Embed Size (px)
Citation preview

Review
Rho-dependent termination and ATPases in transcript termination
John P. Richardson *
Department of Chemistry, Indiana University, 800 East Kirkwood Avenue, Bloomington, IN 47405, USA
Received 21 June 2002; accepted 21 June 2002
Abstract
Transcription factor Rho is a ring-shaped, homohexameric protein that causes transcript termination through actions on nascent RNAs that
are coupled to ATP hydrolysis. The Rho polypeptide has a distinct RNA-binding domain (RNA-BD) of known structure as well as an ATP-
binding domain (ATP-BD) for which a structure has been proposed based on homology modeling. A model is proposed in which Rho first
makes an interaction with a nascent RNA on a C-rich, primarily single-stranded rut region of the transcript as that region emerges from the
exit site of RNA polymerase. A subsequent step involves a temporary release of one subunit of the hexamer to allow the 3Vsegment of the
nascent transcript to enter the central channel of the Rho ring. Actions of the Rho structure in the channel on the 3Vsegment that are coupled
to ATP hydrolysis pull the RNA from its contacts with the template and RNA polymerase, thus causing termination of its synthesis.
D 2002 Elsevier Science B.V. All rights reserved.
Keywords: Rho factor; Transcript termination; Hexameric helicase; RNA-binding protein; NusG; Factor 2
1. Introduction
Rho factor is a protein used in bacteria for termination of
transcripts at certain sites. It serves an active role in
termination, employing the energy from ATP hydrolysis to
mediate the dissociation of a nascent transcript. In Escher-
ichia coli, Rho-dependent terminators are used at the ends of
genes and operons and at regulatory sites (attenuators)
preceding genes [1]. The RNA polymerase in bacteria can
also terminate transcription at other sites without the assis-
tance of Rho factor [1]. These intrinsic terminators are also
used at ends of genes and at attenuators. Both types of
terminators are used in E. coli and other bacteria for the
orderly expression of the genetic information.
RNA polymerases are well adapted to making complete
transcripts of genes. Once committed to the transcription of
a gene sequence, they form stable transcriptional elongation
complexes (TECs) with the DNA and nascent RNA. The
stability of the TEC is consistently high for most sequences,
but decreases greatly upon transcription of the DNA of an
intrinsic terminator [2–5]. A combination of features near
the 3V end of the nascent transcript and the interactions of
that part of the transcript with other components of the TEC
cause RNA polymerase to release its hold on the RNA, thus
allowing spontaneous release of the transcript. The sequen-
ces of an intrinsic terminator conform to a consensus, which
contains a 20 base pair stretch of DNA that encodes an RNA
that can form a stable stem-loop structure [6]. This sequence
is followed by seven to eight A residues in the template
strand. Release of the transcript occurs when the final seven
to eight U residues of the transcript are paired with the seven
to eight A residues of the template. A hybrid helix of rU
residues with dA residues is much less stable than hybrid
helices with other sequences [7]. This property, combined
with the formation of a stem-loop structure in the RNA in
the exit channel of RNA polymerase, is sufficient for the
release of the transcript to dissociate from the complex
before the next nucleotide is added.
Rho-dependent terminators are very different and are not
definable by a simple consensus sequence. Rho serves the
function of mediating release of the transcript at a DNA
sequence at which the transcription complex is too stable for
spontaneous release. Thus, a Rho-dependent terminator can
occur at sites where the sequence of an intrinsic terminator
would be incompatible with other functions of the sequence.
To mediate release, Rho binds to the RNA and uses
mechanical actions on the RNA that are coupled to ATP
hydrolysis to wrest the transcript from RNA polymerase and
the DNA template [8,9]. Rho is thus an ATP-driven machine.
The purpose of this review is to present the recent
advances in the structure and mechanism of Rho that have
0167-4781/02/$ - see front matter D 2002 Elsevier Science B.V. All rights reserved.
PII: S0167 -4781 (02 )00456 -6
* Tel.: +1-812-855-1520; fax: +1-812-855-8300.
E-mail address: [email protected] (J.P. Richardson).
www.bba-direct.com
Biochimica et Biophysica Acta 1577 (2002) 251–260

provided insights into how this machine terminates tran-
scription. Also, some aspects of the biology of the function
of Rho in bacteria will be considered, as will the properties
of some ATP hydrolysis-driven release factors for eukary-
otic RNA polymerases.
2. Structure
Rho functions as an assemblage of six identical, wedge-
shaped subunits arranged around a symmetry axis like the
segments of an orange [10]. The attempts that have been
made to get diffraction grade crystals of Rho protein have
not been successful. However, the morphology of the
protein has been determined by a three-dimensional recon-
struction from the electron micrographic images of nega-
tively stained protein adsorbed onto carbon grids [10]. The
images are primarily ring-shaped with six-fold symmetry.
Although some earlier studies suggested that Rho might
have D3 symmetry, these electron micrographic images
were more consistent with a six-fold or pseudo six-fold
rotational (C6) symmetry. The diameter of the rings at their
widest point was about 120 A. The height along the six-fold
axis was about 90 A.
In the EM analysis, most rings had six subunits but about
10% had only five subunits arranged in a gapped ring
structure. From sedimentation studies, it is known that Rho
readily dissociates into smaller assemblies at concentrations
below 1 AM [11,12], which is about the concentration in the
cell and just under the concentration used for the EM studies.
Thus, the gapped structure could be very relevant for Rho
function. In the current model for the mechanism of Rho
action, a segment of the RNA needs to be captured in the
center of a ring under conditions in which the protein would
not be able to thread onto a closed ring structure [13,14].
This could readily happen by capturing the RNA in the
center of the ring of a temporarily gapped structure.
A single subunit of E. coli Rho has 419 amino acid
residues [15]. An N-terminal segment consisting of the first
129 residues has been isolated and crystallized [16,17]. This
segment binds RNA by itself and is called the RNA-binding
domain (RNA-BD). The remaining part of Rho has a site for
binding ATP [18,19]. It has been termed the ATP binding
domain (ATP-BD), although it has not been isolated.
Molecular level structures of the free RNA-BD both in
crystals [17] and in solution [20] reveal that its first 50
residues are organized into an a-helical bundle while the
remaining part is a h-barrel having an OB fold. The OB fold
is a common structural motif in many proteins that bind
single-stranded polynucleotides. Other proteins with OB
folds are ribosomal protein S1, bacterial cold-shock proteins
and the single-stranded DNA-binding proteins. One of the
hallmarks of Rho function is a requirement for single-
stranded segments in the RNA. These single-stranded seg-
ments bind across the OB-fold segments of multiple sub-
units in the Rho hexamer.
Fig. 1. Views of two opposing subunits of the Rho hexamer. Two modeled subunits are arranged with rotational symmetry about the vertical axis with the plane
of the paper. The N-terminal 123 residues are represented by a black ribbon, while the C-terminal region (residues 126–419) is represented by a gray ribbon.
