Embed Size (px)
Citation preview

ENVIRONMENTAL BIOTECHNOLOGY
Rhodococcus lactonase with organophosphate hydrolase (OPH)activity and His6-tagged OPH with lactonase activity: evolutionaryproximity of the enzymes and new possibilities in their application
Mariia Sirotkina & Elena N. Efremenko
Received: 20 May 2013 /Revised: 30 August 2013 /Accepted: 9 September 2013 /Published online: 22 September 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Decontamination of soils with complex pollutionusing natural strains of microorganisms is a matter of greatimportance. Here we report that oil-oxidizing bacteriaRhodococcus erythropolis AC-1514D and Rhodococcusruber AC-1513D can degrade various organophosphorouspesticides (OP). Cell-mediated degradation of five dif-ferent OP is apparently associated with the presence ofN -acylhomoserine lactonase, which is pronouncedly simi-lar (46–50 %) to the well-known enzyme organophosphatehydrolase (OPH), a hydrolysis catalyst for a wide variety oforganophosphorous compounds. Additionally, we demon-strated the high lactonase activity of hexahistidine-taggedorganophosphate hydrolase (His6-OPH) with respect to vari-ous N-acylhomoserine lactones, and we determined the cata-lytic constants of His6-OPH towards these compounds. Theseexperimental data and theoretical analysis confirmed the hy-pothesis about the evolutionary proximity of OPH andlactonases. Using Rhodococcus cells, we carried out effectivesimultaneous biodegradation of pesticide paraoxon (88 mg/kg) and oil hydrocarbon hexadecane (6.3 g/kg) in the soil.Furthermore, the discovered high lactonase activity of His6-OPH offers new possibilities for developing an efficient strat-egy of combating resistant populations of Gram-negativebacterial cells.
Keywords Organophosphate hydrolase . Lactonase .
Evolution . Organophosphorous pesticides . Bacterialdegradation
Introduction
According to the Food and Agriculture Organization (FAO) ofthe UN, worldwide annual crop losses caused by weeds andinsect pests make up approximately 35 % of the potentialoverall production volume and are estimated at over US$75billion (Fleseriu et al. 2010). Annual pesticide use in theagricultural industry allows the saving of a substantial partof the harvest and reduces losses.
Organophosphorous pesticides account for approximately40 % of the global production of all agricultural pesticides(Grube et al. 2011; Singh 2009). Due to the fact that degrada-tion of organophosphorous pesticides (OP) in the environmentis slow, they accumulate in soils due to the treatment ofagricultural areas. They are subsequently washed out and entergroundwater and rivers (Hopkins et al. 2000), representing ahealth hazard (Eddleston et al. 2008; Hayden et al. 2010;Reigart and Roberts 1999).
Accumulation of excessive OP levels in soil necessitatesthe use of efficient elimination techniques. From the environ-mental and economic standpoint, it is best to degrade OP insitu using high-performance biocatalysts, which may includeenzymes and microorganisms exhibiting a destructive effectwith respect to toxic OP.
Organophosphate hydrolase (OPH, EC 3.1.8.1) is a well-known enzyme of Pseudomonas diminuta MG that catalyzesthe hydrolysis of various organophosphorous compounds withP―O, P―S, and P―F bonds (Efremenko and Sergeeva 2001).A variety of OPH with a hexahistidine sequence at the Nterminus of the molecule (His6-OPH) is a much more efficientcatalyst for a number of substrates as compared to the nativeOPH (Efremenko et al. 2005a, 2007; Votchitseva et al. 2006).
His6-OPH and cellulose-containing carriers (straw, saw-dust) were used to develop an immobilized biocatalyst to beemployed for detoxifying soils contaminated with various OP(Sirotkina et al. 2012). However, the use of such a biocatalyst
M. Sirotkina : E. N. Efremenko (*)Moscow State University, Lenin’s Hills 1/11, 119991 Moscow,Russian Federatione-mail: [email protected]
Appl Microbiol Biotechnol (2014) 98:2647–2656DOI 10.1007/s00253-013-5233-y

is only advisable for ultrarapid soil remediation or for re-moving high concentrations of contaminants (>500 mg/g)that are difficult or impossible to eliminate using other cata-lysts. Decontamination of soils with moderate OP levels(<500 mg/g) calls for using natural strains of microorganismsexhibiting OPH activity that can efficiently detoxify soilswhile retaining long-term viability and metabolic activityunder ambient conditions. Thus, the problem of selectingnatural microorganism strains that exhibit OPH activity andare capable of eliminating OP is an important one.
Products based on Rhodococcus spp. cells, which areundergoing approval for introduction into natural ecosystems,have been reportedly used for bioremediation of soils contam-inated with oil hydrocarbons (Efremenko et al. 2005b;Murygina et al. 2005).
Owing to this, the discovery of OPH activity in these cellswould offer new opportunities for environmentally safe com-prehensive bioremediation of ecosystems simultaneously con-taminated with OP and oil hydrocarbons. It is very importantfrom the practical standpoint since these microorganisms arewell adaptable to diverse living conditions and form a part ofmany biocenoses (Efremenko et al. 2005b). Meanwhile, theprocess of OP degradation could be controlled using knowl-edge about the functioning of enzymatic systems in cells ofthe genus Rhodococcus .
It must be noted that the OPH activity of Rhodococcuscells with respect to various OP is almost unexplored. Thus,there is only one report mentioning an enzyme present inRhodococcus erythropolis W2 cells that slowly decomposedparaoxon (Afriat et al. 2006). However, it is difficult to assessthe OP degrading potential ofRhodococcus cells based on thisfact alone since multiple other sources of enzymes capable ofcatalyzing the hydrolysis of a single organophosphorous sub-strate have been reported (Efremenko and Varfolomeev 2004).This capability alone, however, is insufficient for applyingsuch enzymes and the microorganisms containing them to OPbiodegradation in an efficient and well-grounded manner.
