Embed Size (px)
Citation preview

Review ArticleKilling Me Softly Connotations to Unfolded ProteinResponse and Oxidative Stress in Alzheimerrsquos Disease
Beata Pajdk12 Elhbieta Kania1 and Arkadiusz Orzechowski12
1Electron Microscopy Platform Mossakowski Medical Research Centre Polish Academy of Sciences Pawinskiego 502-106 Warsaw Poland2Department of Physiological Sciences Faculty of Veterinary Medicine Warsaw University of Life Sciences (SGGW)Nowoursynowska 159 02-776 Warsaw Poland
Correspondence should be addressed to Arkadiusz Orzechowski orzechowski arkadiuszwppl
Received 16 October 2015 Revised 28 November 2015 Accepted 7 December 2015
Academic Editor Federica Rizzi
Copyright copy 2016 Beata Pająk et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
This review is focused on the possible causes of mitochondrial dysfunction in AD underlying molecular mechanisms of thismalfunction possible causes and known consequences of APP A120573 and hyperphosphorylated tau presence in mitochondria andthe contribution of altered lipid metabolism (nonsterol isoprenoids) to pathological processes leading to increased formationand accumulation of the aforementioned hallmarks of AD Abnormal protein folding and unfolded protein response seem tobe the outcomes of impaired glycosylation due to metabolic disturbances in geranylgeraniol intermediary metabolism Theorigin and consecutive fate of APP A120573 and tau are emphasized on intracellular trafficking apparently influenced by inaccurateposttranslational modifications We hypothesize that incorrect intracellular processing of APP determines protein translocation tomitochondria in AD Similarly without obvious reasons the passage of A120573 and tau to mitochondria is observed APP targetedto mitochondria blocks the activity of protein translocase complex resulting in poor import of proteins central to oxidativephosphorylation Besides APP A120573 and neurofibrillary tangles of tau directly or indirectly impair mitochondrial biochemistryand bioenergetics with concomitant generation of oxidativenitrosative stress Limited protective mechanisms are inadequate toprevent the free radical-mediated lesions Finally neuronal loss is observed in AD-affected brains typically by pathologic apoptosis
1 Introduction
Alzheimerrsquos disease (AD) is known for almost 120 years as aprogressive fatal human neurodegenerative disease featuredby decline in bothmemory and cognitive functions [1] Early-onset familial AD (FAD) which accounts for less than 5 ofcases is linked to mutations in APP gene on chromosome21 or genes encoding components of 120574-secretase (presenilin1 presenilin 2) resulting in increased A120573
4240ratio where
A12057342
is highly fibrillogenic [2ndash4] Sporadic or late-onsetAD (SAD) which is a major form (over 95 of cases) hasunknown etiology While one out of nine people aged 65 orolder has Alzheimerrsquos nearly one out of three people aged85 or older has the disease [5] The underlying molecularmechanisms that cause the formation of the hallmarks of FADand SAD namely amyloid-120573- (A120573-) containing plaques and
microtubule-associated protein tau-containing neurofibril-lary tangles (NFTs) are not yet fully clarifiedThe A120573 peptideis a cleavage product of amyloid precursor protein (APP) bysequential action of 120573- and 120574-secretases which release 39ndash43-amino-acid peptide from the C-terminal (cytoplasmic)end of transmembrane protein [6] The outcome of theend-stage intracellular lesions in FAD and SAD is loss ofneurons (brain atrophy) with most affected regions as frontalcortex hippocampus and amygdala [7] Severe injuries arevery selective and restricted to neurons as shown by themorphometric analyses of brain tissue slices obtained atautopsy from cases with diagnosis of AD faced up withcases with no clinical or pathological history of neurologicaldisease [8] Bulk of changes is characterized by significantreduction in mitochondria density accumulation of mtDNAand proteins in cytoplasm and in the vacuoles associated
Hindawi Publishing CorporationOxidative Medicine and Cellular LongevityVolume 2016 Article ID 1805304 17 pageshttpdxdoiorg10115520161805304
2 Oxidative Medicine and Cellular Longevity
with lipofuscin once involved in mitophagy [9] These obser-vations show increased mitochondrial degradation productseither by autophagy or by messed up proteolytic systemsMitochondrial abnormalities to milder extent were alsofound in other cell types (endothelium fibroblasts) obtainedfrom patients with AD [10 11] We also observed extensiveautophagy in cellular model of FAD and SAD [12]
For decades the hypothesis of AD (ldquoamyloid cascadehypothesisrdquo) of extracellular amyloid-120573 plaques and intracel-lular NFTs accumulations as clues in AD pathogenesis havebeen extensively examined with conflicting results In thelast decade however a new attractive hypothesis emergedfrom studies concerning mitochondria as key organelles formaintaining neuron functions and survival A growing bodyof evidence supports the idea that dysfunctional mitochon-dria cause development of synaptic abnormalities neuronaldegeneration and ultimately cell death as a consequenceof unbearable oxidative stress in AD [13ndash16] Numerous invitro and in vivo experiments substantiated the so-calledldquovicious cycle hypothesisrdquo pointing to the importance ofmitochondria in the pathogenesis of AD [17ndash28] Due totheir limited glycolytic capacity (lack of salvage pathway)neurons are highly dependent on mitochondrial function forenergy release and severely affected by the limited oxygenand glucose supply making them especially susceptible toenergy dyshomeostasis [29] Moreover mitochondria whichproduce almost entire energy in neurons have recently beenfound to be targeted by APP and A120573 [15 17 30ndash35] Thepresence of APP and A120573 in mitochondria has detrimentalconsequences as both constituents cause perturbations incellular energy homeostasis
2 Neurons Cells Extremely Susceptible toEnergy Dyshomeostasis
Excitability is a basic attribute of neurons (as well as otherexcitable cells) as it encompasses the primary task to receiveanalyze and dispatch electronic signals within the neuronalnetwork or to their cognate effectors This function isachieved by the generation of electric currents some of whichare of high frequencies These electric currents are evokedby the ion fluxes (Na+ K+ Ca2+ and Clminus) through channelslocated in the plasma membrane Any change in the concen-tration ofK+ orNa+ at the extra- or intracellular site of plasmamembrane respectively activates Na+K+-ATPase whichrestores the concentration gradient essential for excitabilityand also controls the cell volumeThe active transport againstthe concentration gradient is entirely dependent on ATPdelivered toNa+K+-ATPase and other pumps (Ca2+-ATPaseH+-ATPase) ATP is hydrolyzed leading to phosphorylationof the pump at a highly conserved aspartate residue andsubsequent release of ADP Energy generation and energyconsumption are tightly coupled to neuronal activity at thecellular level Na+K+-ATPase a major energy-consumingenzyme is well expressed in neurons rich in cytochrome119888 oxidase an important enzyme of the energy-generatingmachinery and glutamatergic receptors that are mediatorsof neuronal activity [36] Na+K+-ATPase enzyme consumesthe bulk of energy in the brain [37ndash39] Nervous cells are
highly enriched in mitochondria the main energy supplyingorganelle which provide ATP once they are sufficientlysupplied with oxygen Mitochondrial ATP is exchanged withcytosolic ADP through inner membrane adenine nucleotidetranslocase so the intracellular location of mitochondria iscrucial for availability of ATP and accelerated by cytosolicADP To meet energy requirements mitochondria moveregularly along the microtubular meshwork to the sites ofhigher ATP demand (high concentration of ADP) wherethey undergo the fusion process Mitochondrial ATP is alsoindispensable energy donor for dynamins (kinesin dynein)the proteins responsible for microtubule-associated axonaltransport of the secretory vesicles Obviously any substantialinterruption of the mitochondrial function distribution andfusion would affect the ATP delivery with resultant defects inneuronal activity
3 APP Processing and A120573 Formation
First widely expressedAPP is a transmembrane glycoproteinsynthesized on three different templates (APP695 APP751and APP770) which resulted from alternative splicing ofthe transcript [40 41] Second after APP is synthesized onpolysomes this protein undergoes N-glycosylation in the EROnce it is N-glycosylated the APP is then transported tothe Golgi apparatus Golgi apparatus is a second chief siteof APP posttranslational modifications including O- and N-glycosylations phosphorylations and sulphonations [42 43]Great deal of mature APP protein is stored in Golgi andtrans-Golgi network (TGN) while approximately 10ofAPPis unidirectionally (anterograde) transported by kinesins inTGN vesicles or in elongated tubular structures along micro-tubules in soma dendrites and axons [44 45] Third APPglycoprotein embedded to plasmamembrane is preferentiallycleaved in the nonamyloidogenic pathway alternatively itcould be internalized via endocytosis [46] Endosomic APPprotein aswell as its processed fragments can return to plasmamembrane can be proteolytically degraded in the lyso-some or can be transported from early endosome to TGNRetention of APP in the endoplasmic reticulumintermediatecompartment (ERIC) eliminated production of intracellularA12057340
but did not alter synthesis of fibrillogenic form (A12057342)
[47] Interestingly the production of intracellular A120573 fromwild-type APP695 appears to be a unique characteristic ofpostmitotic neurons since intracellular A120573 was not detectedin several nonneuronal cell lines [48]WhetherAPP retromer(transported from early endosome to TGN) is also cleavedvia amyloidogenic pathway is not clear due to conflictingobservations [49 50] In neurons APP695 is the major iso-form and could be subject to sequential proteolytic cleavageby 120573- and 120574-secretase to free A120573 The 120573-secretase (BACE1transmembrane aspartyl protease) initiates endoproteolyticcleavage giving rise to N-terminus of A120573 (120573-secretase cleavedAPP to 120573CTF as the intermediate) followed by 120574-secretase(membrane-embedded aspartyl protease complex consistingof presenilin PS) presenilin enhancer-2 (Pen-2) anteriorpharynx defective-1 (Aph-1) and nicastrin which reveals theC-terminus of A120573 [51] Given that two PS (PS1 and PS2) andAph-1 (Aph-1A and Aph-1B) variants exist the processing
Oxidative Medicine and Cellular Longevity 3
P3A
ICD
AIC
D120574-Secretase120574-Secretase
APP-CTF83
120572-Secretase
BACE1120573-Secretase
APPsAPP-120573 sAPP-120572
APP-CTF99
Amyloidogenic NonamyloidogenicA120573
A120573A12057340A
12057342
Figure 1 A diagram of amyloid precursor protein (APP) processing pathway The transmembrane protein APP (membrane indicated inblue) can be processed by two pathways the nonamyloidogenic 120572-secretase pathway and the amyloidogenic 120573-secretase pathway In thenonamyloidogenic pathway 120572-secretase cleaves in the middle of the 120573-amyloid (A120573) region (red) to release the soluble APP-fragment sAPP-120572 The APP C-terminal fragment 83 (APP-CTF83 120572CTF) is then cleaved by 120574-secretase to release the APP intracellular domain (AICD) andP3 fragment In the amyloidogenic pathway 120573-secretase cleaves APP to produce the soluble fragment sAPP-120573 APP-CTF99 (120573CTF) is thencleaved by 120574-secretase to produce A120573
40 A12057342 and AICD Adopted from [179]
of APP by four different human 120574-secretase complexes eachacting at more than one 120573CTF site (120576- 120577- and 120574-) leads toformation of several A120573 (A120573
37ndash43) with A12057340 and A12057342 beingpredominant species Finally three end products are formed(sAPP-120573 A120573 and amyloid precursor protein intracellulardomain AICD) The key neuron 120572-secretase (ADAM10)cleaves APP inside A120573 polypeptide chain to 120572CTF as theintermediate so after subsequent 120574-secretase action on120572CTFthree nonpathogenic fragments are formed (sAPP-120572 P3fragment and AICD) (Figure 1)
As aforementioned FAD is caused by mutations in APPand PSEN genes located on chromosomes 21 and 14 respec-tively but the incidence of AD is also higher in dominantlyinherited duplications of the APP locus in elderly individualswith Downrsquos syndrome (trisomy of the 21st chromosome)pointing to important role played by APP and A120573 in ADMutations in APP located near the 120573-secretase cleavage siteincrease production ofA120573 whereas those near the 120574-secretasecleavage site result in an increased ratio of A120573
42to A120573
40
[52] The 1205764 allele of apolipoprotein E is the major risk factorfor SAD Thus this particular APOE gene polymorphismincreases disease risk in a dose-dependentmanner and lowersthe age of onset as shown by Corder et al [53] One copyof APOE4 increases the risk of AD about fourfold (com-pared with the more common APOE1205763APOE31205763 genotype)whereas two copies of APOE4 increase the risk of AD about12-fold The mechanism by which the amino acid differencebetweenAPOE3 andAPOE4 increases the risk ofAD remainsto be established
Widespread occurrence ofAPP andA120573 in nervous systembrought about the assumption that both components mightplay physiological roles More than few possible conceptshave emerged some validated by experimental data TheAPP protein overexpression led to enhanced survival andgrowth of some cell types [54 55] Furthermore secreted
forms of APP (APPss sAPP120572 and sAPP120573) were antiapoptotic[56] and have a potent neuroprotective action in culturedrat hippocampal and septal neurons and in human corti-cal neurons [57] APPs
695and APPs
751protected neurons
against hypoglycemic damage and the neuroprotection wasabolished by antibodies to a specific region common toboth APPs
695and APPs
751 Thus APPss may normally play
excitoprotective and neuromodulatory roles AccordinglyAPP was shown to stimulate axon branching and the main-tenance and formation of synapses neuronal survival andneuritic outgrowth [58ndash60] APP protein is highly expressedin axons and interacts with extracellular matrix components[61ndash64] Similar to APP A120573 was demonstrated to play aphysiological role in synaptic plasticity as minute quantitiesof the peptide stimulated neurons and enhanced the releaseof neurotransmitter [65 66] Everything can changewhen thethings go awry
4 Perturbations in ER
In healthy cells including neurons ER is a fundamentalorganelle for protein quality control in the secretory pathwaywhich prevents protein aberrant folding and aggregation[67] A bulk of evidence shows the importance of ERin APP maturation and processing With regard to APPintracellular processing both secretases (120572- and 120573-) havebeen identified in the ER together with 120574-secretase whichis present in mitochondria-associated membrane (MAM)subcompartment [48 68] This distinctive intracellular lipid-raft-like structure is involved in cholesterol and phospholipidmetabolism Ca2+ metabolism and mitochondrial dynamicsand becomes markedly augmented in AD [69] MAM isresponsible for the communication between the ER and themitochondria with efficient transfer of Ca2+ from the ERto mitochondria supporting metabolic functions and cell
4 Oxidative Medicine and Cellular Longevity
ER membrane
6
ERAD
Proteasome26S
Ca2+
Ca2+
IP3R
VDAC
MCU
IMMOMM
ATP
GRP
75
MFN
1
MFN
2M
FN2
MFN
2
Sig1R
UPR
7
Mitochondria
Nucleus CHOP ATF6
XBP1ATF4
Gene expression
3
Transcriptionfactors
2
IRE1120572 PERK ATF6
1 GRP78
Chaperones
ER Misfolded proteins
Autophagosome
5
4
ERGolgitraffic
Golgi cisternae
Antioxidant responseApoptosisERAD and autophagyER chaperonesER and Golgi biosynthesis
Figure 2 ER stress triggered by misfolded proteins in several neurodegenerative diseases Abnormal conformations of the proteins APPA120573 and tau are implicated in the pathogenesis of AD Alterations in the function of ER chaperones and UPR-related components ERADERGolgi trafficking and ER-to-mitochondria Ca2+ transfer have been suggested as underlying mechanisms of ER stress triggered by thesedisease-associated proteins These proteins can accumulate and aggregate at the ER and their stable interaction with ER chaperones such asGRP78BiP and PDI may trap ER chaperones altering protein folding with concomitant ER stress In addition these proteins can lead to theoxidative modification of the active site of PDIs by nitrosylation leading to their enzymatic inactivation Furthermore some of these proteinsalter the activity of the UPR stress sensors (IRE1120572 PERK and ATF6) as well as the activitylevels of downstream signaling mediators andtranscription factors including cleaved ATF6 ATF4 and spliced XBP1 As a result genes implicated in autophagy and ERAD antioxidantresponse ER chaperones and organellersquos biosynthesis are upregulatedMoreover these proteins block the exit of vesicles from the ER and alterthe trafficking between ER andGolgi of properly folded proteinsThe cellular responses controlled byUPR transcription factors including themodulation of autophagy mediated degradation of protein aggregates become compromised Disease-related proteins can also interact withERAD components precluding the translocation of ERAD substrates from the ER to the cytosol leading to the accumulation of abnormallyfolded proteins at the ER Finally Ca2+ released from the ERmainly through the IP
3R and its transfer tomitochondria can be impaired in the
presence of disease-related proteins leading to mitochondrial Ca2+ overload and activation of apoptotic cell death pathways AD Alzheimerrsquosdisease ATF6 activating transcription factor 6 ATF4 activating transcription factor 4 ER endoplasmic reticulum ERAD endoplasmic-reticulum-associated protein degradation IP
3R inositol triphosphate receptor IRE1120572 inositol-requiring enzyme 1 alpha PERK protein
kinase R- (PKR-) like ER kinase UPR unfolded protein response XBP1 Xbox binding protein 1 Adopted from [67]
viability [70] The molecular bridges between ER inositol145-triphosphate receptor (IP
3R) and the voltage-dependent
anion channels in the outer mitochondrial membrane arebrought together through the cytosolic chaperone glucose-regulated protein 75 (GRP75) (Figure 2) Additionally thedynamin-relatedGTPasemitofusin 2 (Mfn2) proteins locatedon the ER intermingle with Mfn1 or Mfn2 on mitochondria
to tighten the connection The distance between ER andmitochondria controlled by the phosphofurin acidic clustersorting protein 2 (PACS-2) of ER and the dynamin-relatedGTPase protein 1 (Drp1) is crucial for cell survival as eithertoo long (lack of Ca2+ flux) or too short distance (Ca2+ over-load) might lead to apoptosis [71] Alternatively impairedmitochondrial bioenergetics with reduced cellular ATP
Oxidative Medicine and Cellular Longevity 5
levels stimulate autophagyThe molecular mechanism of ER-mediated autophagy is accurately regulated by Beclin 1 as wellas ER membrane bound protein Bax inhibitor 1 (BI-1) Bothproteins are capable of promoting autophagy through IP
3R-
dependent mechanism [72]
5 APP Processing and ER Stress Response
The nature of APP processing is determined by the com-position of membrane with cholesterol rich lipid rafts asthe site of amyloidogenic cleavage [73] Consequently onemight expect that fate of APP is at least partly establishedby the representation of lipid rafts and possible access to 120573-versus120572-secretase Irrespective of the type of cleavage there isone underestimated biochemical step in ER that might bringabout damaging upshot It is apparently the effectiveness ofER situated N-glycosylation which makes the APP moleculesuitably folded If misfoldedmalfolded protein(s) accumu-late in the ER complex cascade of reactions known as theunfolded protein response (UPR) is triggered with the so-called endoplasmic reticulum stress response (ERS) Today itis widely accepted that during UPR the ER sensors proteinkinase R- (PKR-) like ER kinase (PERK) activating proteinkinase 6 (ATF6) and inositol-requiring enzyme 1 alpha(IRE1120572) are freed fromGRP78BiP protein repression whichhereafter become activated Without going into details ofindividual sensor action a number of reactions occur at boththe genomic and cytoplasmic level with selective degradationof mRNAs encoding protein(s) with abnormal folding andinhibition of protein translation except for genes importantfor UPR redox homeostasis energy metabolism and pro-tein folding [67] On the one hand PERK phosphorylateseukaryotic initiation factor 2 alpha (eIF2120572) to stop entrance ofmethionyl-tRNA to the ribosome on the other hand it allowstranslation of activating transcription factor 4 (ATF4) geneIRE1120572 sets off alternative splicing of Xbox binding protein 1(XBP1) transcript leading to activation of the transcriptionfactor liable to stimulate ERGolgi biogenesis and formationof proteins involved in endoplasmic-reticulum-associatedprotein degradation known as ERAD (Erdj4 p58IPK EDEMRAMP-4 PDI-P5 and HEDJ for details see [67]) FinallyATF6 is activated in the Golgi complex through proteolyticcleavage and translocates to nucleus where it cooperates withXBP1 in upregulation of chaperones and ERAD-related genes[43] In principle UPR is activated to restore ER homeostasisand stop the accumulation of aberrantly formed protein(s)but if the strength of ER stress is unbearable (meaning thatit cannot be compensated by UPR) there is a path to activateapoptosis Among other routes the most central role isplayed by the major proapoptotic transcription factor CEBPhomologous protein CHOPgrowth arrest and DNA damageinduced gene 153 GADD153 It downregulates the antiapop-totic protein Bcl-2 and upregulates proapoptotic Bax and Bak[74] CHOPGADD153 leads to excessive production of reac-tive oxygen species (ROS) within ER subsequent depletionof reduced glutathione (GSH) and Ca2+ flux from the ER tocytoplasm through the IP
3R [75] One hitherto unresolved
issue is the physiologic importance of APP and GRP78BiPinteraction disclosed in coprecipitation study carried out by
Yamamoto et al [76] Bulk ofAPP associatedwithGRP78BiPwas immature protein Given that GRP78BiP expressionlevels declined in samples of brain tissue obtained fromFAD patients as demonstrated by Katayama et al [77] fewdistinct scenarios are possible First binding to GRP78BiPsuggests ER accumulation of immature APP Second APPinteraction with GRP78BiP is a noticeable sign of UPRwhich is further validated by downregulation of GRP78BiPin FAD patients Finally retention of APP in ER is mostlikely a result of amyloidogenic APP processing and as such itprobably occurs in MAM the cholesterol enriched domainsER-mitochondria crossing point is therefore of particularinterest in deciphering the links between the ER placed APPprocessing UPR and resulting cellular responses such asautophagy apoptosis and inflammatory reaction observed inAD (Figure 2)
6 Pathology of Tau Protein
Microtubule-associated protein tau controls assembly andprevents microtubules from severing Microtubular networkis fundamental component of cytoskeleton essential forintracellular transport of secretory vesicles and organelles(ie mitochondria) Glycogen synthase kinase 3 beta- (GSK-3120573-) targeted hyperphosphorylation of tau causes this proteinto dissociate frommicrotubules Consequently microtubulesbecome fragmented and microtubule-dependent transportsystem fails Furthermore hyperphosphorylated tau (P-tau)is prone to form oligomers and toxic filaments known asNFTs or tauopathy [78] A variety of tau conformers werereported to exist pointing to different tauopathies capa-ble of self-propagation [79 80] Interestingly intracellulartau inclusions define AD as clinical symptoms of diseaseobserved when tauopathy is abundant together with intra-cellular A120573 deposits in neocortex [51 81 82] People withabundant A120573 plaques but no or only a few neurofibrillarylesions do not have AD Clinicopathological correlationstudies have been crucial to generate hypotheses about thepathophysiology of the disease by establishing the fact thatthere is a continuum between ldquonormalrdquo aging and ADdementia and that the amyloid plaque buildup occurs primar-ily before the onset of cognitive deficits while neurofibrillarytangles neuron loss and particularly synaptic loss parallel theprogression of cognitive decline [83] Thus misfolded pro-teins and descendant toxic filaments with a number of inter-mediates are critical for manifestation of AD as fibrillogenicAPP processing is not enough for onset of disease Althoughthe molecular mechanism of tauopathy is not deciphered infull recent reports suggest ER stress as the starting point[84 85] This idea is substantiated by the elevated levels ofERS and UPR markers together with P-tau and GSK-3120573 inbrains affected by AD [86] Therefore on the one hand theincidence of UPR is strongly correlated with the presence ofNFTs on the other hand aggregation of P-tau induces ERSwith resultant UPR Some lines of evidence confirmed UPRactivation near the beginning of NFTs formation and pointto the functional link between malformed tau protein andUPRThe in vitro experiments with phosphatase 2A inhibitoror phosphorylation activator demonstrated enhanced P-tau
6 Oxidative Medicine and Cellular Longevity
formation in neurons in concert with the increased levelsof PERK eIF2120572 and XBP1 transcript apparent markers ofUPR [87] Furthermore GRP78BiP was found to encouragetau phosphorylation through facilitated substrate capture byGSK-3120573 [88] GSK-3120573 seems to play dual role first this kinaseprotects neurons from apoptosis as P-tau accumulation isstrong molecular signal to trigger UPR with subsequentautophagy Second UPR raises GSK-3120573 activity throughlysosomal degradation of inactive GSK-3120573 (P-Ser9-GSK-3120573)To sum up ERS and UPR are important molecular machinesused to prevent cell viability turned on by tauopathy
7 Oxidative Stress in ER
Sacs and tubes of ER delineate the compartment where thenewly synthesized proteins undergo maturation to nativestate Native state indicates properly folded fully functionalprotein Important reactions essential for protein folding ofunbranched polypeptide chains include amino acids oxida-tion and glycosylation As a result redox homeostasis inER is shifted to oxidative state so as to promote disulfidebond formation between adjacent cysteines Oxidation ofsulfhydryl groups required to make disulfide bonds is con-trolled by ER oxidase 1120572 (ERO1120572) Next disulfide bondscould be subject to posttranslational modification disulfideexchange by protein disulfide isomerase (PDI) PDI is ableto correct mispaired thiol residues by catalyzing the breakageand formation of correct disulfide bonds These enzymes arefundamental for protein folding Oxidation allows twistingof proteins which is followed by N-glycosylation andor C-and O-mannosylation Protein N-glycosylation in eukary-otes is a complex process divided into several steps Firstthere is a ldquocall forrdquo carrier lipids (polyisoprenyl phosphatessuch as dolichyl phosphates) the membrane lipids knownto function as glycosyl transporters In mammalian cellsthe limiting substrate for dolichol biosynthesis is geranyl-geraniol (GGOH) of mevalonate pathway Dolichols thelongest aliphatic molecules synthesized in animal cellshave 18ndash21 120572-isoprene saturated units (C90ndash105) criticalfor their recognition by the enzymes (glycosyltransferases)that glucosylate dolichyl phosphates [89] Once dolichylmonophosphates (Dol-P) are formed in the ER membranethe precursor oligosaccharide donor (GLc
3Man9GLCNAc
2-
P-P-dolichol) for protein N-glycosylation can be synthesizedon the lumenal leaflet of ER First three sugar intermediatesare produced (Man-P-Dol Glc-P-Dol GlcNAc-P-P-Dol andMan5GlcNAc-P-P-Dol) on the cytoplasmic leaflet of the
ER Next enzyme flippases mediate transbilayer movementof the aforementioned intermediates to lumenal side ofER where conversion to Glc
3Man9GlcNAc
2-P-P-Dol could
be completed Glc3Man9GlcNAc
2-P-P-Dol is also used for
biosynthesis of glycosylphosphatidylinositol (GPI) anchorsTaken together lipid-mediated glycosylation plays a vital
role in the appropriate protein folding and intracellulartranslocation of N-linked glycoproteins [90] Likewise it isimportant for protein O- and C-mannosylation and GPIanchorage Moreover Dol-P availability in the ER is the rate-limiting factor in the production of glycolipid intermediatesand N-glycosylation
8 ER Stress and Apoptosis
It was shown that during ERS the ER resident proapop-totic cysteine protease known as caspase (caspase-12 in ratcaspase-4 in humans) is activated through cleavage As aresult caspase cascade is started via caspase-9 that in turnstimulates effector caspase-3 [91 92] The central role playedby ER in programmed cell death is achieved by PERK branchwhere ATF4 induces the expression of CHOPGADD153which represses antiapoptotic Bcl-2 family proteins andsimultaneously shuffles ER membrane Bax and Bak proteinsinto outer mitochondrial membrane Consequently poresare formed to leak the components of apoptosome frommitochondrial intermembrane space [93] Another impor-tant mechanism of ERS-induced apoptosis is led by Ca2+-dependent ERO1120572-IP
3R pathway where ERO1120572 collaborates
with IP3R in Ca2+ efflux from ER to mitochondria via MAM
[75 94] Accordingly Ca2+ influx facilitates cytochrome 119888release frommitochondria besides cytochrome 119888 can bind toER IP
3R and the complex amplifies the apoptotic signal in a
feedforward manner [95] Last but not least ERS-associatedapoptotic programme is set off by c-Jun N-terminal kinase(JNK) as the effect of IRE1120572 complexed with TNF-receptor-associated factor 2 (TRAF2) activation of apoptosis-signal-regulating kinase 1 (ASK1) [96 97]
What does ERS drive to induce apoptotic death inneurons Actually many reports indicate that APP and A120573 aswell as hyperphosphorylated tau have been shown to blockmitochondrial transport which results in impaired energystorage and oxidative stress [98ndash101] Indeed accumulationof APP A120573 and NFTs in mitochondria led to reducedactivity of some enzymes involved in substrate oxidation(tricarboxylic acid cycle) electron transport chain (ETC)and ATP synthase as well as severely diminishing import ofnuclear-encoded proteins [26 27 102ndash104] One may ask ifthere is any additional link between ER and mitochondriaother than MAM which could account for apoptotic signalThough not directly ER significantly contributes to oxidativestress in mitochondria of AD-affected subjects
9 Abnormal APP Processing and TraffickingCulminate in ER Pathology of AD
From the morphological point of view as neurons arehighly specialized cells soma dendrites and neurites areconsiderably distinct structures Proteins needed by thesecompartments are delivered via microtubules once proteinshave suitable sorting signals (ie APP trafficking from ERto plasma membrane is associated with several posttrans-lational modifications with oxidation and N-glycosylation)Additionally the Golgi apparatus follows ER in subsequentAPP adjustment (O- and N-glycosylation phosphorylationand sulphonation) [42] Any inaccurate alteration of the APPmolecule is potentially hazardous as protein final destiny ismissed causing its retention in ER or trans-Golgi network(TGN) In addition other unusual settings for APP arepossible as this large protein has few signal sequences hiddenwhen APP is correctly folded In the cells transfected withAPP this protein enters coat protein complex I (COPI)
Oxidative Medicine and Cellular Longevity 7
vesicles and undergoes retrograde transport from cis endof the Golgi complex back to the ER [76] Such responsecauses accumulation of APP in the tubulocisternal ER systemtogether with aberrant intracellular translocation of theprotein Interestingly the issue whether APP is subject toretrograde transport with successive fibrillogenic processingbecause of UPR and its interaction with GRP78BiP is notclear as the observations are inconsistent [77] Nonethelessaccumulation of misfoldedmalfolded proteins in the ERsuggests disorganized process of posttranslational change Asanticipated the accretion of proteins of anomalous patternsignals ERS and UPR followed by increased vulnerability toapoptotic cell death Prior to decay however APP proteinlevels in the ER lessen by dint of cleavage with 120573- and120574-secretase [105 106] Products of this cleavage (sAPP120573A120573 and AICD fragment) all appreciably influence neuronalsurvival most likely through nonnative form of the APPsubstrate
10 APP A120573 and NFTs MarkMitochondria as Targeted in AD
Mitochondrial import of A12057340
and A12057342
peptides throughthe translocase TOM complex was blocked by preincubationof isolated mitochondria with antibodies raised againstTOM proteins (TOM20 TOM40 and TOM70) [107]Neither VDAC inhibition with antagonist antibodies norinhibition of mitochondrial permeability transition pores(MPTP) or fall of mitochondrial membrane potential(MMP) affected uptake of A120573 [17] With regard to APPelegant study performed by Anandatheerthavarada andhis colleagues [30] revealed that C-terminal truncatedAPP (lacking A120573) targets mitochondria in cholinergicGABAergic dopaminergic and glutamatergic neurons ofAD brain by clogging up mitochondrial protein translocasecomplex TOM40TIM22 Authors propose that occlusionof translocase is followed by blunted import of nuclear-encoded proteins vitally important for energy homeostasisMitochondrial APP protein transmembrane orientationindicates NH
2-terminal inside in contact with translocase
whereas COOH-terminal is facing cytoplasmic side NH2-
terminus has mitochondrial signal sequence Astonishinglymitochondrial APP molecules were nonglycosylated givingrise to speculation that protein molecules that arrivedat mitochondria have not achieved molecular maturity[31] The accumulation of nonglycosylated APP species inmitochondrial import channels of AD brain was directlyrelated to decreased mitochondrial functions as validatedby the decline in cytochrome 119888 oxidase activity (complexIV) and elevated levels of H
2O2 Furthermore in AD
brain the mitochondrial accumulation of nonglycosylatedAPP went along with a corresponding reduction in plasmamembrane-associatedAPP It suggests that ADbrain hasAPPprocessing and trafficking severely affected by incompleteN-glycosylation ensuing ER protein accumulation andexposure of the cryptic mitochondrial targeting signalfor assisting chaperone proteins Several lines of evidenceindicate that not fully formed proteins are phosphorylatedand bind to the cytosolic proteins required for movement
from ER to mitochondria [108ndash110] Moreover ERS andUPR in AD seem to be incompetent and inefficient inelimination of malfolded proteins In addition to APPalso A120573 was frequently reported to occupy mitochondriaalthough its origin and mechanism of mitochondriatargeting mostly remain unexplored (except for involvementof protein translocase TOM complex) The possibilityof mitochondrial A120573 generation has to be ruled out asmembrane orientation of arrested APP does not allow accessto 120574-secretase (120574-secretase activity in mitochondria wasdetected by independent study) [111] Probably A120573 speciesis derived from APP prior to its translocation (ER) soA120573 may be transported to mitochondria independentlyof APP Collectively observations showing mitochondrialpresence of APP A120573 and tau in aberrant configuration pointtoward the anomaly of protein folding at the level of ER andGolgi apparatus Nonglycosylated molecule of APP suggestsdefective transfer of sugar core from dolichyl phosphate(s)and further modifications such as O- and N-glycosylationsOne may admit that the lack of glycosyl residue bringsabout pathologic processing and trafficking of APP and itsfragments In point of fact dolichol derivatives mixture ofpolyprenols (acyclic isoprenoid alcohols) known as Ropren(Solagran Limited Melbourne Australia) commerciallyused to treat liver diseases were tested in the treatment ofAD in two separate trials conducted in 2005 and 2006 withpromising results [httpwwwasxcomauasxpdf20071119pdf315x5nh4hm8wv7pdf httpwwwasxcomauasxpdf20070221pdf3111ztwcbqzkk9pdf] Further studies areurgently needed to test how important the glycosylationprocess is in the pathogenesis of AD Dolichols areobtained from geranylgeraniol (GGOH) and the latter is anintermediate of mevalonate pathway GGOH is a commonsubstrate for dolichol and ubiquinone synthesis but it isalso necessary for protein prenylation As both GGOHand farnesol (FOH) are engaged in protein prenylationmore concern should be laid on the importance of thesecompounds in AD pathogenesis Observations demonstrat-ing mitochondrial relocalization of other proteins withoutposttranslational modification seemingly point to increasedmitochondrial targeting of immature molecules resultingin mitochondrial dysfunction and acceleration of diseaseprogression [34 112ndash114]
11 Oxidative Stress in Mitochondria ofAD Brains
Normal physiological functions of APP are thought to beinvolved in the stabilizing contact points between synapsesand maintaining mitochondrial functions [60 115] Mito-chondrial dysfunction was often observed regardless of theexperimental model used to study AD [8 29 116ndash119]It includes defects in oxidative phosphorylation decreasedATP decreased membrane potential increased productionof ROSRNS and perturbation in mitochondrial fusion andfission [15 30 31 115 120ndash122] Hyperphosphorylated tau wasalso reported to impair mitochondrial functions [123] Usingproteomic approach the strongest defects of the respiratorycapacity were observed mainly at complexes I IV and ATP
8 Oxidative Medicine and Cellular Longevity
synthase (complex V) at both protein and activity level [124]While APP A120573 and hyperphosphorylated tau are potentinhibitors of mitochondrial import of nuclear-encoded pro-teins apparently the pathology of each leads to metabolicharm in different way In freshly isolated mitochondriafrom AD brains the APP inhibited mitochondrial importof cytochrome 119888 oxidase (COX) subunits IV and Vb [31]Dysfunction of COX increases ROS production (incompletereduction of oxygen molecules) reduces energy stores anddisturbs energy metabolism Accordingly in AD patientsdeficiency of COX was found in brains and platelets [119125 126] Similar to APP A120573 was found in mitochondriaof transgenic mice and cellular and human AD models [1719 33 103 127ndash130] At present it is not clear whether theobservedmitochondrial toxicity is due toAPP orA120573 orNFTsaccumulation Anyway some regularity is observed withrespect to most affected components of respiratory chainNADH-ubiquinone oxidoreductase (complex I) activity isreduced to the utmost by hyperphosphorylated tau whereasdecreased activity of cytochrome 119888 oxidase (complex IV)that resulted in mitochondrial dysfunction was observedduring A120573 and APP accumulation [14 15 31 32 34]Concomitantly rise in the activity of antioxidant enzymesmanganese superoxide dismutase (Mn-SOD) and catalase(CAT) was demonstrated in response to elevated levelsof free radicals including superoxide anion radical (O
2
∙minus)hydroperoxyl radical (HO
2
∙) hydroxyl radical (OH∙) andnitric oxide radical (NO∙) [15] Consistent with observationsof chronic respiratory chain dysfunction and mitochondrialoxidative stress there are reports showing their contributionto tau pathology in AD [131] In any case free radicals thatoverride antioxidant defense react with a wide variety oforganic components causing lipid peroxidation to advancedlipid oxidation end products (ALE) cross-linking of proteinsnitrosylation of proteins and mutations in DNA Mitochon-drial circular DNA (mtDNA) of sim16 kbp is devoid of repairsystems meaning the buildup of lesions There are 37 geneslocated in mtDNA with those encoding protein subunits ofcomplex I (7) complex II (1) and complex IV (3) but notcomplex III Interestingly hallmarks of AD (APP A120573 andNFTs)mostly affectmembers of electron transfer chain (com-plexes I and IV) which rely exclusively on mitochondriallypredetermined subunits Maybe it is not simple coincidencebut the effect of inhibited import to mitochondria of nuclear-encoded subunits of complexes I and IV gives explanationfor toxicity of APP or A120573 or NFTs in mitochondria Thereare additional findings in mitochondria affected by AD suchas lower activity of pyruvate dehydrogenase (PDH) andoxoglutarate dehydrogenase (OGDH) [104 132] Inhibitionof OGDH the enzyme of tricarboxylic acid cycle minimizesthe NADH pool and electron number needed for ETCand mitochondrial membrane potential (Δ120595
119898) to create and
maintain proton gradient obligatory for ATP synthesisThereare also lines of evidence for direct inhibitory action of solubleoligomeric A120573 species on ABAD (A120573-binding alcohol dehy-drogenase) and internal membrane cyclophilin D (CypD)resulting in increased mitochondrial membrane permeabil-ity (MPTP) potentiated ROS production synaptic lossdiminished activity of mitochondrial respiration and finally
cell death [129 132ndash134] CypD knockout prevents mito-chondrial and neuronal perturbations and improves mito-chondrial function in Alzheimerrsquos disease mouse model [128135] It has to be emphasized that oxidativenitrosative stressaffects the fusion and fission process ofmitochondria Fusionthat is speedup by small GTPases mitofusins (Mfn1Mfn2)improves efficiency of mitochondrial respiration and ATPproduction Mitochondrial dynamics are severely imbal-anced in AD cases in favour of fission through elevatedexpression of the fission protein DLP1 (dynamin-like protein1) associatedwith nitrosative stress stirred up byA120573 [136 137]Collectively these observations indicate that toxic intracel-lular A120573
42-43 oligomers differ in action from extracellularaggregates found in amyloid plaques of AD brains [138](Figure 3)
12 Proteostasis in Mitochondria
Mitochondrial protein turnover grants the well-organizedreplacement of nonfunctional proteins into operational oneThe first task is attained through proteolytic degradation ofinner membrane and matrix proteins with local mitochon-drial proteases [139] or outer membrane proteins throughubiquitin-proteasome system [140] The second task is metby intramitochondrial protein synthesis but as almost 1500different nuclear-encoded proteins have to be importedto complete mitochondrial proteome a matter of capableimport is fundamental for the function of the organelleIn extreme cases of cellular injuries observed in neurode-generative diseases damaged mitochondria with extendedloss of the electrochemical potential are selectively removedby autophagy known as mitophagy [112] APP and A120573accumulation has also something to do with altered mito-chondrial dynamics as fission takes advantage of fusion withresultant dysfunction of mitochondria and neurons [137]AD neurons demonstrated selective mechanisms of prote-olytic clearance of oxidatively and nitrosatively modifiedproteins in mitochondria Insulin degrading enzyme (IDE)prevents formation of toxic insoluble fibrils from A120573 asit cleaves A120573 prior to aggregation [141ndash143] A novel zinc-metallopeptidase Presequence Protease (PreP) a member ofpitrilysin oligopeptidase family degrades either intramito-chondrially stored A120573
40or A120573
42proteinThis protease seems
to be highly sensitive to oxidative stress as disulfide bridgeformed between two proximal cysteine residues blocks itscatalytic activity [144ndash149] Another mitochondrial serineprotease HtrA2Omi occupies intermembrane space whereit can cleave APP locked up in protein translocase complex[121] Even though it is well established that HtrA2Omiis released to the cytosol to amplify apoptosis through thedegradation of antiapoptotic proteins and caspase activation[150ndash152] it is also implicated in proteolytic deletion ofmalfoldedAPP in ER [151] Fromknockout studies onmice itis obvious that HtrA2Omi plays a significant shielding roleas mice deficient in this protease exhibit neurodegenerativephenotype with weight loss and premature death [153]Taken together understanding the mechanisms of clearanceof the unwanted proteins including APP A120573 and tau isvital in an attempt to get rid of them from mitochondria
Oxidative Medicine and Cellular Longevity 9
VDAC
ANT
A120573
A120573
A120573
A120573
A120573
A120573
A120573A120573
A120573
A120573
A120573 CypD
CypD ProApCyt c Ca2+
IMM
OMM
IMSΔΨ
+
+
+
minus
minus
minus
ABAD
ROS
ROS
TIM
TOM
Proteinimport
NAD
NAD
NAD
NAD
H+
H+
H+
H+
H+
ADP
Mn-SOD
CuZn-SOD
mtDNA
V
Matrix
UCP
O2∙minus
O2∙minus
IV
III
II
I
O2
O2
DLP1Fis1
Mfn1Mfn2
Opa1
mHsp70
NADH
NADH
NADH
TCAcycle
PyruvateAcetyl-CoA
120572KGDH
120572-KG
PDH
Isocitrate
CitrateOAA
Malate
Fumarate
Succinyl-CoASuccinate
GDPGTP
FAD
FAOH
ATP
Figure 3 Amyloid-120573-related mitochondrial impairment Mitochondria were found to be the target for amyloid-120573 (A120573) which interactswith several proteins leading to mitochondrial dysfunction Indeed A120573 was found in the outer mitochondrial membrane (OMM) andinner mitochondrial membrane (IMM) as well as in the matrix The interaction of A120573 with the OMM affects the transport of nuclear-encoded mitochondrial proteins such as subunits of the electron transport chain complex IV into the organelle via the translocase of theouter membrane (TOM) import machinery Moreover A120573 disturbs the activity of several enzymes such as pyruvate dehydrogenase (PDH)and oxoglutarate dehydrogenase (OGDH) decreasing NADH reduction and the electron transport chain enzyme complex IV reducing theamount of hydrogen that is translocated from the matrix to the intermembrane space (IMS) thus impairing the mitochondrial membranepotential (MMP) Taken together these events cause abnormal mitochondrial electron activities leading to decreased complex V activityand so to a drop in ATP levels in addition to increasing reactive oxygen species (ROS) generation Moreover ROS induce peroxidationof several mitochondrial macromolecules such as mitochondrial DNA (mtDNA) and mitochondrial lipids contributing to mitochondrialimpairment in themitochondrial matrixThe complex of A120573 bound to binding alcohol dehydrogenase (ABAD) impairs the binding of NAD+to ABAD changes mitochondrial membrane permeability and reduces activities of respiratory enzymes inducing further ROS productionand leading to mitochondrial failure A120573 binding also activates Fis1 (fission protein) and promotes increased mitochondrial fragmentationthis increased mitochondrial fragmentation produces defective mitochondria that ultimately damage neurons Furthermore A120573 binding tocyclophilin D (CypD) enhances the protein translocation to the inner membrane favouring the opening of the mitochondrial permeabilitytransition pore formed by the adenine nucleotide translocator (ANT) and voltage-dependent anion channels (VDACs) Cyt 119888 cytochrome 119888DLP1 dynamin-like protein 1 PDH pyruvate dehydrogenase ProAp proapoptotic factors SOD superoxide dismutase TCA tricarboxylicacid TIM translocase of the inner membrane Adopted from [14]
Some hopes are associated with the application of at presentindefinite modulators of proteolytic activity HtrA2Omi isfor example activated by PTEN-induced putative kinase 1(PINK1) upon phosphorylation at Ser142 residue [154] Alter-natively accent has to be put on mitophagy of dysfunctionalmitochondria and mitochondriogenesis
13 Aging versus AD
Themost commonneurodegenerative disorder is representedby Alzheimerrsquos disease characterized by declining memoryreduced cognitive capacity and progressive dementia whichare often fatal to elderly individuals above 65 years of age
10 Oxidative Medicine and Cellular Longevity
It is ranked as the fourth leading cause of death in modernsocieties where average life span increased greatly in the lasttwo decades As 95 of AD cases are diagnosed in olderpeople one might think that a causal relationship existsbetween aging and the onset of disease Certainly a numberof similarities between getting old and being affected withAD could be listed Historically the free radical theory ofaging by Harman [155] suggested aging as ldquoside effectrdquo ofreactive oxygen species formed in mitochondrial respiratorychain Apparently free radicals commonly generated byincomplete reduction of oxygen molecule at complexes Iand III of mitochondrial ETC are capable of damagingDNA RNA and proteins They impair energy storage andlead to operational failure of mitochondria with progres-sive decline of cell viability Almost identical conditionsaccompany AD and are an explicit step in pathogenesisof disease As mtDNA is deprived of repair mechanismsROS-induced DNA strand breaks tend to accumulate withage or AD Thus mtDNA is a vulnerable target for ROSbut the reverse the ROS generation due to the mutatedmtDNA is not convincingly confirmed [156] Moreover theevidence that mutated mtDNA accelerates the progress ofaging is also questioned based on the results from studycarried out on transgenic mice model [157] Although someauthors show inconsistency between the free radical theoryand observations cumulative evaluation of the scientificreports points to antioxidant defense systems as importantfactors in protection from premature aging [158] Othermitochondrial components important for their function arehampered with age adenine nucleotide translocase (ANT)nitric oxide synthase (NOS) and carnitine acyltransferase(CT) [159ndash161] Actually NOS activity is elevated in AD asreported from study performed on cellular model of disease[15] We could not find any information about CT activityin AD whereas ANT activity is noticeably inhibited byA120573 or hyperphosphorylated tau and this effect is reversedby mersalyl a reversible alkylating agent of thiol groups[162] Mitochondrial dysfunction observed in transgenicmice models of AD and aging demonstrates higher activ-ity of genes controlling energy metabolism and apoptosisTaken together physiological aging and AD are associatedwith broad-spectrum dysfunction of mitochondria but thefoundations of mitochondrial decline are dissimilar Morediscrepancies between physiological aging and AD werefound with respect to ERS and UPR which play a significantrole in cellular proteostasis Dolichol was selected as an agingmarker because the progressive increase in dolichol levelwas observed in aging brain [163] In contrast ubiquinoneconcentration which is also synthesized from geranylgeranioldiminishes with agingwhereas cholesterol and dolichyl phos-phate concentrations remain unaltered In AD decreasedlevels of dolichol were observed and increased levels ofubiquinone and dolichyl phosphate without any changesin brain cholesterol AD cannot be regarded as a resultof premature aging The drop in dolichol and augmenteddolichyl phosphate concentration points toward disturbedglycosylation in ER of diseased brain while the increase inubiquinone suggests efforts to protect the brain from oxida-tive stress induced by lipid peroxidation [164 165]
14 Targeting ER Stress in AD Therapy
As ERS is a recognized factor in AD drugs that interfere withERS would theoretically have great therapeutic potentialThere are several compounds grouped in classes thatinteract directly with components of the ERS (salubrinalBiP inducer X (BIX) salicylamide analogs flavonoidsguanabenz and STF083010) chemical chaperons (4-phenyl-butyric acid (PBA) tauroursodeoxycholic acid (TUDCA)and trimethylamine oxide (TMAO)) chemicals that inhibitprotein degradation (Eeyarestatin MG132 and Bortezomib)compounds with antioxidant activity (Edaravone diben-zoylmethane derivatives and N-acetyl cysteine (NAC)) anddrugs controlling calcium signaling (dantrolene and carba-zole derivatives) [166] They may act by inducing transienttranslation arrest upregulation of chaperone proteins andaugmented degradation of ER-associated misfolded proteinsFundamental approach in the development of new therapyis the selection of appropriate molecular targets In ER stresssignaling the aim is to alter the expression of ER stress-associated molecules that can rescue cells from the toxiceffect of ERS Recent efforts in establishing new promisingdrugs against AD are pointing to chemical chaperones suchas PBA TUDCA or trimethylamine oxide (TMAO) Thesesubstances improve protein folding and alleviate nativeprotein conformation [166] It was shown on mouse modelsof AD that PBA TUDCA and TMAO stop A120573 accumulationand avoid the loss of dendritic spines [167] Some observa-tions even demonstrated improved memory and cognitivefunctions [168] associated with improved cell survival [169]Salubrinal ((2E)-3-phenyl-N-[222-trichloro-1-[[(8-quinoli-nylamino)thioxomethyl]amino]ethyl]-2-propenamide Sal)selectively inhibits growth arrest and DNA damage inducedgene 34- (GADD34-) phosphatase complex (GADD34associates with protein phosphatase 1 (PP1)) and promotes invitro dephosphorylation of the alpha subunit of eIF-2120572 andIRE1120572ASK1JNK signaling pathway being protective againstERS even induced by tunicamycin Tm [170] In a great deal ofexperiments testing Sal in cultured cells and animal modelsof AD this substance increased the viability of neuronal cellsand A120573 toxicity [171 172] BIX (2-(34-dihydroxyphenyl)-2-oxoethyl ester thiocyanic acid) preferentially inducedBiP mRNA in an ATF6-dependent manner leading toreduced Tm-induced death of neuronal cells [173] AlsoDBM derivative 14ndash26 (221015840-dimethoxydibenzoylmethane)was found to be neuroprotective for SH-SY5Y and PC-12cells by decreasing expression of BiP and CHOP [174]Dantrolene a ryanodine receptor antagonist that inhibitsabnormal calcium release from the ER inhibited expressionof both phosphorylated PERK and eIF2120572 It also reducedCHOP expression and attenuated thapsigargin-inducedapoptosis in PC-12 cells [175] Neuroprotective effects similarto dantrolene were observed for ([9-(3-cyanobenzyl)-14-dimethylcarbazole]) This substance suppressed increases inintracellular Ca2+ in PC-12 cells treated with thapsigarginand reduced levels of BiP and CHOP [176]
Aforementioned compounds were chosen from othersas the most potent ER stress inhibitors and persuasivelyprotective to neuronal cells 18 of 42 different compounds
Oxidative Medicine and Cellular Longevity 11
were exploited in in vivo and in vitro models of centralnervous system disorders with in fact improved cell or tissueviability [166] Thus the brain is the most frequently investi-gated organ in the context of ERS From these experimentsit becomes clear that CHOP functions as proapoptotic factorThe roles of other specific ER stress molecules as moleculartargets for pharmacological intervention are less clear andvary depending on cell type and context
There are few underestimated modulations in APP pro-cessing that shed more light on current dogma of AD patho-genesis The modulation of mevalonate pathway and choles-terol synthesis were reported to stimulate nonamyloidogenicpathway of APP processing [177] Additionally cholesterolderivative 27-hydroxycholesterol (27-OHC) was shown toinduce ER stress which attenuated leptin-dependent viabilityby activating CHOP in SH-SY5Y neuroblastoma cells [178]Irrespective of a number of compounds examined in AD thecall for new drugs modulating ER stress with healing effect isstill waiting to be revealed
Abbreviations
APOE Apolipoprotein EAPPs Secreted form of APPCHOPGADD153 CEBP homologous protein
growth arrest and DNA damageinduced gene 153
Drp1 Dynamin-1-like proteinERAD Endoplasmic-reticulum-
associated protein degradationERO1120572 ER oxidase 1120572ERS Endoplasmic reticulum stressFOH FarnesolGGOH GeranylgeraniolGRP75 Glucose-regulated protein 75GRP78BiP 78 kDa glucose-regulated proteinGSK-3120573 Synthase kinase 3 betaIP3R Inositol triphosphate receptor
IRE1120572 Inositol-requiring enzyme 1 alphaMAM Mitochondria-associated
membraneMAP LC3 Microtubule-associated protein
light chain 3Mfn2 GTPase mitofusin 2MMP Mitochondrial membrane
potentialMPTP Mitochondrial permeability
transition poreNFT Neurofibrillary tanglesPACS-2 Phosphofurin acidic cluster
sorting protein 2PDI Protein disulfide isomerasePERK Protein kinase R- (PKR-) like
ER kinaseRNS Reactive nitrogen speciesROS Reactive oxygen speciesRyR Ryanodine receptorTOM Transporter outer membraneTIM Transporter inner membrane
UPR Unfolded protein responseVDAC Voltage-dependent anion channelXBP-1 Xbox binding protein 1
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
Support for this work was provided by Grant no UMO-201103BNZ70238 from the National Science Centre inPoland
References
[1] R L Nussbaum andC E Ellis ldquoAlzheimerrsquos disease and Parkin-sonrsquos diseaserdquoTheNewEngland Journal ofMedicine vol 348 no14 pp 1356ndash1364 2003
[2] A L Brunkan and A M Goate ldquoPresenilin function and 120574-secretase activityrdquoThe Journal of Neurochemistry vol 93 no 4pp 769ndash792 2005
[3] V Crentsil ldquoThe pharmacogenomics of Alzheimerrsquos diseaserdquoAgeing Research Reviews vol 3 no 2 pp 153ndash169 2004
[4] D L Price and S S Sisodia ldquoMutant genes in familialAlzheimerrsquos disease and transgenic modelsrdquo Annual Review ofNeuroscience vol 21 pp 479ndash505 1998
[5] D Harman ldquoAlzheimerrsquos disease pathogenesis role of agingrdquoAnnals of the New York Academy of Sciences vol 1067 no 1 pp454ndash460 2006
[6] D J Selkoe ldquoTranslating cell biology into therapeutic advancesin Alzheimerrsquos diseaserdquo Nature vol 399 pp A23ndashA31 1999
[7] H Braak and E Braak ldquoNeuropathological stageing of Alz-heimer-related changesrdquo Acta Neuropathologica vol 82 no 4pp 239ndash259 1991
[8] K Hirai G Aliev A Nunomara et al ldquoMitochondrial abnor-malities inAlzheimerrsquos diseaserdquoTheJournal ofNeuroscience vol21 pp 3017ndash3023 2001
[9] U T Brunk C B Jones and R S Sohal ldquoA novel hypothesisof lipofuscinogenesis and cellular aging based on interac-tions between oxidative stress and autophagocytosisrdquoMutationResearchDNAging vol 275 no 3ndash6 pp 395ndash403 1992
[10] P A Stewart K Hayakawa M-A Akers and H V Vinters ldquoAmorphometric study of the blood-brain barrier in Alzheimerrsquosdiseaserdquo Laboratory Investigation vol 67 no 6 pp 734ndash7421992
[11] J P Blass and G E Gibson ldquoThe role of oxidative abnor-malities in the pathophysiology of Alzheimerrsquos diseaserdquo RevueNeurologique vol 147 no 6-7 pp 513ndash525 1991
[12] B Pająk E Kania and A Orzechowski ldquoNucleofection of ratpheochromocytoma PC-12 cells with human mutated beta-amyloid precursor protein gene (APP-sw) leads to reducedreduced viability autophagy-like process and increased expres-sion and secretion of beta amyloidrdquo BioMed Research Interna-tional vol 2015 Article ID 746092 10 pages 2015
[13] J P Blass ldquoCerebrometabolic abnormalities in AlzheimerrsquosdiseaserdquoNeurological Research vol 25 no 6 pp 556ndash566 2003
12 Oxidative Medicine and Cellular Longevity
[14] A Eckert K Schmitt and J Gotz ldquoMitochondrial dysfunc-tionmdashthe beginning of the end in Alzheimerrsquos disease Sep-arate and synergistic modes of tau and amyloid-120573 toxicityrdquoAlzheimerrsquos Research ampTherapy vol 3 article 15 pp 1ndash11 2011
[15] U Keil A Bonert C A Marques et al ldquoAmyloid 120573-inducedchanges in nitric oxide production and mitochondrial activitylead to apoptosisrdquoThe Journal of Biological Chemistry vol 279no 48 pp 50310ndash50320 2004
[16] A M Swomley and D A Butterfield ldquoOxidative stress inAlzheimer disease and mild cognitive impairment evidencefrom human data provided by redox proteomicsrdquo Archives ofToxicology vol 89 no 10 pp 1669ndash1680 2015
[17] C Caspersen N Wang J Yao et al ldquoMitochondrial A120573 apotential focal point for neuronal metabolic dysfunction inAlzheimerrsquos diseaserdquo The FASEB Journal vol 19 no 14 pp2040ndash2041 2005
[18] G E Gibson K-F R Sheu and J P Blass ldquoAbnormalities ofmitochondrial enzymes inAlzheimer diseaserdquo Journal of NeuralTransmission vol 105 no 8-9 pp 855ndash870 1998
[19] A Eckert S Hauptmann I Scherping et al ldquoSoluble beta-amyloid leads to mitochondrial defects in amyloid precursorprotein and tau transgenic micerdquo Neurodegenerative Diseasesvol 5 no 3-4 pp 157ndash159 2008
[20] A Eckert K L Schulz V Rhein and J Gotz ldquoConvergenceof amyloid-120573 and tau pathologies on mitochondria in vivordquoMolecular Neurobiology vol 41 no 2-3 pp 107ndash114 2010
[21] S Hauptmann I Scherping S Drose et al ldquoMitochondrial dys-function an early event in Alzheimer pathology accumulateswith age in AD transgenic micerdquoNeurobiology of Aging vol 30no 10 pp 1574ndash1586 2009
[22] M Manczak B S Park Y Jung and P H Reddy ldquoDifferentialexpression of oxidative phosphorylation genes in patients withAlzheimerrsquos disease implications for early mitochondrial dys-function and oxidative damagerdquo NeuroMolecular Medicine vol5 no 2 pp 147ndash162 2004
[23] M P Mattson M Gleichmann and A Cheng ldquoMitochondriain neuroplasticity and neurological disordersrdquo Neuron vol 60no 5 pp 748ndash766 2008
[24] P I Moreira M S Santos and C R Oliveira ldquoAlzheimerrsquosdisease a lesson frommitochondrial dysfunctionrdquoAntioxidantsamp Redox Signaling vol 9 no 10 pp 1621ndash1630 2007
[25] P H Reddy ldquoMitochondrial dysfunction in aging and Alz-heimerrsquos disease strategies to protect neuronsrdquo Antioxidants ampRedox Signaling vol 9 no 10 pp 1647ndash1658 2007
[26] V Rhein G Baysang S Rao et al ldquoAmyloid-beta leads toimpaired cellular respiration energy production andmitochon-drial electron chain complex activities in humanneuroblastomacellsrdquo Cellular and Molecular Neurobiology vol 29 no 6-7 pp1063ndash1071 2009
[27] V Rhein X Song A Wiesner et al ldquoAmyloid-120573 and tausynergistically impair the oxidative phosphorylation system intriple transgenic Alzheimerrsquos disease micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 47 pp 20057ndash20062 2009
[28] R X Santos S C Correia X Wang et al ldquoAlzheimerrsquos diseasediverse aspects ofmitochondrial malfunctioningrdquo InternationalJournal of Clinical and Experimental Pathology vol 3 no 6 pp570ndash581 2010
[29] N R Sims ldquoEnergy metabolism oxidative stress and neuronaldegeneration in Alzheimerrsquos diseaserdquoNeurodegeneration vol 5no 4 pp 435ndash440 1996
[30] H K Anandatheerthavarada G Biswas M-A Robin and NG Avadhani ldquoMitochondrial targeting and a novel transmem-brane arrest of Alzheimerrsquos amyloid precursor protein impairsmitochondrial function in neuronal cellsrdquo The Journal of CellBiology vol 161 no 1 pp 41ndash54 2003
[31] L Devi B M Prabhu D F Galati N G Avadhani and HK Anandatheerthavarada ldquoAccumulation of amyloid precur-sor protein in the mitochondrial import channels of humanAlzheimerrsquos disease brain is associated with mitochondrialdysfunctionrdquo The Journal of Neuroscience vol 26 no 35 pp9057ndash9068 2006
[32] L Devi and H K Anandatheerthavarada ldquoMitochondrialtrafficking of APP and alpha synuclein relevance to mito-chondrial dysfunction in Alzheimerrsquos and Parkinsonrsquos diseasesrdquoBiochimica et Biophysica Acta vol 1802 no 1 pp 11ndash19 2010
