 hr. The elec- trophoresis chamber used has been described by RUSHIZKY (1967). The separated nucleotides were located under u.v.
Ribosomes during metamorphosis of Calliphora vicina 573
30
20
t d JI I I
I I
t t
, t x
2 6 10 14 18 FRACTION NUMBER
I 22
"V I
JL i
[ 26S B ? 0 +
,.,F 18S 4p
, t l t
l_i - ~ l~,e., lb. e . l . o . 0p. I . o.eD,.~lp_i~. °
1 2 3 4 5 CMS
Fig. 2. Fig. 2A: G-200 Sephadex chromatography of extracted RNA. RNA was extracted from third instar lar- vae raised on a semi-synthetic diet containing [3H]-uri- dine. RNA dissolved in 0.3ml of 0.1 M NaCI-0.15M sodium acetate, pH 4.7, was applied to a G-200 Sephadex (0.9 x 27 cm) column and elUted with the same buffer at 4°C. One ml fractions were collected and their radioacti- vity ~sayed by co-precipitation of RNA and carrier DNA with 10% TCA. Precipitates were collected on glass fiber filters and counted as decribed. Fig. 2B: Polyacrylamide Gel electrophoresis of RNA purified by G-200 Sephadex chromatography. Fractions 5 to 7 obtained from the analy- sis shown in Fig. 2A were combined and precipitated with ethanol. The RNA precipitate dissolved in E buffer was
resolved for 65 min by PAGE.
* We have previously referred to the rRNA of the large ribosomal sub-unit as 28S (SR£DHARA and L~VEtCaOOK, 1974). More recent evidence (SnrNE and DELGARNO, 1973; LAVA-SANCHEZ and Pm, Po, 1975) suggests that this com- ponent in Diptera is more correctly characterized as 26S, a value arbitrarily adopted in this paper.
light, and eluted with 0.1 N HC1-0.3 M LiC1. Following centrifugation, the extracts were examined as described for UTP above.
Measurement of radioactivity Radioactivity measurements were performed on either
a Packard model 3380 or model 3330 liquid scintillation counter, both of which were equipped with an automatic external standard for quench correction. The scintillation mixture consisted of xylene-Triton X-100 (2:1 v/v) contain- ing 7 g/10mnittuor (NEN).
RESULTS
Nucleotide composition of C. vicina rRNA and 'pellet RNA'
Ribosomal RNA and 'pellet RNA' from late 3rd instar larvae were extracted and purified as described. The RNA's thus prepared were hydrolyzed to mononucleotides with ribonuclease T2, yielding the base ratios shown in Table 1. The values for rRNA are in good agreement with those o f Price (1965), except that G M P is slightly higher and CMP some- what lower. However, the sum of G + C, about 40, is typical of most insect rRNA's (SH~B and DAL- G~NO, 1973). The nucleotide composition of 'pellet RNA' is very similar to that of the rRNA, as might be anticipated if the former were derived from the latter.
Half-life of rRNA
The relative constancy of both rRNA titer (PRo- TZEL and LEWl, mOOK, 1976) and total ribosomes (SRmRARA and L E w ~ o o ~ 1974) during pharate adult development of C. vicina indicates that any mas- sive destruction of high molecular weight RNA, in- cluding rRNA, is unlikely, even though this is what seems to occur in Drosophila (ALoNSO, 1973). To test for this possibility nevertheless, total RNA was extracted from pharate adults derived from I-5-31-1] - uridine fed larvae and analyzed by PAGE. Only three major peaks were obtained for both absorbance and radioactivity, corresponding to 26S* and 18S, and low mol. wt components, throughout adult development (cf. Fig. 1).
