Embed Size (px)
Citation preview

APPLIE MICRoBIoLoGY, Sept. 1968, p. 1331-1336Copyright @ 1968 American Society for Microbiology
Vol. 16, No. 9Printed in U.S.A.
Rhinovirus Plaque Formation in WI-38 Cells withMethylcellulose Overlay
TIMOTHY M. DOLAN,' JAMES D. FENTERS,2 PATRICIA A. FORDYCE,3 AND JACOB C. HOLPER
Devartment of Infectious Diseases, Scientific Division, Abbott Laboratories, North Chicago, Illinois 60064
Received for publication 13 June 1968
A sensitive, reliable plaque assay system is described for five rhinoviruses usingfreshly prepared methylcellulose overlay and human embryonic diploid cells. Circu-lar plaques with irregular edges, 2 mm in size, were formed by rhinoviruses 1A, 2, 6,and 13 after 6 or 7 days of incubation. A fifth rhinovirus, 17, formed a 1- to 2-mmfeather plaque after 14 days of incubation. Plaque counts of rhinoviruses 1A and 13were not affected by varying thepH of the overlay from 6.9 to 7.5. Plaque sizes andplaque-forming unit values of high passage rhinoviruses IA and 13 were equivalentwhen tested at 26, 31, or 36 C. The rhinoviruses tested were sensitive to incubationat 40 C or heating at 50 C. Enhancement of plaques was observed when Mg++ was
incorporated into agar overlays, but enhancement did not occur when Mg++ was
added to methylcellulose overlays.
The plaque assay for animal viruses was firstdescribed by Dulbecco (2) and has since foundwidespread application to many virus-cell sys-tems, incluiding rhinoviruses (5, 11, 13, 14, 18).The original method of observing small foci ofdegenerating cells infected by rhinoviruses underthe microscope (13) was replaced by the produc-tion of plaques under agar (14). Plaque charac-teristics such as shape, size, and degree of lysisof cells caused by different rhinoviruses may beobserved under agar (5, 11, 14, 18). Variationsin the use of primary tissues or human heteroploidcells, methylcellulose or agar overlay, or additivessuch as diethylaminoethyl (DEAE)-dextran orMg++ to the overlay have been described (5).Genetic markers such as the bicarbonate (d)marker (16) or temperature (T) markers (12)may be performed with a plaque assay system.
Methylcellulose is an agar substitute first de-scribed by Hotchin (7) in plaque assays of herpessimplex virus. The advantages of it are that it isnontoxic (7), chemically defined (15), avoidsinhibitory effect of agar (15), and it is relativelyeasy to handle (7).The formation of plaques with rhinovirus 2 in
heteroploid cells using methylcellulose has beenreported (5). The purpose of this paper is topresent a sensitive, reliable assay system for five
1 Present address: University of North Dakota,Department of Microbiology, Grand Forks, N.D.
21Present address: Life Sciences, IIT Research In-stitute, Chicago, Ill. 60616.
3Present address: G. D. Searle, P.O. Box 5110,Chicago, Ill. 60680.
rhinoviruses using the methylcellulosesystem with human diploid cells.
assay
MATERIALS ANDW METHODSTissue culture. The human embryonic diploid cell
strain, WI-38, used in these experiments was originallyobtained from Flow Laboratories, Inc., Rockville,Md. and from L. Hayflick of the Wistar InstitutePhiladelphia, Pa. The methods used for the passage ofWI-38 cells were as described by Hayflick and Moor-head (6). Growth medium was Eagle's basal essentialmedium (BME; Flow Laboratories, Inc.) with theaddition of 10% fetal bovine serum, 50 units per ml ofpolymyxin, and 100lg per ml of neomycin sulfate.Confluent monolayers of WI-38 cells were obtainedin flasks (Falcon Plastics, Los Angeles, Calif.).