[33] M Manczak T S Anekonda E Henson B S Park J Quinnand P H Reddy ldquoMitochondria are a direct site of A120573 accumu-lation in Alzheimerrsquos disease neurons implications for free rad-ical generation and oxidative damage in disease progressionrdquoHuman Molecular Genetics vol 15 no 9 pp 1437ndash1449 2006
[34] P F Pavlov C H Petersen E Glaser and M AnkarcronaldquoMitochondrial accumulation of APP and A120573 significancefor Alzheimer disease pathogenesisrdquo Journal of Cellular andMolecular Medicine vol 13 no 10 pp 4137ndash4145 2009
[35] C Schmidt E Lepsverdize S L Chi et al ldquoAmyloid precursorprotein and amyloid 120573-peptide bind to ATP synthase andregulate its activity at the surface of neural cellsrdquo MolecularPsychiatry vol 13 no 10 pp 953ndash969 2008
[36] K Johar A Priya and M T T Wong-Riley ldquoRegulation ofNa+K+-ATPase by nuclear respiratory factor 1 implication inthe tight coupling of neuronal activity energy generation andenergy consumptionrdquo The Journal of Biological Chemistry vol287 no 48 pp 40381ndash40390 2012
[37] J Astrup P M Sorensen and H R Sorensen ldquoOxygen andglucose consumption related to Na+-K+ transport in caninebrainrdquo Stroke vol 12 no 6 pp 726ndash730 1981
[38] M Erecinska and I A Silver ldquoATP and brain functionrdquo Journalof Cerebral Blood Flow and Metabolism vol 9 no 1 pp 2ndash191989
[39] MMata D J Fink H Gainer et al ldquoActivity-dependent energymetabolism in rat posterior pituitary primarily reflects sodiumpump activityrdquo Journal of Neurochemistry vol 34 no 1 pp 213ndash215 1980
[40] J Kang H-G Lemaire A Unterbeck et al ldquoThe precursor ofAlzheimerrsquos disease amyloid A4 protein resembles a cell-surfacereceptorrdquo Nature vol 325 no 6106 pp 733ndash736 1987
[41] C L Masters and D J Selkoe ldquoBiochemistry of amyloid 120573-protein and amyloid deposits in Alzheimer diseaserdquoCold SpringHarbour Perspectives in Medicine vol 2 no 6 Article IDa006262 2012
[42] A Lai S S Sisodia and I S Trowbridge ldquoCharacterization ofsorting signals in the beta-amyloid precursor protein cytoplas-mic domainrdquo The Journal of Biological Chemistry vol 270 no8 pp 3565ndash3573 1995
[43] A I Placido C M F Pereira A I Duarte et al ldquoThe role ofendoplasmic reticulum in amyloid precursor protein processingand trafficking implications for Alzheimerrsquos diseaserdquo Biochim-ica et Biophysica ActamdashMolecular Basis of Disease vol 1842 no9 pp 1444ndash1453 2014
Oxidative Medicine and Cellular Longevity 13
[44] C Kaether P Skehel and C G Dotti ldquoAxonal membraneproteins are transported in distinct carriers a two-color videomicroscopy study in cultured hippocampal neuronsrdquoMolecularBiology of the Cell vol 11 no 4 pp 1213ndash1224 2000
[45] A Kamal A Almenar-Queralt J F LeBlanc E A Robertsand L S B Goldstein ldquoKinesin-mediated axonal transportof a membrane compartment containing 120573-secretase andpresenilin-1 requires APPrdquo Nature vol 414 no 6864 pp 643ndash648 2001
[46] R G Perez S Soriano J D Hayes et al ldquoMutagenesis identi-fies new signals for 120573-amyloid precursor protein endocytosisturnover and the generation of secreted fragments includingA12057342rdquo The Journal of Biological Chemistry vol 274 no 27 pp18851ndash18856 1999
[47] D G Cook M S Forman J C Sung et al ldquoAlzheimerrsquos A120573(1ndash42) is generated in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo Nature Medicine vol 3 no 9 pp1021ndash1023 1997
[48] A S C Chyung B D Greenberg D G Cook R W Domsand V M-Y Lee ldquoNovel 120573-secretase cleavage of 120573-amyloidprecursor protein in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo The Journal of Cell Biology vol138 no 3 pp 671ndash680 1997
[49] RW-Y Choy Z Cheng and R Schekman ldquoAmyloid precursorprotein (APP) traffics from the cell surface via endosomesfor amyloid 120573 (A120573) production in the trans-Golgi networkrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 30 pp E2077ndashE2082 2012
[50] S A Small and S Gandy ldquoSorting through the cell biologyof Alzheimerrsquos disease intracellular pathways to pathogenesisrdquoNeuron vol 52 no 1 pp 15ndash31 2006
[51] M Goedert ldquoAlzheimerrsquos and Parkinsonrsquos diseases the prionconcept in relation to assembled A120573 tau and 120572-synucleinrdquoScience vol 349 no 6248 2015
[52] I Benilova E Karran and B De Strooper ldquoThe toxic A120573oligomer and Alzheimerrsquos disease an emperor in need ofclothesrdquo Nature Neuroscience vol 15 no 3 pp 349ndash357 2012
[53] E H Corder A M Saunders W J Strittmatter et al ldquoGenedose of apolipoprotein E type 4 allele and the risk of Alzheimerrsquosdisease in late onset familiesrdquo Science vol 261 no 5123 pp 921ndash923 1993
[54] S Brunholz S Sisodia A Lorenzo C Deyts S Kins and GMorfini ldquoAxonal transport of APP and the spatial regulationof APP cleavage and function in neuronal cellsrdquo ExperimentalBrain Research vol 217 no 3-4 pp 353ndash364 2012
[55] G Thinakaran and E H Koo ldquoAmyloid precursor proteintrafficking processing and functionrdquo The Journal of BiologicalChemistry vol 283 no 44 pp 29615ndash29619 2008
[56] T Saitoh M Sundsmo J-M Roch et al ldquoSecreted formof amyloid beta protein precursor is involved in the growthregulation of fibroblastsrdquo Cell vol 58 no 4 pp 615ndash622 1989
[57] M PMattson B Cheng A R Culwell F S Esch I Lieberburgand R E Rydel ldquoEvidence for excitoprotective and intraneu-ronal calcium-regulating roles for secreted forms of the 120573-amyloid precursor proteinrdquo Neuron vol 10 no 2 pp 243ndash2541993
[58] K J Lee C E H Moussa Y Lee et al ldquoBeta amyloid-independent role of amyloid precursor protein in generationandmaintenance of dendritic spinesrdquoNeuroscience vol 169 no1 pp 344ndash356 2010
[59] H Meziane J-C Dodart C Mathis et al ldquoMemory-enhancingeffects of secreted forms of the 120573-amyloid precursor proteinin normal and amnestic micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no21 pp 12683ndash12688 1998
[60] P R Turner K OrsquoConnor W P Tate and W C AbrahamldquoRoles of amyloid precursor protein and its fragments inregulating neural activity plasticity and memoryrdquo Progress inNeurobiology vol 70 no 1 pp 1ndash32 2003
[61] D Beher L Hesse C L Masters and GMulthaup ldquoRegulationof Amyloid Protein Precursor (APP) binding to collagen andmapping of the binding sites on APP and collagen type Irdquo TheJournal of Biological Chemistry vol 271 no 3 pp 1613ndash16201996
[62] S L Sabo A F Ikin J D Buxbaum and P Greengard ldquoTheamyloid precursor protein and its regulatory protein FE65 ingrowth cones and synapses in vitro and in vivordquo The Journal ofNeuroscience vol 23 no 13 pp 5407ndash5415 2003
[63] D H Small V Nurcombe G Reed et al ldquoA heparin-bindingdomain in the amyloid protein precursor of Alzheimerrsquos diseaseis involved in the regulation of neurite outgrowthrdquoThe Journalof Neuroscience vol 14 no 4 pp 2117ndash2127 1994
[64] T G Williamson S S Mok A Henry et al ldquoSecreted glypicanbinds to the amyloid precursor protein of Alzheimerrsquos disease(APP) and inhibits APP-induced neurite outgrowthrdquoThe Jour-nal of Biological Chemistry vol 271 no 49 pp 31215ndash312211996
[65] T Ondrejcak I Klyubin N-W Hu A E Barry W K Cullenand M J Rowan ldquoAlzheimerrsquos disease amyloid 120573-protein andsynaptic functionrdquo NeuroMolecular Medicine vol 12 no 1 pp13ndash26 2010
[66] D Puzzo L Privitera and A Palmeri ldquoHormetic effect ofamyloid-beta peptide in synaptic plasticity and memoryrdquo Neu-robiology of Aging vol 33 no 7 pp 1484e15ndash1484e24 2012
[67] C M F Pereira ldquoCrosstalk between endoplasmic reticulumstress and protein misfolding in neurodegenerative diseasesrdquoISRN Cell Biology vol 2013 Article ID 256404 22 pages 2013
[68] R-W Shin T C Saido M Maeda and T Kitamoto ldquoNovel 120572-secretase cleavage ofAlzheimerrsquos amyloid120573 precursor protein inthe endoplasmic reticulum of COS7 cellsrdquoNeuroscience Lettersvol 376 no 1 pp 14ndash19 2005
[69] E A Schon and E Area-Gomez ldquoMitochondria-associatedER membranes in Alzheimer diseaserdquo Molecular and CellularNeuroscience vol 55 pp 26ndash36 2013
[70] R Rizzuto P Pinton W Carrington et al ldquoClose contacts withthe endoplasmic reticulum as determinants of mitochondrialCa2+ responsesrdquo Science vol 280 no 5370 pp 1763ndash1766 1998
[71] S E Logue P Cleary S Saveljeva and A Samali ldquoNewdirections in ER-stress induced cell deathrdquoApoptosis vol 18 no5 pp 537ndash546 2013
[72] J P Decuypere KWelkenhuyzen T Luyten et al ldquoIns(145)P3receptor-mediated BI-1 Ca2+ signaling and autophagy induc-tion are interrelatedrdquo Autophagy vol 7 pp 1472ndash1489 2011
[73] K S Vetrivel and G Thinakaran ldquoMembrane rafts in Alz-heimerrsquos disease beta-amyloid productionrdquo Biochimica et Bio-physica Acta vol 1801 no 8 pp 860ndash867 2010
[74] H Urra E Dufey F Lisbona D Rojas-Rivera and C HetzldquoWhen ER stress reaches a dead endrdquo Biochimica et BiophysicaActa (BBA) Molecular Cell Research vol 1833 no 12 pp 3507ndash3517 2013
14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Oxidative Medicine and Cellular Longevity
with lipofuscin once involved in mitophagy [9] These obser-vations show increased mitochondrial degradation productseither by autophagy or by messed up proteolytic systemsMitochondrial abnormalities to milder extent were alsofound in other cell types (endothelium fibroblasts) obtainedfrom patients with AD [10 11] We also observed extensiveautophagy in cellular model of FAD and SAD [12]
For decades the hypothesis of AD (ldquoamyloid cascadehypothesisrdquo) of extracellular amyloid-120573 plaques and intracel-lular NFTs accumulations as clues in AD pathogenesis havebeen extensively examined with conflicting results In thelast decade however a new attractive hypothesis emergedfrom studies concerning mitochondria as key organelles formaintaining neuron functions and survival A growing bodyof evidence supports the idea that dysfunctional mitochon-dria cause development of synaptic abnormalities neuronaldegeneration and ultimately cell death as a consequenceof unbearable oxidative stress in AD [13ndash16] Numerous invitro and in vivo experiments substantiated the so-calledldquovicious cycle hypothesisrdquo pointing to the importance ofmitochondria in the pathogenesis of AD [17ndash28] Due totheir limited glycolytic capacity (lack of salvage pathway)neurons are highly dependent on mitochondrial function forenergy release and severely affected by the limited oxygenand glucose supply making them especially susceptible toenergy dyshomeostasis [29] Moreover mitochondria whichproduce almost entire energy in neurons have recently beenfound to be targeted by APP and A120573 [15 17 30ndash35] Thepresence of APP and A120573 in mitochondria has detrimentalconsequences as both constituents cause perturbations incellular energy homeostasis
2 Neurons Cells Extremely Susceptible toEnergy Dyshomeostasis
Excitability is a basic attribute of neurons (as well as otherexcitable cells) as it encompasses the primary task to receiveanalyze and dispatch electronic signals within the neuronalnetwork or to their cognate effectors This function isachieved by the generation of electric currents some of whichare of high frequencies These electric currents are evokedby the ion fluxes (Na+ K+ Ca2+ and Clminus) through channelslocated in the plasma membrane Any change in the concen-tration ofK+ orNa+ at the extra- or intracellular site of plasmamembrane respectively activates Na+K+-ATPase whichrestores the concentration gradient essential for excitabilityand also controls the cell volumeThe active transport againstthe concentration gradient is entirely dependent on ATPdelivered toNa+K+-ATPase and other pumps (Ca2+-ATPaseH+-ATPase) ATP is hydrolyzed leading to phosphorylationof the pump at a highly conserved aspartate residue andsubsequent release of ADP Energy generation and energyconsumption are tightly coupled to neuronal activity at thecellular level Na+K+-ATPase a major energy-consumingenzyme is well expressed in neurons rich in cytochrome119888 oxidase an important enzyme of the energy-generatingmachinery and glutamatergic receptors that are mediatorsof neuronal activity [36] Na+K+-ATPase enzyme consumesthe bulk of energy in the brain [37ndash39] Nervous cells are
highly enriched in mitochondria the main energy supplyingorganelle which provide ATP once they are sufficientlysupplied with oxygen Mitochondrial ATP is exchanged withcytosolic ADP through inner membrane adenine nucleotidetranslocase so the intracellular location of mitochondria iscrucial for availability of ATP and accelerated by cytosolicADP To meet energy requirements mitochondria moveregularly along the microtubular meshwork to the sites ofhigher ATP demand (high concentration of ADP) wherethey undergo the fusion process Mitochondrial ATP is alsoindispensable energy donor for dynamins (kinesin dynein)the proteins responsible for microtubule-associated axonaltransport of the secretory vesicles Obviously any substantialinterruption of the mitochondrial function distribution andfusion would affect the ATP delivery with resultant defects inneuronal activity
3 APP Processing and A120573 Formation
First widely expressedAPP is a transmembrane glycoproteinsynthesized on three different templates (APP695 APP751and APP770) which resulted from alternative splicing ofthe transcript [40 41] Second after APP is synthesized onpolysomes this protein undergoes N-glycosylation in the EROnce it is N-glycosylated the APP is then transported tothe Golgi apparatus Golgi apparatus is a second chief siteof APP posttranslational modifications including O- and N-glycosylations phosphorylations and sulphonations [42 43]Great deal of mature APP protein is stored in Golgi andtrans-Golgi network (TGN) while approximately 10ofAPPis unidirectionally (anterograde) transported by kinesins inTGN vesicles or in elongated tubular structures along micro-tubules in soma dendrites and axons [44 45] Third APPglycoprotein embedded to plasmamembrane is preferentiallycleaved in the nonamyloidogenic pathway alternatively itcould be internalized via endocytosis [46] Endosomic APPprotein aswell as its processed fragments can return to plasmamembrane can be proteolytically degraded in the lyso-some or can be transported from early endosome to TGNRetention of APP in the endoplasmic reticulumintermediatecompartment (ERIC) eliminated production of intracellularA12057340
but did not alter synthesis of fibrillogenic form (A12057342)
[47] Interestingly the production of intracellular A120573 fromwild-type APP695 appears to be a unique characteristic ofpostmitotic neurons since intracellular A120573 was not detectedin several nonneuronal cell lines [48]WhetherAPP retromer(transported from early endosome to TGN) is also cleavedvia amyloidogenic pathway is not clear due to conflictingobservations [49 50] In neurons APP695 is the major iso-form and could be subject to sequential proteolytic cleavageby 120573- and 120574-secretase to free A120573 The 120573-secretase (BACE1transmembrane aspartyl protease) initiates endoproteolyticcleavage giving rise to N-terminus of A120573 (120573-secretase cleavedAPP to 120573CTF as the intermediate) followed by 120574-secretase(membrane-embedded aspartyl protease complex consistingof presenilin PS) presenilin enhancer-2 (Pen-2) anteriorpharynx defective-1 (Aph-1) and nicastrin which reveals theC-terminus of A120573 [51] Given that two PS (PS1 and PS2) andAph-1 (Aph-1A and Aph-1B) variants exist the processing
Oxidative Medicine and Cellular Longevity 3
P3A
ICD
AIC
D120574-Secretase120574-Secretase
APP-CTF83
120572-Secretase
BACE1120573-Secretase
APPsAPP-120573 sAPP-120572
APP-CTF99
Amyloidogenic NonamyloidogenicA120573
A120573A12057340A
12057342
Figure 1 A diagram of amyloid precursor protein (APP) processing pathway The transmembrane protein APP (membrane indicated inblue) can be processed by two pathways the nonamyloidogenic 120572-secretase pathway and the amyloidogenic 120573-secretase pathway In thenonamyloidogenic pathway 120572-secretase cleaves in the middle of the 120573-amyloid (A120573) region (red) to release the soluble APP-fragment sAPP-120572 The APP C-terminal fragment 83 (APP-CTF83 120572CTF) is then cleaved by 120574-secretase to release the APP intracellular domain (AICD) andP3 fragment In the amyloidogenic pathway 120573-secretase cleaves APP to produce the soluble fragment sAPP-120573 APP-CTF99 (120573CTF) is thencleaved by 120574-secretase to produce A120573
40 A12057342 and AICD Adopted from [179]
of APP by four different human 120574-secretase complexes eachacting at more than one 120573CTF site (120576- 120577- and 120574-) leads toformation of several A120573 (A120573
37ndash43) with A12057340 and A12057342 beingpredominant species Finally three end products are formed(sAPP-120573 A120573 and amyloid precursor protein intracellulardomain AICD) The key neuron 120572-secretase (ADAM10)cleaves APP inside A120573 polypeptide chain to 120572CTF as theintermediate so after subsequent 120574-secretase action on120572CTFthree nonpathogenic fragments are formed (sAPP-120572 P3fragment and AICD) (Figure 1)
As aforementioned FAD is caused by mutations in APPand PSEN genes located on chromosomes 21 and 14 respec-tively but the incidence of AD is also higher in dominantlyinherited duplications of the APP locus in elderly individualswith Downrsquos syndrome (trisomy of the 21st chromosome)pointing to important role played by APP and A120573 in ADMutations in APP located near the 120573-secretase cleavage siteincrease production ofA120573 whereas those near the 120574-secretasecleavage site result in an increased ratio of A120573
42to A120573
40
[52] The 1205764 allele of apolipoprotein E is the major risk factorfor SAD Thus this particular APOE gene polymorphismincreases disease risk in a dose-dependentmanner and lowersthe age of onset as shown by Corder et al [53] One copyof APOE4 increases the risk of AD about fourfold (com-pared with the more common APOE1205763APOE31205763 genotype)whereas two copies of APOE4 increase the risk of AD about12-fold The mechanism by which the amino acid differencebetweenAPOE3 andAPOE4 increases the risk ofAD remainsto be established
Widespread occurrence ofAPP andA120573 in nervous systembrought about the assumption that both components mightplay physiological roles More than few possible conceptshave emerged some validated by experimental data TheAPP protein overexpression led to enhanced survival andgrowth of some cell types [54 55] Furthermore secreted
forms of APP (APPss sAPP120572 and sAPP120573) were antiapoptotic[56] and have a potent neuroprotective action in culturedrat hippocampal and septal neurons and in human corti-cal neurons [57] APPs
695and APPs
751protected neurons
against hypoglycemic damage and the neuroprotection wasabolished by antibodies to a specific region common toboth APPs
695and APPs
751 Thus APPss may normally play
excitoprotective and neuromodulatory roles AccordinglyAPP was shown to stimulate axon branching and the main-tenance and formation of synapses neuronal survival andneuritic outgrowth [58ndash60] APP protein is highly expressedin axons and interacts with extracellular matrix components[61ndash64] Similar to APP A120573 was demonstrated to play aphysiological role in synaptic plasticity as minute quantitiesof the peptide stimulated neurons and enhanced the releaseof neurotransmitter [65 66] Everything can changewhen thethings go awry
4 Perturbations in ER
In healthy cells including neurons ER is a fundamentalorganelle for protein quality control in the secretory pathwaywhich prevents protein aberrant folding and aggregation[67] A bulk of evidence shows the importance of ERin APP maturation and processing With regard to APPintracellular processing both secretases (120572- and 120573-) havebeen identified in the ER together with 120574-secretase whichis present in mitochondria-associated membrane (MAM)subcompartment [48 68] This distinctive intracellular lipid-raft-like structure is involved in cholesterol and phospholipidmetabolism Ca2+ metabolism and mitochondrial dynamicsand becomes markedly augmented in AD [69] MAM isresponsible for the communication between the ER and themitochondria with efficient transfer of Ca2+ from the ERto mitochondria supporting metabolic functions and cell
4 Oxidative Medicine and Cellular Longevity
ER membrane
6
ERAD
Proteasome26S
Ca2+
Ca2+
IP3R
VDAC
MCU
IMMOMM
ATP
GRP
75
MFN
1
MFN
2M
FN2
MFN
2
Sig1R
UPR
7
Mitochondria
Nucleus CHOP ATF6
XBP1ATF4
Gene expression
3
Transcriptionfactors
2
IRE1120572 PERK ATF6
1 GRP78
Chaperones
ER Misfolded proteins
Autophagosome
5
4
ERGolgitraffic
Golgi cisternae
Antioxidant responseApoptosisERAD and autophagyER chaperonesER and Golgi biosynthesis
Figure 2 ER stress triggered by misfolded proteins in several neurodegenerative diseases Abnormal conformations of the proteins APPA120573 and tau are implicated in the pathogenesis of AD Alterations in the function of ER chaperones and UPR-related components ERADERGolgi trafficking and ER-to-mitochondria Ca2+ transfer have been suggested as underlying mechanisms of ER stress triggered by thesedisease-associated proteins These proteins can accumulate and aggregate at the ER and their stable interaction with ER chaperones such asGRP78BiP and PDI may trap ER chaperones altering protein folding with concomitant ER stress In addition these proteins can lead to theoxidative modification of the active site of PDIs by nitrosylation leading to their enzymatic inactivation Furthermore some of these proteinsalter the activity of the UPR stress sensors (IRE1120572 PERK and ATF6) as well as the activitylevels of downstream signaling mediators andtranscription factors including cleaved ATF6 ATF4 and spliced XBP1 As a result genes implicated in autophagy and ERAD antioxidantresponse ER chaperones and organellersquos biosynthesis are upregulatedMoreover these proteins block the exit of vesicles from the ER and alterthe trafficking between ER andGolgi of properly folded proteinsThe cellular responses controlled byUPR transcription factors including themodulation of autophagy mediated degradation of protein aggregates become compromised Disease-related proteins can also interact withERAD components precluding the translocation of ERAD substrates from the ER to the cytosol leading to the accumulation of abnormallyfolded proteins at the ER Finally Ca2+ released from the ERmainly through the IP
3R and its transfer tomitochondria can be impaired in the
presence of disease-related proteins leading to mitochondrial Ca2+ overload and activation of apoptotic cell death pathways AD Alzheimerrsquosdisease ATF6 activating transcription factor 6 ATF4 activating transcription factor 4 ER endoplasmic reticulum ERAD endoplasmic-reticulum-associated protein degradation IP
3R inositol triphosphate receptor IRE1120572 inositol-requiring enzyme 1 alpha PERK protein
kinase R- (PKR-) like ER kinase UPR unfolded protein response XBP1 Xbox binding protein 1 Adopted from [67]
viability [70] The molecular bridges between ER inositol145-triphosphate receptor (IP
3R) and the voltage-dependent
anion channels in the outer mitochondrial membrane arebrought together through the cytosolic chaperone glucose-regulated protein 75 (GRP75) (Figure 2) Additionally thedynamin-relatedGTPasemitofusin 2 (Mfn2) proteins locatedon the ER intermingle with Mfn1 or Mfn2 on mitochondria
to tighten the connection The distance between ER andmitochondria controlled by the phosphofurin acidic clustersorting protein 2 (PACS-2) of ER and the dynamin-relatedGTPase protein 1 (Drp1) is crucial for cell survival as eithertoo long (lack of Ca2+ flux) or too short distance (Ca2+ over-load) might lead to apoptosis [71] Alternatively impairedmitochondrial bioenergetics with reduced cellular ATP
Oxidative Medicine and Cellular Longevity 5
levels stimulate autophagyThe molecular mechanism of ER-mediated autophagy is accurately regulated by Beclin 1 as wellas ER membrane bound protein Bax inhibitor 1 (BI-1) Bothproteins are capable of promoting autophagy through IP
3R-
dependent mechanism [72]
5 APP Processing and ER Stress Response
The nature of APP processing is determined by the com-position of membrane with cholesterol rich lipid rafts asthe site of amyloidogenic cleavage [73] Consequently onemight expect that fate of APP is at least partly establishedby the representation of lipid rafts and possible access to 120573-versus120572-secretase Irrespective of the type of cleavage there isone underestimated biochemical step in ER that might bringabout damaging upshot It is apparently the effectiveness ofER situated N-glycosylation which makes the APP moleculesuitably folded If misfoldedmalfolded protein(s) accumu-late in the ER complex cascade of reactions known as theunfolded protein response (UPR) is triggered with the so-called endoplasmic reticulum stress response (ERS) Today itis widely accepted that during UPR the ER sensors proteinkinase R- (PKR-) like ER kinase (PERK) activating proteinkinase 6 (ATF6) and inositol-requiring enzyme 1 alpha(IRE1120572) are freed fromGRP78BiP protein repression whichhereafter become activated Without going into details ofindividual sensor action a number of reactions occur at boththe genomic and cytoplasmic level with selective degradationof mRNAs encoding protein(s) with abnormal folding andinhibition of protein translation except for genes importantfor UPR redox homeostasis energy metabolism and pro-tein folding [67] On the one hand PERK phosphorylateseukaryotic initiation factor 2 alpha (eIF2120572) to stop entrance ofmethionyl-tRNA to the ribosome on the other hand it allowstranslation of activating transcription factor 4 (ATF4) geneIRE1120572 sets off alternative splicing of Xbox binding protein 1(XBP1) transcript leading to activation of the transcriptionfactor liable to stimulate ERGolgi biogenesis and formationof proteins involved in endoplasmic-reticulum-associatedprotein degradation known as ERAD (Erdj4 p58IPK EDEMRAMP-4 PDI-P5 and HEDJ for details see [67]) FinallyATF6 is activated in the Golgi complex through proteolyticcleavage and translocates to nucleus where it cooperates withXBP1 in upregulation of chaperones and ERAD-related genes[43] In principle UPR is activated to restore ER homeostasisand stop the accumulation of aberrantly formed protein(s)but if the strength of ER stress is unbearable (meaning thatit cannot be compensated by UPR) there is a path to activateapoptosis Among other routes the most central role isplayed by the major proapoptotic transcription factor CEBPhomologous protein CHOPgrowth arrest and DNA damageinduced gene 153 GADD153 It downregulates the antiapop-totic protein Bcl-2 and upregulates proapoptotic Bax and Bak[74] CHOPGADD153 leads to excessive production of reac-tive oxygen species (ROS) within ER subsequent depletionof reduced glutathione (GSH) and Ca2+ flux from the ER tocytoplasm through the IP
3R [75] One hitherto unresolved
issue is the physiologic importance of APP and GRP78BiPinteraction disclosed in coprecipitation study carried out by
Yamamoto et al [76] Bulk ofAPP associatedwithGRP78BiPwas immature protein Given that GRP78BiP expressionlevels declined in samples of brain tissue obtained fromFAD patients as demonstrated by Katayama et al [77] fewdistinct scenarios are possible First binding to GRP78BiPsuggests ER accumulation of immature APP Second APPinteraction with GRP78BiP is a noticeable sign of UPRwhich is further validated by downregulation of GRP78BiPin FAD patients Finally retention of APP in ER is mostlikely a result of amyloidogenic APP processing and as such itprobably occurs in MAM the cholesterol enriched domainsER-mitochondria crossing point is therefore of particularinterest in deciphering the links between the ER placed APPprocessing UPR and resulting cellular responses such asautophagy apoptosis and inflammatory reaction observed inAD (Figure 2)
6 Pathology of Tau Protein
Microtubule-associated protein tau controls assembly andprevents microtubules from severing Microtubular networkis fundamental component of cytoskeleton essential forintracellular transport of secretory vesicles and organelles(ie mitochondria) Glycogen synthase kinase 3 beta- (GSK-3120573-) targeted hyperphosphorylation of tau causes this proteinto dissociate frommicrotubules Consequently microtubulesbecome fragmented and microtubule-dependent transportsystem fails Furthermore hyperphosphorylated tau (P-tau)is prone to form oligomers and toxic filaments known asNFTs or tauopathy [78] A variety of tau conformers werereported to exist pointing to different tauopathies capa-ble of self-propagation [79 80] Interestingly intracellulartau inclusions define AD as clinical symptoms of diseaseobserved when tauopathy is abundant together with intra-cellular A120573 deposits in neocortex [51 81 82] People withabundant A120573 plaques but no or only a few neurofibrillarylesions do not have AD Clinicopathological correlationstudies have been crucial to generate hypotheses about thepathophysiology of the disease by establishing the fact thatthere is a continuum between ldquonormalrdquo aging and ADdementia and that the amyloid plaque buildup occurs primar-ily before the onset of cognitive deficits while neurofibrillarytangles neuron loss and particularly synaptic loss parallel theprogression of cognitive decline [83] Thus misfolded pro-teins and descendant toxic filaments with a number of inter-mediates are critical for manifestation of AD as fibrillogenicAPP processing is not enough for onset of disease Althoughthe molecular mechanism of tauopathy is not deciphered infull recent reports suggest ER stress as the starting point[84 85] This idea is substantiated by the elevated levels ofERS and UPR markers together with P-tau and GSK-3120573 inbrains affected by AD [86] Therefore on the one hand theincidence of UPR is strongly correlated with the presence ofNFTs on the other hand aggregation of P-tau induces ERSwith resultant UPR Some lines of evidence confirmed UPRactivation near the beginning of NFTs formation and pointto the functional link between malformed tau protein andUPRThe in vitro experiments with phosphatase 2A inhibitoror phosphorylation activator demonstrated enhanced P-tau
6 Oxidative Medicine and Cellular Longevity
formation in neurons in concert with the increased levelsof PERK eIF2120572 and XBP1 transcript apparent markers ofUPR [87] Furthermore GRP78BiP was found to encouragetau phosphorylation through facilitated substrate capture byGSK-3120573 [88] GSK-3120573 seems to play dual role first this kinaseprotects neurons from apoptosis as P-tau accumulation isstrong molecular signal to trigger UPR with subsequentautophagy Second UPR raises GSK-3120573 activity throughlysosomal degradation of inactive GSK-3120573 (P-Ser9-GSK-3120573)To sum up ERS and UPR are important molecular machinesused to prevent cell viability turned on by tauopathy
7 Oxidative Stress in ER
Sacs and tubes of ER delineate the compartment where thenewly synthesized proteins undergo maturation to nativestate Native state indicates properly folded fully functionalprotein Important reactions essential for protein folding ofunbranched polypeptide chains include amino acids oxida-tion and glycosylation As a result redox homeostasis inER is shifted to oxidative state so as to promote disulfidebond formation between adjacent cysteines Oxidation ofsulfhydryl groups required to make disulfide bonds is con-trolled by ER oxidase 1120572 (ERO1120572) Next disulfide bondscould be subject to posttranslational modification disulfideexchange by protein disulfide isomerase (PDI) PDI is ableto correct mispaired thiol residues by catalyzing the breakageand formation of correct disulfide bonds These enzymes arefundamental for protein folding Oxidation allows twistingof proteins which is followed by N-glycosylation andor C-and O-mannosylation Protein N-glycosylation in eukary-otes is a complex process divided into several steps Firstthere is a ldquocall forrdquo carrier lipids (polyisoprenyl phosphatessuch as dolichyl phosphates) the membrane lipids knownto function as glycosyl transporters In mammalian cellsthe limiting substrate for dolichol biosynthesis is geranyl-geraniol (GGOH) of mevalonate pathway Dolichols thelongest aliphatic molecules synthesized in animal cellshave 18ndash21 120572-isoprene saturated units (C90ndash105) criticalfor their recognition by the enzymes (glycosyltransferases)that glucosylate dolichyl phosphates [89] Once dolichylmonophosphates (Dol-P) are formed in the ER membranethe precursor oligosaccharide donor (GLc
3Man9GLCNAc
2-
P-P-dolichol) for protein N-glycosylation can be synthesizedon the lumenal leaflet of ER First three sugar intermediatesare produced (Man-P-Dol Glc-P-Dol GlcNAc-P-P-Dol andMan5GlcNAc-P-P-Dol) on the cytoplasmic leaflet of the
ER Next enzyme flippases mediate transbilayer movementof the aforementioned intermediates to lumenal side ofER where conversion to Glc
3Man9GlcNAc
2-P-P-Dol could
be completed Glc3Man9GlcNAc
2-P-P-Dol is also used for
biosynthesis of glycosylphosphatidylinositol (GPI) anchorsTaken together lipid-mediated glycosylation plays a vital
role in the appropriate protein folding and intracellulartranslocation of N-linked glycoproteins [90] Likewise it isimportant for protein O- and C-mannosylation and GPIanchorage Moreover Dol-P availability in the ER is the rate-limiting factor in the production of glycolipid intermediatesand N-glycosylation
8 ER Stress and Apoptosis
It was shown that during ERS the ER resident proapop-totic cysteine protease known as caspase (caspase-12 in ratcaspase-4 in humans) is activated through cleavage As aresult caspase cascade is started via caspase-9 that in turnstimulates effector caspase-3 [91 92] The central role playedby ER in programmed cell death is achieved by PERK branchwhere ATF4 induces the expression of CHOPGADD153which represses antiapoptotic Bcl-2 family proteins andsimultaneously shuffles ER membrane Bax and Bak proteinsinto outer mitochondrial membrane Consequently poresare formed to leak the components of apoptosome frommitochondrial intermembrane space [93] Another impor-tant mechanism of ERS-induced apoptosis is led by Ca2+-dependent ERO1120572-IP
3R pathway where ERO1120572 collaborates
with IP3R in Ca2+ efflux from ER to mitochondria via MAM
[75 94] Accordingly Ca2+ influx facilitates cytochrome 119888release frommitochondria besides cytochrome 119888 can bind toER IP
3R and the complex amplifies the apoptotic signal in a
feedforward manner [95] Last but not least ERS-associatedapoptotic programme is set off by c-Jun N-terminal kinase(JNK) as the effect of IRE1120572 complexed with TNF-receptor-associated factor 2 (TRAF2) activation of apoptosis-signal-regulating kinase 1 (ASK1) [96 97]
What does ERS drive to induce apoptotic death inneurons Actually many reports indicate that APP and A120573 aswell as hyperphosphorylated tau have been shown to blockmitochondrial transport which results in impaired energystorage and oxidative stress [98ndash101] Indeed accumulationof APP A120573 and NFTs in mitochondria led to reducedactivity of some enzymes involved in substrate oxidation(tricarboxylic acid cycle) electron transport chain (ETC)and ATP synthase as well as severely diminishing import ofnuclear-encoded proteins [26 27 102ndash104] One may ask ifthere is any additional link between ER and mitochondriaother than MAM which could account for apoptotic signalThough not directly ER significantly contributes to oxidativestress in mitochondria of AD-affected subjects
9 Abnormal APP Processing and TraffickingCulminate in ER Pathology of AD
From the morphological point of view as neurons arehighly specialized cells soma dendrites and neurites areconsiderably distinct structures Proteins needed by thesecompartments are delivered via microtubules once proteinshave suitable sorting signals (ie APP trafficking from ERto plasma membrane is associated with several posttrans-lational modifications with oxidation and N-glycosylation)Additionally the Golgi apparatus follows ER in subsequentAPP adjustment (O- and N-glycosylation phosphorylationand sulphonation) [42] Any inaccurate alteration of the APPmolecule is potentially hazardous as protein final destiny ismissed causing its retention in ER or trans-Golgi network(TGN) In addition other unusual settings for APP arepossible as this large protein has few signal sequences hiddenwhen APP is correctly folded In the cells transfected withAPP this protein enters coat protein complex I (COPI)
Oxidative Medicine and Cellular Longevity 7
vesicles and undergoes retrograde transport from cis endof the Golgi complex back to the ER [76] Such responsecauses accumulation of APP in the tubulocisternal ER systemtogether with aberrant intracellular translocation of theprotein Interestingly the issue whether APP is subject toretrograde transport with successive fibrillogenic processingbecause of UPR and its interaction with GRP78BiP is notclear as the observations are inconsistent [77] Nonethelessaccumulation of misfoldedmalfolded proteins in the ERsuggests disorganized process of posttranslational change Asanticipated the accretion of proteins of anomalous patternsignals ERS and UPR followed by increased vulnerability toapoptotic cell death Prior to decay however APP proteinlevels in the ER lessen by dint of cleavage with 120573- and120574-secretase [105 106] Products of this cleavage (sAPP120573A120573 and AICD fragment) all appreciably influence neuronalsurvival most likely through nonnative form of the APPsubstrate
10 APP A120573 and NFTs MarkMitochondria as Targeted in AD
Mitochondrial import of A12057340
and A12057342
peptides throughthe translocase TOM complex was blocked by preincubationof isolated mitochondria with antibodies raised againstTOM proteins (TOM20 TOM40 and TOM70) [107]Neither VDAC inhibition with antagonist antibodies norinhibition of mitochondrial permeability transition pores(MPTP) or fall of mitochondrial membrane potential(MMP) affected uptake of A120573 [17] With regard to APPelegant study performed by Anandatheerthavarada andhis colleagues [30] revealed that C-terminal truncatedAPP (lacking A120573) targets mitochondria in cholinergicGABAergic dopaminergic and glutamatergic neurons ofAD brain by clogging up mitochondrial protein translocasecomplex TOM40TIM22 Authors propose that occlusionof translocase is followed by blunted import of nuclear-encoded proteins vitally important for energy homeostasisMitochondrial APP protein transmembrane orientationindicates NH
2-terminal inside in contact with translocase
whereas COOH-terminal is facing cytoplasmic side NH2-
terminus has mitochondrial signal sequence Astonishinglymitochondrial APP molecules were nonglycosylated givingrise to speculation that protein molecules that arrivedat mitochondria have not achieved molecular maturity[31] The accumulation of nonglycosylated APP species inmitochondrial import channels of AD brain was directlyrelated to decreased mitochondrial functions as validatedby the decline in cytochrome 119888 oxidase activity (complexIV) and elevated levels of H
2O2 Furthermore in AD
brain the mitochondrial accumulation of nonglycosylatedAPP went along with a corresponding reduction in plasmamembrane-associatedAPP It suggests that ADbrain hasAPPprocessing and trafficking severely affected by incompleteN-glycosylation ensuing ER protein accumulation andexposure of the cryptic mitochondrial targeting signalfor assisting chaperone proteins Several lines of evidenceindicate that not fully formed proteins are phosphorylatedand bind to the cytosolic proteins required for movement
from ER to mitochondria [108ndash110] Moreover ERS andUPR in AD seem to be incompetent and inefficient inelimination of malfolded proteins In addition to APPalso A120573 was frequently reported to occupy mitochondriaalthough its origin and mechanism of mitochondriatargeting mostly remain unexplored (except for involvementof protein translocase TOM complex) The possibilityof mitochondrial A120573 generation has to be ruled out asmembrane orientation of arrested APP does not allow accessto 120574-secretase (120574-secretase activity in mitochondria wasdetected by independent study) [111] Probably A120573 speciesis derived from APP prior to its translocation (ER) soA120573 may be transported to mitochondria independentlyof APP Collectively observations showing mitochondrialpresence of APP A120573 and tau in aberrant configuration pointtoward the anomaly of protein folding at the level of ER andGolgi apparatus Nonglycosylated molecule of APP suggestsdefective transfer of sugar core from dolichyl phosphate(s)and further modifications such as O- and N-glycosylationsOne may admit that the lack of glycosyl residue bringsabout pathologic processing and trafficking of APP and itsfragments In point of fact dolichol derivatives mixture ofpolyprenols (acyclic isoprenoid alcohols) known as Ropren(Solagran Limited Melbourne Australia) commerciallyused to treat liver diseases were tested in the treatment ofAD in two separate trials conducted in 2005 and 2006 withpromising results [httpwwwasxcomauasxpdf20071119pdf315x5nh4hm8wv7pdf httpwwwasxcomauasxpdf20070221pdf3111ztwcbqzkk9pdf] Further studies areurgently needed to test how important the glycosylationprocess is in the pathogenesis of AD Dolichols areobtained from geranylgeraniol (GGOH) and the latter is anintermediate of mevalonate pathway GGOH is a commonsubstrate for dolichol and ubiquinone synthesis but it isalso necessary for protein prenylation As both GGOHand farnesol (FOH) are engaged in protein prenylationmore concern should be laid on the importance of thesecompounds in AD pathogenesis Observations demonstrat-ing mitochondrial relocalization of other proteins withoutposttranslational modification seemingly point to increasedmitochondrial targeting of immature molecules resultingin mitochondrial dysfunction and acceleration of diseaseprogression [34 112ndash114]
11 Oxidative Stress in Mitochondria ofAD Brains
Normal physiological functions of APP are thought to beinvolved in the stabilizing contact points between synapsesand maintaining mitochondrial functions [60 115] Mito-chondrial dysfunction was often observed regardless of theexperimental model used to study AD [8 29 116ndash119]It includes defects in oxidative phosphorylation decreasedATP decreased membrane potential increased productionof ROSRNS and perturbation in mitochondrial fusion andfission [15 30 31 115 120ndash122] Hyperphosphorylated tau wasalso reported to impair mitochondrial functions [123] Usingproteomic approach the strongest defects of the respiratorycapacity were observed mainly at complexes I IV and ATP
8 Oxidative Medicine and Cellular Longevity
synthase (complex V) at both protein and activity level [124]While APP A120573 and hyperphosphorylated tau are potentinhibitors of mitochondrial import of nuclear-encoded pro-teins apparently the pathology of each leads to metabolicharm in different way In freshly isolated mitochondriafrom AD brains the APP inhibited mitochondrial importof cytochrome 119888 oxidase (COX) subunits IV and Vb [31]Dysfunction of COX increases ROS production (incompletereduction of oxygen molecules) reduces energy stores anddisturbs energy metabolism Accordingly in AD patientsdeficiency of COX was found in brains and platelets [119125 126] Similar to APP A120573 was found in mitochondriaof transgenic mice and cellular and human AD models [1719 33 103 127ndash130] At present it is not clear whether theobservedmitochondrial toxicity is due toAPP orA120573 orNFTsaccumulation Anyway some regularity is observed withrespect to most affected components of respiratory chainNADH-ubiquinone oxidoreductase (complex I) activity isreduced to the utmost by hyperphosphorylated tau whereasdecreased activity of cytochrome 119888 oxidase (complex IV)that resulted in mitochondrial dysfunction was observedduring A120573 and APP accumulation [14 15 31 32 34]Concomitantly rise in the activity of antioxidant enzymesmanganese superoxide dismutase (Mn-SOD) and catalase(CAT) was demonstrated in response to elevated levelsof free radicals including superoxide anion radical (O
2
∙minus)hydroperoxyl radical (HO
2
∙) hydroxyl radical (OH∙) andnitric oxide radical (NO∙) [15] Consistent with observationsof chronic respiratory chain dysfunction and mitochondrialoxidative stress there are reports showing their contributionto tau pathology in AD [131] In any case free radicals thatoverride antioxidant defense react with a wide variety oforganic components causing lipid peroxidation to advancedlipid oxidation end products (ALE) cross-linking of proteinsnitrosylation of proteins and mutations in DNA Mitochon-drial circular DNA (mtDNA) of sim16 kbp is devoid of repairsystems meaning the buildup of lesions There are 37 geneslocated in mtDNA with those encoding protein subunits ofcomplex I (7) complex II (1) and complex IV (3) but notcomplex III Interestingly hallmarks of AD (APP A120573 andNFTs)mostly affectmembers of electron transfer chain (com-plexes I and IV) which rely exclusively on mitochondriallypredetermined subunits Maybe it is not simple coincidencebut the effect of inhibited import to mitochondria of nuclear-encoded subunits of complexes I and IV gives explanationfor toxicity of APP or A120573 or NFTs in mitochondria Thereare additional findings in mitochondria affected by AD suchas lower activity of pyruvate dehydrogenase (PDH) andoxoglutarate dehydrogenase (OGDH) [104 132] Inhibitionof OGDH the enzyme of tricarboxylic acid cycle minimizesthe NADH pool and electron number needed for ETCand mitochondrial membrane potential (Δ120595
119898) to create and
maintain proton gradient obligatory for ATP synthesisThereare also lines of evidence for direct inhibitory action of solubleoligomeric A120573 species on ABAD (A120573-binding alcohol dehy-drogenase) and internal membrane cyclophilin D (CypD)resulting in increased mitochondrial membrane permeabil-ity (MPTP) potentiated ROS production synaptic lossdiminished activity of mitochondrial respiration and finally
cell death [129 132ndash134] CypD knockout prevents mito-chondrial and neuronal perturbations and improves mito-chondrial function in Alzheimerrsquos disease mouse model [128135] It has to be emphasized that oxidativenitrosative stressaffects the fusion and fission process ofmitochondria Fusionthat is speedup by small GTPases mitofusins (Mfn1Mfn2)improves efficiency of mitochondrial respiration and ATPproduction Mitochondrial dynamics are severely imbal-anced in AD cases in favour of fission through elevatedexpression of the fission protein DLP1 (dynamin-like protein1) associatedwith nitrosative stress stirred up byA120573 [136 137]Collectively these observations indicate that toxic intracel-lular A120573
42-43 oligomers differ in action from extracellularaggregates found in amyloid plaques of AD brains [138](Figure 3)
12 Proteostasis in Mitochondria
Mitochondrial protein turnover grants the well-organizedreplacement of nonfunctional proteins into operational oneThe first task is attained through proteolytic degradation ofinner membrane and matrix proteins with local mitochon-drial proteases [139] or outer membrane proteins throughubiquitin-proteasome system [140] The second task is metby intramitochondrial protein synthesis but as almost 1500different nuclear-encoded proteins have to be importedto complete mitochondrial proteome a matter of capableimport is fundamental for the function of the organelleIn extreme cases of cellular injuries observed in neurode-generative diseases damaged mitochondria with extendedloss of the electrochemical potential are selectively removedby autophagy known as mitophagy [112] APP and A120573accumulation has also something to do with altered mito-chondrial dynamics as fission takes advantage of fusion withresultant dysfunction of mitochondria and neurons [137]AD neurons demonstrated selective mechanisms of prote-olytic clearance of oxidatively and nitrosatively modifiedproteins in mitochondria Insulin degrading enzyme (IDE)prevents formation of toxic insoluble fibrils from A120573 asit cleaves A120573 prior to aggregation [141ndash143] A novel zinc-metallopeptidase Presequence Protease (PreP) a member ofpitrilysin oligopeptidase family degrades either intramito-chondrially stored A120573
40or A120573
42proteinThis protease seems
to be highly sensitive to oxidative stress as disulfide bridgeformed between two proximal cysteine residues blocks itscatalytic activity [144ndash149] Another mitochondrial serineprotease HtrA2Omi occupies intermembrane space whereit can cleave APP locked up in protein translocase complex[121] Even though it is well established that HtrA2Omiis released to the cytosol to amplify apoptosis through thedegradation of antiapoptotic proteins and caspase activation[150ndash152] it is also implicated in proteolytic deletion ofmalfoldedAPP in ER [151] Fromknockout studies onmice itis obvious that HtrA2Omi plays a significant shielding roleas mice deficient in this protease exhibit neurodegenerativephenotype with weight loss and premature death [153]Taken together understanding the mechanisms of clearanceof the unwanted proteins including APP A120573 and tau isvital in an attempt to get rid of them from mitochondria
Oxidative Medicine and Cellular Longevity 9
VDAC
ANT
A120573
A120573
A120573
A120573
A120573
A120573
A120573A120573
A120573
A120573
A120573 CypD
CypD ProApCyt c Ca2+
IMM
OMM
IMSΔΨ
+
+
+
minus
minus
minus
ABAD
ROS
ROS
TIM
TOM
Proteinimport
NAD
NAD
NAD
NAD
H+
H+
H+
H+
H+
ADP
Mn-SOD
CuZn-SOD
mtDNA
V
Matrix
UCP
O2∙minus
O2∙minus
IV
III
II
I
O2
O2
DLP1Fis1
Mfn1Mfn2
Opa1
mHsp70
NADH
NADH
NADH
TCAcycle
PyruvateAcetyl-CoA
120572KGDH
120572-KG
PDH
Isocitrate
CitrateOAA
Malate
Fumarate
Succinyl-CoASuccinate
GDPGTP
FAD
FAOH
ATP
Figure 3 Amyloid-120573-related mitochondrial impairment Mitochondria were found to be the target for amyloid-120573 (A120573) which interactswith several proteins leading to mitochondrial dysfunction Indeed A120573 was found in the outer mitochondrial membrane (OMM) andinner mitochondrial membrane (IMM) as well as in the matrix The interaction of A120573 with the OMM affects the transport of nuclear-encoded mitochondrial proteins such as subunits of the electron transport chain complex IV into the organelle via the translocase of theouter membrane (TOM) import machinery Moreover A120573 disturbs the activity of several enzymes such as pyruvate dehydrogenase (PDH)and oxoglutarate dehydrogenase (OGDH) decreasing NADH reduction and the electron transport chain enzyme complex IV reducing theamount of hydrogen that is translocated from the matrix to the intermembrane space (IMS) thus impairing the mitochondrial membranepotential (MMP) Taken together these events cause abnormal mitochondrial electron activities leading to decreased complex V activityand so to a drop in ATP levels in addition to increasing reactive oxygen species (ROS) generation Moreover ROS induce peroxidationof several mitochondrial macromolecules such as mitochondrial DNA (mtDNA) and mitochondrial lipids contributing to mitochondrialimpairment in themitochondrial matrixThe complex of A120573 bound to binding alcohol dehydrogenase (ABAD) impairs the binding of NAD+to ABAD changes mitochondrial membrane permeability and reduces activities of respiratory enzymes inducing further ROS productionand leading to mitochondrial failure A120573 binding also activates Fis1 (fission protein) and promotes increased mitochondrial fragmentationthis increased mitochondrial fragmentation produces defective mitochondria that ultimately damage neurons Furthermore A120573 binding tocyclophilin D (CypD) enhances the protein translocation to the inner membrane favouring the opening of the mitochondrial permeabilitytransition pore formed by the adenine nucleotide translocator (ANT) and voltage-dependent anion channels (VDACs) Cyt 119888 cytochrome 119888DLP1 dynamin-like protein 1 PDH pyruvate dehydrogenase ProAp proapoptotic factors SOD superoxide dismutase TCA tricarboxylicacid TIM translocase of the inner membrane Adopted from [14]
Some hopes are associated with the application of at presentindefinite modulators of proteolytic activity HtrA2Omi isfor example activated by PTEN-induced putative kinase 1(PINK1) upon phosphorylation at Ser142 residue [154] Alter-natively accent has to be put on mitophagy of dysfunctionalmitochondria and mitochondriogenesis
13 Aging versus AD
Themost commonneurodegenerative disorder is representedby Alzheimerrsquos disease characterized by declining memoryreduced cognitive capacity and progressive dementia whichare often fatal to elderly individuals above 65 years of age
10 Oxidative Medicine and Cellular Longevity
It is ranked as the fourth leading cause of death in modernsocieties where average life span increased greatly in the lasttwo decades As 95 of AD cases are diagnosed in olderpeople one might think that a causal relationship existsbetween aging and the onset of disease Certainly a numberof similarities between getting old and being affected withAD could be listed Historically the free radical theory ofaging by Harman [155] suggested aging as ldquoside effectrdquo ofreactive oxygen species formed in mitochondrial respiratorychain Apparently free radicals commonly generated byincomplete reduction of oxygen molecule at complexes Iand III of mitochondrial ETC are capable of damagingDNA RNA and proteins They impair energy storage andlead to operational failure of mitochondria with progres-sive decline of cell viability Almost identical conditionsaccompany AD and are an explicit step in pathogenesisof disease As mtDNA is deprived of repair mechanismsROS-induced DNA strand breaks tend to accumulate withage or AD Thus mtDNA is a vulnerable target for ROSbut the reverse the ROS generation due to the mutatedmtDNA is not convincingly confirmed [156] Moreover theevidence that mutated mtDNA accelerates the progress ofaging is also questioned based on the results from studycarried out on transgenic mice model [157] Although someauthors show inconsistency between the free radical theoryand observations cumulative evaluation of the scientificreports points to antioxidant defense systems as importantfactors in protection from premature aging [158] Othermitochondrial components important for their function arehampered with age adenine nucleotide translocase (ANT)nitric oxide synthase (NOS) and carnitine acyltransferase(CT) [159ndash161] Actually NOS activity is elevated in AD asreported from study performed on cellular model of disease[15] We could not find any information about CT activityin AD whereas ANT activity is noticeably inhibited byA120573 or hyperphosphorylated tau and this effect is reversedby mersalyl a reversible alkylating agent of thiol groups[162] Mitochondrial dysfunction observed in transgenicmice models of AD and aging demonstrates higher activ-ity of genes controlling energy metabolism and apoptosisTaken together physiological aging and AD are associatedwith broad-spectrum dysfunction of mitochondria but thefoundations of mitochondrial decline are dissimilar Morediscrepancies between physiological aging and AD werefound with respect to ERS and UPR which play a significantrole in cellular proteostasis Dolichol was selected as an agingmarker because the progressive increase in dolichol levelwas observed in aging brain [163] In contrast ubiquinoneconcentration which is also synthesized from geranylgeranioldiminishes with agingwhereas cholesterol and dolichyl phos-phate concentrations remain unaltered In AD decreasedlevels of dolichol were observed and increased levels ofubiquinone and dolichyl phosphate without any changesin brain cholesterol AD cannot be regarded as a resultof premature aging The drop in dolichol and augmenteddolichyl phosphate concentration points toward disturbedglycosylation in ER of diseased brain while the increase inubiquinone suggests efforts to protect the brain from oxida-tive stress induced by lipid peroxidation [164 165]
14 Targeting ER Stress in AD Therapy
As ERS is a recognized factor in AD drugs that interfere withERS would theoretically have great therapeutic potentialThere are several compounds grouped in classes thatinteract directly with components of the ERS (salubrinalBiP inducer X (BIX) salicylamide analogs flavonoidsguanabenz and STF083010) chemical chaperons (4-phenyl-butyric acid (PBA) tauroursodeoxycholic acid (TUDCA)and trimethylamine oxide (TMAO)) chemicals that inhibitprotein degradation (Eeyarestatin MG132 and Bortezomib)compounds with antioxidant activity (Edaravone diben-zoylmethane derivatives and N-acetyl cysteine (NAC)) anddrugs controlling calcium signaling (dantrolene and carba-zole derivatives) [166] They may act by inducing transienttranslation arrest upregulation of chaperone proteins andaugmented degradation of ER-associated misfolded proteinsFundamental approach in the development of new therapyis the selection of appropriate molecular targets In ER stresssignaling the aim is to alter the expression of ER stress-associated molecules that can rescue cells from the toxiceffect of ERS Recent efforts in establishing new promisingdrugs against AD are pointing to chemical chaperones suchas PBA TUDCA or trimethylamine oxide (TMAO) Thesesubstances improve protein folding and alleviate nativeprotein conformation [166] It was shown on mouse modelsof AD that PBA TUDCA and TMAO stop A120573 accumulationand avoid the loss of dendritic spines [167] Some observa-tions even demonstrated improved memory and cognitivefunctions [168] associated with improved cell survival [169]Salubrinal ((2E)-3-phenyl-N-[222-trichloro-1-[[(8-quinoli-nylamino)thioxomethyl]amino]ethyl]-2-propenamide Sal)selectively inhibits growth arrest and DNA damage inducedgene 34- (GADD34-) phosphatase complex (GADD34associates with protein phosphatase 1 (PP1)) and promotes invitro dephosphorylation of the alpha subunit of eIF-2120572 andIRE1120572ASK1JNK signaling pathway being protective againstERS even induced by tunicamycin Tm [170] In a great deal ofexperiments testing Sal in cultured cells and animal modelsof AD this substance increased the viability of neuronal cellsand A120573 toxicity [171 172] BIX (2-(34-dihydroxyphenyl)-2-oxoethyl ester thiocyanic acid) preferentially inducedBiP mRNA in an ATF6-dependent manner leading toreduced Tm-induced death of neuronal cells [173] AlsoDBM derivative 14ndash26 (221015840-dimethoxydibenzoylmethane)was found to be neuroprotective for SH-SY5Y and PC-12cells by decreasing expression of BiP and CHOP [174]Dantrolene a ryanodine receptor antagonist that inhibitsabnormal calcium release from the ER inhibited expressionof both phosphorylated PERK and eIF2120572 It also reducedCHOP expression and attenuated thapsigargin-inducedapoptosis in PC-12 cells [175] Neuroprotective effects similarto dantrolene were observed for ([9-(3-cyanobenzyl)-14-dimethylcarbazole]) This substance suppressed increases inintracellular Ca2+ in PC-12 cells treated with thapsigarginand reduced levels of BiP and CHOP [176]
Aforementioned compounds were chosen from othersas the most potent ER stress inhibitors and persuasivelyprotective to neuronal cells 18 of 42 different compounds
Oxidative Medicine and Cellular Longevity 11
were exploited in in vivo and in vitro models of centralnervous system disorders with in fact improved cell or tissueviability [166] Thus the brain is the most frequently investi-gated organ in the context of ERS From these experimentsit becomes clear that CHOP functions as proapoptotic factorThe roles of other specific ER stress molecules as moleculartargets for pharmacological intervention are less clear andvary depending on cell type and context
There are few underestimated modulations in APP pro-cessing that shed more light on current dogma of AD patho-genesis The modulation of mevalonate pathway and choles-terol synthesis were reported to stimulate nonamyloidogenicpathway of APP processing [177] Additionally cholesterolderivative 27-hydroxycholesterol (27-OHC) was shown toinduce ER stress which attenuated leptin-dependent viabilityby activating CHOP in SH-SY5Y neuroblastoma cells [178]Irrespective of a number of compounds examined in AD thecall for new drugs modulating ER stress with healing effect isstill waiting to be revealed
Abbreviations
APOE Apolipoprotein EAPPs Secreted form of APPCHOPGADD153 CEBP homologous protein
growth arrest and DNA damageinduced gene 153
Drp1 Dynamin-1-like proteinERAD Endoplasmic-reticulum-
associated protein degradationERO1120572 ER oxidase 1120572ERS Endoplasmic reticulum stressFOH FarnesolGGOH GeranylgeraniolGRP75 Glucose-regulated protein 75GRP78BiP 78 kDa glucose-regulated proteinGSK-3120573 Synthase kinase 3 betaIP3R Inositol triphosphate receptor
IRE1120572 Inositol-requiring enzyme 1 alphaMAM Mitochondria-associated
membraneMAP LC3 Microtubule-associated protein
light chain 3Mfn2 GTPase mitofusin 2MMP Mitochondrial membrane
potentialMPTP Mitochondrial permeability
transition poreNFT Neurofibrillary tanglesPACS-2 Phosphofurin acidic cluster
sorting protein 2PDI Protein disulfide isomerasePERK Protein kinase R- (PKR-) like
ER kinaseRNS Reactive nitrogen speciesROS Reactive oxygen speciesRyR Ryanodine receptorTOM Transporter outer membraneTIM Transporter inner membrane
UPR Unfolded protein responseVDAC Voltage-dependent anion channelXBP-1 Xbox binding protein 1
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
Support for this work was provided by Grant no UMO-201103BNZ70238 from the National Science Centre inPoland
References
[1] R L Nussbaum andC E Ellis ldquoAlzheimerrsquos disease and Parkin-sonrsquos diseaserdquoTheNewEngland Journal ofMedicine vol 348 no14 pp 1356ndash1364 2003
[2] A L Brunkan and A M Goate ldquoPresenilin function and 120574-secretase activityrdquoThe Journal of Neurochemistry vol 93 no 4pp 769ndash792 2005
[3] V Crentsil ldquoThe pharmacogenomics of Alzheimerrsquos diseaserdquoAgeing Research Reviews vol 3 no 2 pp 153ndash169 2004
[4] D L Price and S S Sisodia ldquoMutant genes in familialAlzheimerrsquos disease and transgenic modelsrdquo Annual Review ofNeuroscience vol 21 pp 479ndash505 1998
[5] D Harman ldquoAlzheimerrsquos disease pathogenesis role of agingrdquoAnnals of the New York Academy of Sciences vol 1067 no 1 pp454ndash460 2006
[6] D J Selkoe ldquoTranslating cell biology into therapeutic advancesin Alzheimerrsquos diseaserdquo Nature vol 399 pp A23ndashA31 1999
[7] H Braak and E Braak ldquoNeuropathological stageing of Alz-heimer-related changesrdquo Acta Neuropathologica vol 82 no 4pp 239ndash259 1991
[8] K Hirai G Aliev A Nunomara et al ldquoMitochondrial abnor-malities inAlzheimerrsquos diseaserdquoTheJournal ofNeuroscience vol21 pp 3017ndash3023 2001
[9] U T Brunk C B Jones and R S Sohal ldquoA novel hypothesisof lipofuscinogenesis and cellular aging based on interac-tions between oxidative stress and autophagocytosisrdquoMutationResearchDNAging vol 275 no 3ndash6 pp 395ndash403 1992
[10] P A Stewart K Hayakawa M-A Akers and H V Vinters ldquoAmorphometric study of the blood-brain barrier in Alzheimerrsquosdiseaserdquo Laboratory Investigation vol 67 no 6 pp 734ndash7421992
[11] J P Blass and G E Gibson ldquoThe role of oxidative abnor-malities in the pathophysiology of Alzheimerrsquos diseaserdquo RevueNeurologique vol 147 no 6-7 pp 513ndash525 1991