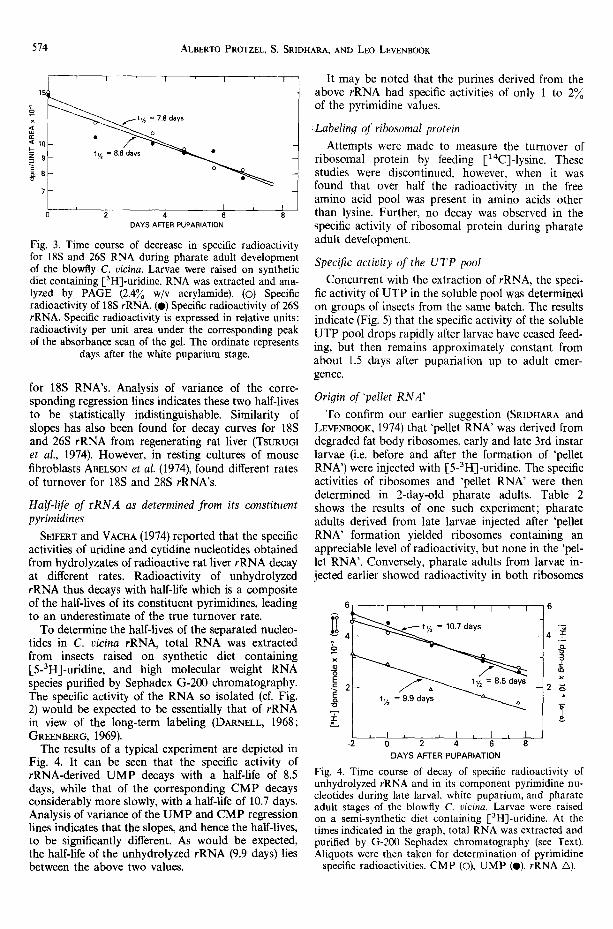
A semilogarithmic plot of the specific activities of 26 and 18S rRNA's as a function of stage of adult development is shown in Fig. 3. It is apparent that the respective activities decay according to first order kinetics, with half-lives of 8.8 for 26S and 7.8 days
Table 1. Nucleotide composition of rRNA and 'pellet RNA' from late 3rd instar larva of C. vicina. Values are the average of two separate analyses
moles % CMP AMP GMP UMP G + C
rRNA 17.8 29.3 23.0 29.5 40.8 'Pellet RNA' 16.0 30.2 23.2 31.0 39.2
5 7 4 A L B E R T O P R O T Z E L , S . S R I D H A R A , A N D L E O L E V E N B O O K
iS~ ' I I I
I < , y2 78o.ys
• ~ ~ t,/~ = 8 8 days
, I , I , I , I 2 4 6 8
DAYS AFTER PUPARIATION
Fig. 3. Time course of decrease in specific radioactivity for 18S and 26S RNA during pharate adult development of the blowfly C. vicina. Larvae were raised on synthetic diet containing [3H]-uridine. RNA was extracted and ana- lyzed by PAGE (2,4~o w/v acrylamide). (©) Specific radioactivity of 18S rRNA. (0) Specific radioactivity of 26S rRNA. Specific radioactivity is expressed in relative units: radioactivity per unit area under the corresponding peak of the absorbance scan of the gel. The ordinate represents
days after the white puparium stage.
for 18S RNA's. Analysis of variance of the corre- sponding regression lines indicates these two half-lives to be statistically indistinguishable. Similarity of slopes has also been found for decay curves for 18S and 26S rRNA from regenerating rat liver (TSURU~I et al., 1974). However, in resting cultures of mouse fibroblasts ABELSON et al. (1974), found different rates of turnover for 18S and 28S rRNA's.
Half-life of rRNA as determined from its constituent p yrimidines
SE[FERT and VACHA (1974) reported that the specific activities of uridine and cytidine nucleotides obtained from hydrolyzates of radioactive rat liver rRNA decay at different rates. Radioactivity of unhydrolyzed rRNA thus decays with half-life which is a composite of the half-lives of its constituent pyrimidines, leading to an underestimate of the true turnover rate.
To determine the half-lives of the separated nucleo- tides in C. vicina rRNA, total RNA was extracted from insects raised on synthetic diet containing [5-3H]-uridine, and high molecular weight RNA species purified by Sephadex G-200 chromatography. The specific activity of the RNA so isolated (cf. Fig. 2) would be expected to be essentially that of rRNA in view of the long-term labeling (DARNELL, 1968; GREENBERG, 1969).
The results of a typical experiment are depicted in Fig. 4. It can be seen that the specific activity of rRNA-derived UMP decays with a half-fife of 8.5 days, while that of the Corresponding CMP decays considerably more slowly, with a half-fife of 10.7 days. Analysis of variance of the UMP and CMP regression lines indicates that the slopes, and hence the half-lives, to be significantly different. As would be expected, the half-life of the unhydrolyzed rRNA (9.9 days) lies between the above two values.
It may be noted that the purines derived from the above rRNA had specific activities of only 1 to 2~o of the pyrimidine values.