The goal of this work was to evaluate the ability ofRhodococcus ruber AC-1513D and R. erythropolis AC-1514Dcells catalyzing the bioremediation of oil-contaminated soils todegrade various OP. This goal included determining the enzy-matic systems responsible for OP degradation in these cells.
Materials and methods
Cell cultivation and His6-OPH isolation
Recombinant Escherichia coli strain SG13009[pREP4]transformed by plasmid pTES-His-OPH encoding His6-OPH(Efremenko et al. 2005a) was used as a source for enzyme
production. The cells were cultivated, and the enzyme wasisolated as previously published (Efremenko et al. 2006).
Oil-oxidizing bacterial cells of R. erythropolis VKM AC-1514D and R. ruber VKMAC-1513D (Murygina et al. 2000),obtained from the All-Russian Collection of Microorganisms(VKM), were used in this work. To accumulate cell biomass,the following medium (pH 6.7–7.2) was used for both cul-tures: glucose 10.0 g/L, yeast extract 1.0 g/L, Na citrate 5.0 g/L, KH2PO4 0.5 g/L, K2HPO4·3H2O 1.0 g/L, NaH2PO4·2H2O0.5 g/L, (NH4)2SO4 1.0 g/L, MgSO4·7H2O 0.1 g/L, CaCl2·6H2O 0.05 g/L, and FeSO4·7H2O 0.02 g/L. pH was adjustedby addition of Na2CO3. Bacteria were cultivated in 200 mL ofmedium in Erlenmeyer flasks (750 mL) on a shaker (AdolfKuhner AG, Switzerland) under aerobic conditions with con-stant agitation (180 rpm) at 28 °C. Cells were separated fromthe medium by centrifugation at 12,000×g over 15 min(Beckman J2-21 centrifuge, USA) and used to prepare cellsuspensions for experiments.
OPH activity of R. erythropolis AC-1514D and R. ruberAC-1513D cells
To determine the OPH activity of Rhodococcus cells, 200 mLof a minimal medium containing 0.2 mM OP (paraoxon,malathion, diazinon, dimethoate, or demeton-S purchasedfrom Sigma, USA) and 20 g/L of R. erythropolis AC-1514Dor R. ruber AC-1513D cells were added to 750-mL flasks.The flasks were then placed on a temperature-controlled shak-er and vigorously agitated (180 rpm). The concentration of OPin the culture medium with cells was determined chromato-graphically every 3 h over a period of 24 h. The data were usedto calculate the initial rate and the degree of OP degradation byR. erythropolis AC-1514D and R. ruber AC-1513D cells.
Chromatographic analysis of OP and hexadecane
Ethyl acetate (5 mL) was added to the analyzed aliquots(5 mL) of media containing OP (paraoxon, diazinon,demeton-S , dimethoate, or malathion), and the OP wereextracted at 25 °C. The extracts were analyzed using HPLC(Knauer Smartline Pump 1000, Germany) by reverse-phasechromatography (Diaspher 110-C18 column, 5 μm, 4.0×250 mm, Biochemmack ST) equipped with a UV detectorusing isocratic elution, with an acetonitrile–water mixture(60:40) as the eluent. The retention time of paraoxon (λ =274 nm), diazinon (λ =225 nm), demeton-S (λ =210 nm),dimethoate (λ =210 nm), and malathion (λ =225 nm) was4.5, 26.2, 6.4, 3.4, and 11.4 min, respectively. The eluent flowrate was 1 mL/min, the detector cell temperature was 25 °C,and the injected sample volume was 20 μL.
In this study, hexadecane was used as a model oil hydro-carbon. Hexane (10 mL) was added to the analyzed soil
2648 Appl Microbiol Biotechnol (2014) 98:2647–2656

samples (3 g) containing hexadecane, and the hydrocarbonwas extracted using a temperature-controlled shaker (180 rpm,25 °C) over 3 h. The extracts were analyzed using Kristal-Lyuks 4,000-m gas chromatograph (Meta-Chrom, Russia)equipped with an OV-101 column (Macherey-Nagel,Germany, 0.5 μm, 0.22×500 mm). Analysis conditions: soft-ware, NetChrom v2.1; detector, FID; detector temperature,300 °C; injector temperature, 280 °C; temperature gradient,80–270 °C; heating rate, 12 °C/min; carrier gas, mixture ofnitrogen (1 atm) and hydrogen (3 atm); the retention time ofhexadecane was 17 min; and the injected sample volume was1 μL.
Amino acid sequence alignment
The analysis of OPH–similar enzymes present inRhodococcuscells was carried out using BLASTP 2.2.27 software (http://www.ncbi.nlm.nih.gov/, Accessed 9 April 2012). To this end,the amino acid sequence of OPH (http://www.uniprot.org/uniprot/P0A434.fasta) was aligned with the amino acidsequences of proteins present in Rhodococcus cells (tax ID:1827). Alignment results were analyzed and presented in theform of information on enzymes with the best-fitting align-ments and the least E values (E value being a parameterdescribing the number of random matches in amino acid ornucleotide sequence alignment). E values in the range of 10−52
to 10−33 were considered.Multiple sequence alignment was carried out using the T-
Coffee application (http://tcoffee.crg.cat/). Amino acidsequences of various enzymes were obtained for alignmentat http://www.uniprot.org: SsoPox (sequence numberQ97VT7), OPH (sequence number P0A434), a putative N-acylhomoserine lactonase from Rhodococcus pyridinivorans(sequence number H0JQF8), a putative phosphotriesterasefrom Rhodococcus imtechensis RKJ300 (sequence numberI0WC66), a putative phosphotriesterase from Rhodococcusopacus M213 (sequence number I5ALT5), and a putativephosphotriesterase from Rhodococcus equi 103S (sequencenumber E4WFC4); at http://www.ncbi.nlm.nih.gov: a putativephosphotriesterase from R. opacus PD630 (sequence numberZP_18284315). AhlA amino acid sequence was obtained fromAfriat et al. (2006).
