Embed Size (px)
Citation preview

MPMI Vol. 34, No. 1, 2021, pp. 88–99. https://doi.org/10.1094/MPMI-07-20-0209-R
RibBX of Bradyrhizobium ORS285 Playsan Important Role in Intracellular Persistencein Various Aeschynomene Host Plants
Nico Nouwen,† Jean-Francois Arrighi, Djamel Gully, and Eric Giraud
Laboratoire des Symbioses Tropicales et Mediterraneennes, IRD, CIRAD, SupAgro, INRAE, University of Montpellier,Montpellier, France
Accepted 23 September 2020.
Bradyrhizobium ORS285 forms a nitrogen-fixating symbiosiswith both Nod factor (NF)-dependent and NF-independentAeschynomene spp. The Bradyrhizobium ORS285 ribBA gene en-codes for a putative bifunctional enzymewith 3,4-dihydroxybutanonephosphate (3,4-DHBP) synthase and guanosine triphosphate(GTP) cyclohydrolase II activities, catalyzing the initial stepsin the riboflavin biosynthesis pathway. In this study, we showthat inactivating the ribBA gene does not cause riboflavinauxotrophy under free-living conditions and that, as shownfor RibBAs from other bacteria, the GTP cyclohydrolase IIdomain has no enzymatic activity. For this reason, we haverenamed the annotated ribBA as ribBX. Because we were un-able to identify other ribBA or ribA and ribB homologs in thegenome of Bradyrhizobium ORS285, we hypothesize that theORS285 strain can use unconventional enzymes or an alter-native pathway for the initial steps of riboflavin biosynthesis.Inactivating ribBX has a drastic impact on the interaction ofBradyrhizobium ORS285 with many of the tested Aeschyno-mene spp. In these Aeschynomene spp., the ORS285 ribBXmutant is able to infect the plant host cells but the intracellularinfection is not maintained and the nodules senesce early. Thisphenotype can be complemented by reintroduction of the 3,4-DHBP synthase domain alone. Our results indicate that, inBradyrhizobiumORS285, the RibBX protein is not essential forriboflavin biosynthesis under free-living conditions and wehypothesize that its activity is needed to sustain riboflavinbiosynthesis under certain symbiotic conditions.
Keywords: bacteria-plant symbiosis, mutualism, nodule organogenesis,rhizobium, rhizobium-legume symbiosis, vitamin B2
Bradyrhizobium sp. ORS285 is a photosynthetic Gram-negative bacterium that belongs to the order Rhizobiales. It
invades the base of lateral roots or the adventitious root pri-mordia on the stem of host Aeschynomene spp. and triggers theformation of a new plant organ, the so called nodule. Withinplant cells of the formed nodules, the bacteria are chronicallymaintained and differentiate into dinitrogen-fixing bacteroids.The ORS285–Aeschynomene interaction is a very interestingsymbiotic model because (i) depending on the Aeschynomenespp., the infection and nodule organogenesis does or does notrely on the bacterial synthesis of Nod factors (NFs) (Giraudet al. 2007) and (ii) between NF-independent and NF-dependentAeschynomene spp., a drastic difference in morphology of theORS285 bacteroids is observed (i.e., the bacteroids are sphericalin NF-independent species whereas they are elongated in the NF-dependent ones) (Czernic et al. 2015).Riboflavin (vitamin B2) is the precursor of flavin mono-
nucleotide (FMN) and flavin adenine dinucleotide (FAD). FMNand FAD are cofactors of flavoproteins that are involved in alarge number of reactions concerning energy metabolism suchas degradation of fatty acids, carbohydrates, and amino acids(Abbas and Sibirny 2011). Flavoproteins also mediate reactionsduring biosynthesis and DNA repair, bioluminescence, andlight sensing (Christie et al. 2015; Dunlap 2014; Tagua et al.2015). Moreover, flavin nucleotides play an crucial role in thesynthesis of secondary metabolites and metabolism of othervitamins such as pyridoxine (vitamin B6), cobalamine (vitaminB12), and folate (Haase et al. 2014). Experiments with a ri-boflavin auxotroph of Rhizobium trifolii showed that the bio-availability of riboflavin strongly influences the symbioticinteraction with different red clover (Trifolium pratense) cul-tivars and that the effectiveness correlated with the flavincontent in the nodules (Pankhurst et al. 1974; Schwinghamer1970). Addition of riboflavin to the plant growth medium alsohas been shown to stimulate Medicago sativa root colonizationby Ensifer meliloti (Yang et al. 2002; Yurgel et al. 2014).Bacterial flavin metabolism has also been shown to be impor-tant for the intracellular survival and mouse colonization byBrucella abortus (Bonomi et al. 2010). This shows that ribo-flavin plays an important role in symbiotic and pathogenicmicrobe–host interactions.Plants and the majority of bacteria are able to synthesize ri-
boflavin de novo through the riboflavin biosynthetic pathway. Inthis pathway, one molecule of guanosin-59-triphosphate (GTP)and two molecules of ribulose-5-phosphate are converted intoone molecule of riboflavin (Fig. 1A). The bacterial enzymes(Escherichia coli abbreviations are used for clarity) in thispathway are GTP cyclohydrolase II (RibA), 3,4-dihydroxy-2-butanone 4-phosphate (3,4-DHBP) synthase (RibB), pyrimidinedeaminase/reductase (RibD), 6,7-dimethyl-8-ribityllumazine
†Corresponding author: N. Nouwen; [email protected]
Funding: This work was supported by a grant from the French NationalResearch Agency (grant “SymEffectors”; ANR-16-CE20-0013).
*The e-Xtra logo stands for “electronic extra” and indicates there aresupplementary materials published online.
The author(s) declare no conflict of interest.
Copyright © 2021 The Author(s). This is an open access articledistributed under the CC BY-NC-ND 4.0 International license.
88 / Molecular Plant-Microbe Interactions

synthase (RibH), and riboflavin synthase (RibE). The genomicorganization of rib genes and the presence of paralogs is highlyvariable (Garcıa-Angulo 2017). In some bacteria, the rib genesare clustered whereas, in others, they are found as mono-cistronic elements scattered over the chromosome. Also, bac-teria containing duplications or multiplications of one or morerib genes have been found. In addition, sometimes the GTPcyclohydrolase II and 3,4-DHBP synthase are fused and form ahybrid enzyme encoded by the ribBA gene, and the pyrimidinedeaminase and reductase function of RibD can be encoded bytwo separate genes, ribD (deaminase) and ribG (reductase),respectively. Riboflavin biosynthesis has shown to be regulatedby an FMN riboswitch (also called RFN element), a RNA se-quence present in the 59-untranslated region of some mRNAsencoding riboflavin biosynthesis enzymes (Winkler et al.2002). In general, the binding of FMN to the RFN elementdownregulates transcription or translation of the downstreamgenes. For a long time, RFN element regulation has been seenas the sole mechanism to control riboflavin biosynthesis inbacteria. However, studies in the last decade have shown thatother, more complex regulation mechanisms exist (Balasu-bramanian et al. 2010; da Silva Neto et al. 2013; Tan et al. 2015;Taniguchi and Wendisch 2015), which makes sense given thediverse role flavoproteins play in the cellular metabolism.Scattered over the chromosome, several putative riboflavin
biosynthesis genes have been found in BradyrhizobiumORS285 (Fig. 1B to D). These include (i) the ribD, ribE (calledribC in ORS285), and ribH1 genes which form an operonstructure (Fig. 1B); (ii) a second copy of ribH (ribH2) (Fig.1C); and (iii) a gene annotated as ribBA, encoding a putativebifunctional 3,4-DHBP synthase/GTP cyclohydrolase II whichforms an operon with a gene annotated as gcdH, encoding aputative glutaryl-CoA dehydrogenase, and likely a third gene,encoding a putative b-lactamase (BRAD5118=BRAD285_v2_5118)(Fig. 1D). This genomic context of rib genes in ORS285 is verywell conserved in bradyrhizobia.In this study, we have characterized the annotated ribBA gene
of Bradyrhizobium ORS285 and its role in symbiosis withdifferent Aeschynomene spp. We show that the protein encodedby ribBA does not have GTP cyclohydrolase II activity, as waspreviously shown for several other annotated ribBA genes(Brutinel et al. 2013). In line with these observations, we haverenamed the gene ribBX. Interestingly, inactivation of ribBXdid not lead to riboflavin auxotrophy under free-living condi-tions but had a drastic impact on the intracellular persistence ofthe ORS285 strain, especially in NF-independent Aeschyno-mene spp.
RESULTS
Inactivation of the Bradyrhizobium ORS285 ribBA genedoes not result in riboflavin auxotrophy.A large screen for mutations that affect the symbiosis be-
tween photosynthetic Bradyrhizobium ORS278 and Aeschy-nomene indica showed that a TN5 insertion into a geneannotated as ribBA results in the formation of nodules that areunable to fix nitrogen (Bonaldi et al. 2010). Based on homol-ogy, the ribBA gene of Bradyrhizobium ORS278 is annotated toencode for a bifunctional enzymewith the activities of both 3,4-DHBP synthase (RibB) and GTP cyclohydrolase II (RibA),which catalyze two essential reactions in riboflavin bio-synthesis (Garcıa-Angulo 2017). In bradyrhizobia, the genomiccontext of ribBA is very well conserved (for example, as in thegenetic organization in Bradyrhizobium ORS285) (Fig. 1D).Given its organization, ribBA most probably forms an operonwith the gcdH gene and this operon might also include a thirdgene encoding for a putative b-lactamase (BRAD285_v2_5118).