:Labeling of ribosomal protein
Attempts were made to measure the turnover of ribosomal protein by feeding [14C]-lysine. These studies were discontinued, however, when it was found that over half the radioactivity in the free amino acid pool was present in amino acids other than lysine. Further, no decay was observed in the specific activity of ribosomal protein during pharate adult development.
Specific activity of the UTP pool
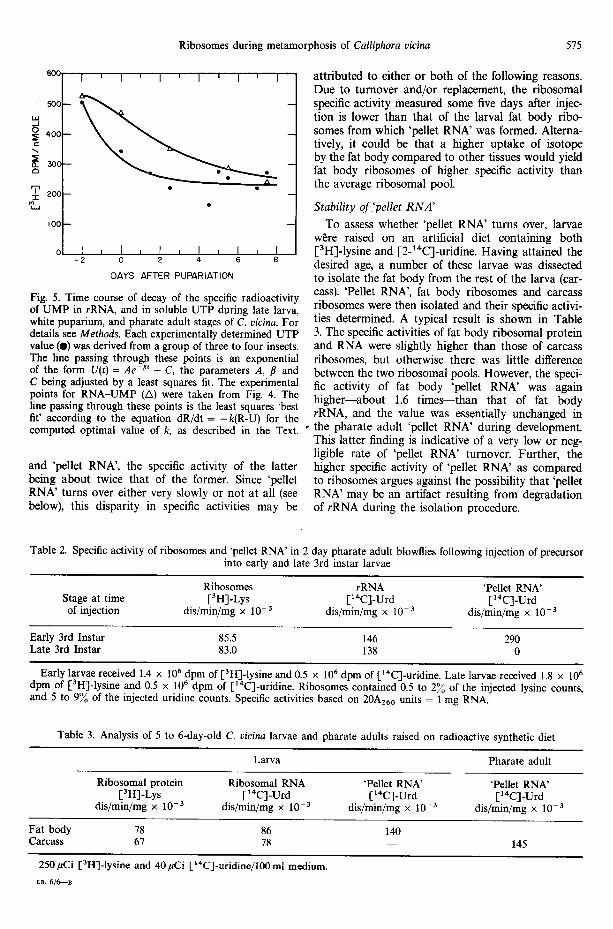
Concurrent with the extraction of rRNA, the speci- fic activity of UTP in the soluble pool was determined on groups of insects from the same batch. The results indicate (Fig. 5) that the specific activity of the soluble UTP pool drops rapidly after larvae have ceased feed- ing, but then remains approximately constant from about 1.5 days after pupariation up to adult emer- gence.
Origin of "pellet RN A'
To confirm our earlier suggestion (SRIDHARA and LEVENBOOK, 1974) that 'pellet RNA' was derived from degraded fat body ribosomes, early and late 3rd instar larvae (i.e. before and after the formation of 'pellet RNA') were injected with [5-3H]-uridine. The specific activities of ribosomes and 'pellet RNA' were then determined in 2-day-old pharate adults. Table 2 shows the results of one such experiment; pharate adults derived from late larvae injected after 'pellet RNA' formation yielded ribosomes containing an appreciable level of radioactivity, but none in the 'pel- let RNA'. Conversely, pharate adults from larvae in- jected earlier showed radioactivity in both ribosomes
6 6 ,k ' I ' I ' I ' I ' I
4 4 -r
t , / l
I , I , I , I , I , I 2 0 2 4 6 8
DAYS AFTER PUPARIATION
Fig. 4. Time course of decay of specific radioactivity of unhydrolyzed rRNA and in its component pyrimidine nu- cleotides during late larval, white puparium, and pharate adult stages of the blowfly C. vicina. Larvae were raised on a semi-synthetic diet containing I-3H]-uridine. At the times indicated in the graph, total RNA was extracted and purified by G-200 Sephadex chromatography (see Text). Aliquots were then taken for determination of pyrimidine
specific radioactivities. CMP (©), UMP (0), rRNA A).