This model is based on the one described in Burgess and Richardson [23].
J.P. Richardson / Biochimica et Biophysica Acta 1577 (2002) 251–260252

To suggest a mechanism of Rho, it has been necessary to
use models for the structures of the ATP-BD and for the
organization of the protein into a hexamer. These models are
based on the known structure of the F1-ATPase, a protein
that is similar to Rho. When compared with all other known
protein sequences in the data base, the sequences that are
most closely similar to those of the Rho protein sequences
are those of the a and h subunits of the F1-ATPase [21].
These subunits are well conserved in a great variety of
organisms and are closely similar to each other. This
similarity is strongest for the ATP-BD of Rho and the
corresponding parts of those subunits. The identity in those
regions is about 21% and the similarity is about 43%. Rho
and the part of the F1-ATPase that contains the three a and
three h subunits are also morphologically very similar [10],
suggesting that the proteins have the same quaternary
organization. For that reason, the tertiary structures of the
a and h subunits can serve as models for the tertiary
structure of Rho, and the conglomerate of the three a and
three h subunits serve as a model for the quaternary
structure [22]. Molecular level models were made by
threading the aligned sequences of the ATP-BD of Rho
onto the coordinates of the corresponding parts of the a and
h subunits of bovine mitochondrial F1-ATPase followed by
an energy minimization to remove steric clashes [23]. The
known coordinates of Rho’s RNA-BD were then docked
onto the model for the six ATP-BDs, replacing the corre-
sponding parts of the a and h subunits at one pole of the
hexamer. The orientation for the RNA-BDs was suggested
from the three-dimensional reconstruction of the Rho struc-
ture from the EM images, to which the known structure of
the RNA-BD could be docked with a unique fit [10]. This
orientation has the clefts for binding RNA facing out toward
the periphery of the globular structure with the normals to
the face of the clefts at about a 45j angle to the six-fold axis.Fig. 1 is a diagram of the ribbon structures of two opposing
subunits.
3. Rho–RNA interactions
Rho factor is known to bind with relatively high affinity
to RNA [24,25] and to the transcription factors NusG
[26,27] and NusA [28]. The binding to RNA is essential
for Rho function and is responsible for the specificity of
Rho action. The interaction of Rho with RNA activates NTP
hydrolysis by Rho [29], a step that is essential for mediating
release of RNA transcripts from the transcription complex.
However, Rho can bind to RNA in the absence of an NTP
[24,25], and this interaction has been used to analyze the
specificity of the binding. Rho has a strong preference for
single-stranded RNA rich in C residues. Thus, it binds very
tightly to poly(C). Poly(C) becomes saturated with Rho at a
ratio of one Rho hexamer per 78 nucleotides or about 13
nucleotides per subunit [24,25]. When complexes of Rho
with poly(C) are treated with ribonuclease, an RNA segment
of about 78 residues is protected from digestion [24,30].
This result suggests that poly(C) is bound in a cleft that
extends continuously across the six subunits.
The interaction of RNA with individual subunits in the
Rho hexamer has been probed by measuring the binding of
oligo(C) molecules that are smaller than 13 nucleotides in
length. For rC10, Rho has three strong and three weak RNA-
binding sites that differ by about 10-fold in their affinities
for oligonucleotides [31,32]. This result indicates that not all
the subunits have identical conformations. The isolated
RNA-BD also binds oligo(C) [16]. However, it does so
with an affinity (the Kd for rC6 is about 2 AM) that is similar
to that for the weak binding sites in the Rho hexamer. Thus,
the organization of Rho into a hexameric structure may
entail a conformational change that puts half of the subunits
into a higher affinity form.
Both isolated RNA-BD and hexameric Rho bind to other
RNA oligonucleotides and RNA homopolymers but with
generally much weaker affinities. The affinity for the next
best RNA homo-oligonucleotide, oligo(U), is more than 10-
fold lower [32]. Both will also bind to single-stranded DNA
as well [16,24]. The isolated RNA-BD binds to oligo(dC)
with an affinity that is about five-fold lower than for the
corresponding oligo(rC) [33]. Thus, the site in the RNA-BD
has a strong preference for C residues and a slight prefer-
ence for RNA over DNA. A crystal structure of a complex
of rC9 with the isolated RNA-BD has revealed the reason
for the strong preference for C residues and for pyrimidines
in general [34]. In the complex, one cytosine base of one
residue makes three H-bond interactions with the side
chains of two key residues in the RNA-BD, two H-bonds
in a bi-dentate interaction with Arg-66 and the third with an
oxygen on the Asp-78 side chain. The base in the next
residue to the 5V side fits into a pocket that is only large
enough for a pyrimidine. Thus, at least two of the inter-
actions in one of the subunits are strongly cytosine or
pyrimidine specific. Evidence from NMR spectra of com-
plexes of oligonucleotides with the isolated RNA-BD indi-
cate that there is another C-specific interaction at residues
88 and 89, which extend from the short a-helix in the OB-
fold of the RNA-BD (T.K. Hitchins and G.S. Rule, personal
communication).
4. Attachment of Rho to a transcript to be terminated
For Rho to terminate the synthesis of a transcript, it first
must become attached in a productive manner. A segment of
the nascent transcript must be available for Rho to bind with
a sufficiently high affinity. In bacteria, ribosomes become
attached to a nascent mRNA as soon the translation initia-
tion site is available and this is usually before Rho can
attach to the RNA. The presence of a ribosome translating
the mRNA at the same rate with which the RNA is being
synthesized effectively prevents Rho from access to the
nascent RNA until the end of the gene or the operon [35].
J.P. Richardson / Biochimica et Biophysica Acta 1577 (2002) 251–260 253

Once the coupling is interrupted, the nascent RNA is then
available, and, if the sequence is appropriate, Rho will
become attached and initiate the termination process. An
appropriate sequence consists of a segment of 40 or more
non-base-paired nucleotides containing a relatively high
proportion of C residues [36,37]. Such a sequence is called
a rut site, named after the genetic designation of the DNA
sequence element called the Rho Utilization site [38]. This
site on the RNA has also been referred to as the rut site for
the RNA attachment role it actually performs [39].
One example of a Rho-dependent terminator is tR1, the
transcript terminator between the cro and cII genes of
bacteriophage E [40]. Its rut sequence starts just down-
stream from the open reading frame for the Cro protein and
just upstream of where transcripts are terminated by Rho
action [38,41]. Rho binds to the E cro RNA containing its
rut sequence with Kdf 30 pM [42]. In contrast the affinity
of Rho to E cro RNAvariants lacking the rut site segment or
to the RNA containing the rut sequence but with it base-
paired to complementary DNA oligonucleotides is about
100-fold weaker [43]. These results correlate with the
findings that deletion of the rut segment of DNA or replace-
ment of the segment of the DNA with one that forms a
highly base-paired structure lead to a loss of termination
function both in vitro and in vivo [38,44]. In addition,
blocking the access to the rut site in vitro by the addition
of DNA oligonucleotides that are complementary specifi-
cally to the rut sequence segments on the nascent RNA also
strongly inhibits Rho-dependent termination [41].