[12] B Pająk E Kania and A Orzechowski ldquoNucleofection of ratpheochromocytoma PC-12 cells with human mutated beta-amyloid precursor protein gene (APP-sw) leads to reducedreduced viability autophagy-like process and increased expres-sion and secretion of beta amyloidrdquo BioMed Research Interna-tional vol 2015 Article ID 746092 10 pages 2015
[13] J P Blass ldquoCerebrometabolic abnormalities in AlzheimerrsquosdiseaserdquoNeurological Research vol 25 no 6 pp 556ndash566 2003
12 Oxidative Medicine and Cellular Longevity
[14] A Eckert K Schmitt and J Gotz ldquoMitochondrial dysfunc-tionmdashthe beginning of the end in Alzheimerrsquos disease Sep-arate and synergistic modes of tau and amyloid-120573 toxicityrdquoAlzheimerrsquos Research ampTherapy vol 3 article 15 pp 1ndash11 2011
[15] U Keil A Bonert C A Marques et al ldquoAmyloid 120573-inducedchanges in nitric oxide production and mitochondrial activitylead to apoptosisrdquoThe Journal of Biological Chemistry vol 279no 48 pp 50310ndash50320 2004
[16] A M Swomley and D A Butterfield ldquoOxidative stress inAlzheimer disease and mild cognitive impairment evidencefrom human data provided by redox proteomicsrdquo Archives ofToxicology vol 89 no 10 pp 1669ndash1680 2015
[17] C Caspersen N Wang J Yao et al ldquoMitochondrial A120573 apotential focal point for neuronal metabolic dysfunction inAlzheimerrsquos diseaserdquo The FASEB Journal vol 19 no 14 pp2040ndash2041 2005
[18] G E Gibson K-F R Sheu and J P Blass ldquoAbnormalities ofmitochondrial enzymes inAlzheimer diseaserdquo Journal of NeuralTransmission vol 105 no 8-9 pp 855ndash870 1998
[19] A Eckert S Hauptmann I Scherping et al ldquoSoluble beta-amyloid leads to mitochondrial defects in amyloid precursorprotein and tau transgenic micerdquo Neurodegenerative Diseasesvol 5 no 3-4 pp 157ndash159 2008
[20] A Eckert K L Schulz V Rhein and J Gotz ldquoConvergenceof amyloid-120573 and tau pathologies on mitochondria in vivordquoMolecular Neurobiology vol 41 no 2-3 pp 107ndash114 2010
[21] S Hauptmann I Scherping S Drose et al ldquoMitochondrial dys-function an early event in Alzheimer pathology accumulateswith age in AD transgenic micerdquoNeurobiology of Aging vol 30no 10 pp 1574ndash1586 2009
[22] M Manczak B S Park Y Jung and P H Reddy ldquoDifferentialexpression of oxidative phosphorylation genes in patients withAlzheimerrsquos disease implications for early mitochondrial dys-function and oxidative damagerdquo NeuroMolecular Medicine vol5 no 2 pp 147ndash162 2004
[23] M P Mattson M Gleichmann and A Cheng ldquoMitochondriain neuroplasticity and neurological disordersrdquo Neuron vol 60no 5 pp 748ndash766 2008
[24] P I Moreira M S Santos and C R Oliveira ldquoAlzheimerrsquosdisease a lesson frommitochondrial dysfunctionrdquoAntioxidantsamp Redox Signaling vol 9 no 10 pp 1621ndash1630 2007
[25] P H Reddy ldquoMitochondrial dysfunction in aging and Alz-heimerrsquos disease strategies to protect neuronsrdquo Antioxidants ampRedox Signaling vol 9 no 10 pp 1647ndash1658 2007
[26] V Rhein G Baysang S Rao et al ldquoAmyloid-beta leads toimpaired cellular respiration energy production andmitochon-drial electron chain complex activities in humanneuroblastomacellsrdquo Cellular and Molecular Neurobiology vol 29 no 6-7 pp1063ndash1071 2009
[27] V Rhein X Song A Wiesner et al ldquoAmyloid-120573 and tausynergistically impair the oxidative phosphorylation system intriple transgenic Alzheimerrsquos disease micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 47 pp 20057ndash20062 2009
[28] R X Santos S C Correia X Wang et al ldquoAlzheimerrsquos diseasediverse aspects ofmitochondrial malfunctioningrdquo InternationalJournal of Clinical and Experimental Pathology vol 3 no 6 pp570ndash581 2010
[29] N R Sims ldquoEnergy metabolism oxidative stress and neuronaldegeneration in Alzheimerrsquos diseaserdquoNeurodegeneration vol 5no 4 pp 435ndash440 1996
[30] H K Anandatheerthavarada G Biswas M-A Robin and NG Avadhani ldquoMitochondrial targeting and a novel transmem-brane arrest of Alzheimerrsquos amyloid precursor protein impairsmitochondrial function in neuronal cellsrdquo The Journal of CellBiology vol 161 no 1 pp 41ndash54 2003
[31] L Devi B M Prabhu D F Galati N G Avadhani and HK Anandatheerthavarada ldquoAccumulation of amyloid precur-sor protein in the mitochondrial import channels of humanAlzheimerrsquos disease brain is associated with mitochondrialdysfunctionrdquo The Journal of Neuroscience vol 26 no 35 pp9057ndash9068 2006
[32] L Devi and H K Anandatheerthavarada ldquoMitochondrialtrafficking of APP and alpha synuclein relevance to mito-chondrial dysfunction in Alzheimerrsquos and Parkinsonrsquos diseasesrdquoBiochimica et Biophysica Acta vol 1802 no 1 pp 11ndash19 2010
[33] M Manczak T S Anekonda E Henson B S Park J Quinnand P H Reddy ldquoMitochondria are a direct site of A120573 accumu-lation in Alzheimerrsquos disease neurons implications for free rad-ical generation and oxidative damage in disease progressionrdquoHuman Molecular Genetics vol 15 no 9 pp 1437ndash1449 2006
[34] P F Pavlov C H Petersen E Glaser and M AnkarcronaldquoMitochondrial accumulation of APP and A120573 significancefor Alzheimer disease pathogenesisrdquo Journal of Cellular andMolecular Medicine vol 13 no 10 pp 4137ndash4145 2009
[35] C Schmidt E Lepsverdize S L Chi et al ldquoAmyloid precursorprotein and amyloid 120573-peptide bind to ATP synthase andregulate its activity at the surface of neural cellsrdquo MolecularPsychiatry vol 13 no 10 pp 953ndash969 2008
[36] K Johar A Priya and M T T Wong-Riley ldquoRegulation ofNa+K+-ATPase by nuclear respiratory factor 1 implication inthe tight coupling of neuronal activity energy generation andenergy consumptionrdquo The Journal of Biological Chemistry vol287 no 48 pp 40381ndash40390 2012
[37] J Astrup P M Sorensen and H R Sorensen ldquoOxygen andglucose consumption related to Na+-K+ transport in caninebrainrdquo Stroke vol 12 no 6 pp 726ndash730 1981
[38] M Erecinska and I A Silver ldquoATP and brain functionrdquo Journalof Cerebral Blood Flow and Metabolism vol 9 no 1 pp 2ndash191989
[39] MMata D J Fink H Gainer et al ldquoActivity-dependent energymetabolism in rat posterior pituitary primarily reflects sodiumpump activityrdquo Journal of Neurochemistry vol 34 no 1 pp 213ndash215 1980
[40] J Kang H-G Lemaire A Unterbeck et al ldquoThe precursor ofAlzheimerrsquos disease amyloid A4 protein resembles a cell-surfacereceptorrdquo Nature vol 325 no 6106 pp 733ndash736 1987
[41] C L Masters and D J Selkoe ldquoBiochemistry of amyloid 120573-protein and amyloid deposits in Alzheimer diseaserdquoCold SpringHarbour Perspectives in Medicine vol 2 no 6 Article IDa006262 2012
[42] A Lai S S Sisodia and I S Trowbridge ldquoCharacterization ofsorting signals in the beta-amyloid precursor protein cytoplas-mic domainrdquo The Journal of Biological Chemistry vol 270 no8 pp 3565ndash3573 1995
[43] A I Placido C M F Pereira A I Duarte et al ldquoThe role ofendoplasmic reticulum in amyloid precursor protein processingand trafficking implications for Alzheimerrsquos diseaserdquo Biochim-ica et Biophysica ActamdashMolecular Basis of Disease vol 1842 no9 pp 1444ndash1453 2014
Oxidative Medicine and Cellular Longevity 13
[44] C Kaether P Skehel and C G Dotti ldquoAxonal membraneproteins are transported in distinct carriers a two-color videomicroscopy study in cultured hippocampal neuronsrdquoMolecularBiology of the Cell vol 11 no 4 pp 1213ndash1224 2000
[45] A Kamal A Almenar-Queralt J F LeBlanc E A Robertsand L S B Goldstein ldquoKinesin-mediated axonal transportof a membrane compartment containing 120573-secretase andpresenilin-1 requires APPrdquo Nature vol 414 no 6864 pp 643ndash648 2001
[46] R G Perez S Soriano J D Hayes et al ldquoMutagenesis identi-fies new signals for 120573-amyloid precursor protein endocytosisturnover and the generation of secreted fragments includingA12057342rdquo The Journal of Biological Chemistry vol 274 no 27 pp18851ndash18856 1999
[47] D G Cook M S Forman J C Sung et al ldquoAlzheimerrsquos A120573(1ndash42) is generated in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo Nature Medicine vol 3 no 9 pp1021ndash1023 1997
[48] A S C Chyung B D Greenberg D G Cook R W Domsand V M-Y Lee ldquoNovel 120573-secretase cleavage of 120573-amyloidprecursor protein in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo The Journal of Cell Biology vol138 no 3 pp 671ndash680 1997
[49] RW-Y Choy Z Cheng and R Schekman ldquoAmyloid precursorprotein (APP) traffics from the cell surface via endosomesfor amyloid 120573 (A120573) production in the trans-Golgi networkrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 30 pp E2077ndashE2082 2012
[50] S A Small and S Gandy ldquoSorting through the cell biologyof Alzheimerrsquos disease intracellular pathways to pathogenesisrdquoNeuron vol 52 no 1 pp 15ndash31 2006
[51] M Goedert ldquoAlzheimerrsquos and Parkinsonrsquos diseases the prionconcept in relation to assembled A120573 tau and 120572-synucleinrdquoScience vol 349 no 6248 2015
[52] I Benilova E Karran and B De Strooper ldquoThe toxic A120573oligomer and Alzheimerrsquos disease an emperor in need ofclothesrdquo Nature Neuroscience vol 15 no 3 pp 349ndash357 2012
[53] E H Corder A M Saunders W J Strittmatter et al ldquoGenedose of apolipoprotein E type 4 allele and the risk of Alzheimerrsquosdisease in late onset familiesrdquo Science vol 261 no 5123 pp 921ndash923 1993
[54] S Brunholz S Sisodia A Lorenzo C Deyts S Kins and GMorfini ldquoAxonal transport of APP and the spatial regulationof APP cleavage and function in neuronal cellsrdquo ExperimentalBrain Research vol 217 no 3-4 pp 353ndash364 2012
[55] G Thinakaran and E H Koo ldquoAmyloid precursor proteintrafficking processing and functionrdquo The Journal of BiologicalChemistry vol 283 no 44 pp 29615ndash29619 2008
[56] T Saitoh M Sundsmo J-M Roch et al ldquoSecreted formof amyloid beta protein precursor is involved in the growthregulation of fibroblastsrdquo Cell vol 58 no 4 pp 615ndash622 1989
[57] M PMattson B Cheng A R Culwell F S Esch I Lieberburgand R E Rydel ldquoEvidence for excitoprotective and intraneu-ronal calcium-regulating roles for secreted forms of the 120573-amyloid precursor proteinrdquo Neuron vol 10 no 2 pp 243ndash2541993
[58] K J Lee C E H Moussa Y Lee et al ldquoBeta amyloid-independent role of amyloid precursor protein in generationandmaintenance of dendritic spinesrdquoNeuroscience vol 169 no1 pp 344ndash356 2010
[59] H Meziane J-C Dodart C Mathis et al ldquoMemory-enhancingeffects of secreted forms of the 120573-amyloid precursor proteinin normal and amnestic micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no21 pp 12683ndash12688 1998
[60] P R Turner K OrsquoConnor W P Tate and W C AbrahamldquoRoles of amyloid precursor protein and its fragments inregulating neural activity plasticity and memoryrdquo Progress inNeurobiology vol 70 no 1 pp 1ndash32 2003
[61] D Beher L Hesse C L Masters and GMulthaup ldquoRegulationof Amyloid Protein Precursor (APP) binding to collagen andmapping of the binding sites on APP and collagen type Irdquo TheJournal of Biological Chemistry vol 271 no 3 pp 1613ndash16201996
[62] S L Sabo A F Ikin J D Buxbaum and P Greengard ldquoTheamyloid precursor protein and its regulatory protein FE65 ingrowth cones and synapses in vitro and in vivordquo The Journal ofNeuroscience vol 23 no 13 pp 5407ndash5415 2003
[63] D H Small V Nurcombe G Reed et al ldquoA heparin-bindingdomain in the amyloid protein precursor of Alzheimerrsquos diseaseis involved in the regulation of neurite outgrowthrdquoThe Journalof Neuroscience vol 14 no 4 pp 2117ndash2127 1994
[64] T G Williamson S S Mok A Henry et al ldquoSecreted glypicanbinds to the amyloid precursor protein of Alzheimerrsquos disease(APP) and inhibits APP-induced neurite outgrowthrdquoThe Jour-nal of Biological Chemistry vol 271 no 49 pp 31215ndash312211996
[65] T Ondrejcak I Klyubin N-W Hu A E Barry W K Cullenand M J Rowan ldquoAlzheimerrsquos disease amyloid 120573-protein andsynaptic functionrdquo NeuroMolecular Medicine vol 12 no 1 pp13ndash26 2010
[66] D Puzzo L Privitera and A Palmeri ldquoHormetic effect ofamyloid-beta peptide in synaptic plasticity and memoryrdquo Neu-robiology of Aging vol 33 no 7 pp 1484e15ndash1484e24 2012
[67] C M F Pereira ldquoCrosstalk between endoplasmic reticulumstress and protein misfolding in neurodegenerative diseasesrdquoISRN Cell Biology vol 2013 Article ID 256404 22 pages 2013
[68] R-W Shin T C Saido M Maeda and T Kitamoto ldquoNovel 120572-secretase cleavage ofAlzheimerrsquos amyloid120573 precursor protein inthe endoplasmic reticulum of COS7 cellsrdquoNeuroscience Lettersvol 376 no 1 pp 14ndash19 2005
[69] E A Schon and E Area-Gomez ldquoMitochondria-associatedER membranes in Alzheimer diseaserdquo Molecular and CellularNeuroscience vol 55 pp 26ndash36 2013
[70] R Rizzuto P Pinton W Carrington et al ldquoClose contacts withthe endoplasmic reticulum as determinants of mitochondrialCa2+ responsesrdquo Science vol 280 no 5370 pp 1763ndash1766 1998
[71] S E Logue P Cleary S Saveljeva and A Samali ldquoNewdirections in ER-stress induced cell deathrdquoApoptosis vol 18 no5 pp 537ndash546 2013
[72] J P Decuypere KWelkenhuyzen T Luyten et al ldquoIns(145)P3receptor-mediated BI-1 Ca2+ signaling and autophagy induc-tion are interrelatedrdquo Autophagy vol 7 pp 1472ndash1489 2011
[73] K S Vetrivel and G Thinakaran ldquoMembrane rafts in Alz-heimerrsquos disease beta-amyloid productionrdquo Biochimica et Bio-physica Acta vol 1801 no 8 pp 860ndash867 2010
[74] H Urra E Dufey F Lisbona D Rojas-Rivera and C HetzldquoWhen ER stress reaches a dead endrdquo Biochimica et BiophysicaActa (BBA) Molecular Cell Research vol 1833 no 12 pp 3507ndash3517 2013
14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 3
P3A
ICD
AIC
D120574-Secretase120574-Secretase
APP-CTF83
120572-Secretase
BACE1120573-Secretase
APPsAPP-120573 sAPP-120572
APP-CTF99
Amyloidogenic NonamyloidogenicA120573
A120573A12057340A
12057342
Figure 1 A diagram of amyloid precursor protein (APP) processing pathway The transmembrane protein APP (membrane indicated inblue) can be processed by two pathways the nonamyloidogenic 120572-secretase pathway and the amyloidogenic 120573-secretase pathway In thenonamyloidogenic pathway 120572-secretase cleaves in the middle of the 120573-amyloid (A120573) region (red) to release the soluble APP-fragment sAPP-120572 The APP C-terminal fragment 83 (APP-CTF83 120572CTF) is then cleaved by 120574-secretase to release the APP intracellular domain (AICD) andP3 fragment In the amyloidogenic pathway 120573-secretase cleaves APP to produce the soluble fragment sAPP-120573 APP-CTF99 (120573CTF) is thencleaved by 120574-secretase to produce A120573
40 A12057342 and AICD Adopted from [179]
of APP by four different human 120574-secretase complexes eachacting at more than one 120573CTF site (120576- 120577- and 120574-) leads toformation of several A120573 (A120573
37ndash43) with A12057340 and A12057342 beingpredominant species Finally three end products are formed(sAPP-120573 A120573 and amyloid precursor protein intracellulardomain AICD) The key neuron 120572-secretase (ADAM10)cleaves APP inside A120573 polypeptide chain to 120572CTF as theintermediate so after subsequent 120574-secretase action on120572CTFthree nonpathogenic fragments are formed (sAPP-120572 P3fragment and AICD) (Figure 1)
As aforementioned FAD is caused by mutations in APPand PSEN genes located on chromosomes 21 and 14 respec-tively but the incidence of AD is also higher in dominantlyinherited duplications of the APP locus in elderly individualswith Downrsquos syndrome (trisomy of the 21st chromosome)pointing to important role played by APP and A120573 in ADMutations in APP located near the 120573-secretase cleavage siteincrease production ofA120573 whereas those near the 120574-secretasecleavage site result in an increased ratio of A120573
42to A120573
40
[52] The 1205764 allele of apolipoprotein E is the major risk factorfor SAD Thus this particular APOE gene polymorphismincreases disease risk in a dose-dependentmanner and lowersthe age of onset as shown by Corder et al [53] One copyof APOE4 increases the risk of AD about fourfold (com-pared with the more common APOE1205763APOE31205763 genotype)whereas two copies of APOE4 increase the risk of AD about12-fold The mechanism by which the amino acid differencebetweenAPOE3 andAPOE4 increases the risk ofAD remainsto be established
Widespread occurrence ofAPP andA120573 in nervous systembrought about the assumption that both components mightplay physiological roles More than few possible conceptshave emerged some validated by experimental data TheAPP protein overexpression led to enhanced survival andgrowth of some cell types [54 55] Furthermore secreted
forms of APP (APPss sAPP120572 and sAPP120573) were antiapoptotic[56] and have a potent neuroprotective action in culturedrat hippocampal and septal neurons and in human corti-cal neurons [57] APPs
695and APPs
751protected neurons
against hypoglycemic damage and the neuroprotection wasabolished by antibodies to a specific region common toboth APPs
695and APPs
751 Thus APPss may normally play
excitoprotective and neuromodulatory roles AccordinglyAPP was shown to stimulate axon branching and the main-tenance and formation of synapses neuronal survival andneuritic outgrowth [58ndash60] APP protein is highly expressedin axons and interacts with extracellular matrix components[61ndash64] Similar to APP A120573 was demonstrated to play aphysiological role in synaptic plasticity as minute quantitiesof the peptide stimulated neurons and enhanced the releaseof neurotransmitter [65 66] Everything can changewhen thethings go awry
4 Perturbations in ER
In healthy cells including neurons ER is a fundamentalorganelle for protein quality control in the secretory pathwaywhich prevents protein aberrant folding and aggregation[67] A bulk of evidence shows the importance of ERin APP maturation and processing With regard to APPintracellular processing both secretases (120572- and 120573-) havebeen identified in the ER together with 120574-secretase whichis present in mitochondria-associated membrane (MAM)subcompartment [48 68] This distinctive intracellular lipid-raft-like structure is involved in cholesterol and phospholipidmetabolism Ca2+ metabolism and mitochondrial dynamicsand becomes markedly augmented in AD [69] MAM isresponsible for the communication between the ER and themitochondria with efficient transfer of Ca2+ from the ERto mitochondria supporting metabolic functions and cell
4 Oxidative Medicine and Cellular Longevity
ER membrane
6
ERAD
Proteasome26S
Ca2+
Ca2+
IP3R
VDAC
MCU
IMMOMM
ATP
GRP
75
MFN
1
MFN
2M
FN2
MFN
2
Sig1R
UPR
7
Mitochondria
Nucleus CHOP ATF6
XBP1ATF4
Gene expression
3
Transcriptionfactors
2
IRE1120572 PERK ATF6
1 GRP78
Chaperones
ER Misfolded proteins
Autophagosome
5
4
ERGolgitraffic
Golgi cisternae
Antioxidant responseApoptosisERAD and autophagyER chaperonesER and Golgi biosynthesis
Figure 2 ER stress triggered by misfolded proteins in several neurodegenerative diseases Abnormal conformations of the proteins APPA120573 and tau are implicated in the pathogenesis of AD Alterations in the function of ER chaperones and UPR-related components ERADERGolgi trafficking and ER-to-mitochondria Ca2+ transfer have been suggested as underlying mechanisms of ER stress triggered by thesedisease-associated proteins These proteins can accumulate and aggregate at the ER and their stable interaction with ER chaperones such asGRP78BiP and PDI may trap ER chaperones altering protein folding with concomitant ER stress In addition these proteins can lead to theoxidative modification of the active site of PDIs by nitrosylation leading to their enzymatic inactivation Furthermore some of these proteinsalter the activity of the UPR stress sensors (IRE1120572 PERK and ATF6) as well as the activitylevels of downstream signaling mediators andtranscription factors including cleaved ATF6 ATF4 and spliced XBP1 As a result genes implicated in autophagy and ERAD antioxidantresponse ER chaperones and organellersquos biosynthesis are upregulatedMoreover these proteins block the exit of vesicles from the ER and alterthe trafficking between ER andGolgi of properly folded proteinsThe cellular responses controlled byUPR transcription factors including themodulation of autophagy mediated degradation of protein aggregates become compromised Disease-related proteins can also interact withERAD components precluding the translocation of ERAD substrates from the ER to the cytosol leading to the accumulation of abnormallyfolded proteins at the ER Finally Ca2+ released from the ERmainly through the IP
3R and its transfer tomitochondria can be impaired in the
presence of disease-related proteins leading to mitochondrial Ca2+ overload and activation of apoptotic cell death pathways AD Alzheimerrsquosdisease ATF6 activating transcription factor 6 ATF4 activating transcription factor 4 ER endoplasmic reticulum ERAD endoplasmic-reticulum-associated protein degradation IP
3R inositol triphosphate receptor IRE1120572 inositol-requiring enzyme 1 alpha PERK protein
kinase R- (PKR-) like ER kinase UPR unfolded protein response XBP1 Xbox binding protein 1 Adopted from [67]
viability [70] The molecular bridges between ER inositol145-triphosphate receptor (IP
3R) and the voltage-dependent
anion channels in the outer mitochondrial membrane arebrought together through the cytosolic chaperone glucose-regulated protein 75 (GRP75) (Figure 2) Additionally thedynamin-relatedGTPasemitofusin 2 (Mfn2) proteins locatedon the ER intermingle with Mfn1 or Mfn2 on mitochondria
to tighten the connection The distance between ER andmitochondria controlled by the phosphofurin acidic clustersorting protein 2 (PACS-2) of ER and the dynamin-relatedGTPase protein 1 (Drp1) is crucial for cell survival as eithertoo long (lack of Ca2+ flux) or too short distance (Ca2+ over-load) might lead to apoptosis [71] Alternatively impairedmitochondrial bioenergetics with reduced cellular ATP
Oxidative Medicine and Cellular Longevity 5
levels stimulate autophagyThe molecular mechanism of ER-mediated autophagy is accurately regulated by Beclin 1 as wellas ER membrane bound protein Bax inhibitor 1 (BI-1) Bothproteins are capable of promoting autophagy through IP
3R-
dependent mechanism [72]
5 APP Processing and ER Stress Response
The nature of APP processing is determined by the com-position of membrane with cholesterol rich lipid rafts asthe site of amyloidogenic cleavage [73] Consequently onemight expect that fate of APP is at least partly establishedby the representation of lipid rafts and possible access to 120573-versus120572-secretase Irrespective of the type of cleavage there isone underestimated biochemical step in ER that might bringabout damaging upshot It is apparently the effectiveness ofER situated N-glycosylation which makes the APP moleculesuitably folded If misfoldedmalfolded protein(s) accumu-late in the ER complex cascade of reactions known as theunfolded protein response (UPR) is triggered with the so-called endoplasmic reticulum stress response (ERS) Today itis widely accepted that during UPR the ER sensors proteinkinase R- (PKR-) like ER kinase (PERK) activating proteinkinase 6 (ATF6) and inositol-requiring enzyme 1 alpha(IRE1120572) are freed fromGRP78BiP protein repression whichhereafter become activated Without going into details ofindividual sensor action a number of reactions occur at boththe genomic and cytoplasmic level with selective degradationof mRNAs encoding protein(s) with abnormal folding andinhibition of protein translation except for genes importantfor UPR redox homeostasis energy metabolism and pro-tein folding [67] On the one hand PERK phosphorylateseukaryotic initiation factor 2 alpha (eIF2120572) to stop entrance ofmethionyl-tRNA to the ribosome on the other hand it allowstranslation of activating transcription factor 4 (ATF4) geneIRE1120572 sets off alternative splicing of Xbox binding protein 1(XBP1) transcript leading to activation of the transcriptionfactor liable to stimulate ERGolgi biogenesis and formationof proteins involved in endoplasmic-reticulum-associatedprotein degradation known as ERAD (Erdj4 p58IPK EDEMRAMP-4 PDI-P5 and HEDJ for details see [67]) FinallyATF6 is activated in the Golgi complex through proteolyticcleavage and translocates to nucleus where it cooperates withXBP1 in upregulation of chaperones and ERAD-related genes[43] In principle UPR is activated to restore ER homeostasisand stop the accumulation of aberrantly formed protein(s)but if the strength of ER stress is unbearable (meaning thatit cannot be compensated by UPR) there is a path to activateapoptosis Among other routes the most central role isplayed by the major proapoptotic transcription factor CEBPhomologous protein CHOPgrowth arrest and DNA damageinduced gene 153 GADD153 It downregulates the antiapop-totic protein Bcl-2 and upregulates proapoptotic Bax and Bak[74] CHOPGADD153 leads to excessive production of reac-tive oxygen species (ROS) within ER subsequent depletionof reduced glutathione (GSH) and Ca2+ flux from the ER tocytoplasm through the IP
3R [75] One hitherto unresolved
issue is the physiologic importance of APP and GRP78BiPinteraction disclosed in coprecipitation study carried out by
Yamamoto et al [76] Bulk ofAPP associatedwithGRP78BiPwas immature protein Given that GRP78BiP expressionlevels declined in samples of brain tissue obtained fromFAD patients as demonstrated by Katayama et al [77] fewdistinct scenarios are possible First binding to GRP78BiPsuggests ER accumulation of immature APP Second APPinteraction with GRP78BiP is a noticeable sign of UPRwhich is further validated by downregulation of GRP78BiPin FAD patients Finally retention of APP in ER is mostlikely a result of amyloidogenic APP processing and as such itprobably occurs in MAM the cholesterol enriched domainsER-mitochondria crossing point is therefore of particularinterest in deciphering the links between the ER placed APPprocessing UPR and resulting cellular responses such asautophagy apoptosis and inflammatory reaction observed inAD (Figure 2)
6 Pathology of Tau Protein
Microtubule-associated protein tau controls assembly andprevents microtubules from severing Microtubular networkis fundamental component of cytoskeleton essential forintracellular transport of secretory vesicles and organelles(ie mitochondria) Glycogen synthase kinase 3 beta- (GSK-3120573-) targeted hyperphosphorylation of tau causes this proteinto dissociate frommicrotubules Consequently microtubulesbecome fragmented and microtubule-dependent transportsystem fails Furthermore hyperphosphorylated tau (P-tau)is prone to form oligomers and toxic filaments known asNFTs or tauopathy [78] A variety of tau conformers werereported to exist pointing to different tauopathies capa-ble of self-propagation [79 80] Interestingly intracellulartau inclusions define AD as clinical symptoms of diseaseobserved when tauopathy is abundant together with intra-cellular A120573 deposits in neocortex [51 81 82] People withabundant A120573 plaques but no or only a few neurofibrillarylesions do not have AD Clinicopathological correlationstudies have been crucial to generate hypotheses about thepathophysiology of the disease by establishing the fact thatthere is a continuum between ldquonormalrdquo aging and ADdementia and that the amyloid plaque buildup occurs primar-ily before the onset of cognitive deficits while neurofibrillarytangles neuron loss and particularly synaptic loss parallel theprogression of cognitive decline [83] Thus misfolded pro-teins and descendant toxic filaments with a number of inter-mediates are critical for manifestation of AD as fibrillogenicAPP processing is not enough for onset of disease Althoughthe molecular mechanism of tauopathy is not deciphered infull recent reports suggest ER stress as the starting point[84 85] This idea is substantiated by the elevated levels ofERS and UPR markers together with P-tau and GSK-3120573 inbrains affected by AD [86] Therefore on the one hand theincidence of UPR is strongly correlated with the presence ofNFTs on the other hand aggregation of P-tau induces ERSwith resultant UPR Some lines of evidence confirmed UPRactivation near the beginning of NFTs formation and pointto the functional link between malformed tau protein andUPRThe in vitro experiments with phosphatase 2A inhibitoror phosphorylation activator demonstrated enhanced P-tau
6 Oxidative Medicine and Cellular Longevity
formation in neurons in concert with the increased levelsof PERK eIF2120572 and XBP1 transcript apparent markers ofUPR [87] Furthermore GRP78BiP was found to encouragetau phosphorylation through facilitated substrate capture byGSK-3120573 [88] GSK-3120573 seems to play dual role first this kinaseprotects neurons from apoptosis as P-tau accumulation isstrong molecular signal to trigger UPR with subsequentautophagy Second UPR raises GSK-3120573 activity throughlysosomal degradation of inactive GSK-3120573 (P-Ser9-GSK-3120573)To sum up ERS and UPR are important molecular machinesused to prevent cell viability turned on by tauopathy
7 Oxidative Stress in ER
Sacs and tubes of ER delineate the compartment where thenewly synthesized proteins undergo maturation to nativestate Native state indicates properly folded fully functionalprotein Important reactions essential for protein folding ofunbranched polypeptide chains include amino acids oxida-tion and glycosylation As a result redox homeostasis inER is shifted to oxidative state so as to promote disulfidebond formation between adjacent cysteines Oxidation ofsulfhydryl groups required to make disulfide bonds is con-trolled by ER oxidase 1120572 (ERO1120572) Next disulfide bondscould be subject to posttranslational modification disulfideexchange by protein disulfide isomerase (PDI) PDI is ableto correct mispaired thiol residues by catalyzing the breakageand formation of correct disulfide bonds These enzymes arefundamental for protein folding Oxidation allows twistingof proteins which is followed by N-glycosylation andor C-and O-mannosylation Protein N-glycosylation in eukary-otes is a complex process divided into several steps Firstthere is a ldquocall forrdquo carrier lipids (polyisoprenyl phosphatessuch as dolichyl phosphates) the membrane lipids knownto function as glycosyl transporters In mammalian cellsthe limiting substrate for dolichol biosynthesis is geranyl-geraniol (GGOH) of mevalonate pathway Dolichols thelongest aliphatic molecules synthesized in animal cellshave 18ndash21 120572-isoprene saturated units (C90ndash105) criticalfor their recognition by the enzymes (glycosyltransferases)that glucosylate dolichyl phosphates [89] Once dolichylmonophosphates (Dol-P) are formed in the ER membranethe precursor oligosaccharide donor (GLc
3Man9GLCNAc
2-
P-P-dolichol) for protein N-glycosylation can be synthesizedon the lumenal leaflet of ER First three sugar intermediatesare produced (Man-P-Dol Glc-P-Dol GlcNAc-P-P-Dol andMan5GlcNAc-P-P-Dol) on the cytoplasmic leaflet of the
ER Next enzyme flippases mediate transbilayer movementof the aforementioned intermediates to lumenal side ofER where conversion to Glc
3Man9GlcNAc
2-P-P-Dol could
be completed Glc3Man9GlcNAc
2-P-P-Dol is also used for
biosynthesis of glycosylphosphatidylinositol (GPI) anchorsTaken together lipid-mediated glycosylation plays a vital
role in the appropriate protein folding and intracellulartranslocation of N-linked glycoproteins [90] Likewise it isimportant for protein O- and C-mannosylation and GPIanchorage Moreover Dol-P availability in the ER is the rate-limiting factor in the production of glycolipid intermediatesand N-glycosylation
8 ER Stress and Apoptosis
It was shown that during ERS the ER resident proapop-totic cysteine protease known as caspase (caspase-12 in ratcaspase-4 in humans) is activated through cleavage As aresult caspase cascade is started via caspase-9 that in turnstimulates effector caspase-3 [91 92] The central role playedby ER in programmed cell death is achieved by PERK branchwhere ATF4 induces the expression of CHOPGADD153which represses antiapoptotic Bcl-2 family proteins andsimultaneously shuffles ER membrane Bax and Bak proteinsinto outer mitochondrial membrane Consequently poresare formed to leak the components of apoptosome frommitochondrial intermembrane space [93] Another impor-tant mechanism of ERS-induced apoptosis is led by Ca2+-dependent ERO1120572-IP
3R pathway where ERO1120572 collaborates
with IP3R in Ca2+ efflux from ER to mitochondria via MAM
[75 94] Accordingly Ca2+ influx facilitates cytochrome 119888release frommitochondria besides cytochrome 119888 can bind toER IP
3R and the complex amplifies the apoptotic signal in a
feedforward manner [95] Last but not least ERS-associatedapoptotic programme is set off by c-Jun N-terminal kinase(JNK) as the effect of IRE1120572 complexed with TNF-receptor-associated factor 2 (TRAF2) activation of apoptosis-signal-regulating kinase 1 (ASK1) [96 97]
What does ERS drive to induce apoptotic death inneurons Actually many reports indicate that APP and A120573 aswell as hyperphosphorylated tau have been shown to blockmitochondrial transport which results in impaired energystorage and oxidative stress [98ndash101] Indeed accumulationof APP A120573 and NFTs in mitochondria led to reducedactivity of some enzymes involved in substrate oxidation(tricarboxylic acid cycle) electron transport chain (ETC)and ATP synthase as well as severely diminishing import ofnuclear-encoded proteins [26 27 102ndash104] One may ask ifthere is any additional link between ER and mitochondriaother than MAM which could account for apoptotic signalThough not directly ER significantly contributes to oxidativestress in mitochondria of AD-affected subjects
9 Abnormal APP Processing and TraffickingCulminate in ER Pathology of AD
From the morphological point of view as neurons arehighly specialized cells soma dendrites and neurites areconsiderably distinct structures Proteins needed by thesecompartments are delivered via microtubules once proteinshave suitable sorting signals (ie APP trafficking from ERto plasma membrane is associated with several posttrans-lational modifications with oxidation and N-glycosylation)Additionally the Golgi apparatus follows ER in subsequentAPP adjustment (O- and N-glycosylation phosphorylationand sulphonation) [42] Any inaccurate alteration of the APPmolecule is potentially hazardous as protein final destiny ismissed causing its retention in ER or trans-Golgi network(TGN) In addition other unusual settings for APP arepossible as this large protein has few signal sequences hiddenwhen APP is correctly folded In the cells transfected withAPP this protein enters coat protein complex I (COPI)
Oxidative Medicine and Cellular Longevity 7
vesicles and undergoes retrograde transport from cis endof the Golgi complex back to the ER [76] Such responsecauses accumulation of APP in the tubulocisternal ER systemtogether with aberrant intracellular translocation of theprotein Interestingly the issue whether APP is subject toretrograde transport with successive fibrillogenic processingbecause of UPR and its interaction with GRP78BiP is notclear as the observations are inconsistent [77] Nonethelessaccumulation of misfoldedmalfolded proteins in the ERsuggests disorganized process of posttranslational change Asanticipated the accretion of proteins of anomalous patternsignals ERS and UPR followed by increased vulnerability toapoptotic cell death Prior to decay however APP proteinlevels in the ER lessen by dint of cleavage with 120573- and120574-secretase [105 106] Products of this cleavage (sAPP120573A120573 and AICD fragment) all appreciably influence neuronalsurvival most likely through nonnative form of the APPsubstrate
10 APP A120573 and NFTs MarkMitochondria as Targeted in AD
Mitochondrial import of A12057340
and A12057342
peptides throughthe translocase TOM complex was blocked by preincubationof isolated mitochondria with antibodies raised againstTOM proteins (TOM20 TOM40 and TOM70) [107]Neither VDAC inhibition with antagonist antibodies norinhibition of mitochondrial permeability transition pores(MPTP) or fall of mitochondrial membrane potential(MMP) affected uptake of A120573 [17] With regard to APPelegant study performed by Anandatheerthavarada andhis colleagues [30] revealed that C-terminal truncatedAPP (lacking A120573) targets mitochondria in cholinergicGABAergic dopaminergic and glutamatergic neurons ofAD brain by clogging up mitochondrial protein translocasecomplex TOM40TIM22 Authors propose that occlusionof translocase is followed by blunted import of nuclear-encoded proteins vitally important for energy homeostasisMitochondrial APP protein transmembrane orientationindicates NH
2-terminal inside in contact with translocase
whereas COOH-terminal is facing cytoplasmic side NH2-
terminus has mitochondrial signal sequence Astonishinglymitochondrial APP molecules were nonglycosylated givingrise to speculation that protein molecules that arrivedat mitochondria have not achieved molecular maturity[31] The accumulation of nonglycosylated APP species inmitochondrial import channels of AD brain was directlyrelated to decreased mitochondrial functions as validatedby the decline in cytochrome 119888 oxidase activity (complexIV) and elevated levels of H
2O2 Furthermore in AD
brain the mitochondrial accumulation of nonglycosylatedAPP went along with a corresponding reduction in plasmamembrane-associatedAPP It suggests that ADbrain hasAPPprocessing and trafficking severely affected by incompleteN-glycosylation ensuing ER protein accumulation andexposure of the cryptic mitochondrial targeting signalfor assisting chaperone proteins Several lines of evidenceindicate that not fully formed proteins are phosphorylatedand bind to the cytosolic proteins required for movement
from ER to mitochondria [108ndash110] Moreover ERS andUPR in AD seem to be incompetent and inefficient inelimination of malfolded proteins In addition to APPalso A120573 was frequently reported to occupy mitochondriaalthough its origin and mechanism of mitochondriatargeting mostly remain unexplored (except for involvementof protein translocase TOM complex) The possibilityof mitochondrial A120573 generation has to be ruled out asmembrane orientation of arrested APP does not allow accessto 120574-secretase (120574-secretase activity in mitochondria wasdetected by independent study) [111] Probably A120573 speciesis derived from APP prior to its translocation (ER) soA120573 may be transported to mitochondria independentlyof APP Collectively observations showing mitochondrialpresence of APP A120573 and tau in aberrant configuration pointtoward the anomaly of protein folding at the level of ER andGolgi apparatus Nonglycosylated molecule of APP suggestsdefective transfer of sugar core from dolichyl phosphate(s)and further modifications such as O- and N-glycosylationsOne may admit that the lack of glycosyl residue bringsabout pathologic processing and trafficking of APP and itsfragments In point of fact dolichol derivatives mixture ofpolyprenols (acyclic isoprenoid alcohols) known as Ropren(Solagran Limited Melbourne Australia) commerciallyused to treat liver diseases were tested in the treatment ofAD in two separate trials conducted in 2005 and 2006 withpromising results [httpwwwasxcomauasxpdf20071119pdf315x5nh4hm8wv7pdf httpwwwasxcomauasxpdf20070221pdf3111ztwcbqzkk9pdf] Further studies areurgently needed to test how important the glycosylationprocess is in the pathogenesis of AD Dolichols areobtained from geranylgeraniol (GGOH) and the latter is anintermediate of mevalonate pathway GGOH is a commonsubstrate for dolichol and ubiquinone synthesis but it isalso necessary for protein prenylation As both GGOHand farnesol (FOH) are engaged in protein prenylationmore concern should be laid on the importance of thesecompounds in AD pathogenesis Observations demonstrat-ing mitochondrial relocalization of other proteins withoutposttranslational modification seemingly point to increasedmitochondrial targeting of immature molecules resultingin mitochondrial dysfunction and acceleration of diseaseprogression [34 112ndash114]
11 Oxidative Stress in Mitochondria ofAD Brains
Normal physiological functions of APP are thought to beinvolved in the stabilizing contact points between synapsesand maintaining mitochondrial functions [60 115] Mito-chondrial dysfunction was often observed regardless of theexperimental model used to study AD [8 29 116ndash119]It includes defects in oxidative phosphorylation decreasedATP decreased membrane potential increased productionof ROSRNS and perturbation in mitochondrial fusion andfission [15 30 31 115 120ndash122] Hyperphosphorylated tau wasalso reported to impair mitochondrial functions [123] Usingproteomic approach the strongest defects of the respiratorycapacity were observed mainly at complexes I IV and ATP
8 Oxidative Medicine and Cellular Longevity
synthase (complex V) at both protein and activity level [124]While APP A120573 and hyperphosphorylated tau are potentinhibitors of mitochondrial import of nuclear-encoded pro-teins apparently the pathology of each leads to metabolicharm in different way In freshly isolated mitochondriafrom AD brains the APP inhibited mitochondrial importof cytochrome 119888 oxidase (COX) subunits IV and Vb [31]Dysfunction of COX increases ROS production (incompletereduction of oxygen molecules) reduces energy stores anddisturbs energy metabolism Accordingly in AD patientsdeficiency of COX was found in brains and platelets [119125 126] Similar to APP A120573 was found in mitochondriaof transgenic mice and cellular and human AD models [1719 33 103 127ndash130] At present it is not clear whether theobservedmitochondrial toxicity is due toAPP orA120573 orNFTsaccumulation Anyway some regularity is observed withrespect to most affected components of respiratory chainNADH-ubiquinone oxidoreductase (complex I) activity isreduced to the utmost by hyperphosphorylated tau whereasdecreased activity of cytochrome 119888 oxidase (complex IV)that resulted in mitochondrial dysfunction was observedduring A120573 and APP accumulation [14 15 31 32 34]Concomitantly rise in the activity of antioxidant enzymesmanganese superoxide dismutase (Mn-SOD) and catalase(CAT) was demonstrated in response to elevated levelsof free radicals including superoxide anion radical (O
2
∙minus)hydroperoxyl radical (HO
2
∙) hydroxyl radical (OH∙) andnitric oxide radical (NO∙) [15] Consistent with observationsof chronic respiratory chain dysfunction and mitochondrialoxidative stress there are reports showing their contributionto tau pathology in AD [131] In any case free radicals thatoverride antioxidant defense react with a wide variety oforganic components causing lipid peroxidation to advancedlipid oxidation end products (ALE) cross-linking of proteinsnitrosylation of proteins and mutations in DNA Mitochon-drial circular DNA (mtDNA) of sim16 kbp is devoid of repairsystems meaning the buildup of lesions There are 37 geneslocated in mtDNA with those encoding protein subunits ofcomplex I (7) complex II (1) and complex IV (3) but notcomplex III Interestingly hallmarks of AD (APP A120573 andNFTs)mostly affectmembers of electron transfer chain (com-plexes I and IV) which rely exclusively on mitochondriallypredetermined subunits Maybe it is not simple coincidencebut the effect of inhibited import to mitochondria of nuclear-encoded subunits of complexes I and IV gives explanationfor toxicity of APP or A120573 or NFTs in mitochondria Thereare additional findings in mitochondria affected by AD suchas lower activity of pyruvate dehydrogenase (PDH) andoxoglutarate dehydrogenase (OGDH) [104 132] Inhibitionof OGDH the enzyme of tricarboxylic acid cycle minimizesthe NADH pool and electron number needed for ETCand mitochondrial membrane potential (Δ120595
119898) to create and
maintain proton gradient obligatory for ATP synthesisThereare also lines of evidence for direct inhibitory action of solubleoligomeric A120573 species on ABAD (A120573-binding alcohol dehy-drogenase) and internal membrane cyclophilin D (CypD)resulting in increased mitochondrial membrane permeabil-ity (MPTP) potentiated ROS production synaptic lossdiminished activity of mitochondrial respiration and finally
cell death [129 132ndash134] CypD knockout prevents mito-chondrial and neuronal perturbations and improves mito-chondrial function in Alzheimerrsquos disease mouse model [128135] It has to be emphasized that oxidativenitrosative stressaffects the fusion and fission process ofmitochondria Fusionthat is speedup by small GTPases mitofusins (Mfn1Mfn2)improves efficiency of mitochondrial respiration and ATPproduction Mitochondrial dynamics are severely imbal-anced in AD cases in favour of fission through elevatedexpression of the fission protein DLP1 (dynamin-like protein1) associatedwith nitrosative stress stirred up byA120573 [136 137]Collectively these observations indicate that toxic intracel-lular A120573
42-43 oligomers differ in action from extracellularaggregates found in amyloid plaques of AD brains [138](Figure 3)
12 Proteostasis in Mitochondria
Mitochondrial protein turnover grants the well-organizedreplacement of nonfunctional proteins into operational oneThe first task is attained through proteolytic degradation ofinner membrane and matrix proteins with local mitochon-drial proteases [139] or outer membrane proteins throughubiquitin-proteasome system [140] The second task is metby intramitochondrial protein synthesis but as almost 1500different nuclear-encoded proteins have to be importedto complete mitochondrial proteome a matter of capableimport is fundamental for the function of the organelleIn extreme cases of cellular injuries observed in neurode-generative diseases damaged mitochondria with extendedloss of the electrochemical potential are selectively removedby autophagy known as mitophagy [112] APP and A120573accumulation has also something to do with altered mito-chondrial dynamics as fission takes advantage of fusion withresultant dysfunction of mitochondria and neurons [137]AD neurons demonstrated selective mechanisms of prote-olytic clearance of oxidatively and nitrosatively modifiedproteins in mitochondria Insulin degrading enzyme (IDE)prevents formation of toxic insoluble fibrils from A120573 asit cleaves A120573 prior to aggregation [141ndash143] A novel zinc-metallopeptidase Presequence Protease (PreP) a member ofpitrilysin oligopeptidase family degrades either intramito-chondrially stored A120573
40or A120573
42proteinThis protease seems
to be highly sensitive to oxidative stress as disulfide bridgeformed between two proximal cysteine residues blocks itscatalytic activity [144ndash149] Another mitochondrial serineprotease HtrA2Omi occupies intermembrane space whereit can cleave APP locked up in protein translocase complex[121] Even though it is well established that HtrA2Omiis released to the cytosol to amplify apoptosis through thedegradation of antiapoptotic proteins and caspase activation[150ndash152] it is also implicated in proteolytic deletion ofmalfoldedAPP in ER [151] Fromknockout studies onmice itis obvious that HtrA2Omi plays a significant shielding roleas mice deficient in this protease exhibit neurodegenerativephenotype with weight loss and premature death [153]Taken together understanding the mechanisms of clearanceof the unwanted proteins including APP A120573 and tau isvital in an attempt to get rid of them from mitochondria
Oxidative Medicine and Cellular Longevity 9
VDAC
ANT
A120573
A120573
A120573
A120573
A120573
A120573
A120573A120573
A120573
A120573
A120573 CypD
CypD ProApCyt c Ca2+
IMM
OMM
IMSΔΨ
+
+
+
minus
minus
minus
ABAD
ROS
ROS
TIM
TOM
Proteinimport
NAD
NAD
NAD
NAD
H+
H+
H+
H+
H+
ADP
Mn-SOD
CuZn-SOD
mtDNA
V
Matrix
UCP
O2∙minus
O2∙minus
IV
III
II
I
O2
O2
DLP1Fis1
Mfn1Mfn2
Opa1
mHsp70
NADH
NADH
NADH
TCAcycle
PyruvateAcetyl-CoA
120572KGDH
120572-KG
PDH
Isocitrate
CitrateOAA
Malate
Fumarate
Succinyl-CoASuccinate
GDPGTP
FAD
FAOH
ATP
Figure 3 Amyloid-120573-related mitochondrial impairment Mitochondria were found to be the target for amyloid-120573 (A120573) which interactswith several proteins leading to mitochondrial dysfunction Indeed A120573 was found in the outer mitochondrial membrane (OMM) andinner mitochondrial membrane (IMM) as well as in the matrix The interaction of A120573 with the OMM affects the transport of nuclear-encoded mitochondrial proteins such as subunits of the electron transport chain complex IV into the organelle via the translocase of theouter membrane (TOM) import machinery Moreover A120573 disturbs the activity of several enzymes such as pyruvate dehydrogenase (PDH)and oxoglutarate dehydrogenase (OGDH) decreasing NADH reduction and the electron transport chain enzyme complex IV reducing theamount of hydrogen that is translocated from the matrix to the intermembrane space (IMS) thus impairing the mitochondrial membranepotential (MMP) Taken together these events cause abnormal mitochondrial electron activities leading to decreased complex V activityand so to a drop in ATP levels in addition to increasing reactive oxygen species (ROS) generation Moreover ROS induce peroxidationof several mitochondrial macromolecules such as mitochondrial DNA (mtDNA) and mitochondrial lipids contributing to mitochondrialimpairment in themitochondrial matrixThe complex of A120573 bound to binding alcohol dehydrogenase (ABAD) impairs the binding of NAD+to ABAD changes mitochondrial membrane permeability and reduces activities of respiratory enzymes inducing further ROS productionand leading to mitochondrial failure A120573 binding also activates Fis1 (fission protein) and promotes increased mitochondrial fragmentationthis increased mitochondrial fragmentation produces defective mitochondria that ultimately damage neurons Furthermore A120573 binding tocyclophilin D (CypD) enhances the protein translocation to the inner membrane favouring the opening of the mitochondrial permeabilitytransition pore formed by the adenine nucleotide translocator (ANT) and voltage-dependent anion channels (VDACs) Cyt 119888 cytochrome 119888DLP1 dynamin-like protein 1 PDH pyruvate dehydrogenase ProAp proapoptotic factors SOD superoxide dismutase TCA tricarboxylicacid TIM translocase of the inner membrane Adopted from [14]
Some hopes are associated with the application of at presentindefinite modulators of proteolytic activity HtrA2Omi isfor example activated by PTEN-induced putative kinase 1(PINK1) upon phosphorylation at Ser142 residue [154] Alter-natively accent has to be put on mitophagy of dysfunctionalmitochondria and mitochondriogenesis
13 Aging versus AD
Themost commonneurodegenerative disorder is representedby Alzheimerrsquos disease characterized by declining memoryreduced cognitive capacity and progressive dementia whichare often fatal to elderly individuals above 65 years of age
10 Oxidative Medicine and Cellular Longevity
It is ranked as the fourth leading cause of death in modernsocieties where average life span increased greatly in the lasttwo decades As 95 of AD cases are diagnosed in olderpeople one might think that a causal relationship existsbetween aging and the onset of disease Certainly a numberof similarities between getting old and being affected withAD could be listed Historically the free radical theory ofaging by Harman [155] suggested aging as ldquoside effectrdquo ofreactive oxygen species formed in mitochondrial respiratorychain Apparently free radicals commonly generated byincomplete reduction of oxygen molecule at complexes Iand III of mitochondrial ETC are capable of damagingDNA RNA and proteins They impair energy storage andlead to operational failure of mitochondria with progres-sive decline of cell viability Almost identical conditionsaccompany AD and are an explicit step in pathogenesisof disease As mtDNA is deprived of repair mechanismsROS-induced DNA strand breaks tend to accumulate withage or AD Thus mtDNA is a vulnerable target for ROSbut the reverse the ROS generation due to the mutatedmtDNA is not convincingly confirmed [156] Moreover theevidence that mutated mtDNA accelerates the progress ofaging is also questioned based on the results from studycarried out on transgenic mice model [157] Although someauthors show inconsistency between the free radical theoryand observations cumulative evaluation of the scientificreports points to antioxidant defense systems as importantfactors in protection from premature aging [158] Othermitochondrial components important for their function arehampered with age adenine nucleotide translocase (ANT)nitric oxide synthase (NOS) and carnitine acyltransferase(CT) [159ndash161] Actually NOS activity is elevated in AD asreported from study performed on cellular model of disease[15] We could not find any information about CT activityin AD whereas ANT activity is noticeably inhibited byA120573 or hyperphosphorylated tau and this effect is reversedby mersalyl a reversible alkylating agent of thiol groups[162] Mitochondrial dysfunction observed in transgenicmice models of AD and aging demonstrates higher activ-ity of genes controlling energy metabolism and apoptosisTaken together physiological aging and AD are associatedwith broad-spectrum dysfunction of mitochondria but thefoundations of mitochondrial decline are dissimilar Morediscrepancies between physiological aging and AD werefound with respect to ERS and UPR which play a significantrole in cellular proteostasis Dolichol was selected as an agingmarker because the progressive increase in dolichol levelwas observed in aging brain [163] In contrast ubiquinoneconcentration which is also synthesized from geranylgeranioldiminishes with agingwhereas cholesterol and dolichyl phos-phate concentrations remain unaltered In AD decreasedlevels of dolichol were observed and increased levels ofubiquinone and dolichyl phosphate without any changesin brain cholesterol AD cannot be regarded as a resultof premature aging The drop in dolichol and augmenteddolichyl phosphate concentration points toward disturbedglycosylation in ER of diseased brain while the increase inubiquinone suggests efforts to protect the brain from oxida-tive stress induced by lipid peroxidation [164 165]
14 Targeting ER Stress in AD Therapy
As ERS is a recognized factor in AD drugs that interfere withERS would theoretically have great therapeutic potentialThere are several compounds grouped in classes thatinteract directly with components of the ERS (salubrinalBiP inducer X (BIX) salicylamide analogs flavonoidsguanabenz and STF083010) chemical chaperons (4-phenyl-butyric acid (PBA) tauroursodeoxycholic acid (TUDCA)and trimethylamine oxide (TMAO)) chemicals that inhibitprotein degradation (Eeyarestatin MG132 and Bortezomib)compounds with antioxidant activity (Edaravone diben-zoylmethane derivatives and N-acetyl cysteine (NAC)) anddrugs controlling calcium signaling (dantrolene and carba-zole derivatives) [166] They may act by inducing transienttranslation arrest upregulation of chaperone proteins andaugmented degradation of ER-associated misfolded proteinsFundamental approach in the development of new therapyis the selection of appropriate molecular targets In ER stresssignaling the aim is to alter the expression of ER stress-associated molecules that can rescue cells from the toxiceffect of ERS Recent efforts in establishing new promisingdrugs against AD are pointing to chemical chaperones suchas PBA TUDCA or trimethylamine oxide (TMAO) Thesesubstances improve protein folding and alleviate nativeprotein conformation [166] It was shown on mouse modelsof AD that PBA TUDCA and TMAO stop A120573 accumulationand avoid the loss of dendritic spines [167] Some observa-tions even demonstrated improved memory and cognitivefunctions [168] associated with improved cell survival [169]Salubrinal ((2E)-3-phenyl-N-[222-trichloro-1-[[(8-quinoli-nylamino)thioxomethyl]amino]ethyl]-2-propenamide Sal)selectively inhibits growth arrest and DNA damage inducedgene 34- (GADD34-) phosphatase complex (GADD34associates with protein phosphatase 1 (PP1)) and promotes invitro dephosphorylation of the alpha subunit of eIF-2120572 andIRE1120572ASK1JNK signaling pathway being protective againstERS even induced by tunicamycin Tm [170] In a great deal ofexperiments testing Sal in cultured cells and animal modelsof AD this substance increased the viability of neuronal cellsand A120573 toxicity [171 172] BIX (2-(34-dihydroxyphenyl)-2-oxoethyl ester thiocyanic acid) preferentially inducedBiP mRNA in an ATF6-dependent manner leading toreduced Tm-induced death of neuronal cells [173] AlsoDBM derivative 14ndash26 (221015840-dimethoxydibenzoylmethane)was found to be neuroprotective for SH-SY5Y and PC-12cells by decreasing expression of BiP and CHOP [174]Dantrolene a ryanodine receptor antagonist that inhibitsabnormal calcium release from the ER inhibited expressionof both phosphorylated PERK and eIF2120572 It also reducedCHOP expression and attenuated thapsigargin-inducedapoptosis in PC-12 cells [175] Neuroprotective effects similarto dantrolene were observed for ([9-(3-cyanobenzyl)-14-dimethylcarbazole]) This substance suppressed increases inintracellular Ca2+ in PC-12 cells treated with thapsigarginand reduced levels of BiP and CHOP [176]
Aforementioned compounds were chosen from othersas the most potent ER stress inhibitors and persuasivelyprotective to neuronal cells 18 of 42 different compounds
Oxidative Medicine and Cellular Longevity 11
were exploited in in vivo and in vitro models of centralnervous system disorders with in fact improved cell or tissueviability [166] Thus the brain is the most frequently investi-gated organ in the context of ERS From these experimentsit becomes clear that CHOP functions as proapoptotic factorThe roles of other specific ER stress molecules as moleculartargets for pharmacological intervention are less clear andvary depending on cell type and context
There are few underestimated modulations in APP pro-cessing that shed more light on current dogma of AD patho-genesis The modulation of mevalonate pathway and choles-terol synthesis were reported to stimulate nonamyloidogenicpathway of APP processing [177] Additionally cholesterolderivative 27-hydroxycholesterol (27-OHC) was shown toinduce ER stress which attenuated leptin-dependent viabilityby activating CHOP in SH-SY5Y neuroblastoma cells [178]Irrespective of a number of compounds examined in AD thecall for new drugs modulating ER stress with healing effect isstill waiting to be revealed
Abbreviations
APOE Apolipoprotein EAPPs Secreted form of APPCHOPGADD153 CEBP homologous protein
growth arrest and DNA damageinduced gene 153
Drp1 Dynamin-1-like proteinERAD Endoplasmic-reticulum-
associated protein degradationERO1120572 ER oxidase 1120572ERS Endoplasmic reticulum stressFOH FarnesolGGOH GeranylgeraniolGRP75 Glucose-regulated protein 75GRP78BiP 78 kDa glucose-regulated proteinGSK-3120573 Synthase kinase 3 betaIP3R Inositol triphosphate receptor
IRE1120572 Inositol-requiring enzyme 1 alphaMAM Mitochondria-associated
membraneMAP LC3 Microtubule-associated protein
light chain 3Mfn2 GTPase mitofusin 2MMP Mitochondrial membrane
potentialMPTP Mitochondrial permeability
transition poreNFT Neurofibrillary tanglesPACS-2 Phosphofurin acidic cluster
sorting protein 2PDI Protein disulfide isomerasePERK Protein kinase R- (PKR-) like
ER kinaseRNS Reactive nitrogen speciesROS Reactive oxygen speciesRyR Ryanodine receptorTOM Transporter outer membraneTIM Transporter inner membrane
UPR Unfolded protein responseVDAC Voltage-dependent anion channelXBP-1 Xbox binding protein 1
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
Support for this work was provided by Grant no UMO-201103BNZ70238 from the National Science Centre inPoland
References
[1] R L Nussbaum andC E Ellis ldquoAlzheimerrsquos disease and Parkin-sonrsquos diseaserdquoTheNewEngland Journal ofMedicine vol 348 no14 pp 1356ndash1364 2003
[2] A L Brunkan and A M Goate ldquoPresenilin function and 120574-secretase activityrdquoThe Journal of Neurochemistry vol 93 no 4pp 769ndash792 2005
[3] V Crentsil ldquoThe pharmacogenomics of Alzheimerrsquos diseaserdquoAgeing Research Reviews vol 3 no 2 pp 153ndash169 2004
[4] D L Price and S S Sisodia ldquoMutant genes in familialAlzheimerrsquos disease and transgenic modelsrdquo Annual Review ofNeuroscience vol 21 pp 479ndash505 1998
[5] D Harman ldquoAlzheimerrsquos disease pathogenesis role of agingrdquoAnnals of the New York Academy of Sciences vol 1067 no 1 pp454ndash460 2006
[6] D J Selkoe ldquoTranslating cell biology into therapeutic advancesin Alzheimerrsquos diseaserdquo Nature vol 399 pp A23ndashA31 1999
[7] H Braak and E Braak ldquoNeuropathological stageing of Alz-heimer-related changesrdquo Acta Neuropathologica vol 82 no 4pp 239ndash259 1991
[8] K Hirai G Aliev A Nunomara et al ldquoMitochondrial abnor-malities inAlzheimerrsquos diseaserdquoTheJournal ofNeuroscience vol21 pp 3017ndash3023 2001
[9] U T Brunk C B Jones and R S Sohal ldquoA novel hypothesisof lipofuscinogenesis and cellular aging based on interac-tions between oxidative stress and autophagocytosisrdquoMutationResearchDNAging vol 275 no 3ndash6 pp 395ndash403 1992
[10] P A Stewart K Hayakawa M-A Akers and H V Vinters ldquoAmorphometric study of the blood-brain barrier in Alzheimerrsquosdiseaserdquo Laboratory Investigation vol 67 no 6 pp 734ndash7421992
[11] J P Blass and G E Gibson ldquoThe role of oxidative abnor-malities in the pathophysiology of Alzheimerrsquos diseaserdquo RevueNeurologique vol 147 no 6-7 pp 513ndash525 1991
[12] B Pająk E Kania and A Orzechowski ldquoNucleofection of ratpheochromocytoma PC-12 cells with human mutated beta-amyloid precursor protein gene (APP-sw) leads to reducedreduced viability autophagy-like process and increased expres-sion and secretion of beta amyloidrdquo BioMed Research Interna-tional vol 2015 Article ID 746092 10 pages 2015
[13] J P Blass ldquoCerebrometabolic abnormalities in AlzheimerrsquosdiseaserdquoNeurological Research vol 25 no 6 pp 556ndash566 2003
12 Oxidative Medicine and Cellular Longevity
[14] A Eckert K Schmitt and J Gotz ldquoMitochondrial dysfunc-tionmdashthe beginning of the end in Alzheimerrsquos disease Sep-arate and synergistic modes of tau and amyloid-120573 toxicityrdquoAlzheimerrsquos Research ampTherapy vol 3 article 15 pp 1ndash11 2011
[15] U Keil A Bonert C A Marques et al ldquoAmyloid 120573-inducedchanges in nitric oxide production and mitochondrial activitylead to apoptosisrdquoThe Journal of Biological Chemistry vol 279no 48 pp 50310ndash50320 2004
[16] A M Swomley and D A Butterfield ldquoOxidative stress inAlzheimer disease and mild cognitive impairment evidencefrom human data provided by redox proteomicsrdquo Archives ofToxicology vol 89 no 10 pp 1669ndash1680 2015
[17] C Caspersen N Wang J Yao et al ldquoMitochondrial A120573 apotential focal point for neuronal metabolic dysfunction inAlzheimerrsquos diseaserdquo The FASEB Journal vol 19 no 14 pp2040ndash2041 2005
[18] G E Gibson K-F R Sheu and J P Blass ldquoAbnormalities ofmitochondrial enzymes inAlzheimer diseaserdquo Journal of NeuralTransmission vol 105 no 8-9 pp 855ndash870 1998
[19] A Eckert S Hauptmann I Scherping et al ldquoSoluble beta-amyloid leads to mitochondrial defects in amyloid precursorprotein and tau transgenic micerdquo Neurodegenerative Diseasesvol 5 no 3-4 pp 157ndash159 2008