Ribosomes during metamorphosis of Calliphora vicina 575
600 "I 500
.~ 400
T 2O0
1 0 0 D
I - 2
I ' I ' I ' I ' I ' I
• • A
I i I i I J I f I 0 2 4 6 8
DAYS AFTER PUPARIATION
Fig. 5. Time course of decay of the specific radioactivity of UMP in rRNA, and in soluble UTP during late larva, white puparium, and pharate adult stages of C. vicina. For details see Methods. Each experimentally determined UTP value (0) was derived from a group of three to four insects. The line passing through these points is an exponential of the form U(t) = Ae -at + C, the parameters A, // and C being adjusted by a least squares fit. The experimental points for RNA-UMP (A) were taken from Fig. 4. The line passing through these points is the least squares 'best fit' according to the equation dR/dt = -k(R-U) for the computed optimal value of k, as described in the Text.
and 'pellet RNA', the specific activity of the latter being about twice that of the former. Since 'pellet RNA' turns over either very slowly or not at all (see below), this disparity in specific activities may be
attributed to either or both of the following reasons. Due to turnover and/or replacement, the ribosomal specific activity measured some five days after injec- tion is lower than that of the larval fat body ribo- somes from which 'pellet RNA' was formed. Alterna- tively, it could be that a higher uptake of isotope by the fat body compared to other tissues would yield fat body ribosomes of higher specific activity than the average ribosomal pool.
Stability o f 'pe l le t R N A "
To assess whether 'pellet RNA' turns over, larvae wbre raised on an artificial diet containing both [3H]-lysine and [2-14C]-uridine. Having attained the desired age, a number of these larvae was dissected to isolate the fat body from the rest of the larva (car- cass). 'Pellet RNA', fat body ribosomes and carcass ribosomes were then isolated and their specific activi- ties determined. A typical result is shown in Table 3. The specific activities of fat body ribosomal protein and RNA were slightly higher than those of carcass ribosomes, but otherwise there was little difference between the two ribosomal pools. However, the speci- fic activity of fat body 'pellet RNA' was again higher--about 1.6 t imes-- than that of fat body rRNA, and the value was essentially unchanged in the pharate adult 'pellet RNA' during development. This latter finding is indicative of a very low or neg- ligible rate of 'pellet RNA' turnover. Further, the higher specific activity of 'pellet RNA' as compared to ribosomes argues against the possibility that 'pellet RNA' may be an artifact resulting from degradation of rRNA during the isolation procedure.
Table 2. Specific activity of ribosomes and 'pellet RNA' in 2 day pharate adult blowflies following injection of precursor into early and late 3rd instar larvae
Ribosomes rRNA 'Pellet RNA' Stage at time [3H]-Lys [~4C]-Urd [14C].Ur d of injection dis/min/mg x 10 -3 dis/min/mg x 10 -3 dis/min/mg x 10 -3
Early 3rd Instar 85.5 146 290 Late 3rd Instar 83.0 138 0
Early larvae received 1.4 x 1 0 6 dpm of [3H]-lysine and 0.5 x 1 0 6 dpm of [14C]-uridine. Late larvae received 1.8 × 1 0 6
dpm of [3H]-lysine and 0.5 x 106 dpm of [14C]-uridine. Ribosomes contained 0.5 to 2~ of the injected lysine counts, and 5 to 9~o of the injected uridine counts. Specific activities based on 20A26o units = 1 mg RNA.
Table 3. Analysis of 5 to 6-day-old C. vicina larvae and pharate adults raised on radioactive synthetic diet
Larva Pharate adult
Ribosomal protein Ribosomal RNA 'Pellet RNA' 'Pellet RNA' [3H]_Lys [14C].Urd [14C].Urd [14C].Urd
dis/min/mg x 10 -3 dis/min/mg x 10 -3 dis/min/mg x 10 -3 dis/min/mg x 10 -3
Fat body 78 86 140 Carcass 67 78 - - 145
250/tCi [3H]-lysine and 40/iCi [14C]-uridine/10Oml medium. LB. 6 / 6 - - B

576 ALBERTO PROTZEL, S. SRIDHARA, AND LEO LEVENBOOK
Table 4. Excretion of 'pellet RNA' by adult flies
'Pellet RNA'
dis/min/mg x 10 -3 dis/min/insect x 10 -3 'Jar wash'
dis/min/fly x 10 -3
Larvae raised on synthetic diet 150 21 16
Larva injected during early 3rd instar 290 39 35
Larvae were either raised on a synthetic diet containing 7.5 #Ci of [14C]-uridine or early 3rd instar larvae were injected with 0.25/~Ci of [14C]-uridine. The radioactivity in pharate adult 'pellet RNA' and in 'jar wash' remaining in jars after adult emergence was then determined. 'Jar wash' is the material recoverable from the jars in which adults had emerged (see Text). The pool of 'pellet RNA' was 0.14 mg per insect.