5. Termination sites
A Rho-dependent terminator consists of two parts, the rut
region and the region where transcripts are terminated,
called the tsp region [45]. This second part can extend for
nearly 100 base pairs, thus giving an extent of nearly 150 bp
overall for a Rho-dependent terminator. The broad extent of
the tsp region is because the transcripts are terminated at
several positions often in clusters of five to eight successive
stop points separated by regions of 20–30 bp where few
transcripts are terminated [40,46,47]. This dispersion of the
stop points differs from the relatively precise position of the
stop points for an intrinsic terminator, in which most tran-
scripts end at a single base pair with a small fraction at an
adjacent base pair [2].
The positions of the stop points in a Rho-dependent
terminator are determined by the sequence features that
control the rate of nucleotide addition by RNA polymerase
at each residue. The rate for addition of nucleotides by RNA
polymerases can vary considerably from one position to the
other, giving rise to characteristic pause points that are
distributed very much like the stop points at a Rho-depend-
ent terminator [48]. These pause points are dictated by
several features of the DNA sequence in and around the
base pairs at which the pausing occurs [49,50]. Comparing
the distribution of natural pause points in a tsp region of a
Rho-dependent terminator during transcription of the region
in the absence of Rho with the distribution of stop points of
the RNA molecules synthesized in the presence of Rho
shows a very good correlation [51,52]. Thus, termination
occurs at pause sites that are downstream from a rut region.
Pause sites occur naturally and with similar frequencies
in most DNA sequences. As a consequence, when a new
DNA sequence not from a natural termination site is
positioned downstream from a rut region, Rho will cause
transcripts to be terminated at the natural pause sites in the
inserted DNA [45]. This characteristic implies that the most
unique determinant of a Rho-dependent terminator is the rut
region. Also, because Rho does not bind stably to an RNA
until most of the rut region is available [53], it acts very
efficiently at the first pause sites encountered by RNA
polymerase just after the rut region is available. This means
that the distance between where Rho loads on the RNA and
where the transcript is terminated is relatively short, in a
range from 20 to 40 nucleotides. Models showing Rho
binding to a far upstream site and having it migrate along
scores of RNA nucleotides until it encounters a paused RNA
polymerase are probably not good representations.
Mutant forms of Rho that act more slowly than normal
Rho are very poor at terminating transcription with a normal
RNA polymerase under normal transcription conditions
[54]. However, the function of these mutants can be recov-
ered when they are used with a mutant RNA polymerase
that elongates RNA chains more slowly or with a normal
RNA polymerase under conditions of lower NTP concen-
trations [55]. These observations reveal a kinetic coupling
between Rho action and transcript elongation. A critical
aspect of this kinetic coupling is the timing of how long it
takes Rho to start tracking along a transcript once it has
bound tightly to the transcript. Even if the distance to the
pause site is not far, a delay in acting on the RNAwill cause
Rho to lose its chance to dissociate the transcript before
RNA polymerase has moved beyond the pause site.
6. ATP binding and hydrolysis
Rho–RNA complexes catalyze the hydrolysis of all four
nucleoside triphosphates [56]. However, its preferred sub-
strate is ATP. Rho will bind to ATP in the absence of RNA.
Each hexamer has three sites that bind ATP with Kdf 0.5
AM [57]. Although there was some evidence that Rho could
bind three more ATP molecules with a much weaker affinity
[58,59], the existence of these sites has been difficult to
verify [60]. Pre-steady state kinetic measurements using a
rapid mix and quench device showed that addition of RNA
(poly(C)) to the Rho–ATP3 complex led to hydrolysis of 1
ATP in less than 5 ms and at least 1.5 more ATP by 200 ms
[60]. These results and others measuring the rates of
hydrolysis of labeled ATP added to a presaturated Rho–
ATP3 complex have been simulated by a model with the
J.P. Richardson / Biochimica et Biophysica Acta 1577 (2002) 251–260254

following properties [60]. Rho has three catalytic sites in
which ATP can bind. Addition of RNA to the Rho–ATP3complex causes rapid hydrolysis (rate constant of 300 s� 1)
of one ATP. This reaction is followed by a rate-limiting step
(either product release or protein conformational change) at
27 s � 1. Additional bound ATP molecules sequentially
undergo similar rapid chemical hydrolysis followed by a
similar slow step. This model is consistent with the steady-
state hydrolysis rate (turnover number) of 30 s � 1 per
hexamer. The rapid quench results were not consistent with
an alternative model that has been proposed in which ATP
bound at slow exchanging sites (i.e. the high affinity sites) is
hydrolyzed at a relatively slow rate of 1.8 s� 1 [14]. That
alternative model posits that the high-affinity sites are not
catalytic and that turnover ATP hydrolysis is carried out in
the putative low-affinity sites. Attempts to reproduce the
experiments that were consistent with this alternative model
in another laboratory were not successful [60], suggesting
that those results may have been a characteristic of a
particular preparation of Rho protein.
7. The role of NTP hydrolysis
As noted above, Rho can bind to RNA in the absence of
an NTP. However, in the cell and in transcription assay
mixtures, NTPs are present and used by Rho as substrates to
mediate the termination process. The interaction of Rho
with RNA activates catalytic hydrolysis of NTPs to the
corresponding NDP and inorganic phosphate (Pi). When the
RNA is a nascent transcript, the hydrolysis reaction
becomes coupled to the release of the transcript and hence
its termination [8,9]. However, even after the transcript has
been released, Rho will continue to interact with it and to
hydrolyze ATP at a steady-state turnover rate of as much as
60 NTPs s� 1 per Rho [29,45]. The hydrolysis reaction is
absolutely dependent on RNA. Even though some single-
stranded DNA molecules can bind to Rho nearly as tightly
as RNA can, those DNA molecules do not activate any ATP
hydrolysis [24]. On the other hand, the release reaction is
not limited to polymerase and DNA template attached to the
3V end of a transcript. Rho will also catalyze the release of
DNA molecules or DNA oligonucleotides that are paired
with a segment (20–40 base pairs) of the RNA, as long as
the DNA is on the 3V side of the rut segment of the RNA
[61,62]. This release is again coupled to NTP hydrolysis.
Thus, this ability to act as a hybrid helicase indicates that
Rho can move on a transcript in a direction that will bring it
into contact with the RNA polymerase that is synthesizing
the RNA. This tracking action on the RNA could also be the
driving force that pulls the transcript from its complex with
the polymerase and the DNA template.