[20] A Eckert K L Schulz V Rhein and J Gotz ldquoConvergenceof amyloid-120573 and tau pathologies on mitochondria in vivordquoMolecular Neurobiology vol 41 no 2-3 pp 107ndash114 2010
[21] S Hauptmann I Scherping S Drose et al ldquoMitochondrial dys-function an early event in Alzheimer pathology accumulateswith age in AD transgenic micerdquoNeurobiology of Aging vol 30no 10 pp 1574ndash1586 2009
[22] M Manczak B S Park Y Jung and P H Reddy ldquoDifferentialexpression of oxidative phosphorylation genes in patients withAlzheimerrsquos disease implications for early mitochondrial dys-function and oxidative damagerdquo NeuroMolecular Medicine vol5 no 2 pp 147ndash162 2004
[23] M P Mattson M Gleichmann and A Cheng ldquoMitochondriain neuroplasticity and neurological disordersrdquo Neuron vol 60no 5 pp 748ndash766 2008
[24] P I Moreira M S Santos and C R Oliveira ldquoAlzheimerrsquosdisease a lesson frommitochondrial dysfunctionrdquoAntioxidantsamp Redox Signaling vol 9 no 10 pp 1621ndash1630 2007
[25] P H Reddy ldquoMitochondrial dysfunction in aging and Alz-heimerrsquos disease strategies to protect neuronsrdquo Antioxidants ampRedox Signaling vol 9 no 10 pp 1647ndash1658 2007
[26] V Rhein G Baysang S Rao et al ldquoAmyloid-beta leads toimpaired cellular respiration energy production andmitochon-drial electron chain complex activities in humanneuroblastomacellsrdquo Cellular and Molecular Neurobiology vol 29 no 6-7 pp1063ndash1071 2009
[27] V Rhein X Song A Wiesner et al ldquoAmyloid-120573 and tausynergistically impair the oxidative phosphorylation system intriple transgenic Alzheimerrsquos disease micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 47 pp 20057ndash20062 2009
[28] R X Santos S C Correia X Wang et al ldquoAlzheimerrsquos diseasediverse aspects ofmitochondrial malfunctioningrdquo InternationalJournal of Clinical and Experimental Pathology vol 3 no 6 pp570ndash581 2010
[29] N R Sims ldquoEnergy metabolism oxidative stress and neuronaldegeneration in Alzheimerrsquos diseaserdquoNeurodegeneration vol 5no 4 pp 435ndash440 1996
[30] H K Anandatheerthavarada G Biswas M-A Robin and NG Avadhani ldquoMitochondrial targeting and a novel transmem-brane arrest of Alzheimerrsquos amyloid precursor protein impairsmitochondrial function in neuronal cellsrdquo The Journal of CellBiology vol 161 no 1 pp 41ndash54 2003
[31] L Devi B M Prabhu D F Galati N G Avadhani and HK Anandatheerthavarada ldquoAccumulation of amyloid precur-sor protein in the mitochondrial import channels of humanAlzheimerrsquos disease brain is associated with mitochondrialdysfunctionrdquo The Journal of Neuroscience vol 26 no 35 pp9057ndash9068 2006
[32] L Devi and H K Anandatheerthavarada ldquoMitochondrialtrafficking of APP and alpha synuclein relevance to mito-chondrial dysfunction in Alzheimerrsquos and Parkinsonrsquos diseasesrdquoBiochimica et Biophysica Acta vol 1802 no 1 pp 11ndash19 2010
[33] M Manczak T S Anekonda E Henson B S Park J Quinnand P H Reddy ldquoMitochondria are a direct site of A120573 accumu-lation in Alzheimerrsquos disease neurons implications for free rad-ical generation and oxidative damage in disease progressionrdquoHuman Molecular Genetics vol 15 no 9 pp 1437ndash1449 2006
[34] P F Pavlov C H Petersen E Glaser and M AnkarcronaldquoMitochondrial accumulation of APP and A120573 significancefor Alzheimer disease pathogenesisrdquo Journal of Cellular andMolecular Medicine vol 13 no 10 pp 4137ndash4145 2009
[35] C Schmidt E Lepsverdize S L Chi et al ldquoAmyloid precursorprotein and amyloid 120573-peptide bind to ATP synthase andregulate its activity at the surface of neural cellsrdquo MolecularPsychiatry vol 13 no 10 pp 953ndash969 2008
[36] K Johar A Priya and M T T Wong-Riley ldquoRegulation ofNa+K+-ATPase by nuclear respiratory factor 1 implication inthe tight coupling of neuronal activity energy generation andenergy consumptionrdquo The Journal of Biological Chemistry vol287 no 48 pp 40381ndash40390 2012
[37] J Astrup P M Sorensen and H R Sorensen ldquoOxygen andglucose consumption related to Na+-K+ transport in caninebrainrdquo Stroke vol 12 no 6 pp 726ndash730 1981
[38] M Erecinska and I A Silver ldquoATP and brain functionrdquo Journalof Cerebral Blood Flow and Metabolism vol 9 no 1 pp 2ndash191989
[39] MMata D J Fink H Gainer et al ldquoActivity-dependent energymetabolism in rat posterior pituitary primarily reflects sodiumpump activityrdquo Journal of Neurochemistry vol 34 no 1 pp 213ndash215 1980
[40] J Kang H-G Lemaire A Unterbeck et al ldquoThe precursor ofAlzheimerrsquos disease amyloid A4 protein resembles a cell-surfacereceptorrdquo Nature vol 325 no 6106 pp 733ndash736 1987
[41] C L Masters and D J Selkoe ldquoBiochemistry of amyloid 120573-protein and amyloid deposits in Alzheimer diseaserdquoCold SpringHarbour Perspectives in Medicine vol 2 no 6 Article IDa006262 2012
[42] A Lai S S Sisodia and I S Trowbridge ldquoCharacterization ofsorting signals in the beta-amyloid precursor protein cytoplas-mic domainrdquo The Journal of Biological Chemistry vol 270 no8 pp 3565ndash3573 1995
[43] A I Placido C M F Pereira A I Duarte et al ldquoThe role ofendoplasmic reticulum in amyloid precursor protein processingand trafficking implications for Alzheimerrsquos diseaserdquo Biochim-ica et Biophysica ActamdashMolecular Basis of Disease vol 1842 no9 pp 1444ndash1453 2014
Oxidative Medicine and Cellular Longevity 13
[44] C Kaether P Skehel and C G Dotti ldquoAxonal membraneproteins are transported in distinct carriers a two-color videomicroscopy study in cultured hippocampal neuronsrdquoMolecularBiology of the Cell vol 11 no 4 pp 1213ndash1224 2000
[45] A Kamal A Almenar-Queralt J F LeBlanc E A Robertsand L S B Goldstein ldquoKinesin-mediated axonal transportof a membrane compartment containing 120573-secretase andpresenilin-1 requires APPrdquo Nature vol 414 no 6864 pp 643ndash648 2001
[46] R G Perez S Soriano J D Hayes et al ldquoMutagenesis identi-fies new signals for 120573-amyloid precursor protein endocytosisturnover and the generation of secreted fragments includingA12057342rdquo The Journal of Biological Chemistry vol 274 no 27 pp18851ndash18856 1999
[47] D G Cook M S Forman J C Sung et al ldquoAlzheimerrsquos A120573(1ndash42) is generated in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo Nature Medicine vol 3 no 9 pp1021ndash1023 1997
[48] A S C Chyung B D Greenberg D G Cook R W Domsand V M-Y Lee ldquoNovel 120573-secretase cleavage of 120573-amyloidprecursor protein in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo The Journal of Cell Biology vol138 no 3 pp 671ndash680 1997
[49] RW-Y Choy Z Cheng and R Schekman ldquoAmyloid precursorprotein (APP) traffics from the cell surface via endosomesfor amyloid 120573 (A120573) production in the trans-Golgi networkrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 30 pp E2077ndashE2082 2012
[50] S A Small and S Gandy ldquoSorting through the cell biologyof Alzheimerrsquos disease intracellular pathways to pathogenesisrdquoNeuron vol 52 no 1 pp 15ndash31 2006
[51] M Goedert ldquoAlzheimerrsquos and Parkinsonrsquos diseases the prionconcept in relation to assembled A120573 tau and 120572-synucleinrdquoScience vol 349 no 6248 2015
[52] I Benilova E Karran and B De Strooper ldquoThe toxic A120573oligomer and Alzheimerrsquos disease an emperor in need ofclothesrdquo Nature Neuroscience vol 15 no 3 pp 349ndash357 2012
[53] E H Corder A M Saunders W J Strittmatter et al ldquoGenedose of apolipoprotein E type 4 allele and the risk of Alzheimerrsquosdisease in late onset familiesrdquo Science vol 261 no 5123 pp 921ndash923 1993
[54] S Brunholz S Sisodia A Lorenzo C Deyts S Kins and GMorfini ldquoAxonal transport of APP and the spatial regulationof APP cleavage and function in neuronal cellsrdquo ExperimentalBrain Research vol 217 no 3-4 pp 353ndash364 2012
[55] G Thinakaran and E H Koo ldquoAmyloid precursor proteintrafficking processing and functionrdquo The Journal of BiologicalChemistry vol 283 no 44 pp 29615ndash29619 2008
[56] T Saitoh M Sundsmo J-M Roch et al ldquoSecreted formof amyloid beta protein precursor is involved in the growthregulation of fibroblastsrdquo Cell vol 58 no 4 pp 615ndash622 1989
[57] M PMattson B Cheng A R Culwell F S Esch I Lieberburgand R E Rydel ldquoEvidence for excitoprotective and intraneu-ronal calcium-regulating roles for secreted forms of the 120573-amyloid precursor proteinrdquo Neuron vol 10 no 2 pp 243ndash2541993
[58] K J Lee C E H Moussa Y Lee et al ldquoBeta amyloid-independent role of amyloid precursor protein in generationandmaintenance of dendritic spinesrdquoNeuroscience vol 169 no1 pp 344ndash356 2010
[59] H Meziane J-C Dodart C Mathis et al ldquoMemory-enhancingeffects of secreted forms of the 120573-amyloid precursor proteinin normal and amnestic micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no21 pp 12683ndash12688 1998
[60] P R Turner K OrsquoConnor W P Tate and W C AbrahamldquoRoles of amyloid precursor protein and its fragments inregulating neural activity plasticity and memoryrdquo Progress inNeurobiology vol 70 no 1 pp 1ndash32 2003
[61] D Beher L Hesse C L Masters and GMulthaup ldquoRegulationof Amyloid Protein Precursor (APP) binding to collagen andmapping of the binding sites on APP and collagen type Irdquo TheJournal of Biological Chemistry vol 271 no 3 pp 1613ndash16201996
[62] S L Sabo A F Ikin J D Buxbaum and P Greengard ldquoTheamyloid precursor protein and its regulatory protein FE65 ingrowth cones and synapses in vitro and in vivordquo The Journal ofNeuroscience vol 23 no 13 pp 5407ndash5415 2003
[63] D H Small V Nurcombe G Reed et al ldquoA heparin-bindingdomain in the amyloid protein precursor of Alzheimerrsquos diseaseis involved in the regulation of neurite outgrowthrdquoThe Journalof Neuroscience vol 14 no 4 pp 2117ndash2127 1994
[64] T G Williamson S S Mok A Henry et al ldquoSecreted glypicanbinds to the amyloid precursor protein of Alzheimerrsquos disease(APP) and inhibits APP-induced neurite outgrowthrdquoThe Jour-nal of Biological Chemistry vol 271 no 49 pp 31215ndash312211996
[65] T Ondrejcak I Klyubin N-W Hu A E Barry W K Cullenand M J Rowan ldquoAlzheimerrsquos disease amyloid 120573-protein andsynaptic functionrdquo NeuroMolecular Medicine vol 12 no 1 pp13ndash26 2010
[66] D Puzzo L Privitera and A Palmeri ldquoHormetic effect ofamyloid-beta peptide in synaptic plasticity and memoryrdquo Neu-robiology of Aging vol 33 no 7 pp 1484e15ndash1484e24 2012
[67] C M F Pereira ldquoCrosstalk between endoplasmic reticulumstress and protein misfolding in neurodegenerative diseasesrdquoISRN Cell Biology vol 2013 Article ID 256404 22 pages 2013
[68] R-W Shin T C Saido M Maeda and T Kitamoto ldquoNovel 120572-secretase cleavage ofAlzheimerrsquos amyloid120573 precursor protein inthe endoplasmic reticulum of COS7 cellsrdquoNeuroscience Lettersvol 376 no 1 pp 14ndash19 2005
[69] E A Schon and E Area-Gomez ldquoMitochondria-associatedER membranes in Alzheimer diseaserdquo Molecular and CellularNeuroscience vol 55 pp 26ndash36 2013
[70] R Rizzuto P Pinton W Carrington et al ldquoClose contacts withthe endoplasmic reticulum as determinants of mitochondrialCa2+ responsesrdquo Science vol 280 no 5370 pp 1763ndash1766 1998
[71] S E Logue P Cleary S Saveljeva and A Samali ldquoNewdirections in ER-stress induced cell deathrdquoApoptosis vol 18 no5 pp 537ndash546 2013
[72] J P Decuypere KWelkenhuyzen T Luyten et al ldquoIns(145)P3receptor-mediated BI-1 Ca2+ signaling and autophagy induc-tion are interrelatedrdquo Autophagy vol 7 pp 1472ndash1489 2011
[73] K S Vetrivel and G Thinakaran ldquoMembrane rafts in Alz-heimerrsquos disease beta-amyloid productionrdquo Biochimica et Bio-physica Acta vol 1801 no 8 pp 860ndash867 2010
[74] H Urra E Dufey F Lisbona D Rojas-Rivera and C HetzldquoWhen ER stress reaches a dead endrdquo Biochimica et BiophysicaActa (BBA) Molecular Cell Research vol 1833 no 12 pp 3507ndash3517 2013
14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
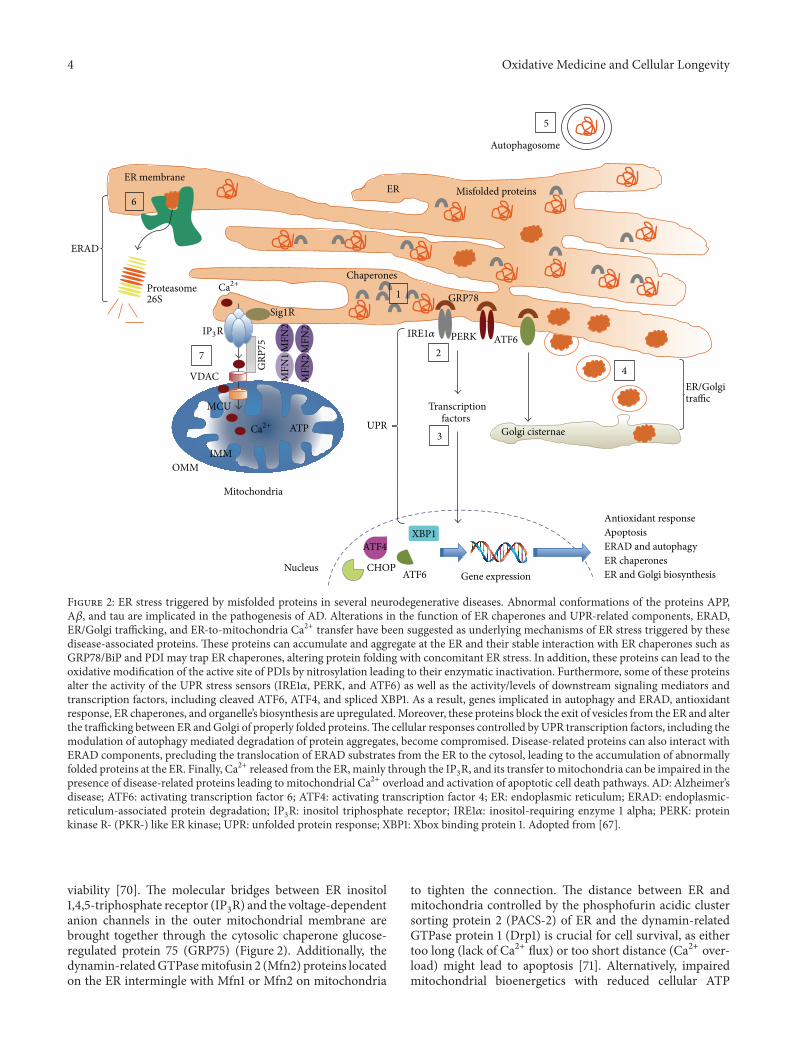
4 Oxidative Medicine and Cellular Longevity
ER membrane
6
ERAD
Proteasome26S
Ca2+
Ca2+
IP3R
VDAC
MCU
IMMOMM
ATP
GRP
75
MFN
1
MFN
2M
FN2
MFN
2
Sig1R
UPR
7
Mitochondria
Nucleus CHOP ATF6
XBP1ATF4
Gene expression
3
Transcriptionfactors
2
IRE1120572 PERK ATF6
1 GRP78
Chaperones
ER Misfolded proteins
Autophagosome
5
4
ERGolgitraffic
Golgi cisternae
Antioxidant responseApoptosisERAD and autophagyER chaperonesER and Golgi biosynthesis
Figure 2 ER stress triggered by misfolded proteins in several neurodegenerative diseases Abnormal conformations of the proteins APPA120573 and tau are implicated in the pathogenesis of AD Alterations in the function of ER chaperones and UPR-related components ERADERGolgi trafficking and ER-to-mitochondria Ca2+ transfer have been suggested as underlying mechanisms of ER stress triggered by thesedisease-associated proteins These proteins can accumulate and aggregate at the ER and their stable interaction with ER chaperones such asGRP78BiP and PDI may trap ER chaperones altering protein folding with concomitant ER stress In addition these proteins can lead to theoxidative modification of the active site of PDIs by nitrosylation leading to their enzymatic inactivation Furthermore some of these proteinsalter the activity of the UPR stress sensors (IRE1120572 PERK and ATF6) as well as the activitylevels of downstream signaling mediators andtranscription factors including cleaved ATF6 ATF4 and spliced XBP1 As a result genes implicated in autophagy and ERAD antioxidantresponse ER chaperones and organellersquos biosynthesis are upregulatedMoreover these proteins block the exit of vesicles from the ER and alterthe trafficking between ER andGolgi of properly folded proteinsThe cellular responses controlled byUPR transcription factors including themodulation of autophagy mediated degradation of protein aggregates become compromised Disease-related proteins can also interact withERAD components precluding the translocation of ERAD substrates from the ER to the cytosol leading to the accumulation of abnormallyfolded proteins at the ER Finally Ca2+ released from the ERmainly through the IP
3R and its transfer tomitochondria can be impaired in the
presence of disease-related proteins leading to mitochondrial Ca2+ overload and activation of apoptotic cell death pathways AD Alzheimerrsquosdisease ATF6 activating transcription factor 6 ATF4 activating transcription factor 4 ER endoplasmic reticulum ERAD endoplasmic-reticulum-associated protein degradation IP
3R inositol triphosphate receptor IRE1120572 inositol-requiring enzyme 1 alpha PERK protein
kinase R- (PKR-) like ER kinase UPR unfolded protein response XBP1 Xbox binding protein 1 Adopted from [67]
viability [70] The molecular bridges between ER inositol145-triphosphate receptor (IP
3R) and the voltage-dependent
anion channels in the outer mitochondrial membrane arebrought together through the cytosolic chaperone glucose-regulated protein 75 (GRP75) (Figure 2) Additionally thedynamin-relatedGTPasemitofusin 2 (Mfn2) proteins locatedon the ER intermingle with Mfn1 or Mfn2 on mitochondria
to tighten the connection The distance between ER andmitochondria controlled by the phosphofurin acidic clustersorting protein 2 (PACS-2) of ER and the dynamin-relatedGTPase protein 1 (Drp1) is crucial for cell survival as eithertoo long (lack of Ca2+ flux) or too short distance (Ca2+ over-load) might lead to apoptosis [71] Alternatively impairedmitochondrial bioenergetics with reduced cellular ATP
Oxidative Medicine and Cellular Longevity 5
levels stimulate autophagyThe molecular mechanism of ER-mediated autophagy is accurately regulated by Beclin 1 as wellas ER membrane bound protein Bax inhibitor 1 (BI-1) Bothproteins are capable of promoting autophagy through IP
3R-
dependent mechanism [72]
5 APP Processing and ER Stress Response
The nature of APP processing is determined by the com-position of membrane with cholesterol rich lipid rafts asthe site of amyloidogenic cleavage [73] Consequently onemight expect that fate of APP is at least partly establishedby the representation of lipid rafts and possible access to 120573-versus120572-secretase Irrespective of the type of cleavage there isone underestimated biochemical step in ER that might bringabout damaging upshot It is apparently the effectiveness ofER situated N-glycosylation which makes the APP moleculesuitably folded If misfoldedmalfolded protein(s) accumu-late in the ER complex cascade of reactions known as theunfolded protein response (UPR) is triggered with the so-called endoplasmic reticulum stress response (ERS) Today itis widely accepted that during UPR the ER sensors proteinkinase R- (PKR-) like ER kinase (PERK) activating proteinkinase 6 (ATF6) and inositol-requiring enzyme 1 alpha(IRE1120572) are freed fromGRP78BiP protein repression whichhereafter become activated Without going into details ofindividual sensor action a number of reactions occur at boththe genomic and cytoplasmic level with selective degradationof mRNAs encoding protein(s) with abnormal folding andinhibition of protein translation except for genes importantfor UPR redox homeostasis energy metabolism and pro-tein folding [67] On the one hand PERK phosphorylateseukaryotic initiation factor 2 alpha (eIF2120572) to stop entrance ofmethionyl-tRNA to the ribosome on the other hand it allowstranslation of activating transcription factor 4 (ATF4) geneIRE1120572 sets off alternative splicing of Xbox binding protein 1(XBP1) transcript leading to activation of the transcriptionfactor liable to stimulate ERGolgi biogenesis and formationof proteins involved in endoplasmic-reticulum-associatedprotein degradation known as ERAD (Erdj4 p58IPK EDEMRAMP-4 PDI-P5 and HEDJ for details see [67]) FinallyATF6 is activated in the Golgi complex through proteolyticcleavage and translocates to nucleus where it cooperates withXBP1 in upregulation of chaperones and ERAD-related genes[43] In principle UPR is activated to restore ER homeostasisand stop the accumulation of aberrantly formed protein(s)but if the strength of ER stress is unbearable (meaning thatit cannot be compensated by UPR) there is a path to activateapoptosis Among other routes the most central role isplayed by the major proapoptotic transcription factor CEBPhomologous protein CHOPgrowth arrest and DNA damageinduced gene 153 GADD153 It downregulates the antiapop-totic protein Bcl-2 and upregulates proapoptotic Bax and Bak[74] CHOPGADD153 leads to excessive production of reac-tive oxygen species (ROS) within ER subsequent depletionof reduced glutathione (GSH) and Ca2+ flux from the ER tocytoplasm through the IP
3R [75] One hitherto unresolved
issue is the physiologic importance of APP and GRP78BiPinteraction disclosed in coprecipitation study carried out by
Yamamoto et al [76] Bulk ofAPP associatedwithGRP78BiPwas immature protein Given that GRP78BiP expressionlevels declined in samples of brain tissue obtained fromFAD patients as demonstrated by Katayama et al [77] fewdistinct scenarios are possible First binding to GRP78BiPsuggests ER accumulation of immature APP Second APPinteraction with GRP78BiP is a noticeable sign of UPRwhich is further validated by downregulation of GRP78BiPin FAD patients Finally retention of APP in ER is mostlikely a result of amyloidogenic APP processing and as such itprobably occurs in MAM the cholesterol enriched domainsER-mitochondria crossing point is therefore of particularinterest in deciphering the links between the ER placed APPprocessing UPR and resulting cellular responses such asautophagy apoptosis and inflammatory reaction observed inAD (Figure 2)
6 Pathology of Tau Protein
Microtubule-associated protein tau controls assembly andprevents microtubules from severing Microtubular networkis fundamental component of cytoskeleton essential forintracellular transport of secretory vesicles and organelles(ie mitochondria) Glycogen synthase kinase 3 beta- (GSK-3120573-) targeted hyperphosphorylation of tau causes this proteinto dissociate frommicrotubules Consequently microtubulesbecome fragmented and microtubule-dependent transportsystem fails Furthermore hyperphosphorylated tau (P-tau)is prone to form oligomers and toxic filaments known asNFTs or tauopathy [78] A variety of tau conformers werereported to exist pointing to different tauopathies capa-ble of self-propagation [79 80] Interestingly intracellulartau inclusions define AD as clinical symptoms of diseaseobserved when tauopathy is abundant together with intra-cellular A120573 deposits in neocortex [51 81 82] People withabundant A120573 plaques but no or only a few neurofibrillarylesions do not have AD Clinicopathological correlationstudies have been crucial to generate hypotheses about thepathophysiology of the disease by establishing the fact thatthere is a continuum between ldquonormalrdquo aging and ADdementia and that the amyloid plaque buildup occurs primar-ily before the onset of cognitive deficits while neurofibrillarytangles neuron loss and particularly synaptic loss parallel theprogression of cognitive decline [83] Thus misfolded pro-teins and descendant toxic filaments with a number of inter-mediates are critical for manifestation of AD as fibrillogenicAPP processing is not enough for onset of disease Althoughthe molecular mechanism of tauopathy is not deciphered infull recent reports suggest ER stress as the starting point[84 85] This idea is substantiated by the elevated levels ofERS and UPR markers together with P-tau and GSK-3120573 inbrains affected by AD [86] Therefore on the one hand theincidence of UPR is strongly correlated with the presence ofNFTs on the other hand aggregation of P-tau induces ERSwith resultant UPR Some lines of evidence confirmed UPRactivation near the beginning of NFTs formation and pointto the functional link between malformed tau protein andUPRThe in vitro experiments with phosphatase 2A inhibitoror phosphorylation activator demonstrated enhanced P-tau
6 Oxidative Medicine and Cellular Longevity
formation in neurons in concert with the increased levelsof PERK eIF2120572 and XBP1 transcript apparent markers ofUPR [87] Furthermore GRP78BiP was found to encouragetau phosphorylation through facilitated substrate capture byGSK-3120573 [88] GSK-3120573 seems to play dual role first this kinaseprotects neurons from apoptosis as P-tau accumulation isstrong molecular signal to trigger UPR with subsequentautophagy Second UPR raises GSK-3120573 activity throughlysosomal degradation of inactive GSK-3120573 (P-Ser9-GSK-3120573)To sum up ERS and UPR are important molecular machinesused to prevent cell viability turned on by tauopathy
7 Oxidative Stress in ER
Sacs and tubes of ER delineate the compartment where thenewly synthesized proteins undergo maturation to nativestate Native state indicates properly folded fully functionalprotein Important reactions essential for protein folding ofunbranched polypeptide chains include amino acids oxida-tion and glycosylation As a result redox homeostasis inER is shifted to oxidative state so as to promote disulfidebond formation between adjacent cysteines Oxidation ofsulfhydryl groups required to make disulfide bonds is con-trolled by ER oxidase 1120572 (ERO1120572) Next disulfide bondscould be subject to posttranslational modification disulfideexchange by protein disulfide isomerase (PDI) PDI is ableto correct mispaired thiol residues by catalyzing the breakageand formation of correct disulfide bonds These enzymes arefundamental for protein folding Oxidation allows twistingof proteins which is followed by N-glycosylation andor C-and O-mannosylation Protein N-glycosylation in eukary-otes is a complex process divided into several steps Firstthere is a ldquocall forrdquo carrier lipids (polyisoprenyl phosphatessuch as dolichyl phosphates) the membrane lipids knownto function as glycosyl transporters In mammalian cellsthe limiting substrate for dolichol biosynthesis is geranyl-geraniol (GGOH) of mevalonate pathway Dolichols thelongest aliphatic molecules synthesized in animal cellshave 18ndash21 120572-isoprene saturated units (C90ndash105) criticalfor their recognition by the enzymes (glycosyltransferases)that glucosylate dolichyl phosphates [89] Once dolichylmonophosphates (Dol-P) are formed in the ER membranethe precursor oligosaccharide donor (GLc
3Man9GLCNAc
2-
P-P-dolichol) for protein N-glycosylation can be synthesizedon the lumenal leaflet of ER First three sugar intermediatesare produced (Man-P-Dol Glc-P-Dol GlcNAc-P-P-Dol andMan5GlcNAc-P-P-Dol) on the cytoplasmic leaflet of the
ER Next enzyme flippases mediate transbilayer movementof the aforementioned intermediates to lumenal side ofER where conversion to Glc
3Man9GlcNAc
2-P-P-Dol could
be completed Glc3Man9GlcNAc
2-P-P-Dol is also used for
biosynthesis of glycosylphosphatidylinositol (GPI) anchorsTaken together lipid-mediated glycosylation plays a vital
role in the appropriate protein folding and intracellulartranslocation of N-linked glycoproteins [90] Likewise it isimportant for protein O- and C-mannosylation and GPIanchorage Moreover Dol-P availability in the ER is the rate-limiting factor in the production of glycolipid intermediatesand N-glycosylation
8 ER Stress and Apoptosis
It was shown that during ERS the ER resident proapop-totic cysteine protease known as caspase (caspase-12 in ratcaspase-4 in humans) is activated through cleavage As aresult caspase cascade is started via caspase-9 that in turnstimulates effector caspase-3 [91 92] The central role playedby ER in programmed cell death is achieved by PERK branchwhere ATF4 induces the expression of CHOPGADD153which represses antiapoptotic Bcl-2 family proteins andsimultaneously shuffles ER membrane Bax and Bak proteinsinto outer mitochondrial membrane Consequently poresare formed to leak the components of apoptosome frommitochondrial intermembrane space [93] Another impor-tant mechanism of ERS-induced apoptosis is led by Ca2+-dependent ERO1120572-IP
3R pathway where ERO1120572 collaborates
with IP3R in Ca2+ efflux from ER to mitochondria via MAM
[75 94] Accordingly Ca2+ influx facilitates cytochrome 119888release frommitochondria besides cytochrome 119888 can bind toER IP
3R and the complex amplifies the apoptotic signal in a
feedforward manner [95] Last but not least ERS-associatedapoptotic programme is set off by c-Jun N-terminal kinase(JNK) as the effect of IRE1120572 complexed with TNF-receptor-associated factor 2 (TRAF2) activation of apoptosis-signal-regulating kinase 1 (ASK1) [96 97]
What does ERS drive to induce apoptotic death inneurons Actually many reports indicate that APP and A120573 aswell as hyperphosphorylated tau have been shown to blockmitochondrial transport which results in impaired energystorage and oxidative stress [98ndash101] Indeed accumulationof APP A120573 and NFTs in mitochondria led to reducedactivity of some enzymes involved in substrate oxidation(tricarboxylic acid cycle) electron transport chain (ETC)and ATP synthase as well as severely diminishing import ofnuclear-encoded proteins [26 27 102ndash104] One may ask ifthere is any additional link between ER and mitochondriaother than MAM which could account for apoptotic signalThough not directly ER significantly contributes to oxidativestress in mitochondria of AD-affected subjects
9 Abnormal APP Processing and TraffickingCulminate in ER Pathology of AD
From the morphological point of view as neurons arehighly specialized cells soma dendrites and neurites areconsiderably distinct structures Proteins needed by thesecompartments are delivered via microtubules once proteinshave suitable sorting signals (ie APP trafficking from ERto plasma membrane is associated with several posttrans-lational modifications with oxidation and N-glycosylation)Additionally the Golgi apparatus follows ER in subsequentAPP adjustment (O- and N-glycosylation phosphorylationand sulphonation) [42] Any inaccurate alteration of the APPmolecule is potentially hazardous as protein final destiny ismissed causing its retention in ER or trans-Golgi network(TGN) In addition other unusual settings for APP arepossible as this large protein has few signal sequences hiddenwhen APP is correctly folded In the cells transfected withAPP this protein enters coat protein complex I (COPI)
Oxidative Medicine and Cellular Longevity 7
vesicles and undergoes retrograde transport from cis endof the Golgi complex back to the ER [76] Such responsecauses accumulation of APP in the tubulocisternal ER systemtogether with aberrant intracellular translocation of theprotein Interestingly the issue whether APP is subject toretrograde transport with successive fibrillogenic processingbecause of UPR and its interaction with GRP78BiP is notclear as the observations are inconsistent [77] Nonethelessaccumulation of misfoldedmalfolded proteins in the ERsuggests disorganized process of posttranslational change Asanticipated the accretion of proteins of anomalous patternsignals ERS and UPR followed by increased vulnerability toapoptotic cell death Prior to decay however APP proteinlevels in the ER lessen by dint of cleavage with 120573- and120574-secretase [105 106] Products of this cleavage (sAPP120573A120573 and AICD fragment) all appreciably influence neuronalsurvival most likely through nonnative form of the APPsubstrate
10 APP A120573 and NFTs MarkMitochondria as Targeted in AD
Mitochondrial import of A12057340
and A12057342
peptides throughthe translocase TOM complex was blocked by preincubationof isolated mitochondria with antibodies raised againstTOM proteins (TOM20 TOM40 and TOM70) [107]Neither VDAC inhibition with antagonist antibodies norinhibition of mitochondrial permeability transition pores(MPTP) or fall of mitochondrial membrane potential(MMP) affected uptake of A120573 [17] With regard to APPelegant study performed by Anandatheerthavarada andhis colleagues [30] revealed that C-terminal truncatedAPP (lacking A120573) targets mitochondria in cholinergicGABAergic dopaminergic and glutamatergic neurons ofAD brain by clogging up mitochondrial protein translocasecomplex TOM40TIM22 Authors propose that occlusionof translocase is followed by blunted import of nuclear-encoded proteins vitally important for energy homeostasisMitochondrial APP protein transmembrane orientationindicates NH
2-terminal inside in contact with translocase
whereas COOH-terminal is facing cytoplasmic side NH2-
terminus has mitochondrial signal sequence Astonishinglymitochondrial APP molecules were nonglycosylated givingrise to speculation that protein molecules that arrivedat mitochondria have not achieved molecular maturity[31] The accumulation of nonglycosylated APP species inmitochondrial import channels of AD brain was directlyrelated to decreased mitochondrial functions as validatedby the decline in cytochrome 119888 oxidase activity (complexIV) and elevated levels of H
2O2 Furthermore in AD
brain the mitochondrial accumulation of nonglycosylatedAPP went along with a corresponding reduction in plasmamembrane-associatedAPP It suggests that ADbrain hasAPPprocessing and trafficking severely affected by incompleteN-glycosylation ensuing ER protein accumulation andexposure of the cryptic mitochondrial targeting signalfor assisting chaperone proteins Several lines of evidenceindicate that not fully formed proteins are phosphorylatedand bind to the cytosolic proteins required for movement
from ER to mitochondria [108ndash110] Moreover ERS andUPR in AD seem to be incompetent and inefficient inelimination of malfolded proteins In addition to APPalso A120573 was frequently reported to occupy mitochondriaalthough its origin and mechanism of mitochondriatargeting mostly remain unexplored (except for involvementof protein translocase TOM complex) The possibilityof mitochondrial A120573 generation has to be ruled out asmembrane orientation of arrested APP does not allow accessto 120574-secretase (120574-secretase activity in mitochondria wasdetected by independent study) [111] Probably A120573 speciesis derived from APP prior to its translocation (ER) soA120573 may be transported to mitochondria independentlyof APP Collectively observations showing mitochondrialpresence of APP A120573 and tau in aberrant configuration pointtoward the anomaly of protein folding at the level of ER andGolgi apparatus Nonglycosylated molecule of APP suggestsdefective transfer of sugar core from dolichyl phosphate(s)and further modifications such as O- and N-glycosylationsOne may admit that the lack of glycosyl residue bringsabout pathologic processing and trafficking of APP and itsfragments In point of fact dolichol derivatives mixture ofpolyprenols (acyclic isoprenoid alcohols) known as Ropren(Solagran Limited Melbourne Australia) commerciallyused to treat liver diseases were tested in the treatment ofAD in two separate trials conducted in 2005 and 2006 withpromising results [httpwwwasxcomauasxpdf20071119pdf315x5nh4hm8wv7pdf httpwwwasxcomauasxpdf20070221pdf3111ztwcbqzkk9pdf] Further studies areurgently needed to test how important the glycosylationprocess is in the pathogenesis of AD Dolichols areobtained from geranylgeraniol (GGOH) and the latter is anintermediate of mevalonate pathway GGOH is a commonsubstrate for dolichol and ubiquinone synthesis but it isalso necessary for protein prenylation As both GGOHand farnesol (FOH) are engaged in protein prenylationmore concern should be laid on the importance of thesecompounds in AD pathogenesis Observations demonstrat-ing mitochondrial relocalization of other proteins withoutposttranslational modification seemingly point to increasedmitochondrial targeting of immature molecules resultingin mitochondrial dysfunction and acceleration of diseaseprogression [34 112ndash114]
11 Oxidative Stress in Mitochondria ofAD Brains
Normal physiological functions of APP are thought to beinvolved in the stabilizing contact points between synapsesand maintaining mitochondrial functions [60 115] Mito-chondrial dysfunction was often observed regardless of theexperimental model used to study AD [8 29 116ndash119]It includes defects in oxidative phosphorylation decreasedATP decreased membrane potential increased productionof ROSRNS and perturbation in mitochondrial fusion andfission [15 30 31 115 120ndash122] Hyperphosphorylated tau wasalso reported to impair mitochondrial functions [123] Usingproteomic approach the strongest defects of the respiratorycapacity were observed mainly at complexes I IV and ATP
8 Oxidative Medicine and Cellular Longevity
synthase (complex V) at both protein and activity level [124]While APP A120573 and hyperphosphorylated tau are potentinhibitors of mitochondrial import of nuclear-encoded pro-teins apparently the pathology of each leads to metabolicharm in different way In freshly isolated mitochondriafrom AD brains the APP inhibited mitochondrial importof cytochrome 119888 oxidase (COX) subunits IV and Vb [31]Dysfunction of COX increases ROS production (incompletereduction of oxygen molecules) reduces energy stores anddisturbs energy metabolism Accordingly in AD patientsdeficiency of COX was found in brains and platelets [119125 126] Similar to APP A120573 was found in mitochondriaof transgenic mice and cellular and human AD models [1719 33 103 127ndash130] At present it is not clear whether theobservedmitochondrial toxicity is due toAPP orA120573 orNFTsaccumulation Anyway some regularity is observed withrespect to most affected components of respiratory chainNADH-ubiquinone oxidoreductase (complex I) activity isreduced to the utmost by hyperphosphorylated tau whereasdecreased activity of cytochrome 119888 oxidase (complex IV)that resulted in mitochondrial dysfunction was observedduring A120573 and APP accumulation [14 15 31 32 34]Concomitantly rise in the activity of antioxidant enzymesmanganese superoxide dismutase (Mn-SOD) and catalase(CAT) was demonstrated in response to elevated levelsof free radicals including superoxide anion radical (O
2
∙minus)hydroperoxyl radical (HO
2
∙) hydroxyl radical (OH∙) andnitric oxide radical (NO∙) [15] Consistent with observationsof chronic respiratory chain dysfunction and mitochondrialoxidative stress there are reports showing their contributionto tau pathology in AD [131] In any case free radicals thatoverride antioxidant defense react with a wide variety oforganic components causing lipid peroxidation to advancedlipid oxidation end products (ALE) cross-linking of proteinsnitrosylation of proteins and mutations in DNA Mitochon-drial circular DNA (mtDNA) of sim16 kbp is devoid of repairsystems meaning the buildup of lesions There are 37 geneslocated in mtDNA with those encoding protein subunits ofcomplex I (7) complex II (1) and complex IV (3) but notcomplex III Interestingly hallmarks of AD (APP A120573 andNFTs)mostly affectmembers of electron transfer chain (com-plexes I and IV) which rely exclusively on mitochondriallypredetermined subunits Maybe it is not simple coincidencebut the effect of inhibited import to mitochondria of nuclear-encoded subunits of complexes I and IV gives explanationfor toxicity of APP or A120573 or NFTs in mitochondria Thereare additional findings in mitochondria affected by AD suchas lower activity of pyruvate dehydrogenase (PDH) andoxoglutarate dehydrogenase (OGDH) [104 132] Inhibitionof OGDH the enzyme of tricarboxylic acid cycle minimizesthe NADH pool and electron number needed for ETCand mitochondrial membrane potential (Δ120595
119898) to create and
maintain proton gradient obligatory for ATP synthesisThereare also lines of evidence for direct inhibitory action of solubleoligomeric A120573 species on ABAD (A120573-binding alcohol dehy-drogenase) and internal membrane cyclophilin D (CypD)resulting in increased mitochondrial membrane permeabil-ity (MPTP) potentiated ROS production synaptic lossdiminished activity of mitochondrial respiration and finally
cell death [129 132ndash134] CypD knockout prevents mito-chondrial and neuronal perturbations and improves mito-chondrial function in Alzheimerrsquos disease mouse model [128135] It has to be emphasized that oxidativenitrosative stressaffects the fusion and fission process ofmitochondria Fusionthat is speedup by small GTPases mitofusins (Mfn1Mfn2)improves efficiency of mitochondrial respiration and ATPproduction Mitochondrial dynamics are severely imbal-anced in AD cases in favour of fission through elevatedexpression of the fission protein DLP1 (dynamin-like protein1) associatedwith nitrosative stress stirred up byA120573 [136 137]Collectively these observations indicate that toxic intracel-lular A120573
42-43 oligomers differ in action from extracellularaggregates found in amyloid plaques of AD brains [138](Figure 3)
12 Proteostasis in Mitochondria
Mitochondrial protein turnover grants the well-organizedreplacement of nonfunctional proteins into operational oneThe first task is attained through proteolytic degradation ofinner membrane and matrix proteins with local mitochon-drial proteases [139] or outer membrane proteins throughubiquitin-proteasome system [140] The second task is metby intramitochondrial protein synthesis but as almost 1500different nuclear-encoded proteins have to be importedto complete mitochondrial proteome a matter of capableimport is fundamental for the function of the organelleIn extreme cases of cellular injuries observed in neurode-generative diseases damaged mitochondria with extendedloss of the electrochemical potential are selectively removedby autophagy known as mitophagy [112] APP and A120573accumulation has also something to do with altered mito-chondrial dynamics as fission takes advantage of fusion withresultant dysfunction of mitochondria and neurons [137]AD neurons demonstrated selective mechanisms of prote-olytic clearance of oxidatively and nitrosatively modifiedproteins in mitochondria Insulin degrading enzyme (IDE)prevents formation of toxic insoluble fibrils from A120573 asit cleaves A120573 prior to aggregation [141ndash143] A novel zinc-metallopeptidase Presequence Protease (PreP) a member ofpitrilysin oligopeptidase family degrades either intramito-chondrially stored A120573
40or A120573
42proteinThis protease seems
to be highly sensitive to oxidative stress as disulfide bridgeformed between two proximal cysteine residues blocks itscatalytic activity [144ndash149] Another mitochondrial serineprotease HtrA2Omi occupies intermembrane space whereit can cleave APP locked up in protein translocase complex[121] Even though it is well established that HtrA2Omiis released to the cytosol to amplify apoptosis through thedegradation of antiapoptotic proteins and caspase activation[150ndash152] it is also implicated in proteolytic deletion ofmalfoldedAPP in ER [151] Fromknockout studies onmice itis obvious that HtrA2Omi plays a significant shielding roleas mice deficient in this protease exhibit neurodegenerativephenotype with weight loss and premature death [153]Taken together understanding the mechanisms of clearanceof the unwanted proteins including APP A120573 and tau isvital in an attempt to get rid of them from mitochondria
Oxidative Medicine and Cellular Longevity 9
VDAC
ANT
A120573
A120573
A120573
A120573
A120573
A120573
A120573A120573
A120573
A120573
A120573 CypD
CypD ProApCyt c Ca2+
IMM
OMM
IMSΔΨ
+
+
+
minus
minus
minus
ABAD
ROS
ROS
TIM
TOM
Proteinimport
NAD
NAD
NAD
NAD
H+
H+
H+
H+
H+
ADP
Mn-SOD
CuZn-SOD
mtDNA
V
Matrix
UCP
O2∙minus
O2∙minus
IV
III
II
I
O2
O2
DLP1Fis1
Mfn1Mfn2
Opa1
mHsp70
NADH
NADH
NADH
TCAcycle
PyruvateAcetyl-CoA
120572KGDH
120572-KG
PDH
Isocitrate
CitrateOAA
Malate
Fumarate
Succinyl-CoASuccinate
GDPGTP
FAD
FAOH
ATP
Figure 3 Amyloid-120573-related mitochondrial impairment Mitochondria were found to be the target for amyloid-120573 (A120573) which interactswith several proteins leading to mitochondrial dysfunction Indeed A120573 was found in the outer mitochondrial membrane (OMM) andinner mitochondrial membrane (IMM) as well as in the matrix The interaction of A120573 with the OMM affects the transport of nuclear-encoded mitochondrial proteins such as subunits of the electron transport chain complex IV into the organelle via the translocase of theouter membrane (TOM) import machinery Moreover A120573 disturbs the activity of several enzymes such as pyruvate dehydrogenase (PDH)and oxoglutarate dehydrogenase (OGDH) decreasing NADH reduction and the electron transport chain enzyme complex IV reducing theamount of hydrogen that is translocated from the matrix to the intermembrane space (IMS) thus impairing the mitochondrial membranepotential (MMP) Taken together these events cause abnormal mitochondrial electron activities leading to decreased complex V activityand so to a drop in ATP levels in addition to increasing reactive oxygen species (ROS) generation Moreover ROS induce peroxidationof several mitochondrial macromolecules such as mitochondrial DNA (mtDNA) and mitochondrial lipids contributing to mitochondrialimpairment in themitochondrial matrixThe complex of A120573 bound to binding alcohol dehydrogenase (ABAD) impairs the binding of NAD+to ABAD changes mitochondrial membrane permeability and reduces activities of respiratory enzymes inducing further ROS productionand leading to mitochondrial failure A120573 binding also activates Fis1 (fission protein) and promotes increased mitochondrial fragmentationthis increased mitochondrial fragmentation produces defective mitochondria that ultimately damage neurons Furthermore A120573 binding tocyclophilin D (CypD) enhances the protein translocation to the inner membrane favouring the opening of the mitochondrial permeabilitytransition pore formed by the adenine nucleotide translocator (ANT) and voltage-dependent anion channels (VDACs) Cyt 119888 cytochrome 119888DLP1 dynamin-like protein 1 PDH pyruvate dehydrogenase ProAp proapoptotic factors SOD superoxide dismutase TCA tricarboxylicacid TIM translocase of the inner membrane Adopted from [14]
Some hopes are associated with the application of at presentindefinite modulators of proteolytic activity HtrA2Omi isfor example activated by PTEN-induced putative kinase 1(PINK1) upon phosphorylation at Ser142 residue [154] Alter-natively accent has to be put on mitophagy of dysfunctionalmitochondria and mitochondriogenesis
13 Aging versus AD
Themost commonneurodegenerative disorder is representedby Alzheimerrsquos disease characterized by declining memoryreduced cognitive capacity and progressive dementia whichare often fatal to elderly individuals above 65 years of age
10 Oxidative Medicine and Cellular Longevity
It is ranked as the fourth leading cause of death in modernsocieties where average life span increased greatly in the lasttwo decades As 95 of AD cases are diagnosed in olderpeople one might think that a causal relationship existsbetween aging and the onset of disease Certainly a numberof similarities between getting old and being affected withAD could be listed Historically the free radical theory ofaging by Harman [155] suggested aging as ldquoside effectrdquo ofreactive oxygen species formed in mitochondrial respiratorychain Apparently free radicals commonly generated byincomplete reduction of oxygen molecule at complexes Iand III of mitochondrial ETC are capable of damagingDNA RNA and proteins They impair energy storage andlead to operational failure of mitochondria with progres-sive decline of cell viability Almost identical conditionsaccompany AD and are an explicit step in pathogenesisof disease As mtDNA is deprived of repair mechanismsROS-induced DNA strand breaks tend to accumulate withage or AD Thus mtDNA is a vulnerable target for ROSbut the reverse the ROS generation due to the mutatedmtDNA is not convincingly confirmed [156] Moreover theevidence that mutated mtDNA accelerates the progress ofaging is also questioned based on the results from studycarried out on transgenic mice model [157] Although someauthors show inconsistency between the free radical theoryand observations cumulative evaluation of the scientificreports points to antioxidant defense systems as importantfactors in protection from premature aging [158] Othermitochondrial components important for their function arehampered with age adenine nucleotide translocase (ANT)nitric oxide synthase (NOS) and carnitine acyltransferase(CT) [159ndash161] Actually NOS activity is elevated in AD asreported from study performed on cellular model of disease[15] We could not find any information about CT activityin AD whereas ANT activity is noticeably inhibited byA120573 or hyperphosphorylated tau and this effect is reversedby mersalyl a reversible alkylating agent of thiol groups[162] Mitochondrial dysfunction observed in transgenicmice models of AD and aging demonstrates higher activ-ity of genes controlling energy metabolism and apoptosisTaken together physiological aging and AD are associatedwith broad-spectrum dysfunction of mitochondria but thefoundations of mitochondrial decline are dissimilar Morediscrepancies between physiological aging and AD werefound with respect to ERS and UPR which play a significantrole in cellular proteostasis Dolichol was selected as an agingmarker because the progressive increase in dolichol levelwas observed in aging brain [163] In contrast ubiquinoneconcentration which is also synthesized from geranylgeranioldiminishes with agingwhereas cholesterol and dolichyl phos-phate concentrations remain unaltered In AD decreasedlevels of dolichol were observed and increased levels ofubiquinone and dolichyl phosphate without any changesin brain cholesterol AD cannot be regarded as a resultof premature aging The drop in dolichol and augmenteddolichyl phosphate concentration points toward disturbedglycosylation in ER of diseased brain while the increase inubiquinone suggests efforts to protect the brain from oxida-tive stress induced by lipid peroxidation [164 165]
14 Targeting ER Stress in AD Therapy
As ERS is a recognized factor in AD drugs that interfere withERS would theoretically have great therapeutic potentialThere are several compounds grouped in classes thatinteract directly with components of the ERS (salubrinalBiP inducer X (BIX) salicylamide analogs flavonoidsguanabenz and STF083010) chemical chaperons (4-phenyl-butyric acid (PBA) tauroursodeoxycholic acid (TUDCA)and trimethylamine oxide (TMAO)) chemicals that inhibitprotein degradation (Eeyarestatin MG132 and Bortezomib)compounds with antioxidant activity (Edaravone diben-zoylmethane derivatives and N-acetyl cysteine (NAC)) anddrugs controlling calcium signaling (dantrolene and carba-zole derivatives) [166] They may act by inducing transienttranslation arrest upregulation of chaperone proteins andaugmented degradation of ER-associated misfolded proteinsFundamental approach in the development of new therapyis the selection of appropriate molecular targets In ER stresssignaling the aim is to alter the expression of ER stress-associated molecules that can rescue cells from the toxiceffect of ERS Recent efforts in establishing new promisingdrugs against AD are pointing to chemical chaperones suchas PBA TUDCA or trimethylamine oxide (TMAO) Thesesubstances improve protein folding and alleviate nativeprotein conformation [166] It was shown on mouse modelsof AD that PBA TUDCA and TMAO stop A120573 accumulationand avoid the loss of dendritic spines [167] Some observa-tions even demonstrated improved memory and cognitivefunctions [168] associated with improved cell survival [169]Salubrinal ((2E)-3-phenyl-N-[222-trichloro-1-[[(8-quinoli-nylamino)thioxomethyl]amino]ethyl]-2-propenamide Sal)selectively inhibits growth arrest and DNA damage inducedgene 34- (GADD34-) phosphatase complex (GADD34associates with protein phosphatase 1 (PP1)) and promotes invitro dephosphorylation of the alpha subunit of eIF-2120572 andIRE1120572ASK1JNK signaling pathway being protective againstERS even induced by tunicamycin Tm [170] In a great deal ofexperiments testing Sal in cultured cells and animal modelsof AD this substance increased the viability of neuronal cellsand A120573 toxicity [171 172] BIX (2-(34-dihydroxyphenyl)-2-oxoethyl ester thiocyanic acid) preferentially inducedBiP mRNA in an ATF6-dependent manner leading toreduced Tm-induced death of neuronal cells [173] AlsoDBM derivative 14ndash26 (221015840-dimethoxydibenzoylmethane)was found to be neuroprotective for SH-SY5Y and PC-12cells by decreasing expression of BiP and CHOP [174]Dantrolene a ryanodine receptor antagonist that inhibitsabnormal calcium release from the ER inhibited expressionof both phosphorylated PERK and eIF2120572 It also reducedCHOP expression and attenuated thapsigargin-inducedapoptosis in PC-12 cells [175] Neuroprotective effects similarto dantrolene were observed for ([9-(3-cyanobenzyl)-14-dimethylcarbazole]) This substance suppressed increases inintracellular Ca2+ in PC-12 cells treated with thapsigarginand reduced levels of BiP and CHOP [176]
Aforementioned compounds were chosen from othersas the most potent ER stress inhibitors and persuasivelyprotective to neuronal cells 18 of 42 different compounds
Oxidative Medicine and Cellular Longevity 11
were exploited in in vivo and in vitro models of centralnervous system disorders with in fact improved cell or tissueviability [166] Thus the brain is the most frequently investi-gated organ in the context of ERS From these experimentsit becomes clear that CHOP functions as proapoptotic factorThe roles of other specific ER stress molecules as moleculartargets for pharmacological intervention are less clear andvary depending on cell type and context
There are few underestimated modulations in APP pro-cessing that shed more light on current dogma of AD patho-genesis The modulation of mevalonate pathway and choles-terol synthesis were reported to stimulate nonamyloidogenicpathway of APP processing [177] Additionally cholesterolderivative 27-hydroxycholesterol (27-OHC) was shown toinduce ER stress which attenuated leptin-dependent viabilityby activating CHOP in SH-SY5Y neuroblastoma cells [178]Irrespective of a number of compounds examined in AD thecall for new drugs modulating ER stress with healing effect isstill waiting to be revealed
Abbreviations
APOE Apolipoprotein EAPPs Secreted form of APPCHOPGADD153 CEBP homologous protein
growth arrest and DNA damageinduced gene 153
Drp1 Dynamin-1-like proteinERAD Endoplasmic-reticulum-
associated protein degradationERO1120572 ER oxidase 1120572ERS Endoplasmic reticulum stressFOH FarnesolGGOH GeranylgeraniolGRP75 Glucose-regulated protein 75GRP78BiP 78 kDa glucose-regulated proteinGSK-3120573 Synthase kinase 3 betaIP3R Inositol triphosphate receptor
IRE1120572 Inositol-requiring enzyme 1 alphaMAM Mitochondria-associated
membraneMAP LC3 Microtubule-associated protein
light chain 3Mfn2 GTPase mitofusin 2MMP Mitochondrial membrane
potentialMPTP Mitochondrial permeability
transition poreNFT Neurofibrillary tanglesPACS-2 Phosphofurin acidic cluster
sorting protein 2PDI Protein disulfide isomerasePERK Protein kinase R- (PKR-) like
ER kinaseRNS Reactive nitrogen speciesROS Reactive oxygen speciesRyR Ryanodine receptorTOM Transporter outer membraneTIM Transporter inner membrane
UPR Unfolded protein responseVDAC Voltage-dependent anion channelXBP-1 Xbox binding protein 1
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
Support for this work was provided by Grant no UMO-201103BNZ70238 from the National Science Centre inPoland
References
[1] R L Nussbaum andC E Ellis ldquoAlzheimerrsquos disease and Parkin-sonrsquos diseaserdquoTheNewEngland Journal ofMedicine vol 348 no14 pp 1356ndash1364 2003
[2] A L Brunkan and A M Goate ldquoPresenilin function and 120574-secretase activityrdquoThe Journal of Neurochemistry vol 93 no 4pp 769ndash792 2005
[3] V Crentsil ldquoThe pharmacogenomics of Alzheimerrsquos diseaserdquoAgeing Research Reviews vol 3 no 2 pp 153ndash169 2004
[4] D L Price and S S Sisodia ldquoMutant genes in familialAlzheimerrsquos disease and transgenic modelsrdquo Annual Review ofNeuroscience vol 21 pp 479ndash505 1998
[5] D Harman ldquoAlzheimerrsquos disease pathogenesis role of agingrdquoAnnals of the New York Academy of Sciences vol 1067 no 1 pp454ndash460 2006
[6] D J Selkoe ldquoTranslating cell biology into therapeutic advancesin Alzheimerrsquos diseaserdquo Nature vol 399 pp A23ndashA31 1999
[7] H Braak and E Braak ldquoNeuropathological stageing of Alz-heimer-related changesrdquo Acta Neuropathologica vol 82 no 4pp 239ndash259 1991
[8] K Hirai G Aliev A Nunomara et al ldquoMitochondrial abnor-malities inAlzheimerrsquos diseaserdquoTheJournal ofNeuroscience vol21 pp 3017ndash3023 2001
[9] U T Brunk C B Jones and R S Sohal ldquoA novel hypothesisof lipofuscinogenesis and cellular aging based on interac-tions between oxidative stress and autophagocytosisrdquoMutationResearchDNAging vol 275 no 3ndash6 pp 395ndash403 1992
[10] P A Stewart K Hayakawa M-A Akers and H V Vinters ldquoAmorphometric study of the blood-brain barrier in Alzheimerrsquosdiseaserdquo Laboratory Investigation vol 67 no 6 pp 734ndash7421992
[11] J P Blass and G E Gibson ldquoThe role of oxidative abnor-malities in the pathophysiology of Alzheimerrsquos diseaserdquo RevueNeurologique vol 147 no 6-7 pp 513ndash525 1991
[12] B Pająk E Kania and A Orzechowski ldquoNucleofection of ratpheochromocytoma PC-12 cells with human mutated beta-amyloid precursor protein gene (APP-sw) leads to reducedreduced viability autophagy-like process and increased expres-sion and secretion of beta amyloidrdquo BioMed Research Interna-tional vol 2015 Article ID 746092 10 pages 2015
[13] J P Blass ldquoCerebrometabolic abnormalities in AlzheimerrsquosdiseaserdquoNeurological Research vol 25 no 6 pp 556ndash566 2003
12 Oxidative Medicine and Cellular Longevity
[14] A Eckert K Schmitt and J Gotz ldquoMitochondrial dysfunc-tionmdashthe beginning of the end in Alzheimerrsquos disease Sep-arate and synergistic modes of tau and amyloid-120573 toxicityrdquoAlzheimerrsquos Research ampTherapy vol 3 article 15 pp 1ndash11 2011
[15] U Keil A Bonert C A Marques et al ldquoAmyloid 120573-inducedchanges in nitric oxide production and mitochondrial activitylead to apoptosisrdquoThe Journal of Biological Chemistry vol 279no 48 pp 50310ndash50320 2004
[16] A M Swomley and D A Butterfield ldquoOxidative stress inAlzheimer disease and mild cognitive impairment evidencefrom human data provided by redox proteomicsrdquo Archives ofToxicology vol 89 no 10 pp 1669ndash1680 2015
[17] C Caspersen N Wang J Yao et al ldquoMitochondrial A120573 apotential focal point for neuronal metabolic dysfunction inAlzheimerrsquos diseaserdquo The FASEB Journal vol 19 no 14 pp2040ndash2041 2005
[18] G E Gibson K-F R Sheu and J P Blass ldquoAbnormalities ofmitochondrial enzymes inAlzheimer diseaserdquo Journal of NeuralTransmission vol 105 no 8-9 pp 855ndash870 1998
[19] A Eckert S Hauptmann I Scherping et al ldquoSoluble beta-amyloid leads to mitochondrial defects in amyloid precursorprotein and tau transgenic micerdquo Neurodegenerative Diseasesvol 5 no 3-4 pp 157ndash159 2008
[20] A Eckert K L Schulz V Rhein and J Gotz ldquoConvergenceof amyloid-120573 and tau pathologies on mitochondria in vivordquoMolecular Neurobiology vol 41 no 2-3 pp 107ndash114 2010
[21] S Hauptmann I Scherping S Drose et al ldquoMitochondrial dys-function an early event in Alzheimer pathology accumulateswith age in AD transgenic micerdquoNeurobiology of Aging vol 30no 10 pp 1574ndash1586 2009
[22] M Manczak B S Park Y Jung and P H Reddy ldquoDifferentialexpression of oxidative phosphorylation genes in patients withAlzheimerrsquos disease implications for early mitochondrial dys-function and oxidative damagerdquo NeuroMolecular Medicine vol5 no 2 pp 147ndash162 2004
[23] M P Mattson M Gleichmann and A Cheng ldquoMitochondriain neuroplasticity and neurological disordersrdquo Neuron vol 60no 5 pp 748ndash766 2008
[24] P I Moreira M S Santos and C R Oliveira ldquoAlzheimerrsquosdisease a lesson frommitochondrial dysfunctionrdquoAntioxidantsamp Redox Signaling vol 9 no 10 pp 1621ndash1630 2007
[25] P H Reddy ldquoMitochondrial dysfunction in aging and Alz-heimerrsquos disease strategies to protect neuronsrdquo Antioxidants ampRedox Signaling vol 9 no 10 pp 1647ndash1658 2007
[26] V Rhein G Baysang S Rao et al ldquoAmyloid-beta leads toimpaired cellular respiration energy production andmitochon-drial electron chain complex activities in humanneuroblastomacellsrdquo Cellular and Molecular Neurobiology vol 29 no 6-7 pp1063ndash1071 2009
[27] V Rhein X Song A Wiesner et al ldquoAmyloid-120573 and tausynergistically impair the oxidative phosphorylation system intriple transgenic Alzheimerrsquos disease micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 47 pp 20057ndash20062 2009
[28] R X Santos S C Correia X Wang et al ldquoAlzheimerrsquos diseasediverse aspects ofmitochondrial malfunctioningrdquo InternationalJournal of Clinical and Experimental Pathology vol 3 no 6 pp570ndash581 2010
[29] N R Sims ldquoEnergy metabolism oxidative stress and neuronaldegeneration in Alzheimerrsquos diseaserdquoNeurodegeneration vol 5no 4 pp 435ndash440 1996
[30] H K Anandatheerthavarada G Biswas M-A Robin and NG Avadhani ldquoMitochondrial targeting and a novel transmem-brane arrest of Alzheimerrsquos amyloid precursor protein impairsmitochondrial function in neuronal cellsrdquo The Journal of CellBiology vol 161 no 1 pp 41ndash54 2003
[31] L Devi B M Prabhu D F Galati N G Avadhani and HK Anandatheerthavarada ldquoAccumulation of amyloid precur-sor protein in the mitochondrial import channels of humanAlzheimerrsquos disease brain is associated with mitochondrialdysfunctionrdquo The Journal of Neuroscience vol 26 no 35 pp9057ndash9068 2006
[32] L Devi and H K Anandatheerthavarada ldquoMitochondrialtrafficking of APP and alpha synuclein relevance to mito-chondrial dysfunction in Alzheimerrsquos and Parkinsonrsquos diseasesrdquoBiochimica et Biophysica Acta vol 1802 no 1 pp 11ndash19 2010
[33] M Manczak T S Anekonda E Henson B S Park J Quinnand P H Reddy ldquoMitochondria are a direct site of A120573 accumu-lation in Alzheimerrsquos disease neurons implications for free rad-ical generation and oxidative damage in disease progressionrdquoHuman Molecular Genetics vol 15 no 9 pp 1437ndash1449 2006
[34] P F Pavlov C H Petersen E Glaser and M AnkarcronaldquoMitochondrial accumulation of APP and A120573 significancefor Alzheimer disease pathogenesisrdquo Journal of Cellular andMolecular Medicine vol 13 no 10 pp 4137ndash4145 2009
[35] C Schmidt E Lepsverdize S L Chi et al ldquoAmyloid precursorprotein and amyloid 120573-peptide bind to ATP synthase andregulate its activity at the surface of neural cellsrdquo MolecularPsychiatry vol 13 no 10 pp 953ndash969 2008
[36] K Johar A Priya and M T T Wong-Riley ldquoRegulation ofNa+K+-ATPase by nuclear respiratory factor 1 implication inthe tight coupling of neuronal activity energy generation andenergy consumptionrdquo The Journal of Biological Chemistry vol287 no 48 pp 40381ndash40390 2012
[37] J Astrup P M Sorensen and H R Sorensen ldquoOxygen andglucose consumption related to Na+-K+ transport in caninebrainrdquo Stroke vol 12 no 6 pp 726ndash730 1981
[38] M Erecinska and I A Silver ldquoATP and brain functionrdquo Journalof Cerebral Blood Flow and Metabolism vol 9 no 1 pp 2ndash191989
[39] MMata D J Fink H Gainer et al ldquoActivity-dependent energymetabolism in rat posterior pituitary primarily reflects sodiumpump activityrdquo Journal of Neurochemistry vol 34 no 1 pp 213ndash215 1980
[40] J Kang H-G Lemaire A Unterbeck et al ldquoThe precursor ofAlzheimerrsquos disease amyloid A4 protein resembles a cell-surfacereceptorrdquo Nature vol 325 no 6106 pp 733ndash736 1987
[41] C L Masters and D J Selkoe ldquoBiochemistry of amyloid 120573-protein and amyloid deposits in Alzheimer diseaserdquoCold SpringHarbour Perspectives in Medicine vol 2 no 6 Article IDa006262 2012
[42] A Lai S S Sisodia and I S Trowbridge ldquoCharacterization ofsorting signals in the beta-amyloid precursor protein cytoplas-mic domainrdquo The Journal of Biological Chemistry vol 270 no8 pp 3565ndash3573 1995
[43] A I Placido C M F Pereira A I Duarte et al ldquoThe role ofendoplasmic reticulum in amyloid precursor protein processingand trafficking implications for Alzheimerrsquos diseaserdquo Biochim-ica et Biophysica ActamdashMolecular Basis of Disease vol 1842 no9 pp 1444ndash1453 2014
Oxidative Medicine and Cellular Longevity 13
[44] C Kaether P Skehel and C G Dotti ldquoAxonal membraneproteins are transported in distinct carriers a two-color videomicroscopy study in cultured hippocampal neuronsrdquoMolecularBiology of the Cell vol 11 no 4 pp 1213ndash1224 2000
[45] A Kamal A Almenar-Queralt J F LeBlanc E A Robertsand L S B Goldstein ldquoKinesin-mediated axonal transportof a membrane compartment containing 120573-secretase andpresenilin-1 requires APPrdquo Nature vol 414 no 6864 pp 643ndash648 2001
[46] R G Perez S Soriano J D Hayes et al ldquoMutagenesis identi-fies new signals for 120573-amyloid precursor protein endocytosisturnover and the generation of secreted fragments includingA12057342rdquo The Journal of Biological Chemistry vol 274 no 27 pp18851ndash18856 1999
[47] D G Cook M S Forman J C Sung et al ldquoAlzheimerrsquos A120573(1ndash42) is generated in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo Nature Medicine vol 3 no 9 pp1021ndash1023 1997
[48] A S C Chyung B D Greenberg D G Cook R W Domsand V M-Y Lee ldquoNovel 120573-secretase cleavage of 120573-amyloidprecursor protein in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo The Journal of Cell Biology vol138 no 3 pp 671ndash680 1997