Overlays. A 3% stock methylcellulose (400 centi-poises, Fisher Chemical Co., Pittsburgh, Pa.) solutionwas prepared with triple distilled water and auto-claved at 124 C for 20 min. As the methylcellulosecooled to room temperature, it was occasionallyrotated by hand to effect the solution. Immediatelyafter the methyl-cellulose dissolved, an equal volumeof twofold concentration of media with 10% fetalcalf serum (Hyland Laboratories, Los Angeles, Calif.)was added. The concentrated media consisted ofdouble volumes of vitamins, L-glutamines, and essen-tial amino acids in a 2X solution of Earle's BSS (2XBME). Modified lactalbumin hydrolysate (14), modi-fied Earle's BSS (10), or a 2X solution of 199 wereused in some experiments. Final concentrations of1.25% Noble agar or Difco agar were used in com-bination with different media. DEAE-dextran (Phar-macia, Uppsala, Sweden) or 30 mm MgC12 was in-corporated into some experiments to note if plaquesize enhancement occurred.
Plaque assays. Monolayers of WI-38 cells were used
1331
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
m o
n 14
Oct
ober
202
1 by
197
.153
.84.
81.

DOLAN ET AL.
4 to 6 days after seeding the flasks. Prior to virusinoculation, the cell sheet was washed once with 2 mlof Earle's BSS, the supernatant fluid was decanted,and 2 ml of Earle's BSS was added. An 0.1-ml amountof the appropriate dilution of virus was added to eachof three flasks, with absorption for 2 hr at room tem-perature. The overlays were added to the flasks in 5-mivolumes using a Cornwall automatic syringe. At theend of the 6- to 7-day incubation period at 33 C, themethylcellulose overlay was decanted. A 4-nd amountof absolute methanol was added to each flask for 10min. The alcohol was removed, and 4 ml of a 1:10stock solution of Giemsa stain (Difco) was added for30 min. In experiments using agar, a second overlaycontaining a 1:20,000 final concentration of neutralred was added, and the flasks were incubated for 4hr at 36 C.
Linearity and temperature experiments. Linearityexperiments were performed according to the methodof Dulbecco and Vogt (3) using twofold dilutions ofvirus. The temperature genetic markers, T26, T30,and T40, were determined by incubating the test for7 days at the stated temperature (12). The T50 markerwas determined by the method of Hozinski et al. (8),except that the viruses were heated for 30 min at 50 Cin our system before plaque titrations.
Viruses. Rhinoviruses 1A, 2, 6, 13, and 17 wereused. The history of the viruses was given in an earlierpublication (4). The nomenclature used is as approvedby the Directors of the World Health OrganizationRespiratory and Enterovirus Reference Centers (9).Rhinoviruses 1 and 13, which had been passed twicein WI cells, were kindly supplied by Vernon Knight,College of Medicine, Baylor University, Houston,Tex.
Neutralization tests. An 0.3-nl amount of a 1:20dilution of heat-inactivated (56C, 30 min) bovineantisera (Abbott Laboratories, North Chicago, Ill.)was added to 0.3 ml of twofold dilutions of viruses,incubated at room temperature for 2 hr. An 0.1-mlamount of this mixture was inoculated into each ofthree flasks containing WI-38 cells. Absorption wasfor 2 hr at room temperature. After decanting thefluids, the BME-methylcellulose overlay was added,and the test was incubated for 7 days at 33 C. The cellswere then stained with Giemsa stain, and the plaqueswere counted.
RESULTS
Determination of overlay for rhinovirus plaques.Rhinoviruses 1A, 2, 13, and 17 produced con-sistent macroplaques with BME and methyl-cellulose on WI-38 cells (Table 1). The use ofmodified lactalbumin hydrolysate or modified 199with methylcellulose led to the formation ofmicroplaques, except with rhinovirus 2 whichformed macroplaques- with modified 199 andmethylcellulose. By contrast, under agar, onlymicroplaques or faint hazy macroplaques wereproduced by four rhinoviruses. Due to the lackof a strong background stain with neutral red,the plaques were difficult to observe, and the
TABLE 1. Selection of overlay for rhinovirus plaquesI~~~~~~~~~~~~~
Overlay'
LH, FCS, NA..LH, FCS, MC..199, FCS, NA..199, FCS, MC..BME, FCS, NABME, FCS,NA, D.......