Determination of the lactonase activity of His6-OPH and R.erythropolis AC-1514D and R. ruber AC-1513D cells
Lactonase activity of R. erythropolis AC-1514D and R. ruberAC-1513D cells and His6-OPH was determined by colorime-try (spectrophotometer Agilent UV-853, Germany) using apH-sensitive indicator (Cresol Red, Sigma, USA) (Afriatet al. 2006; Freeman et al. 2009; Larsson et al. 1996).
To determine the lactonase activity of His6-OPH, Eppendorftubes were charged with 983–915 μL of bicine buffer (pH 8.2;0.625 mM) containing 0.1 MNaCl, 5 μL of Cresol Red from astock DMSO solution (12.5 mM), 25 μL of a His6-OPHsolution in bicine buffer (pH 8.2, 1.25 mM), and 12–80 μLof the substrate. In this work, the following N-acylhomoserinelactones were used as substrates: L-homoserine lactone hydro-chloride, N -butyryl-D ,L -homoserine lactone, N -hexyl-L -homoserine lactone, N -(3-oxooctyl)-L-homoserine lactone,N-(3-oxodecyl)-L-homoserine lactone, and N-(3-oxododecyl)-L-homoserine lactone (Sigma, USA). Spectrophotometric mea-surements (OD570) of the reaction mixture were carried outevery 15 min. Catalytic constants were calculated from theinitial rates of the enzymatic reaction corresponding to thevarious initial substrate concentrations using Lineweaver–Burk plots (1/V0−1/S ). The unit of lactonase activity wasdefined as the enzyme concentration that brings about thehydrolysis of 1 μmol of substrate over 1 min at 25 °C (pH 8.2).
Lactonase activity in cells of R. erythropolis AC-1514Dand R. ruber AC-1513D was determined as follows:Eppendorf tubes were charged with 1 mL of cell suspension,cells were precipitated by centrifugation (6,000 rpm, 15 min),and the weight of the precipitated bacterial cells was mea-sured. Cell moisture content (%) was determined by dryingone sample of the precipitated bacterial cells to constantweight. Other samples were used for activity measurement:the cells were resuspended in 945 μL of bicine buffer (pH 8.2,0.625 mM) containing 0.1 M NaCl, to which 5 μL of CreosolRed stock solution in DMSO (12.5 mM) and 50 μL ofaqueous stock solution of an N-acylhomoserine lactone wereadded. The total volume of the reaction mixture was 1 mL.Spectrophotometric measurements (OD570) of the reactionmixture were performed every 15 min after precipitating thecells by centrifugation (5,000 rpm, 10 min). The unit of cellactivity was defined as the enzyme concentration in dried cellsthat brings about the hydrolysis of 1 μmol of substrate perminute at 25 °C (pH 8.2).
OP and hexadecane degradation in soil under the action of R.erythropolis AC-1514D and R. ruber AC-1513D cells
In order to carry out complex bioremediation, bacterial cells ofR. erythropolis AC-1514D and R. ruber AC-1513D at theconcentration of 106 cells/g were introduced into soil (cher-nozem, pH 7.0, humidity 75 %, humus 6.7 %) that wassimultaneously contaminated with paraoxon (88 mg/kg) andhexadecane (6.3 g/kg) (Sigma, USA). The concentration ofthe OP and oil hydrocarbon was measured chromatographi-cally after collecting soil samples and extracting the com-pounds of interest. The data were used to determine the degreeof OP and hexadecane degradation by R. erythropolis AC-1514D and R. ruber AC-1513D cells.
Appl Microbiol Biotechnol (2014) 98:2647–2656 2649
Results
OPH activity of Rhodococcus cells
Investigation of OPH activity in cells of R. erythropolis AC-1514D and R. ruber AC-1513D was carried out in a minimalmedium using various OP (paraoxon, malathion, Diazinon,demeton-S , or dimethoate) as the primary source of carbon.The bacterial cells of interest were found to degrade all OPunder investigation. The data obtained were used to calculatethe initial rates and degrees of OP degradation (Table 1). Thecells of R. erythropolis AC-1514D and R. ruber AC-1513Ddemonstrated the highest efficiency in degrading malathion.
Screening of similar enzymes in Rhodococcus cells
In order to find the enzyme responsible for OP degradation byR. erythropolis AC-1514D and R. ruber AC-1513D cells, wecarried out a comparative analysis of amino acid sequences ofproteins present in Rhodococcus cells (according to the well-known database http://www.ncbi.nlm.nih.gov/) and OPHfrom P. diminuta MG (3.1.8.1). ОРН is the most thoroughlycharacterized bacterial enzyme that efficiently catalyzes thehydrolysis of a wide range of OP (UniProt, http://www.uniprot.org, code P0A434).