The gcdH gene encodes for a glutaryl-CoA dehydrogenase, anenzyme that can play a role in many different pathways (fattyacid/lysine/benzoate degradation, tryptophan metabolism, andmetabolism of secondary metabolites). Interestingly, in the re-action catalyzed by GcdH, a derivative of riboflavin, FAD,functions as cofactor by accepting the electron in the oxidation ofthe substrate, and its reoxidation by an electron-transfer flavo-protein completes the catalytic cycle of the enzyme. Transcribedin the opposite direction of ribBA is a gene that is annotated inBradyrhizobium genomes as putative 2-aminoadipate trans-aminase or transaminase. However, closer examination of thegene product showed that, in addition to the transaminase do-main, it also contains a helix-turn-helix DNA-binding domain.For this reason, we hypothesized that the gene product is not anenzyme but a transcriptional regulator belonging to the GntRfamily (Rigali et al. 2002) and, as such, could be implicated inthe expression of the ribBA-gcdH(-BRAD5118) operon. To in-vestigate the role of ribBA and the surrounding genes in thesymbiotic interaction of Bradyrhizobium ORS285 with Aeschy-nomene legumes, we have inactivated the ribBA, gcdH,BRAD5118, and gntR-like genes by integration of the non-replicative plasmid pJG194(-gusA) (Griffitts and Long 2008). Allmutant strains were able to grow on solid minimal growth me-dium without riboflavin supplementation (Fig. 1E) and hadsimilar growth rates compared with the wild-type (WT) strainwhen grown in equivalent liquid medium (Fig. 1F). This obser-vation is remarkable because RibB and RibA are the only known
Fig. 1. A, Riboflavin biosynthesis pathway and enzymes involved in thedifferent reactions in Escherichia coli: 1 = guanosine triphosphate (GTP);2 = 2,5-diamino-5-ribosylamino-4(3H)-pyrimidone-59-phosphate; 3 = 5-amino-6-ribosylamino-2,4(1H,3H)-pyrimidinione-59-phosphate; 4 = 5-amino-6-ribitylamino-2,4(1H,3H)-pyrimidinione-59-phosphate; 5 = 5-amino-6-ribitylamino-2,4(1H,3H)-pyrimidinione; 6 = ribulose-5-phosphate (Ru5P);7 = 3,4-dihydroxy-2-butanone-4-phosphate (3,4-DHBP); 8 = 6,7-dimethyl-8-ribityllumazine; and 9 = riboflavin. B to D, Schematic representation ofthe different genomic regions of putative rib genes in BradyrhizobiumORS285. The insertion of the nonreplicative plasmid to inactivate the dif-ferent genes in the ribBA region are indicated by an arrow in D. E, Growthof BradyrhizobiumORS285 and derivatives on minimal medium agar plateswithout riboflavin supplementation. F, Representative growth curves ofBradyrhizobium ORS285 and derivatives in minimal medium without ri-boflavin supplementation.
Vol. 34, No. 1, 2021 / 89
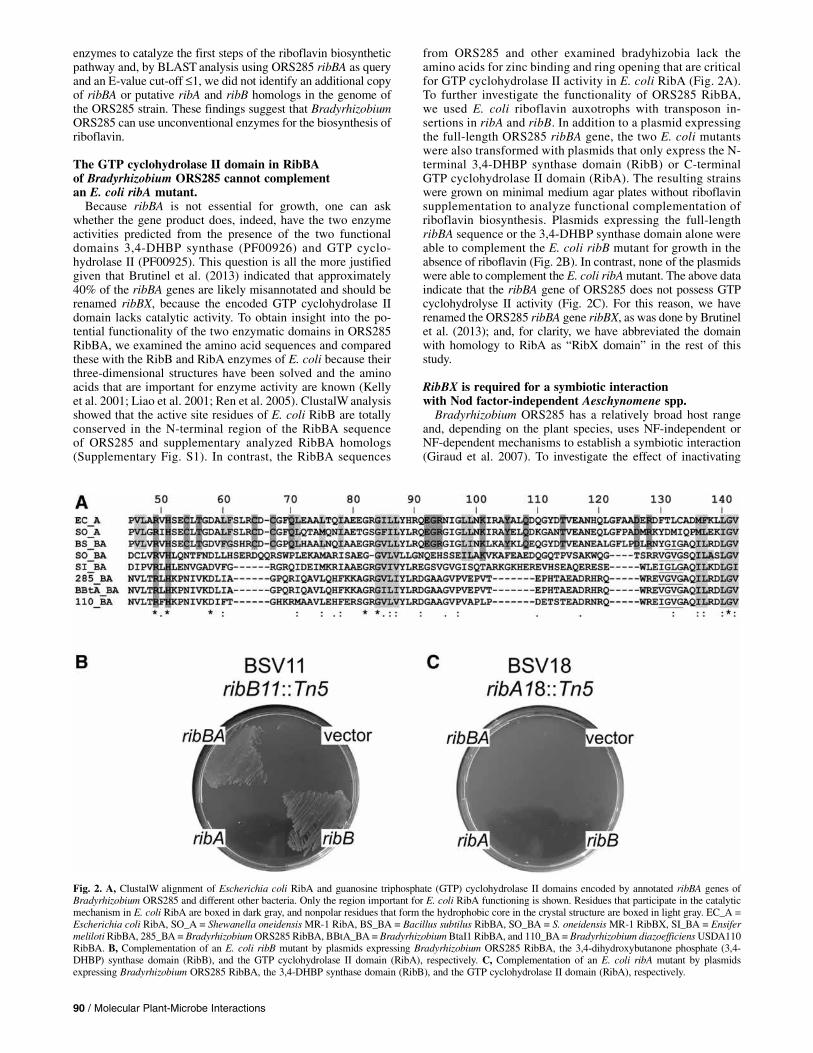
enzymes to catalyze the first steps of the riboflavin biosyntheticpathway and, by BLAST analysis using ORS285 ribBA as queryand an E-value cut-off £1, we did not identify an additional copyof ribBA or putative ribA and ribB homologs in the genome ofthe ORS285 strain. These findings suggest that BradyrhizobiumORS285 can use unconventional enzymes for the biosynthesis ofriboflavin.
The GTP cyclohydrolase II domain in RibBAof Bradyrhizobium ORS285 cannot complementan E. coli ribA mutant.Because ribBA is not essential for growth, one can ask
whether the gene product does, indeed, have the two enzymeactivities predicted from the presence of the two functionaldomains 3,4-DHBP synthase (PF00926) and GTP cyclo-hydrolase II (PF00925). This question is all the more justifiedgiven that Brutinel et al. (2013) indicated that approximately40% of the ribBA genes are likely misannotated and should berenamed ribBX, because the encoded GTP cyclohydrolase IIdomain lacks catalytic activity. To obtain insight into the po-tential functionality of the two enzymatic domains in ORS285RibBA, we examined the amino acid sequences and comparedthese with the RibB and RibA enzymes of E. coli because theirthree-dimensional structures have been solved and the aminoacids that are important for enzyme activity are known (Kellyet al. 2001; Liao et al. 2001; Ren et al. 2005). ClustalWanalysisshowed that the active site residues of E. coli RibB are totallyconserved in the N-terminal region of the RibBA sequenceof ORS285 and supplementary analyzed RibBA homologs(Supplementary Fig. S1). In contrast, the RibBA sequences
from ORS285 and other examined bradyhizobia lack theamino acids for zinc binding and ring opening that are criticalfor GTP cyclohydrolase II activity in E. coli RibA (Fig. 2A).To further investigate the functionality of ORS285 RibBA,we used E. coli riboflavin auxotrophs with transposon in-sertions in ribA and ribB. In addition to a plasmid expressingthe full-length ORS285 ribBA gene, the two E. coli mutantswere also transformed with plasmids that only express the N-terminal 3,4-DHBP synthase domain (RibB) or C-terminalGTP cyclohydrolase II domain (RibA). The resulting strainswere grown on minimal medium agar plates without riboflavinsupplementation to analyze functional complementation ofriboflavin biosynthesis. Plasmids expressing the full-lengthribBA sequence or the 3,4-DHBP synthase domain alone wereable to complement the E. coli ribB mutant for growth in theabsence of riboflavin (Fig. 2B). In contrast, none of the plasmidswere able to complement the E. coli ribAmutant. The above dataindicate that the ribBA gene of ORS285 does not possess GTPcyclohydrolyse II activity (Fig. 2C). For this reason, we haverenamed the ORS285 ribBA gene ribBX, as was done by Brutinelet al. (2013); and, for clarity, we have abbreviated the domainwith homology to RibA as “RibX domain” in the rest of thisstudy.
RibBX is required for a symbiotic interactionwith Nod factor-independent Aeschynomene spp.Bradyrhizobium ORS285 has a relatively broad host range
and, depending on the plant species, uses NF-independent orNF-dependent mechanisms to establish a symbiotic interaction(Giraud et al. 2007). To investigate the effect of inactivating
Fig. 2. A, ClustalW alignment of Escherichia coli RibA and guanosine triphosphate (GTP) cyclohydrolase II domains encoded by annotated ribBA genes ofBradyrhizobium ORS285 and different other bacteria. Only the region important for E. coli RibA functioning is shown. Residues that participate in the catalyticmechanism in E. coli RibA are boxed in dark gray, and nonpolar residues that form the hydrophobic core in the crystal structure are boxed in light gray. EC_A =Escherichia coli RibA, SO_A = Shewanella oneidensisMR-1 RibA, BS_BA = Bacillus subtilus RibBA, SO_BA = S. oneidensis MR-1 RibBX, SI_BA = EnsifermelilotiRibBA, 285_BA = BradyrhizobiumORS285 RibBA, BBtA_BA = BradyrhizobiumBtaI1 RibBA, and 110_BA = Bradyrhizobium diazoefficiensUSDA110RibBA. B, Complementation of an E. coli ribB mutant by plasmids expressing Bradyrhizobium ORS285 RibBA, the 3,4-dihydroxybutanone phosphate (3,4-DHBP) synthase domain (RibB), and the GTP cyclohydrolase II domain (RibA), respectively. C, Complementation of an E. coli ribA mutant by plasmidsexpressing Bradyrhizobium ORS285 RibBA, the 3,4-DHBP synthase domain (RibB), and the GTP cyclohydrolase II domain (RibA), respectively.