Currently, there is a dispute over the nature of the
tracking reaction. In one interpretation, which is supported
by an elegant experiment [63], Rho maintains a contact with
the rut site used for its initial tight binding to the transcript
[64]. In this mechanism, Rho remains tethered to the rut site
on the RNA while it is tracking toward the 3V end of that
same RNA [63]. In an alternative view [65], Rho is
proposed to migrate down the RNA without maintaining
its original contact. Because it is difficult to prove that one
Rho is making separate contacts at two distinct regions of an
RNA during the migration process, neither mechanism can
be eliminated at this point.
Part of the dispute is on the number and placement of
sites on Rho for binding to RNA. Rho appears to have two
kinds of sites for binding RNA. This became evident with
the finding that ATP hydrolysis by Rho could be activated
by combinations of single-stranded DNA and short oligor-
ibonucleotides [66]. One kind of site binds both RNA and
DNA while the other site is specific for RNA. NTP
hydrolysis would then be dependent upon interactions at
both sites. It is well established that both RNA and DNA
can bind to the OB-fold segment of the RNA-BD, thus
placing the primary site in that domain [33]. However, until
recently, the location of the secondary site was not known.
One reasonable suggestion was that the two kinds of sites
could be different conformations of the site, the OB fold, in
the RNA-BD in different subunits of hexameric Rho [65].
This interpretation is supported by the finding that three of
the subunits can bind oligo(C) with high affinity and three
bind it with lower affinity [31]. However, two lines of recent
evidence suggest that RNA can also interact with a site that
is in the ATP-BD.
The first line of evidence is from cross-linking studies, in
which a photoactivatable chemical group on a loop region
(the R-loop, see Fig. 1) of the ATP-BD form cross-links to
RNA if the RNA has a rut sequence and an RNA segment
on the 3V of the rut sequence [23]. In contrast, an RNA that
has its 3V end at the end of the rut sequence forms cross-
links only with a group placed in the OB fold of the RNA-
BD. The position of the R loop in the subunits of the
homology-modeled structure of Rho is in the hole in the
center of the ring-shaped structure. This result suggests that
the rut segment of the RNA makes contacts in the RNA-BD
while RNA to the 3V of rut passes through the hole of the
Rho hexamer. This passage of a polynucleotide segment
through the hole of a ring-shaped hexamer is a characteristic
of hexameric DNA helicases, proteins that are structurally
and functionally similar to Rho [67–69].
The second line of evidence has come from studies of the
protection of Rho from cleavage by reactive peroxyl-species
generated by the interaction of chelated iron with hydrogen
peroxide [70]. An exposed part of Rho on a loop in the ATP-
BD, in this case the Q-loop (see Fig. 1), which also faces
into the hole of the Rho hexamer, becomes protected
specifically by RNA, not DNA. On the other hand, both
RNA and DNA gave extensive protection of accessible parts
of the OB-fold in the RNA-BD. These results suggest the
presence of an RNA-specific site on or near the Q-loop. The
amino acid residues on this loop are well conserved in the
Rho homologues, and mutational studies have now demon-
J.P. Richardson / Biochimica et Biophysica Acta 1577 (2002) 251–260 255
strated that changes of any of the four Q-loop-residues that
can make hydrogen bonds or ionic interactions to alanines
abrogate Rho termination function [70a]. More specifically,
these mutants are very defective in their interactions with
RNA in their secondary site but not in their primary site.
Taken together, the results of the protection studies and the
functional analysis of the Q-loop mutations indicate that the
Q-loop plays an important role in the interactions of Rho
with RNA. They are also consistent with the model that has
the 3V segment of RNA passing through the hole in the ring-
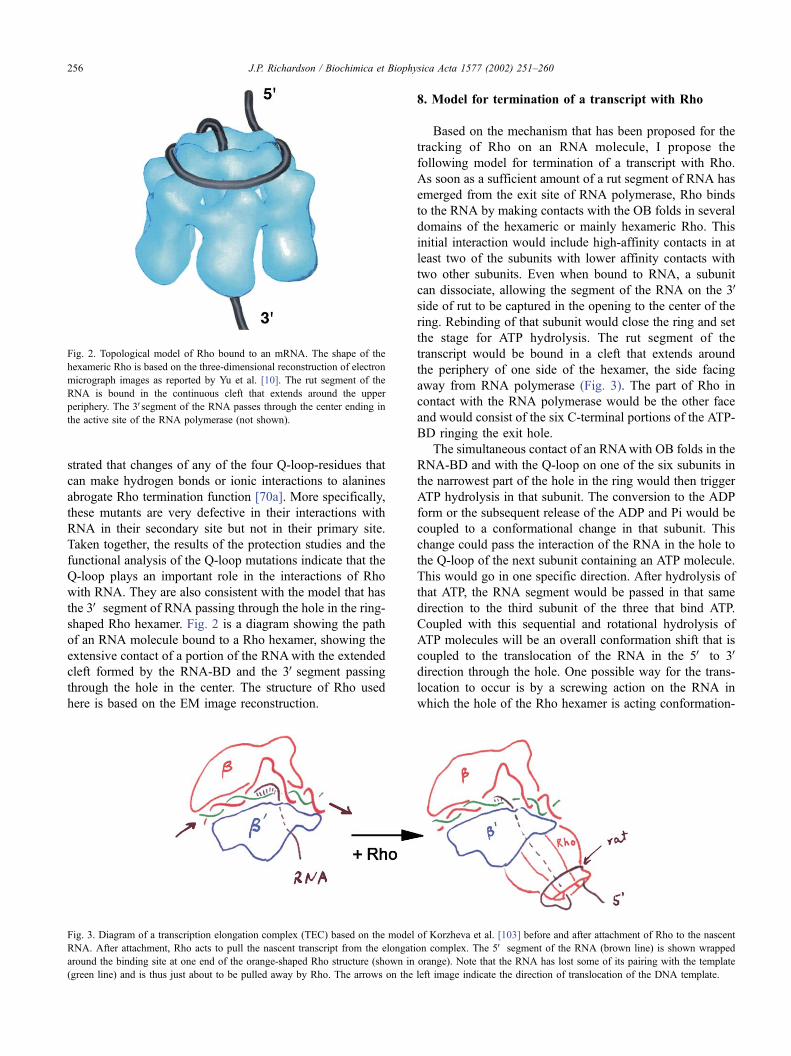
shaped Rho hexamer. Fig. 2 is a diagram showing the path
of an RNA molecule bound to a Rho hexamer, showing the
extensive contact of a portion of the RNAwith the extended
cleft formed by the RNA-BD and the 3Vsegment passing
through the hole in the center. The structure of Rho used
here is based on the EM image reconstruction.
8. Model for termination of a transcript with Rho
Based on the mechanism that has been proposed for the
tracking of Rho on an RNA molecule, I propose the
following model for termination of a transcript with Rho.