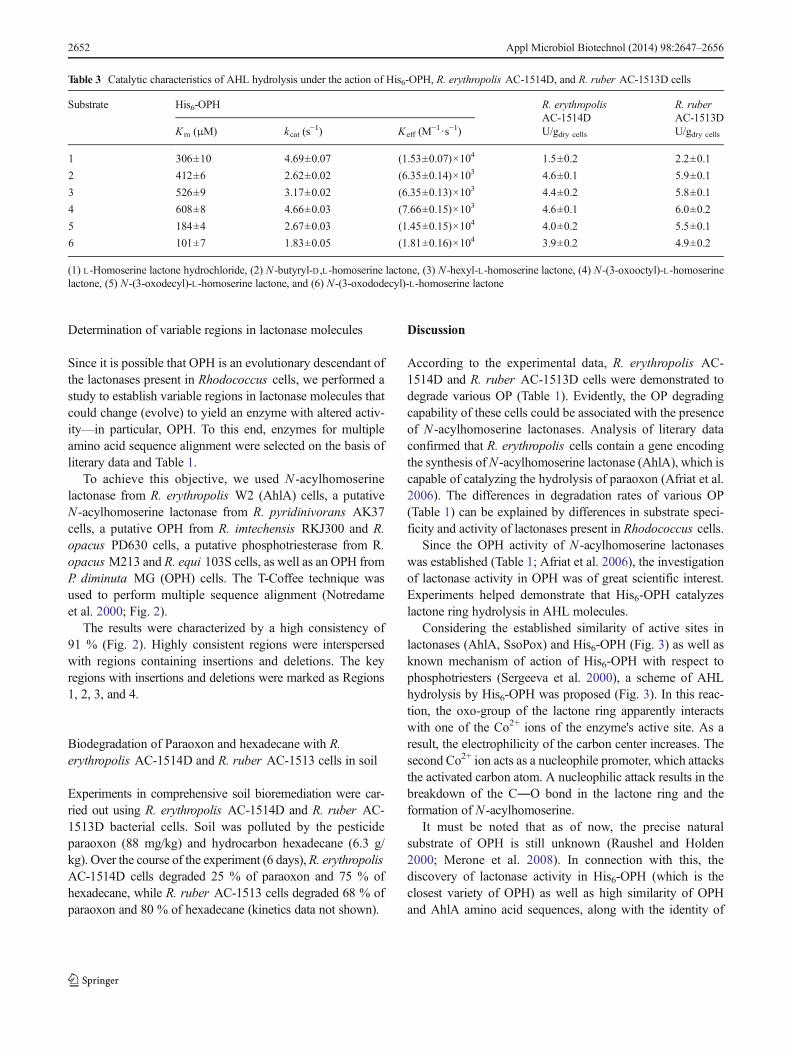
The comparison demonstrated that Rhodococcus cells con-tain proteins that are 46–51 % similar and 28–34 % identicalto OPH (Table 2). “Identity” is the content (%) of identicalamino acid residues at matching positions relative to the totalnumber of positions. “Similarity” is the content (%) of similaramino acid residues at matching positions relative to the totalnumber of positions. The positions of all conservative aminoacid residues making up the active site of OPH (H55, H57,K169, H201, H230, D301) perfectly matched those in amino acidsequences of Rhodococcus proteins.
Further comparison of amino acid sequences demon-strated that most Rhodococcus enzymes are similar to OPHN-acylhomoserine lactonases (EC 3.1.1.81). These enzymescatalyze the hydrolysis of N -acylhomoserine lactones (AHL),
which are “quorum sensing” response auto-inducers in Gram-negative bacterial cells (Hao 2009; Miller and Bassler 2001;Whitehead et al. 2001).
The key activity of N -acylhomoserine lactonase is thecatalysis of lactone ring hydrolysis in various AHL molecules(Afriat et al. 2006; Uroz et al. 2008). At the same time, it wasanN-acylhomoserine lactonase (AhlA) that was demonstratedto slowly hydrolyze paraoxon inR. erythropolis W2 cells witha Keff of 0.5 M−1 s−1 (Afriat et al. 2006).
Additionally, the analysis of literary data showed that OPHcan, in turn, exhibit hydrolytic activity with respect to variouslactones (not of the N -acylhomoserine series, however),cleaving C―O and C―S bonds (Afriat et al. 2006; Roodveldtand Tawfik 2005).
In connection with such substrate “cross-specificity” ofOPH and AhlA, it would be interesting to compare the struc-tural organization of the active sites of these two enzymes.However, since the 3D structure of AhlA is still unresolved,direct structural comparison of the substrate-binding sites of
Table 1 Initial rates and degrees of destruction of various OP (0.2 mM)over 24 h under the action of R. erythropolis AC-1514D and R. ruberAC-1513D cells (18 gcells/L)
OP R. erythropolis AC-1514D R. ruber AC-1513D
V0 (μM/h) Degradationdegree (%)
V0 (μM/h) Degradationdegree (%)
Paraoxon 1.8±0.1 12 1.8±0.1 35
Malathion 8.4±0.1 89 12.0±0.2 95
Demeton-S 3.6±0.1 26 2.4±0.1 27
Diazinon 2.4±0.2 24 7.8±0.2 60
Dimethoate 1.8±0.1 14 2.4±0.1 16
Table 2 Results obtained after amino acid sequence alignment of OPHand proteins present in Rhodococcus cells according to BLASTP 2.2.27(Altschul et al. 1997, 2005) (Accessed 9 April 2012)
Microorganism Putative enzyme Identity(%)
Homology(%)
Sum ofamino acidresidues
R. equi 103S Phosphotriesterase 33 50 317
R. equi ATCC33707
Phosphotriesterase 33 50 317
R. imtechensisRKJ300
Phosphotriesterase 31 51 330
R. opacusPD630
N-acylhomoserinelactonase
31 51 330
R. jostii RHA1 Phosphotriesterase 31 51 330
R. opacusM213
Phosphotriesterase 32 50 329
Rhodococcussp. JVH1
Phosphotriesterase 32 50 330
R. opacusM213
Aryldialkylphosphatase
32 51 325
R.pyridinivora-ns AK37
N-acylhomoserinelactonase
31 49 323
R. imtechensisRKJ300
Aryldialkylphosphatase
29 47 327
R. erythropolisSK121
N-acylhomoserinelactonase
28 47 323
Rhodococcussp. MP50
N-acylhomoserinelactonase
28 47 323
R. erythropolisPR4
N-acylhomoserinelactonase
30 49 323
Rhodococcussp. SQ1
N-acylhomoserinelactonase
29 48 323
Rhodococcussp. 3008
N-acylhomoserinelactonase
29 48 323
R. opacusPD630
Phosphotriesterase 34 46 317
2650 Appl Microbiol Biotechnol (2014) 98:2647–2656

OPH andAhlA is impossible. Due to this, OPHwas comparedto another well-known lactonase, SsoPox, which had beenisolated from the hyperthermophilic archaebacteriumSulfolobus solfataricus (Merone et al. 2005). This choice ofobject for comparative analysis was informed by the fact thatSsoPox exhibits both OPH and AhlA activities (Merone et al.2008).