Since adult flies do not contain 'pellet RNA' (SRID- HARA and LEVENBOOK, 1974) its ultimate fate is a matter of interest. From 76 to 90~o of 'pellet RNA' radioactivity could be accounted for in aqueous extracts of meconia, excreta and empty pupal cases remaining in jars ('jar wash') in which the flies had emerged (Table 4). In view of the absence of high mol. wt material (SRIDHARA and LEVENBOOK, 1974) the radioactive extracts were analyzed on a Dowex-1 (formate) column (PLAISTED and REGGIO, 1963); most of the radioactivity appeared in the nucleic acid base + nucleoside fraction, while none was associ- ated with individual nucleotides. Thus, assuming that 'jar wash' radioactivity is derived from 'pellet RNA', it would seem to be extensively degraded at the time of adult emergence.
DISCUSSION
Ribosomal turnover
The specific activity of C. vicina rRNA, derived from larvae either fed or injected with labeled uridine, progressively decreases during the insect's metamor- phosis. Making certain assumptions, the rate of this decrease can be used to compute the replacement rate of the ribosomal pool during pharate adult develop- ment. It should be noted that the rRNA pool is almost constant during metamorphosis (PROTZEL and LEVENaOOK, 1976).
The simplest model for such computation consists of a steady-state homogeneous pool of labeled rRNA decaying exponentially, with unlabeled input but radioactive output (SHIPLEY and CLARK, 1972). The apparent half-life of unhydrolyzed C. vicina rRNA based on such a model is 9.9 days. This value is a composite of the respective half-lives of its component pyrimidines CMP and UMP, which are significantly different. Based on ribosomal UMP, rRNA decays with a shorter half-life of 8.5 days. The rate of rRNA turnover calculated from this half-life would be grossly underestimated, since the UTP precursor pool (input) is not unlabeled as conjectured above, but is
in fact radioactive throughout development (cf. Fig. 5).
To account for this fact in computing the rRNA turnover rate, the time-cours e o f rRN A -U M P specific activity may be described by the differential equation:
dr _ - k ( R - U ) (1) dt
where at any time t, k is the fraction of the rRNA pool replaced per unit time, and R and U are the specific activities of rRNA-UMP and the soluble UTP pool respectively. Further, the time-course of U can itself be specified by fitting an exponential curve of the form:
U = Ae- ~' + C (2)
to the determined values of U. Such an exponential curve appeared to fit the experimental points (Fig. 5), more complex curve fitting not being justified by the quality of the data. In any event, small variations in the precise shape of the curve have only a minor effect on the derived value of k.
The parameters of equation (2), obtained by a least- squares, reiterative, curve-fitting computer program (MLAB), (KNOTT and REECE, 1972; COOPER, 1973), had the following values: A, 277.5; fl, 0.57; C, 230.7. To obtain the required value for k, equations (1) and (2) were combined, and the MLAB program employed to compute the best theoretical curve for the time-course of R when fitted to the actual exper- imental R values (Fig. 5).
It should be noted that in the determination of k, the source of labeled input into the UTP pool, includ- ing any recycling of label from rRNA breakdown, is immaterial since the UTP specific activities during development are known. However, an intrinsic error may exist if, due to the presence of multiple UTP pools (PLAGEMANN, 1972), the UTP as measured and hence defined by the theoretical fitted curve, does not accurately represent the true specific activity of the actual rRNA precursor(s).
Figure 5 illustrates the theoretical best fit for R thus obtained, computed from the final k value of

Ribosomes during metamorphosis of Calliphora vicina 577
0.31, and leads to a calculated rRNA half-life of 2.2 days in marked contrast to the 8.5 days when the radioactivity of the precursor pool is ignored. It may then be calculated that during metamorphosi s about 31% of the rRNA pool turns over per day, or that at least 93%/0 of the ribosomal pool has been synthe- sized de novo during pharate adult development.
It is well known that adult tissues are formed from the proliferation and differentiation of imaginal cells (cf. WlGGLESWORTH, 1972). In rapidly proliferating cells (WEBER, 1972; ABELSON et al., 1974) there is little or no detectable ribosomal turnover; it is entirely possible, therefore, that in dividing imaginal cells, ribosomes turn over either very slowly or not at all. Further, with the progressive formation of adult tis-
CHEN P. S. and LEVENBOOK L. (1966) Studies on the hemo- lymph proteins of the blowfly Phormia regina--II. Syn- thesis and breakdown as revealed by isotopic labeling. J. Insect Physiol. 12, 1611-1627.