BME, FCS,DA, D.......
BME, FCS,MC ........
BME, CS, MC..Earle's, CS,DA, D.......
Rhino- Rhino- Rhino- Rhino-virus virus virus virusIA 2 6 13
4b
0
x
x
x
x
-+-
+ +
x
x
Rbino-virus17
+
a"LH, modified lactalbumin hydrolysate; FCS,fetal calf serum; NA, Noble agar; MC, methyl-cellulose; BME, basal medium, Eagle; D, DEAE-dextran; DA, Difco agar; CS, agamma chickenserum.
b Symbols: +, macroplaques; i, microplaques;0, negative;-, not tested; X, faint hazy macro-plaques.
method was discarded in favor of the methyl-cellulose overlay.Optimal number of plaques of rhinoviruses
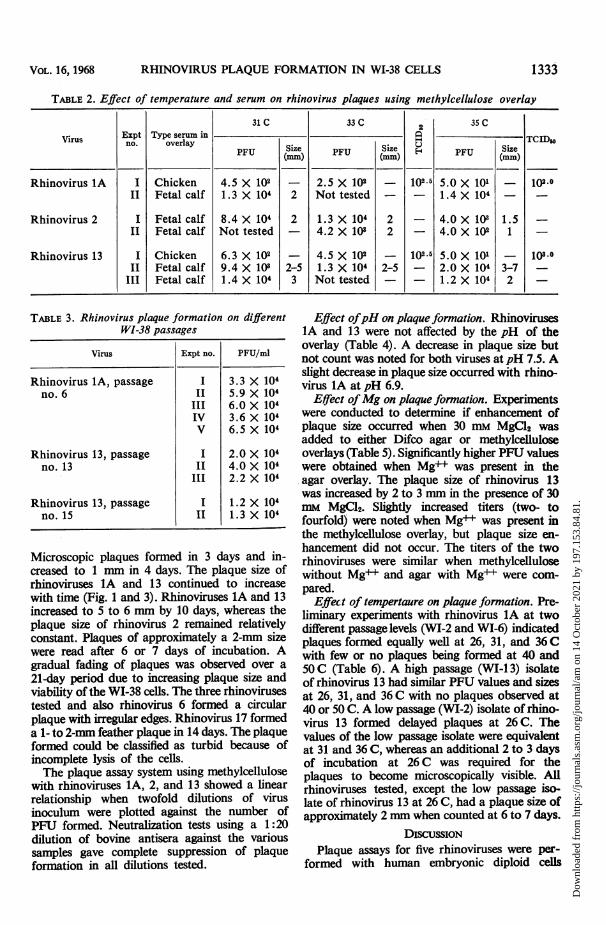
were produced at 31 or 33 C using methylcellu-lose, BME, and either agammaglobulin chickenserum or fetal calf serum (Table 2). Using fetalcalf serum in the overlay, equivalent plaque-forming unit (PFU) values were obtained at 31,33, and 35 C with rhinoviruses IA and 13. Withthe same overlay, the PFU values of rhinovirus 2appeared to decrease with increasing temperature.The PFU values of rhinoviruses lA and 13 alsodecreased with increased temperature with theagammaglobulin chicken serum overlay. ThePFU to TCID50 ratio was approximately 1:1 at33 C but ranged from 1:2 to 1:20 at 35 C withrhinoviruses 1A and 13, respectively, whenagamma chicken serum was used. Some granula-tion was observed in the monolayers containingchicken serum which was not observed with fetalbovine serum.
Virus stock of either rhinovirus lA or 13 didnot vary more than twofold when titrated ondifferent passages of WI-38 cells over a 6-monthperiod of time (Table 3).
Figures 1, 2, and 3 demonstrate the number ofmacroplaques formed and their size with rhino-viruses 1A, 2, and 13. Maximal plaque titer wasobtained in 4 to 5 days with the three rhino-viruses at 33 C using BME and methylcellulose.