The analysis of amino acid sequence alignment resultsshowed that all amino acids (H55, H57, K169, H201, H230,D301) participating in binding metal ions at the active site arestrictly conservative and perfectly match across the threeenzymes in question: SsoPox, AhlA, and OPH (Fig. 1). Thedegree of identity and similarity of SsoPox and AhlAwas 40and 62 %, respectively, while the identity and similarity ofSsoPox and OPHwere 31 and 50%, respectively. Pronouncedsimilarity of SsoPox and AhlA allowed us to conclude that theconfiguration of substrate-binding sites in these two enzymesis almost identical.
The key difference between OPH and SsoPox (and, appar-ently, AhlA) lies in the substrate binding site, which comprisesloop 1, loop 7, and loop 8 (Fig. 1). SsoPox and AhlA arecharacterized by an elongated loop 8 and shortened loop 7(Elias et al. 2008); in OPH, the pattern is reversed, with loop 7being 15 amino acid residues longer (Raushel 2002). Due tothis, the substrate binding site in OPH is best suited for thephosphotriester leaving group, whereas the hydrophobicchannel formed by loop 8, along with the shortened loop 7
present in SsoPox and AhlA lactonases, allows to bind AHLmolecules more efficiently than phosphotriester molecules.
Lactonase activity of His6-OPH, R. erythropolis AC-1514Dand R. ruber AC-1513D cells
Lactonase activity was investigated in His6-OPH as well as inbacterial cells of R. erythropolis AC-1514D and R. ruber AC-1513D. We demonstrated His6-OPH to catalyze lactone ringhydrolysis in AHL molecules (Table 3). Analysis of experi-mental data allowed the calculation of the catalytic character-istics of this enzyme with respect to these substrates.
The efficiency constant of His6-OPHwas found to increasewith the length of acyl radical in the AHL molecule.Additionally, His6-OPH was demonstrated to catalyze thehydrolysis of AHLs that contain the 3-oxo group in the acylradical and those that do not contain it.
The cells of R. erythropolis AC-1514D and R. ruber AC-1513D bacteria also degraded AHL molecules (Table 3),which was probably associated with the presence of N -acylhomoserine lactonases in the cells. However, unlikeHis6-OPH, they became less active with acyl radical lengthincreasing from C8 to C10. Additionally, their activity alsodeteriorated upon transition from C4 to C0. Thus, the substratespecificity of lactonase and enzymes in Rhodococcus cellswas demonstrated to differ.
Fig. 1 Amino acid sequence alignment of AhlA lactonase from R.erythropolis W2, SsoPox lactonase from S. solfataricus, and OPH fromP. diminuta MG. Arrows indicate amino acids incorporated into the
active sites of enzymes; horizontal lines outline substrate binding loop.Alignment was performed using T-Coffee (http://tcoffee.crg.cat/)
Appl Microbiol Biotechnol (2014) 98:2647–2656 2651
Determination of variable regions in lactonase molecules
Since it is possible that OPH is an evolutionary descendant ofthe lactonases present in Rhodococcus cells, we performed astudy to establish variable regions in lactonase molecules thatcould change (evolve) to yield an enzyme with altered activ-ity—in particular, OPH. To this end, enzymes for multipleamino acid sequence alignment were selected on the basis ofliterary data and Table 1.
To achieve this objective, we used N -acylhomoserinelactonase from R. erythropolis W2 (AhlA) cells, a putativeN -acylhomoserine lactonase from R. pyridinivorans AK37cells, a putative OPH from R. imtechensis RKJ300 and R.opacus PD630 cells, a putative phosphotriesterase from R.opacus M213 and R. equi 103S cells, as well as an OPH fromP. diminuta MG (OPH) cells. The T-Coffee technique wasused to perform multiple sequence alignment (Notredameet al. 2000; Fig. 2).
The results were characterized by a high consistency of91 % (Fig. 2). Highly consistent regions were interspersedwith regions containing insertions and deletions. The keyregions with insertions and deletions were marked as Regions1, 2, 3, and 4.
Biodegradation of Paraoxon and hexadecane with R.erythropolis AC-1514D and R. ruber AC-1513 cells in soil
Experiments in comprehensive soil bioremediation were car-ried out using R. erythropolis AC-1514D and R. ruber AC-1513D bacterial cells. Soil was polluted by the pesticideparaoxon (88 mg/kg) and hydrocarbon hexadecane (6.3 g/kg). Over the course of the experiment (6 days), R. erythropolisAC-1514D cells degraded 25 % of paraoxon and 75 % ofhexadecane, while R. ruber AC-1513 cells degraded 68 % ofparaoxon and 80 % of hexadecane (kinetics data not shown).
Discussion
According to the experimental data, R. erythropolis AC-1514D and R. ruber AC-1513D cells were demonstrated todegrade various OP (Table 1). Evidently, the OP degradingcapability of these cells could be associated with the presenceof N -acylhomoserine lactonases. Analysis of literary dataconfirmed that R. erythropolis cells contain a gene encodingthe synthesis ofN -acylhomoserine lactonase (AhlA), which iscapable of catalyzing the hydrolysis of paraoxon (Afriat et al.2006). The differences in degradation rates of various OP(Table 1) can be explained by differences in substrate speci-ficity and activity of lactonases present in Rhodococcus cells.