90 / Molecular Plant-Microbe Interactions
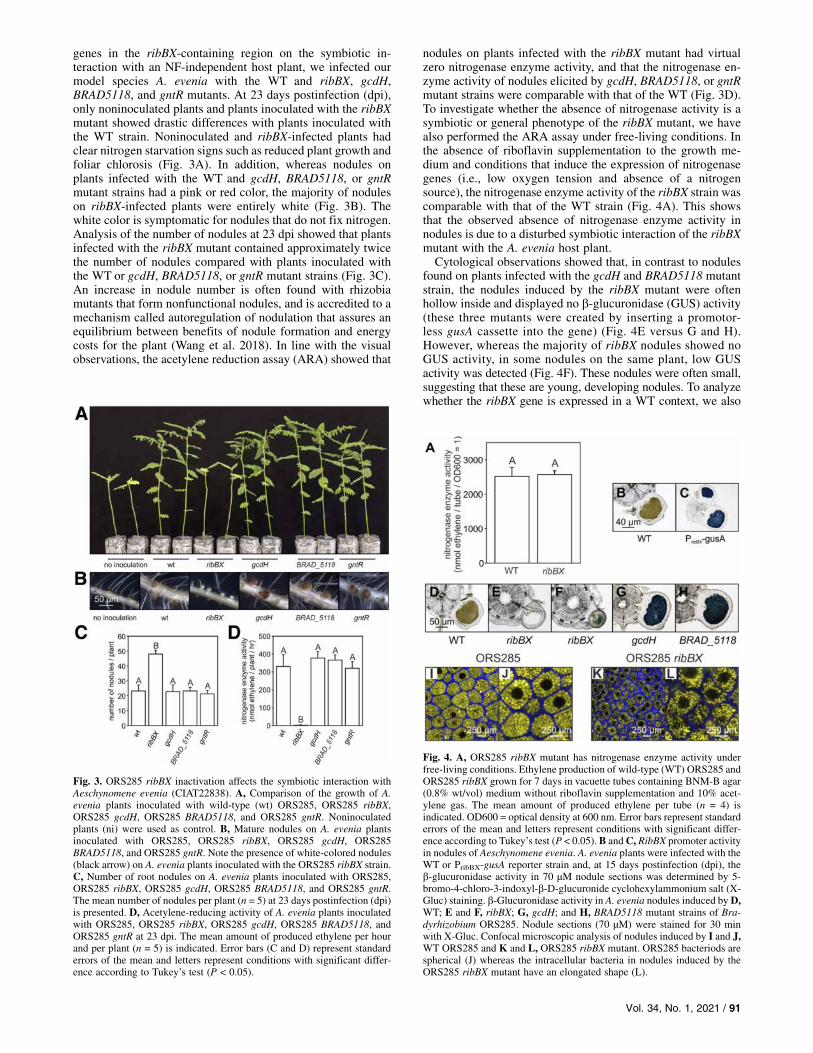
genes in the ribBX-containing region on the symbiotic in-teraction with an NF-independent host plant, we infected ourmodel species A. evenia with the WT and ribBX, gcdH,BRAD5118, and gntR mutants. At 23 days postinfection (dpi),only noninoculated plants and plants inoculated with the ribBXmutant showed drastic differences with plants inoculated withthe WT strain. Noninoculated and ribBX-infected plants hadclear nitrogen starvation signs such as reduced plant growth andfoliar chlorosis (Fig. 3A). In addition, whereas nodules onplants infected with the WT and gcdH, BRAD5118, or gntRmutant strains had a pink or red color, the majority of noduleson ribBX-infected plants were entirely white (Fig. 3B). Thewhite color is symptomatic for nodules that do not fix nitrogen.Analysis of the number of nodules at 23 dpi showed that plantsinfected with the ribBX mutant contained approximately twicethe number of nodules compared with plants inoculated withthe WT or gcdH, BRAD5118, or gntR mutant strains (Fig. 3C).An increase in nodule number is often found with rhizobiamutants that form nonfunctional nodules, and is accredited to amechanism called autoregulation of nodulation that assures anequilibrium between benefits of nodule formation and energycosts for the plant (Wang et al. 2018). In line with the visualobservations, the acetylene reduction assay (ARA) showed that
nodules on plants infected with the ribBX mutant had virtualzero nitrogenase enzyme activity, and that the nitrogenase en-zyme activity of nodules elicited by gcdH, BRAD5118, or gntRmutant strains were comparable with that of the WT (Fig. 3D).To investigate whether the absence of nitrogenase activity is asymbiotic or general phenotype of the ribBX mutant, we havealso performed the ARA assay under free-living conditions. Inthe absence of riboflavin supplementation to the growth me-dium and conditions that induce the expression of nitrogenasegenes (i.e., low oxygen tension and absence of a nitrogensource), the nitrogenase enzyme activity of the ribBX strain wascomparable with that of the WT strain (Fig. 4A). This showsthat the observed absence of nitrogenase enzyme activity innodules is due to a disturbed symbiotic interaction of the ribBXmutant with the A. evenia host plant.Cytological observations showed that, in contrast to nodules
found on plants infected with the gcdH and BRAD5118 mutantstrain, the nodules induced by the ribBX mutant were oftenhollow inside and displayed no b-glucuronidase (GUS) activity(these three mutants were created by inserting a promotor-less gusA cassette into the gene) (Fig. 4E versus G and H).However, whereas the majority of ribBX nodules showed noGUS activity, in some nodules on the same plant, low GUSactivity was detected (Fig. 4F). These nodules were often small,suggesting that these are young, developing nodules. To analyzewhether the ribBX gene is expressed in a WT context, we also
Fig. 3. ORS285 ribBX inactivation affects the symbiotic interaction withAeschynomene evenia (CIAT22838). A, Comparison of the growth of A.evenia plants inoculated with wild-type (wt) ORS285, ORS285 ribBX,ORS285 gcdH, ORS285 BRAD5118, and ORS285 gntR. Noninoculatedplants (ni) were used as control. B, Mature nodules on A. evenia plantsinoculated with ORS285, ORS285 ribBX, ORS285 gcdH, ORS285BRAD5118, and ORS285 gntR. Note the presence of white-colored nodules(black arrow) on A. evenia plants inoculated with the ORS285 ribBX strain.C, Number of root nodules on A. evenia plants inoculated with ORS285,ORS285 ribBX, ORS285 gcdH, ORS285 BRAD5118, and ORS285 gntR.The mean number of nodules per plant (n = 5) at 23 days postinfection (dpi)is presented. D, Acetylene-reducing activity of A. evenia plants inoculatedwith ORS285, ORS285 ribBX, ORS285 gcdH, ORS285 BRAD5118, andORS285 gntR at 23 dpi. The mean amount of produced ethylene per hourand per plant (n = 5) is indicated. Error bars (C and D) represent standarderrors of the mean and letters represent conditions with significant differ-ence according to Tukey’s test (P < 0.05).
Fig. 4. A, ORS285 ribBX mutant has nitrogenase enzyme activity underfree-living conditions. Ethylene production of wild-type (WT) ORS285 andORS285 ribBX grown for 7 days in vacuette tubes containing BNM-B agar(0.8% wt/vol) medium without riboflavin supplementation and 10% acet-ylene gas. The mean amount of produced ethylene per tube (n = 4) isindicated. OD600 = optical density at 600 nm. Error bars represent standarderrors of the mean and letters represent conditions with significant differ-ence according to Tukey’s test (P < 0.05).B andC, RibBX promoter activityin nodules of Aeschynomene evenia. A. evenia plants were infected with theWT or PribBX-gusA reporter strain and, at 15 days postinfection (dpi), theb-glucuronidase activity in 70 µM nodule sections was determined by 5-bromo-4-chloro-3-indoxyl-b-D-glucuronide cyclohexylammonium salt (X-Gluc) staining. b-Glucuronidase activity in A. evenia nodules induced byD,WT; E and F, ribBX; G, gcdH; and H, BRAD5118 mutant strains of Bra-dyrhizobium ORS285. Nodule sections (70 µM) were stained for 30 minwith X-Gluc. Confocal microscopic analysis of nodules induced by I and J,WT ORS285 and K and L, ORS285 ribBX mutant. ORS285 bacteriods arespherical (J) whereas the intracellular bacteria in nodules induced by theORS285 ribBX mutant have an elongated shape (L).
Vol. 34, No. 1, 2021 / 91
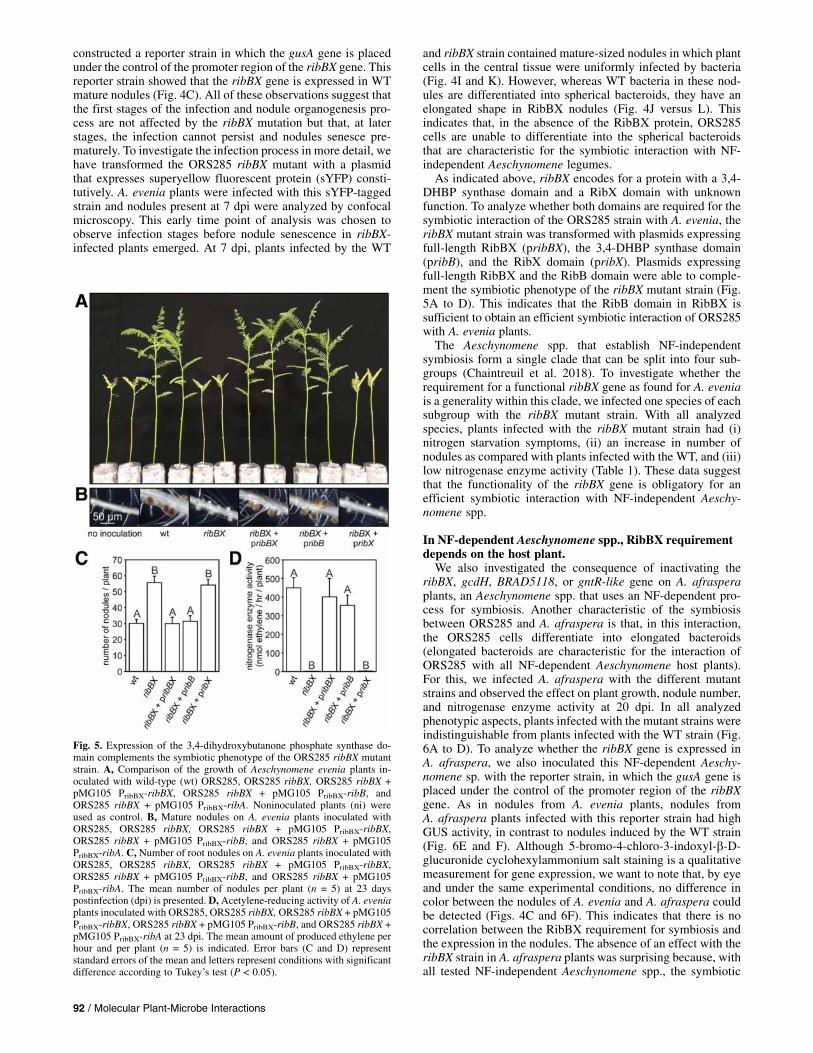
constructed a reporter strain in which the gusA gene is placedunder the control of the promoter region of the ribBX gene. Thisreporter strain showed that the ribBX gene is expressed in WTmature nodules (Fig. 4C). All of these observations suggest thatthe first stages of the infection and nodule organogenesis pro-cess are not affected by the ribBX mutation but that, at laterstages, the infection cannot persist and nodules senesce pre-maturely. To investigate the infection process in more detail, wehave transformed the ORS285 ribBX mutant with a plasmidthat expresses superyellow fluorescent protein (sYFP) consti-tutively. A. evenia plants were infected with this sYFP-taggedstrain and nodules present at 7 dpi were analyzed by confocalmicroscopy. This early time point of analysis was chosen toobserve infection stages before nodule senescence in ribBX-infected plants emerged. At 7 dpi, plants infected by the WT
and ribBX strain contained mature-sized nodules in which plantcells in the central tissue were uniformly infected by bacteria(Fig. 4I and K). However, whereas WT bacteria in these nod-ules are differentiated into spherical bacteroids, they have anelongated shape in RibBX nodules (Fig. 4J versus L). Thisindicates that, in the absence of the RibBX protein, ORS285cells are unable to differentiate into the spherical bacteroidsthat are characteristic for the symbiotic interaction with NF-independent Aeschynomene legumes.As indicated above, ribBX encodes for a protein with a 3,4-
DHBP synthase domain and a RibX domain with unknownfunction. To analyze whether both domains are required for thesymbiotic interaction of the ORS285 strain with A. evenia, theribBX mutant strain was transformed with plasmids expressingfull-length RibBX (pribBX), the 3,4-DHBP synthase domain(pribB), and the RibX domain (pribX). Plasmids expressingfull-length RibBX and the RibB domain were able to comple-ment the symbiotic phenotype of the ribBX mutant strain (Fig.5A to D). This indicates that the RibB domain in RibBX issufficient to obtain an efficient symbiotic interaction of ORS285with A. evenia plants.The Aeschynomene spp. that establish NF-independent
symbiosis form a single clade that can be split into four sub-groups (Chaintreuil et al. 2018). To investigate whether therequirement for a functional ribBX gene as found for A. eveniais a generality within this clade, we infected one species of eachsubgroup with the ribBX mutant strain. With all analyzedspecies, plants infected with the ribBX mutant strain had (i)nitrogen starvation symptoms, (ii) an increase in number ofnodules as compared with plants infected with the WT, and (iii)low nitrogenase enzyme activity (Table 1). These data suggestthat the functionality of the ribBX gene is obligatory for anefficient symbiotic interaction with NF-independent Aeschy-nomene spp.