[49] RW-Y Choy Z Cheng and R Schekman ldquoAmyloid precursorprotein (APP) traffics from the cell surface via endosomesfor amyloid 120573 (A120573) production in the trans-Golgi networkrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 30 pp E2077ndashE2082 2012
[50] S A Small and S Gandy ldquoSorting through the cell biologyof Alzheimerrsquos disease intracellular pathways to pathogenesisrdquoNeuron vol 52 no 1 pp 15ndash31 2006
[51] M Goedert ldquoAlzheimerrsquos and Parkinsonrsquos diseases the prionconcept in relation to assembled A120573 tau and 120572-synucleinrdquoScience vol 349 no 6248 2015
[52] I Benilova E Karran and B De Strooper ldquoThe toxic A120573oligomer and Alzheimerrsquos disease an emperor in need ofclothesrdquo Nature Neuroscience vol 15 no 3 pp 349ndash357 2012
[53] E H Corder A M Saunders W J Strittmatter et al ldquoGenedose of apolipoprotein E type 4 allele and the risk of Alzheimerrsquosdisease in late onset familiesrdquo Science vol 261 no 5123 pp 921ndash923 1993
[54] S Brunholz S Sisodia A Lorenzo C Deyts S Kins and GMorfini ldquoAxonal transport of APP and the spatial regulationof APP cleavage and function in neuronal cellsrdquo ExperimentalBrain Research vol 217 no 3-4 pp 353ndash364 2012
[55] G Thinakaran and E H Koo ldquoAmyloid precursor proteintrafficking processing and functionrdquo The Journal of BiologicalChemistry vol 283 no 44 pp 29615ndash29619 2008
[56] T Saitoh M Sundsmo J-M Roch et al ldquoSecreted formof amyloid beta protein precursor is involved in the growthregulation of fibroblastsrdquo Cell vol 58 no 4 pp 615ndash622 1989
[57] M PMattson B Cheng A R Culwell F S Esch I Lieberburgand R E Rydel ldquoEvidence for excitoprotective and intraneu-ronal calcium-regulating roles for secreted forms of the 120573-amyloid precursor proteinrdquo Neuron vol 10 no 2 pp 243ndash2541993
[58] K J Lee C E H Moussa Y Lee et al ldquoBeta amyloid-independent role of amyloid precursor protein in generationandmaintenance of dendritic spinesrdquoNeuroscience vol 169 no1 pp 344ndash356 2010
[59] H Meziane J-C Dodart C Mathis et al ldquoMemory-enhancingeffects of secreted forms of the 120573-amyloid precursor proteinin normal and amnestic micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no21 pp 12683ndash12688 1998
[60] P R Turner K OrsquoConnor W P Tate and W C AbrahamldquoRoles of amyloid precursor protein and its fragments inregulating neural activity plasticity and memoryrdquo Progress inNeurobiology vol 70 no 1 pp 1ndash32 2003
[61] D Beher L Hesse C L Masters and GMulthaup ldquoRegulationof Amyloid Protein Precursor (APP) binding to collagen andmapping of the binding sites on APP and collagen type Irdquo TheJournal of Biological Chemistry vol 271 no 3 pp 1613ndash16201996
[62] S L Sabo A F Ikin J D Buxbaum and P Greengard ldquoTheamyloid precursor protein and its regulatory protein FE65 ingrowth cones and synapses in vitro and in vivordquo The Journal ofNeuroscience vol 23 no 13 pp 5407ndash5415 2003
[63] D H Small V Nurcombe G Reed et al ldquoA heparin-bindingdomain in the amyloid protein precursor of Alzheimerrsquos diseaseis involved in the regulation of neurite outgrowthrdquoThe Journalof Neuroscience vol 14 no 4 pp 2117ndash2127 1994
[64] T G Williamson S S Mok A Henry et al ldquoSecreted glypicanbinds to the amyloid precursor protein of Alzheimerrsquos disease(APP) and inhibits APP-induced neurite outgrowthrdquoThe Jour-nal of Biological Chemistry vol 271 no 49 pp 31215ndash312211996
[65] T Ondrejcak I Klyubin N-W Hu A E Barry W K Cullenand M J Rowan ldquoAlzheimerrsquos disease amyloid 120573-protein andsynaptic functionrdquo NeuroMolecular Medicine vol 12 no 1 pp13ndash26 2010
[66] D Puzzo L Privitera and A Palmeri ldquoHormetic effect ofamyloid-beta peptide in synaptic plasticity and memoryrdquo Neu-robiology of Aging vol 33 no 7 pp 1484e15ndash1484e24 2012
[67] C M F Pereira ldquoCrosstalk between endoplasmic reticulumstress and protein misfolding in neurodegenerative diseasesrdquoISRN Cell Biology vol 2013 Article ID 256404 22 pages 2013
[68] R-W Shin T C Saido M Maeda and T Kitamoto ldquoNovel 120572-secretase cleavage ofAlzheimerrsquos amyloid120573 precursor protein inthe endoplasmic reticulum of COS7 cellsrdquoNeuroscience Lettersvol 376 no 1 pp 14ndash19 2005
[69] E A Schon and E Area-Gomez ldquoMitochondria-associatedER membranes in Alzheimer diseaserdquo Molecular and CellularNeuroscience vol 55 pp 26ndash36 2013
[70] R Rizzuto P Pinton W Carrington et al ldquoClose contacts withthe endoplasmic reticulum as determinants of mitochondrialCa2+ responsesrdquo Science vol 280 no 5370 pp 1763ndash1766 1998
[71] S E Logue P Cleary S Saveljeva and A Samali ldquoNewdirections in ER-stress induced cell deathrdquoApoptosis vol 18 no5 pp 537ndash546 2013
[72] J P Decuypere KWelkenhuyzen T Luyten et al ldquoIns(145)P3receptor-mediated BI-1 Ca2+ signaling and autophagy induc-tion are interrelatedrdquo Autophagy vol 7 pp 1472ndash1489 2011
[73] K S Vetrivel and G Thinakaran ldquoMembrane rafts in Alz-heimerrsquos disease beta-amyloid productionrdquo Biochimica et Bio-physica Acta vol 1801 no 8 pp 860ndash867 2010
[74] H Urra E Dufey F Lisbona D Rojas-Rivera and C HetzldquoWhen ER stress reaches a dead endrdquo Biochimica et BiophysicaActa (BBA) Molecular Cell Research vol 1833 no 12 pp 3507ndash3517 2013
14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 5
levels stimulate autophagyThe molecular mechanism of ER-mediated autophagy is accurately regulated by Beclin 1 as wellas ER membrane bound protein Bax inhibitor 1 (BI-1) Bothproteins are capable of promoting autophagy through IP
3R-
dependent mechanism [72]
5 APP Processing and ER Stress Response
The nature of APP processing is determined by the com-position of membrane with cholesterol rich lipid rafts asthe site of amyloidogenic cleavage [73] Consequently onemight expect that fate of APP is at least partly establishedby the representation of lipid rafts and possible access to 120573-versus120572-secretase Irrespective of the type of cleavage there isone underestimated biochemical step in ER that might bringabout damaging upshot It is apparently the effectiveness ofER situated N-glycosylation which makes the APP moleculesuitably folded If misfoldedmalfolded protein(s) accumu-late in the ER complex cascade of reactions known as theunfolded protein response (UPR) is triggered with the so-called endoplasmic reticulum stress response (ERS) Today itis widely accepted that during UPR the ER sensors proteinkinase R- (PKR-) like ER kinase (PERK) activating proteinkinase 6 (ATF6) and inositol-requiring enzyme 1 alpha(IRE1120572) are freed fromGRP78BiP protein repression whichhereafter become activated Without going into details ofindividual sensor action a number of reactions occur at boththe genomic and cytoplasmic level with selective degradationof mRNAs encoding protein(s) with abnormal folding andinhibition of protein translation except for genes importantfor UPR redox homeostasis energy metabolism and pro-tein folding [67] On the one hand PERK phosphorylateseukaryotic initiation factor 2 alpha (eIF2120572) to stop entrance ofmethionyl-tRNA to the ribosome on the other hand it allowstranslation of activating transcription factor 4 (ATF4) geneIRE1120572 sets off alternative splicing of Xbox binding protein 1(XBP1) transcript leading to activation of the transcriptionfactor liable to stimulate ERGolgi biogenesis and formationof proteins involved in endoplasmic-reticulum-associatedprotein degradation known as ERAD (Erdj4 p58IPK EDEMRAMP-4 PDI-P5 and HEDJ for details see [67]) FinallyATF6 is activated in the Golgi complex through proteolyticcleavage and translocates to nucleus where it cooperates withXBP1 in upregulation of chaperones and ERAD-related genes[43] In principle UPR is activated to restore ER homeostasisand stop the accumulation of aberrantly formed protein(s)but if the strength of ER stress is unbearable (meaning thatit cannot be compensated by UPR) there is a path to activateapoptosis Among other routes the most central role isplayed by the major proapoptotic transcription factor CEBPhomologous protein CHOPgrowth arrest and DNA damageinduced gene 153 GADD153 It downregulates the antiapop-totic protein Bcl-2 and upregulates proapoptotic Bax and Bak[74] CHOPGADD153 leads to excessive production of reac-tive oxygen species (ROS) within ER subsequent depletionof reduced glutathione (GSH) and Ca2+ flux from the ER tocytoplasm through the IP
3R [75] One hitherto unresolved
issue is the physiologic importance of APP and GRP78BiPinteraction disclosed in coprecipitation study carried out by
Yamamoto et al [76] Bulk ofAPP associatedwithGRP78BiPwas immature protein Given that GRP78BiP expressionlevels declined in samples of brain tissue obtained fromFAD patients as demonstrated by Katayama et al [77] fewdistinct scenarios are possible First binding to GRP78BiPsuggests ER accumulation of immature APP Second APPinteraction with GRP78BiP is a noticeable sign of UPRwhich is further validated by downregulation of GRP78BiPin FAD patients Finally retention of APP in ER is mostlikely a result of amyloidogenic APP processing and as such itprobably occurs in MAM the cholesterol enriched domainsER-mitochondria crossing point is therefore of particularinterest in deciphering the links between the ER placed APPprocessing UPR and resulting cellular responses such asautophagy apoptosis and inflammatory reaction observed inAD (Figure 2)
6 Pathology of Tau Protein
Microtubule-associated protein tau controls assembly andprevents microtubules from severing Microtubular networkis fundamental component of cytoskeleton essential forintracellular transport of secretory vesicles and organelles(ie mitochondria) Glycogen synthase kinase 3 beta- (GSK-3120573-) targeted hyperphosphorylation of tau causes this proteinto dissociate frommicrotubules Consequently microtubulesbecome fragmented and microtubule-dependent transportsystem fails Furthermore hyperphosphorylated tau (P-tau)is prone to form oligomers and toxic filaments known asNFTs or tauopathy [78] A variety of tau conformers werereported to exist pointing to different tauopathies capa-ble of self-propagation [79 80] Interestingly intracellulartau inclusions define AD as clinical symptoms of diseaseobserved when tauopathy is abundant together with intra-cellular A120573 deposits in neocortex [51 81 82] People withabundant A120573 plaques but no or only a few neurofibrillarylesions do not have AD Clinicopathological correlationstudies have been crucial to generate hypotheses about thepathophysiology of the disease by establishing the fact thatthere is a continuum between ldquonormalrdquo aging and ADdementia and that the amyloid plaque buildup occurs primar-ily before the onset of cognitive deficits while neurofibrillarytangles neuron loss and particularly synaptic loss parallel theprogression of cognitive decline [83] Thus misfolded pro-teins and descendant toxic filaments with a number of inter-mediates are critical for manifestation of AD as fibrillogenicAPP processing is not enough for onset of disease Althoughthe molecular mechanism of tauopathy is not deciphered infull recent reports suggest ER stress as the starting point[84 85] This idea is substantiated by the elevated levels ofERS and UPR markers together with P-tau and GSK-3120573 inbrains affected by AD [86] Therefore on the one hand theincidence of UPR is strongly correlated with the presence ofNFTs on the other hand aggregation of P-tau induces ERSwith resultant UPR Some lines of evidence confirmed UPRactivation near the beginning of NFTs formation and pointto the functional link between malformed tau protein andUPRThe in vitro experiments with phosphatase 2A inhibitoror phosphorylation activator demonstrated enhanced P-tau
6 Oxidative Medicine and Cellular Longevity
formation in neurons in concert with the increased levelsof PERK eIF2120572 and XBP1 transcript apparent markers ofUPR [87] Furthermore GRP78BiP was found to encouragetau phosphorylation through facilitated substrate capture byGSK-3120573 [88] GSK-3120573 seems to play dual role first this kinaseprotects neurons from apoptosis as P-tau accumulation isstrong molecular signal to trigger UPR with subsequentautophagy Second UPR raises GSK-3120573 activity throughlysosomal degradation of inactive GSK-3120573 (P-Ser9-GSK-3120573)To sum up ERS and UPR are important molecular machinesused to prevent cell viability turned on by tauopathy
7 Oxidative Stress in ER
Sacs and tubes of ER delineate the compartment where thenewly synthesized proteins undergo maturation to nativestate Native state indicates properly folded fully functionalprotein Important reactions essential for protein folding ofunbranched polypeptide chains include amino acids oxida-tion and glycosylation As a result redox homeostasis inER is shifted to oxidative state so as to promote disulfidebond formation between adjacent cysteines Oxidation ofsulfhydryl groups required to make disulfide bonds is con-trolled by ER oxidase 1120572 (ERO1120572) Next disulfide bondscould be subject to posttranslational modification disulfideexchange by protein disulfide isomerase (PDI) PDI is ableto correct mispaired thiol residues by catalyzing the breakageand formation of correct disulfide bonds These enzymes arefundamental for protein folding Oxidation allows twistingof proteins which is followed by N-glycosylation andor C-and O-mannosylation Protein N-glycosylation in eukary-otes is a complex process divided into several steps Firstthere is a ldquocall forrdquo carrier lipids (polyisoprenyl phosphatessuch as dolichyl phosphates) the membrane lipids knownto function as glycosyl transporters In mammalian cellsthe limiting substrate for dolichol biosynthesis is geranyl-geraniol (GGOH) of mevalonate pathway Dolichols thelongest aliphatic molecules synthesized in animal cellshave 18ndash21 120572-isoprene saturated units (C90ndash105) criticalfor their recognition by the enzymes (glycosyltransferases)that glucosylate dolichyl phosphates [89] Once dolichylmonophosphates (Dol-P) are formed in the ER membranethe precursor oligosaccharide donor (GLc
3Man9GLCNAc
2-
P-P-dolichol) for protein N-glycosylation can be synthesizedon the lumenal leaflet of ER First three sugar intermediatesare produced (Man-P-Dol Glc-P-Dol GlcNAc-P-P-Dol andMan5GlcNAc-P-P-Dol) on the cytoplasmic leaflet of the
ER Next enzyme flippases mediate transbilayer movementof the aforementioned intermediates to lumenal side ofER where conversion to Glc
3Man9GlcNAc
2-P-P-Dol could
be completed Glc3Man9GlcNAc
2-P-P-Dol is also used for
biosynthesis of glycosylphosphatidylinositol (GPI) anchorsTaken together lipid-mediated glycosylation plays a vital
role in the appropriate protein folding and intracellulartranslocation of N-linked glycoproteins [90] Likewise it isimportant for protein O- and C-mannosylation and GPIanchorage Moreover Dol-P availability in the ER is the rate-limiting factor in the production of glycolipid intermediatesand N-glycosylation
8 ER Stress and Apoptosis
It was shown that during ERS the ER resident proapop-totic cysteine protease known as caspase (caspase-12 in ratcaspase-4 in humans) is activated through cleavage As aresult caspase cascade is started via caspase-9 that in turnstimulates effector caspase-3 [91 92] The central role playedby ER in programmed cell death is achieved by PERK branchwhere ATF4 induces the expression of CHOPGADD153which represses antiapoptotic Bcl-2 family proteins andsimultaneously shuffles ER membrane Bax and Bak proteinsinto outer mitochondrial membrane Consequently poresare formed to leak the components of apoptosome frommitochondrial intermembrane space [93] Another impor-tant mechanism of ERS-induced apoptosis is led by Ca2+-dependent ERO1120572-IP
3R pathway where ERO1120572 collaborates
with IP3R in Ca2+ efflux from ER to mitochondria via MAM
[75 94] Accordingly Ca2+ influx facilitates cytochrome 119888release frommitochondria besides cytochrome 119888 can bind toER IP
3R and the complex amplifies the apoptotic signal in a
feedforward manner [95] Last but not least ERS-associatedapoptotic programme is set off by c-Jun N-terminal kinase(JNK) as the effect of IRE1120572 complexed with TNF-receptor-associated factor 2 (TRAF2) activation of apoptosis-signal-regulating kinase 1 (ASK1) [96 97]
What does ERS drive to induce apoptotic death inneurons Actually many reports indicate that APP and A120573 aswell as hyperphosphorylated tau have been shown to blockmitochondrial transport which results in impaired energystorage and oxidative stress [98ndash101] Indeed accumulationof APP A120573 and NFTs in mitochondria led to reducedactivity of some enzymes involved in substrate oxidation(tricarboxylic acid cycle) electron transport chain (ETC)and ATP synthase as well as severely diminishing import ofnuclear-encoded proteins [26 27 102ndash104] One may ask ifthere is any additional link between ER and mitochondriaother than MAM which could account for apoptotic signalThough not directly ER significantly contributes to oxidativestress in mitochondria of AD-affected subjects
9 Abnormal APP Processing and TraffickingCulminate in ER Pathology of AD
From the morphological point of view as neurons arehighly specialized cells soma dendrites and neurites areconsiderably distinct structures Proteins needed by thesecompartments are delivered via microtubules once proteinshave suitable sorting signals (ie APP trafficking from ERto plasma membrane is associated with several posttrans-lational modifications with oxidation and N-glycosylation)Additionally the Golgi apparatus follows ER in subsequentAPP adjustment (O- and N-glycosylation phosphorylationand sulphonation) [42] Any inaccurate alteration of the APPmolecule is potentially hazardous as protein final destiny ismissed causing its retention in ER or trans-Golgi network(TGN) In addition other unusual settings for APP arepossible as this large protein has few signal sequences hiddenwhen APP is correctly folded In the cells transfected withAPP this protein enters coat protein complex I (COPI)
Oxidative Medicine and Cellular Longevity 7
vesicles and undergoes retrograde transport from cis endof the Golgi complex back to the ER [76] Such responsecauses accumulation of APP in the tubulocisternal ER systemtogether with aberrant intracellular translocation of theprotein Interestingly the issue whether APP is subject toretrograde transport with successive fibrillogenic processingbecause of UPR and its interaction with GRP78BiP is notclear as the observations are inconsistent [77] Nonethelessaccumulation of misfoldedmalfolded proteins in the ERsuggests disorganized process of posttranslational change Asanticipated the accretion of proteins of anomalous patternsignals ERS and UPR followed by increased vulnerability toapoptotic cell death Prior to decay however APP proteinlevels in the ER lessen by dint of cleavage with 120573- and120574-secretase [105 106] Products of this cleavage (sAPP120573A120573 and AICD fragment) all appreciably influence neuronalsurvival most likely through nonnative form of the APPsubstrate
10 APP A120573 and NFTs MarkMitochondria as Targeted in AD
Mitochondrial import of A12057340
and A12057342
peptides throughthe translocase TOM complex was blocked by preincubationof isolated mitochondria with antibodies raised againstTOM proteins (TOM20 TOM40 and TOM70) [107]Neither VDAC inhibition with antagonist antibodies norinhibition of mitochondrial permeability transition pores(MPTP) or fall of mitochondrial membrane potential(MMP) affected uptake of A120573 [17] With regard to APPelegant study performed by Anandatheerthavarada andhis colleagues [30] revealed that C-terminal truncatedAPP (lacking A120573) targets mitochondria in cholinergicGABAergic dopaminergic and glutamatergic neurons ofAD brain by clogging up mitochondrial protein translocasecomplex TOM40TIM22 Authors propose that occlusionof translocase is followed by blunted import of nuclear-encoded proteins vitally important for energy homeostasisMitochondrial APP protein transmembrane orientationindicates NH
2-terminal inside in contact with translocase
whereas COOH-terminal is facing cytoplasmic side NH2-
terminus has mitochondrial signal sequence Astonishinglymitochondrial APP molecules were nonglycosylated givingrise to speculation that protein molecules that arrivedat mitochondria have not achieved molecular maturity[31] The accumulation of nonglycosylated APP species inmitochondrial import channels of AD brain was directlyrelated to decreased mitochondrial functions as validatedby the decline in cytochrome 119888 oxidase activity (complexIV) and elevated levels of H
2O2 Furthermore in AD
brain the mitochondrial accumulation of nonglycosylatedAPP went along with a corresponding reduction in plasmamembrane-associatedAPP It suggests that ADbrain hasAPPprocessing and trafficking severely affected by incompleteN-glycosylation ensuing ER protein accumulation andexposure of the cryptic mitochondrial targeting signalfor assisting chaperone proteins Several lines of evidenceindicate that not fully formed proteins are phosphorylatedand bind to the cytosolic proteins required for movement
from ER to mitochondria [108ndash110] Moreover ERS andUPR in AD seem to be incompetent and inefficient inelimination of malfolded proteins In addition to APPalso A120573 was frequently reported to occupy mitochondriaalthough its origin and mechanism of mitochondriatargeting mostly remain unexplored (except for involvementof protein translocase TOM complex) The possibilityof mitochondrial A120573 generation has to be ruled out asmembrane orientation of arrested APP does not allow accessto 120574-secretase (120574-secretase activity in mitochondria wasdetected by independent study) [111] Probably A120573 speciesis derived from APP prior to its translocation (ER) soA120573 may be transported to mitochondria independentlyof APP Collectively observations showing mitochondrialpresence of APP A120573 and tau in aberrant configuration pointtoward the anomaly of protein folding at the level of ER andGolgi apparatus Nonglycosylated molecule of APP suggestsdefective transfer of sugar core from dolichyl phosphate(s)and further modifications such as O- and N-glycosylationsOne may admit that the lack of glycosyl residue bringsabout pathologic processing and trafficking of APP and itsfragments In point of fact dolichol derivatives mixture ofpolyprenols (acyclic isoprenoid alcohols) known as Ropren(Solagran Limited Melbourne Australia) commerciallyused to treat liver diseases were tested in the treatment ofAD in two separate trials conducted in 2005 and 2006 withpromising results [httpwwwasxcomauasxpdf20071119pdf315x5nh4hm8wv7pdf httpwwwasxcomauasxpdf20070221pdf3111ztwcbqzkk9pdf] Further studies areurgently needed to test how important the glycosylationprocess is in the pathogenesis of AD Dolichols areobtained from geranylgeraniol (GGOH) and the latter is anintermediate of mevalonate pathway GGOH is a commonsubstrate for dolichol and ubiquinone synthesis but it isalso necessary for protein prenylation As both GGOHand farnesol (FOH) are engaged in protein prenylationmore concern should be laid on the importance of thesecompounds in AD pathogenesis Observations demonstrat-ing mitochondrial relocalization of other proteins withoutposttranslational modification seemingly point to increasedmitochondrial targeting of immature molecules resultingin mitochondrial dysfunction and acceleration of diseaseprogression [34 112ndash114]
11 Oxidative Stress in Mitochondria ofAD Brains
Normal physiological functions of APP are thought to beinvolved in the stabilizing contact points between synapsesand maintaining mitochondrial functions [60 115] Mito-chondrial dysfunction was often observed regardless of theexperimental model used to study AD [8 29 116ndash119]It includes defects in oxidative phosphorylation decreasedATP decreased membrane potential increased productionof ROSRNS and perturbation in mitochondrial fusion andfission [15 30 31 115 120ndash122] Hyperphosphorylated tau wasalso reported to impair mitochondrial functions [123] Usingproteomic approach the strongest defects of the respiratorycapacity were observed mainly at complexes I IV and ATP
8 Oxidative Medicine and Cellular Longevity
synthase (complex V) at both protein and activity level [124]While APP A120573 and hyperphosphorylated tau are potentinhibitors of mitochondrial import of nuclear-encoded pro-teins apparently the pathology of each leads to metabolicharm in different way In freshly isolated mitochondriafrom AD brains the APP inhibited mitochondrial importof cytochrome 119888 oxidase (COX) subunits IV and Vb [31]Dysfunction of COX increases ROS production (incompletereduction of oxygen molecules) reduces energy stores anddisturbs energy metabolism Accordingly in AD patientsdeficiency of COX was found in brains and platelets [119125 126] Similar to APP A120573 was found in mitochondriaof transgenic mice and cellular and human AD models [1719 33 103 127ndash130] At present it is not clear whether theobservedmitochondrial toxicity is due toAPP orA120573 orNFTsaccumulation Anyway some regularity is observed withrespect to most affected components of respiratory chainNADH-ubiquinone oxidoreductase (complex I) activity isreduced to the utmost by hyperphosphorylated tau whereasdecreased activity of cytochrome 119888 oxidase (complex IV)that resulted in mitochondrial dysfunction was observedduring A120573 and APP accumulation [14 15 31 32 34]Concomitantly rise in the activity of antioxidant enzymesmanganese superoxide dismutase (Mn-SOD) and catalase(CAT) was demonstrated in response to elevated levelsof free radicals including superoxide anion radical (O
2
∙minus)hydroperoxyl radical (HO
2
∙) hydroxyl radical (OH∙) andnitric oxide radical (NO∙) [15] Consistent with observationsof chronic respiratory chain dysfunction and mitochondrialoxidative stress there are reports showing their contributionto tau pathology in AD [131] In any case free radicals thatoverride antioxidant defense react with a wide variety oforganic components causing lipid peroxidation to advancedlipid oxidation end products (ALE) cross-linking of proteinsnitrosylation of proteins and mutations in DNA Mitochon-drial circular DNA (mtDNA) of sim16 kbp is devoid of repairsystems meaning the buildup of lesions There are 37 geneslocated in mtDNA with those encoding protein subunits ofcomplex I (7) complex II (1) and complex IV (3) but notcomplex III Interestingly hallmarks of AD (APP A120573 andNFTs)mostly affectmembers of electron transfer chain (com-plexes I and IV) which rely exclusively on mitochondriallypredetermined subunits Maybe it is not simple coincidencebut the effect of inhibited import to mitochondria of nuclear-encoded subunits of complexes I and IV gives explanationfor toxicity of APP or A120573 or NFTs in mitochondria Thereare additional findings in mitochondria affected by AD suchas lower activity of pyruvate dehydrogenase (PDH) andoxoglutarate dehydrogenase (OGDH) [104 132] Inhibitionof OGDH the enzyme of tricarboxylic acid cycle minimizesthe NADH pool and electron number needed for ETCand mitochondrial membrane potential (Δ120595
119898) to create and
maintain proton gradient obligatory for ATP synthesisThereare also lines of evidence for direct inhibitory action of solubleoligomeric A120573 species on ABAD (A120573-binding alcohol dehy-drogenase) and internal membrane cyclophilin D (CypD)resulting in increased mitochondrial membrane permeabil-ity (MPTP) potentiated ROS production synaptic lossdiminished activity of mitochondrial respiration and finally
cell death [129 132ndash134] CypD knockout prevents mito-chondrial and neuronal perturbations and improves mito-chondrial function in Alzheimerrsquos disease mouse model [128135] It has to be emphasized that oxidativenitrosative stressaffects the fusion and fission process ofmitochondria Fusionthat is speedup by small GTPases mitofusins (Mfn1Mfn2)improves efficiency of mitochondrial respiration and ATPproduction Mitochondrial dynamics are severely imbal-anced in AD cases in favour of fission through elevatedexpression of the fission protein DLP1 (dynamin-like protein1) associatedwith nitrosative stress stirred up byA120573 [136 137]Collectively these observations indicate that toxic intracel-lular A120573
42-43 oligomers differ in action from extracellularaggregates found in amyloid plaques of AD brains [138](Figure 3)
12 Proteostasis in Mitochondria
Mitochondrial protein turnover grants the well-organizedreplacement of nonfunctional proteins into operational oneThe first task is attained through proteolytic degradation ofinner membrane and matrix proteins with local mitochon-drial proteases [139] or outer membrane proteins throughubiquitin-proteasome system [140] The second task is metby intramitochondrial protein synthesis but as almost 1500different nuclear-encoded proteins have to be importedto complete mitochondrial proteome a matter of capableimport is fundamental for the function of the organelleIn extreme cases of cellular injuries observed in neurode-generative diseases damaged mitochondria with extendedloss of the electrochemical potential are selectively removedby autophagy known as mitophagy [112] APP and A120573accumulation has also something to do with altered mito-chondrial dynamics as fission takes advantage of fusion withresultant dysfunction of mitochondria and neurons [137]AD neurons demonstrated selective mechanisms of prote-olytic clearance of oxidatively and nitrosatively modifiedproteins in mitochondria Insulin degrading enzyme (IDE)prevents formation of toxic insoluble fibrils from A120573 asit cleaves A120573 prior to aggregation [141ndash143] A novel zinc-metallopeptidase Presequence Protease (PreP) a member ofpitrilysin oligopeptidase family degrades either intramito-chondrially stored A120573
40or A120573
42proteinThis protease seems
to be highly sensitive to oxidative stress as disulfide bridgeformed between two proximal cysteine residues blocks itscatalytic activity [144ndash149] Another mitochondrial serineprotease HtrA2Omi occupies intermembrane space whereit can cleave APP locked up in protein translocase complex[121] Even though it is well established that HtrA2Omiis released to the cytosol to amplify apoptosis through thedegradation of antiapoptotic proteins and caspase activation[150ndash152] it is also implicated in proteolytic deletion ofmalfoldedAPP in ER [151] Fromknockout studies onmice itis obvious that HtrA2Omi plays a significant shielding roleas mice deficient in this protease exhibit neurodegenerativephenotype with weight loss and premature death [153]Taken together understanding the mechanisms of clearanceof the unwanted proteins including APP A120573 and tau isvital in an attempt to get rid of them from mitochondria
Oxidative Medicine and Cellular Longevity 9
VDAC
ANT
A120573
A120573
A120573
A120573
A120573
A120573
A120573A120573
A120573
A120573
A120573 CypD
CypD ProApCyt c Ca2+
IMM
OMM
IMSΔΨ
+
+
+
minus
minus
minus
ABAD
ROS
ROS
TIM
TOM
Proteinimport
NAD
NAD
NAD
NAD
H+
H+
H+
H+
H+
ADP
Mn-SOD
CuZn-SOD
mtDNA
V
Matrix
UCP
O2∙minus
O2∙minus
IV
III
II
I
O2
O2
DLP1Fis1
Mfn1Mfn2
Opa1
mHsp70
NADH
NADH
NADH
TCAcycle
PyruvateAcetyl-CoA
120572KGDH
120572-KG
PDH
Isocitrate
CitrateOAA
Malate
Fumarate
Succinyl-CoASuccinate
GDPGTP
FAD
FAOH
ATP
Figure 3 Amyloid-120573-related mitochondrial impairment Mitochondria were found to be the target for amyloid-120573 (A120573) which interactswith several proteins leading to mitochondrial dysfunction Indeed A120573 was found in the outer mitochondrial membrane (OMM) andinner mitochondrial membrane (IMM) as well as in the matrix The interaction of A120573 with the OMM affects the transport of nuclear-encoded mitochondrial proteins such as subunits of the electron transport chain complex IV into the organelle via the translocase of theouter membrane (TOM) import machinery Moreover A120573 disturbs the activity of several enzymes such as pyruvate dehydrogenase (PDH)and oxoglutarate dehydrogenase (OGDH) decreasing NADH reduction and the electron transport chain enzyme complex IV reducing theamount of hydrogen that is translocated from the matrix to the intermembrane space (IMS) thus impairing the mitochondrial membranepotential (MMP) Taken together these events cause abnormal mitochondrial electron activities leading to decreased complex V activityand so to a drop in ATP levels in addition to increasing reactive oxygen species (ROS) generation Moreover ROS induce peroxidationof several mitochondrial macromolecules such as mitochondrial DNA (mtDNA) and mitochondrial lipids contributing to mitochondrialimpairment in themitochondrial matrixThe complex of A120573 bound to binding alcohol dehydrogenase (ABAD) impairs the binding of NAD+to ABAD changes mitochondrial membrane permeability and reduces activities of respiratory enzymes inducing further ROS productionand leading to mitochondrial failure A120573 binding also activates Fis1 (fission protein) and promotes increased mitochondrial fragmentationthis increased mitochondrial fragmentation produces defective mitochondria that ultimately damage neurons Furthermore A120573 binding tocyclophilin D (CypD) enhances the protein translocation to the inner membrane favouring the opening of the mitochondrial permeabilitytransition pore formed by the adenine nucleotide translocator (ANT) and voltage-dependent anion channels (VDACs) Cyt 119888 cytochrome 119888DLP1 dynamin-like protein 1 PDH pyruvate dehydrogenase ProAp proapoptotic factors SOD superoxide dismutase TCA tricarboxylicacid TIM translocase of the inner membrane Adopted from [14]
Some hopes are associated with the application of at presentindefinite modulators of proteolytic activity HtrA2Omi isfor example activated by PTEN-induced putative kinase 1(PINK1) upon phosphorylation at Ser142 residue [154] Alter-natively accent has to be put on mitophagy of dysfunctionalmitochondria and mitochondriogenesis
13 Aging versus AD
Themost commonneurodegenerative disorder is representedby Alzheimerrsquos disease characterized by declining memoryreduced cognitive capacity and progressive dementia whichare often fatal to elderly individuals above 65 years of age
10 Oxidative Medicine and Cellular Longevity
It is ranked as the fourth leading cause of death in modernsocieties where average life span increased greatly in the lasttwo decades As 95 of AD cases are diagnosed in olderpeople one might think that a causal relationship existsbetween aging and the onset of disease Certainly a numberof similarities between getting old and being affected withAD could be listed Historically the free radical theory ofaging by Harman [155] suggested aging as ldquoside effectrdquo ofreactive oxygen species formed in mitochondrial respiratorychain Apparently free radicals commonly generated byincomplete reduction of oxygen molecule at complexes Iand III of mitochondrial ETC are capable of damagingDNA RNA and proteins They impair energy storage andlead to operational failure of mitochondria with progres-sive decline of cell viability Almost identical conditionsaccompany AD and are an explicit step in pathogenesisof disease As mtDNA is deprived of repair mechanismsROS-induced DNA strand breaks tend to accumulate withage or AD Thus mtDNA is a vulnerable target for ROSbut the reverse the ROS generation due to the mutatedmtDNA is not convincingly confirmed [156] Moreover theevidence that mutated mtDNA accelerates the progress ofaging is also questioned based on the results from studycarried out on transgenic mice model [157] Although someauthors show inconsistency between the free radical theoryand observations cumulative evaluation of the scientificreports points to antioxidant defense systems as importantfactors in protection from premature aging [158] Othermitochondrial components important for their function arehampered with age adenine nucleotide translocase (ANT)nitric oxide synthase (NOS) and carnitine acyltransferase(CT) [159ndash161] Actually NOS activity is elevated in AD asreported from study performed on cellular model of disease[15] We could not find any information about CT activityin AD whereas ANT activity is noticeably inhibited byA120573 or hyperphosphorylated tau and this effect is reversedby mersalyl a reversible alkylating agent of thiol groups[162] Mitochondrial dysfunction observed in transgenicmice models of AD and aging demonstrates higher activ-ity of genes controlling energy metabolism and apoptosisTaken together physiological aging and AD are associatedwith broad-spectrum dysfunction of mitochondria but thefoundations of mitochondrial decline are dissimilar Morediscrepancies between physiological aging and AD werefound with respect to ERS and UPR which play a significantrole in cellular proteostasis Dolichol was selected as an agingmarker because the progressive increase in dolichol levelwas observed in aging brain [163] In contrast ubiquinoneconcentration which is also synthesized from geranylgeranioldiminishes with agingwhereas cholesterol and dolichyl phos-phate concentrations remain unaltered In AD decreasedlevels of dolichol were observed and increased levels ofubiquinone and dolichyl phosphate without any changesin brain cholesterol AD cannot be regarded as a resultof premature aging The drop in dolichol and augmenteddolichyl phosphate concentration points toward disturbedglycosylation in ER of diseased brain while the increase inubiquinone suggests efforts to protect the brain from oxida-tive stress induced by lipid peroxidation [164 165]
14 Targeting ER Stress in AD Therapy
As ERS is a recognized factor in AD drugs that interfere withERS would theoretically have great therapeutic potentialThere are several compounds grouped in classes thatinteract directly with components of the ERS (salubrinalBiP inducer X (BIX) salicylamide analogs flavonoidsguanabenz and STF083010) chemical chaperons (4-phenyl-butyric acid (PBA) tauroursodeoxycholic acid (TUDCA)and trimethylamine oxide (TMAO)) chemicals that inhibitprotein degradation (Eeyarestatin MG132 and Bortezomib)compounds with antioxidant activity (Edaravone diben-zoylmethane derivatives and N-acetyl cysteine (NAC)) anddrugs controlling calcium signaling (dantrolene and carba-zole derivatives) [166] They may act by inducing transienttranslation arrest upregulation of chaperone proteins andaugmented degradation of ER-associated misfolded proteinsFundamental approach in the development of new therapyis the selection of appropriate molecular targets In ER stresssignaling the aim is to alter the expression of ER stress-associated molecules that can rescue cells from the toxiceffect of ERS Recent efforts in establishing new promisingdrugs against AD are pointing to chemical chaperones suchas PBA TUDCA or trimethylamine oxide (TMAO) Thesesubstances improve protein folding and alleviate nativeprotein conformation [166] It was shown on mouse modelsof AD that PBA TUDCA and TMAO stop A120573 accumulationand avoid the loss of dendritic spines [167] Some observa-tions even demonstrated improved memory and cognitivefunctions [168] associated with improved cell survival [169]Salubrinal ((2E)-3-phenyl-N-[222-trichloro-1-[[(8-quinoli-nylamino)thioxomethyl]amino]ethyl]-2-propenamide Sal)selectively inhibits growth arrest and DNA damage inducedgene 34- (GADD34-) phosphatase complex (GADD34associates with protein phosphatase 1 (PP1)) and promotes invitro dephosphorylation of the alpha subunit of eIF-2120572 andIRE1120572ASK1JNK signaling pathway being protective againstERS even induced by tunicamycin Tm [170] In a great deal ofexperiments testing Sal in cultured cells and animal modelsof AD this substance increased the viability of neuronal cellsand A120573 toxicity [171 172] BIX (2-(34-dihydroxyphenyl)-2-oxoethyl ester thiocyanic acid) preferentially inducedBiP mRNA in an ATF6-dependent manner leading toreduced Tm-induced death of neuronal cells [173] AlsoDBM derivative 14ndash26 (221015840-dimethoxydibenzoylmethane)was found to be neuroprotective for SH-SY5Y and PC-12cells by decreasing expression of BiP and CHOP [174]Dantrolene a ryanodine receptor antagonist that inhibitsabnormal calcium release from the ER inhibited expressionof both phosphorylated PERK and eIF2120572 It also reducedCHOP expression and attenuated thapsigargin-inducedapoptosis in PC-12 cells [175] Neuroprotective effects similarto dantrolene were observed for ([9-(3-cyanobenzyl)-14-dimethylcarbazole]) This substance suppressed increases inintracellular Ca2+ in PC-12 cells treated with thapsigarginand reduced levels of BiP and CHOP [176]
Aforementioned compounds were chosen from othersas the most potent ER stress inhibitors and persuasivelyprotective to neuronal cells 18 of 42 different compounds
Oxidative Medicine and Cellular Longevity 11
were exploited in in vivo and in vitro models of centralnervous system disorders with in fact improved cell or tissueviability [166] Thus the brain is the most frequently investi-gated organ in the context of ERS From these experimentsit becomes clear that CHOP functions as proapoptotic factorThe roles of other specific ER stress molecules as moleculartargets for pharmacological intervention are less clear andvary depending on cell type and context
There are few underestimated modulations in APP pro-cessing that shed more light on current dogma of AD patho-genesis The modulation of mevalonate pathway and choles-terol synthesis were reported to stimulate nonamyloidogenicpathway of APP processing [177] Additionally cholesterolderivative 27-hydroxycholesterol (27-OHC) was shown toinduce ER stress which attenuated leptin-dependent viabilityby activating CHOP in SH-SY5Y neuroblastoma cells [178]Irrespective of a number of compounds examined in AD thecall for new drugs modulating ER stress with healing effect isstill waiting to be revealed
Abbreviations
APOE Apolipoprotein EAPPs Secreted form of APPCHOPGADD153 CEBP homologous protein
growth arrest and DNA damageinduced gene 153
Drp1 Dynamin-1-like proteinERAD Endoplasmic-reticulum-
associated protein degradationERO1120572 ER oxidase 1120572ERS Endoplasmic reticulum stressFOH FarnesolGGOH GeranylgeraniolGRP75 Glucose-regulated protein 75GRP78BiP 78 kDa glucose-regulated proteinGSK-3120573 Synthase kinase 3 betaIP3R Inositol triphosphate receptor
IRE1120572 Inositol-requiring enzyme 1 alphaMAM Mitochondria-associated
membraneMAP LC3 Microtubule-associated protein
light chain 3Mfn2 GTPase mitofusin 2MMP Mitochondrial membrane
potentialMPTP Mitochondrial permeability
transition poreNFT Neurofibrillary tanglesPACS-2 Phosphofurin acidic cluster
sorting protein 2PDI Protein disulfide isomerasePERK Protein kinase R- (PKR-) like
ER kinaseRNS Reactive nitrogen speciesROS Reactive oxygen speciesRyR Ryanodine receptorTOM Transporter outer membraneTIM Transporter inner membrane
UPR Unfolded protein responseVDAC Voltage-dependent anion channelXBP-1 Xbox binding protein 1
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
Support for this work was provided by Grant no UMO-201103BNZ70238 from the National Science Centre inPoland
References
[1] R L Nussbaum andC E Ellis ldquoAlzheimerrsquos disease and Parkin-sonrsquos diseaserdquoTheNewEngland Journal ofMedicine vol 348 no14 pp 1356ndash1364 2003
[2] A L Brunkan and A M Goate ldquoPresenilin function and 120574-secretase activityrdquoThe Journal of Neurochemistry vol 93 no 4pp 769ndash792 2005
[3] V Crentsil ldquoThe pharmacogenomics of Alzheimerrsquos diseaserdquoAgeing Research Reviews vol 3 no 2 pp 153ndash169 2004
[4] D L Price and S S Sisodia ldquoMutant genes in familialAlzheimerrsquos disease and transgenic modelsrdquo Annual Review ofNeuroscience vol 21 pp 479ndash505 1998
[5] D Harman ldquoAlzheimerrsquos disease pathogenesis role of agingrdquoAnnals of the New York Academy of Sciences vol 1067 no 1 pp454ndash460 2006
[6] D J Selkoe ldquoTranslating cell biology into therapeutic advancesin Alzheimerrsquos diseaserdquo Nature vol 399 pp A23ndashA31 1999
[7] H Braak and E Braak ldquoNeuropathological stageing of Alz-heimer-related changesrdquo Acta Neuropathologica vol 82 no 4pp 239ndash259 1991
[8] K Hirai G Aliev A Nunomara et al ldquoMitochondrial abnor-malities inAlzheimerrsquos diseaserdquoTheJournal ofNeuroscience vol21 pp 3017ndash3023 2001
[9] U T Brunk C B Jones and R S Sohal ldquoA novel hypothesisof lipofuscinogenesis and cellular aging based on interac-tions between oxidative stress and autophagocytosisrdquoMutationResearchDNAging vol 275 no 3ndash6 pp 395ndash403 1992
[10] P A Stewart K Hayakawa M-A Akers and H V Vinters ldquoAmorphometric study of the blood-brain barrier in Alzheimerrsquosdiseaserdquo Laboratory Investigation vol 67 no 6 pp 734ndash7421992
[11] J P Blass and G E Gibson ldquoThe role of oxidative abnor-malities in the pathophysiology of Alzheimerrsquos diseaserdquo RevueNeurologique vol 147 no 6-7 pp 513ndash525 1991
[12] B Pająk E Kania and A Orzechowski ldquoNucleofection of ratpheochromocytoma PC-12 cells with human mutated beta-amyloid precursor protein gene (APP-sw) leads to reducedreduced viability autophagy-like process and increased expres-sion and secretion of beta amyloidrdquo BioMed Research Interna-tional vol 2015 Article ID 746092 10 pages 2015
[13] J P Blass ldquoCerebrometabolic abnormalities in AlzheimerrsquosdiseaserdquoNeurological Research vol 25 no 6 pp 556ndash566 2003
12 Oxidative Medicine and Cellular Longevity
[14] A Eckert K Schmitt and J Gotz ldquoMitochondrial dysfunc-tionmdashthe beginning of the end in Alzheimerrsquos disease Sep-arate and synergistic modes of tau and amyloid-120573 toxicityrdquoAlzheimerrsquos Research ampTherapy vol 3 article 15 pp 1ndash11 2011
[15] U Keil A Bonert C A Marques et al ldquoAmyloid 120573-inducedchanges in nitric oxide production and mitochondrial activitylead to apoptosisrdquoThe Journal of Biological Chemistry vol 279no 48 pp 50310ndash50320 2004
[16] A M Swomley and D A Butterfield ldquoOxidative stress inAlzheimer disease and mild cognitive impairment evidencefrom human data provided by redox proteomicsrdquo Archives ofToxicology vol 89 no 10 pp 1669ndash1680 2015
[17] C Caspersen N Wang J Yao et al ldquoMitochondrial A120573 apotential focal point for neuronal metabolic dysfunction inAlzheimerrsquos diseaserdquo The FASEB Journal vol 19 no 14 pp2040ndash2041 2005
[18] G E Gibson K-F R Sheu and J P Blass ldquoAbnormalities ofmitochondrial enzymes inAlzheimer diseaserdquo Journal of NeuralTransmission vol 105 no 8-9 pp 855ndash870 1998
[19] A Eckert S Hauptmann I Scherping et al ldquoSoluble beta-amyloid leads to mitochondrial defects in amyloid precursorprotein and tau transgenic micerdquo Neurodegenerative Diseasesvol 5 no 3-4 pp 157ndash159 2008
[20] A Eckert K L Schulz V Rhein and J Gotz ldquoConvergenceof amyloid-120573 and tau pathologies on mitochondria in vivordquoMolecular Neurobiology vol 41 no 2-3 pp 107ndash114 2010
[21] S Hauptmann I Scherping S Drose et al ldquoMitochondrial dys-function an early event in Alzheimer pathology accumulateswith age in AD transgenic micerdquoNeurobiology of Aging vol 30no 10 pp 1574ndash1586 2009
[22] M Manczak B S Park Y Jung and P H Reddy ldquoDifferentialexpression of oxidative phosphorylation genes in patients withAlzheimerrsquos disease implications for early mitochondrial dys-function and oxidative damagerdquo NeuroMolecular Medicine vol5 no 2 pp 147ndash162 2004
[23] M P Mattson M Gleichmann and A Cheng ldquoMitochondriain neuroplasticity and neurological disordersrdquo Neuron vol 60no 5 pp 748ndash766 2008
[24] P I Moreira M S Santos and C R Oliveira ldquoAlzheimerrsquosdisease a lesson frommitochondrial dysfunctionrdquoAntioxidantsamp Redox Signaling vol 9 no 10 pp 1621ndash1630 2007
[25] P H Reddy ldquoMitochondrial dysfunction in aging and Alz-heimerrsquos disease strategies to protect neuronsrdquo Antioxidants ampRedox Signaling vol 9 no 10 pp 1647ndash1658 2007
[26] V Rhein G Baysang S Rao et al ldquoAmyloid-beta leads toimpaired cellular respiration energy production andmitochon-drial electron chain complex activities in humanneuroblastomacellsrdquo Cellular and Molecular Neurobiology vol 29 no 6-7 pp1063ndash1071 2009
[27] V Rhein X Song A Wiesner et al ldquoAmyloid-120573 and tausynergistically impair the oxidative phosphorylation system intriple transgenic Alzheimerrsquos disease micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 47 pp 20057ndash20062 2009
[28] R X Santos S C Correia X Wang et al ldquoAlzheimerrsquos diseasediverse aspects ofmitochondrial malfunctioningrdquo InternationalJournal of Clinical and Experimental Pathology vol 3 no 6 pp570ndash581 2010
[29] N R Sims ldquoEnergy metabolism oxidative stress and neuronaldegeneration in Alzheimerrsquos diseaserdquoNeurodegeneration vol 5no 4 pp 435ndash440 1996
[30] H K Anandatheerthavarada G Biswas M-A Robin and NG Avadhani ldquoMitochondrial targeting and a novel transmem-brane arrest of Alzheimerrsquos amyloid precursor protein impairsmitochondrial function in neuronal cellsrdquo The Journal of CellBiology vol 161 no 1 pp 41ndash54 2003
[31] L Devi B M Prabhu D F Galati N G Avadhani and HK Anandatheerthavarada ldquoAccumulation of amyloid precur-sor protein in the mitochondrial import channels of humanAlzheimerrsquos disease brain is associated with mitochondrialdysfunctionrdquo The Journal of Neuroscience vol 26 no 35 pp9057ndash9068 2006
[32] L Devi and H K Anandatheerthavarada ldquoMitochondrialtrafficking of APP and alpha synuclein relevance to mito-chondrial dysfunction in Alzheimerrsquos and Parkinsonrsquos diseasesrdquoBiochimica et Biophysica Acta vol 1802 no 1 pp 11ndash19 2010
[33] M Manczak T S Anekonda E Henson B S Park J Quinnand P H Reddy ldquoMitochondria are a direct site of A120573 accumu-lation in Alzheimerrsquos disease neurons implications for free rad-ical generation and oxidative damage in disease progressionrdquoHuman Molecular Genetics vol 15 no 9 pp 1437ndash1449 2006
[34] P F Pavlov C H Petersen E Glaser and M AnkarcronaldquoMitochondrial accumulation of APP and A120573 significancefor Alzheimer disease pathogenesisrdquo Journal of Cellular andMolecular Medicine vol 13 no 10 pp 4137ndash4145 2009
[35] C Schmidt E Lepsverdize S L Chi et al ldquoAmyloid precursorprotein and amyloid 120573-peptide bind to ATP synthase andregulate its activity at the surface of neural cellsrdquo MolecularPsychiatry vol 13 no 10 pp 953ndash969 2008
[36] K Johar A Priya and M T T Wong-Riley ldquoRegulation ofNa+K+-ATPase by nuclear respiratory factor 1 implication inthe tight coupling of neuronal activity energy generation andenergy consumptionrdquo The Journal of Biological Chemistry vol287 no 48 pp 40381ndash40390 2012
[37] J Astrup P M Sorensen and H R Sorensen ldquoOxygen andglucose consumption related to Na+-K+ transport in caninebrainrdquo Stroke vol 12 no 6 pp 726ndash730 1981
[38] M Erecinska and I A Silver ldquoATP and brain functionrdquo Journalof Cerebral Blood Flow and Metabolism vol 9 no 1 pp 2ndash191989
[39] MMata D J Fink H Gainer et al ldquoActivity-dependent energymetabolism in rat posterior pituitary primarily reflects sodiumpump activityrdquo Journal of Neurochemistry vol 34 no 1 pp 213ndash215 1980
[40] J Kang H-G Lemaire A Unterbeck et al ldquoThe precursor ofAlzheimerrsquos disease amyloid A4 protein resembles a cell-surfacereceptorrdquo Nature vol 325 no 6106 pp 733ndash736 1987
[41] C L Masters and D J Selkoe ldquoBiochemistry of amyloid 120573-protein and amyloid deposits in Alzheimer diseaserdquoCold SpringHarbour Perspectives in Medicine vol 2 no 6 Article IDa006262 2012
[42] A Lai S S Sisodia and I S Trowbridge ldquoCharacterization ofsorting signals in the beta-amyloid precursor protein cytoplas-mic domainrdquo The Journal of Biological Chemistry vol 270 no8 pp 3565ndash3573 1995
[43] A I Placido C M F Pereira A I Duarte et al ldquoThe role ofendoplasmic reticulum in amyloid precursor protein processingand trafficking implications for Alzheimerrsquos diseaserdquo Biochim-ica et Biophysica ActamdashMolecular Basis of Disease vol 1842 no9 pp 1444ndash1453 2014
Oxidative Medicine and Cellular Longevity 13
[44] C Kaether P Skehel and C G Dotti ldquoAxonal membraneproteins are transported in distinct carriers a two-color videomicroscopy study in cultured hippocampal neuronsrdquoMolecularBiology of the Cell vol 11 no 4 pp 1213ndash1224 2000
[45] A Kamal A Almenar-Queralt J F LeBlanc E A Robertsand L S B Goldstein ldquoKinesin-mediated axonal transportof a membrane compartment containing 120573-secretase andpresenilin-1 requires APPrdquo Nature vol 414 no 6864 pp 643ndash648 2001
[46] R G Perez S Soriano J D Hayes et al ldquoMutagenesis identi-fies new signals for 120573-amyloid precursor protein endocytosisturnover and the generation of secreted fragments includingA12057342rdquo The Journal of Biological Chemistry vol 274 no 27 pp18851ndash18856 1999
[47] D G Cook M S Forman J C Sung et al ldquoAlzheimerrsquos A120573(1ndash42) is generated in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo Nature Medicine vol 3 no 9 pp1021ndash1023 1997
[48] A S C Chyung B D Greenberg D G Cook R W Domsand V M-Y Lee ldquoNovel 120573-secretase cleavage of 120573-amyloidprecursor protein in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo The Journal of Cell Biology vol138 no 3 pp 671ndash680 1997
[49] RW-Y Choy Z Cheng and R Schekman ldquoAmyloid precursorprotein (APP) traffics from the cell surface via endosomesfor amyloid 120573 (A120573) production in the trans-Golgi networkrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 30 pp E2077ndashE2082 2012
[50] S A Small and S Gandy ldquoSorting through the cell biologyof Alzheimerrsquos disease intracellular pathways to pathogenesisrdquoNeuron vol 52 no 1 pp 15ndash31 2006
[51] M Goedert ldquoAlzheimerrsquos and Parkinsonrsquos diseases the prionconcept in relation to assembled A120573 tau and 120572-synucleinrdquoScience vol 349 no 6248 2015
[52] I Benilova E Karran and B De Strooper ldquoThe toxic A120573oligomer and Alzheimerrsquos disease an emperor in need ofclothesrdquo Nature Neuroscience vol 15 no 3 pp 349ndash357 2012
[53] E H Corder A M Saunders W J Strittmatter et al ldquoGenedose of apolipoprotein E type 4 allele and the risk of Alzheimerrsquosdisease in late onset familiesrdquo Science vol 261 no 5123 pp 921ndash923 1993
[54] S Brunholz S Sisodia A Lorenzo C Deyts S Kins and GMorfini ldquoAxonal transport of APP and the spatial regulationof APP cleavage and function in neuronal cellsrdquo ExperimentalBrain Research vol 217 no 3-4 pp 353ndash364 2012
[55] G Thinakaran and E H Koo ldquoAmyloid precursor proteintrafficking processing and functionrdquo The Journal of BiologicalChemistry vol 283 no 44 pp 29615ndash29619 2008
[56] T Saitoh M Sundsmo J-M Roch et al ldquoSecreted formof amyloid beta protein precursor is involved in the growthregulation of fibroblastsrdquo Cell vol 58 no 4 pp 615ndash622 1989
[57] M PMattson B Cheng A R Culwell F S Esch I Lieberburgand R E Rydel ldquoEvidence for excitoprotective and intraneu-ronal calcium-regulating roles for secreted forms of the 120573-amyloid precursor proteinrdquo Neuron vol 10 no 2 pp 243ndash2541993
[58] K J Lee C E H Moussa Y Lee et al ldquoBeta amyloid-independent role of amyloid precursor protein in generationandmaintenance of dendritic spinesrdquoNeuroscience vol 169 no1 pp 344ndash356 2010
[59] H Meziane J-C Dodart C Mathis et al ldquoMemory-enhancingeffects of secreted forms of the 120573-amyloid precursor proteinin normal and amnestic micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no21 pp 12683ndash12688 1998
[60] P R Turner K OrsquoConnor W P Tate and W C AbrahamldquoRoles of amyloid precursor protein and its fragments inregulating neural activity plasticity and memoryrdquo Progress inNeurobiology vol 70 no 1 pp 1ndash32 2003
[61] D Beher L Hesse C L Masters and GMulthaup ldquoRegulationof Amyloid Protein Precursor (APP) binding to collagen andmapping of the binding sites on APP and collagen type Irdquo TheJournal of Biological Chemistry vol 271 no 3 pp 1613ndash16201996
[62] S L Sabo A F Ikin J D Buxbaum and P Greengard ldquoTheamyloid precursor protein and its regulatory protein FE65 ingrowth cones and synapses in vitro and in vivordquo The Journal ofNeuroscience vol 23 no 13 pp 5407ndash5415 2003
[63] D H Small V Nurcombe G Reed et al ldquoA heparin-bindingdomain in the amyloid protein precursor of Alzheimerrsquos diseaseis involved in the regulation of neurite outgrowthrdquoThe Journalof Neuroscience vol 14 no 4 pp 2117ndash2127 1994
[64] T G Williamson S S Mok A Henry et al ldquoSecreted glypicanbinds to the amyloid precursor protein of Alzheimerrsquos disease(APP) and inhibits APP-induced neurite outgrowthrdquoThe Jour-nal of Biological Chemistry vol 271 no 49 pp 31215ndash312211996
[65] T Ondrejcak I Klyubin N-W Hu A E Barry W K Cullenand M J Rowan ldquoAlzheimerrsquos disease amyloid 120573-protein andsynaptic functionrdquo NeuroMolecular Medicine vol 12 no 1 pp13ndash26 2010
[66] D Puzzo L Privitera and A Palmeri ldquoHormetic effect ofamyloid-beta peptide in synaptic plasticity and memoryrdquo Neu-robiology of Aging vol 33 no 7 pp 1484e15ndash1484e24 2012
[67] C M F Pereira ldquoCrosstalk between endoplasmic reticulumstress and protein misfolding in neurodegenerative diseasesrdquoISRN Cell Biology vol 2013 Article ID 256404 22 pages 2013
[68] R-W Shin T C Saido M Maeda and T Kitamoto ldquoNovel 120572-secretase cleavage ofAlzheimerrsquos amyloid120573 precursor protein inthe endoplasmic reticulum of COS7 cellsrdquoNeuroscience Lettersvol 376 no 1 pp 14ndash19 2005
[69] E A Schon and E Area-Gomez ldquoMitochondria-associatedER membranes in Alzheimer diseaserdquo Molecular and CellularNeuroscience vol 55 pp 26ndash36 2013
[70] R Rizzuto P Pinton W Carrington et al ldquoClose contacts withthe endoplasmic reticulum as determinants of mitochondrialCa2+ responsesrdquo Science vol 280 no 5370 pp 1763ndash1766 1998
[71] S E Logue P Cleary S Saveljeva and A Samali ldquoNewdirections in ER-stress induced cell deathrdquoApoptosis vol 18 no5 pp 537ndash546 2013
[72] J P Decuypere KWelkenhuyzen T Luyten et al ldquoIns(145)P3receptor-mediated BI-1 Ca2+ signaling and autophagy induc-tion are interrelatedrdquo Autophagy vol 7 pp 1472ndash1489 2011
[73] K S Vetrivel and G Thinakaran ldquoMembrane rafts in Alz-heimerrsquos disease beta-amyloid productionrdquo Biochimica et Bio-physica Acta vol 1801 no 8 pp 860ndash867 2010
[74] H Urra E Dufey F Lisbona D Rojas-Rivera and C HetzldquoWhen ER stress reaches a dead endrdquo Biochimica et BiophysicaActa (BBA) Molecular Cell Research vol 1833 no 12 pp 3507ndash3517 2013
14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 Oxidative Medicine and Cellular Longevity
formation in neurons in concert with the increased levelsof PERK eIF2120572 and XBP1 transcript apparent markers ofUPR [87] Furthermore GRP78BiP was found to encouragetau phosphorylation through facilitated substrate capture byGSK-3120573 [88] GSK-3120573 seems to play dual role first this kinaseprotects neurons from apoptosis as P-tau accumulation isstrong molecular signal to trigger UPR with subsequentautophagy Second UPR raises GSK-3120573 activity throughlysosomal degradation of inactive GSK-3120573 (P-Ser9-GSK-3120573)To sum up ERS and UPR are important molecular machinesused to prevent cell viability turned on by tauopathy
7 Oxidative Stress in ER
Sacs and tubes of ER delineate the compartment where thenewly synthesized proteins undergo maturation to nativestate Native state indicates properly folded fully functionalprotein Important reactions essential for protein folding ofunbranched polypeptide chains include amino acids oxida-tion and glycosylation As a result redox homeostasis inER is shifted to oxidative state so as to promote disulfidebond formation between adjacent cysteines Oxidation ofsulfhydryl groups required to make disulfide bonds is con-trolled by ER oxidase 1120572 (ERO1120572) Next disulfide bondscould be subject to posttranslational modification disulfideexchange by protein disulfide isomerase (PDI) PDI is ableto correct mispaired thiol residues by catalyzing the breakageand formation of correct disulfide bonds These enzymes arefundamental for protein folding Oxidation allows twistingof proteins which is followed by N-glycosylation andor C-and O-mannosylation Protein N-glycosylation in eukary-otes is a complex process divided into several steps Firstthere is a ldquocall forrdquo carrier lipids (polyisoprenyl phosphatessuch as dolichyl phosphates) the membrane lipids knownto function as glycosyl transporters In mammalian cellsthe limiting substrate for dolichol biosynthesis is geranyl-geraniol (GGOH) of mevalonate pathway Dolichols thelongest aliphatic molecules synthesized in animal cellshave 18ndash21 120572-isoprene saturated units (C90ndash105) criticalfor their recognition by the enzymes (glycosyltransferases)that glucosylate dolichyl phosphates [89] Once dolichylmonophosphates (Dol-P) are formed in the ER membranethe precursor oligosaccharide donor (GLc
3Man9GLCNAc
2-
P-P-dolichol) for protein N-glycosylation can be synthesizedon the lumenal leaflet of ER First three sugar intermediatesare produced (Man-P-Dol Glc-P-Dol GlcNAc-P-P-Dol andMan5GlcNAc-P-P-Dol) on the cytoplasmic leaflet of the
ER Next enzyme flippases mediate transbilayer movementof the aforementioned intermediates to lumenal side ofER where conversion to Glc
3Man9GlcNAc
2-P-P-Dol could
be completed Glc3Man9GlcNAc
2-P-P-Dol is also used for
biosynthesis of glycosylphosphatidylinositol (GPI) anchorsTaken together lipid-mediated glycosylation plays a vital
role in the appropriate protein folding and intracellulartranslocation of N-linked glycoproteins [90] Likewise it isimportant for protein O- and C-mannosylation and GPIanchorage Moreover Dol-P availability in the ER is the rate-limiting factor in the production of glycolipid intermediatesand N-glycosylation
8 ER Stress and Apoptosis
It was shown that during ERS the ER resident proapop-totic cysteine protease known as caspase (caspase-12 in ratcaspase-4 in humans) is activated through cleavage As aresult caspase cascade is started via caspase-9 that in turnstimulates effector caspase-3 [91 92] The central role playedby ER in programmed cell death is achieved by PERK branchwhere ATF4 induces the expression of CHOPGADD153which represses antiapoptotic Bcl-2 family proteins andsimultaneously shuffles ER membrane Bax and Bak proteinsinto outer mitochondrial membrane Consequently poresare formed to leak the components of apoptosome frommitochondrial intermembrane space [93] Another impor-tant mechanism of ERS-induced apoptosis is led by Ca2+-dependent ERO1120572-IP
3R pathway where ERO1120572 collaborates
with IP3R in Ca2+ efflux from ER to mitochondria via MAM
[75 94] Accordingly Ca2+ influx facilitates cytochrome 119888release frommitochondria besides cytochrome 119888 can bind toER IP
3R and the complex amplifies the apoptotic signal in a
feedforward manner [95] Last but not least ERS-associatedapoptotic programme is set off by c-Jun N-terminal kinase(JNK) as the effect of IRE1120572 complexed with TNF-receptor-associated factor 2 (TRAF2) activation of apoptosis-signal-regulating kinase 1 (ASK1) [96 97]