1332 APPL. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
m o
n 14
Oct
ober
202
1 by
197
.153
.84.
81.
RHINOVIRUS PLAQUE FORMATION IN WI-38 CELLS
TABLE 2. Effect of temperature and serum on rhinovirus plaques using methylcellulose overlay
31C 33C 35C
Expt Type serum in
PU Size PU Size H F SizePU (MM) PU (MM) (mm)U
Rhinovirus 1A I Chicken 4.5 X 102 - 2.5 X 102 - 1O25 5.0 X 101 - 102.0II Fetal calf 1. 3 X 104 2 Not tested - - 1.4 X 104 -
Rhinovirus 2 I Fetal calf 8.4 X 104 2 1.3 X 104 2 - 4.0 X 103 1.5II Fetal calf Not tested - 4.2 X 108 2 - 4.0 X 102 1
Rhinovirus 13 I Chicken 6.3 X 102 - 4.5 X 102 - 102.5 5.0 X 101 - 103-II Fetal calf 9.4 X 102 2-5 1.3 X 104 2-5 - 2.0 X 104 3-7
III Fetal calf 1.4 X 104 3 Not tested - - 1.2 X 104 2 -
TABLE 3. Rhinovirus plaque formation on differentWI-38 passages
Virus Expt no. PFUFrml
Rhinovirus 1A, passage I 3.3 X 104no. 6 II 5.9 X 104
III 6.0 X 104IV 3.6 X 104V 6.5X 104
Rhinovirus 13, passage I 2.0 X 104no. 13 II 4.0 X 104
III 2.2 X 104
Rhinovirus 13, passage I 1.2 X 104no. 15 II 1.3 X 104
Microscopic plaques formed in 3 days and in-creased to 1 mm in 4 days. The plaque size ofrhinoviruses 1A and 13 continued to increasewith time (Fig. 1 and 3). Rhinoviruses IA and 13increased to 5 to 6 mm by 10 days, whereas theplaque size of rhinovirus 2 remained relativelyconstant. Plaques of approximately a 2-mm sizewere read after 6 or 7 days of incubation. Agradual fading of plaques was observed over a
21-day period due to increasing plaque size andviability of the WI-38 cells. The three rhinovirusestested and also rhinovirus 6 formed a circularplaque with irregular edges. Rhinovirus 17 formeda 1- to 2-mm feather plaque in 14 days. The plaqueformed could be classified as turbid because ofincomplete lysis of the cells.The plaque assay system using methylcellulose
with rhinoviruses 1A, 2, and 13 showed a linearrelationship when twofold dilutions of virusinoculum were plotted against the number ofPFU formed. Neutralization tests using a 1:20dilution of bovine antisera against the varioussamples gave complete suppression of plaqueformation in all dilutions tested.
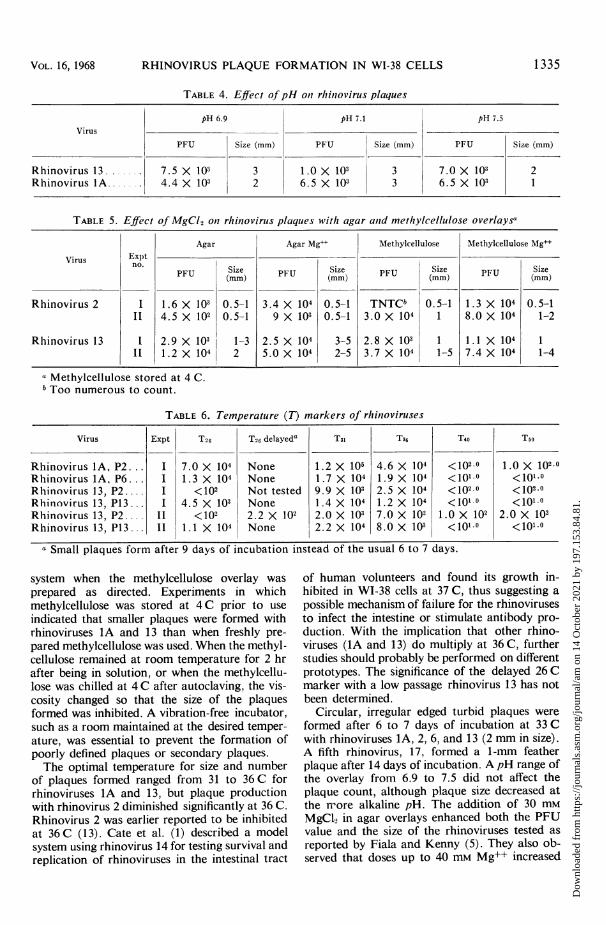
Effect ofpH on plaque formation. Rhinoviruses1A and 13 were not affected by the pH of theoverlay (Table 4). A decrease in plaque size butnot count was noted for both viruses at pH 7.5. Aslight decrease in plaque size occurred with rhino-virus 1A at pH 6.9.