In NF-dependent Aeschynomene spp., RibBX requirementdepends on the host plant.We also investigated the consequence of inactivating the
ribBX, gcdH, BRAD5118, or gntR-like gene on A. afrasperaplants, an Aeschynomene spp. that uses an NF-dependent pro-cess for symbiosis. Another characteristic of the symbiosisbetween ORS285 and A. afraspera is that, in this interaction,the ORS285 cells differentiate into elongated bacteroids(elongated bacteroids are characteristic for the interaction ofORS285 with all NF-dependent Aeschynomene host plants).For this, we infected A. afraspera with the different mutantstrains and observed the effect on plant growth, nodule number,and nitrogenase enzyme activity at 20 dpi. In all analyzedphenotypic aspects, plants infected with the mutant strains wereindistinguishable from plants infected with the WT strain (Fig.6A to D). To analyze whether the ribBX gene is expressed inA. afraspera, we also inoculated this NF-dependent Aeschy-nomene sp. with the reporter strain, in which the gusA gene isplaced under the control of the promoter region of the ribBXgene. As in nodules from A. evenia plants, nodules fromA. afraspera plants infected with this reporter strain had highGUS activity, in contrast to nodules induced by the WT strain(Fig. 6E and F). Although 5-bromo-4-chloro-3-indoxyl-b-D-glucuronide cyclohexylammonium salt staining is a qualitativemeasurement for gene expression, we want to note that, by eyeand under the same experimental conditions, no difference incolor between the nodules of A. evenia and A. afraspera couldbe detected (Figs. 4C and 6F). This indicates that there is nocorrelation between the RibBX requirement for symbiosis andthe expression in the nodules. The absence of an effect with theribBX strain in A. afraspera plants was surprising because, withall tested NF-independent Aeschynomene spp., the symbiotic
Fig. 5. Expression of the 3,4-dihydroxybutanone phosphate synthase do-main complements the symbiotic phenotype of the ORS285 ribBX mutantstrain. A, Comparison of the growth of Aeschynomene evenia plants in-oculated with wild-type (wt) ORS285, ORS285 ribBX, ORS285 ribBX +pMG105 PribBX-ribBX, ORS285 ribBX + pMG105 PribBX-ribB, andORS285 ribBX + pMG105 PribBX-ribA. Noninoculated plants (ni) wereused as control. B, Mature nodules on A. evenia plants inoculated withORS285, ORS285 ribBX, ORS285 ribBX + pMG105 PribBX-ribBX,ORS285 ribBX + pMG105 PribBX-ribB, and ORS285 ribBX + pMG105PribBX-ribA. C, Number of root nodules on A. evenia plants inoculated withORS285, ORS285 ribBX, ORS285 ribBX + pMG105 PribBX-ribBX,ORS285 ribBX + pMG105 PribBX-ribB, and ORS285 ribBX + pMG105PribBX-ribA. The mean number of nodules per plant (n = 5) at 23 dayspostinfection (dpi) is presented. D,Acetylene-reducing activity of A. eveniaplants inoculated with ORS285, ORS285 ribBX, ORS285 ribBX + pMG105PribBX-ribBX, ORS285 ribBX + pMG105 PribBX-ribB, and ORS285 ribBX +pMG105 PribBX-ribA at 23 dpi. The mean amount of produced ethylene perhour and per plant (n = 5) is indicated. Error bars (C and D) representstandard errors of the mean and letters represent conditions with significantdifference according to Tukey’s test (P < 0.05).
92 / Molecular Plant-Microbe Interactions

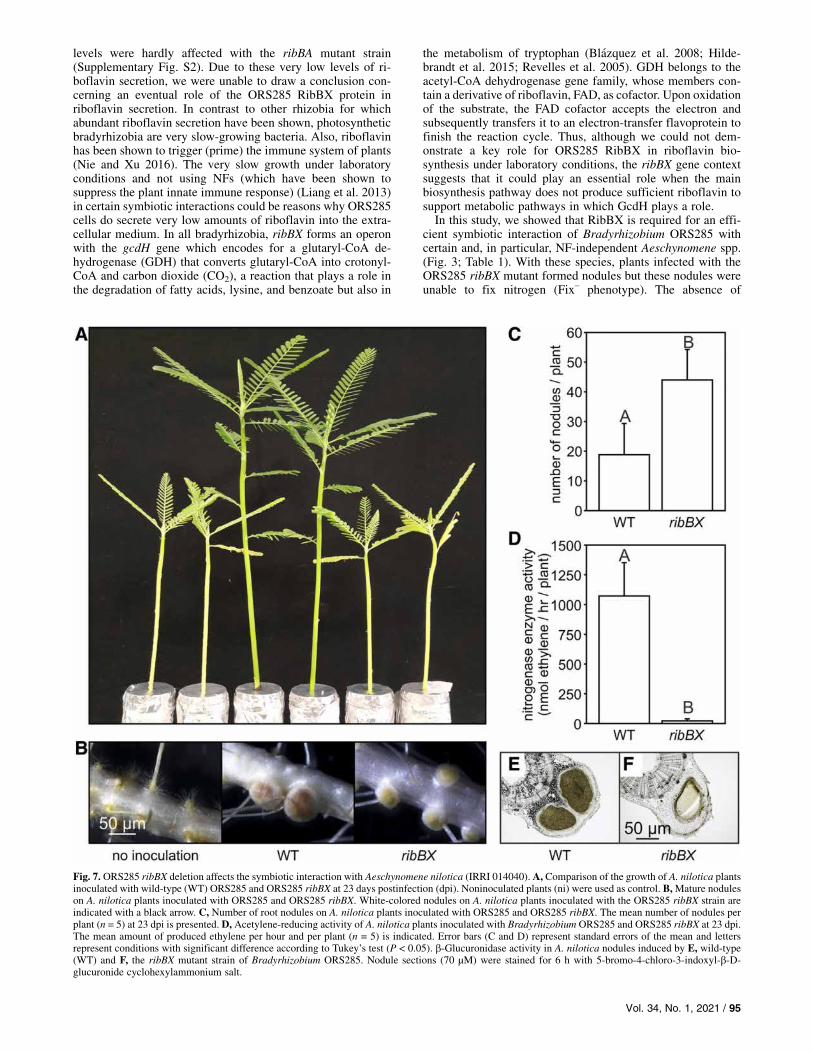
interaction of this mutant was drastically affected (Fig. 3;Table 1). For this reason, we tested the ribBX mutant withanother NF-dependent Aeschynomene sp., A. nilotica, whichestablishes an efficient symbiosis with the WT ORS285 strain.In contrast to A. afraspera, A. nilotica plants had nitrogenstarvation symptoms, increased nodule numbers, and no ni-trogenase enzyme activity when infected with the ribBXmutant(Fig. 7A to D). This indicates that, in the group of NF-dependent Aeschynomene spp., the requirement of RibBX ac-tivity to establish an efficient symbiosis varies with the hostplant.To investigate the contrasting phenotype of the ribBX mutant
in symbiosis with A. afraspera and A. nilotica in more detail,we used the YFP-tagged version of the WT and ribBX mutantstrain to study the bacteroid differentiation. In nodules of bothNF-dependent Aeschynomene spp., ORS285 ribBXmutant cellsdifferentiated like the WT cells into elongated bacteroids (Fig.8B, D, F, and H). However, despite the differentiation of theribBX mutant into elongated bacteroids in A. nilotica plants asobserved at 7 dpi, most of the ribBX mutant-induced nodulesobserved at 21 dpi were hollow inside and displayed no GUSactivity (Fig. 7F). This indicates that, despite the differentiationof the ribBX mutant into elongated bacteroids, the bacterialinfection was not maintained and the nodules senesced veryshortly after their formation.