As soon as a sufficient amount of a rut segment of RNA has
emerged from the exit site of RNA polymerase, Rho binds
to the RNA by making contacts with the OB folds in several
domains of the hexameric or mainly hexameric Rho. This
initial interaction would include high-affinity contacts in at
least two of the subunits with lower affinity contacts with
two other subunits. Even when bound to RNA, a subunit
can dissociate, allowing the segment of the RNA on the 3Vside of rut to be captured in the opening to the center of the
ring. Rebinding of that subunit would close the ring and set
the stage for ATP hydrolysis. The rut segment of the
transcript would be bound in a cleft that extends around
the periphery of one side of the hexamer, the side facing
away from RNA polymerase (Fig. 3). The part of Rho in
contact with the RNA polymerase would be the other face
and would consist of the six C-terminal portions of the ATP-
BD ringing the exit hole.
The simultaneous contact of an RNAwith OB folds in the
RNA-BD and with the Q-loop on one of the six subunits in
the narrowest part of the hole in the ring would then trigger
ATP hydrolysis in that subunit. The conversion to the ADP
form or the subsequent release of the ADP and Pi would be
coupled to a conformational change in that subunit. This
change could pass the interaction of the RNA in the hole to
the Q-loop of the next subunit containing an ATP molecule.
This would go in one specific direction. After hydrolysis of
that ATP, the RNA segment would be passed in that same
direction to the third subunit of the three that bind ATP.
Coupled with this sequential and rotational hydrolysis of
ATP molecules will be an overall conformation shift that is
coupled to the translocation of the RNA in the 5V to 3Vdirection through the hole. One possible way for the trans-
location to occur is by a screwing action on the RNA in
which the hole of the Rho hexamer is acting conformation-
Fig. 2. Topological model of Rho bound to an mRNA. The shape of the
hexameric Rho is based on the three-dimensional reconstruction of electron
micrograph images as reported by Yu et al. [10]. The rut segment of the
RNA is bound in the continuous cleft that extends around the upper
periphery. The 3Vsegment of the RNA passes through the center ending in
the active site of the RNA polymerase (not shown).
Fig. 3. Diagram of a transcription elongation complex (TEC) based on the model of Korzheva et al. [103] before and after attachment of Rho to the nascent
RNA. After attachment, Rho acts to pull the nascent transcript from the elongation complex. The 5V segment of the RNA (brown line) is shown wrapped
around the binding site at one end of the orange-shaped Rho structure (shown in orange). Note that the RNA has lost some of its pairing with the template
(green line) and is thus just about to be pulled away by Rho. The arrows on the left image indicate the direction of translocation of the DNA template.
J.P. Richardson / Biochimica et Biophysica Acta 1577 (2002) 251–260256

ally like a nut on a helical single-stranded RNA. This action
could pull the RNA through with a step of about one
nucleotide per ATP hydrolyzed, as has been measured
[71], or at a linear translocation rate as fast as 60 nucleotides
per second (based on the ATP hydrolysis turnover number).
This estimated rate is in the right range to be a bit faster than
the average rate of transcript elongation of about 50 nucleo-
tides per second [1]. Hence, when RNA polymerase reaches
a pause site after Rho is firmly attached, the rate would be
fast enough to pull the RNA out of the RNA polymerase.
The dissociation of the RNA from its base-pairing inter-
actions with the template DNA could be by one of the
proposed mechanisms for helicases [72]. The bases paired
at the ends of double helices are known to separate briefly as
a result of thermal motions (called breathing). If Rho pulls
the RNA into its hole by a nucleotide step during the
temporary base pair separation, it will capture the RNA in
a form that will not be able to reform that terminal pair with
the DNA template. This could happen as Rho is pulling at the
RNA at the exit site with the contact (perhaps nonspecific) of
Rho with RNA polymerase acting as the leverage for the
pulling action. Step by step the pairing with the template is
broken and the RNA pulled completely through the exit site
to affect the release of the transcript. Fig. 3 shows a diagram
of a Rho molecule pulling the transcript partially away from
its interactions with the DNA template in RNA polymerase.
9. NusG is a Rho cofactor
In the cell, the function of Rho at some terminators
depends on the presence of a 21 kDa monomeric protein
called NusG [73]. This protein was discovered because of its
function as a host cell cofactor for the transcription anti-
termination function of bacteriophage E protein N [74].
However, it has since been found to be a transcript elonga-
tion factor [75], which acts to enhance the escape of RNA
polymerase from class II pause sites [49] and has the general
effect of increasing the rate of transcript elongation by about
25% [76]. NusG is an essential protein for viability of E.
coli [77]. In cells that are experimentally depleted of NusG,
some Rho-dependent terminators no longer function prop-
erly, though others seem less affected [73]. A requirement of
NusG for Rho function was overlooked, presumably
because the conditions that were used to study Rho function
in vitro had been optimized for Rho function by reduction of
the NTP concentration. When transcripts are elongated in
vitro at the same rate that occurs in vivo, some terminators
do show a dependence on NusG for efficient Rho-mediated
termination [78]. These results suggest that NusG is over-
coming a kinetic deficiency of Rho to act by itself under
certain conditions.
NusG binds both RNA polymerase [79] and Rho [26],
although it is not known yet whether it can bind both
simultaneously. Assuming that it can, it could act either to
bring Rho into close proximity to the emerging transcript or
to stabilize the complex of Rho with the nascent transcript
and RNA polymerase [80]. By forming a complex of Rho
with RNA polymerase before the rut site emerges, the
binding of the rut segment to the primary RNA-binding site
would become a unimolecular reaction. Thus, this hypoth-
esis is consistent with the evidence that NusG is acting to
overcome a kinetic deficiency of Rho to act by itself.
NusG has been shown to make a 1:1 complex with
hexameric Rho [27]. The position of this unique binding
site on the Rho hexamer has not been located, but because it
is unique, it is likely to be in one of the two entrances to the
hole. If a NusG bound to RNA polymerase also binds to
Rho on the side with the RNA-BD, it would orient Rho so
that the emerging transcript would be in close proximity to
the cleft of the RNA-BD. However, once the transcript is
bound tightly in part of that cleft, that interaction with NusG
would have to be released to allow the RNA to enter the
hole. In addition Rho would have to rotate 180j end to end
to have the hole at the C-terminal domains facing RNA
polymerase as is shown in the diagram in Fig. 3. This would
be some fancy footwork. An alternative suggested role for
NusG is that it favors an RNA polymerase conformation
that enhances RNA release [27,81].
NusA protein has been shown to bind to Rho and RNA
polymerase and can do so simultaneously [28]. NusA, like
NusG, is a host factor for bacteriophage E protein N-
mediated anti-termination. It is also a transcript elongation
factor, but unlike NusG, NusA acts to enhance pausing at a
certain class of pause sites and thus slows the rate of
transcript elongation [49,51]. Its action in Rho-dependent
termination has been ambiguous. In most assays, it acts to
inhibit Rho function, perhaps by competing with Rho for rut
segment interactions [40,76,82]. However, there is also a
report in which NusA is shown to enhance Rho action [83],
as might be expected from its ability to enhance pausing.