What does ERS drive to induce apoptotic death inneurons Actually many reports indicate that APP and A120573 aswell as hyperphosphorylated tau have been shown to blockmitochondrial transport which results in impaired energystorage and oxidative stress [98ndash101] Indeed accumulationof APP A120573 and NFTs in mitochondria led to reducedactivity of some enzymes involved in substrate oxidation(tricarboxylic acid cycle) electron transport chain (ETC)and ATP synthase as well as severely diminishing import ofnuclear-encoded proteins [26 27 102ndash104] One may ask ifthere is any additional link between ER and mitochondriaother than MAM which could account for apoptotic signalThough not directly ER significantly contributes to oxidativestress in mitochondria of AD-affected subjects
9 Abnormal APP Processing and TraffickingCulminate in ER Pathology of AD
From the morphological point of view as neurons arehighly specialized cells soma dendrites and neurites areconsiderably distinct structures Proteins needed by thesecompartments are delivered via microtubules once proteinshave suitable sorting signals (ie APP trafficking from ERto plasma membrane is associated with several posttrans-lational modifications with oxidation and N-glycosylation)Additionally the Golgi apparatus follows ER in subsequentAPP adjustment (O- and N-glycosylation phosphorylationand sulphonation) [42] Any inaccurate alteration of the APPmolecule is potentially hazardous as protein final destiny ismissed causing its retention in ER or trans-Golgi network(TGN) In addition other unusual settings for APP arepossible as this large protein has few signal sequences hiddenwhen APP is correctly folded In the cells transfected withAPP this protein enters coat protein complex I (COPI)
Oxidative Medicine and Cellular Longevity 7
vesicles and undergoes retrograde transport from cis endof the Golgi complex back to the ER [76] Such responsecauses accumulation of APP in the tubulocisternal ER systemtogether with aberrant intracellular translocation of theprotein Interestingly the issue whether APP is subject toretrograde transport with successive fibrillogenic processingbecause of UPR and its interaction with GRP78BiP is notclear as the observations are inconsistent [77] Nonethelessaccumulation of misfoldedmalfolded proteins in the ERsuggests disorganized process of posttranslational change Asanticipated the accretion of proteins of anomalous patternsignals ERS and UPR followed by increased vulnerability toapoptotic cell death Prior to decay however APP proteinlevels in the ER lessen by dint of cleavage with 120573- and120574-secretase [105 106] Products of this cleavage (sAPP120573A120573 and AICD fragment) all appreciably influence neuronalsurvival most likely through nonnative form of the APPsubstrate
10 APP A120573 and NFTs MarkMitochondria as Targeted in AD
Mitochondrial import of A12057340
and A12057342
peptides throughthe translocase TOM complex was blocked by preincubationof isolated mitochondria with antibodies raised againstTOM proteins (TOM20 TOM40 and TOM70) [107]Neither VDAC inhibition with antagonist antibodies norinhibition of mitochondrial permeability transition pores(MPTP) or fall of mitochondrial membrane potential(MMP) affected uptake of A120573 [17] With regard to APPelegant study performed by Anandatheerthavarada andhis colleagues [30] revealed that C-terminal truncatedAPP (lacking A120573) targets mitochondria in cholinergicGABAergic dopaminergic and glutamatergic neurons ofAD brain by clogging up mitochondrial protein translocasecomplex TOM40TIM22 Authors propose that occlusionof translocase is followed by blunted import of nuclear-encoded proteins vitally important for energy homeostasisMitochondrial APP protein transmembrane orientationindicates NH
2-terminal inside in contact with translocase
whereas COOH-terminal is facing cytoplasmic side NH2-
terminus has mitochondrial signal sequence Astonishinglymitochondrial APP molecules were nonglycosylated givingrise to speculation that protein molecules that arrivedat mitochondria have not achieved molecular maturity[31] The accumulation of nonglycosylated APP species inmitochondrial import channels of AD brain was directlyrelated to decreased mitochondrial functions as validatedby the decline in cytochrome 119888 oxidase activity (complexIV) and elevated levels of H
2O2 Furthermore in AD
brain the mitochondrial accumulation of nonglycosylatedAPP went along with a corresponding reduction in plasmamembrane-associatedAPP It suggests that ADbrain hasAPPprocessing and trafficking severely affected by incompleteN-glycosylation ensuing ER protein accumulation andexposure of the cryptic mitochondrial targeting signalfor assisting chaperone proteins Several lines of evidenceindicate that not fully formed proteins are phosphorylatedand bind to the cytosolic proteins required for movement
from ER to mitochondria [108ndash110] Moreover ERS andUPR in AD seem to be incompetent and inefficient inelimination of malfolded proteins In addition to APPalso A120573 was frequently reported to occupy mitochondriaalthough its origin and mechanism of mitochondriatargeting mostly remain unexplored (except for involvementof protein translocase TOM complex) The possibilityof mitochondrial A120573 generation has to be ruled out asmembrane orientation of arrested APP does not allow accessto 120574-secretase (120574-secretase activity in mitochondria wasdetected by independent study) [111] Probably A120573 speciesis derived from APP prior to its translocation (ER) soA120573 may be transported to mitochondria independentlyof APP Collectively observations showing mitochondrialpresence of APP A120573 and tau in aberrant configuration pointtoward the anomaly of protein folding at the level of ER andGolgi apparatus Nonglycosylated molecule of APP suggestsdefective transfer of sugar core from dolichyl phosphate(s)and further modifications such as O- and N-glycosylationsOne may admit that the lack of glycosyl residue bringsabout pathologic processing and trafficking of APP and itsfragments In point of fact dolichol derivatives mixture ofpolyprenols (acyclic isoprenoid alcohols) known as Ropren(Solagran Limited Melbourne Australia) commerciallyused to treat liver diseases were tested in the treatment ofAD in two separate trials conducted in 2005 and 2006 withpromising results [httpwwwasxcomauasxpdf20071119pdf315x5nh4hm8wv7pdf httpwwwasxcomauasxpdf20070221pdf3111ztwcbqzkk9pdf] Further studies areurgently needed to test how important the glycosylationprocess is in the pathogenesis of AD Dolichols areobtained from geranylgeraniol (GGOH) and the latter is anintermediate of mevalonate pathway GGOH is a commonsubstrate for dolichol and ubiquinone synthesis but it isalso necessary for protein prenylation As both GGOHand farnesol (FOH) are engaged in protein prenylationmore concern should be laid on the importance of thesecompounds in AD pathogenesis Observations demonstrat-ing mitochondrial relocalization of other proteins withoutposttranslational modification seemingly point to increasedmitochondrial targeting of immature molecules resultingin mitochondrial dysfunction and acceleration of diseaseprogression [34 112ndash114]
11 Oxidative Stress in Mitochondria ofAD Brains
Normal physiological functions of APP are thought to beinvolved in the stabilizing contact points between synapsesand maintaining mitochondrial functions [60 115] Mito-chondrial dysfunction was often observed regardless of theexperimental model used to study AD [8 29 116ndash119]It includes defects in oxidative phosphorylation decreasedATP decreased membrane potential increased productionof ROSRNS and perturbation in mitochondrial fusion andfission [15 30 31 115 120ndash122] Hyperphosphorylated tau wasalso reported to impair mitochondrial functions [123] Usingproteomic approach the strongest defects of the respiratorycapacity were observed mainly at complexes I IV and ATP
8 Oxidative Medicine and Cellular Longevity
synthase (complex V) at both protein and activity level [124]While APP A120573 and hyperphosphorylated tau are potentinhibitors of mitochondrial import of nuclear-encoded pro-teins apparently the pathology of each leads to metabolicharm in different way In freshly isolated mitochondriafrom AD brains the APP inhibited mitochondrial importof cytochrome 119888 oxidase (COX) subunits IV and Vb [31]Dysfunction of COX increases ROS production (incompletereduction of oxygen molecules) reduces energy stores anddisturbs energy metabolism Accordingly in AD patientsdeficiency of COX was found in brains and platelets [119125 126] Similar to APP A120573 was found in mitochondriaof transgenic mice and cellular and human AD models [1719 33 103 127ndash130] At present it is not clear whether theobservedmitochondrial toxicity is due toAPP orA120573 orNFTsaccumulation Anyway some regularity is observed withrespect to most affected components of respiratory chainNADH-ubiquinone oxidoreductase (complex I) activity isreduced to the utmost by hyperphosphorylated tau whereasdecreased activity of cytochrome 119888 oxidase (complex IV)that resulted in mitochondrial dysfunction was observedduring A120573 and APP accumulation [14 15 31 32 34]Concomitantly rise in the activity of antioxidant enzymesmanganese superoxide dismutase (Mn-SOD) and catalase(CAT) was demonstrated in response to elevated levelsof free radicals including superoxide anion radical (O
2
∙minus)hydroperoxyl radical (HO
2
∙) hydroxyl radical (OH∙) andnitric oxide radical (NO∙) [15] Consistent with observationsof chronic respiratory chain dysfunction and mitochondrialoxidative stress there are reports showing their contributionto tau pathology in AD [131] In any case free radicals thatoverride antioxidant defense react with a wide variety oforganic components causing lipid peroxidation to advancedlipid oxidation end products (ALE) cross-linking of proteinsnitrosylation of proteins and mutations in DNA Mitochon-drial circular DNA (mtDNA) of sim16 kbp is devoid of repairsystems meaning the buildup of lesions There are 37 geneslocated in mtDNA with those encoding protein subunits ofcomplex I (7) complex II (1) and complex IV (3) but notcomplex III Interestingly hallmarks of AD (APP A120573 andNFTs)mostly affectmembers of electron transfer chain (com-plexes I and IV) which rely exclusively on mitochondriallypredetermined subunits Maybe it is not simple coincidencebut the effect of inhibited import to mitochondria of nuclear-encoded subunits of complexes I and IV gives explanationfor toxicity of APP or A120573 or NFTs in mitochondria Thereare additional findings in mitochondria affected by AD suchas lower activity of pyruvate dehydrogenase (PDH) andoxoglutarate dehydrogenase (OGDH) [104 132] Inhibitionof OGDH the enzyme of tricarboxylic acid cycle minimizesthe NADH pool and electron number needed for ETCand mitochondrial membrane potential (Δ120595
119898) to create and
maintain proton gradient obligatory for ATP synthesisThereare also lines of evidence for direct inhibitory action of solubleoligomeric A120573 species on ABAD (A120573-binding alcohol dehy-drogenase) and internal membrane cyclophilin D (CypD)resulting in increased mitochondrial membrane permeabil-ity (MPTP) potentiated ROS production synaptic lossdiminished activity of mitochondrial respiration and finally
cell death [129 132ndash134] CypD knockout prevents mito-chondrial and neuronal perturbations and improves mito-chondrial function in Alzheimerrsquos disease mouse model [128135] It has to be emphasized that oxidativenitrosative stressaffects the fusion and fission process ofmitochondria Fusionthat is speedup by small GTPases mitofusins (Mfn1Mfn2)improves efficiency of mitochondrial respiration and ATPproduction Mitochondrial dynamics are severely imbal-anced in AD cases in favour of fission through elevatedexpression of the fission protein DLP1 (dynamin-like protein1) associatedwith nitrosative stress stirred up byA120573 [136 137]Collectively these observations indicate that toxic intracel-lular A120573
42-43 oligomers differ in action from extracellularaggregates found in amyloid plaques of AD brains [138](Figure 3)
12 Proteostasis in Mitochondria
Mitochondrial protein turnover grants the well-organizedreplacement of nonfunctional proteins into operational oneThe first task is attained through proteolytic degradation ofinner membrane and matrix proteins with local mitochon-drial proteases [139] or outer membrane proteins throughubiquitin-proteasome system [140] The second task is metby intramitochondrial protein synthesis but as almost 1500different nuclear-encoded proteins have to be importedto complete mitochondrial proteome a matter of capableimport is fundamental for the function of the organelleIn extreme cases of cellular injuries observed in neurode-generative diseases damaged mitochondria with extendedloss of the electrochemical potential are selectively removedby autophagy known as mitophagy [112] APP and A120573accumulation has also something to do with altered mito-chondrial dynamics as fission takes advantage of fusion withresultant dysfunction of mitochondria and neurons [137]AD neurons demonstrated selective mechanisms of prote-olytic clearance of oxidatively and nitrosatively modifiedproteins in mitochondria Insulin degrading enzyme (IDE)prevents formation of toxic insoluble fibrils from A120573 asit cleaves A120573 prior to aggregation [141ndash143] A novel zinc-metallopeptidase Presequence Protease (PreP) a member ofpitrilysin oligopeptidase family degrades either intramito-chondrially stored A120573
40or A120573
42proteinThis protease seems
to be highly sensitive to oxidative stress as disulfide bridgeformed between two proximal cysteine residues blocks itscatalytic activity [144ndash149] Another mitochondrial serineprotease HtrA2Omi occupies intermembrane space whereit can cleave APP locked up in protein translocase complex[121] Even though it is well established that HtrA2Omiis released to the cytosol to amplify apoptosis through thedegradation of antiapoptotic proteins and caspase activation[150ndash152] it is also implicated in proteolytic deletion ofmalfoldedAPP in ER [151] Fromknockout studies onmice itis obvious that HtrA2Omi plays a significant shielding roleas mice deficient in this protease exhibit neurodegenerativephenotype with weight loss and premature death [153]Taken together understanding the mechanisms of clearanceof the unwanted proteins including APP A120573 and tau isvital in an attempt to get rid of them from mitochondria
Oxidative Medicine and Cellular Longevity 9
VDAC
ANT
A120573
A120573
A120573
A120573
A120573
A120573
A120573A120573
A120573
A120573
A120573 CypD
CypD ProApCyt c Ca2+
IMM
OMM
IMSΔΨ
+
+
+
minus
minus
minus
ABAD
ROS
ROS
TIM
TOM
Proteinimport
NAD
NAD
NAD
NAD
H+
H+
H+
H+
H+
ADP
Mn-SOD
CuZn-SOD
mtDNA
V
Matrix
UCP
O2∙minus
O2∙minus
IV
III
II
I
O2
O2
DLP1Fis1
Mfn1Mfn2
Opa1
mHsp70
NADH
NADH
NADH
TCAcycle
PyruvateAcetyl-CoA
120572KGDH
120572-KG
PDH
Isocitrate
CitrateOAA
Malate
Fumarate
Succinyl-CoASuccinate
GDPGTP
FAD
FAOH
ATP
Figure 3 Amyloid-120573-related mitochondrial impairment Mitochondria were found to be the target for amyloid-120573 (A120573) which interactswith several proteins leading to mitochondrial dysfunction Indeed A120573 was found in the outer mitochondrial membrane (OMM) andinner mitochondrial membrane (IMM) as well as in the matrix The interaction of A120573 with the OMM affects the transport of nuclear-encoded mitochondrial proteins such as subunits of the electron transport chain complex IV into the organelle via the translocase of theouter membrane (TOM) import machinery Moreover A120573 disturbs the activity of several enzymes such as pyruvate dehydrogenase (PDH)and oxoglutarate dehydrogenase (OGDH) decreasing NADH reduction and the electron transport chain enzyme complex IV reducing theamount of hydrogen that is translocated from the matrix to the intermembrane space (IMS) thus impairing the mitochondrial membranepotential (MMP) Taken together these events cause abnormal mitochondrial electron activities leading to decreased complex V activityand so to a drop in ATP levels in addition to increasing reactive oxygen species (ROS) generation Moreover ROS induce peroxidationof several mitochondrial macromolecules such as mitochondrial DNA (mtDNA) and mitochondrial lipids contributing to mitochondrialimpairment in themitochondrial matrixThe complex of A120573 bound to binding alcohol dehydrogenase (ABAD) impairs the binding of NAD+to ABAD changes mitochondrial membrane permeability and reduces activities of respiratory enzymes inducing further ROS productionand leading to mitochondrial failure A120573 binding also activates Fis1 (fission protein) and promotes increased mitochondrial fragmentationthis increased mitochondrial fragmentation produces defective mitochondria that ultimately damage neurons Furthermore A120573 binding tocyclophilin D (CypD) enhances the protein translocation to the inner membrane favouring the opening of the mitochondrial permeabilitytransition pore formed by the adenine nucleotide translocator (ANT) and voltage-dependent anion channels (VDACs) Cyt 119888 cytochrome 119888DLP1 dynamin-like protein 1 PDH pyruvate dehydrogenase ProAp proapoptotic factors SOD superoxide dismutase TCA tricarboxylicacid TIM translocase of the inner membrane Adopted from [14]
Some hopes are associated with the application of at presentindefinite modulators of proteolytic activity HtrA2Omi isfor example activated by PTEN-induced putative kinase 1(PINK1) upon phosphorylation at Ser142 residue [154] Alter-natively accent has to be put on mitophagy of dysfunctionalmitochondria and mitochondriogenesis
13 Aging versus AD
Themost commonneurodegenerative disorder is representedby Alzheimerrsquos disease characterized by declining memoryreduced cognitive capacity and progressive dementia whichare often fatal to elderly individuals above 65 years of age
10 Oxidative Medicine and Cellular Longevity
It is ranked as the fourth leading cause of death in modernsocieties where average life span increased greatly in the lasttwo decades As 95 of AD cases are diagnosed in olderpeople one might think that a causal relationship existsbetween aging and the onset of disease Certainly a numberof similarities between getting old and being affected withAD could be listed Historically the free radical theory ofaging by Harman [155] suggested aging as ldquoside effectrdquo ofreactive oxygen species formed in mitochondrial respiratorychain Apparently free radicals commonly generated byincomplete reduction of oxygen molecule at complexes Iand III of mitochondrial ETC are capable of damagingDNA RNA and proteins They impair energy storage andlead to operational failure of mitochondria with progres-sive decline of cell viability Almost identical conditionsaccompany AD and are an explicit step in pathogenesisof disease As mtDNA is deprived of repair mechanismsROS-induced DNA strand breaks tend to accumulate withage or AD Thus mtDNA is a vulnerable target for ROSbut the reverse the ROS generation due to the mutatedmtDNA is not convincingly confirmed [156] Moreover theevidence that mutated mtDNA accelerates the progress ofaging is also questioned based on the results from studycarried out on transgenic mice model [157] Although someauthors show inconsistency between the free radical theoryand observations cumulative evaluation of the scientificreports points to antioxidant defense systems as importantfactors in protection from premature aging [158] Othermitochondrial components important for their function arehampered with age adenine nucleotide translocase (ANT)nitric oxide synthase (NOS) and carnitine acyltransferase(CT) [159ndash161] Actually NOS activity is elevated in AD asreported from study performed on cellular model of disease[15] We could not find any information about CT activityin AD whereas ANT activity is noticeably inhibited byA120573 or hyperphosphorylated tau and this effect is reversedby mersalyl a reversible alkylating agent of thiol groups[162] Mitochondrial dysfunction observed in transgenicmice models of AD and aging demonstrates higher activ-ity of genes controlling energy metabolism and apoptosisTaken together physiological aging and AD are associatedwith broad-spectrum dysfunction of mitochondria but thefoundations of mitochondrial decline are dissimilar Morediscrepancies between physiological aging and AD werefound with respect to ERS and UPR which play a significantrole in cellular proteostasis Dolichol was selected as an agingmarker because the progressive increase in dolichol levelwas observed in aging brain [163] In contrast ubiquinoneconcentration which is also synthesized from geranylgeranioldiminishes with agingwhereas cholesterol and dolichyl phos-phate concentrations remain unaltered In AD decreasedlevels of dolichol were observed and increased levels ofubiquinone and dolichyl phosphate without any changesin brain cholesterol AD cannot be regarded as a resultof premature aging The drop in dolichol and augmenteddolichyl phosphate concentration points toward disturbedglycosylation in ER of diseased brain while the increase inubiquinone suggests efforts to protect the brain from oxida-tive stress induced by lipid peroxidation [164 165]
14 Targeting ER Stress in AD Therapy
As ERS is a recognized factor in AD drugs that interfere withERS would theoretically have great therapeutic potentialThere are several compounds grouped in classes thatinteract directly with components of the ERS (salubrinalBiP inducer X (BIX) salicylamide analogs flavonoidsguanabenz and STF083010) chemical chaperons (4-phenyl-butyric acid (PBA) tauroursodeoxycholic acid (TUDCA)and trimethylamine oxide (TMAO)) chemicals that inhibitprotein degradation (Eeyarestatin MG132 and Bortezomib)compounds with antioxidant activity (Edaravone diben-zoylmethane derivatives and N-acetyl cysteine (NAC)) anddrugs controlling calcium signaling (dantrolene and carba-zole derivatives) [166] They may act by inducing transienttranslation arrest upregulation of chaperone proteins andaugmented degradation of ER-associated misfolded proteinsFundamental approach in the development of new therapyis the selection of appropriate molecular targets In ER stresssignaling the aim is to alter the expression of ER stress-associated molecules that can rescue cells from the toxiceffect of ERS Recent efforts in establishing new promisingdrugs against AD are pointing to chemical chaperones suchas PBA TUDCA or trimethylamine oxide (TMAO) Thesesubstances improve protein folding and alleviate nativeprotein conformation [166] It was shown on mouse modelsof AD that PBA TUDCA and TMAO stop A120573 accumulationand avoid the loss of dendritic spines [167] Some observa-tions even demonstrated improved memory and cognitivefunctions [168] associated with improved cell survival [169]Salubrinal ((2E)-3-phenyl-N-[222-trichloro-1-[[(8-quinoli-nylamino)thioxomethyl]amino]ethyl]-2-propenamide Sal)selectively inhibits growth arrest and DNA damage inducedgene 34- (GADD34-) phosphatase complex (GADD34associates with protein phosphatase 1 (PP1)) and promotes invitro dephosphorylation of the alpha subunit of eIF-2120572 andIRE1120572ASK1JNK signaling pathway being protective againstERS even induced by tunicamycin Tm [170] In a great deal ofexperiments testing Sal in cultured cells and animal modelsof AD this substance increased the viability of neuronal cellsand A120573 toxicity [171 172] BIX (2-(34-dihydroxyphenyl)-2-oxoethyl ester thiocyanic acid) preferentially inducedBiP mRNA in an ATF6-dependent manner leading toreduced Tm-induced death of neuronal cells [173] AlsoDBM derivative 14ndash26 (221015840-dimethoxydibenzoylmethane)was found to be neuroprotective for SH-SY5Y and PC-12cells by decreasing expression of BiP and CHOP [174]Dantrolene a ryanodine receptor antagonist that inhibitsabnormal calcium release from the ER inhibited expressionof both phosphorylated PERK and eIF2120572 It also reducedCHOP expression and attenuated thapsigargin-inducedapoptosis in PC-12 cells [175] Neuroprotective effects similarto dantrolene were observed for ([9-(3-cyanobenzyl)-14-dimethylcarbazole]) This substance suppressed increases inintracellular Ca2+ in PC-12 cells treated with thapsigarginand reduced levels of BiP and CHOP [176]
Aforementioned compounds were chosen from othersas the most potent ER stress inhibitors and persuasivelyprotective to neuronal cells 18 of 42 different compounds
Oxidative Medicine and Cellular Longevity 11
were exploited in in vivo and in vitro models of centralnervous system disorders with in fact improved cell or tissueviability [166] Thus the brain is the most frequently investi-gated organ in the context of ERS From these experimentsit becomes clear that CHOP functions as proapoptotic factorThe roles of other specific ER stress molecules as moleculartargets for pharmacological intervention are less clear andvary depending on cell type and context
There are few underestimated modulations in APP pro-cessing that shed more light on current dogma of AD patho-genesis The modulation of mevalonate pathway and choles-terol synthesis were reported to stimulate nonamyloidogenicpathway of APP processing [177] Additionally cholesterolderivative 27-hydroxycholesterol (27-OHC) was shown toinduce ER stress which attenuated leptin-dependent viabilityby activating CHOP in SH-SY5Y neuroblastoma cells [178]Irrespective of a number of compounds examined in AD thecall for new drugs modulating ER stress with healing effect isstill waiting to be revealed
Abbreviations
APOE Apolipoprotein EAPPs Secreted form of APPCHOPGADD153 CEBP homologous protein
growth arrest and DNA damageinduced gene 153
Drp1 Dynamin-1-like proteinERAD Endoplasmic-reticulum-
associated protein degradationERO1120572 ER oxidase 1120572ERS Endoplasmic reticulum stressFOH FarnesolGGOH GeranylgeraniolGRP75 Glucose-regulated protein 75GRP78BiP 78 kDa glucose-regulated proteinGSK-3120573 Synthase kinase 3 betaIP3R Inositol triphosphate receptor
IRE1120572 Inositol-requiring enzyme 1 alphaMAM Mitochondria-associated
membraneMAP LC3 Microtubule-associated protein
light chain 3Mfn2 GTPase mitofusin 2MMP Mitochondrial membrane
potentialMPTP Mitochondrial permeability
transition poreNFT Neurofibrillary tanglesPACS-2 Phosphofurin acidic cluster
sorting protein 2PDI Protein disulfide isomerasePERK Protein kinase R- (PKR-) like
ER kinaseRNS Reactive nitrogen speciesROS Reactive oxygen speciesRyR Ryanodine receptorTOM Transporter outer membraneTIM Transporter inner membrane
UPR Unfolded protein responseVDAC Voltage-dependent anion channelXBP-1 Xbox binding protein 1
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
Support for this work was provided by Grant no UMO-201103BNZ70238 from the National Science Centre inPoland
References
[1] R L Nussbaum andC E Ellis ldquoAlzheimerrsquos disease and Parkin-sonrsquos diseaserdquoTheNewEngland Journal ofMedicine vol 348 no14 pp 1356ndash1364 2003
[2] A L Brunkan and A M Goate ldquoPresenilin function and 120574-secretase activityrdquoThe Journal of Neurochemistry vol 93 no 4pp 769ndash792 2005
[3] V Crentsil ldquoThe pharmacogenomics of Alzheimerrsquos diseaserdquoAgeing Research Reviews vol 3 no 2 pp 153ndash169 2004
[4] D L Price and S S Sisodia ldquoMutant genes in familialAlzheimerrsquos disease and transgenic modelsrdquo Annual Review ofNeuroscience vol 21 pp 479ndash505 1998
[5] D Harman ldquoAlzheimerrsquos disease pathogenesis role of agingrdquoAnnals of the New York Academy of Sciences vol 1067 no 1 pp454ndash460 2006
[6] D J Selkoe ldquoTranslating cell biology into therapeutic advancesin Alzheimerrsquos diseaserdquo Nature vol 399 pp A23ndashA31 1999
[7] H Braak and E Braak ldquoNeuropathological stageing of Alz-heimer-related changesrdquo Acta Neuropathologica vol 82 no 4pp 239ndash259 1991
[8] K Hirai G Aliev A Nunomara et al ldquoMitochondrial abnor-malities inAlzheimerrsquos diseaserdquoTheJournal ofNeuroscience vol21 pp 3017ndash3023 2001
[9] U T Brunk C B Jones and R S Sohal ldquoA novel hypothesisof lipofuscinogenesis and cellular aging based on interac-tions between oxidative stress and autophagocytosisrdquoMutationResearchDNAging vol 275 no 3ndash6 pp 395ndash403 1992
[10] P A Stewart K Hayakawa M-A Akers and H V Vinters ldquoAmorphometric study of the blood-brain barrier in Alzheimerrsquosdiseaserdquo Laboratory Investigation vol 67 no 6 pp 734ndash7421992
[11] J P Blass and G E Gibson ldquoThe role of oxidative abnor-malities in the pathophysiology of Alzheimerrsquos diseaserdquo RevueNeurologique vol 147 no 6-7 pp 513ndash525 1991
[12] B Pająk E Kania and A Orzechowski ldquoNucleofection of ratpheochromocytoma PC-12 cells with human mutated beta-amyloid precursor protein gene (APP-sw) leads to reducedreduced viability autophagy-like process and increased expres-sion and secretion of beta amyloidrdquo BioMed Research Interna-tional vol 2015 Article ID 746092 10 pages 2015
[13] J P Blass ldquoCerebrometabolic abnormalities in AlzheimerrsquosdiseaserdquoNeurological Research vol 25 no 6 pp 556ndash566 2003
12 Oxidative Medicine and Cellular Longevity
[14] A Eckert K Schmitt and J Gotz ldquoMitochondrial dysfunc-tionmdashthe beginning of the end in Alzheimerrsquos disease Sep-arate and synergistic modes of tau and amyloid-120573 toxicityrdquoAlzheimerrsquos Research ampTherapy vol 3 article 15 pp 1ndash11 2011
[15] U Keil A Bonert C A Marques et al ldquoAmyloid 120573-inducedchanges in nitric oxide production and mitochondrial activitylead to apoptosisrdquoThe Journal of Biological Chemistry vol 279no 48 pp 50310ndash50320 2004
[16] A M Swomley and D A Butterfield ldquoOxidative stress inAlzheimer disease and mild cognitive impairment evidencefrom human data provided by redox proteomicsrdquo Archives ofToxicology vol 89 no 10 pp 1669ndash1680 2015
[17] C Caspersen N Wang J Yao et al ldquoMitochondrial A120573 apotential focal point for neuronal metabolic dysfunction inAlzheimerrsquos diseaserdquo The FASEB Journal vol 19 no 14 pp2040ndash2041 2005
[18] G E Gibson K-F R Sheu and J P Blass ldquoAbnormalities ofmitochondrial enzymes inAlzheimer diseaserdquo Journal of NeuralTransmission vol 105 no 8-9 pp 855ndash870 1998
[19] A Eckert S Hauptmann I Scherping et al ldquoSoluble beta-amyloid leads to mitochondrial defects in amyloid precursorprotein and tau transgenic micerdquo Neurodegenerative Diseasesvol 5 no 3-4 pp 157ndash159 2008
[20] A Eckert K L Schulz V Rhein and J Gotz ldquoConvergenceof amyloid-120573 and tau pathologies on mitochondria in vivordquoMolecular Neurobiology vol 41 no 2-3 pp 107ndash114 2010
[21] S Hauptmann I Scherping S Drose et al ldquoMitochondrial dys-function an early event in Alzheimer pathology accumulateswith age in AD transgenic micerdquoNeurobiology of Aging vol 30no 10 pp 1574ndash1586 2009
[22] M Manczak B S Park Y Jung and P H Reddy ldquoDifferentialexpression of oxidative phosphorylation genes in patients withAlzheimerrsquos disease implications for early mitochondrial dys-function and oxidative damagerdquo NeuroMolecular Medicine vol5 no 2 pp 147ndash162 2004
[23] M P Mattson M Gleichmann and A Cheng ldquoMitochondriain neuroplasticity and neurological disordersrdquo Neuron vol 60no 5 pp 748ndash766 2008
[24] P I Moreira M S Santos and C R Oliveira ldquoAlzheimerrsquosdisease a lesson frommitochondrial dysfunctionrdquoAntioxidantsamp Redox Signaling vol 9 no 10 pp 1621ndash1630 2007
[25] P H Reddy ldquoMitochondrial dysfunction in aging and Alz-heimerrsquos disease strategies to protect neuronsrdquo Antioxidants ampRedox Signaling vol 9 no 10 pp 1647ndash1658 2007
[26] V Rhein G Baysang S Rao et al ldquoAmyloid-beta leads toimpaired cellular respiration energy production andmitochon-drial electron chain complex activities in humanneuroblastomacellsrdquo Cellular and Molecular Neurobiology vol 29 no 6-7 pp1063ndash1071 2009
[27] V Rhein X Song A Wiesner et al ldquoAmyloid-120573 and tausynergistically impair the oxidative phosphorylation system intriple transgenic Alzheimerrsquos disease micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 47 pp 20057ndash20062 2009
[28] R X Santos S C Correia X Wang et al ldquoAlzheimerrsquos diseasediverse aspects ofmitochondrial malfunctioningrdquo InternationalJournal of Clinical and Experimental Pathology vol 3 no 6 pp570ndash581 2010
[29] N R Sims ldquoEnergy metabolism oxidative stress and neuronaldegeneration in Alzheimerrsquos diseaserdquoNeurodegeneration vol 5no 4 pp 435ndash440 1996
[30] H K Anandatheerthavarada G Biswas M-A Robin and NG Avadhani ldquoMitochondrial targeting and a novel transmem-brane arrest of Alzheimerrsquos amyloid precursor protein impairsmitochondrial function in neuronal cellsrdquo The Journal of CellBiology vol 161 no 1 pp 41ndash54 2003
[31] L Devi B M Prabhu D F Galati N G Avadhani and HK Anandatheerthavarada ldquoAccumulation of amyloid precur-sor protein in the mitochondrial import channels of humanAlzheimerrsquos disease brain is associated with mitochondrialdysfunctionrdquo The Journal of Neuroscience vol 26 no 35 pp9057ndash9068 2006
[32] L Devi and H K Anandatheerthavarada ldquoMitochondrialtrafficking of APP and alpha synuclein relevance to mito-chondrial dysfunction in Alzheimerrsquos and Parkinsonrsquos diseasesrdquoBiochimica et Biophysica Acta vol 1802 no 1 pp 11ndash19 2010
[33] M Manczak T S Anekonda E Henson B S Park J Quinnand P H Reddy ldquoMitochondria are a direct site of A120573 accumu-lation in Alzheimerrsquos disease neurons implications for free rad-ical generation and oxidative damage in disease progressionrdquoHuman Molecular Genetics vol 15 no 9 pp 1437ndash1449 2006
[34] P F Pavlov C H Petersen E Glaser and M AnkarcronaldquoMitochondrial accumulation of APP and A120573 significancefor Alzheimer disease pathogenesisrdquo Journal of Cellular andMolecular Medicine vol 13 no 10 pp 4137ndash4145 2009
[35] C Schmidt E Lepsverdize S L Chi et al ldquoAmyloid precursorprotein and amyloid 120573-peptide bind to ATP synthase andregulate its activity at the surface of neural cellsrdquo MolecularPsychiatry vol 13 no 10 pp 953ndash969 2008
[36] K Johar A Priya and M T T Wong-Riley ldquoRegulation ofNa+K+-ATPase by nuclear respiratory factor 1 implication inthe tight coupling of neuronal activity energy generation andenergy consumptionrdquo The Journal of Biological Chemistry vol287 no 48 pp 40381ndash40390 2012
[37] J Astrup P M Sorensen and H R Sorensen ldquoOxygen andglucose consumption related to Na+-K+ transport in caninebrainrdquo Stroke vol 12 no 6 pp 726ndash730 1981
[38] M Erecinska and I A Silver ldquoATP and brain functionrdquo Journalof Cerebral Blood Flow and Metabolism vol 9 no 1 pp 2ndash191989
[39] MMata D J Fink H Gainer et al ldquoActivity-dependent energymetabolism in rat posterior pituitary primarily reflects sodiumpump activityrdquo Journal of Neurochemistry vol 34 no 1 pp 213ndash215 1980
[40] J Kang H-G Lemaire A Unterbeck et al ldquoThe precursor ofAlzheimerrsquos disease amyloid A4 protein resembles a cell-surfacereceptorrdquo Nature vol 325 no 6106 pp 733ndash736 1987
[41] C L Masters and D J Selkoe ldquoBiochemistry of amyloid 120573-protein and amyloid deposits in Alzheimer diseaserdquoCold SpringHarbour Perspectives in Medicine vol 2 no 6 Article IDa006262 2012
[42] A Lai S S Sisodia and I S Trowbridge ldquoCharacterization ofsorting signals in the beta-amyloid precursor protein cytoplas-mic domainrdquo The Journal of Biological Chemistry vol 270 no8 pp 3565ndash3573 1995
[43] A I Placido C M F Pereira A I Duarte et al ldquoThe role ofendoplasmic reticulum in amyloid precursor protein processingand trafficking implications for Alzheimerrsquos diseaserdquo Biochim-ica et Biophysica ActamdashMolecular Basis of Disease vol 1842 no9 pp 1444ndash1453 2014
Oxidative Medicine and Cellular Longevity 13
[44] C Kaether P Skehel and C G Dotti ldquoAxonal membraneproteins are transported in distinct carriers a two-color videomicroscopy study in cultured hippocampal neuronsrdquoMolecularBiology of the Cell vol 11 no 4 pp 1213ndash1224 2000
[45] A Kamal A Almenar-Queralt J F LeBlanc E A Robertsand L S B Goldstein ldquoKinesin-mediated axonal transportof a membrane compartment containing 120573-secretase andpresenilin-1 requires APPrdquo Nature vol 414 no 6864 pp 643ndash648 2001
[46] R G Perez S Soriano J D Hayes et al ldquoMutagenesis identi-fies new signals for 120573-amyloid precursor protein endocytosisturnover and the generation of secreted fragments includingA12057342rdquo The Journal of Biological Chemistry vol 274 no 27 pp18851ndash18856 1999
[47] D G Cook M S Forman J C Sung et al ldquoAlzheimerrsquos A120573(1ndash42) is generated in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo Nature Medicine vol 3 no 9 pp1021ndash1023 1997
[48] A S C Chyung B D Greenberg D G Cook R W Domsand V M-Y Lee ldquoNovel 120573-secretase cleavage of 120573-amyloidprecursor protein in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo The Journal of Cell Biology vol138 no 3 pp 671ndash680 1997
[49] RW-Y Choy Z Cheng and R Schekman ldquoAmyloid precursorprotein (APP) traffics from the cell surface via endosomesfor amyloid 120573 (A120573) production in the trans-Golgi networkrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 30 pp E2077ndashE2082 2012
[50] S A Small and S Gandy ldquoSorting through the cell biologyof Alzheimerrsquos disease intracellular pathways to pathogenesisrdquoNeuron vol 52 no 1 pp 15ndash31 2006
[51] M Goedert ldquoAlzheimerrsquos and Parkinsonrsquos diseases the prionconcept in relation to assembled A120573 tau and 120572-synucleinrdquoScience vol 349 no 6248 2015
[52] I Benilova E Karran and B De Strooper ldquoThe toxic A120573oligomer and Alzheimerrsquos disease an emperor in need ofclothesrdquo Nature Neuroscience vol 15 no 3 pp 349ndash357 2012
[53] E H Corder A M Saunders W J Strittmatter et al ldquoGenedose of apolipoprotein E type 4 allele and the risk of Alzheimerrsquosdisease in late onset familiesrdquo Science vol 261 no 5123 pp 921ndash923 1993
[54] S Brunholz S Sisodia A Lorenzo C Deyts S Kins and GMorfini ldquoAxonal transport of APP and the spatial regulationof APP cleavage and function in neuronal cellsrdquo ExperimentalBrain Research vol 217 no 3-4 pp 353ndash364 2012
[55] G Thinakaran and E H Koo ldquoAmyloid precursor proteintrafficking processing and functionrdquo The Journal of BiologicalChemistry vol 283 no 44 pp 29615ndash29619 2008
[56] T Saitoh M Sundsmo J-M Roch et al ldquoSecreted formof amyloid beta protein precursor is involved in the growthregulation of fibroblastsrdquo Cell vol 58 no 4 pp 615ndash622 1989
[57] M PMattson B Cheng A R Culwell F S Esch I Lieberburgand R E Rydel ldquoEvidence for excitoprotective and intraneu-ronal calcium-regulating roles for secreted forms of the 120573-amyloid precursor proteinrdquo Neuron vol 10 no 2 pp 243ndash2541993
[58] K J Lee C E H Moussa Y Lee et al ldquoBeta amyloid-independent role of amyloid precursor protein in generationandmaintenance of dendritic spinesrdquoNeuroscience vol 169 no1 pp 344ndash356 2010
[59] H Meziane J-C Dodart C Mathis et al ldquoMemory-enhancingeffects of secreted forms of the 120573-amyloid precursor proteinin normal and amnestic micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no21 pp 12683ndash12688 1998
[60] P R Turner K OrsquoConnor W P Tate and W C AbrahamldquoRoles of amyloid precursor protein and its fragments inregulating neural activity plasticity and memoryrdquo Progress inNeurobiology vol 70 no 1 pp 1ndash32 2003
[61] D Beher L Hesse C L Masters and GMulthaup ldquoRegulationof Amyloid Protein Precursor (APP) binding to collagen andmapping of the binding sites on APP and collagen type Irdquo TheJournal of Biological Chemistry vol 271 no 3 pp 1613ndash16201996
[62] S L Sabo A F Ikin J D Buxbaum and P Greengard ldquoTheamyloid precursor protein and its regulatory protein FE65 ingrowth cones and synapses in vitro and in vivordquo The Journal ofNeuroscience vol 23 no 13 pp 5407ndash5415 2003
[63] D H Small V Nurcombe G Reed et al ldquoA heparin-bindingdomain in the amyloid protein precursor of Alzheimerrsquos diseaseis involved in the regulation of neurite outgrowthrdquoThe Journalof Neuroscience vol 14 no 4 pp 2117ndash2127 1994
[64] T G Williamson S S Mok A Henry et al ldquoSecreted glypicanbinds to the amyloid precursor protein of Alzheimerrsquos disease(APP) and inhibits APP-induced neurite outgrowthrdquoThe Jour-nal of Biological Chemistry vol 271 no 49 pp 31215ndash312211996
[65] T Ondrejcak I Klyubin N-W Hu A E Barry W K Cullenand M J Rowan ldquoAlzheimerrsquos disease amyloid 120573-protein andsynaptic functionrdquo NeuroMolecular Medicine vol 12 no 1 pp13ndash26 2010
[66] D Puzzo L Privitera and A Palmeri ldquoHormetic effect ofamyloid-beta peptide in synaptic plasticity and memoryrdquo Neu-robiology of Aging vol 33 no 7 pp 1484e15ndash1484e24 2012
[67] C M F Pereira ldquoCrosstalk between endoplasmic reticulumstress and protein misfolding in neurodegenerative diseasesrdquoISRN Cell Biology vol 2013 Article ID 256404 22 pages 2013
[68] R-W Shin T C Saido M Maeda and T Kitamoto ldquoNovel 120572-secretase cleavage ofAlzheimerrsquos amyloid120573 precursor protein inthe endoplasmic reticulum of COS7 cellsrdquoNeuroscience Lettersvol 376 no 1 pp 14ndash19 2005
[69] E A Schon and E Area-Gomez ldquoMitochondria-associatedER membranes in Alzheimer diseaserdquo Molecular and CellularNeuroscience vol 55 pp 26ndash36 2013
[70] R Rizzuto P Pinton W Carrington et al ldquoClose contacts withthe endoplasmic reticulum as determinants of mitochondrialCa2+ responsesrdquo Science vol 280 no 5370 pp 1763ndash1766 1998
[71] S E Logue P Cleary S Saveljeva and A Samali ldquoNewdirections in ER-stress induced cell deathrdquoApoptosis vol 18 no5 pp 537ndash546 2013
[72] J P Decuypere KWelkenhuyzen T Luyten et al ldquoIns(145)P3receptor-mediated BI-1 Ca2+ signaling and autophagy induc-tion are interrelatedrdquo Autophagy vol 7 pp 1472ndash1489 2011
[73] K S Vetrivel and G Thinakaran ldquoMembrane rafts in Alz-heimerrsquos disease beta-amyloid productionrdquo Biochimica et Bio-physica Acta vol 1801 no 8 pp 860ndash867 2010
[74] H Urra E Dufey F Lisbona D Rojas-Rivera and C HetzldquoWhen ER stress reaches a dead endrdquo Biochimica et BiophysicaActa (BBA) Molecular Cell Research vol 1833 no 12 pp 3507ndash3517 2013
14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 7
vesicles and undergoes retrograde transport from cis endof the Golgi complex back to the ER [76] Such responsecauses accumulation of APP in the tubulocisternal ER systemtogether with aberrant intracellular translocation of theprotein Interestingly the issue whether APP is subject toretrograde transport with successive fibrillogenic processingbecause of UPR and its interaction with GRP78BiP is notclear as the observations are inconsistent [77] Nonethelessaccumulation of misfoldedmalfolded proteins in the ERsuggests disorganized process of posttranslational change Asanticipated the accretion of proteins of anomalous patternsignals ERS and UPR followed by increased vulnerability toapoptotic cell death Prior to decay however APP proteinlevels in the ER lessen by dint of cleavage with 120573- and120574-secretase [105 106] Products of this cleavage (sAPP120573A120573 and AICD fragment) all appreciably influence neuronalsurvival most likely through nonnative form of the APPsubstrate
10 APP A120573 and NFTs MarkMitochondria as Targeted in AD
Mitochondrial import of A12057340
and A12057342
peptides throughthe translocase TOM complex was blocked by preincubationof isolated mitochondria with antibodies raised againstTOM proteins (TOM20 TOM40 and TOM70) [107]Neither VDAC inhibition with antagonist antibodies norinhibition of mitochondrial permeability transition pores(MPTP) or fall of mitochondrial membrane potential(MMP) affected uptake of A120573 [17] With regard to APPelegant study performed by Anandatheerthavarada andhis colleagues [30] revealed that C-terminal truncatedAPP (lacking A120573) targets mitochondria in cholinergicGABAergic dopaminergic and glutamatergic neurons ofAD brain by clogging up mitochondrial protein translocasecomplex TOM40TIM22 Authors propose that occlusionof translocase is followed by blunted import of nuclear-encoded proteins vitally important for energy homeostasisMitochondrial APP protein transmembrane orientationindicates NH
2-terminal inside in contact with translocase
whereas COOH-terminal is facing cytoplasmic side NH2-
terminus has mitochondrial signal sequence Astonishinglymitochondrial APP molecules were nonglycosylated givingrise to speculation that protein molecules that arrivedat mitochondria have not achieved molecular maturity[31] The accumulation of nonglycosylated APP species inmitochondrial import channels of AD brain was directlyrelated to decreased mitochondrial functions as validatedby the decline in cytochrome 119888 oxidase activity (complexIV) and elevated levels of H
2O2 Furthermore in AD
brain the mitochondrial accumulation of nonglycosylatedAPP went along with a corresponding reduction in plasmamembrane-associatedAPP It suggests that ADbrain hasAPPprocessing and trafficking severely affected by incompleteN-glycosylation ensuing ER protein accumulation andexposure of the cryptic mitochondrial targeting signalfor assisting chaperone proteins Several lines of evidenceindicate that not fully formed proteins are phosphorylatedand bind to the cytosolic proteins required for movement
from ER to mitochondria [108ndash110] Moreover ERS andUPR in AD seem to be incompetent and inefficient inelimination of malfolded proteins In addition to APPalso A120573 was frequently reported to occupy mitochondriaalthough its origin and mechanism of mitochondriatargeting mostly remain unexplored (except for involvementof protein translocase TOM complex) The possibilityof mitochondrial A120573 generation has to be ruled out asmembrane orientation of arrested APP does not allow accessto 120574-secretase (120574-secretase activity in mitochondria wasdetected by independent study) [111] Probably A120573 speciesis derived from APP prior to its translocation (ER) soA120573 may be transported to mitochondria independentlyof APP Collectively observations showing mitochondrialpresence of APP A120573 and tau in aberrant configuration pointtoward the anomaly of protein folding at the level of ER andGolgi apparatus Nonglycosylated molecule of APP suggestsdefective transfer of sugar core from dolichyl phosphate(s)and further modifications such as O- and N-glycosylationsOne may admit that the lack of glycosyl residue bringsabout pathologic processing and trafficking of APP and itsfragments In point of fact dolichol derivatives mixture ofpolyprenols (acyclic isoprenoid alcohols) known as Ropren(Solagran Limited Melbourne Australia) commerciallyused to treat liver diseases were tested in the treatment ofAD in two separate trials conducted in 2005 and 2006 withpromising results [httpwwwasxcomauasxpdf20071119pdf315x5nh4hm8wv7pdf httpwwwasxcomauasxpdf20070221pdf3111ztwcbqzkk9pdf] Further studies areurgently needed to test how important the glycosylationprocess is in the pathogenesis of AD Dolichols areobtained from geranylgeraniol (GGOH) and the latter is anintermediate of mevalonate pathway GGOH is a commonsubstrate for dolichol and ubiquinone synthesis but it isalso necessary for protein prenylation As both GGOHand farnesol (FOH) are engaged in protein prenylationmore concern should be laid on the importance of thesecompounds in AD pathogenesis Observations demonstrat-ing mitochondrial relocalization of other proteins withoutposttranslational modification seemingly point to increasedmitochondrial targeting of immature molecules resultingin mitochondrial dysfunction and acceleration of diseaseprogression [34 112ndash114]
11 Oxidative Stress in Mitochondria ofAD Brains
Normal physiological functions of APP are thought to beinvolved in the stabilizing contact points between synapsesand maintaining mitochondrial functions [60 115] Mito-chondrial dysfunction was often observed regardless of theexperimental model used to study AD [8 29 116ndash119]It includes defects in oxidative phosphorylation decreasedATP decreased membrane potential increased productionof ROSRNS and perturbation in mitochondrial fusion andfission [15 30 31 115 120ndash122] Hyperphosphorylated tau wasalso reported to impair mitochondrial functions [123] Usingproteomic approach the strongest defects of the respiratorycapacity were observed mainly at complexes I IV and ATP
8 Oxidative Medicine and Cellular Longevity
synthase (complex V) at both protein and activity level [124]While APP A120573 and hyperphosphorylated tau are potentinhibitors of mitochondrial import of nuclear-encoded pro-teins apparently the pathology of each leads to metabolicharm in different way In freshly isolated mitochondriafrom AD brains the APP inhibited mitochondrial importof cytochrome 119888 oxidase (COX) subunits IV and Vb [31]Dysfunction of COX increases ROS production (incompletereduction of oxygen molecules) reduces energy stores anddisturbs energy metabolism Accordingly in AD patientsdeficiency of COX was found in brains and platelets [119125 126] Similar to APP A120573 was found in mitochondriaof transgenic mice and cellular and human AD models [1719 33 103 127ndash130] At present it is not clear whether theobservedmitochondrial toxicity is due toAPP orA120573 orNFTsaccumulation Anyway some regularity is observed withrespect to most affected components of respiratory chainNADH-ubiquinone oxidoreductase (complex I) activity isreduced to the utmost by hyperphosphorylated tau whereasdecreased activity of cytochrome 119888 oxidase (complex IV)that resulted in mitochondrial dysfunction was observedduring A120573 and APP accumulation [14 15 31 32 34]Concomitantly rise in the activity of antioxidant enzymesmanganese superoxide dismutase (Mn-SOD) and catalase(CAT) was demonstrated in response to elevated levelsof free radicals including superoxide anion radical (O
2
∙minus)hydroperoxyl radical (HO
2
∙) hydroxyl radical (OH∙) andnitric oxide radical (NO∙) [15] Consistent with observationsof chronic respiratory chain dysfunction and mitochondrialoxidative stress there are reports showing their contributionto tau pathology in AD [131] In any case free radicals thatoverride antioxidant defense react with a wide variety oforganic components causing lipid peroxidation to advancedlipid oxidation end products (ALE) cross-linking of proteinsnitrosylation of proteins and mutations in DNA Mitochon-drial circular DNA (mtDNA) of sim16 kbp is devoid of repairsystems meaning the buildup of lesions There are 37 geneslocated in mtDNA with those encoding protein subunits ofcomplex I (7) complex II (1) and complex IV (3) but notcomplex III Interestingly hallmarks of AD (APP A120573 andNFTs)mostly affectmembers of electron transfer chain (com-plexes I and IV) which rely exclusively on mitochondriallypredetermined subunits Maybe it is not simple coincidencebut the effect of inhibited import to mitochondria of nuclear-encoded subunits of complexes I and IV gives explanationfor toxicity of APP or A120573 or NFTs in mitochondria Thereare additional findings in mitochondria affected by AD suchas lower activity of pyruvate dehydrogenase (PDH) andoxoglutarate dehydrogenase (OGDH) [104 132] Inhibitionof OGDH the enzyme of tricarboxylic acid cycle minimizesthe NADH pool and electron number needed for ETCand mitochondrial membrane potential (Δ120595
119898) to create and
maintain proton gradient obligatory for ATP synthesisThereare also lines of evidence for direct inhibitory action of solubleoligomeric A120573 species on ABAD (A120573-binding alcohol dehy-drogenase) and internal membrane cyclophilin D (CypD)resulting in increased mitochondrial membrane permeabil-ity (MPTP) potentiated ROS production synaptic lossdiminished activity of mitochondrial respiration and finally
cell death [129 132ndash134] CypD knockout prevents mito-chondrial and neuronal perturbations and improves mito-chondrial function in Alzheimerrsquos disease mouse model [128135] It has to be emphasized that oxidativenitrosative stressaffects the fusion and fission process ofmitochondria Fusionthat is speedup by small GTPases mitofusins (Mfn1Mfn2)improves efficiency of mitochondrial respiration and ATPproduction Mitochondrial dynamics are severely imbal-anced in AD cases in favour of fission through elevatedexpression of the fission protein DLP1 (dynamin-like protein1) associatedwith nitrosative stress stirred up byA120573 [136 137]Collectively these observations indicate that toxic intracel-lular A120573
42-43 oligomers differ in action from extracellularaggregates found in amyloid plaques of AD brains [138](Figure 3)
12 Proteostasis in Mitochondria
Mitochondrial protein turnover grants the well-organizedreplacement of nonfunctional proteins into operational oneThe first task is attained through proteolytic degradation ofinner membrane and matrix proteins with local mitochon-drial proteases [139] or outer membrane proteins throughubiquitin-proteasome system [140] The second task is metby intramitochondrial protein synthesis but as almost 1500different nuclear-encoded proteins have to be importedto complete mitochondrial proteome a matter of capableimport is fundamental for the function of the organelleIn extreme cases of cellular injuries observed in neurode-generative diseases damaged mitochondria with extendedloss of the electrochemical potential are selectively removedby autophagy known as mitophagy [112] APP and A120573accumulation has also something to do with altered mito-chondrial dynamics as fission takes advantage of fusion withresultant dysfunction of mitochondria and neurons [137]AD neurons demonstrated selective mechanisms of prote-olytic clearance of oxidatively and nitrosatively modifiedproteins in mitochondria Insulin degrading enzyme (IDE)prevents formation of toxic insoluble fibrils from A120573 asit cleaves A120573 prior to aggregation [141ndash143] A novel zinc-metallopeptidase Presequence Protease (PreP) a member ofpitrilysin oligopeptidase family degrades either intramito-chondrially stored A120573
40or A120573
42proteinThis protease seems
to be highly sensitive to oxidative stress as disulfide bridgeformed between two proximal cysteine residues blocks itscatalytic activity [144ndash149] Another mitochondrial serineprotease HtrA2Omi occupies intermembrane space whereit can cleave APP locked up in protein translocase complex[121] Even though it is well established that HtrA2Omiis released to the cytosol to amplify apoptosis through thedegradation of antiapoptotic proteins and caspase activation[150ndash152] it is also implicated in proteolytic deletion ofmalfoldedAPP in ER [151] Fromknockout studies onmice itis obvious that HtrA2Omi plays a significant shielding roleas mice deficient in this protease exhibit neurodegenerativephenotype with weight loss and premature death [153]Taken together understanding the mechanisms of clearanceof the unwanted proteins including APP A120573 and tau isvital in an attempt to get rid of them from mitochondria
Oxidative Medicine and Cellular Longevity 9
VDAC
ANT
A120573
A120573
A120573
A120573
A120573
A120573
A120573A120573
A120573
A120573
A120573 CypD
CypD ProApCyt c Ca2+
IMM
OMM
IMSΔΨ
+
+
+
minus
minus
minus
ABAD
ROS
ROS
TIM
TOM
Proteinimport
NAD
NAD
NAD
NAD
H+
H+
H+
H+
H+
ADP
Mn-SOD
CuZn-SOD
mtDNA
V
Matrix
UCP
O2∙minus
O2∙minus
IV
III
II
I
O2
O2
DLP1Fis1
Mfn1Mfn2
Opa1
mHsp70
NADH
NADH
NADH
TCAcycle
PyruvateAcetyl-CoA
120572KGDH
120572-KG
PDH
Isocitrate
CitrateOAA
Malate
Fumarate
Succinyl-CoASuccinate
GDPGTP
FAD
FAOH
ATP
Figure 3 Amyloid-120573-related mitochondrial impairment Mitochondria were found to be the target for amyloid-120573 (A120573) which interactswith several proteins leading to mitochondrial dysfunction Indeed A120573 was found in the outer mitochondrial membrane (OMM) andinner mitochondrial membrane (IMM) as well as in the matrix The interaction of A120573 with the OMM affects the transport of nuclear-encoded mitochondrial proteins such as subunits of the electron transport chain complex IV into the organelle via the translocase of theouter membrane (TOM) import machinery Moreover A120573 disturbs the activity of several enzymes such as pyruvate dehydrogenase (PDH)and oxoglutarate dehydrogenase (OGDH) decreasing NADH reduction and the electron transport chain enzyme complex IV reducing theamount of hydrogen that is translocated from the matrix to the intermembrane space (IMS) thus impairing the mitochondrial membranepotential (MMP) Taken together these events cause abnormal mitochondrial electron activities leading to decreased complex V activityand so to a drop in ATP levels in addition to increasing reactive oxygen species (ROS) generation Moreover ROS induce peroxidationof several mitochondrial macromolecules such as mitochondrial DNA (mtDNA) and mitochondrial lipids contributing to mitochondrialimpairment in themitochondrial matrixThe complex of A120573 bound to binding alcohol dehydrogenase (ABAD) impairs the binding of NAD+to ABAD changes mitochondrial membrane permeability and reduces activities of respiratory enzymes inducing further ROS productionand leading to mitochondrial failure A120573 binding also activates Fis1 (fission protein) and promotes increased mitochondrial fragmentationthis increased mitochondrial fragmentation produces defective mitochondria that ultimately damage neurons Furthermore A120573 binding tocyclophilin D (CypD) enhances the protein translocation to the inner membrane favouring the opening of the mitochondrial permeabilitytransition pore formed by the adenine nucleotide translocator (ANT) and voltage-dependent anion channels (VDACs) Cyt 119888 cytochrome 119888DLP1 dynamin-like protein 1 PDH pyruvate dehydrogenase ProAp proapoptotic factors SOD superoxide dismutase TCA tricarboxylicacid TIM translocase of the inner membrane Adopted from [14]
Some hopes are associated with the application of at presentindefinite modulators of proteolytic activity HtrA2Omi isfor example activated by PTEN-induced putative kinase 1(PINK1) upon phosphorylation at Ser142 residue [154] Alter-natively accent has to be put on mitophagy of dysfunctionalmitochondria and mitochondriogenesis
13 Aging versus AD
Themost commonneurodegenerative disorder is representedby Alzheimerrsquos disease characterized by declining memoryreduced cognitive capacity and progressive dementia whichare often fatal to elderly individuals above 65 years of age
10 Oxidative Medicine and Cellular Longevity
It is ranked as the fourth leading cause of death in modernsocieties where average life span increased greatly in the lasttwo decades As 95 of AD cases are diagnosed in olderpeople one might think that a causal relationship existsbetween aging and the onset of disease Certainly a numberof similarities between getting old and being affected withAD could be listed Historically the free radical theory ofaging by Harman [155] suggested aging as ldquoside effectrdquo ofreactive oxygen species formed in mitochondrial respiratorychain Apparently free radicals commonly generated byincomplete reduction of oxygen molecule at complexes Iand III of mitochondrial ETC are capable of damagingDNA RNA and proteins They impair energy storage andlead to operational failure of mitochondria with progres-sive decline of cell viability Almost identical conditionsaccompany AD and are an explicit step in pathogenesisof disease As mtDNA is deprived of repair mechanismsROS-induced DNA strand breaks tend to accumulate withage or AD Thus mtDNA is a vulnerable target for ROSbut the reverse the ROS generation due to the mutatedmtDNA is not convincingly confirmed [156] Moreover theevidence that mutated mtDNA accelerates the progress ofaging is also questioned based on the results from studycarried out on transgenic mice model [157] Although someauthors show inconsistency between the free radical theoryand observations cumulative evaluation of the scientificreports points to antioxidant defense systems as importantfactors in protection from premature aging [158] Othermitochondrial components important for their function arehampered with age adenine nucleotide translocase (ANT)nitric oxide synthase (NOS) and carnitine acyltransferase(CT) [159ndash161] Actually NOS activity is elevated in AD asreported from study performed on cellular model of disease[15] We could not find any information about CT activityin AD whereas ANT activity is noticeably inhibited byA120573 or hyperphosphorylated tau and this effect is reversedby mersalyl a reversible alkylating agent of thiol groups[162] Mitochondrial dysfunction observed in transgenicmice models of AD and aging demonstrates higher activ-ity of genes controlling energy metabolism and apoptosisTaken together physiological aging and AD are associatedwith broad-spectrum dysfunction of mitochondria but thefoundations of mitochondrial decline are dissimilar Morediscrepancies between physiological aging and AD werefound with respect to ERS and UPR which play a significantrole in cellular proteostasis Dolichol was selected as an agingmarker because the progressive increase in dolichol levelwas observed in aging brain [163] In contrast ubiquinoneconcentration which is also synthesized from geranylgeranioldiminishes with agingwhereas cholesterol and dolichyl phos-phate concentrations remain unaltered In AD decreasedlevels of dolichol were observed and increased levels ofubiquinone and dolichyl phosphate without any changesin brain cholesterol AD cannot be regarded as a resultof premature aging The drop in dolichol and augmenteddolichyl phosphate concentration points toward disturbedglycosylation in ER of diseased brain while the increase inubiquinone suggests efforts to protect the brain from oxida-tive stress induced by lipid peroxidation [164 165]
14 Targeting ER Stress in AD Therapy
As ERS is a recognized factor in AD drugs that interfere withERS would theoretically have great therapeutic potentialThere are several compounds grouped in classes thatinteract directly with components of the ERS (salubrinalBiP inducer X (BIX) salicylamide analogs flavonoidsguanabenz and STF083010) chemical chaperons (4-phenyl-butyric acid (PBA) tauroursodeoxycholic acid (TUDCA)and trimethylamine oxide (TMAO)) chemicals that inhibitprotein degradation (Eeyarestatin MG132 and Bortezomib)compounds with antioxidant activity (Edaravone diben-zoylmethane derivatives and N-acetyl cysteine (NAC)) anddrugs controlling calcium signaling (dantrolene and carba-zole derivatives) [166] They may act by inducing transienttranslation arrest upregulation of chaperone proteins andaugmented degradation of ER-associated misfolded proteinsFundamental approach in the development of new therapyis the selection of appropriate molecular targets In ER stresssignaling the aim is to alter the expression of ER stress-associated molecules that can rescue cells from the toxiceffect of ERS Recent efforts in establishing new promisingdrugs against AD are pointing to chemical chaperones suchas PBA TUDCA or trimethylamine oxide (TMAO) Thesesubstances improve protein folding and alleviate nativeprotein conformation [166] It was shown on mouse modelsof AD that PBA TUDCA and TMAO stop A120573 accumulationand avoid the loss of dendritic spines [167] Some observa-tions even demonstrated improved memory and cognitivefunctions [168] associated with improved cell survival [169]Salubrinal ((2E)-3-phenyl-N-[222-trichloro-1-[[(8-quinoli-nylamino)thioxomethyl]amino]ethyl]-2-propenamide Sal)selectively inhibits growth arrest and DNA damage inducedgene 34- (GADD34-) phosphatase complex (GADD34associates with protein phosphatase 1 (PP1)) and promotes invitro dephosphorylation of the alpha subunit of eIF-2120572 andIRE1120572ASK1JNK signaling pathway being protective againstERS even induced by tunicamycin Tm [170] In a great deal ofexperiments testing Sal in cultured cells and animal modelsof AD this substance increased the viability of neuronal cellsand A120573 toxicity [171 172] BIX (2-(34-dihydroxyphenyl)-2-oxoethyl ester thiocyanic acid) preferentially inducedBiP mRNA in an ATF6-dependent manner leading toreduced Tm-induced death of neuronal cells [173] AlsoDBM derivative 14ndash26 (221015840-dimethoxydibenzoylmethane)was found to be neuroprotective for SH-SY5Y and PC-12cells by decreasing expression of BiP and CHOP [174]Dantrolene a ryanodine receptor antagonist that inhibitsabnormal calcium release from the ER inhibited expressionof both phosphorylated PERK and eIF2120572 It also reducedCHOP expression and attenuated thapsigargin-inducedapoptosis in PC-12 cells [175] Neuroprotective effects similarto dantrolene were observed for ([9-(3-cyanobenzyl)-14-dimethylcarbazole]) This substance suppressed increases inintracellular Ca2+ in PC-12 cells treated with thapsigarginand reduced levels of BiP and CHOP [176]