DISCUSSION
The riboflavin biosynthesis pathway has been extensivelystudied, and all enzymes that compose this pathway are thoughtto be identified and well conserved in bacteria. Similar to otherbacteria, the Bradyrhizobium ORS285 strain contains sequencehomologs of the RibA and RibB enzymes that catalyze theinitial step of two separate reactions that converge in the ri-boflavin synthesis pathway (Fig. 1A). However, as found insome bacteria, the RibB and RibA domains are fused andencoded by a single gene that was annotated as ribBA. Here, weshow that the growth of an ORS285 strain in which the ribBAgene is inactivated is indistinguishable from the growth of theWT strain, indicating that the ribBA mutant still producedsufficient riboflavin to sustain growth (Fig. 1E and F). In ad-dition, sequence analysis of the RibA domain and comple-mentation experiments using an E. coli ribA mutant strainindicated that this domain does not have GTP cyclohydrolase IIactivity (Fig. 2A and B). The latter has also been observed forribBA homologs from Shewanella oneidensis, Pseudomonasputida, Vibrio parahaemolyticus, and Burkholderia cen-ocepacia which, for that reason, have been renamed as ribBX
(Brutinel et al. 2013). In accordance, we have renamed theORS285 ribBA gene ribBX and the domain with homology toRibA the RibX domain. For S. oneidensis RibBX, it has beenshown that the presence of the RibX domain decreases the 3,4-DHBP synthase activity of the RibB domain (Brutinel et al.2013). In addition, recent experiments using purified Acineto-bacter baumannii RibBX showed that FMN (one of the endproducts of riboflavin biosynthesis) drastically inhibits the 3,4-DHBP synthase activity (Wang et al. 2019). Structural modelingof FMN on the A. baumannii RibBX crystal structure indicatedthat the RibX domain can bind FMN. Based on this finding, it ishypothesized that FMN binding to the RibX domain allosteri-cally regulates the enzyme activity of the RibB domain (Wanget al. 2019). These results indicate that the RibX domain has aregulatory function and explains why some of these domainsoriginally annotated as “RibA” do not display the predicted GTPcyclohydrolase II enzyme activity.The above results, and the absence of other genes encoding
proteins with homology to RibA and RibB, suggest thatORS285 has other unknown enzymes with GTP cyclohydrolaseII and 3,4-DHBP synthase activity or uses a different bio-synthesis pathway to produce riboflavin. With respect to theRibB enzyme activity, studies have indicated that diacetyl (=butanedione/butane-2,3-dione) is able to replace 3,4-dihy-droxybutanone in the riboflavin biosynthesis pathway (Cross-ley et al. 2007; Nakajima and Mitsuda 1984). Diacetyl issynthesized from pyruvate and the first step of the biosynthesispathway is mediated by thiamine pyrophosphate (TPP). In-terestingly, in Bradyrhizobium ORS285 and several otherbacteria, the gene thiL, encoding the enzyme catalyzing the laststep in TPP biosynthesis, is found close to rib genes (Fig. 1B).Thus, using diacetyl instead of 3,4-dihydroxybutanone to syn-thesize riboflavin could, in part, explain why the absence of3,4-DHBP synthase activity in the ORS285 ribBX strain doesnot lead to riboflavin auxotrophy. Interestingly, an Ensifermeliloti ribBAmutant has also been shown not to be a riboflavinauxotroph and no additional ribBA, ribA, or ribB genes could beidentified (Yurgel et al. 2014). Thus, the presence of an un-conventional route or alternative enzymes to synthesize ribo-flavin could be more common in rhizobia and requires furtherresearch in the future.In several bacteria, ribBA/ribBX mutants have been shown to
secrete less or no riboflavin into the extracellular medium(Brutinel et al. 2013; Worst et al. 1998; Yang et al. 2002; Yurgelet al. 2014). In contrast to E. meliloti, laboratory-grownORS285 WT cells secrete very low levels of riboflavin into theculture medium (riboflavin fluorescence just above the fluo-rescent levels of the noninoculated growth medium), and these
Table 1. Characteristics of the symbiotic interaction between the wild type (WT) and the ribBX mutant of Bradyrhizobium ORS285 and different Nod-factor(NF)-independent and NF-dependent Aeschynomene spp.a
Growthstimulation plants
Mean number ofnodules/plant Leghemoglobin
Nitrogenase enzymeactivity (nmol
ethylene/plant/h)
Plant species WT ribBX WT ribBX WT ribBX WT ribBX
NF-independentAeschynomene evenia +++ _ 29 ± 4 56 ± 8 Yes No 450 ± 34 4 ± 1A. sensitiva +++ _ 29 ± 5 43 ± 4 Yes No 319 ± 91 83 ± 30A. deamii +++ _ 48 ± 8 53 ± 4 Yes No 618 ± 293 162 ± 70A. tambacoudensis +++ _ 8 ± 3 20 ± 11 Yes No 70 ± 30 nd
NF-dependentA. afraspera +++ +++ 25 ± 2 25 ± 1 Yes Yes 702 ± 33 790 ± 96A. nilotica +++ _ 12 ± 3 42 ± 2 Yes No 577 ± 106 33 ± 12
a Seedlings of different Aeschynomene spp. were inoculated with wild-type (WT) ORS285 and ORS285 ribBX. At 23 days postinfection (dpi), the plant growthwas compared with noninoculated control plants and the mean nodule number, mature nodule phenotype, and mean nitrogenase enzyme activity, as analyzedby the acetylene reduction assay, was determined. Symbols: +++ indicates no nitrogen-starvation signs and plants are much better developed than thenoninoculated control plants, _ indicates no difference with noninoculated control plants, nd = not detectable, and ± indicates the standard error of the mean.
Vol. 34, No. 1, 2021 / 93
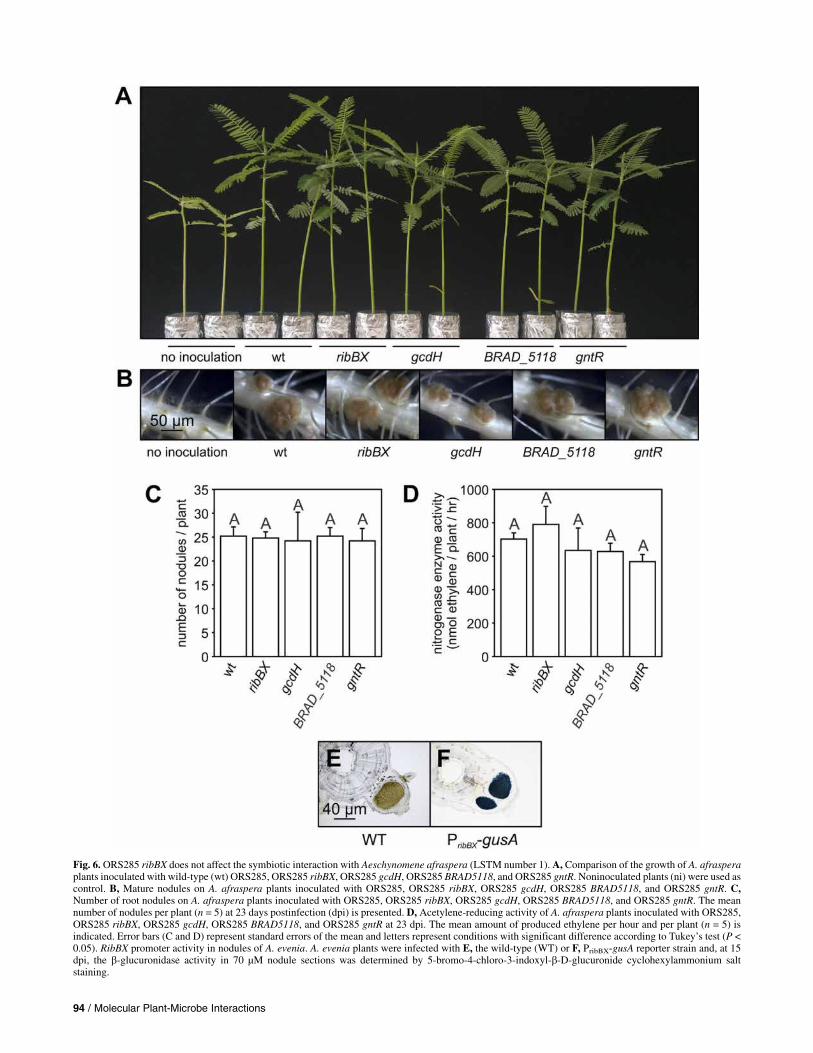
Fig. 6. ORS285 ribBX does not affect the symbiotic interaction with Aeschynomene afraspera (LSTM number 1).A, Comparison of the growth of A. afrasperaplants inoculated with wild-type (wt) ORS285, ORS285 ribBX, ORS285 gcdH, ORS285 BRAD5118, and ORS285 gntR. Noninoculated plants (ni) were used ascontrol. B, Mature nodules on A. afraspera plants inoculated with ORS285, ORS285 ribBX, ORS285 gcdH, ORS285 BRAD5118, and ORS285 gntR. C,Number of root nodules on A. afraspera plants inoculated with ORS285, ORS285 ribBX, ORS285 gcdH, ORS285 BRAD5118, and ORS285 gntR. The meannumber of nodules per plant (n = 5) at 23 days postinfection (dpi) is presented. D, Acetylene-reducing activity of A. afraspera plants inoculated with ORS285,ORS285 ribBX, ORS285 gcdH, ORS285 BRAD5118, and ORS285 gntR at 23 dpi. The mean amount of produced ethylene per hour and per plant (n = 5) isindicated. Error bars (C and D) represent standard errors of the mean and letters represent conditions with significant difference according to Tukey’s test (P <0.05). RibBX promoter activity in nodules of A. evenia. A. evenia plants were infected with E, the wild-type (WT) or F, PribBX-gusA reporter strain and, at 15dpi, the b-glucuronidase activity in 70 µM nodule sections was determined by 5-bromo-4-chloro-3-indoxyl-b-D-glucuronide cyclohexylammonium saltstaining.