Effect ofMg on plaque formation. Experimentswere conducted to determine if enhancement ofplaque size occurred when 30 mm MgC12 wasadded to either Difco agar or methylcelluloseoverlays (Table 5). Significantly higher PFU valueswere obtained when Mg++ was present in theagar overlay. The plaque size of rhinovirus 13was increased by 2 to 3 mm in the presence of 30mM MgC92. Slightly increased titers (two- tofourfold) were noted when Mg++ was present inthe methylcellulose overlay, but plaque size en-hancement did not occur. The titers of the tworhinoviruses were similar when methylcellulosewithout Mg+ and agar with Mg+ were com-pared.
Effect of tempertaure on plaque formation. Pre-liminary experiments with rhinovirus IA at twodifferent passage levels (WI-2 and WI-6) indicatedplaques formed equally well at 26, 31, and 36 Cwith few or no plaques being formed at 40 and50 C (Table 6). A high passage (WI-I 3) isolateof rhinovirus 13 had similar PFU values and sizesat 26, 31, and 36 C with no plaques observed at40 or 50 C. A low passage (WI-2) isolate of rhino-virus 13 formed delayed plaques at 26 C. Thevalues of the low passage isolate were equivalentat 31 and 36 C, whereas an additional 2 to 3 daysof incubation at 26 C was required for theplaques to become microscopically visible. Allrhinoviruses tested, except the low passage iso-late of rhinovirus 13 at 26 C, had a plaque size ofapproximately 2 mm when counted at 6 to 7 days.
DIscussIoNPlaque assays for five rhinoviruses were per-
formed with human embryonic diploid cells
1333VOL. 16, 1968
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
m o
n 14
Oct
ober
202
1 by
197
.153
.84.
81.

DOLAN ET AL.
FIG. 1. Rhinovirus IA plaque formation at different days. From left to right, bottles represent days 2, 3, 4, 6, 7,9, 10, 21.
FIG. 2. Rhinovirus 2 plaque formation at different days. From left to right, bottles represent days 2, 3, 4, 6, 7,9, 10, 21.
FIG. 3. Rhinovirus 13 plaque formation at different days. From left to right, bottles represent days 2, 3, 4, 6, 7,9, 10, 21.
(WI-38) and a simple overlay medium consistingof BME, methylcellulose, and fetal bovine serum.The critical factor for plaque production appearedto be the preparation of methylcellulose. Fiala
and Kenny (5) found that rhinovirus 2 produceda plaque in calf HeLa cells of only 0.5 mm withmethylcellulose, whereas we observed that 2-mmplaques were produced in the human diploid
1334 APPL. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
m o
n 14
Oct
ober
202
1 by
197
.153
.84.
81.