NusA is essential for viability of E. coli [84]. However, a
conditionally lethal mutation in nusA can be suppressed by
certain mutations in rho, namely mutations that allow
expression of an active but defective Rho protein [85]. This
observation suggests that the functions of the two proteins
are competitive. One possible role of NusA is to ensure a
close coupling of translation with transcription, perhaps by
retarding transcript elongation to prevent RNA polymerase
from moving ahead of a translating ribosome. Although this
is an attractive model and is consistent with the suppression
of a NusA defect by a Rho defect, direct evidence for this
function is lacking.
10. Biological role of Rho
As mentioned in the introduction, E. coli uses two kinds
of transcript terminators. This is not true for all bacteria.
With the advent of genome sequencing, it is now known that
a few bacteria, a Mycoplasma and a Synecchocystis for
instance, lack a rho gene. On the other hand, most bacteria
J.P. Richardson / Biochimica et Biophysica Acta 1577 (2002) 251–260 257

have a rho gene. It is present in organisms in lineages that
diverged from that of E. coli over a billion years ago,
indicating that it is not a recent addition [21]. However,
there is no evidence yet for a rho-like gene in archael or
eukaroytic organisms. Thus, Rho is not essential for life or
even life in the bacteria, but it arose early in evolution,
suggesting that it has a useful function for most bacteria.
The question of whether eukaroytic cells use an ATP-
dependent mechanism for termination of transcripts is
considered in the final section of this review.
The rho gene is essential for E. coli and a few other
organisms [86,87]. In general, cells that are sensitive to the
Rho-specific antibiotic bicyclomycin probably have essen-
tial rho genes [88,89]. However, rho is not essential for
Bacillus subtilis [90] or for Staphylococcus aureus [91]. In
B. subtilis, rho is expressed but the level of Rho in the cells
is 50-fold lower than the level of Rho in E. coli [92]. The
Rho that is present does function at least to regulate its own
synthesis. Altogether, these observations suggest that B.
subtilis does not make extensive use of Rho-dependent
terminators for the orderly expression of its genes, but
nonetheless maintains the gene and expresses it for some
function.
The extent to which E. coli makes use of its two kinds
of transcript terminators is not known. Putative intrinsic
terminators can be identified by genome analysis, but the
actual efficiency of functioning of a sequence as a termi-
nator is still difficult to predict. For example, a sequence
for an intrinsic terminator is found at the end of the trp
operon of E. coli [93]. However, functional studies sub-
sequently showed that that terminator was very weak and
that a much more efficient Rho-dependent terminator was
responsible for terminating most transcripts for that operon
[47]. A similar organization was found in a number of other
operons [94]. Because Rho-dependent terminators lack a
readily identifiable consensus sequence, it is not known
how frequently they are used. Depletion of a cell of Rho
may be lethal because of the catastrophic effect of the
expression of many different sequences that normally are
not expressed or because products of the inadvertent
expression are toxic.
The fact that the rho gene arose early in the natural
history of the bacteria suggests that it gives some selective
advantage to a cell. One likely advantage is that under
conditions of stress, it prevents the continued synthesis of
an RNA that cannot be translated [35]. In E. coli, Rho has
been shown to terminate the synthesis of transcripts very
early in the lac operon when the cells are starved for amino
acids [95]. This comes about because the lac operon, like
many other operons, contains latent Rho-dependent termi-
nators within the early part of the gene [96]. These intra-
genic terminators do not function under conditions of
normal expression, presumably because a ribosome is
present translating the RNA as it emerges from the exit
site of RNA polymerase. This ribosome would block the
access of Rho to the RNA in a way that would allow it to
terminate transcription. However, when the movement of
the ribosome is slowed or blocked because of the absence
of an amino acid, a segment of a transcript containing a rut
site would become exposed allowing Rho to bind and
terminate the partial transcript by its normal mechanism.
The selective advantage this would have is that it would
prevent the loss of energy in making a transcript that will
not be translated.
The broad sequence specificity of a Rho dependent
terminator as well as the dependence of access of Rho to
the transcript makes this type of terminator well suited for
this stress-protection function. These features also made
Rho-dependent terminators very suitable for placement at
the end of a gene or operon.
11. Is a Rho factor present in eukaryotic cells?
A factor that is analogous to Rho has not yet been
discovered in eukaryotic cells. These cells certainly lack a
gene that has the strongly conserved sequence features that
are characteristic of the rho genes in bacteria. However,
one aspect of transcript termination with RNA polymerase
II is similar to the Rho-dependent mechanism and that is
termination occurs in broad regions downstream from a
sequence signal [97,98]. Thus, these cells could have
another kind of protein with the ability to bind to a
transcript and act as a molecular machine to translocate
along the RNA and wrest the transcript from RNA poly-
merase. An ATPase, called factor 2, which acts to disso-
ciate transcripts from RNA polymerase II, has been isolated
[99], but its function may be involved with release of
abortive products at the early stage of the transcription
process. It is not an RNA-binding protein. Instead, its
ATPase activity is stimulated by DNA [100]. Also, its
sequence makes it related to chromatin remodeling proteins
[101]. RNA polymerase I also makes use of a protein
factor, called Reb1p in yeast, to mediate release of tran-
scripts at terminators, but this factor binds to DNA and
does not appear to require ATP hydrolysis [102].
Acknowledgements
I thank Ronnie Wei for preparation of Figs. 1 and 2 and
Lislott Richardson, R. Wei and Brandt Burgess for
discussions. Research cited from my laboratory was
supported by NIH Grant GM56095.
References
[1] J.P. Richardson, J. Greenblatt, in: F.C. Neidhardt, R. Curtiss III, J.L.
Ingraham, E.C.C. Lin, K.B. Low, B. Magasanik, W.S. Reznikoff, M.
Riley, M. Schaechter, H.E. Umbarger (Eds.), Escherichia coli and
Salmonella: Cellular and Molecular Biology, 2nd Ed., Washington,
DC, 1996, pp. 822–848.
J.P. Richardson / Biochimica et Biophysica Acta 1577 (2002) 251–260258

[2] K.S. Wilson, P.H. von Hippel, J. Mol. Biol. 244 (1994) 36–51.
[3] R.A. Mooney, I. Artsimovitch, R. Landick, J. Bacteriol. 180 (1998)
3265–3275.
[4] I. Gusarov, E. Nudler, Mol. Cell 3 (1999) 495–504.
[5] W.S. Yarnell, J.W. Roberts, Science 284 (1999) 611–615.
[6] Y. dVAubenton Carafa, E. Brody, C. Thermes, J. Mol. Biol. 216
(1990) 835–858.
[7] F.H. Martin , I. Tinoco Jr., Nucleic Acids Res. 8 (1980) 2295–2299.