94 / Molecular Plant-Microbe Interactions
levels were hardly affected with the ribBA mutant strain(Supplementary Fig. S2). Due to these very low levels of ri-boflavin secretion, we were unable to draw a conclusion con-cerning an eventual role of the ORS285 RibBX protein inriboflavin secretion. In contrast to other rhizobia for whichabundant riboflavin secretion have been shown, photosyntheticbradyrhizobia are very slow-growing bacteria. Also, riboflavinhas been shown to trigger (prime) the immune system of plants(Nie and Xu 2016). The very slow growth under laboratoryconditions and not using NFs (which have been shown tosuppress the plant innate immune response) (Liang et al. 2013)in certain symbiotic interactions could be reasons why ORS285cells do secrete very low amounts of riboflavin into the extra-cellular medium. In all bradyrhizobia, ribBX forms an operonwith the gcdH gene which encodes for a glutaryl-CoA de-hydrogenase (GDH) that converts glutaryl-CoA into crotonyl-CoA and carbon dioxide (CO2), a reaction that plays a role inthe degradation of fatty acids, lysine, and benzoate but also in
the metabolism of tryptophan (Blazquez et al. 2008; Hilde-brandt et al. 2015; Revelles et al. 2005). GDH belongs to theacetyl-CoA dehydrogenase gene family, whose members con-tain a derivative of riboflavin, FAD, as cofactor. Upon oxidationof the substrate, the FAD cofactor accepts the electron andsubsequently transfers it to an electron-transfer flavoprotein tofinish the reaction cycle. Thus, although we could not dem-onstrate a key role for ORS285 RibBX in riboflavin bio-synthesis under laboratory conditions, the ribBX gene contextsuggests that it could play an essential role when the mainbiosynthesis pathway does not produce sufficient riboflavin tosupport metabolic pathways in which GcdH plays a role.In this study, we showed that RibBX is required for an effi-
cient symbiotic interaction of Bradyrhizobium ORS285 withcertain and, in particular, NF-independent Aeschynomene spp.(Fig. 3; Table 1). With these species, plants infected with theORS285 ribBX mutant formed nodules but these nodules wereunable to fix nitrogen (Fix
_phenotype). The absence of
Fig. 7. ORS285 ribBX deletion affects the symbiotic interaction with Aeschynomene nilotica (IRRI 014040). A, Comparison of the growth of A. nilotica plantsinoculated with wild-type (WT) ORS285 and ORS285 ribBX at 23 days postinfection (dpi). Noninoculated plants (ni) were used as control. B,Mature noduleson A. nilotica plants inoculated with ORS285 and ORS285 ribBX. White-colored nodules on A. nilotica plants inoculated with the ORS285 ribBX strain areindicated with a black arrow. C, Number of root nodules on A. nilotica plants inoculated with ORS285 and ORS285 ribBX. The mean number of nodules perplant (n = 5) at 23 dpi is presented. D, Acetylene-reducing activity of A. nilotica plants inoculated with Bradyrhizobium ORS285 and ORS285 ribBX at 23 dpi.The mean amount of produced ethylene per hour and per plant (n = 5) is indicated. Error bars (C and D) represent standard errors of the mean and lettersrepresent conditions with significant difference according to Tukey’s test (P < 0.05). b-Glucuronidase activity in A. nilotica nodules induced by E, wild-type(WT) and F, the ribBX mutant strain of Bradyrhizobium ORS285. Nodule sections (70 µM) were stained for 6 h with 5-bromo-4-chloro-3-indoxyl-b-D-glucuronide cyclohexylammonium salt.
Vol. 34, No. 1, 2021 / 95

nitrogenase enzyme activity seems not be related to a dys-function of the nitrogenase enzyme complex itself because theORS285 ribBXmutant is able to fix nitrogen when grown underfree-living conditions (Fig. 4A). The Fix
_phenotype of the
ribBX mutant could be complemented by expressing the com-plete RibBX protein or the RibB domain but not by the RibXdomain (Fig. 5). This suggests that the observed symbioticdefect was due to the absence of the RibB domain alone. Mi-croscopic analysis of NF-independent Aeschynomene evenianodules showed that the ribBX mutant infects plant cells in-tracellularly but that the bacteria did not differentiate intospherical bacteroids, which are characteristic for NF-independentAeschynomene spp. (Fig. 4L). In addition, the nodules senescedearly and, at 21 dpi, the majority of nodules on ribBX-infectedplants were completely devoid of bacteria (Fig. 4E). Thus,ORS285 RibBX or, rather, the RibB domain seems to be requiredfor the persistence of the intracellular infection in Aeschynomeneplants. Interestingly, a link between riboflavin biosynthesis andintracellular survival in eukaryotic cells has previously beenshown for Brucella abortus (Bonomi et al. 2010). As indicatedabove, the gene context of ribBX suggests that riboflavin bio-synthesis via RibBX is employed when the bacterium uses met-abolic pathways that require GcdH enzyme activity. However,inactivation of the gcdH gene had no effect on the symbioticinteraction of ORS285 with Aeschynomene legumes (Figs. 3 and6). In this context, it should be noted that BradyrhizobiumORS285, in addition to gcdH, contains many other genes thatencode for proteins which belong to the same gene family asGcdH and of which the function is unknown. Thus, it is highlypossible that the absence of a symbiotic phenotype for the gcdHmutant is due to functional redundancy and that one of theseputative acetyl-CoA dehydrogenases converts the formed glutaryl-CoA during symbiosis. Alternatively, Bradyrhizobium ORS285could contain an additional glutarate catabolic pathway, as re-cently discovered in P. putida KT2440. In this pathway, glutarateis not acetylated into glutaryl-CoA but in a reaction with 2-keto-glutarate converted into L-2-hydroxyglutarate and succinate(Zhang et al. 2018). Taken together, as was done by the authors inthe B. abortus study, at this stage we are unable to pinpoint aspecific biological process during the intracellular infection forwhich riboflavin biosynthesis is required.Intriguingly, when the NF-dependent Aeschynomene sp.
A. afraspera was infected with the ORS285 ribBX mutant, nosymbiotic defect was observed (Fig. 6). This means that ORS285does not need RibBX activity for interacting with A. afraspera,
or that this plant species supplies the bacterium with the requiredriboflavin. A similar difference in symbiotic interaction haspreviously described for an ORS285 nifV mutant (Nouwenet al. 2017). In this case, we were able to demonstrate thatA. afraspera expresses a nodule-specific homocitrate synthase thatprovides the bacteria with homocitrate so that it can form an activenitrogenase enzyme complex. Plants synthesize riboflavin usingenzymes that are similar to the ones found in bacteria (Fischer andBacher 2006). Analysis of genomic and transcriptomic data ofA. evenia and A. afraspera showed that they have multiple copiesof genes encoding RibBA-like proteins (Supplementary Table S4).In both A. evenia and A. afraspera, some of these genes showed asomewhat increased expression in nodule tissue. However, none ofthe RibBA orthologs showed a nodule-specific expression profilesuch as we have found for FEN1 (Nouwen et al. 2017). Thus, thenodule-specific expression of an RibBA ortholog in A. afrasperato provide riboflavin to the symbiotic bacterium is likely not thereason why no effect is observed in the interaction of this plantspecies with the ORS285 ribBX mutant.The nonrequirement of bacterial RibBX in symbiosis is not a
universal characteristic in NF-dependent Aeschynomene spp.because, in the second species tested, A. nilotica, the ribBXmutation had a drastic negative impact on the symbiotic in-teraction (Fig. 7). Such variation has also been observed, forinstance, for the interaction of an R. trifolii riboflavin auxotrophwith different clover species (genus Trifolium) (Pankhurst et al.1974; Schwinghamer 1970). This indicates that bacterial ribo-flavin requirements in certain NF-dependent interactions aredependent on the host plant genotype. As an essential compo-nent of flavoproteins, riboflavin derivatives play importantroles in many different physiological processes. Consequently,environmental factors such as oxidative or nitrosative stress,iron or oxygen availability, available growth substrates, andeventual toxic compounds to be inactivated are expected tohave a great impact on how much riboflavin has to be synthe-sized by the bacterium. A specific environmental condition inthe nodule tissue of certain Aeschynomene spp. could mean thatthe bacterium requires the RibBX protein to increase the totalamount of riboflavin synthesized or, alternatively, to compen-sate for a reduced synthesis via a thus-far-unidentified pathway.Given the complexity of the rhizobium–legume interaction, weexpect that the total amount of riboflavin required during sym-biosis (and the pathway used to synthesize it) is determined bymultiple factors. This could explain the host-dependent pheno-type as observed in NF-dependent symbiosis but also means thatthe existence of NF-independent interactions in which bacterialRibBX is not required cannot be completely excluded.Taken together, this study shows that the RibBX protein
plays an important role in the symbiotic interaction of Bra-dyrhizobium ORS285 with the majority of the tested Aeschy-nomene spp. In addition, as found for E. meliloti (Yurgel et al.2014), inactivation of ribB(A)X did not result in riboflavinauxotrophy, whereas no other copies of genes homologous toribA and ribB are present. Future studies are required to obtaina better understanding of riboflavin biosynthesis in rhizobia andbradyrhizobia in general and the requirement of riboflavinduring symbiosis.
MATERIALS AND METHODS
Bacterial strains and growth conditions.Tables of strains and plasmids used in this study can be found
in the supplementary materials (Supplementary Tables S1 andS2). Bradyrhizobium ORS285 (Molouba et al. 1999) and de-rivatives were grown at 28�C in modified yeast extract mannitolmedium (YM) (Giraud et al. 2000) or a minimal BNM-Bmediumto which no riboflavin was added (Podlesakova et al. 2013).
Fig. 8. Confocal microscopic analysis of nodules induced by wild-type(WT) ORS285 and ORS285 ribBX mutant on A to D, Aeschynomeneafraspera and E to H, A. nilotica plants. The ORS285 and ORS285 ribBXbacteriods are elongated in both Aeschynomene spp.
96 / Molecular Plant-Microbe Interactions

Escherichia coli riboflavin auxotrophs (Bandrin et al. 1983)were obtained from the Coli Genetic Stock Center. E. colistrains were grown in Luria-Bertani medium or a minimal M9medium at 37�C. When required, the media were supplementedwith ampicillin (50 to 100 µg/ml), kanamycin (50 to100 µg/ml), cefotaxime (20 µg/ml), spectinomycine (20 to200 µg/ml), or 500 µM riboflavin.