Since the OPH activity of N -acylhomoserine lactonaseswas established (Table 1; Afriat et al. 2006), the investigationof lactonase activity in OPH was of great scientific interest.Experiments helped demonstrate that His6-OPH catalyzeslactone ring hydrolysis in AHL molecules.
Considering the established similarity of active sites inlactonases (AhlA, SsoPox) and His6-OPH (Fig. 3) as well asknown mechanism of action of His6-OPH with respect tophosphotriesters (Sergeeva et al. 2000), a scheme of AHLhydrolysis by His6-OPH was proposed (Fig. 3). In this reac-tion, the oxo-group of the lactone ring apparently interactswith one of the Co2+ ions of the enzyme's active site. As aresult, the electrophilicity of the carbon center increases. Thesecond Co2+ ion acts as a nucleophile promoter, which attacksthe activated carbon atom. A nucleophilic attack results in thebreakdown of the C―O bond in the lactone ring and theformation of N -acylhomoserine.
It must be noted that as of now, the precise naturalsubstrate of OPH is still unknown (Raushel and Holden2000; Merone et al. 2008). In connection with this, thediscovery of lactonase activity in His6-OPH (which is theclosest variety of OPH) as well as high similarity of OPHand AhlA amino acid sequences, along with the identity of
Table 3 Catalytic characteristics of AHL hydrolysis under the action of His6-OPH, R. erythropolis AC-1514D, and R. ruber AC-1513D cells
Substrate His6-OPH R. erythropolisAC-1514D
R. ruberAC-1513D
Km (μМ) kcat (s−1) Keff (М
−1·s−1) U/gdry cells U/gdry cells
1 306±10 4.69±0.07 (1.53±0.07)×104 1.5±0.2 2.2±0.1
2 412±6 2.62±0.02 (6.35±0.14)×103 4.6±0.1 5.9±0.1
3 526±9 3.17±0.02 (6.35±0.13)×103 4.4±0.2 5.8±0.1
4 608±8 4.66±0.03 (7.66±0.15)×103 4.6±0.1 6.0±0.2
5 184±4 2.67±0.03 (1.45±0.15)×104 4.0±0.2 5.5±0.1
6 101±7 1.83±0.05 (1.81±0.16)×104 3.9±0.2 4.9±0.2
(1) L-Homoserine lactone hydrochloride, (2) N-butyryl-D,L-homoserine lactone, (3) N-hexyl-L-homoserine lactone, (4) N-(3-oxooctyl)-L-homoserinelactone, (5) N-(3-oxodecyl)-L-homoserine lactone, and (6) N-(3-oxododecyl)-L-homoserine lactone
2652 Appl Microbiol Biotechnol (2014) 98:2647–2656

their active sites, allowed us to hypothesize that an evolu-tionary link may exist between these enzymes.
In investigating such a link between OPH and AhlA, wedecided to follow three criteria proposed in the literature for
establishing evolutionary relationships between enzymes(Gerlt and Raushel 2003): the overall structure of the activesite, high similarity of amino acid sequences (for divergentevolution), and substrate specificity, with an implication that
Fig. 2 Amino acid sequence alignment of enzymes AhlA from R.erythropolis W2 (AhlA), putative N-acylhomoserine lactonase from R.pyridinivorans AK37 (AK37), putative phosphotriesterase from R.imtechensis RKJ300 (RKJ_1), putative phosphotriesterase from R.
opacus M213 (M213_1), putative phosphotriesterase enzyme from R.equi 103S (103S), OPH from P. diminuta MG (OPH), and putativephosphotriesterase from R. opacus PD630 (PD_2). Alignment wasperformed using T-Coffee technique (http://tcoffee.crg.cat/)
Appl Microbiol Biotechnol (2014) 98:2647–2656 2653

the evolutionary ancestor and its descendant enzyme cancatalyze identical chemical reactions but exhibit different sub-strate specificity.
Experimental data and their analysis demonstrated thatOPH and AhlA meet all the criteria for establishing an evolu-tionary “kinship”. According to the results of amino acidsequence alignment, the position of amino acid residues inthe active center of OPH perfectly matched that of amino acidresidues in AhlA (Fig. 1). Meanwhile, the degrees of similar-ity and identity of the enzymes were 47 and 28 %, respective-ly. OPH and AhlA catalyzed identical chemical reactions, butin the case of OPH, greater enzyme efficiency constants wereobtained for phosphotriesters (e.g., paraoxon), whereas in thecase of AhlA, greater values were observed with AHL.
Comparative analysis (Fig. 2) of Rhodococcus enzymessimilar to OPH (Table 2) and AhlA lactonase allowed us toformulate a hypothesis about the localization of evolutionaryvariable regions in the enzyme molecules of Rhodococcuscells. Changes in these specific regions could lead to OPHdevelopment from lactonases. This hypothesis agreed withreported data of experiments in directed in vitro evolution,which demonstrated that enzymes can rapidly develop a“new” activity owing to a limited number of mutations underconditions conductive to evolution (Aharoni et al. 2005). Inthis connection, lactonase activity of OPH could be a residualtrait of an ancestor enzyme—lactonase. This hypothesis waspreviously proposed in the literature (Afriat et al. 2006), butexperimental facts confirming that OPH exhibits lactonaseactivity were lacking. In connection with this, the results ofthis work are of major importance for basic science.
Comparison of results of two amino acid sequence align-ments (Fig. 2, Table 2) revealed that region 1 corresponds
to loop 1, region 3 corresponds to loop 7, and region 4 corre-sponds to loop 8. As noted above, loops 1, 7, and 8 areresponsible for organizing the substrate binding site in OPH,SsoPox, and apparently, AhlA (Elias et al. 2008; Raushel 2002).