[8] K. Shigesada, C.W. Wu, Nucleic Acids Res. 8 (1980) 3355–3369.
[9] J.P. Richardson, R. Conaway, Biochemistry 19 (1980) 4293–4299.
[10] X. Yu, T. Horiguchi, K. Shigesada, E.H. Egelman, J. Mol. Biol. 299
(2000) 1279–1287.
[11] L.R. Finger, J.P. Richardson, J. Mol. Biol. 156 (1982) 203–219.
[12] J. Geiselmann, T.D. Yager, S.C. Gill, P. Calmettes, P.H. von Hippel,
Biochemistry 31 (1992) 111–121.
[13] B.R. Burgess, J.P. Richardson, J. Biol. Chem. 276 (2001)
17106–17110.
[14] D.E. Kim, S.S. Patel, J. Biol. Chem. 274 (1999) 32667–32671.
[15] J.L. Pinkham, T. Platt, Nucleic Acids Res. 11 (1983) 3531–3545.
[16] D. Modrak, J.P. Richardson, Biochemistry 33 (1994) 8292–8299.
[17] T.J. Allison, T.C. Wood, D.M. Briercheck, F. Rastinejad, J.P.
Richardson, G.S. Rule, Nat. Struct. Biol. 5 (1998) 352–356.
[18] A.J. Dombroski, J.R. LaDine, R.L. Cross, T. Platt, J. Biol. Chem.
263 (1988) 18810–18815.
[19] J.W. Dolan, N.F. Marshall, J.P. Richardson, J. Biol. Chem. 265
(1990) 5747–5754.
[20] D.M. Briercheck, T.C. Wood, T.J. Allison, J.P. Richardson, G.S.
Rule, Nat. Struct. Biol. 5 (1998) 393–399.
[21] T. Opperman, J.P. Richardson, J. Bacteriol. 176 (1994) 5033–5043.
[22] Y. Miwa, T. Horiguchi, K. Shigesada, J. Mol. Biol. 254 (1995)
815–837.
[23] B.R. Burgess, J.P. Richardson, J. Biol. Chem. 276 (2000) 4182–4189.
[24] G.R. Galluppi, J.P. Richardson, J. Mol. Biol. 138 (1980) 513–539.
[25] J.A. McSwiggen, D.G. Bear, P.H. von Hippel, J. Mol. Biol. 199
(1988) 609–622.
[26] J. Li, S.W. Mason, J. Greenblatt, Genes Dev. 7 (1993) 161–172.
[27] Z. Pasman, P.H. von Hippel, Biochemistry 39 (2000) 5573–5585.
[28] M.C. Schmidt, M.J. Chamberlin, J. Biol. Chem. 259 (1984)
15000–15002.
[29] C. Lowery-Goldhammer, J.P. Richardson, Proc. Natl. Acad. Sci.
U. S. A. 71 (1974) 2003–2007.
[30] D.G. Bear, P.S. Hicks, K.W. Escudero, C.L. Andrews, J.A.
McSwiggen, P.H. von Hippel, J. Mol. Biol. 199 (1988) 623–635.
[31] J. Geiselmann, T.D. Yager, P.H. von Hippel, Protein Sci. 1 (1992)
861–873.
[32] Y. Wang, P.H. von Hippel, J. Biol. Chem. 268 (1993) 13947–13955.
[33] D.M. Briercheck, T.J. Allison, J.P. Richardson, J.F. Ellena, T.C.
Wood, G.S. Rule, J. Biomol. NMR 8 (1996) 429–444.
[34] C.E. Bogden, D. Fass, N. Bergman, M.D. Nichols, J.M. Berger, Mol.
Cell 3 (1999) 487–493.
[35] J.P. Richardson, Cell 64 (1991) 1047–1049.
[36] P. Alifano, F. Rivellini, D. Limauro, C.B. Bruni, M.S. Carlomagno,
Cell 64 (1991) 553–563.
[37] W.D. Morgan, D.G. Bear, B.L. Litchman, P.H. von Hippel, Nucleic
Acids Res. 13 (1985) 3739–3754.
[38] C.Y. Chen, J.P. Richardson, J. Biol. Chem. 262 (1987) 11292–11299.
[39] D.G. Bear, D.S. Peabody, Trends Biochem. Sci. 13 (1988) 343–347.
[40] L.F. Lau, J.W. Roberts, R. Wu, Proc. Natl. Acad. Sci. U. S. A. 79
(1981) 6171–6175.
[41] C.Y. Chen, G.R. Galluppi, J.P. Richardson, Cell 46 (1986) 1023–1028.
[42] E. Gan, J.P. Richardson, Biochemistry 38 (1999) 16882–16888.
[43] I. Faus, J.P. Richardson, Biochemistry 28 (1989) 3510–3517.
[44] J.E. Graham, J.P. Richardson, J. Biol. Chem. 273 (1998) 20764–20769.
[45] L.V. Richardson, J.P. Richardson, J. Biol. Chem. 271 (1996)
21597–21603.
[46] W.D. Morgan, D.G. Bear, P.H. von Hippel, J. Biol. Chem. 258
(1983) 9553–9564.
[47] A.M. Wu, G.E. Christie, T. Platt, Proc. Natl. Acad. Sci. U. S. A. 78
(1981) 2913–2917.
[48] J.R. Levin, M.J. Chamberlin, J. Mol. Biol. 196 (1987) 61–84.
[49] I. Artsimovitch, R. Landick, Proc. Natl. Acad. Sci. U. S. A. 97
(2000) 7090–7095.
[50] C.L. Chan, R. Landick, J. Mol. Biol. 233 (1993) 25–42.
[51] L.F. Lau, J.W. Roberts, R. Wu, J. Biol. Chem. 258 (1983) 9391–9397.
[52] W.D. Morgan, D.G. Bear, P.H. von Hippel, J. Biol. Chem. 258
(1983) 9565–9574.
[53] M.A. Ceruzzi, S.L. Bektesh, J.P. Richardson, J. Biol. Chem. 260
(1985) 9412–9418.
[54] J.P. Richardson, J.L. Carey, J. Biol. Chem. 257 (1982) 5767–5771.
[55] D.J. Jin, R.R. Burgess, J.P. Richardson, C.A. Gross, Proc. Natl.
Acad. Sci. U. S. A. 89 (1992) 1453–1457.
[56] C. Lowery, J.P. Richardson, J. Biol. Chem. 252 (1977) 1375–1380.
[57] B.L. Stitt, J. Biol. Chem. 263 (1988) 11130–11137.
[58] J. Geiselmann, P.H. von Hippel, Protein Sci. 1 (1992) 850–860.
[59] D.E. Kim, K. Shigesada, S.S. Patel, J. Biol. Chem. 274 (1999)
11623–11628.
[60] B.L. Stitt, Biochemistry 40 (2001) 2276–2281.