Construction of mutant and reporter strainsof Bradyrhizobium ORS285.All DNA fragments were amplified using primers as listed in
Supplementary Table S3. To construct ribBX, gcdH, BRAD5118,and gntR insertional mutants, 312 to 382 bp of the internal se-quence of each gene was amplified by PCR. The resultingfragments were isolated, cloned into plasmid pGEM-T Easy(Promega Corp.), and transformed into thermocompetent E. coliXL2-Blue cells (Agilent). Correct clones were verified by se-quence analysis. The internal gene fragments in pGEM-T Easywere excised with SalI-BamHI and ligated into suicide vectorpJG194-PnodA-gusA (ribBX, gcdH, and BRAD5118) (Supple-mentary Materials) or pJG194 (gntR) (Griffitts and Long 2008)digested with SalI-BamHI. After transformation into E. coliXL2-Blue cells, correct clones were selected via kanamycinresistance (50 µg/ml) and subsequent DNA restriction enzymeanalysis. To construct a ribBX reporter strain, the 276-bp regionupstream of ribBX was amplified by PCR, cloned into pGEM-TEasy, and transformed into E. coli XL2-Blue cells. After verifi-cation by sequence analysis, the promoter fragment was excisedwith SalI-BamHI and ligated into suicide vector pJG194-PnodA-gusA digested with the same enzymes. After transformationinto E. coli XL2-Blue cells, correct clones were selected asdescribed above. Plasmids were transformed into CaCl2-competent E. coli S17-1 cells (Simon et al. 1983) and mo-bilized into Bradyrhizobium ORS285 using the biparentalmating protocol, as previously described (Podlesakova et al.2013). Correct insertion of the plasmids was verified by PCR.To construct sYFP-tagged derivatives, plasmid pMG105-Paph-sYFP-specR (for construction, see SupplementaryMaterials) was transformed into electrocompetent Bradyrhi-zobium ORS285 (mutant) cells and, after transformation,plasmid-containing clones were selected via spectinomycineresistance (200 µg/ml) and sYFP fluorescence.
Complementation of the ORS285 ribBX mutant.Complementation of the ORS285 ribBX mutant was tested
with different DNA constructs: (i) construct A, consisting of theribBX promoter region plus complete ribBX gene; (ii) constructB, consisting of the ribBX promoter region plus the RibB do-main; and (iii) the ribBX promoter region plus the RibX do-main. These DNA regions were PCR amplified using primersthat can be found in Supplementary Table S3 and cloned intothe pMG105-specR plasmid (Supplementary Materials) thatcontains the spectinomycine resistance gene. The pMG105plasmid replicates and is stable in the ORS285 strain. To con-struct the plasmid in which the RibB domain has been deleted,the promoter region plus the first 12 nucleotides of ribBX andthe RibX domain were PCR amplified and subsequently fusedusing overlapping PCR. The constructed plasmids (pMG105-PribBX-ribBX, pMG105-PribBX-ribB, and pMG105-PribBX-ribX)were introduced into electrocompetent cells of the ORS285ribBX mutant by electroporation. Strains containing plasmidswere selected on YM plates supplemented with spectinomycineat 200 µg/ml and kanamycine at 50 µg/ml.
Complementation of E. coli ribA and ribB mutants.For complementation of E. coli riboflavin auxotroph mu-
tants, ribBX, the ribB domain, and the ribX domain were PCR
amplified from plasmids pMG105-PribBX-ribBX, pMG105-PribBX-ribB, and pMG105-PribBX-ribX, respectively, using pri-mers as indicated in Supplementary Table S3. The resultingfragments were cloned into pGEM-T Easy and transformed intoE. coli XL2-Blue cells, and correct clones were verified usingDNA restriction enzyme analysis and DNA sequencing. Sub-sequently, the genes were excised from pGEM-T Easy byBamHI-HindIII digestion and cloned into plasmid pJF118 E/H(Furste et al. 1986) digested with the same enzymes. Aftertransformation into E. coli XL2-Blue cells, correct clones wereselected using DNA restriction enzyme analysis. For comple-mentation studies, the different plasmids and pJF118 E/H weretransformed into CaCl2-competent cells of BSV11 (ribB11::Tn5) and BSV18 (ribA18::Tn5), respectively.
Plant growth and acetylene reduction assay.Aeschynomene spp. used in this study were A. evenia
(CIAT22838; Malawi), A. deamii (LSTM number 24; Mexico),A. tambacoudensis (LSTM number 60; Senegal), A sensitiva(LSTM number 28; Senegal), A. afraspera (LSTM number 1;Senegal), and A. nilotica (IRRI 014040; Senegal). Sterilizationof seed, germination, plant growth, and inoculation with bac-terial strains were as described (Nouwen et al. 2017). At theindicated times after inoculation (as specified in the figurelegends), photos of plants were taken, the number of nodules onthe roots were counted, and the ARA was used to measure thenitrogenase enzyme activity (Podlesakova et al. 2013). Timepoints of analysis were chosen such that there was a cleardifference in growth phenotype between noninoculated and(WT) inoculated plants.
Microscopy.For confocal microscopy, root sections containing nod-
ules were harvested and rinsed with distilled water. Therinsed root sections were fixed overnight with 4% (wt/vol)paraformaldehyde in phosphate-buffered saline (PBS) atroom temperature. The fixed root sections were washedthree times with PBS, embedded in 5% agar, and sectioned(50 to 70 µM) using a vibratome (VT1000S; Leica, Nanterre,France). Nodules induced by sYFP-tagged strains were an-alyzed as described by Bonaldi et al. (2011). To determinethe GUS activity in nodules formed by the mutant andPribBX-reporter strains, fresh nodules were embedded in 5%agarose and sectioned (50 to 70 µM) using a vibratome(VT1000S; Leica). Nodule sections were incubated for30 min in GUS assay buffer (Bonaldi et al. 2011) at 37�C.After staining, the sections were washed three times withwater, mounted on microscope slides, and observed underbright-field illumination with a macroscope (Nikon AZ100;Champigny-sur-Marne, France).
In vitro nitrogenase enzyme activity.To determine the nitrogenase enzyme activity under free-
living conditions, Bradyrhizobium ORS285 and ORS285ribBX were grown in 9-ml vacuette tubes (reference 455001;Greiner Bio-One) containing 2 ml of BNM-B mediumwithout riboflavin supplementation. To avoid overpressure,0.9 ml of air was removed before injecting the same volumeof 100% acetylene. The cultures were incubated at 28�Cand, at 7 dpi, 1-ml gas samples were analyzed for ethyleneproduction by gas chromatography and the optical density(at 600 nm) of the culture was determined (Nouwen et al.2017).
AUTHOR-RECOMMENDED INTERNET RESOURCE
Coli Genetic Stock Center: https://cgsc.biology.yale.edu
Vol. 34, No. 1, 2021 / 97

LITERATURE CITED
Abbas, C. A., and Sibirny, A. A. 2011. Genetic control of biosynthesis andtransport of riboflavin and flavin nucleotides and construction of robustbiotechnological producers. Microbiol. Mol. Biol. Rev. 75:321-360.
Balasubramanian, R., Levinson, B. T., and Rosenzweig, A. C. 2010.Secretion of flavins by three species of methanotrophic bacteria. Appl.Environ. Microbiol. 76:7356-7358.
Bandrin, S. V., Rabinovich, P. M., and Stepanov, A. I. 1983. Tri gruppystsepleniia genov biosinteza riboflavina Escherichia coli [Three linkagegroups of the genes of riboflavin biosynthesis in Escherichia coli].Genetika 19:1419-1425.
Blazquez, B., Carmona, M., Garcıa, J. L., and Dıaz, E. 2008. Identificationand analysis of a glutaryl-CoA dehydrogenase-encoding gene and itscognate transcriptional regulator from Azoarcus sp. CIB. Environ.Microbiol. 10:474-482.
Bonaldi, K., Gargani, D., Prin, Y., Fardoux, J., Gully, D., Nouwen, N.,Goormachtig, S., and Giraud, E. 2011. Nodulation of Aeschynomeneafraspera and A. indica by photosynthetic Bradyrhizobium Sp. strainORS285: The nod-dependent versus the nod-independent symbioticinteraction. Mol. Plant-Microbe Interact. 24:1359-1371.
Bonaldi, K., Gourion, B., Fardoux, J., Hannibal, L., Cartieaux, F., Boursot,M., Vallenet, D., Chaintreuil, C., Prin, Y., Nouwen, N., and Giraud, E.2010. Large-scale transposon mutagenesis of photosynthetic Bradyrhi-zobium sp. strain ORS278 reveals new genetic loci putatively importantfor nod-independent symbiosis with Aeschynomene indica. Mol. Plant-Microbe Interact. 23:760-770.
Bonomi, H. R., Marchesini, M. I., Klinke, S., Ugalde, J. E., Zylberman, V.,Ugalde, R. A., Comerci, D. J., and Goldbaum, F. A. 2010. An atypicalriboflavin pathway is essential for Brucella abortus virulence. PLoS One5:e9435. doi:
Brutinel, E. D., Dean, A. M., and Gralnick, J. A. 2013. Description of ariboflavin biosynthetic gene variant prevalent in the phylum Proteobac-teria. J. Bacteriol. 195:5479-5486.
Chaintreuil, C., Perrier, X., Martin, G., Fardoux, J., Lewis, G. P., Brottier,L., Rivallan, R., Gomez-Pacheco, M., Bourges, M., Lamy, L., Thibaud,B., Ramanankierana, H., Randriambanona, H., Vandrot, H., Mournet, P.,Giraud, E., and Arrighi, J.-F. 2018. Naturally occurring variations in thenod-independent model legume Aeschynomene evenia and relatives: Aresource for nodulation genetics. BMC Plant Biol. 18:54.
Christie, J. M., Blackwood, L., Petersen, J., and Sullivan, S. 2015. Plantflavoprotein photoreceptors. Plant Cell Physiol. 56:401-413.
Crossley, R. A., Gaskin, D. J. H., Holmes, K., Mulholland, F., Wells, J. M.,Kelly, D. J., van Vliet, A. H. M., and Walton, N. J. 2007. Riboflavinbiosynthesis is associated with assimilatory ferric reduction and ironacquisition by Campylobacter jejuni. Appl. Environ. Microbiol. 73:7819-7825.
Czernic, P., Gully, D., Cartieaux, F., Moulin, L., Guefrachi, I., Patrel, D.,Pierre, O., Fardoux, J., Chaintreuil, C., Nguyen, P., Gressent, F., DaSilva, C., Poulain, J., Wincker, P., Rofidal, V., Hem, S., Barriere, Q.,Arrighi, J.-F., Mergaert, P., and Giraud, E. 2015. Convergent evolution ofendosymbiont differentiation in dalbergioid and inverted repeat-lackingclade legumes mediated by nodule-specific cysteine-rich peptides. PlantPhysiol. 169:1254-1265.
da Silva Neto, J. F., Lourenco, R. F., and Marques, M. V. 2013. Globaltranscriptional response of Caulobacter crescentus to iron availability.BMC Genomics 14:549.