RHINOVIRUS PLAQUE FORMATION IN WI-38 CELLS
TABLE 4. Effect of pH on1 rhinovirus plaques
TABLE 5. Effect of MgCl2 otn rhinovirus plaques with agar anid methylcellulose overlaysa
Agar Agar Mgd' Methylcellulose Methylcellulose Mg++
Virus ExptVirusExpt'FUSize Size Size Size
PFU (mm) PU (mm) PU (mm) PU (m
Rhinovirus 2 I 1.6 X 102 0.5-1 3.4 X 104 0.5-1 TNTCb 0.5-1 1.3 X 104 0.5-1Il 4.5 X 102 0.5-1 9 X 103 0.5-1 3.0 X 104 1 8.0 X 104 1-2
Rhinovirus 13 1 2.9 X 103 1-3 2.5 X 104 3-5 2.8 X 103 1 1.1 X 104 1_ 1.2 X 104 2 5.0 X 104 2-5 3.7 X 104 1-5 7.4 X 104 1-4
a Methylcellulose stored at 4 C.Too numerous to count.
TABLE 6. Temperature (T) markers of rhin7oviruses
Virus Expt T26 T26 delayeda T31 T36 T40 T50
Rhinovirus lA, P2... I 7.0 X 104 None 1.2 X 105 4.6 X 104 <102.0 1.0 X 102.0Rhinovirus1A, P6.. . I 1.3 X 104 None 1.7 X 104 1.9 X 104 <1010 <101.0Rhinovirus 13, P2 .... I <102 Not tested 9.9 X 103 2.5 X 104 <102.0 <102.0Rhinovirus 13, P13 ... I 4.5 X 103 None 1.4 X 104 1.2 X 104 <1010 <1010.Rhinovirus 13, P2 .... II <102 2.2 X 102 2.0 X 103 7.0 X 102 1.0 X 102 2.0 X 102Rhinovirus 13, P13... II 1.1 X 104 None 2.2 X 104 8.0 X 102 <101.0 <101.0
a Small plaques form after 9 days of incubation instead of the usual 6 to 7 days.
system when the methylcellulose overlay wasprepared as directed. Experiments in whichmethylcellulose was stored at 4C prior to useindicated that smaller plaques were formed withrhinoviruses IA and 13 than when freshly pre-pared methylcellulose was used. When the methyl-cellulose remained at room temperature for 2 hrafter being in solution, or when the methylcellu-lose was chilled at 4 C after autoclaving, the vis-cosity changed so that the size of the plaquesformed was inhibited. A vibration-free incubator,such as a room maintained at the desired temper-ature, was essential to prevent the formation ofpoorly defined plaques or secondary plaques.The optimal temperature for size and number
of plaques formed ranged from 31 to 36 C forrhinoviruses IA and 13, but plaque productionwith rhinovirus 2 diminished significantly at 36 C.Rhinovirus 2 was earlier reported to be inhibitedat 36 C (13). Cate et al. (1) described a modelsystem using rhinovirus 14 for testing survival andreplication of rhinoviruses in the intestinal tract
of human volunteers and found its growth in-hibited in WI-38 cells at 37 C, thus suggesting apossible mechanism of failure for the rhinovirusesto infect the intestine or stimulate antibody pro-duction. With the implication that other rhino-viruses (1A and 13) do multiply at 36 C, furtherstudies should probably be performed on differentprototypes. The significance of the delayed 26 Cmarker with a low passage rhinovirus 13 has notbeen determined.
Circular, irregular edged turbid plaques wereformed after 6 to 7 days of incubation at 33 Cwith rhinoviruses 1A, 2, 6, and 13 (2 mm in size).A fifth rhinovirus, 17, formed a 1-mm featherplaque after 14 days of incubation. A pH range ofthe overlay from 6.9 to 7.5 did not affect theplaque count, although plaque size decreased atthe more alkaline pH. The addition of 30 mMMgCl, in agar overlays enhanced both the PFUvalue and the size of the rhinoviruses tested asreported by Fiala and Kenny (5). They also ob-served that doses up to 40 mm Mg++ increased
1 335VOL. 16, 1968
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
m o
n 14
Oct
ober
202
1 by
197
.153
.84.
81.

DOLAN ET AL.
rhinovirus 2 plaque size when using methyl-cellulose overlay and a sensitive human hetero-ploid cell line. In our studies with the humandiploid cell strain WI-38, the incorporation ofMg++ into the methylcellulose overlay did notenhance titers or plaque sizes significantly. Thissuggests that the Mg++ ion has its effect with theagar or agar-cell system as reported by Walfiset al. (17) with enteroviruses.