[61] C.A. Brennan, A.J. Dombroski, T. Platt, Cell 48 (1987) 945–952.
[62] K.M. Walstrom, J.M. Dozono, S. Robic, P.H. von Hippel, Biochem-
istry 36 (1997) 7980–7992.
[63] E.J. Steinmetz, T. Platt, Proc. Natl. Acad. Sci. U. S. A. 91 (1994)
1401–1405.
[64] I. Faus, J.P. Richardson, J. Mol. Biol. 212 (1990) 53–66.
[65] J. Geiselmann, Y. Wang, S.E. Seifried, P.H. von Hippel, Proc. Natl.
Acad. Sci. U. S. A. 90 (1993) 7754–7758.
[66] J.P. Richardson, J. Biol. Chem. 257 (1982) 5760–5766.
[67] M.J. Jezewska, S. Rajendran, W. Bujalowski, Biochemistry 37
(1998) 3116–3136.
[68] H.H. Egelman, X. Yu, R. Wild, M.M. Hingorani, S.S. Patel, Proc.
Natl. Acad. Sci. U. S. A. 92 (1995) 3869–3873.
[69] S.S. Patel, K.M. Picha, Annu. Rev. Biochem. 69 (2000) 651–697.
[70] R.R. Wei, J.P. Richardson, J. Biol. Chem. 276 (2001) 28380–28387.
[71] R.R. Wei, J.P. Richardson, J. Mol. Biol. 314 (2001) 1007–1015.
[72] K.M. Walstrom, J.M. Dozono, P.H. von Hippel, Biochemistry 36
(1997) 7993–8004.
[73] P.H. von Hippel, E. Delagoutte, Cell 104 (2001) 177–190.
[74] S.L. Sullivan, M.E. Gottesman, Cell 68 (1992) 989–994.
[75] S.W. Mason, J. Greenblatt, Genes Dev. 5 (1991) 1504–1512.
[76] E. Burova, S.C. Hung, V. Sagitov, B.L. Stitt, M.E. Gottesman, J.
Bacteriol. 177 (1995) 1388–1392.
[77] C.M. Burns, L.V. Richardson, J.P. Richardson, J. Mol. Biol. 278
(1998) 307–316.
[78] W.L. Downing, S.L. Sullivan, M.E. Gottesman, P.P. Dennis, J. Bac-
teriol. 172 (1990) 1621–1627.
[79] C.M. Burns, J.P. Richardson, Proc. Natl. Acad. Sci. U. S. A. 92
(1995) 4738–4742.
[80] J. Li, R. Horwitz, S. McCracken, J. Greenblatt, J. Biol. Chem. 267
(1992) 6012–6019.
[81] K.W. Nehrke, T. Platt, J. Mol. Biol. 243 (1994) 830–839.
[82] C.M. Burns, W.L. Nowatzke, J.P. Richardson, J. Biol. Chem. 274
(1999) 5245–5251.
[83] C.D. Sigmund, E.A. Morgan, Biochemistry 27 (1988) 5622–5627.
[84] M. Kainz, R.L. Gourse, J. Mol. Biol. 284 (1998) 1379–1390.
[85] M.G. Craven, D.I. Friedman, J. Bacteriol. 173 (1991) 1485–1491.
[86] C. Zheng, D.I. Friedman, Proc. Natl. Acad. Sci. U. S. A. 91 (1994)
7543–7547.
[87] S. Brown, E.R. Brickman, J. Beckwith, J. Bacteriol. 146 (1981)
422–425.
[88] A. Das, D. Court, S. Adhya, Proc. Natl. Acad. Sci. U. S. A. 73
(1976) 1959–1963.
[89] A. Zwiefka, H. Kohn, W.R. Widger, Biochemistry 32 (1993)
3564–3570.
[90] W.L. Nowatzke, E. Keller, G. Koch, J.P. Richardson, J. Bacteriol. 179
(1997) 5238–5240.
J.P. Richardson / Biochimica et Biophysica Acta 1577 (2002) 251–260 259

[91] P.G. Quirk , E.A. Dunkley Jr., P. Lee, T.A. Krulwich, J. Bacteriol. 175
(1993) 647–654.
[92] R.S. Washburn, A. Marra, A.P. Bryant, M. Rosenberg, D.R. Gentry,
Antimicrob. Agents Chemother. 45 (2001) 1099–1103.
[93] C.J. Ingham, J. Dennis, P.A. Furneaux, Mol. Microbiol. 31 (1999)
651–663.
[94] A.M.Wu, T. Platt, Proc.Natl. Acad. Sci. U. S.A. 75 (1978) 5442–5446.
[95] J.H. Sameshima, R.C. Wek, G.W. Hatfield, J. Biol. Chem. 264
(1989) 1224–1231.
[96] E.C. Ruteshouser, J.P. Richardson, J. Mol. Biol. 208 (1989) 23–43.
[97] S. Adhya, M. Gottesman, B. de Crombrugghe, D. Court, in: M.
Chamberlin, R. Losick (Eds.), Transcription Termination Regulates
Gene Expression, Cold SpringHarbor, NewYork, 1976, pp. 719–730.
[98] N.J. Proudfoot, Trends Biochem. Sci. 14 (1989) 105–110.
[99] S. Connelly, J.L. Manley, Genes Dev. 2 (1988) 440–452.
[100] Z. Xie, D.H. Price, J. Biol. Chem. 271 (1996) 11043–11046.
[101] Z. Xie, D.H. Price, J. Biol. Chem. 273 (1998) 3771–3777.
[102] M. Liu, Z. Xie, D.H. Price, J. Biol. Chem. 273 (1998) 25541–25544.
[103] W.H. Lang, R.H. Reeder, Proc. Natl. Acad. Sci. U. S. A. 92 (1995)
9781–9785.
[104] N. Korzheva, A. Mustaev, M. Kozlov, A. Malhotra, V. Nikiforov, A.
Goldfarb, S.A. Darst, Science 289 (2000) 619–625.
J.P. Richardson / Biochimica et Biophysica Acta 1577 (2002) 251–260260












![P4-ATPases: lipid flippases in cell membranes · recently treated different aspects of P4-ATPases [ 18, 77, 86, 94]. In this review, we will first provide an overview on the functional](https://img.pdfslide.us/doc/110x75/5ede6227ad6a402d6669b49d/p4-atpases-lipid-flippases-in-cell-membranes-recently-treated-different-aspects.jpg)
![Review V-ATPases and osteoclasts: ambiguous future of V ...thno.org/v08p5379.pdfosteoclasts, which is a key factor for bone resorption [2]. The V-ATPases-related regulation of extracellular](https://img.pdfslide.us/doc/110x75/5ee15f47ad6a402d666c473b/review-v-atpases-and-osteoclasts-ambiguous-future-of-v-thnoorg-osteoclasts.jpg)