Dunlap, P. 2014. Biochemistry and genetics of bacterial bioluminescence.Adv. Biochem. Eng. Biotechnol. 144:37-64.
Fischer, M., and Bacher, A. 2006. Biosynthesis of vitamin B2 in plants.Physiol. Plant. 126:304-318.
Furste, J. P., Pansegrau, W., Frank, R., Blocker, H., Scholz, P., Bagdasarian,M., and Lanka, E. 1986. Molecular cloning of the plasmid RP4 primaseregion in a multi-host-range tacP expression vector. Gene 48:119-131.
Garcıa-Angulo, V. A. 2017. Overlapping riboflavin supply pathways inbacteria. Crit. Rev. Microbiol. 43:196-209.
Giraud, E., Hannibal, L., Fardoux, J., Vermeglio, A., and Dreyfus, B. 2000.Effect ofBradyrhizobium photosynthesis on stem nodulation ofAeschynomenesensitiva. Proc. Natl. Acad. Sci. U.S.A. 97:14795-14800.
Giraud, E., Moulin, L., Vallenet, D., Barbe, V., Cytryn, E., Avarre, J.-C.,Jaubert, M., Simon, D., Cartieaux, F., Prin, Y., Bena, G., Hannibal, L.,Fardoux, J., Kojadinovic, M., Vuillet, L., Lajus, A., Cruveiller, S., Rouy,Z., Mangenot, S., Segurens, B., Dossat, C., Franck, W. L., Chang, W.-S.,Saunders, E., Bruce, D., Richardson, P., Normand, P., Dreyfus, B.,Pignol, D., Stacey, G., Emerich, D., Vermeglio, A., Medigue, C., andSadowsky, M. 2007. Legumes symbioses: Absence of Nod genes inphotosynthetic bradyrhizobia. Science 316:1307-1312.
Griffitts, J. S., and Long, S. R. 2008. A symbiotic mutant of Sinorhizobiummeliloti reveals a novel genetic pathway involving succinoglycanbiosynthetic functions. Mol. Microbiol. 67:1292-1306.
Haase, I., Grawert, T., Illarionov, B., Bacher, A., and Fischer, M. 2014.Recent advances in riboflavin biosynthesis. Pages 15-40 in: Flavins andFlavoproteins. S. Weber and E. Schleicher, eds. Methods in MolecularBiology, vol. 1146. Humana Press, New York, NY, U.S.A.
Hildebrandt, T. M., Nunes Nesi, A., Araujo, W. L., and Braun, H. P. 2015.Amino acid catabolism in plants. Mol. Plant 8:1563-1579.
Kelly, M. J. S., Ball, L. J., Krieger, C., Yu, Y., Fischer, M., Schiffmann, S.,Schmieder, P., Kuhne, R., Bermel, W., Bacher, A., Richter, G., andOschkinat, H. 2001. The NMR structure of the 47-kDa dimeric enzyme3,4-dihydroxy-2-butanone-4-phosphate synthase and ligand bindingstudies reveal the location of the active site. Proc. Natl. Acad. Sci.U.S.A. 98:13025-13030.
Liang, Y., Cao, Y., Tanaka, K., Thibivilliers, S., Wan, J., Choi, J., Kang, Ch.,Qiu, J., and Stacey, G. 2013. Nonlegumes respond to rhizobial Nod factorsby suppressing the innate immune response. Science 341:1384-1387.
Liao, D.-I., Calabrese, J. C., Wawrzak, Z., Viitanen, P. V., and Jordan, D. B.2001. Crystal structure of 3,4-dihydroxy-2-butanone 4-phosphatesynthase of riboflavin biosynthesis. Structure 9:11-18.
Molouba, F., Lorquin, J., Willems, A., Hoste, B., Giraud, E., Dreyfus, B.,Gillis, M., de Lajudie, P., and Masson-Boivin, C. 1999. Photosyntheticbradyrhizobia from Aeschynomene spp. are specific to stem-nodulatedspecies and form a separate 16S ribosomal DNA restriction fragmentlength polymorphism group. Appl. Environ. Microbiol. 65:3084-3094.
Nakajima, K., and Mitsuda, H. 1984. Possibility of diacetyl and relatedcompounds as the 4-carbon compound necessary for the formation ofriboflavin in Ashbya gossypii. Acta Vitaminol. Enzymol. 6:271-282.
Nie, S., and Xu, H. 2016. Riboflavin-induced disease resistance requires themitogen-activated protein kinases 3 and 6 in Arabidopsis thaliana. PLoSOne 11:e0153175. doi:
Nouwen, N., Arrighi, J.-F., Cartieaux, F., Chaintreuil, C., Gully, D., Klopp,C., and Giraud, E. 2017. The role of rhizobial (NifV) and plant (FEN1)homocitrate synthases in Aeschynomene/photosynthetic Bradyrhizobiumsymbiosis. Sci. Rep. 7:448.
Pankhurst, C. E., Schwinghamer, E. A., Thorne, S. W., and Bergersen, F. J.1974. The flavin content of clovers relative to symbiosis with a riboflavin-requiring mutant of Rhizobium trifoli. Plant Physiol. 53:198-205.
Podlesakova, K., Fardoux, J., Patrel, D., Bonaldi, K., Novak, O., Strnad, M.,Giraud, E., Spıchal, L., and Nouwen, N. 2013. Rhizobial synthesizedcytokinins contribute to but are not essential for the symbiotic interactionbetween photosynthetic Bradyrhizobia and Aeschynomene legumes.Mol. Plant-Microbe Interact. 26:1232-1238.
Ren, J., Kotaka, M., Lockyer, M., Lamb, H. K., Hawkins, A. R., andStammers, D. K. 2005. GTP cyclohydrolase II structure and mechanism.J. Biol. Chem. 280:36912-36919.
Revelles, O., Espinosa-Urgel, M., Fuhrer, T., Sauer, U., and Ramos, J. L.2005. Multiple and interconnected pathways for L-lysine catabolism inPseudomonas putida KT2440. J. Bacteriol. 187:7500-7510.
Rigali, S., Derouaux, A., Giannotta, F., and Dusart, J. 2002. Subdivision ofthe helix-turn-helix GntR family of bacterial regulators in the FadR,HutC, MocR, and YtrA subfamilies. J. Biol. Chem. 277:12507-12515.
Schwinghamer, E. 1970. Requirement for riboflavin for effective symbiosison clover by an auxotrophic mutant strain of Rhizobium trifolii. Aust.J. Biol. Sci. 23:1187.
Simon, R., Priefer, U., and Puhler, A. 1983. A broad host range mobilizationsystem for in vivo genetic engineering: Transposon mutagenesis in Gramnegative bacteria. Bio/Technology 1:784-791.
Tagua, V. G., Pausch, M., Eckel, M., Gutierrez, G., Miralles-Duran, A.,Sanz, C., Eslava, A. P., Pokorny, R., Corrochano, L. M., and Batschauer,A. 2015. Fungal cryptochrome with DNA repair activity reveals an earlystage in cryptochrome evolution. Proc. Natl. Acad. Sci. U.S.A. 112:15130-15135.
Tan, Y., Liu, Z.-Y., Liu, Z., Zheng, H.-J., and Li, F.-L. 2015. Comparativetranscriptome analysis between csrA-disruption Clostridium acetobuty-licum and its parent strain. Mol. Biosyst. 11:1434-1442.
Taniguchi, H., and Wendisch, V. F. 2015. Exploring the role of sigma factorgene expression on production by Corynebacterium glutamicum: Sigmafactor H and FMN as example. Front. Microbiol. 6:740.
Wang, C., Reid, J. B., and Foo, E. 2018. The art of self-control—Autoregulation of plant–microbe symbioses. Front. Plant Sci. 9:988.
Wang, J., Lonergan, Z. R., Gonzalez-Gutierrez, G., Nairn, B. L., Maxwell,C. N., Zhang, Y., Andreini, C., Karty, J. A., Chazin, W. J., Trinidad, J. C.,Skaar, E. P., and Giedroc, D. P. 2019. Multi-metal restriction byCalprotectin impacts de novo flavin biosynthesis in Acinetobacterbaumannii. Cell Chem. Biol. 26:745-755.e7.
98 / Molecular Plant-Microbe Interactions

Winkler, W. C., Cohen-Chalamish, S., and Breaker, R. R. 2002. An mRNAstructure that controls gene expression by binding FMN. Proc. Natl.Acad. Sci. U.S.A. 99:15908-15913.
Worst, D. J., Gerrits, M. M., Vandenbroucke-Grauls, C. M. J. E., andKusters, J. G. 1998. Helicobacter pylori ribBA-mediated riboflavinproduction is involved in iron acquisition. J. Bacteriol. 180:1473-1479.
Yang, G., Bhuvaneswari, T. V., Joseph, C. M., King, M. D., and Phillips,D. A. 2002. Roles for riboflavin in the Sinorhizobium-alfalfa association.Mol. Plant-Microbe Interact. 15:456-462.
Yurgel, S. N., Rice, J., Domreis, E., Lynch, J., Sa, N., Qamar, Z., Rajamani,S., Gao, M., Roje, S., and Bauer, W. D. 2014. Sinorhizobium melilotiflavin secretion and bacteria-host interaction: Role of the bifunctionalRibBA protein. Mol. Plant-Microbe Interact. 27:437-445.
Zhang, M., Gao, C., Guo, X., Guo, S., Kang, Z., Xiao, D., Yan, J., Tao, F.,Zhang, W., Dong, W., Liu, P., Yang, C., Ma, C., and Xu, P. 2018.Increased glutarate production by blocking the glutaryl-CoA dehydro-genation pathway and a catabolic pathway involving L-2-hydroxyglu-tarate. Nat. Commun. 9:2114.
Vol. 34, No. 1, 2021 / 99


























![80 Great Ideas for Role Plays[1]](https://img.pdfslide.us/doc/110x75/577d2a191a28ab4e1ea8a6e9/80-great-ideas-for-role-plays1.jpg)