Thus, the greatest evolutionary change occurred in the loopresponsible for substrate binding. These changesmay give riseto new enzymes that are different from the original enzyme interms of substrate specificity. Changes in amino acid compo-sition that do not involve the enzyme's active site may have acompensatory nature or represent coordinated substitutionsaimed at maintaining the integrity of the enzyme's structureand catalytic properties after evolutionary substitutions ofspecific amino acid residues (Pazos et al. 1997).
The newly established ability of oil-oxidizing R. erythropolisAC-1514D and R. ruber AC-1513D cells to degrade a widerange of OP is an important practical advantage of these micro-organisms since OP are known to be used as antimicrobialadditives to lubricant oils and petroleum (Singh 2009); addition-ally, some pesticide products are manufactured with addedpetroleum products (Chariot TM Insecticides 2011), and petro-leum products per se can be used as pesticides (Koul et al.2008). In this connection, Rhodococcus bacterial cells appearto be promising biocatalysts for comprehensive simultaneousdegradation of OP and the accompanying hydrocarbons invarious media and ecosystems.
In conclusion, it must be emphasized that not only the OPHactivity of Rhodococcus cells was demonstrated in this workwith respect to five different OP, but also the lactonase activityof His6-OPH was established for the first time. These resultsoffer new opportunities in using the enzyme to suppress“quorum sensing” response in Gram-negative bacterial cells.The lactonase activity of His6-OPH can be used to develop
Fig. 3 Hypothesized schema ofthe mechanism of His6-OPHaction in the hydrolysis of AHL
2654 Appl Microbiol Biotechnol (2014) 98:2647–2656

new strategies of combating resistant bacterial associates thatinduce inflammatory processes, biocorrosion, etc. Theoreticalconsiderations helped demonstrate that the emergence of OPHfrom lactonases through changes in amino acid composition incertain evolutionarily variable regions of the proteins isprobable.
Acknowledgments This work was supported by the Russian Founda-tion of Basic Research (application no. 13-04-01086, project typeA_13_04-01086).Wewish to thank I. Lyagin for the help with alignments.
References
Afriat L, Roodveldt C, Manco G, Tawfik DS (2006) The latent promis-cuity of newly identified microbial lactonases is linked to a recentlydiverged phosphotriesterase. Biochemistry 45:13677–13686. doi:10.1021/bi061268r
Aharoni A, Gaidukov L, Khersonsky O, McQGould S, Roodveldt C,Tawfik DS (2005) The “evolvability” of promiscuous protein func-tion. Nat Genet 37:73–76. doi:10.1038/ng1482
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W,Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new gener-ation of protein database search programs. Nucleic Acids Res 25:3389–3402. doi:10.1093/nar/25.17.3389
Altschul SF, Wootton JC, Gertz EM, Agarwala R, Morgulis A, SchäfferAA, Yu YK (2005) Protein database searches using compositionallyadjusted substitution matrices. FEBS J 272:5101–5109. doi:10.1111/j.1742-4658.2005.04945.x
Chariot TM Insecticides (2011) Material safety data sheet, 10 p. FMCCorporation Agricultural Products Group, Philadelphia
Eddleston M, Buckley NA, Eyer P, Dawson AH (2008) Management ofacute organophosphorus pesticide poisoning. Lancet 371:597–607.doi:10.1016/S0140-6736(07)61202-1
Efremenko EN, Sergeeva VS (2001) Organophosphate hydrolase - anenzyme catalyzing degradation of phosphorus-containing toxinsand pesticides. Russ Chem Bull 50:1826–1832. doi:10.1023/A:1014377912147
Efremenko EN, Varfolomeev SD (2004) Enzymes destructingphosphororganic neurotoxins. Uspekhi Biologicheskoi Khimmii(Ru) 44:307–340
Efremenko EN, Votchitseva JA, Aliev TK, Varfolomeev SD (2005a)Recombinant plasmid DNA pTES-His-OPH and producer ofoligohistidine-containing organophosphate hydrolase. RU patent2255975, 19 Dec 2003
Efremenko EN, Azizov RE, Raeva AA, Abbasov VM, Varfolomeyev SD(2005b) An approach to the rapid control of oil spill bioremediationby bioluminescent method of intracellular ATP determination. IntBiodeter Biodegr 56:94–100. doi:10.1016/j.ibiod.2005.06.004
Efremenko E, Votchitseva Y, Plieva F, Galaev I, Mattiasson B (2006)Purification of His6-organophosphate hydrolase using monolithicsupermacroporous polyacrylamide cryogels developed forimmobilized metal affinity chromatography. Appl Microbiol Biot70:558–563. doi:10.1007/s00253-005-0103-x
Efremenko E, Lyagin I, Votchitseva Y, Sirotkina M, Varfolomeyev S(2007) Polyhistidine-containing organophosphorus hydrolase withoutstanding properties. Biocatal Biotransfor 25:103–108. doi:10.1080/10242420601141796
Elias M, Dupuy J, Merone L, Mandrich L, Porzio E, Moniot S,Rochu D, Lecomte C, Rossi M, Masson P, Manco G, Chabriere E(2008) Structural basis for natural lactonase and promiscuous
phosphotriesterase activities. J Mol Biol 379:1017–1028. doi:10.1016/j.jmb.2008.04.022