Aforementioned compounds were chosen from othersas the most potent ER stress inhibitors and persuasivelyprotective to neuronal cells 18 of 42 different compounds
Oxidative Medicine and Cellular Longevity 11
were exploited in in vivo and in vitro models of centralnervous system disorders with in fact improved cell or tissueviability [166] Thus the brain is the most frequently investi-gated organ in the context of ERS From these experimentsit becomes clear that CHOP functions as proapoptotic factorThe roles of other specific ER stress molecules as moleculartargets for pharmacological intervention are less clear andvary depending on cell type and context
There are few underestimated modulations in APP pro-cessing that shed more light on current dogma of AD patho-genesis The modulation of mevalonate pathway and choles-terol synthesis were reported to stimulate nonamyloidogenicpathway of APP processing [177] Additionally cholesterolderivative 27-hydroxycholesterol (27-OHC) was shown toinduce ER stress which attenuated leptin-dependent viabilityby activating CHOP in SH-SY5Y neuroblastoma cells [178]Irrespective of a number of compounds examined in AD thecall for new drugs modulating ER stress with healing effect isstill waiting to be revealed
Abbreviations
APOE Apolipoprotein EAPPs Secreted form of APPCHOPGADD153 CEBP homologous protein
growth arrest and DNA damageinduced gene 153
Drp1 Dynamin-1-like proteinERAD Endoplasmic-reticulum-
associated protein degradationERO1120572 ER oxidase 1120572ERS Endoplasmic reticulum stressFOH FarnesolGGOH GeranylgeraniolGRP75 Glucose-regulated protein 75GRP78BiP 78 kDa glucose-regulated proteinGSK-3120573 Synthase kinase 3 betaIP3R Inositol triphosphate receptor
IRE1120572 Inositol-requiring enzyme 1 alphaMAM Mitochondria-associated
membraneMAP LC3 Microtubule-associated protein
light chain 3Mfn2 GTPase mitofusin 2MMP Mitochondrial membrane
potentialMPTP Mitochondrial permeability
transition poreNFT Neurofibrillary tanglesPACS-2 Phosphofurin acidic cluster
sorting protein 2PDI Protein disulfide isomerasePERK Protein kinase R- (PKR-) like
ER kinaseRNS Reactive nitrogen speciesROS Reactive oxygen speciesRyR Ryanodine receptorTOM Transporter outer membraneTIM Transporter inner membrane
UPR Unfolded protein responseVDAC Voltage-dependent anion channelXBP-1 Xbox binding protein 1
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
Support for this work was provided by Grant no UMO-201103BNZ70238 from the National Science Centre inPoland
References
[1] R L Nussbaum andC E Ellis ldquoAlzheimerrsquos disease and Parkin-sonrsquos diseaserdquoTheNewEngland Journal ofMedicine vol 348 no14 pp 1356ndash1364 2003
[2] A L Brunkan and A M Goate ldquoPresenilin function and 120574-secretase activityrdquoThe Journal of Neurochemistry vol 93 no 4pp 769ndash792 2005
[3] V Crentsil ldquoThe pharmacogenomics of Alzheimerrsquos diseaserdquoAgeing Research Reviews vol 3 no 2 pp 153ndash169 2004
[4] D L Price and S S Sisodia ldquoMutant genes in familialAlzheimerrsquos disease and transgenic modelsrdquo Annual Review ofNeuroscience vol 21 pp 479ndash505 1998
[5] D Harman ldquoAlzheimerrsquos disease pathogenesis role of agingrdquoAnnals of the New York Academy of Sciences vol 1067 no 1 pp454ndash460 2006
[6] D J Selkoe ldquoTranslating cell biology into therapeutic advancesin Alzheimerrsquos diseaserdquo Nature vol 399 pp A23ndashA31 1999
[7] H Braak and E Braak ldquoNeuropathological stageing of Alz-heimer-related changesrdquo Acta Neuropathologica vol 82 no 4pp 239ndash259 1991
[8] K Hirai G Aliev A Nunomara et al ldquoMitochondrial abnor-malities inAlzheimerrsquos diseaserdquoTheJournal ofNeuroscience vol21 pp 3017ndash3023 2001
[9] U T Brunk C B Jones and R S Sohal ldquoA novel hypothesisof lipofuscinogenesis and cellular aging based on interac-tions between oxidative stress and autophagocytosisrdquoMutationResearchDNAging vol 275 no 3ndash6 pp 395ndash403 1992
[10] P A Stewart K Hayakawa M-A Akers and H V Vinters ldquoAmorphometric study of the blood-brain barrier in Alzheimerrsquosdiseaserdquo Laboratory Investigation vol 67 no 6 pp 734ndash7421992
[11] J P Blass and G E Gibson ldquoThe role of oxidative abnor-malities in the pathophysiology of Alzheimerrsquos diseaserdquo RevueNeurologique vol 147 no 6-7 pp 513ndash525 1991
[12] B Pająk E Kania and A Orzechowski ldquoNucleofection of ratpheochromocytoma PC-12 cells with human mutated beta-amyloid precursor protein gene (APP-sw) leads to reducedreduced viability autophagy-like process and increased expres-sion and secretion of beta amyloidrdquo BioMed Research Interna-tional vol 2015 Article ID 746092 10 pages 2015
[13] J P Blass ldquoCerebrometabolic abnormalities in AlzheimerrsquosdiseaserdquoNeurological Research vol 25 no 6 pp 556ndash566 2003
12 Oxidative Medicine and Cellular Longevity
[14] A Eckert K Schmitt and J Gotz ldquoMitochondrial dysfunc-tionmdashthe beginning of the end in Alzheimerrsquos disease Sep-arate and synergistic modes of tau and amyloid-120573 toxicityrdquoAlzheimerrsquos Research ampTherapy vol 3 article 15 pp 1ndash11 2011
[15] U Keil A Bonert C A Marques et al ldquoAmyloid 120573-inducedchanges in nitric oxide production and mitochondrial activitylead to apoptosisrdquoThe Journal of Biological Chemistry vol 279no 48 pp 50310ndash50320 2004
[16] A M Swomley and D A Butterfield ldquoOxidative stress inAlzheimer disease and mild cognitive impairment evidencefrom human data provided by redox proteomicsrdquo Archives ofToxicology vol 89 no 10 pp 1669ndash1680 2015
[17] C Caspersen N Wang J Yao et al ldquoMitochondrial A120573 apotential focal point for neuronal metabolic dysfunction inAlzheimerrsquos diseaserdquo The FASEB Journal vol 19 no 14 pp2040ndash2041 2005
[18] G E Gibson K-F R Sheu and J P Blass ldquoAbnormalities ofmitochondrial enzymes inAlzheimer diseaserdquo Journal of NeuralTransmission vol 105 no 8-9 pp 855ndash870 1998
[19] A Eckert S Hauptmann I Scherping et al ldquoSoluble beta-amyloid leads to mitochondrial defects in amyloid precursorprotein and tau transgenic micerdquo Neurodegenerative Diseasesvol 5 no 3-4 pp 157ndash159 2008
[20] A Eckert K L Schulz V Rhein and J Gotz ldquoConvergenceof amyloid-120573 and tau pathologies on mitochondria in vivordquoMolecular Neurobiology vol 41 no 2-3 pp 107ndash114 2010
[21] S Hauptmann I Scherping S Drose et al ldquoMitochondrial dys-function an early event in Alzheimer pathology accumulateswith age in AD transgenic micerdquoNeurobiology of Aging vol 30no 10 pp 1574ndash1586 2009
[22] M Manczak B S Park Y Jung and P H Reddy ldquoDifferentialexpression of oxidative phosphorylation genes in patients withAlzheimerrsquos disease implications for early mitochondrial dys-function and oxidative damagerdquo NeuroMolecular Medicine vol5 no 2 pp 147ndash162 2004
[23] M P Mattson M Gleichmann and A Cheng ldquoMitochondriain neuroplasticity and neurological disordersrdquo Neuron vol 60no 5 pp 748ndash766 2008
[24] P I Moreira M S Santos and C R Oliveira ldquoAlzheimerrsquosdisease a lesson frommitochondrial dysfunctionrdquoAntioxidantsamp Redox Signaling vol 9 no 10 pp 1621ndash1630 2007
[25] P H Reddy ldquoMitochondrial dysfunction in aging and Alz-heimerrsquos disease strategies to protect neuronsrdquo Antioxidants ampRedox Signaling vol 9 no 10 pp 1647ndash1658 2007
[26] V Rhein G Baysang S Rao et al ldquoAmyloid-beta leads toimpaired cellular respiration energy production andmitochon-drial electron chain complex activities in humanneuroblastomacellsrdquo Cellular and Molecular Neurobiology vol 29 no 6-7 pp1063ndash1071 2009
[27] V Rhein X Song A Wiesner et al ldquoAmyloid-120573 and tausynergistically impair the oxidative phosphorylation system intriple transgenic Alzheimerrsquos disease micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 47 pp 20057ndash20062 2009
[28] R X Santos S C Correia X Wang et al ldquoAlzheimerrsquos diseasediverse aspects ofmitochondrial malfunctioningrdquo InternationalJournal of Clinical and Experimental Pathology vol 3 no 6 pp570ndash581 2010
[29] N R Sims ldquoEnergy metabolism oxidative stress and neuronaldegeneration in Alzheimerrsquos diseaserdquoNeurodegeneration vol 5no 4 pp 435ndash440 1996
[30] H K Anandatheerthavarada G Biswas M-A Robin and NG Avadhani ldquoMitochondrial targeting and a novel transmem-brane arrest of Alzheimerrsquos amyloid precursor protein impairsmitochondrial function in neuronal cellsrdquo The Journal of CellBiology vol 161 no 1 pp 41ndash54 2003
[31] L Devi B M Prabhu D F Galati N G Avadhani and HK Anandatheerthavarada ldquoAccumulation of amyloid precur-sor protein in the mitochondrial import channels of humanAlzheimerrsquos disease brain is associated with mitochondrialdysfunctionrdquo The Journal of Neuroscience vol 26 no 35 pp9057ndash9068 2006
[32] L Devi and H K Anandatheerthavarada ldquoMitochondrialtrafficking of APP and alpha synuclein relevance to mito-chondrial dysfunction in Alzheimerrsquos and Parkinsonrsquos diseasesrdquoBiochimica et Biophysica Acta vol 1802 no 1 pp 11ndash19 2010
[33] M Manczak T S Anekonda E Henson B S Park J Quinnand P H Reddy ldquoMitochondria are a direct site of A120573 accumu-lation in Alzheimerrsquos disease neurons implications for free rad-ical generation and oxidative damage in disease progressionrdquoHuman Molecular Genetics vol 15 no 9 pp 1437ndash1449 2006
[34] P F Pavlov C H Petersen E Glaser and M AnkarcronaldquoMitochondrial accumulation of APP and A120573 significancefor Alzheimer disease pathogenesisrdquo Journal of Cellular andMolecular Medicine vol 13 no 10 pp 4137ndash4145 2009
[35] C Schmidt E Lepsverdize S L Chi et al ldquoAmyloid precursorprotein and amyloid 120573-peptide bind to ATP synthase andregulate its activity at the surface of neural cellsrdquo MolecularPsychiatry vol 13 no 10 pp 953ndash969 2008
[36] K Johar A Priya and M T T Wong-Riley ldquoRegulation ofNa+K+-ATPase by nuclear respiratory factor 1 implication inthe tight coupling of neuronal activity energy generation andenergy consumptionrdquo The Journal of Biological Chemistry vol287 no 48 pp 40381ndash40390 2012
[37] J Astrup P M Sorensen and H R Sorensen ldquoOxygen andglucose consumption related to Na+-K+ transport in caninebrainrdquo Stroke vol 12 no 6 pp 726ndash730 1981
[38] M Erecinska and I A Silver ldquoATP and brain functionrdquo Journalof Cerebral Blood Flow and Metabolism vol 9 no 1 pp 2ndash191989
[39] MMata D J Fink H Gainer et al ldquoActivity-dependent energymetabolism in rat posterior pituitary primarily reflects sodiumpump activityrdquo Journal of Neurochemistry vol 34 no 1 pp 213ndash215 1980
[40] J Kang H-G Lemaire A Unterbeck et al ldquoThe precursor ofAlzheimerrsquos disease amyloid A4 protein resembles a cell-surfacereceptorrdquo Nature vol 325 no 6106 pp 733ndash736 1987
[41] C L Masters and D J Selkoe ldquoBiochemistry of amyloid 120573-protein and amyloid deposits in Alzheimer diseaserdquoCold SpringHarbour Perspectives in Medicine vol 2 no 6 Article IDa006262 2012
[42] A Lai S S Sisodia and I S Trowbridge ldquoCharacterization ofsorting signals in the beta-amyloid precursor protein cytoplas-mic domainrdquo The Journal of Biological Chemistry vol 270 no8 pp 3565ndash3573 1995
[43] A I Placido C M F Pereira A I Duarte et al ldquoThe role ofendoplasmic reticulum in amyloid precursor protein processingand trafficking implications for Alzheimerrsquos diseaserdquo Biochim-ica et Biophysica ActamdashMolecular Basis of Disease vol 1842 no9 pp 1444ndash1453 2014
Oxidative Medicine and Cellular Longevity 13
[44] C Kaether P Skehel and C G Dotti ldquoAxonal membraneproteins are transported in distinct carriers a two-color videomicroscopy study in cultured hippocampal neuronsrdquoMolecularBiology of the Cell vol 11 no 4 pp 1213ndash1224 2000
[45] A Kamal A Almenar-Queralt J F LeBlanc E A Robertsand L S B Goldstein ldquoKinesin-mediated axonal transportof a membrane compartment containing 120573-secretase andpresenilin-1 requires APPrdquo Nature vol 414 no 6864 pp 643ndash648 2001
[46] R G Perez S Soriano J D Hayes et al ldquoMutagenesis identi-fies new signals for 120573-amyloid precursor protein endocytosisturnover and the generation of secreted fragments includingA12057342rdquo The Journal of Biological Chemistry vol 274 no 27 pp18851ndash18856 1999
[47] D G Cook M S Forman J C Sung et al ldquoAlzheimerrsquos A120573(1ndash42) is generated in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo Nature Medicine vol 3 no 9 pp1021ndash1023 1997
[48] A S C Chyung B D Greenberg D G Cook R W Domsand V M-Y Lee ldquoNovel 120573-secretase cleavage of 120573-amyloidprecursor protein in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo The Journal of Cell Biology vol138 no 3 pp 671ndash680 1997
[49] RW-Y Choy Z Cheng and R Schekman ldquoAmyloid precursorprotein (APP) traffics from the cell surface via endosomesfor amyloid 120573 (A120573) production in the trans-Golgi networkrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 30 pp E2077ndashE2082 2012
[50] S A Small and S Gandy ldquoSorting through the cell biologyof Alzheimerrsquos disease intracellular pathways to pathogenesisrdquoNeuron vol 52 no 1 pp 15ndash31 2006
[51] M Goedert ldquoAlzheimerrsquos and Parkinsonrsquos diseases the prionconcept in relation to assembled A120573 tau and 120572-synucleinrdquoScience vol 349 no 6248 2015
[52] I Benilova E Karran and B De Strooper ldquoThe toxic A120573oligomer and Alzheimerrsquos disease an emperor in need ofclothesrdquo Nature Neuroscience vol 15 no 3 pp 349ndash357 2012
[53] E H Corder A M Saunders W J Strittmatter et al ldquoGenedose of apolipoprotein E type 4 allele and the risk of Alzheimerrsquosdisease in late onset familiesrdquo Science vol 261 no 5123 pp 921ndash923 1993
[54] S Brunholz S Sisodia A Lorenzo C Deyts S Kins and GMorfini ldquoAxonal transport of APP and the spatial regulationof APP cleavage and function in neuronal cellsrdquo ExperimentalBrain Research vol 217 no 3-4 pp 353ndash364 2012
[55] G Thinakaran and E H Koo ldquoAmyloid precursor proteintrafficking processing and functionrdquo The Journal of BiologicalChemistry vol 283 no 44 pp 29615ndash29619 2008
[56] T Saitoh M Sundsmo J-M Roch et al ldquoSecreted formof amyloid beta protein precursor is involved in the growthregulation of fibroblastsrdquo Cell vol 58 no 4 pp 615ndash622 1989
[57] M PMattson B Cheng A R Culwell F S Esch I Lieberburgand R E Rydel ldquoEvidence for excitoprotective and intraneu-ronal calcium-regulating roles for secreted forms of the 120573-amyloid precursor proteinrdquo Neuron vol 10 no 2 pp 243ndash2541993
[58] K J Lee C E H Moussa Y Lee et al ldquoBeta amyloid-independent role of amyloid precursor protein in generationandmaintenance of dendritic spinesrdquoNeuroscience vol 169 no1 pp 344ndash356 2010
[59] H Meziane J-C Dodart C Mathis et al ldquoMemory-enhancingeffects of secreted forms of the 120573-amyloid precursor proteinin normal and amnestic micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no21 pp 12683ndash12688 1998
[60] P R Turner K OrsquoConnor W P Tate and W C AbrahamldquoRoles of amyloid precursor protein and its fragments inregulating neural activity plasticity and memoryrdquo Progress inNeurobiology vol 70 no 1 pp 1ndash32 2003
[61] D Beher L Hesse C L Masters and GMulthaup ldquoRegulationof Amyloid Protein Precursor (APP) binding to collagen andmapping of the binding sites on APP and collagen type Irdquo TheJournal of Biological Chemistry vol 271 no 3 pp 1613ndash16201996
[62] S L Sabo A F Ikin J D Buxbaum and P Greengard ldquoTheamyloid precursor protein and its regulatory protein FE65 ingrowth cones and synapses in vitro and in vivordquo The Journal ofNeuroscience vol 23 no 13 pp 5407ndash5415 2003
[63] D H Small V Nurcombe G Reed et al ldquoA heparin-bindingdomain in the amyloid protein precursor of Alzheimerrsquos diseaseis involved in the regulation of neurite outgrowthrdquoThe Journalof Neuroscience vol 14 no 4 pp 2117ndash2127 1994
[64] T G Williamson S S Mok A Henry et al ldquoSecreted glypicanbinds to the amyloid precursor protein of Alzheimerrsquos disease(APP) and inhibits APP-induced neurite outgrowthrdquoThe Jour-nal of Biological Chemistry vol 271 no 49 pp 31215ndash312211996
[65] T Ondrejcak I Klyubin N-W Hu A E Barry W K Cullenand M J Rowan ldquoAlzheimerrsquos disease amyloid 120573-protein andsynaptic functionrdquo NeuroMolecular Medicine vol 12 no 1 pp13ndash26 2010
[66] D Puzzo L Privitera and A Palmeri ldquoHormetic effect ofamyloid-beta peptide in synaptic plasticity and memoryrdquo Neu-robiology of Aging vol 33 no 7 pp 1484e15ndash1484e24 2012
[67] C M F Pereira ldquoCrosstalk between endoplasmic reticulumstress and protein misfolding in neurodegenerative diseasesrdquoISRN Cell Biology vol 2013 Article ID 256404 22 pages 2013
[68] R-W Shin T C Saido M Maeda and T Kitamoto ldquoNovel 120572-secretase cleavage ofAlzheimerrsquos amyloid120573 precursor protein inthe endoplasmic reticulum of COS7 cellsrdquoNeuroscience Lettersvol 376 no 1 pp 14ndash19 2005
[69] E A Schon and E Area-Gomez ldquoMitochondria-associatedER membranes in Alzheimer diseaserdquo Molecular and CellularNeuroscience vol 55 pp 26ndash36 2013
[70] R Rizzuto P Pinton W Carrington et al ldquoClose contacts withthe endoplasmic reticulum as determinants of mitochondrialCa2+ responsesrdquo Science vol 280 no 5370 pp 1763ndash1766 1998
[71] S E Logue P Cleary S Saveljeva and A Samali ldquoNewdirections in ER-stress induced cell deathrdquoApoptosis vol 18 no5 pp 537ndash546 2013
[72] J P Decuypere KWelkenhuyzen T Luyten et al ldquoIns(145)P3receptor-mediated BI-1 Ca2+ signaling and autophagy induc-tion are interrelatedrdquo Autophagy vol 7 pp 1472ndash1489 2011
[73] K S Vetrivel and G Thinakaran ldquoMembrane rafts in Alz-heimerrsquos disease beta-amyloid productionrdquo Biochimica et Bio-physica Acta vol 1801 no 8 pp 860ndash867 2010
[74] H Urra E Dufey F Lisbona D Rojas-Rivera and C HetzldquoWhen ER stress reaches a dead endrdquo Biochimica et BiophysicaActa (BBA) Molecular Cell Research vol 1833 no 12 pp 3507ndash3517 2013
14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 Oxidative Medicine and Cellular Longevity
synthase (complex V) at both protein and activity level [124]While APP A120573 and hyperphosphorylated tau are potentinhibitors of mitochondrial import of nuclear-encoded pro-teins apparently the pathology of each leads to metabolicharm in different way In freshly isolated mitochondriafrom AD brains the APP inhibited mitochondrial importof cytochrome 119888 oxidase (COX) subunits IV and Vb [31]Dysfunction of COX increases ROS production (incompletereduction of oxygen molecules) reduces energy stores anddisturbs energy metabolism Accordingly in AD patientsdeficiency of COX was found in brains and platelets [119125 126] Similar to APP A120573 was found in mitochondriaof transgenic mice and cellular and human AD models [1719 33 103 127ndash130] At present it is not clear whether theobservedmitochondrial toxicity is due toAPP orA120573 orNFTsaccumulation Anyway some regularity is observed withrespect to most affected components of respiratory chainNADH-ubiquinone oxidoreductase (complex I) activity isreduced to the utmost by hyperphosphorylated tau whereasdecreased activity of cytochrome 119888 oxidase (complex IV)that resulted in mitochondrial dysfunction was observedduring A120573 and APP accumulation [14 15 31 32 34]Concomitantly rise in the activity of antioxidant enzymesmanganese superoxide dismutase (Mn-SOD) and catalase(CAT) was demonstrated in response to elevated levelsof free radicals including superoxide anion radical (O
2
∙minus)hydroperoxyl radical (HO
2
∙) hydroxyl radical (OH∙) andnitric oxide radical (NO∙) [15] Consistent with observationsof chronic respiratory chain dysfunction and mitochondrialoxidative stress there are reports showing their contributionto tau pathology in AD [131] In any case free radicals thatoverride antioxidant defense react with a wide variety oforganic components causing lipid peroxidation to advancedlipid oxidation end products (ALE) cross-linking of proteinsnitrosylation of proteins and mutations in DNA Mitochon-drial circular DNA (mtDNA) of sim16 kbp is devoid of repairsystems meaning the buildup of lesions There are 37 geneslocated in mtDNA with those encoding protein subunits ofcomplex I (7) complex II (1) and complex IV (3) but notcomplex III Interestingly hallmarks of AD (APP A120573 andNFTs)mostly affectmembers of electron transfer chain (com-plexes I and IV) which rely exclusively on mitochondriallypredetermined subunits Maybe it is not simple coincidencebut the effect of inhibited import to mitochondria of nuclear-encoded subunits of complexes I and IV gives explanationfor toxicity of APP or A120573 or NFTs in mitochondria Thereare additional findings in mitochondria affected by AD suchas lower activity of pyruvate dehydrogenase (PDH) andoxoglutarate dehydrogenase (OGDH) [104 132] Inhibitionof OGDH the enzyme of tricarboxylic acid cycle minimizesthe NADH pool and electron number needed for ETCand mitochondrial membrane potential (Δ120595
119898) to create and
maintain proton gradient obligatory for ATP synthesisThereare also lines of evidence for direct inhibitory action of solubleoligomeric A120573 species on ABAD (A120573-binding alcohol dehy-drogenase) and internal membrane cyclophilin D (CypD)resulting in increased mitochondrial membrane permeabil-ity (MPTP) potentiated ROS production synaptic lossdiminished activity of mitochondrial respiration and finally
cell death [129 132ndash134] CypD knockout prevents mito-chondrial and neuronal perturbations and improves mito-chondrial function in Alzheimerrsquos disease mouse model [128135] It has to be emphasized that oxidativenitrosative stressaffects the fusion and fission process ofmitochondria Fusionthat is speedup by small GTPases mitofusins (Mfn1Mfn2)improves efficiency of mitochondrial respiration and ATPproduction Mitochondrial dynamics are severely imbal-anced in AD cases in favour of fission through elevatedexpression of the fission protein DLP1 (dynamin-like protein1) associatedwith nitrosative stress stirred up byA120573 [136 137]Collectively these observations indicate that toxic intracel-lular A120573
42-43 oligomers differ in action from extracellularaggregates found in amyloid plaques of AD brains [138](Figure 3)
12 Proteostasis in Mitochondria
Mitochondrial protein turnover grants the well-organizedreplacement of nonfunctional proteins into operational oneThe first task is attained through proteolytic degradation ofinner membrane and matrix proteins with local mitochon-drial proteases [139] or outer membrane proteins throughubiquitin-proteasome system [140] The second task is metby intramitochondrial protein synthesis but as almost 1500different nuclear-encoded proteins have to be importedto complete mitochondrial proteome a matter of capableimport is fundamental for the function of the organelleIn extreme cases of cellular injuries observed in neurode-generative diseases damaged mitochondria with extendedloss of the electrochemical potential are selectively removedby autophagy known as mitophagy [112] APP and A120573accumulation has also something to do with altered mito-chondrial dynamics as fission takes advantage of fusion withresultant dysfunction of mitochondria and neurons [137]AD neurons demonstrated selective mechanisms of prote-olytic clearance of oxidatively and nitrosatively modifiedproteins in mitochondria Insulin degrading enzyme (IDE)prevents formation of toxic insoluble fibrils from A120573 asit cleaves A120573 prior to aggregation [141ndash143] A novel zinc-metallopeptidase Presequence Protease (PreP) a member ofpitrilysin oligopeptidase family degrades either intramito-chondrially stored A120573
40or A120573
42proteinThis protease seems
to be highly sensitive to oxidative stress as disulfide bridgeformed between two proximal cysteine residues blocks itscatalytic activity [144ndash149] Another mitochondrial serineprotease HtrA2Omi occupies intermembrane space whereit can cleave APP locked up in protein translocase complex[121] Even though it is well established that HtrA2Omiis released to the cytosol to amplify apoptosis through thedegradation of antiapoptotic proteins and caspase activation[150ndash152] it is also implicated in proteolytic deletion ofmalfoldedAPP in ER [151] Fromknockout studies onmice itis obvious that HtrA2Omi plays a significant shielding roleas mice deficient in this protease exhibit neurodegenerativephenotype with weight loss and premature death [153]Taken together understanding the mechanisms of clearanceof the unwanted proteins including APP A120573 and tau isvital in an attempt to get rid of them from mitochondria
Oxidative Medicine and Cellular Longevity 9
VDAC
ANT
A120573
A120573
A120573
A120573
A120573
A120573
A120573A120573
A120573
A120573
A120573 CypD
CypD ProApCyt c Ca2+
IMM
OMM
IMSΔΨ
+
+
+
minus
minus
minus
ABAD
ROS
ROS
TIM
TOM
Proteinimport
NAD
NAD
NAD
NAD
H+
H+
H+
H+
H+
ADP
Mn-SOD
CuZn-SOD
mtDNA
V
Matrix
UCP
O2∙minus
O2∙minus
IV
III
II
I
O2
O2
DLP1Fis1
Mfn1Mfn2
Opa1
mHsp70
NADH
NADH
NADH
TCAcycle
PyruvateAcetyl-CoA
120572KGDH
120572-KG
PDH
Isocitrate
CitrateOAA
Malate
Fumarate
Succinyl-CoASuccinate
GDPGTP
FAD
FAOH
ATP
Figure 3 Amyloid-120573-related mitochondrial impairment Mitochondria were found to be the target for amyloid-120573 (A120573) which interactswith several proteins leading to mitochondrial dysfunction Indeed A120573 was found in the outer mitochondrial membrane (OMM) andinner mitochondrial membrane (IMM) as well as in the matrix The interaction of A120573 with the OMM affects the transport of nuclear-encoded mitochondrial proteins such as subunits of the electron transport chain complex IV into the organelle via the translocase of theouter membrane (TOM) import machinery Moreover A120573 disturbs the activity of several enzymes such as pyruvate dehydrogenase (PDH)and oxoglutarate dehydrogenase (OGDH) decreasing NADH reduction and the electron transport chain enzyme complex IV reducing theamount of hydrogen that is translocated from the matrix to the intermembrane space (IMS) thus impairing the mitochondrial membranepotential (MMP) Taken together these events cause abnormal mitochondrial electron activities leading to decreased complex V activityand so to a drop in ATP levels in addition to increasing reactive oxygen species (ROS) generation Moreover ROS induce peroxidationof several mitochondrial macromolecules such as mitochondrial DNA (mtDNA) and mitochondrial lipids contributing to mitochondrialimpairment in themitochondrial matrixThe complex of A120573 bound to binding alcohol dehydrogenase (ABAD) impairs the binding of NAD+to ABAD changes mitochondrial membrane permeability and reduces activities of respiratory enzymes inducing further ROS productionand leading to mitochondrial failure A120573 binding also activates Fis1 (fission protein) and promotes increased mitochondrial fragmentationthis increased mitochondrial fragmentation produces defective mitochondria that ultimately damage neurons Furthermore A120573 binding tocyclophilin D (CypD) enhances the protein translocation to the inner membrane favouring the opening of the mitochondrial permeabilitytransition pore formed by the adenine nucleotide translocator (ANT) and voltage-dependent anion channels (VDACs) Cyt 119888 cytochrome 119888DLP1 dynamin-like protein 1 PDH pyruvate dehydrogenase ProAp proapoptotic factors SOD superoxide dismutase TCA tricarboxylicacid TIM translocase of the inner membrane Adopted from [14]
Some hopes are associated with the application of at presentindefinite modulators of proteolytic activity HtrA2Omi isfor example activated by PTEN-induced putative kinase 1(PINK1) upon phosphorylation at Ser142 residue [154] Alter-natively accent has to be put on mitophagy of dysfunctionalmitochondria and mitochondriogenesis
13 Aging versus AD
Themost commonneurodegenerative disorder is representedby Alzheimerrsquos disease characterized by declining memoryreduced cognitive capacity and progressive dementia whichare often fatal to elderly individuals above 65 years of age
10 Oxidative Medicine and Cellular Longevity
It is ranked as the fourth leading cause of death in modernsocieties where average life span increased greatly in the lasttwo decades As 95 of AD cases are diagnosed in olderpeople one might think that a causal relationship existsbetween aging and the onset of disease Certainly a numberof similarities between getting old and being affected withAD could be listed Historically the free radical theory ofaging by Harman [155] suggested aging as ldquoside effectrdquo ofreactive oxygen species formed in mitochondrial respiratorychain Apparently free radicals commonly generated byincomplete reduction of oxygen molecule at complexes Iand III of mitochondrial ETC are capable of damagingDNA RNA and proteins They impair energy storage andlead to operational failure of mitochondria with progres-sive decline of cell viability Almost identical conditionsaccompany AD and are an explicit step in pathogenesisof disease As mtDNA is deprived of repair mechanismsROS-induced DNA strand breaks tend to accumulate withage or AD Thus mtDNA is a vulnerable target for ROSbut the reverse the ROS generation due to the mutatedmtDNA is not convincingly confirmed [156] Moreover theevidence that mutated mtDNA accelerates the progress ofaging is also questioned based on the results from studycarried out on transgenic mice model [157] Although someauthors show inconsistency between the free radical theoryand observations cumulative evaluation of the scientificreports points to antioxidant defense systems as importantfactors in protection from premature aging [158] Othermitochondrial components important for their function arehampered with age adenine nucleotide translocase (ANT)nitric oxide synthase (NOS) and carnitine acyltransferase(CT) [159ndash161] Actually NOS activity is elevated in AD asreported from study performed on cellular model of disease[15] We could not find any information about CT activityin AD whereas ANT activity is noticeably inhibited byA120573 or hyperphosphorylated tau and this effect is reversedby mersalyl a reversible alkylating agent of thiol groups[162] Mitochondrial dysfunction observed in transgenicmice models of AD and aging demonstrates higher activ-ity of genes controlling energy metabolism and apoptosisTaken together physiological aging and AD are associatedwith broad-spectrum dysfunction of mitochondria but thefoundations of mitochondrial decline are dissimilar Morediscrepancies between physiological aging and AD werefound with respect to ERS and UPR which play a significantrole in cellular proteostasis Dolichol was selected as an agingmarker because the progressive increase in dolichol levelwas observed in aging brain [163] In contrast ubiquinoneconcentration which is also synthesized from geranylgeranioldiminishes with agingwhereas cholesterol and dolichyl phos-phate concentrations remain unaltered In AD decreasedlevels of dolichol were observed and increased levels ofubiquinone and dolichyl phosphate without any changesin brain cholesterol AD cannot be regarded as a resultof premature aging The drop in dolichol and augmenteddolichyl phosphate concentration points toward disturbedglycosylation in ER of diseased brain while the increase inubiquinone suggests efforts to protect the brain from oxida-tive stress induced by lipid peroxidation [164 165]
14 Targeting ER Stress in AD Therapy
As ERS is a recognized factor in AD drugs that interfere withERS would theoretically have great therapeutic potentialThere are several compounds grouped in classes thatinteract directly with components of the ERS (salubrinalBiP inducer X (BIX) salicylamide analogs flavonoidsguanabenz and STF083010) chemical chaperons (4-phenyl-butyric acid (PBA) tauroursodeoxycholic acid (TUDCA)and trimethylamine oxide (TMAO)) chemicals that inhibitprotein degradation (Eeyarestatin MG132 and Bortezomib)compounds with antioxidant activity (Edaravone diben-zoylmethane derivatives and N-acetyl cysteine (NAC)) anddrugs controlling calcium signaling (dantrolene and carba-zole derivatives) [166] They may act by inducing transienttranslation arrest upregulation of chaperone proteins andaugmented degradation of ER-associated misfolded proteinsFundamental approach in the development of new therapyis the selection of appropriate molecular targets In ER stresssignaling the aim is to alter the expression of ER stress-associated molecules that can rescue cells from the toxiceffect of ERS Recent efforts in establishing new promisingdrugs against AD are pointing to chemical chaperones suchas PBA TUDCA or trimethylamine oxide (TMAO) Thesesubstances improve protein folding and alleviate nativeprotein conformation [166] It was shown on mouse modelsof AD that PBA TUDCA and TMAO stop A120573 accumulationand avoid the loss of dendritic spines [167] Some observa-tions even demonstrated improved memory and cognitivefunctions [168] associated with improved cell survival [169]Salubrinal ((2E)-3-phenyl-N-[222-trichloro-1-[[(8-quinoli-nylamino)thioxomethyl]amino]ethyl]-2-propenamide Sal)selectively inhibits growth arrest and DNA damage inducedgene 34- (GADD34-) phosphatase complex (GADD34associates with protein phosphatase 1 (PP1)) and promotes invitro dephosphorylation of the alpha subunit of eIF-2120572 andIRE1120572ASK1JNK signaling pathway being protective againstERS even induced by tunicamycin Tm [170] In a great deal ofexperiments testing Sal in cultured cells and animal modelsof AD this substance increased the viability of neuronal cellsand A120573 toxicity [171 172] BIX (2-(34-dihydroxyphenyl)-2-oxoethyl ester thiocyanic acid) preferentially inducedBiP mRNA in an ATF6-dependent manner leading toreduced Tm-induced death of neuronal cells [173] AlsoDBM derivative 14ndash26 (221015840-dimethoxydibenzoylmethane)was found to be neuroprotective for SH-SY5Y and PC-12cells by decreasing expression of BiP and CHOP [174]Dantrolene a ryanodine receptor antagonist that inhibitsabnormal calcium release from the ER inhibited expressionof both phosphorylated PERK and eIF2120572 It also reducedCHOP expression and attenuated thapsigargin-inducedapoptosis in PC-12 cells [175] Neuroprotective effects similarto dantrolene were observed for ([9-(3-cyanobenzyl)-14-dimethylcarbazole]) This substance suppressed increases inintracellular Ca2+ in PC-12 cells treated with thapsigarginand reduced levels of BiP and CHOP [176]
Aforementioned compounds were chosen from othersas the most potent ER stress inhibitors and persuasivelyprotective to neuronal cells 18 of 42 different compounds
Oxidative Medicine and Cellular Longevity 11
were exploited in in vivo and in vitro models of centralnervous system disorders with in fact improved cell or tissueviability [166] Thus the brain is the most frequently investi-gated organ in the context of ERS From these experimentsit becomes clear that CHOP functions as proapoptotic factorThe roles of other specific ER stress molecules as moleculartargets for pharmacological intervention are less clear andvary depending on cell type and context
There are few underestimated modulations in APP pro-cessing that shed more light on current dogma of AD patho-genesis The modulation of mevalonate pathway and choles-terol synthesis were reported to stimulate nonamyloidogenicpathway of APP processing [177] Additionally cholesterolderivative 27-hydroxycholesterol (27-OHC) was shown toinduce ER stress which attenuated leptin-dependent viabilityby activating CHOP in SH-SY5Y neuroblastoma cells [178]Irrespective of a number of compounds examined in AD thecall for new drugs modulating ER stress with healing effect isstill waiting to be revealed
Abbreviations
APOE Apolipoprotein EAPPs Secreted form of APPCHOPGADD153 CEBP homologous protein
growth arrest and DNA damageinduced gene 153
Drp1 Dynamin-1-like proteinERAD Endoplasmic-reticulum-
associated protein degradationERO1120572 ER oxidase 1120572ERS Endoplasmic reticulum stressFOH FarnesolGGOH GeranylgeraniolGRP75 Glucose-regulated protein 75GRP78BiP 78 kDa glucose-regulated proteinGSK-3120573 Synthase kinase 3 betaIP3R Inositol triphosphate receptor
IRE1120572 Inositol-requiring enzyme 1 alphaMAM Mitochondria-associated
membraneMAP LC3 Microtubule-associated protein
light chain 3Mfn2 GTPase mitofusin 2MMP Mitochondrial membrane
potentialMPTP Mitochondrial permeability
transition poreNFT Neurofibrillary tanglesPACS-2 Phosphofurin acidic cluster
sorting protein 2PDI Protein disulfide isomerasePERK Protein kinase R- (PKR-) like
ER kinaseRNS Reactive nitrogen speciesROS Reactive oxygen speciesRyR Ryanodine receptorTOM Transporter outer membraneTIM Transporter inner membrane
UPR Unfolded protein responseVDAC Voltage-dependent anion channelXBP-1 Xbox binding protein 1
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
Support for this work was provided by Grant no UMO-201103BNZ70238 from the National Science Centre inPoland
References
[1] R L Nussbaum andC E Ellis ldquoAlzheimerrsquos disease and Parkin-sonrsquos diseaserdquoTheNewEngland Journal ofMedicine vol 348 no14 pp 1356ndash1364 2003
[2] A L Brunkan and A M Goate ldquoPresenilin function and 120574-secretase activityrdquoThe Journal of Neurochemistry vol 93 no 4pp 769ndash792 2005
[3] V Crentsil ldquoThe pharmacogenomics of Alzheimerrsquos diseaserdquoAgeing Research Reviews vol 3 no 2 pp 153ndash169 2004
[4] D L Price and S S Sisodia ldquoMutant genes in familialAlzheimerrsquos disease and transgenic modelsrdquo Annual Review ofNeuroscience vol 21 pp 479ndash505 1998
[5] D Harman ldquoAlzheimerrsquos disease pathogenesis role of agingrdquoAnnals of the New York Academy of Sciences vol 1067 no 1 pp454ndash460 2006
[6] D J Selkoe ldquoTranslating cell biology into therapeutic advancesin Alzheimerrsquos diseaserdquo Nature vol 399 pp A23ndashA31 1999
[7] H Braak and E Braak ldquoNeuropathological stageing of Alz-heimer-related changesrdquo Acta Neuropathologica vol 82 no 4pp 239ndash259 1991
[8] K Hirai G Aliev A Nunomara et al ldquoMitochondrial abnor-malities inAlzheimerrsquos diseaserdquoTheJournal ofNeuroscience vol21 pp 3017ndash3023 2001
[9] U T Brunk C B Jones and R S Sohal ldquoA novel hypothesisof lipofuscinogenesis and cellular aging based on interac-tions between oxidative stress and autophagocytosisrdquoMutationResearchDNAging vol 275 no 3ndash6 pp 395ndash403 1992
[10] P A Stewart K Hayakawa M-A Akers and H V Vinters ldquoAmorphometric study of the blood-brain barrier in Alzheimerrsquosdiseaserdquo Laboratory Investigation vol 67 no 6 pp 734ndash7421992
[11] J P Blass and G E Gibson ldquoThe role of oxidative abnor-malities in the pathophysiology of Alzheimerrsquos diseaserdquo RevueNeurologique vol 147 no 6-7 pp 513ndash525 1991
[12] B Pająk E Kania and A Orzechowski ldquoNucleofection of ratpheochromocytoma PC-12 cells with human mutated beta-amyloid precursor protein gene (APP-sw) leads to reducedreduced viability autophagy-like process and increased expres-sion and secretion of beta amyloidrdquo BioMed Research Interna-tional vol 2015 Article ID 746092 10 pages 2015
[13] J P Blass ldquoCerebrometabolic abnormalities in AlzheimerrsquosdiseaserdquoNeurological Research vol 25 no 6 pp 556ndash566 2003
12 Oxidative Medicine and Cellular Longevity
[14] A Eckert K Schmitt and J Gotz ldquoMitochondrial dysfunc-tionmdashthe beginning of the end in Alzheimerrsquos disease Sep-arate and synergistic modes of tau and amyloid-120573 toxicityrdquoAlzheimerrsquos Research ampTherapy vol 3 article 15 pp 1ndash11 2011
[15] U Keil A Bonert C A Marques et al ldquoAmyloid 120573-inducedchanges in nitric oxide production and mitochondrial activitylead to apoptosisrdquoThe Journal of Biological Chemistry vol 279no 48 pp 50310ndash50320 2004
[16] A M Swomley and D A Butterfield ldquoOxidative stress inAlzheimer disease and mild cognitive impairment evidencefrom human data provided by redox proteomicsrdquo Archives ofToxicology vol 89 no 10 pp 1669ndash1680 2015
[17] C Caspersen N Wang J Yao et al ldquoMitochondrial A120573 apotential focal point for neuronal metabolic dysfunction inAlzheimerrsquos diseaserdquo The FASEB Journal vol 19 no 14 pp2040ndash2041 2005
[18] G E Gibson K-F R Sheu and J P Blass ldquoAbnormalities ofmitochondrial enzymes inAlzheimer diseaserdquo Journal of NeuralTransmission vol 105 no 8-9 pp 855ndash870 1998
[19] A Eckert S Hauptmann I Scherping et al ldquoSoluble beta-amyloid leads to mitochondrial defects in amyloid precursorprotein and tau transgenic micerdquo Neurodegenerative Diseasesvol 5 no 3-4 pp 157ndash159 2008
[20] A Eckert K L Schulz V Rhein and J Gotz ldquoConvergenceof amyloid-120573 and tau pathologies on mitochondria in vivordquoMolecular Neurobiology vol 41 no 2-3 pp 107ndash114 2010
[21] S Hauptmann I Scherping S Drose et al ldquoMitochondrial dys-function an early event in Alzheimer pathology accumulateswith age in AD transgenic micerdquoNeurobiology of Aging vol 30no 10 pp 1574ndash1586 2009
[22] M Manczak B S Park Y Jung and P H Reddy ldquoDifferentialexpression of oxidative phosphorylation genes in patients withAlzheimerrsquos disease implications for early mitochondrial dys-function and oxidative damagerdquo NeuroMolecular Medicine vol5 no 2 pp 147ndash162 2004
[23] M P Mattson M Gleichmann and A Cheng ldquoMitochondriain neuroplasticity and neurological disordersrdquo Neuron vol 60no 5 pp 748ndash766 2008
[24] P I Moreira M S Santos and C R Oliveira ldquoAlzheimerrsquosdisease a lesson frommitochondrial dysfunctionrdquoAntioxidantsamp Redox Signaling vol 9 no 10 pp 1621ndash1630 2007
[25] P H Reddy ldquoMitochondrial dysfunction in aging and Alz-heimerrsquos disease strategies to protect neuronsrdquo Antioxidants ampRedox Signaling vol 9 no 10 pp 1647ndash1658 2007
[26] V Rhein G Baysang S Rao et al ldquoAmyloid-beta leads toimpaired cellular respiration energy production andmitochon-drial electron chain complex activities in humanneuroblastomacellsrdquo Cellular and Molecular Neurobiology vol 29 no 6-7 pp1063ndash1071 2009
[27] V Rhein X Song A Wiesner et al ldquoAmyloid-120573 and tausynergistically impair the oxidative phosphorylation system intriple transgenic Alzheimerrsquos disease micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 47 pp 20057ndash20062 2009
[28] R X Santos S C Correia X Wang et al ldquoAlzheimerrsquos diseasediverse aspects ofmitochondrial malfunctioningrdquo InternationalJournal of Clinical and Experimental Pathology vol 3 no 6 pp570ndash581 2010
[29] N R Sims ldquoEnergy metabolism oxidative stress and neuronaldegeneration in Alzheimerrsquos diseaserdquoNeurodegeneration vol 5no 4 pp 435ndash440 1996
[30] H K Anandatheerthavarada G Biswas M-A Robin and NG Avadhani ldquoMitochondrial targeting and a novel transmem-brane arrest of Alzheimerrsquos amyloid precursor protein impairsmitochondrial function in neuronal cellsrdquo The Journal of CellBiology vol 161 no 1 pp 41ndash54 2003
[31] L Devi B M Prabhu D F Galati N G Avadhani and HK Anandatheerthavarada ldquoAccumulation of amyloid precur-sor protein in the mitochondrial import channels of humanAlzheimerrsquos disease brain is associated with mitochondrialdysfunctionrdquo The Journal of Neuroscience vol 26 no 35 pp9057ndash9068 2006
[32] L Devi and H K Anandatheerthavarada ldquoMitochondrialtrafficking of APP and alpha synuclein relevance to mito-chondrial dysfunction in Alzheimerrsquos and Parkinsonrsquos diseasesrdquoBiochimica et Biophysica Acta vol 1802 no 1 pp 11ndash19 2010
[33] M Manczak T S Anekonda E Henson B S Park J Quinnand P H Reddy ldquoMitochondria are a direct site of A120573 accumu-lation in Alzheimerrsquos disease neurons implications for free rad-ical generation and oxidative damage in disease progressionrdquoHuman Molecular Genetics vol 15 no 9 pp 1437ndash1449 2006
[34] P F Pavlov C H Petersen E Glaser and M AnkarcronaldquoMitochondrial accumulation of APP and A120573 significancefor Alzheimer disease pathogenesisrdquo Journal of Cellular andMolecular Medicine vol 13 no 10 pp 4137ndash4145 2009
[35] C Schmidt E Lepsverdize S L Chi et al ldquoAmyloid precursorprotein and amyloid 120573-peptide bind to ATP synthase andregulate its activity at the surface of neural cellsrdquo MolecularPsychiatry vol 13 no 10 pp 953ndash969 2008
[36] K Johar A Priya and M T T Wong-Riley ldquoRegulation ofNa+K+-ATPase by nuclear respiratory factor 1 implication inthe tight coupling of neuronal activity energy generation andenergy consumptionrdquo The Journal of Biological Chemistry vol287 no 48 pp 40381ndash40390 2012
[37] J Astrup P M Sorensen and H R Sorensen ldquoOxygen andglucose consumption related to Na+-K+ transport in caninebrainrdquo Stroke vol 12 no 6 pp 726ndash730 1981
[38] M Erecinska and I A Silver ldquoATP and brain functionrdquo Journalof Cerebral Blood Flow and Metabolism vol 9 no 1 pp 2ndash191989
[39] MMata D J Fink H Gainer et al ldquoActivity-dependent energymetabolism in rat posterior pituitary primarily reflects sodiumpump activityrdquo Journal of Neurochemistry vol 34 no 1 pp 213ndash215 1980
[40] J Kang H-G Lemaire A Unterbeck et al ldquoThe precursor ofAlzheimerrsquos disease amyloid A4 protein resembles a cell-surfacereceptorrdquo Nature vol 325 no 6106 pp 733ndash736 1987
[41] C L Masters and D J Selkoe ldquoBiochemistry of amyloid 120573-protein and amyloid deposits in Alzheimer diseaserdquoCold SpringHarbour Perspectives in Medicine vol 2 no 6 Article IDa006262 2012
[42] A Lai S S Sisodia and I S Trowbridge ldquoCharacterization ofsorting signals in the beta-amyloid precursor protein cytoplas-mic domainrdquo The Journal of Biological Chemistry vol 270 no8 pp 3565ndash3573 1995
[43] A I Placido C M F Pereira A I Duarte et al ldquoThe role ofendoplasmic reticulum in amyloid precursor protein processingand trafficking implications for Alzheimerrsquos diseaserdquo Biochim-ica et Biophysica ActamdashMolecular Basis of Disease vol 1842 no9 pp 1444ndash1453 2014
Oxidative Medicine and Cellular Longevity 13
[44] C Kaether P Skehel and C G Dotti ldquoAxonal membraneproteins are transported in distinct carriers a two-color videomicroscopy study in cultured hippocampal neuronsrdquoMolecularBiology of the Cell vol 11 no 4 pp 1213ndash1224 2000
[45] A Kamal A Almenar-Queralt J F LeBlanc E A Robertsand L S B Goldstein ldquoKinesin-mediated axonal transportof a membrane compartment containing 120573-secretase andpresenilin-1 requires APPrdquo Nature vol 414 no 6864 pp 643ndash648 2001
[46] R G Perez S Soriano J D Hayes et al ldquoMutagenesis identi-fies new signals for 120573-amyloid precursor protein endocytosisturnover and the generation of secreted fragments includingA12057342rdquo The Journal of Biological Chemistry vol 274 no 27 pp18851ndash18856 1999
[47] D G Cook M S Forman J C Sung et al ldquoAlzheimerrsquos A120573(1ndash42) is generated in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo Nature Medicine vol 3 no 9 pp1021ndash1023 1997
[48] A S C Chyung B D Greenberg D G Cook R W Domsand V M-Y Lee ldquoNovel 120573-secretase cleavage of 120573-amyloidprecursor protein in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo The Journal of Cell Biology vol138 no 3 pp 671ndash680 1997
[49] RW-Y Choy Z Cheng and R Schekman ldquoAmyloid precursorprotein (APP) traffics from the cell surface via endosomesfor amyloid 120573 (A120573) production in the trans-Golgi networkrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 30 pp E2077ndashE2082 2012
[50] S A Small and S Gandy ldquoSorting through the cell biologyof Alzheimerrsquos disease intracellular pathways to pathogenesisrdquoNeuron vol 52 no 1 pp 15ndash31 2006
[51] M Goedert ldquoAlzheimerrsquos and Parkinsonrsquos diseases the prionconcept in relation to assembled A120573 tau and 120572-synucleinrdquoScience vol 349 no 6248 2015
[52] I Benilova E Karran and B De Strooper ldquoThe toxic A120573oligomer and Alzheimerrsquos disease an emperor in need ofclothesrdquo Nature Neuroscience vol 15 no 3 pp 349ndash357 2012
[53] E H Corder A M Saunders W J Strittmatter et al ldquoGenedose of apolipoprotein E type 4 allele and the risk of Alzheimerrsquosdisease in late onset familiesrdquo Science vol 261 no 5123 pp 921ndash923 1993
[54] S Brunholz S Sisodia A Lorenzo C Deyts S Kins and GMorfini ldquoAxonal transport of APP and the spatial regulationof APP cleavage and function in neuronal cellsrdquo ExperimentalBrain Research vol 217 no 3-4 pp 353ndash364 2012
[55] G Thinakaran and E H Koo ldquoAmyloid precursor proteintrafficking processing and functionrdquo The Journal of BiologicalChemistry vol 283 no 44 pp 29615ndash29619 2008
[56] T Saitoh M Sundsmo J-M Roch et al ldquoSecreted formof amyloid beta protein precursor is involved in the growthregulation of fibroblastsrdquo Cell vol 58 no 4 pp 615ndash622 1989
[57] M PMattson B Cheng A R Culwell F S Esch I Lieberburgand R E Rydel ldquoEvidence for excitoprotective and intraneu-ronal calcium-regulating roles for secreted forms of the 120573-amyloid precursor proteinrdquo Neuron vol 10 no 2 pp 243ndash2541993
[58] K J Lee C E H Moussa Y Lee et al ldquoBeta amyloid-independent role of amyloid precursor protein in generationandmaintenance of dendritic spinesrdquoNeuroscience vol 169 no1 pp 344ndash356 2010
[59] H Meziane J-C Dodart C Mathis et al ldquoMemory-enhancingeffects of secreted forms of the 120573-amyloid precursor proteinin normal and amnestic micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no21 pp 12683ndash12688 1998
[60] P R Turner K OrsquoConnor W P Tate and W C AbrahamldquoRoles of amyloid precursor protein and its fragments inregulating neural activity plasticity and memoryrdquo Progress inNeurobiology vol 70 no 1 pp 1ndash32 2003
[61] D Beher L Hesse C L Masters and GMulthaup ldquoRegulationof Amyloid Protein Precursor (APP) binding to collagen andmapping of the binding sites on APP and collagen type Irdquo TheJournal of Biological Chemistry vol 271 no 3 pp 1613ndash16201996
[62] S L Sabo A F Ikin J D Buxbaum and P Greengard ldquoTheamyloid precursor protein and its regulatory protein FE65 ingrowth cones and synapses in vitro and in vivordquo The Journal ofNeuroscience vol 23 no 13 pp 5407ndash5415 2003
[63] D H Small V Nurcombe G Reed et al ldquoA heparin-bindingdomain in the amyloid protein precursor of Alzheimerrsquos diseaseis involved in the regulation of neurite outgrowthrdquoThe Journalof Neuroscience vol 14 no 4 pp 2117ndash2127 1994
[64] T G Williamson S S Mok A Henry et al ldquoSecreted glypicanbinds to the amyloid precursor protein of Alzheimerrsquos disease(APP) and inhibits APP-induced neurite outgrowthrdquoThe Jour-nal of Biological Chemistry vol 271 no 49 pp 31215ndash312211996
[65] T Ondrejcak I Klyubin N-W Hu A E Barry W K Cullenand M J Rowan ldquoAlzheimerrsquos disease amyloid 120573-protein andsynaptic functionrdquo NeuroMolecular Medicine vol 12 no 1 pp13ndash26 2010
[66] D Puzzo L Privitera and A Palmeri ldquoHormetic effect ofamyloid-beta peptide in synaptic plasticity and memoryrdquo Neu-robiology of Aging vol 33 no 7 pp 1484e15ndash1484e24 2012
[67] C M F Pereira ldquoCrosstalk between endoplasmic reticulumstress and protein misfolding in neurodegenerative diseasesrdquoISRN Cell Biology vol 2013 Article ID 256404 22 pages 2013
[68] R-W Shin T C Saido M Maeda and T Kitamoto ldquoNovel 120572-secretase cleavage ofAlzheimerrsquos amyloid120573 precursor protein inthe endoplasmic reticulum of COS7 cellsrdquoNeuroscience Lettersvol 376 no 1 pp 14ndash19 2005
[69] E A Schon and E Area-Gomez ldquoMitochondria-associatedER membranes in Alzheimer diseaserdquo Molecular and CellularNeuroscience vol 55 pp 26ndash36 2013
[70] R Rizzuto P Pinton W Carrington et al ldquoClose contacts withthe endoplasmic reticulum as determinants of mitochondrialCa2+ responsesrdquo Science vol 280 no 5370 pp 1763ndash1766 1998
[71] S E Logue P Cleary S Saveljeva and A Samali ldquoNewdirections in ER-stress induced cell deathrdquoApoptosis vol 18 no5 pp 537ndash546 2013
[72] J P Decuypere KWelkenhuyzen T Luyten et al ldquoIns(145)P3receptor-mediated BI-1 Ca2+ signaling and autophagy induc-tion are interrelatedrdquo Autophagy vol 7 pp 1472ndash1489 2011
[73] K S Vetrivel and G Thinakaran ldquoMembrane rafts in Alz-heimerrsquos disease beta-amyloid productionrdquo Biochimica et Bio-physica Acta vol 1801 no 8 pp 860ndash867 2010
[74] H Urra E Dufey F Lisbona D Rojas-Rivera and C HetzldquoWhen ER stress reaches a dead endrdquo Biochimica et BiophysicaActa (BBA) Molecular Cell Research vol 1833 no 12 pp 3507ndash3517 2013
14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
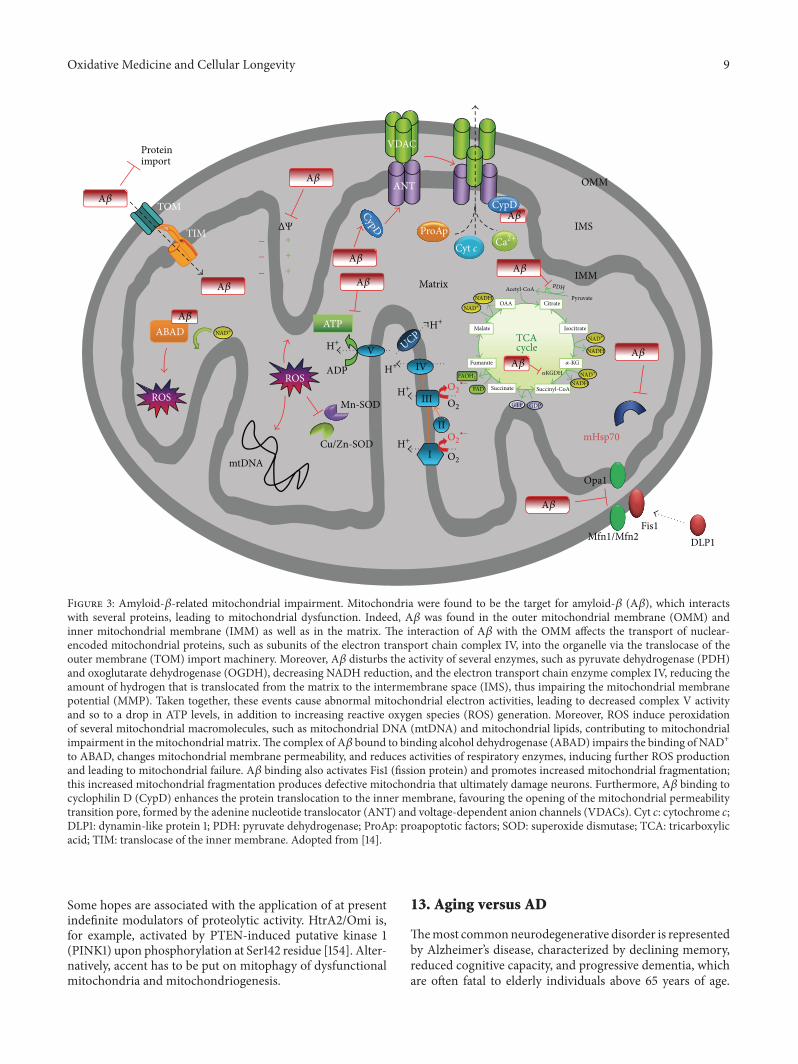
Oxidative Medicine and Cellular Longevity 9
VDAC
ANT
A120573
A120573
A120573
A120573
A120573
A120573
A120573A120573
A120573
A120573
A120573 CypD
CypD ProApCyt c Ca2+
IMM
OMM
IMSΔΨ
+
+
+
minus
minus
minus
ABAD
ROS
ROS
TIM
TOM
Proteinimport
NAD
NAD
NAD
NAD
H+
H+
H+
H+
H+
ADP
Mn-SOD
CuZn-SOD
mtDNA
V
Matrix
UCP
O2∙minus
O2∙minus
IV
III
II
I
O2
O2
DLP1Fis1
Mfn1Mfn2
Opa1
mHsp70
NADH
NADH
NADH
TCAcycle
PyruvateAcetyl-CoA
120572KGDH
120572-KG
PDH
Isocitrate
CitrateOAA
Malate
Fumarate
Succinyl-CoASuccinate
GDPGTP
FAD
FAOH
ATP
Figure 3 Amyloid-120573-related mitochondrial impairment Mitochondria were found to be the target for amyloid-120573 (A120573) which interactswith several proteins leading to mitochondrial dysfunction Indeed A120573 was found in the outer mitochondrial membrane (OMM) andinner mitochondrial membrane (IMM) as well as in the matrix The interaction of A120573 with the OMM affects the transport of nuclear-encoded mitochondrial proteins such as subunits of the electron transport chain complex IV into the organelle via the translocase of theouter membrane (TOM) import machinery Moreover A120573 disturbs the activity of several enzymes such as pyruvate dehydrogenase (PDH)and oxoglutarate dehydrogenase (OGDH) decreasing NADH reduction and the electron transport chain enzyme complex IV reducing theamount of hydrogen that is translocated from the matrix to the intermembrane space (IMS) thus impairing the mitochondrial membranepotential (MMP) Taken together these events cause abnormal mitochondrial electron activities leading to decreased complex V activityand so to a drop in ATP levels in addition to increasing reactive oxygen species (ROS) generation Moreover ROS induce peroxidationof several mitochondrial macromolecules such as mitochondrial DNA (mtDNA) and mitochondrial lipids contributing to mitochondrialimpairment in themitochondrial matrixThe complex of A120573 bound to binding alcohol dehydrogenase (ABAD) impairs the binding of NAD+to ABAD changes mitochondrial membrane permeability and reduces activities of respiratory enzymes inducing further ROS productionand leading to mitochondrial failure A120573 binding also activates Fis1 (fission protein) and promotes increased mitochondrial fragmentationthis increased mitochondrial fragmentation produces defective mitochondria that ultimately damage neurons Furthermore A120573 binding tocyclophilin D (CypD) enhances the protein translocation to the inner membrane favouring the opening of the mitochondrial permeabilitytransition pore formed by the adenine nucleotide translocator (ANT) and voltage-dependent anion channels (VDACs) Cyt 119888 cytochrome 119888DLP1 dynamin-like protein 1 PDH pyruvate dehydrogenase ProAp proapoptotic factors SOD superoxide dismutase TCA tricarboxylicacid TIM translocase of the inner membrane Adopted from [14]
Some hopes are associated with the application of at presentindefinite modulators of proteolytic activity HtrA2Omi isfor example activated by PTEN-induced putative kinase 1(PINK1) upon phosphorylation at Ser142 residue [154] Alter-natively accent has to be put on mitophagy of dysfunctionalmitochondria and mitochondriogenesis
13 Aging versus AD
Themost commonneurodegenerative disorder is representedby Alzheimerrsquos disease characterized by declining memoryreduced cognitive capacity and progressive dementia whichare often fatal to elderly individuals above 65 years of age
10 Oxidative Medicine and Cellular Longevity
It is ranked as the fourth leading cause of death in modernsocieties where average life span increased greatly in the lasttwo decades As 95 of AD cases are diagnosed in olderpeople one might think that a causal relationship existsbetween aging and the onset of disease Certainly a numberof similarities between getting old and being affected withAD could be listed Historically the free radical theory ofaging by Harman [155] suggested aging as ldquoside effectrdquo ofreactive oxygen species formed in mitochondrial respiratorychain Apparently free radicals commonly generated byincomplete reduction of oxygen molecule at complexes Iand III of mitochondrial ETC are capable of damagingDNA RNA and proteins They impair energy storage andlead to operational failure of mitochondria with progres-sive decline of cell viability Almost identical conditionsaccompany AD and are an explicit step in pathogenesisof disease As mtDNA is deprived of repair mechanismsROS-induced DNA strand breaks tend to accumulate withage or AD Thus mtDNA is a vulnerable target for ROSbut the reverse the ROS generation due to the mutatedmtDNA is not convincingly confirmed [156] Moreover theevidence that mutated mtDNA accelerates the progress ofaging is also questioned based on the results from studycarried out on transgenic mice model [157] Although someauthors show inconsistency between the free radical theoryand observations cumulative evaluation of the scientificreports points to antioxidant defense systems as importantfactors in protection from premature aging [158] Othermitochondrial components important for their function arehampered with age adenine nucleotide translocase (ANT)nitric oxide synthase (NOS) and carnitine acyltransferase(CT) [159ndash161] Actually NOS activity is elevated in AD asreported from study performed on cellular model of disease[15] We could not find any information about CT activityin AD whereas ANT activity is noticeably inhibited byA120573 or hyperphosphorylated tau and this effect is reversedby mersalyl a reversible alkylating agent of thiol groups[162] Mitochondrial dysfunction observed in transgenicmice models of AD and aging demonstrates higher activ-ity of genes controlling energy metabolism and apoptosisTaken together physiological aging and AD are associatedwith broad-spectrum dysfunction of mitochondria but thefoundations of mitochondrial decline are dissimilar Morediscrepancies between physiological aging and AD werefound with respect to ERS and UPR which play a significantrole in cellular proteostasis Dolichol was selected as an agingmarker because the progressive increase in dolichol levelwas observed in aging brain [163] In contrast ubiquinoneconcentration which is also synthesized from geranylgeranioldiminishes with agingwhereas cholesterol and dolichyl phos-phate concentrations remain unaltered In AD decreasedlevels of dolichol were observed and increased levels ofubiquinone and dolichyl phosphate without any changesin brain cholesterol AD cannot be regarded as a resultof premature aging The drop in dolichol and augmenteddolichyl phosphate concentration points toward disturbedglycosylation in ER of diseased brain while the increase inubiquinone suggests efforts to protect the brain from oxida-tive stress induced by lipid peroxidation [164 165]
14 Targeting ER Stress in AD Therapy
As ERS is a recognized factor in AD drugs that interfere withERS would theoretically have great therapeutic potentialThere are several compounds grouped in classes thatinteract directly with components of the ERS (salubrinalBiP inducer X (BIX) salicylamide analogs flavonoidsguanabenz and STF083010) chemical chaperons (4-phenyl-butyric acid (PBA) tauroursodeoxycholic acid (TUDCA)and trimethylamine oxide (TMAO)) chemicals that inhibitprotein degradation (Eeyarestatin MG132 and Bortezomib)compounds with antioxidant activity (Edaravone diben-zoylmethane derivatives and N-acetyl cysteine (NAC)) anddrugs controlling calcium signaling (dantrolene and carba-zole derivatives) [166] They may act by inducing transienttranslation arrest upregulation of chaperone proteins andaugmented degradation of ER-associated misfolded proteinsFundamental approach in the development of new therapyis the selection of appropriate molecular targets In ER stresssignaling the aim is to alter the expression of ER stress-associated molecules that can rescue cells from the toxiceffect of ERS Recent efforts in establishing new promisingdrugs against AD are pointing to chemical chaperones suchas PBA TUDCA or trimethylamine oxide (TMAO) Thesesubstances improve protein folding and alleviate nativeprotein conformation [166] It was shown on mouse modelsof AD that PBA TUDCA and TMAO stop A120573 accumulationand avoid the loss of dendritic spines [167] Some observa-tions even demonstrated improved memory and cognitivefunctions [168] associated with improved cell survival [169]Salubrinal ((2E)-3-phenyl-N-[222-trichloro-1-[[(8-quinoli-nylamino)thioxomethyl]amino]ethyl]-2-propenamide Sal)selectively inhibits growth arrest and DNA damage inducedgene 34- (GADD34-) phosphatase complex (GADD34associates with protein phosphatase 1 (PP1)) and promotes invitro dephosphorylation of the alpha subunit of eIF-2120572 andIRE1120572ASK1JNK signaling pathway being protective againstERS even induced by tunicamycin Tm [170] In a great deal ofexperiments testing Sal in cultured cells and animal modelsof AD this substance increased the viability of neuronal cellsand A120573 toxicity [171 172] BIX (2-(34-dihydroxyphenyl)-2-oxoethyl ester thiocyanic acid) preferentially inducedBiP mRNA in an ATF6-dependent manner leading toreduced Tm-induced death of neuronal cells [173] AlsoDBM derivative 14ndash26 (221015840-dimethoxydibenzoylmethane)was found to be neuroprotective for SH-SY5Y and PC-12cells by decreasing expression of BiP and CHOP [174]Dantrolene a ryanodine receptor antagonist that inhibitsabnormal calcium release from the ER inhibited expressionof both phosphorylated PERK and eIF2120572 It also reducedCHOP expression and attenuated thapsigargin-inducedapoptosis in PC-12 cells [175] Neuroprotective effects similarto dantrolene were observed for ([9-(3-cyanobenzyl)-14-dimethylcarbazole]) This substance suppressed increases inintracellular Ca2+ in PC-12 cells treated with thapsigarginand reduced levels of BiP and CHOP [176]