COOPER H. L. (1973) Analysis of precursor pool labelling kinetics without pool size measurements: a computer assisted approach. Anal. Biochem. 53, 49-63.
CROSSLEY A. C. (1972) Ultrastructural changes during tran- sition of larval to adult intersegmental muscle at meta- morphosis in the blowfly Calliphora erythocephala---II. The formation of adult muscle. J. Embryol. exp. Morph. 27, 75-101.
DARrCELL J. E. (1968) Nucleic acids of animal cells. Bact. Revs. 32, 262-290.
DESSEV G. N. and GRANCHOV K. (1973) Degradation of ribonucleic acid in rat liver ribosomes. J. molec. Biol. 76, 425 ~4~.
sues, there is a concomitant increase in rRNA, i.e., GREENBERGJ. R. (1969)Synthesis and properties of riboso- an expanding adult pool. Conversely, in larval cells undergoing histolysis, the cell nuclei become pycnotic (THOMSON, 1973) leading to an impaired capacity for ribosomal biosynthesis, while pre-existing ribosomes are either expelled (CRoSSLEY, 1972) or degraded (SRIDHARA and LEVENaOOK, 1974). Since the total rRNA pool remains almost constant, a net increase in adult tissue rRNA, and a corresponding decrease in larval tissue rRNA is implied; to a first approxima- tion, k is then a measure of the rate of this conversion.
Observations on the larval fat body lead to a direct, albeit approximate value for the rate of fat body ribo- somal degradation. In this tissue most of the ribo- somes are degraded without replacement to form a metabolically stable product, 'pellet RNA'. The time for the fat body ribosomal pool to decrease to one half is about 1.5 days (SRIDHARA and LEVENBOOK, 1974); in the absence of any de novo synthesis, there- fore, this half-life may be used to calculate that the rate of fat-body ribosomal degradation is about 1.5 times that of the remaining larval ribosomes during pharate adult development. 'Pellet RNA' as such, rep- resents an unusually large accumulation of degraded RNA. Degradation products of rRNA have been observed in the fiber cells during chicken lens devel- opment (ZAPISEK and PAPACONSTANTINoU, t973), and in the degenerating silk gland of the silkworm (OKABE et al., 1974). Similarly, in vitro studies on the degrada- tion of rat liver rRNA showed that while 20% of the
R N A is converted into acid soluble products, frag- ments of about 500 nucleotides in length remain en- tirely intact (DESSEV and GRANCHOV, 1973).
REFERENCES
ABELSON H. T., JOHNSON L. F., PENMAN S., and GREEN H. (1974) Changes in RNA in relation to growth of the fibroblast--II. The lifetime of mRNA, rRNA, and tRNA in resting and growing cells. Cell 1, 161-165.
ALONSO C. (1973) RNA metabolism during metamorphosis of Drosophila hydei. J. Insect Physiol. 19, 2135-2142.
BARRIT L. C. and BIRT L. M. (1971) Development of Lu~i- lia cuprina. Correlation of biochemical and morphologi- cal events. J. Insect Physiol. 17, 1169-1183.
mal RNA in Drosophila. J. molec. Biol. 46, 85-98. KNOTT G. D. and REECE D. K. (1972) MLAB: A civilized
curve-fitting system. Proceedings of the ONLINE, 1972 International Conference l, 497-526, Brunel Univ., Eng- land.
LAVA-SANCHEZ P. A. and PuPPO S. (1975) Occurrence of 'hidden breaks' at specific sites of 26S ribosomal RNA of Musca carnaria. J. molec. Biol. 95, 9-20.
LOENING U. E. (1969) The determination of the molecular weight of ribonucleic acid by polyacrylamide-gel electro- phoresis. Biochem. J. 113, 131-138.
Low R. B., WOOL I. G., alad MARTIN T. E. (1969) Skeletal muscle ribosomal proteins: general characteristics and effect of diabetes. Biochim. Biophys. Acta 194, 190- 202.
MOLDAVE K. and SORTER R. P. (1967) Purification of aminoacyl-sRNA by molecular sieve chromatography on Sephadex. Methods in Enzymolooy 12, 598-601.
NEUHARD J., RANDERATH E. and RANDERATH K. (1965) Ion-exchange thin-layer chromatography--XIII. Resolu- tion of complex nucleoside triphosphate mixtures. Anal. Biochem. 13, 211-222.