Fleseriu A, Oroian I, Viman O, Brasovean I (2010) Pest risk analysis - away to counter attack losses caused by plant pathogens insolanaceae’s cultures. ProEnvironment/ProMediu 3:289–291
Freeman L, Buisson M, Tarbouriech N, Van der Heyden A, Labbe P,Burmeister WP (2009) The flexible motif V of Epstein-Barr virusdeoxyuridine 5′-triphosphate pyrophosphatase is essential for catal-ysis. J Biol Chem 284:25280–25289. doi:10.1074/jbc.M109.019315
Gerlt JA, Raushel FM (2003) Evolution of function in (b/a)8-barrelenzymes. Curr Opin Chem Biol 7:252–264. doi:10.1016/S1367-5931(03)00019-X
Grube A, Donaldson D, Kiely T, Wu L (2011) Pesticides industry salesand usage 2006 and 2007. US EPA. http://www.epa.gov/opp00001/pestsales/07pestsales/market_estimates2007.pdf. AccessedFebruary 2011
Hao Y (2009) Two of the mechanisms used by bacteria to modify theenvironment: quorum sensing and acc deaminase. The University ofWaterloo, Dissertation
Hayden K, Norton M, Darcey D, Ostbye T, Zandi P, Breitner J, Welsh-Bohmer K (2010) Occupational exposure to pesticides increases therisk of incident AD: the cache county study. Neurology 74:1524–1530. doi:10.1212/WNL.0b013e3181dd4423
Hopkins EH, Hippe DJ, Frick EA, Buell GR (2000) Organophosphoruspesticide occurrence and distribution in surface and ground water ofthe United States, 1992–97. US Geological Survey Open-FileReport 00–187. http://pubs.usgs.gov/of/2000/ofr00-187/pdf/ofr00-187.pdf. Accessed June 2000
Koul O, Walia S, Dhaliwal GS (2008) Essential oils as green pesticides:potential and constraints. Biopest Int 4:63–84
Larsson G, Nyman PO, Kvassman JO (1996) Kinetic characterization ofdUTPase from Escherichia coli . J Biol Chem 271:24010–24016.doi:10.1074/jbc.271.39.24010
Merone L, Mandrich L, Rossi M, Manco G (2005) A thermostablephosphotriesterase from the archaeon Sulfolobus solfataricus : clon-ing, overexpression and properties. Extremophiles 9:297–305. doi:10.1007/s00792-005-0445-4
Merone L, Mandrich L, Rossi M, Manco G (2008) Enzymes withphosphotriesterase and lactonase activities in Archaea. Curr ChemBiol 2:237–248. doi:10.2174/187231308785739756
Miller MB, Bassler BL (2001) Quorum sensing in bacteria. Annu RevMicrobiol 55:165–199. doi:10.1146/annurev.micro.55.1.165
Murygina V, Arinbasarov M, Kalyuzhnyi S (2000) Bioremediation of oilpolluted aquatic and soils with novel preparation “Rhoder”.Biodegradation 11:385–389. doi:10.1023/A:1011680703911
Murygina VP, Markarova MY, Kalyuzhnyi SV (2005) Application ofbiopreparation "Roder" for remediation of oil polluted polar marshywetlands in Komi Republic. Environ Int 31:163–166. doi:10.1016/j.envint.2004.09.010
Notredame C, Higgins DG, Heringa J (2000) T-Coffee: a novel methodfor fast and accurate multiple sequence alignment. J Mol Biol 302:205–217. doi:10.1006/jmbi.2000.4042
Pazos F, Helmer-Citterich M, Ausiello G, Valencia A (1997) Correlatedmutations contain information about protein-protein interaction. JMol Biol 271:511–523. doi:10.1006/jmbi.1997.1198
Raushel FM, HoldenHM (2000) Phosphotriesterase: an enzyme in searchof its natural substrate. Adv Enzymol Relat Areas Mol Biol 74:51–93. doi:10.1002/9780470123201.ch2
Raushel FM (2002) Bacterial detoxification of organophosphate nerveagents. Curr Opin Microbiol 5:288–295. doi:10.1016/S1369-5274(02)00314-4
Reigart JR, Roberts JR (1999) Recognition and management of pesticidepoisonings. US EPA. http://www.epa.gov/oppfead1/safety/healthcare/handbook/Chap04.pdf. Accessed Mar 1999
Roodveldt C, Tawfik DS (2005) Shared promiscuous activities andevolutionary features in various members of the amidohydrolase
Appl Microbiol Biotechnol (2014) 98:2647–2656 2655

superfamily. Biochemistry 44:12728–12736. doi:10.1021/bi051021e
Sergeeva VS, Efremenko EN, Kazankov GM, Varfolomeyev SD (2000)Double effect of organic amines (activation and inhibition) on thephosphotriesterase. JMol Catal B-Enzym 10:571–576. doi:10.1016/S1381-1177(00)00102-8
Singh BK (2009) Organophosphorus-degrading bacteria: ecology andindustrial applications. Nat Rev Microbiol 7:156–164. doi:10.1038/nrmicro2050
Sirotkina M, Lyagin I, Efremenko E (2012) Hydrolysis of organophos-phorus pesticides in soil: new opportunities with ecocompatibleimmobilized His6-OPH. Int Biodeter Biodegr 68:18–23. doi:10.1016/j.ibiod.2011.12.004
Uroz S, Oger PM, Chapelle E, Adeline MT, Faure D, Dessaux Y(2008) A Rhodococcus qsdA-encoded enzyme defines a nov-el class of large-spectrum quorum-quenching lactonases.Appl Environ Microbiol 74:1357–1366. doi:10.1128/AEM.02014-07
Votchitseva YA, Efremenko EN, Aliev TK, Varfolomeyev SD(2006) Properties of hexahistidine-tagged organophosphatehydrolase. Biochem (Mosc) 71:167–72. doi:10.1134/S0006297906020088
Whitehead NA, Barnard AML, Slater H, Simpson NJL, Salmond GPC(2001) Quorum-sensing in gram-negative bacteria. FEMSMicrobiol Rev 25:365–404. doi:10.1111/j.1574-6976.2001.tb00583.x
2656 Appl Microbiol Biotechnol (2014) 98:2647–2656





