ACKNOWLEDGMENTThis investigation was supported by the Vaccine
Development Board of the National Institutes ofHealth under contract PH 43-62-487.
LITERATURE CITED1. Cate, T. R., R. G. Douglas, Jr., K. M. Johnson,
R. B. Couch, and V. Knight. 1967. Studies onthe inability of rhinovirus to survive and repli-cate in the intestinal tract of volunteers. Proc.Soc. Exptl. Biol. Med. 124:1290-1295.
2. Dulbecco, R. 1952. Production of plaques inmonolayer tissue cultures by single particlesof an animal virus. Proc. Natl. Acad. Sci. U.S.38:747-752.
3. Dulbecco, R., and M. Vogt. 1954. Plaque forma-tion and isolation of pure lines with polio-myelitis viruses. J. Exptl. Med. 99:167-182.
4. Fenters, J. D., S. S. Gillum, J. C. Holper, andG. S. Marquis. 1966. Serotypic relationshipsamong rhinoviruses. Am. J. Epidemiol. 84:10-20.
5. Fiala, M., and G. E. Kenny. 1966. Enhancementof rhinovirus plaque formation in humanheteroploid cell cultures by magnesium andcalcium. J. Bacteriol. 92:1710-1715.
6. Hayflick, L., and P. S. Moorhead. 1961. Theserial cultivation of human diploid cell strains.Exptl. Cell Res. 25:585-621.
7. Hotchin, J. E. 1955. Use of methylcellulose gel asa substitute for agar in tissue culture overlays.Nature 175:352.
8. Hozinski, V. I., V. B. Seibil, N. S. Pantelyeva,S. M. Mazurova, and E. A. Novikova. 1966.The rct4o and T50 markers and characteristicsof some variants of measles virus. Acta Virol.(Prague) 10:20-27.
9. Kapikian, A. Z. (Chairman), et al. 1967. Rhino-viruses: a numbering system. Nature 213:761-763.
10. Kisch, A. L., and K. M. Johnson. 1963. A plaqueassay for respiratory syncytial virus. Proc. Soc.Exptl. Biol. Med. 112:583-589.
11. Kisch, A. L., P. A. Webb, and K. M. Johnson.1964. Further properties of five newly recog-nized picomaviruses (rhinoviruses). Am. J.Hyg. 79:125-133.
12. Lwoff, A., and M. Lwoff. 1958. L' inhibition dudeveloppement du virus poliomyelitique a 390et le probleme du role de l'hyperthermie dansl'evolution des infections virales. Compt.Rend. 246:190-192.
13. Parsons, R., and D. A. J. Tyrrell. 1961. A plaquemethod for assaying some viruses isolatedfrom common colds. Nature 189:640-642.
14. Porterfield, J. S. 1962. Titration of some commoncold viruses (rhinoviruses) and their antisera bya plaque method. Nature 194:1044-1047.
15. Schulze, I. T., and R. W. Schlesinger. 1963.Plaque assay of dengue and other group Barthropod-borne viruses under methylcelluloseoverlay media. Virology 19:40-48.
16. Vogt, M., R. Dulbecco, and H. A. Wenner. 1957.Mutants of poliomyelitis viruses with reducedefficiency of plating in acid medium and reducedneuropathogenicity. Virology 4:141-155.
17. Wallis, C., J. L. Melnick, and M. Bianchi. 1962.Factors influencing enterovirus and reovirusgrowth and plaque formation. Texas Rept.Biol. Med. 20:693-702.
18. Webb, P. A., K. M. Johnson, and M. A. Mufson.1964. A description of two newly-recognizedrhinoviruses of human origin. Proc. Soc. Exptl.Biol. Med. 116:845-852.
1336 APPL. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
m o
n 14
Oct
ober
202
1 by
197
.153
.84.
81.





