OKABE K., KOYANAGI R., and KOGA K. (1975) RNA in the degenerating silk gland of Bombyx mori. J. Insect Physiol. 21, 1305-1309.
PEACOCK A. C. and DINGMAN C. W. (1968) Molecular weight estimation and separation of ribonucleic acid by electrophoresis in agarose-acrylamide composite gels. Biochemistry 7, 668-674.
PEREZ C. (1910) Recherches histologiques sur la mrtamor- phose des muscides Calliphora erythrocephala Mg. Arch. Zool. exp. 9bn. (5) 4, 1-274.
PETRI W. H., FRISTROM J. W., STEWART D. J., and HANLEY E. W. (1971) The in vitro synthesis and characteristics of ribosomal RNA in imaginal discs of Drosophila melano#aster. Molec. #en. Genet. 110, 245-262.
PLAGEMANN P. G. W. (1972) Nucleotide pools in Novikoff rat hepatoma cells growing in suspension culture--III. Effect of nucleosides in medium on levels of nucleotides in separate nucleotide pools for nuclear and cytoplasmic RNA synthesis. J. Cell Biol. 52, 131-146.
PLAISTED P. H. and REGGm R. B. (1963) A rapid column chromatographic separation of acid soluble nucleotides. Contrib. Boyce Thompson Inst. 22, 71-80.
PRICE G. M. (1965) Nucleic acids in the larva of the blowfly Calliphora erythrocephala. J. Insect Physiol. 11, 869-878.
PROTZEL A. and LEVENnOOK L. (1976) Rate of ribosomal RNA synthesis during metamorphosis of the blowfly Calliphora vicina. Insect Biochem. 6, 631-635.

578 ALBERTO PROTZEL, S. SRIDHARA, AND LEO LEVENBOOK
RUSHIZKY G. W. (1967) Mapping of oligonucleotides. Methods in Enzymology 12, 395-398.
SEIFERT J. and VACHA J. (1974) Differences in the turnover of uridylic and cytidylic acids of rat liver cytoplasmic ribosomes. Arch. Biochem. Biophys. 160, 285-288.
SHINE J. and DALGARNO L. (1973) Occurrence of heat dis- - sociable ribosomal RNA in insects. The presence of three
polynucleotide chains in 26S RNA from cultured Aedes aegypti cells. J. molec. Biol. 75, 57-72.
SHIPLEY R. A. and CLARK R. E. (1972) Tracer Methods for In Vivo Kinetics. Theory and Applications. Academic Press, N.Y.
SPITNIK-ELSON P. (1965) The preparation of ribosomal pro- tein from E. coli with lithium chloride and urea. Bio- chem. Biophys. Res. Comm. 18, 557-562.
SI~DHARA S. and LEVENBOOK L. (1974) The contribution of the fat body to RNA and ribosomal changes during
development of the blowfly Calliphora erythrocephala (Meig.). Develop. Biol. 38, 64-72.
THOMSON J. A. (1973) In The Biochemistry of Gene Expres- sion in Higher Organisms (Ed. by POLLAK J. K. and LEE J. W.) pp. 320-332. Australia and New Zealand Book Co., Sydney.
TSURUrI K., MORITA T., and OrATA K. (1974) Mode of degradation of ribosomes in regenerating rat liver in vivo. Eur. J. Biochem. 45, 119-126.
WEBER M. J. (1972) Ribosomal turnover in contact inhi- bited cells. Nature New Biol. 235, 58-61.
WIGGLESWORTH V. B. (1972) In The Principles of Insect Physiology 7th ed., pp. 61-145. Chapman & Hall, Eng- land.
ZAPISEK W. F. and PAPACONSTANTINOU J. (1973) Degrada- tion of ribosomal RNA during chicken lens develop- ment. Biochim. Biophys. Acta 299, 603-611.



















![Optical(Coherence(Tomography:(Age(estimation(of(Calliphora ......Introduction(Pupae,)a)sedentary)and)long]lasting)stage)of)the)blowfly)lifecycle,)are)often)reported)at)crime)scenes)](https://img.pdfslide.us/doc/110x75/60b54e2575eeff0dad7cccb6/opticalcoherencetomographyageestimationofcalliphora-introductionpupaeasedentaryandlonglastingstageoftheblowflylifecycleareoftenreportedatcrimescenes.jpg)