Aforementioned compounds were chosen from othersas the most potent ER stress inhibitors and persuasivelyprotective to neuronal cells 18 of 42 different compounds
Oxidative Medicine and Cellular Longevity 11
were exploited in in vivo and in vitro models of centralnervous system disorders with in fact improved cell or tissueviability [166] Thus the brain is the most frequently investi-gated organ in the context of ERS From these experimentsit becomes clear that CHOP functions as proapoptotic factorThe roles of other specific ER stress molecules as moleculartargets for pharmacological intervention are less clear andvary depending on cell type and context
There are few underestimated modulations in APP pro-cessing that shed more light on current dogma of AD patho-genesis The modulation of mevalonate pathway and choles-terol synthesis were reported to stimulate nonamyloidogenicpathway of APP processing [177] Additionally cholesterolderivative 27-hydroxycholesterol (27-OHC) was shown toinduce ER stress which attenuated leptin-dependent viabilityby activating CHOP in SH-SY5Y neuroblastoma cells [178]Irrespective of a number of compounds examined in AD thecall for new drugs modulating ER stress with healing effect isstill waiting to be revealed
Abbreviations
APOE Apolipoprotein EAPPs Secreted form of APPCHOPGADD153 CEBP homologous protein
growth arrest and DNA damageinduced gene 153
Drp1 Dynamin-1-like proteinERAD Endoplasmic-reticulum-
associated protein degradationERO1120572 ER oxidase 1120572ERS Endoplasmic reticulum stressFOH FarnesolGGOH GeranylgeraniolGRP75 Glucose-regulated protein 75GRP78BiP 78 kDa glucose-regulated proteinGSK-3120573 Synthase kinase 3 betaIP3R Inositol triphosphate receptor
IRE1120572 Inositol-requiring enzyme 1 alphaMAM Mitochondria-associated
membraneMAP LC3 Microtubule-associated protein
light chain 3Mfn2 GTPase mitofusin 2MMP Mitochondrial membrane
potentialMPTP Mitochondrial permeability
transition poreNFT Neurofibrillary tanglesPACS-2 Phosphofurin acidic cluster
sorting protein 2PDI Protein disulfide isomerasePERK Protein kinase R- (PKR-) like
ER kinaseRNS Reactive nitrogen speciesROS Reactive oxygen speciesRyR Ryanodine receptorTOM Transporter outer membraneTIM Transporter inner membrane
UPR Unfolded protein responseVDAC Voltage-dependent anion channelXBP-1 Xbox binding protein 1
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
Support for this work was provided by Grant no UMO-201103BNZ70238 from the National Science Centre inPoland
References
[1] R L Nussbaum andC E Ellis ldquoAlzheimerrsquos disease and Parkin-sonrsquos diseaserdquoTheNewEngland Journal ofMedicine vol 348 no14 pp 1356ndash1364 2003
[2] A L Brunkan and A M Goate ldquoPresenilin function and 120574-secretase activityrdquoThe Journal of Neurochemistry vol 93 no 4pp 769ndash792 2005
[3] V Crentsil ldquoThe pharmacogenomics of Alzheimerrsquos diseaserdquoAgeing Research Reviews vol 3 no 2 pp 153ndash169 2004
[4] D L Price and S S Sisodia ldquoMutant genes in familialAlzheimerrsquos disease and transgenic modelsrdquo Annual Review ofNeuroscience vol 21 pp 479ndash505 1998
[5] D Harman ldquoAlzheimerrsquos disease pathogenesis role of agingrdquoAnnals of the New York Academy of Sciences vol 1067 no 1 pp454ndash460 2006
[6] D J Selkoe ldquoTranslating cell biology into therapeutic advancesin Alzheimerrsquos diseaserdquo Nature vol 399 pp A23ndashA31 1999
[7] H Braak and E Braak ldquoNeuropathological stageing of Alz-heimer-related changesrdquo Acta Neuropathologica vol 82 no 4pp 239ndash259 1991
[8] K Hirai G Aliev A Nunomara et al ldquoMitochondrial abnor-malities inAlzheimerrsquos diseaserdquoTheJournal ofNeuroscience vol21 pp 3017ndash3023 2001
[9] U T Brunk C B Jones and R S Sohal ldquoA novel hypothesisof lipofuscinogenesis and cellular aging based on interac-tions between oxidative stress and autophagocytosisrdquoMutationResearchDNAging vol 275 no 3ndash6 pp 395ndash403 1992
[10] P A Stewart K Hayakawa M-A Akers and H V Vinters ldquoAmorphometric study of the blood-brain barrier in Alzheimerrsquosdiseaserdquo Laboratory Investigation vol 67 no 6 pp 734ndash7421992
[11] J P Blass and G E Gibson ldquoThe role of oxidative abnor-malities in the pathophysiology of Alzheimerrsquos diseaserdquo RevueNeurologique vol 147 no 6-7 pp 513ndash525 1991
[12] B Pająk E Kania and A Orzechowski ldquoNucleofection of ratpheochromocytoma PC-12 cells with human mutated beta-amyloid precursor protein gene (APP-sw) leads to reducedreduced viability autophagy-like process and increased expres-sion and secretion of beta amyloidrdquo BioMed Research Interna-tional vol 2015 Article ID 746092 10 pages 2015
[13] J P Blass ldquoCerebrometabolic abnormalities in AlzheimerrsquosdiseaserdquoNeurological Research vol 25 no 6 pp 556ndash566 2003
12 Oxidative Medicine and Cellular Longevity
[14] A Eckert K Schmitt and J Gotz ldquoMitochondrial dysfunc-tionmdashthe beginning of the end in Alzheimerrsquos disease Sep-arate and synergistic modes of tau and amyloid-120573 toxicityrdquoAlzheimerrsquos Research ampTherapy vol 3 article 15 pp 1ndash11 2011
[15] U Keil A Bonert C A Marques et al ldquoAmyloid 120573-inducedchanges in nitric oxide production and mitochondrial activitylead to apoptosisrdquoThe Journal of Biological Chemistry vol 279no 48 pp 50310ndash50320 2004
[16] A M Swomley and D A Butterfield ldquoOxidative stress inAlzheimer disease and mild cognitive impairment evidencefrom human data provided by redox proteomicsrdquo Archives ofToxicology vol 89 no 10 pp 1669ndash1680 2015
[17] C Caspersen N Wang J Yao et al ldquoMitochondrial A120573 apotential focal point for neuronal metabolic dysfunction inAlzheimerrsquos diseaserdquo The FASEB Journal vol 19 no 14 pp2040ndash2041 2005
[18] G E Gibson K-F R Sheu and J P Blass ldquoAbnormalities ofmitochondrial enzymes inAlzheimer diseaserdquo Journal of NeuralTransmission vol 105 no 8-9 pp 855ndash870 1998
[19] A Eckert S Hauptmann I Scherping et al ldquoSoluble beta-amyloid leads to mitochondrial defects in amyloid precursorprotein and tau transgenic micerdquo Neurodegenerative Diseasesvol 5 no 3-4 pp 157ndash159 2008
[20] A Eckert K L Schulz V Rhein and J Gotz ldquoConvergenceof amyloid-120573 and tau pathologies on mitochondria in vivordquoMolecular Neurobiology vol 41 no 2-3 pp 107ndash114 2010
[21] S Hauptmann I Scherping S Drose et al ldquoMitochondrial dys-function an early event in Alzheimer pathology accumulateswith age in AD transgenic micerdquoNeurobiology of Aging vol 30no 10 pp 1574ndash1586 2009
[22] M Manczak B S Park Y Jung and P H Reddy ldquoDifferentialexpression of oxidative phosphorylation genes in patients withAlzheimerrsquos disease implications for early mitochondrial dys-function and oxidative damagerdquo NeuroMolecular Medicine vol5 no 2 pp 147ndash162 2004
[23] M P Mattson M Gleichmann and A Cheng ldquoMitochondriain neuroplasticity and neurological disordersrdquo Neuron vol 60no 5 pp 748ndash766 2008
[24] P I Moreira M S Santos and C R Oliveira ldquoAlzheimerrsquosdisease a lesson frommitochondrial dysfunctionrdquoAntioxidantsamp Redox Signaling vol 9 no 10 pp 1621ndash1630 2007
[25] P H Reddy ldquoMitochondrial dysfunction in aging and Alz-heimerrsquos disease strategies to protect neuronsrdquo Antioxidants ampRedox Signaling vol 9 no 10 pp 1647ndash1658 2007
[26] V Rhein G Baysang S Rao et al ldquoAmyloid-beta leads toimpaired cellular respiration energy production andmitochon-drial electron chain complex activities in humanneuroblastomacellsrdquo Cellular and Molecular Neurobiology vol 29 no 6-7 pp1063ndash1071 2009
[27] V Rhein X Song A Wiesner et al ldquoAmyloid-120573 and tausynergistically impair the oxidative phosphorylation system intriple transgenic Alzheimerrsquos disease micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 47 pp 20057ndash20062 2009
[28] R X Santos S C Correia X Wang et al ldquoAlzheimerrsquos diseasediverse aspects ofmitochondrial malfunctioningrdquo InternationalJournal of Clinical and Experimental Pathology vol 3 no 6 pp570ndash581 2010
[29] N R Sims ldquoEnergy metabolism oxidative stress and neuronaldegeneration in Alzheimerrsquos diseaserdquoNeurodegeneration vol 5no 4 pp 435ndash440 1996
[30] H K Anandatheerthavarada G Biswas M-A Robin and NG Avadhani ldquoMitochondrial targeting and a novel transmem-brane arrest of Alzheimerrsquos amyloid precursor protein impairsmitochondrial function in neuronal cellsrdquo The Journal of CellBiology vol 161 no 1 pp 41ndash54 2003
[31] L Devi B M Prabhu D F Galati N G Avadhani and HK Anandatheerthavarada ldquoAccumulation of amyloid precur-sor protein in the mitochondrial import channels of humanAlzheimerrsquos disease brain is associated with mitochondrialdysfunctionrdquo The Journal of Neuroscience vol 26 no 35 pp9057ndash9068 2006
[32] L Devi and H K Anandatheerthavarada ldquoMitochondrialtrafficking of APP and alpha synuclein relevance to mito-chondrial dysfunction in Alzheimerrsquos and Parkinsonrsquos diseasesrdquoBiochimica et Biophysica Acta vol 1802 no 1 pp 11ndash19 2010
[33] M Manczak T S Anekonda E Henson B S Park J Quinnand P H Reddy ldquoMitochondria are a direct site of A120573 accumu-lation in Alzheimerrsquos disease neurons implications for free rad-ical generation and oxidative damage in disease progressionrdquoHuman Molecular Genetics vol 15 no 9 pp 1437ndash1449 2006
[34] P F Pavlov C H Petersen E Glaser and M AnkarcronaldquoMitochondrial accumulation of APP and A120573 significancefor Alzheimer disease pathogenesisrdquo Journal of Cellular andMolecular Medicine vol 13 no 10 pp 4137ndash4145 2009
[35] C Schmidt E Lepsverdize S L Chi et al ldquoAmyloid precursorprotein and amyloid 120573-peptide bind to ATP synthase andregulate its activity at the surface of neural cellsrdquo MolecularPsychiatry vol 13 no 10 pp 953ndash969 2008
[36] K Johar A Priya and M T T Wong-Riley ldquoRegulation ofNa+K+-ATPase by nuclear respiratory factor 1 implication inthe tight coupling of neuronal activity energy generation andenergy consumptionrdquo The Journal of Biological Chemistry vol287 no 48 pp 40381ndash40390 2012
[37] J Astrup P M Sorensen and H R Sorensen ldquoOxygen andglucose consumption related to Na+-K+ transport in caninebrainrdquo Stroke vol 12 no 6 pp 726ndash730 1981
[38] M Erecinska and I A Silver ldquoATP and brain functionrdquo Journalof Cerebral Blood Flow and Metabolism vol 9 no 1 pp 2ndash191989
[39] MMata D J Fink H Gainer et al ldquoActivity-dependent energymetabolism in rat posterior pituitary primarily reflects sodiumpump activityrdquo Journal of Neurochemistry vol 34 no 1 pp 213ndash215 1980
[40] J Kang H-G Lemaire A Unterbeck et al ldquoThe precursor ofAlzheimerrsquos disease amyloid A4 protein resembles a cell-surfacereceptorrdquo Nature vol 325 no 6106 pp 733ndash736 1987
[41] C L Masters and D J Selkoe ldquoBiochemistry of amyloid 120573-protein and amyloid deposits in Alzheimer diseaserdquoCold SpringHarbour Perspectives in Medicine vol 2 no 6 Article IDa006262 2012
[42] A Lai S S Sisodia and I S Trowbridge ldquoCharacterization ofsorting signals in the beta-amyloid precursor protein cytoplas-mic domainrdquo The Journal of Biological Chemistry vol 270 no8 pp 3565ndash3573 1995
[43] A I Placido C M F Pereira A I Duarte et al ldquoThe role ofendoplasmic reticulum in amyloid precursor protein processingand trafficking implications for Alzheimerrsquos diseaserdquo Biochim-ica et Biophysica ActamdashMolecular Basis of Disease vol 1842 no9 pp 1444ndash1453 2014
Oxidative Medicine and Cellular Longevity 13
[44] C Kaether P Skehel and C G Dotti ldquoAxonal membraneproteins are transported in distinct carriers a two-color videomicroscopy study in cultured hippocampal neuronsrdquoMolecularBiology of the Cell vol 11 no 4 pp 1213ndash1224 2000
[45] A Kamal A Almenar-Queralt J F LeBlanc E A Robertsand L S B Goldstein ldquoKinesin-mediated axonal transportof a membrane compartment containing 120573-secretase andpresenilin-1 requires APPrdquo Nature vol 414 no 6864 pp 643ndash648 2001
[46] R G Perez S Soriano J D Hayes et al ldquoMutagenesis identi-fies new signals for 120573-amyloid precursor protein endocytosisturnover and the generation of secreted fragments includingA12057342rdquo The Journal of Biological Chemistry vol 274 no 27 pp18851ndash18856 1999
[47] D G Cook M S Forman J C Sung et al ldquoAlzheimerrsquos A120573(1ndash42) is generated in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo Nature Medicine vol 3 no 9 pp1021ndash1023 1997
[48] A S C Chyung B D Greenberg D G Cook R W Domsand V M-Y Lee ldquoNovel 120573-secretase cleavage of 120573-amyloidprecursor protein in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo The Journal of Cell Biology vol138 no 3 pp 671ndash680 1997
[49] RW-Y Choy Z Cheng and R Schekman ldquoAmyloid precursorprotein (APP) traffics from the cell surface via endosomesfor amyloid 120573 (A120573) production in the trans-Golgi networkrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 30 pp E2077ndashE2082 2012
[50] S A Small and S Gandy ldquoSorting through the cell biologyof Alzheimerrsquos disease intracellular pathways to pathogenesisrdquoNeuron vol 52 no 1 pp 15ndash31 2006
[51] M Goedert ldquoAlzheimerrsquos and Parkinsonrsquos diseases the prionconcept in relation to assembled A120573 tau and 120572-synucleinrdquoScience vol 349 no 6248 2015
[52] I Benilova E Karran and B De Strooper ldquoThe toxic A120573oligomer and Alzheimerrsquos disease an emperor in need ofclothesrdquo Nature Neuroscience vol 15 no 3 pp 349ndash357 2012
[53] E H Corder A M Saunders W J Strittmatter et al ldquoGenedose of apolipoprotein E type 4 allele and the risk of Alzheimerrsquosdisease in late onset familiesrdquo Science vol 261 no 5123 pp 921ndash923 1993
[54] S Brunholz S Sisodia A Lorenzo C Deyts S Kins and GMorfini ldquoAxonal transport of APP and the spatial regulationof APP cleavage and function in neuronal cellsrdquo ExperimentalBrain Research vol 217 no 3-4 pp 353ndash364 2012
[55] G Thinakaran and E H Koo ldquoAmyloid precursor proteintrafficking processing and functionrdquo The Journal of BiologicalChemistry vol 283 no 44 pp 29615ndash29619 2008
[56] T Saitoh M Sundsmo J-M Roch et al ldquoSecreted formof amyloid beta protein precursor is involved in the growthregulation of fibroblastsrdquo Cell vol 58 no 4 pp 615ndash622 1989
[57] M PMattson B Cheng A R Culwell F S Esch I Lieberburgand R E Rydel ldquoEvidence for excitoprotective and intraneu-ronal calcium-regulating roles for secreted forms of the 120573-amyloid precursor proteinrdquo Neuron vol 10 no 2 pp 243ndash2541993
[58] K J Lee C E H Moussa Y Lee et al ldquoBeta amyloid-independent role of amyloid precursor protein in generationandmaintenance of dendritic spinesrdquoNeuroscience vol 169 no1 pp 344ndash356 2010
[59] H Meziane J-C Dodart C Mathis et al ldquoMemory-enhancingeffects of secreted forms of the 120573-amyloid precursor proteinin normal and amnestic micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no21 pp 12683ndash12688 1998
[60] P R Turner K OrsquoConnor W P Tate and W C AbrahamldquoRoles of amyloid precursor protein and its fragments inregulating neural activity plasticity and memoryrdquo Progress inNeurobiology vol 70 no 1 pp 1ndash32 2003
[61] D Beher L Hesse C L Masters and GMulthaup ldquoRegulationof Amyloid Protein Precursor (APP) binding to collagen andmapping of the binding sites on APP and collagen type Irdquo TheJournal of Biological Chemistry vol 271 no 3 pp 1613ndash16201996
[62] S L Sabo A F Ikin J D Buxbaum and P Greengard ldquoTheamyloid precursor protein and its regulatory protein FE65 ingrowth cones and synapses in vitro and in vivordquo The Journal ofNeuroscience vol 23 no 13 pp 5407ndash5415 2003
[63] D H Small V Nurcombe G Reed et al ldquoA heparin-bindingdomain in the amyloid protein precursor of Alzheimerrsquos diseaseis involved in the regulation of neurite outgrowthrdquoThe Journalof Neuroscience vol 14 no 4 pp 2117ndash2127 1994
[64] T G Williamson S S Mok A Henry et al ldquoSecreted glypicanbinds to the amyloid precursor protein of Alzheimerrsquos disease(APP) and inhibits APP-induced neurite outgrowthrdquoThe Jour-nal of Biological Chemistry vol 271 no 49 pp 31215ndash312211996
[65] T Ondrejcak I Klyubin N-W Hu A E Barry W K Cullenand M J Rowan ldquoAlzheimerrsquos disease amyloid 120573-protein andsynaptic functionrdquo NeuroMolecular Medicine vol 12 no 1 pp13ndash26 2010
[66] D Puzzo L Privitera and A Palmeri ldquoHormetic effect ofamyloid-beta peptide in synaptic plasticity and memoryrdquo Neu-robiology of Aging vol 33 no 7 pp 1484e15ndash1484e24 2012
[67] C M F Pereira ldquoCrosstalk between endoplasmic reticulumstress and protein misfolding in neurodegenerative diseasesrdquoISRN Cell Biology vol 2013 Article ID 256404 22 pages 2013
[68] R-W Shin T C Saido M Maeda and T Kitamoto ldquoNovel 120572-secretase cleavage ofAlzheimerrsquos amyloid120573 precursor protein inthe endoplasmic reticulum of COS7 cellsrdquoNeuroscience Lettersvol 376 no 1 pp 14ndash19 2005
[69] E A Schon and E Area-Gomez ldquoMitochondria-associatedER membranes in Alzheimer diseaserdquo Molecular and CellularNeuroscience vol 55 pp 26ndash36 2013
[70] R Rizzuto P Pinton W Carrington et al ldquoClose contacts withthe endoplasmic reticulum as determinants of mitochondrialCa2+ responsesrdquo Science vol 280 no 5370 pp 1763ndash1766 1998
[71] S E Logue P Cleary S Saveljeva and A Samali ldquoNewdirections in ER-stress induced cell deathrdquoApoptosis vol 18 no5 pp 537ndash546 2013
[72] J P Decuypere KWelkenhuyzen T Luyten et al ldquoIns(145)P3receptor-mediated BI-1 Ca2+ signaling and autophagy induc-tion are interrelatedrdquo Autophagy vol 7 pp 1472ndash1489 2011
[73] K S Vetrivel and G Thinakaran ldquoMembrane rafts in Alz-heimerrsquos disease beta-amyloid productionrdquo Biochimica et Bio-physica Acta vol 1801 no 8 pp 860ndash867 2010
[74] H Urra E Dufey F Lisbona D Rojas-Rivera and C HetzldquoWhen ER stress reaches a dead endrdquo Biochimica et BiophysicaActa (BBA) Molecular Cell Research vol 1833 no 12 pp 3507ndash3517 2013
14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Oxidative Medicine and Cellular Longevity
It is ranked as the fourth leading cause of death in modernsocieties where average life span increased greatly in the lasttwo decades As 95 of AD cases are diagnosed in olderpeople one might think that a causal relationship existsbetween aging and the onset of disease Certainly a numberof similarities between getting old and being affected withAD could be listed Historically the free radical theory ofaging by Harman [155] suggested aging as ldquoside effectrdquo ofreactive oxygen species formed in mitochondrial respiratorychain Apparently free radicals commonly generated byincomplete reduction of oxygen molecule at complexes Iand III of mitochondrial ETC are capable of damagingDNA RNA and proteins They impair energy storage andlead to operational failure of mitochondria with progres-sive decline of cell viability Almost identical conditionsaccompany AD and are an explicit step in pathogenesisof disease As mtDNA is deprived of repair mechanismsROS-induced DNA strand breaks tend to accumulate withage or AD Thus mtDNA is a vulnerable target for ROSbut the reverse the ROS generation due to the mutatedmtDNA is not convincingly confirmed [156] Moreover theevidence that mutated mtDNA accelerates the progress ofaging is also questioned based on the results from studycarried out on transgenic mice model [157] Although someauthors show inconsistency between the free radical theoryand observations cumulative evaluation of the scientificreports points to antioxidant defense systems as importantfactors in protection from premature aging [158] Othermitochondrial components important for their function arehampered with age adenine nucleotide translocase (ANT)nitric oxide synthase (NOS) and carnitine acyltransferase(CT) [159ndash161] Actually NOS activity is elevated in AD asreported from study performed on cellular model of disease[15] We could not find any information about CT activityin AD whereas ANT activity is noticeably inhibited byA120573 or hyperphosphorylated tau and this effect is reversedby mersalyl a reversible alkylating agent of thiol groups[162] Mitochondrial dysfunction observed in transgenicmice models of AD and aging demonstrates higher activ-ity of genes controlling energy metabolism and apoptosisTaken together physiological aging and AD are associatedwith broad-spectrum dysfunction of mitochondria but thefoundations of mitochondrial decline are dissimilar Morediscrepancies between physiological aging and AD werefound with respect to ERS and UPR which play a significantrole in cellular proteostasis Dolichol was selected as an agingmarker because the progressive increase in dolichol levelwas observed in aging brain [163] In contrast ubiquinoneconcentration which is also synthesized from geranylgeranioldiminishes with agingwhereas cholesterol and dolichyl phos-phate concentrations remain unaltered In AD decreasedlevels of dolichol were observed and increased levels ofubiquinone and dolichyl phosphate without any changesin brain cholesterol AD cannot be regarded as a resultof premature aging The drop in dolichol and augmenteddolichyl phosphate concentration points toward disturbedglycosylation in ER of diseased brain while the increase inubiquinone suggests efforts to protect the brain from oxida-tive stress induced by lipid peroxidation [164 165]
14 Targeting ER Stress in AD Therapy
As ERS is a recognized factor in AD drugs that interfere withERS would theoretically have great therapeutic potentialThere are several compounds grouped in classes thatinteract directly with components of the ERS (salubrinalBiP inducer X (BIX) salicylamide analogs flavonoidsguanabenz and STF083010) chemical chaperons (4-phenyl-butyric acid (PBA) tauroursodeoxycholic acid (TUDCA)and trimethylamine oxide (TMAO)) chemicals that inhibitprotein degradation (Eeyarestatin MG132 and Bortezomib)compounds with antioxidant activity (Edaravone diben-zoylmethane derivatives and N-acetyl cysteine (NAC)) anddrugs controlling calcium signaling (dantrolene and carba-zole derivatives) [166] They may act by inducing transienttranslation arrest upregulation of chaperone proteins andaugmented degradation of ER-associated misfolded proteinsFundamental approach in the development of new therapyis the selection of appropriate molecular targets In ER stresssignaling the aim is to alter the expression of ER stress-associated molecules that can rescue cells from the toxiceffect of ERS Recent efforts in establishing new promisingdrugs against AD are pointing to chemical chaperones suchas PBA TUDCA or trimethylamine oxide (TMAO) Thesesubstances improve protein folding and alleviate nativeprotein conformation [166] It was shown on mouse modelsof AD that PBA TUDCA and TMAO stop A120573 accumulationand avoid the loss of dendritic spines [167] Some observa-tions even demonstrated improved memory and cognitivefunctions [168] associated with improved cell survival [169]Salubrinal ((2E)-3-phenyl-N-[222-trichloro-1-[[(8-quinoli-nylamino)thioxomethyl]amino]ethyl]-2-propenamide Sal)selectively inhibits growth arrest and DNA damage inducedgene 34- (GADD34-) phosphatase complex (GADD34associates with protein phosphatase 1 (PP1)) and promotes invitro dephosphorylation of the alpha subunit of eIF-2120572 andIRE1120572ASK1JNK signaling pathway being protective againstERS even induced by tunicamycin Tm [170] In a great deal ofexperiments testing Sal in cultured cells and animal modelsof AD this substance increased the viability of neuronal cellsand A120573 toxicity [171 172] BIX (2-(34-dihydroxyphenyl)-2-oxoethyl ester thiocyanic acid) preferentially inducedBiP mRNA in an ATF6-dependent manner leading toreduced Tm-induced death of neuronal cells [173] AlsoDBM derivative 14ndash26 (221015840-dimethoxydibenzoylmethane)was found to be neuroprotective for SH-SY5Y and PC-12cells by decreasing expression of BiP and CHOP [174]Dantrolene a ryanodine receptor antagonist that inhibitsabnormal calcium release from the ER inhibited expressionof both phosphorylated PERK and eIF2120572 It also reducedCHOP expression and attenuated thapsigargin-inducedapoptosis in PC-12 cells [175] Neuroprotective effects similarto dantrolene were observed for ([9-(3-cyanobenzyl)-14-dimethylcarbazole]) This substance suppressed increases inintracellular Ca2+ in PC-12 cells treated with thapsigarginand reduced levels of BiP and CHOP [176]
Aforementioned compounds were chosen from othersas the most potent ER stress inhibitors and persuasivelyprotective to neuronal cells 18 of 42 different compounds
Oxidative Medicine and Cellular Longevity 11
were exploited in in vivo and in vitro models of centralnervous system disorders with in fact improved cell or tissueviability [166] Thus the brain is the most frequently investi-gated organ in the context of ERS From these experimentsit becomes clear that CHOP functions as proapoptotic factorThe roles of other specific ER stress molecules as moleculartargets for pharmacological intervention are less clear andvary depending on cell type and context
There are few underestimated modulations in APP pro-cessing that shed more light on current dogma of AD patho-genesis The modulation of mevalonate pathway and choles-terol synthesis were reported to stimulate nonamyloidogenicpathway of APP processing [177] Additionally cholesterolderivative 27-hydroxycholesterol (27-OHC) was shown toinduce ER stress which attenuated leptin-dependent viabilityby activating CHOP in SH-SY5Y neuroblastoma cells [178]Irrespective of a number of compounds examined in AD thecall for new drugs modulating ER stress with healing effect isstill waiting to be revealed
Abbreviations
APOE Apolipoprotein EAPPs Secreted form of APPCHOPGADD153 CEBP homologous protein
growth arrest and DNA damageinduced gene 153
Drp1 Dynamin-1-like proteinERAD Endoplasmic-reticulum-
associated protein degradationERO1120572 ER oxidase 1120572ERS Endoplasmic reticulum stressFOH FarnesolGGOH GeranylgeraniolGRP75 Glucose-regulated protein 75GRP78BiP 78 kDa glucose-regulated proteinGSK-3120573 Synthase kinase 3 betaIP3R Inositol triphosphate receptor
IRE1120572 Inositol-requiring enzyme 1 alphaMAM Mitochondria-associated
membraneMAP LC3 Microtubule-associated protein
light chain 3Mfn2 GTPase mitofusin 2MMP Mitochondrial membrane
potentialMPTP Mitochondrial permeability
transition poreNFT Neurofibrillary tanglesPACS-2 Phosphofurin acidic cluster
sorting protein 2PDI Protein disulfide isomerasePERK Protein kinase R- (PKR-) like
ER kinaseRNS Reactive nitrogen speciesROS Reactive oxygen speciesRyR Ryanodine receptorTOM Transporter outer membraneTIM Transporter inner membrane
UPR Unfolded protein responseVDAC Voltage-dependent anion channelXBP-1 Xbox binding protein 1
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
Support for this work was provided by Grant no UMO-201103BNZ70238 from the National Science Centre inPoland
References
[1] R L Nussbaum andC E Ellis ldquoAlzheimerrsquos disease and Parkin-sonrsquos diseaserdquoTheNewEngland Journal ofMedicine vol 348 no14 pp 1356ndash1364 2003
[2] A L Brunkan and A M Goate ldquoPresenilin function and 120574-secretase activityrdquoThe Journal of Neurochemistry vol 93 no 4pp 769ndash792 2005
[3] V Crentsil ldquoThe pharmacogenomics of Alzheimerrsquos diseaserdquoAgeing Research Reviews vol 3 no 2 pp 153ndash169 2004
[4] D L Price and S S Sisodia ldquoMutant genes in familialAlzheimerrsquos disease and transgenic modelsrdquo Annual Review ofNeuroscience vol 21 pp 479ndash505 1998
[5] D Harman ldquoAlzheimerrsquos disease pathogenesis role of agingrdquoAnnals of the New York Academy of Sciences vol 1067 no 1 pp454ndash460 2006
[6] D J Selkoe ldquoTranslating cell biology into therapeutic advancesin Alzheimerrsquos diseaserdquo Nature vol 399 pp A23ndashA31 1999
[7] H Braak and E Braak ldquoNeuropathological stageing of Alz-heimer-related changesrdquo Acta Neuropathologica vol 82 no 4pp 239ndash259 1991
[8] K Hirai G Aliev A Nunomara et al ldquoMitochondrial abnor-malities inAlzheimerrsquos diseaserdquoTheJournal ofNeuroscience vol21 pp 3017ndash3023 2001
[9] U T Brunk C B Jones and R S Sohal ldquoA novel hypothesisof lipofuscinogenesis and cellular aging based on interac-tions between oxidative stress and autophagocytosisrdquoMutationResearchDNAging vol 275 no 3ndash6 pp 395ndash403 1992
[10] P A Stewart K Hayakawa M-A Akers and H V Vinters ldquoAmorphometric study of the blood-brain barrier in Alzheimerrsquosdiseaserdquo Laboratory Investigation vol 67 no 6 pp 734ndash7421992
[11] J P Blass and G E Gibson ldquoThe role of oxidative abnor-malities in the pathophysiology of Alzheimerrsquos diseaserdquo RevueNeurologique vol 147 no 6-7 pp 513ndash525 1991
[12] B Pająk E Kania and A Orzechowski ldquoNucleofection of ratpheochromocytoma PC-12 cells with human mutated beta-amyloid precursor protein gene (APP-sw) leads to reducedreduced viability autophagy-like process and increased expres-sion and secretion of beta amyloidrdquo BioMed Research Interna-tional vol 2015 Article ID 746092 10 pages 2015
[13] J P Blass ldquoCerebrometabolic abnormalities in AlzheimerrsquosdiseaserdquoNeurological Research vol 25 no 6 pp 556ndash566 2003
12 Oxidative Medicine and Cellular Longevity
[14] A Eckert K Schmitt and J Gotz ldquoMitochondrial dysfunc-tionmdashthe beginning of the end in Alzheimerrsquos disease Sep-arate and synergistic modes of tau and amyloid-120573 toxicityrdquoAlzheimerrsquos Research ampTherapy vol 3 article 15 pp 1ndash11 2011
[15] U Keil A Bonert C A Marques et al ldquoAmyloid 120573-inducedchanges in nitric oxide production and mitochondrial activitylead to apoptosisrdquoThe Journal of Biological Chemistry vol 279no 48 pp 50310ndash50320 2004
[16] A M Swomley and D A Butterfield ldquoOxidative stress inAlzheimer disease and mild cognitive impairment evidencefrom human data provided by redox proteomicsrdquo Archives ofToxicology vol 89 no 10 pp 1669ndash1680 2015
[17] C Caspersen N Wang J Yao et al ldquoMitochondrial A120573 apotential focal point for neuronal metabolic dysfunction inAlzheimerrsquos diseaserdquo The FASEB Journal vol 19 no 14 pp2040ndash2041 2005
[18] G E Gibson K-F R Sheu and J P Blass ldquoAbnormalities ofmitochondrial enzymes inAlzheimer diseaserdquo Journal of NeuralTransmission vol 105 no 8-9 pp 855ndash870 1998
[19] A Eckert S Hauptmann I Scherping et al ldquoSoluble beta-amyloid leads to mitochondrial defects in amyloid precursorprotein and tau transgenic micerdquo Neurodegenerative Diseasesvol 5 no 3-4 pp 157ndash159 2008
[20] A Eckert K L Schulz V Rhein and J Gotz ldquoConvergenceof amyloid-120573 and tau pathologies on mitochondria in vivordquoMolecular Neurobiology vol 41 no 2-3 pp 107ndash114 2010
[21] S Hauptmann I Scherping S Drose et al ldquoMitochondrial dys-function an early event in Alzheimer pathology accumulateswith age in AD transgenic micerdquoNeurobiology of Aging vol 30no 10 pp 1574ndash1586 2009
[22] M Manczak B S Park Y Jung and P H Reddy ldquoDifferentialexpression of oxidative phosphorylation genes in patients withAlzheimerrsquos disease implications for early mitochondrial dys-function and oxidative damagerdquo NeuroMolecular Medicine vol5 no 2 pp 147ndash162 2004
[23] M P Mattson M Gleichmann and A Cheng ldquoMitochondriain neuroplasticity and neurological disordersrdquo Neuron vol 60no 5 pp 748ndash766 2008
[24] P I Moreira M S Santos and C R Oliveira ldquoAlzheimerrsquosdisease a lesson frommitochondrial dysfunctionrdquoAntioxidantsamp Redox Signaling vol 9 no 10 pp 1621ndash1630 2007
[25] P H Reddy ldquoMitochondrial dysfunction in aging and Alz-heimerrsquos disease strategies to protect neuronsrdquo Antioxidants ampRedox Signaling vol 9 no 10 pp 1647ndash1658 2007
[26] V Rhein G Baysang S Rao et al ldquoAmyloid-beta leads toimpaired cellular respiration energy production andmitochon-drial electron chain complex activities in humanneuroblastomacellsrdquo Cellular and Molecular Neurobiology vol 29 no 6-7 pp1063ndash1071 2009
[27] V Rhein X Song A Wiesner et al ldquoAmyloid-120573 and tausynergistically impair the oxidative phosphorylation system intriple transgenic Alzheimerrsquos disease micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 47 pp 20057ndash20062 2009
[28] R X Santos S C Correia X Wang et al ldquoAlzheimerrsquos diseasediverse aspects ofmitochondrial malfunctioningrdquo InternationalJournal of Clinical and Experimental Pathology vol 3 no 6 pp570ndash581 2010
[29] N R Sims ldquoEnergy metabolism oxidative stress and neuronaldegeneration in Alzheimerrsquos diseaserdquoNeurodegeneration vol 5no 4 pp 435ndash440 1996
[30] H K Anandatheerthavarada G Biswas M-A Robin and NG Avadhani ldquoMitochondrial targeting and a novel transmem-brane arrest of Alzheimerrsquos amyloid precursor protein impairsmitochondrial function in neuronal cellsrdquo The Journal of CellBiology vol 161 no 1 pp 41ndash54 2003
[31] L Devi B M Prabhu D F Galati N G Avadhani and HK Anandatheerthavarada ldquoAccumulation of amyloid precur-sor protein in the mitochondrial import channels of humanAlzheimerrsquos disease brain is associated with mitochondrialdysfunctionrdquo The Journal of Neuroscience vol 26 no 35 pp9057ndash9068 2006
[32] L Devi and H K Anandatheerthavarada ldquoMitochondrialtrafficking of APP and alpha synuclein relevance to mito-chondrial dysfunction in Alzheimerrsquos and Parkinsonrsquos diseasesrdquoBiochimica et Biophysica Acta vol 1802 no 1 pp 11ndash19 2010
[33] M Manczak T S Anekonda E Henson B S Park J Quinnand P H Reddy ldquoMitochondria are a direct site of A120573 accumu-lation in Alzheimerrsquos disease neurons implications for free rad-ical generation and oxidative damage in disease progressionrdquoHuman Molecular Genetics vol 15 no 9 pp 1437ndash1449 2006
[34] P F Pavlov C H Petersen E Glaser and M AnkarcronaldquoMitochondrial accumulation of APP and A120573 significancefor Alzheimer disease pathogenesisrdquo Journal of Cellular andMolecular Medicine vol 13 no 10 pp 4137ndash4145 2009
[35] C Schmidt E Lepsverdize S L Chi et al ldquoAmyloid precursorprotein and amyloid 120573-peptide bind to ATP synthase andregulate its activity at the surface of neural cellsrdquo MolecularPsychiatry vol 13 no 10 pp 953ndash969 2008
[36] K Johar A Priya and M T T Wong-Riley ldquoRegulation ofNa+K+-ATPase by nuclear respiratory factor 1 implication inthe tight coupling of neuronal activity energy generation andenergy consumptionrdquo The Journal of Biological Chemistry vol287 no 48 pp 40381ndash40390 2012
[37] J Astrup P M Sorensen and H R Sorensen ldquoOxygen andglucose consumption related to Na+-K+ transport in caninebrainrdquo Stroke vol 12 no 6 pp 726ndash730 1981
[38] M Erecinska and I A Silver ldquoATP and brain functionrdquo Journalof Cerebral Blood Flow and Metabolism vol 9 no 1 pp 2ndash191989
[39] MMata D J Fink H Gainer et al ldquoActivity-dependent energymetabolism in rat posterior pituitary primarily reflects sodiumpump activityrdquo Journal of Neurochemistry vol 34 no 1 pp 213ndash215 1980
[40] J Kang H-G Lemaire A Unterbeck et al ldquoThe precursor ofAlzheimerrsquos disease amyloid A4 protein resembles a cell-surfacereceptorrdquo Nature vol 325 no 6106 pp 733ndash736 1987
[41] C L Masters and D J Selkoe ldquoBiochemistry of amyloid 120573-protein and amyloid deposits in Alzheimer diseaserdquoCold SpringHarbour Perspectives in Medicine vol 2 no 6 Article IDa006262 2012
[42] A Lai S S Sisodia and I S Trowbridge ldquoCharacterization ofsorting signals in the beta-amyloid precursor protein cytoplas-mic domainrdquo The Journal of Biological Chemistry vol 270 no8 pp 3565ndash3573 1995
[43] A I Placido C M F Pereira A I Duarte et al ldquoThe role ofendoplasmic reticulum in amyloid precursor protein processingand trafficking implications for Alzheimerrsquos diseaserdquo Biochim-ica et Biophysica ActamdashMolecular Basis of Disease vol 1842 no9 pp 1444ndash1453 2014
Oxidative Medicine and Cellular Longevity 13
[44] C Kaether P Skehel and C G Dotti ldquoAxonal membraneproteins are transported in distinct carriers a two-color videomicroscopy study in cultured hippocampal neuronsrdquoMolecularBiology of the Cell vol 11 no 4 pp 1213ndash1224 2000
[45] A Kamal A Almenar-Queralt J F LeBlanc E A Robertsand L S B Goldstein ldquoKinesin-mediated axonal transportof a membrane compartment containing 120573-secretase andpresenilin-1 requires APPrdquo Nature vol 414 no 6864 pp 643ndash648 2001
[46] R G Perez S Soriano J D Hayes et al ldquoMutagenesis identi-fies new signals for 120573-amyloid precursor protein endocytosisturnover and the generation of secreted fragments includingA12057342rdquo The Journal of Biological Chemistry vol 274 no 27 pp18851ndash18856 1999
[47] D G Cook M S Forman J C Sung et al ldquoAlzheimerrsquos A120573(1ndash42) is generated in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo Nature Medicine vol 3 no 9 pp1021ndash1023 1997
[48] A S C Chyung B D Greenberg D G Cook R W Domsand V M-Y Lee ldquoNovel 120573-secretase cleavage of 120573-amyloidprecursor protein in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo The Journal of Cell Biology vol138 no 3 pp 671ndash680 1997
[49] RW-Y Choy Z Cheng and R Schekman ldquoAmyloid precursorprotein (APP) traffics from the cell surface via endosomesfor amyloid 120573 (A120573) production in the trans-Golgi networkrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 30 pp E2077ndashE2082 2012
[50] S A Small and S Gandy ldquoSorting through the cell biologyof Alzheimerrsquos disease intracellular pathways to pathogenesisrdquoNeuron vol 52 no 1 pp 15ndash31 2006
[51] M Goedert ldquoAlzheimerrsquos and Parkinsonrsquos diseases the prionconcept in relation to assembled A120573 tau and 120572-synucleinrdquoScience vol 349 no 6248 2015
[52] I Benilova E Karran and B De Strooper ldquoThe toxic A120573oligomer and Alzheimerrsquos disease an emperor in need ofclothesrdquo Nature Neuroscience vol 15 no 3 pp 349ndash357 2012
[53] E H Corder A M Saunders W J Strittmatter et al ldquoGenedose of apolipoprotein E type 4 allele and the risk of Alzheimerrsquosdisease in late onset familiesrdquo Science vol 261 no 5123 pp 921ndash923 1993
[54] S Brunholz S Sisodia A Lorenzo C Deyts S Kins and GMorfini ldquoAxonal transport of APP and the spatial regulationof APP cleavage and function in neuronal cellsrdquo ExperimentalBrain Research vol 217 no 3-4 pp 353ndash364 2012
[55] G Thinakaran and E H Koo ldquoAmyloid precursor proteintrafficking processing and functionrdquo The Journal of BiologicalChemistry vol 283 no 44 pp 29615ndash29619 2008
[56] T Saitoh M Sundsmo J-M Roch et al ldquoSecreted formof amyloid beta protein precursor is involved in the growthregulation of fibroblastsrdquo Cell vol 58 no 4 pp 615ndash622 1989
[57] M PMattson B Cheng A R Culwell F S Esch I Lieberburgand R E Rydel ldquoEvidence for excitoprotective and intraneu-ronal calcium-regulating roles for secreted forms of the 120573-amyloid precursor proteinrdquo Neuron vol 10 no 2 pp 243ndash2541993
[58] K J Lee C E H Moussa Y Lee et al ldquoBeta amyloid-independent role of amyloid precursor protein in generationandmaintenance of dendritic spinesrdquoNeuroscience vol 169 no1 pp 344ndash356 2010
[59] H Meziane J-C Dodart C Mathis et al ldquoMemory-enhancingeffects of secreted forms of the 120573-amyloid precursor proteinin normal and amnestic micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no21 pp 12683ndash12688 1998
[60] P R Turner K OrsquoConnor W P Tate and W C AbrahamldquoRoles of amyloid precursor protein and its fragments inregulating neural activity plasticity and memoryrdquo Progress inNeurobiology vol 70 no 1 pp 1ndash32 2003
[61] D Beher L Hesse C L Masters and GMulthaup ldquoRegulationof Amyloid Protein Precursor (APP) binding to collagen andmapping of the binding sites on APP and collagen type Irdquo TheJournal of Biological Chemistry vol 271 no 3 pp 1613ndash16201996
[62] S L Sabo A F Ikin J D Buxbaum and P Greengard ldquoTheamyloid precursor protein and its regulatory protein FE65 ingrowth cones and synapses in vitro and in vivordquo The Journal ofNeuroscience vol 23 no 13 pp 5407ndash5415 2003
[63] D H Small V Nurcombe G Reed et al ldquoA heparin-bindingdomain in the amyloid protein precursor of Alzheimerrsquos diseaseis involved in the regulation of neurite outgrowthrdquoThe Journalof Neuroscience vol 14 no 4 pp 2117ndash2127 1994
[64] T G Williamson S S Mok A Henry et al ldquoSecreted glypicanbinds to the amyloid precursor protein of Alzheimerrsquos disease(APP) and inhibits APP-induced neurite outgrowthrdquoThe Jour-nal of Biological Chemistry vol 271 no 49 pp 31215ndash312211996
[65] T Ondrejcak I Klyubin N-W Hu A E Barry W K Cullenand M J Rowan ldquoAlzheimerrsquos disease amyloid 120573-protein andsynaptic functionrdquo NeuroMolecular Medicine vol 12 no 1 pp13ndash26 2010
[66] D Puzzo L Privitera and A Palmeri ldquoHormetic effect ofamyloid-beta peptide in synaptic plasticity and memoryrdquo Neu-robiology of Aging vol 33 no 7 pp 1484e15ndash1484e24 2012
[67] C M F Pereira ldquoCrosstalk between endoplasmic reticulumstress and protein misfolding in neurodegenerative diseasesrdquoISRN Cell Biology vol 2013 Article ID 256404 22 pages 2013
[68] R-W Shin T C Saido M Maeda and T Kitamoto ldquoNovel 120572-secretase cleavage ofAlzheimerrsquos amyloid120573 precursor protein inthe endoplasmic reticulum of COS7 cellsrdquoNeuroscience Lettersvol 376 no 1 pp 14ndash19 2005
[69] E A Schon and E Area-Gomez ldquoMitochondria-associatedER membranes in Alzheimer diseaserdquo Molecular and CellularNeuroscience vol 55 pp 26ndash36 2013
[70] R Rizzuto P Pinton W Carrington et al ldquoClose contacts withthe endoplasmic reticulum as determinants of mitochondrialCa2+ responsesrdquo Science vol 280 no 5370 pp 1763ndash1766 1998
[71] S E Logue P Cleary S Saveljeva and A Samali ldquoNewdirections in ER-stress induced cell deathrdquoApoptosis vol 18 no5 pp 537ndash546 2013
[72] J P Decuypere KWelkenhuyzen T Luyten et al ldquoIns(145)P3receptor-mediated BI-1 Ca2+ signaling and autophagy induc-tion are interrelatedrdquo Autophagy vol 7 pp 1472ndash1489 2011
[73] K S Vetrivel and G Thinakaran ldquoMembrane rafts in Alz-heimerrsquos disease beta-amyloid productionrdquo Biochimica et Bio-physica Acta vol 1801 no 8 pp 860ndash867 2010
[74] H Urra E Dufey F Lisbona D Rojas-Rivera and C HetzldquoWhen ER stress reaches a dead endrdquo Biochimica et BiophysicaActa (BBA) Molecular Cell Research vol 1833 no 12 pp 3507ndash3517 2013
14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 11
were exploited in in vivo and in vitro models of centralnervous system disorders with in fact improved cell or tissueviability [166] Thus the brain is the most frequently investi-gated organ in the context of ERS From these experimentsit becomes clear that CHOP functions as proapoptotic factorThe roles of other specific ER stress molecules as moleculartargets for pharmacological intervention are less clear andvary depending on cell type and context
There are few underestimated modulations in APP pro-cessing that shed more light on current dogma of AD patho-genesis The modulation of mevalonate pathway and choles-terol synthesis were reported to stimulate nonamyloidogenicpathway of APP processing [177] Additionally cholesterolderivative 27-hydroxycholesterol (27-OHC) was shown toinduce ER stress which attenuated leptin-dependent viabilityby activating CHOP in SH-SY5Y neuroblastoma cells [178]Irrespective of a number of compounds examined in AD thecall for new drugs modulating ER stress with healing effect isstill waiting to be revealed
Abbreviations
APOE Apolipoprotein EAPPs Secreted form of APPCHOPGADD153 CEBP homologous protein
growth arrest and DNA damageinduced gene 153
Drp1 Dynamin-1-like proteinERAD Endoplasmic-reticulum-
associated protein degradationERO1120572 ER oxidase 1120572ERS Endoplasmic reticulum stressFOH FarnesolGGOH GeranylgeraniolGRP75 Glucose-regulated protein 75GRP78BiP 78 kDa glucose-regulated proteinGSK-3120573 Synthase kinase 3 betaIP3R Inositol triphosphate receptor
IRE1120572 Inositol-requiring enzyme 1 alphaMAM Mitochondria-associated
membraneMAP LC3 Microtubule-associated protein
light chain 3Mfn2 GTPase mitofusin 2MMP Mitochondrial membrane
potentialMPTP Mitochondrial permeability
transition poreNFT Neurofibrillary tanglesPACS-2 Phosphofurin acidic cluster
sorting protein 2PDI Protein disulfide isomerasePERK Protein kinase R- (PKR-) like
ER kinaseRNS Reactive nitrogen speciesROS Reactive oxygen speciesRyR Ryanodine receptorTOM Transporter outer membraneTIM Transporter inner membrane
UPR Unfolded protein responseVDAC Voltage-dependent anion channelXBP-1 Xbox binding protein 1
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
Support for this work was provided by Grant no UMO-201103BNZ70238 from the National Science Centre inPoland
References
[1] R L Nussbaum andC E Ellis ldquoAlzheimerrsquos disease and Parkin-sonrsquos diseaserdquoTheNewEngland Journal ofMedicine vol 348 no14 pp 1356ndash1364 2003
[2] A L Brunkan and A M Goate ldquoPresenilin function and 120574-secretase activityrdquoThe Journal of Neurochemistry vol 93 no 4pp 769ndash792 2005
[3] V Crentsil ldquoThe pharmacogenomics of Alzheimerrsquos diseaserdquoAgeing Research Reviews vol 3 no 2 pp 153ndash169 2004
[4] D L Price and S S Sisodia ldquoMutant genes in familialAlzheimerrsquos disease and transgenic modelsrdquo Annual Review ofNeuroscience vol 21 pp 479ndash505 1998
[5] D Harman ldquoAlzheimerrsquos disease pathogenesis role of agingrdquoAnnals of the New York Academy of Sciences vol 1067 no 1 pp454ndash460 2006
[6] D J Selkoe ldquoTranslating cell biology into therapeutic advancesin Alzheimerrsquos diseaserdquo Nature vol 399 pp A23ndashA31 1999
[7] H Braak and E Braak ldquoNeuropathological stageing of Alz-heimer-related changesrdquo Acta Neuropathologica vol 82 no 4pp 239ndash259 1991
[8] K Hirai G Aliev A Nunomara et al ldquoMitochondrial abnor-malities inAlzheimerrsquos diseaserdquoTheJournal ofNeuroscience vol21 pp 3017ndash3023 2001
[9] U T Brunk C B Jones and R S Sohal ldquoA novel hypothesisof lipofuscinogenesis and cellular aging based on interac-tions between oxidative stress and autophagocytosisrdquoMutationResearchDNAging vol 275 no 3ndash6 pp 395ndash403 1992
[10] P A Stewart K Hayakawa M-A Akers and H V Vinters ldquoAmorphometric study of the blood-brain barrier in Alzheimerrsquosdiseaserdquo Laboratory Investigation vol 67 no 6 pp 734ndash7421992
[11] J P Blass and G E Gibson ldquoThe role of oxidative abnor-malities in the pathophysiology of Alzheimerrsquos diseaserdquo RevueNeurologique vol 147 no 6-7 pp 513ndash525 1991
[12] B Pająk E Kania and A Orzechowski ldquoNucleofection of ratpheochromocytoma PC-12 cells with human mutated beta-amyloid precursor protein gene (APP-sw) leads to reducedreduced viability autophagy-like process and increased expres-sion and secretion of beta amyloidrdquo BioMed Research Interna-tional vol 2015 Article ID 746092 10 pages 2015
[13] J P Blass ldquoCerebrometabolic abnormalities in AlzheimerrsquosdiseaserdquoNeurological Research vol 25 no 6 pp 556ndash566 2003
12 Oxidative Medicine and Cellular Longevity
[14] A Eckert K Schmitt and J Gotz ldquoMitochondrial dysfunc-tionmdashthe beginning of the end in Alzheimerrsquos disease Sep-arate and synergistic modes of tau and amyloid-120573 toxicityrdquoAlzheimerrsquos Research ampTherapy vol 3 article 15 pp 1ndash11 2011
[15] U Keil A Bonert C A Marques et al ldquoAmyloid 120573-inducedchanges in nitric oxide production and mitochondrial activitylead to apoptosisrdquoThe Journal of Biological Chemistry vol 279no 48 pp 50310ndash50320 2004
[16] A M Swomley and D A Butterfield ldquoOxidative stress inAlzheimer disease and mild cognitive impairment evidencefrom human data provided by redox proteomicsrdquo Archives ofToxicology vol 89 no 10 pp 1669ndash1680 2015
[17] C Caspersen N Wang J Yao et al ldquoMitochondrial A120573 apotential focal point for neuronal metabolic dysfunction inAlzheimerrsquos diseaserdquo The FASEB Journal vol 19 no 14 pp2040ndash2041 2005
[18] G E Gibson K-F R Sheu and J P Blass ldquoAbnormalities ofmitochondrial enzymes inAlzheimer diseaserdquo Journal of NeuralTransmission vol 105 no 8-9 pp 855ndash870 1998
[19] A Eckert S Hauptmann I Scherping et al ldquoSoluble beta-amyloid leads to mitochondrial defects in amyloid precursorprotein and tau transgenic micerdquo Neurodegenerative Diseasesvol 5 no 3-4 pp 157ndash159 2008
[20] A Eckert K L Schulz V Rhein and J Gotz ldquoConvergenceof amyloid-120573 and tau pathologies on mitochondria in vivordquoMolecular Neurobiology vol 41 no 2-3 pp 107ndash114 2010
[21] S Hauptmann I Scherping S Drose et al ldquoMitochondrial dys-function an early event in Alzheimer pathology accumulateswith age in AD transgenic micerdquoNeurobiology of Aging vol 30no 10 pp 1574ndash1586 2009
[22] M Manczak B S Park Y Jung and P H Reddy ldquoDifferentialexpression of oxidative phosphorylation genes in patients withAlzheimerrsquos disease implications for early mitochondrial dys-function and oxidative damagerdquo NeuroMolecular Medicine vol5 no 2 pp 147ndash162 2004
[23] M P Mattson M Gleichmann and A Cheng ldquoMitochondriain neuroplasticity and neurological disordersrdquo Neuron vol 60no 5 pp 748ndash766 2008
[24] P I Moreira M S Santos and C R Oliveira ldquoAlzheimerrsquosdisease a lesson frommitochondrial dysfunctionrdquoAntioxidantsamp Redox Signaling vol 9 no 10 pp 1621ndash1630 2007
[25] P H Reddy ldquoMitochondrial dysfunction in aging and Alz-heimerrsquos disease strategies to protect neuronsrdquo Antioxidants ampRedox Signaling vol 9 no 10 pp 1647ndash1658 2007
[26] V Rhein G Baysang S Rao et al ldquoAmyloid-beta leads toimpaired cellular respiration energy production andmitochon-drial electron chain complex activities in humanneuroblastomacellsrdquo Cellular and Molecular Neurobiology vol 29 no 6-7 pp1063ndash1071 2009
[27] V Rhein X Song A Wiesner et al ldquoAmyloid-120573 and tausynergistically impair the oxidative phosphorylation system intriple transgenic Alzheimerrsquos disease micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 47 pp 20057ndash20062 2009
[28] R X Santos S C Correia X Wang et al ldquoAlzheimerrsquos diseasediverse aspects ofmitochondrial malfunctioningrdquo InternationalJournal of Clinical and Experimental Pathology vol 3 no 6 pp570ndash581 2010
[29] N R Sims ldquoEnergy metabolism oxidative stress and neuronaldegeneration in Alzheimerrsquos diseaserdquoNeurodegeneration vol 5no 4 pp 435ndash440 1996
[30] H K Anandatheerthavarada G Biswas M-A Robin and NG Avadhani ldquoMitochondrial targeting and a novel transmem-brane arrest of Alzheimerrsquos amyloid precursor protein impairsmitochondrial function in neuronal cellsrdquo The Journal of CellBiology vol 161 no 1 pp 41ndash54 2003
[31] L Devi B M Prabhu D F Galati N G Avadhani and HK Anandatheerthavarada ldquoAccumulation of amyloid precur-sor protein in the mitochondrial import channels of humanAlzheimerrsquos disease brain is associated with mitochondrialdysfunctionrdquo The Journal of Neuroscience vol 26 no 35 pp9057ndash9068 2006
[32] L Devi and H K Anandatheerthavarada ldquoMitochondrialtrafficking of APP and alpha synuclein relevance to mito-chondrial dysfunction in Alzheimerrsquos and Parkinsonrsquos diseasesrdquoBiochimica et Biophysica Acta vol 1802 no 1 pp 11ndash19 2010
[33] M Manczak T S Anekonda E Henson B S Park J Quinnand P H Reddy ldquoMitochondria are a direct site of A120573 accumu-lation in Alzheimerrsquos disease neurons implications for free rad-ical generation and oxidative damage in disease progressionrdquoHuman Molecular Genetics vol 15 no 9 pp 1437ndash1449 2006
[34] P F Pavlov C H Petersen E Glaser and M AnkarcronaldquoMitochondrial accumulation of APP and A120573 significancefor Alzheimer disease pathogenesisrdquo Journal of Cellular andMolecular Medicine vol 13 no 10 pp 4137ndash4145 2009
[35] C Schmidt E Lepsverdize S L Chi et al ldquoAmyloid precursorprotein and amyloid 120573-peptide bind to ATP synthase andregulate its activity at the surface of neural cellsrdquo MolecularPsychiatry vol 13 no 10 pp 953ndash969 2008
[36] K Johar A Priya and M T T Wong-Riley ldquoRegulation ofNa+K+-ATPase by nuclear respiratory factor 1 implication inthe tight coupling of neuronal activity energy generation andenergy consumptionrdquo The Journal of Biological Chemistry vol287 no 48 pp 40381ndash40390 2012
[37] J Astrup P M Sorensen and H R Sorensen ldquoOxygen andglucose consumption related to Na+-K+ transport in caninebrainrdquo Stroke vol 12 no 6 pp 726ndash730 1981
[38] M Erecinska and I A Silver ldquoATP and brain functionrdquo Journalof Cerebral Blood Flow and Metabolism vol 9 no 1 pp 2ndash191989
[39] MMata D J Fink H Gainer et al ldquoActivity-dependent energymetabolism in rat posterior pituitary primarily reflects sodiumpump activityrdquo Journal of Neurochemistry vol 34 no 1 pp 213ndash215 1980
[40] J Kang H-G Lemaire A Unterbeck et al ldquoThe precursor ofAlzheimerrsquos disease amyloid A4 protein resembles a cell-surfacereceptorrdquo Nature vol 325 no 6106 pp 733ndash736 1987
[41] C L Masters and D J Selkoe ldquoBiochemistry of amyloid 120573-protein and amyloid deposits in Alzheimer diseaserdquoCold SpringHarbour Perspectives in Medicine vol 2 no 6 Article IDa006262 2012
[42] A Lai S S Sisodia and I S Trowbridge ldquoCharacterization ofsorting signals in the beta-amyloid precursor protein cytoplas-mic domainrdquo The Journal of Biological Chemistry vol 270 no8 pp 3565ndash3573 1995
[43] A I Placido C M F Pereira A I Duarte et al ldquoThe role ofendoplasmic reticulum in amyloid precursor protein processingand trafficking implications for Alzheimerrsquos diseaserdquo Biochim-ica et Biophysica ActamdashMolecular Basis of Disease vol 1842 no9 pp 1444ndash1453 2014
Oxidative Medicine and Cellular Longevity 13
[44] C Kaether P Skehel and C G Dotti ldquoAxonal membraneproteins are transported in distinct carriers a two-color videomicroscopy study in cultured hippocampal neuronsrdquoMolecularBiology of the Cell vol 11 no 4 pp 1213ndash1224 2000
[45] A Kamal A Almenar-Queralt J F LeBlanc E A Robertsand L S B Goldstein ldquoKinesin-mediated axonal transportof a membrane compartment containing 120573-secretase andpresenilin-1 requires APPrdquo Nature vol 414 no 6864 pp 643ndash648 2001
[46] R G Perez S Soriano J D Hayes et al ldquoMutagenesis identi-fies new signals for 120573-amyloid precursor protein endocytosisturnover and the generation of secreted fragments includingA12057342rdquo The Journal of Biological Chemistry vol 274 no 27 pp18851ndash18856 1999
[47] D G Cook M S Forman J C Sung et al ldquoAlzheimerrsquos A120573(1ndash42) is generated in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo Nature Medicine vol 3 no 9 pp1021ndash1023 1997
[48] A S C Chyung B D Greenberg D G Cook R W Domsand V M-Y Lee ldquoNovel 120573-secretase cleavage of 120573-amyloidprecursor protein in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo The Journal of Cell Biology vol138 no 3 pp 671ndash680 1997
[49] RW-Y Choy Z Cheng and R Schekman ldquoAmyloid precursorprotein (APP) traffics from the cell surface via endosomesfor amyloid 120573 (A120573) production in the trans-Golgi networkrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 30 pp E2077ndashE2082 2012
[50] S A Small and S Gandy ldquoSorting through the cell biologyof Alzheimerrsquos disease intracellular pathways to pathogenesisrdquoNeuron vol 52 no 1 pp 15ndash31 2006
[51] M Goedert ldquoAlzheimerrsquos and Parkinsonrsquos diseases the prionconcept in relation to assembled A120573 tau and 120572-synucleinrdquoScience vol 349 no 6248 2015
[52] I Benilova E Karran and B De Strooper ldquoThe toxic A120573oligomer and Alzheimerrsquos disease an emperor in need ofclothesrdquo Nature Neuroscience vol 15 no 3 pp 349ndash357 2012
[53] E H Corder A M Saunders W J Strittmatter et al ldquoGenedose of apolipoprotein E type 4 allele and the risk of Alzheimerrsquosdisease in late onset familiesrdquo Science vol 261 no 5123 pp 921ndash923 1993
[54] S Brunholz S Sisodia A Lorenzo C Deyts S Kins and GMorfini ldquoAxonal transport of APP and the spatial regulationof APP cleavage and function in neuronal cellsrdquo ExperimentalBrain Research vol 217 no 3-4 pp 353ndash364 2012
[55] G Thinakaran and E H Koo ldquoAmyloid precursor proteintrafficking processing and functionrdquo The Journal of BiologicalChemistry vol 283 no 44 pp 29615ndash29619 2008
[56] T Saitoh M Sundsmo J-M Roch et al ldquoSecreted formof amyloid beta protein precursor is involved in the growthregulation of fibroblastsrdquo Cell vol 58 no 4 pp 615ndash622 1989
[57] M PMattson B Cheng A R Culwell F S Esch I Lieberburgand R E Rydel ldquoEvidence for excitoprotective and intraneu-ronal calcium-regulating roles for secreted forms of the 120573-amyloid precursor proteinrdquo Neuron vol 10 no 2 pp 243ndash2541993
[58] K J Lee C E H Moussa Y Lee et al ldquoBeta amyloid-independent role of amyloid precursor protein in generationandmaintenance of dendritic spinesrdquoNeuroscience vol 169 no1 pp 344ndash356 2010
[59] H Meziane J-C Dodart C Mathis et al ldquoMemory-enhancingeffects of secreted forms of the 120573-amyloid precursor proteinin normal and amnestic micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no21 pp 12683ndash12688 1998
[60] P R Turner K OrsquoConnor W P Tate and W C AbrahamldquoRoles of amyloid precursor protein and its fragments inregulating neural activity plasticity and memoryrdquo Progress inNeurobiology vol 70 no 1 pp 1ndash32 2003
[61] D Beher L Hesse C L Masters and GMulthaup ldquoRegulationof Amyloid Protein Precursor (APP) binding to collagen andmapping of the binding sites on APP and collagen type Irdquo TheJournal of Biological Chemistry vol 271 no 3 pp 1613ndash16201996
[62] S L Sabo A F Ikin J D Buxbaum and P Greengard ldquoTheamyloid precursor protein and its regulatory protein FE65 ingrowth cones and synapses in vitro and in vivordquo The Journal ofNeuroscience vol 23 no 13 pp 5407ndash5415 2003
[63] D H Small V Nurcombe G Reed et al ldquoA heparin-bindingdomain in the amyloid protein precursor of Alzheimerrsquos diseaseis involved in the regulation of neurite outgrowthrdquoThe Journalof Neuroscience vol 14 no 4 pp 2117ndash2127 1994
[64] T G Williamson S S Mok A Henry et al ldquoSecreted glypicanbinds to the amyloid precursor protein of Alzheimerrsquos disease(APP) and inhibits APP-induced neurite outgrowthrdquoThe Jour-nal of Biological Chemistry vol 271 no 49 pp 31215ndash312211996
[65] T Ondrejcak I Klyubin N-W Hu A E Barry W K Cullenand M J Rowan ldquoAlzheimerrsquos disease amyloid 120573-protein andsynaptic functionrdquo NeuroMolecular Medicine vol 12 no 1 pp13ndash26 2010
[66] D Puzzo L Privitera and A Palmeri ldquoHormetic effect ofamyloid-beta peptide in synaptic plasticity and memoryrdquo Neu-robiology of Aging vol 33 no 7 pp 1484e15ndash1484e24 2012
[67] C M F Pereira ldquoCrosstalk between endoplasmic reticulumstress and protein misfolding in neurodegenerative diseasesrdquoISRN Cell Biology vol 2013 Article ID 256404 22 pages 2013
[68] R-W Shin T C Saido M Maeda and T Kitamoto ldquoNovel 120572-secretase cleavage ofAlzheimerrsquos amyloid120573 precursor protein inthe endoplasmic reticulum of COS7 cellsrdquoNeuroscience Lettersvol 376 no 1 pp 14ndash19 2005
[69] E A Schon and E Area-Gomez ldquoMitochondria-associatedER membranes in Alzheimer diseaserdquo Molecular and CellularNeuroscience vol 55 pp 26ndash36 2013
[70] R Rizzuto P Pinton W Carrington et al ldquoClose contacts withthe endoplasmic reticulum as determinants of mitochondrialCa2+ responsesrdquo Science vol 280 no 5370 pp 1763ndash1766 1998
[71] S E Logue P Cleary S Saveljeva and A Samali ldquoNewdirections in ER-stress induced cell deathrdquoApoptosis vol 18 no5 pp 537ndash546 2013
[72] J P Decuypere KWelkenhuyzen T Luyten et al ldquoIns(145)P3receptor-mediated BI-1 Ca2+ signaling and autophagy induc-tion are interrelatedrdquo Autophagy vol 7 pp 1472ndash1489 2011
[73] K S Vetrivel and G Thinakaran ldquoMembrane rafts in Alz-heimerrsquos disease beta-amyloid productionrdquo Biochimica et Bio-physica Acta vol 1801 no 8 pp 860ndash867 2010
[74] H Urra E Dufey F Lisbona D Rojas-Rivera and C HetzldquoWhen ER stress reaches a dead endrdquo Biochimica et BiophysicaActa (BBA) Molecular Cell Research vol 1833 no 12 pp 3507ndash3517 2013
14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 Oxidative Medicine and Cellular Longevity
[14] A Eckert K Schmitt and J Gotz ldquoMitochondrial dysfunc-tionmdashthe beginning of the end in Alzheimerrsquos disease Sep-arate and synergistic modes of tau and amyloid-120573 toxicityrdquoAlzheimerrsquos Research ampTherapy vol 3 article 15 pp 1ndash11 2011
[15] U Keil A Bonert C A Marques et al ldquoAmyloid 120573-inducedchanges in nitric oxide production and mitochondrial activitylead to apoptosisrdquoThe Journal of Biological Chemistry vol 279no 48 pp 50310ndash50320 2004
[16] A M Swomley and D A Butterfield ldquoOxidative stress inAlzheimer disease and mild cognitive impairment evidencefrom human data provided by redox proteomicsrdquo Archives ofToxicology vol 89 no 10 pp 1669ndash1680 2015
[17] C Caspersen N Wang J Yao et al ldquoMitochondrial A120573 apotential focal point for neuronal metabolic dysfunction inAlzheimerrsquos diseaserdquo The FASEB Journal vol 19 no 14 pp2040ndash2041 2005
[18] G E Gibson K-F R Sheu and J P Blass ldquoAbnormalities ofmitochondrial enzymes inAlzheimer diseaserdquo Journal of NeuralTransmission vol 105 no 8-9 pp 855ndash870 1998
[19] A Eckert S Hauptmann I Scherping et al ldquoSoluble beta-amyloid leads to mitochondrial defects in amyloid precursorprotein and tau transgenic micerdquo Neurodegenerative Diseasesvol 5 no 3-4 pp 157ndash159 2008
[20] A Eckert K L Schulz V Rhein and J Gotz ldquoConvergenceof amyloid-120573 and tau pathologies on mitochondria in vivordquoMolecular Neurobiology vol 41 no 2-3 pp 107ndash114 2010
[21] S Hauptmann I Scherping S Drose et al ldquoMitochondrial dys-function an early event in Alzheimer pathology accumulateswith age in AD transgenic micerdquoNeurobiology of Aging vol 30no 10 pp 1574ndash1586 2009
[22] M Manczak B S Park Y Jung and P H Reddy ldquoDifferentialexpression of oxidative phosphorylation genes in patients withAlzheimerrsquos disease implications for early mitochondrial dys-function and oxidative damagerdquo NeuroMolecular Medicine vol5 no 2 pp 147ndash162 2004
[23] M P Mattson M Gleichmann and A Cheng ldquoMitochondriain neuroplasticity and neurological disordersrdquo Neuron vol 60no 5 pp 748ndash766 2008
[24] P I Moreira M S Santos and C R Oliveira ldquoAlzheimerrsquosdisease a lesson frommitochondrial dysfunctionrdquoAntioxidantsamp Redox Signaling vol 9 no 10 pp 1621ndash1630 2007
[25] P H Reddy ldquoMitochondrial dysfunction in aging and Alz-heimerrsquos disease strategies to protect neuronsrdquo Antioxidants ampRedox Signaling vol 9 no 10 pp 1647ndash1658 2007
[26] V Rhein G Baysang S Rao et al ldquoAmyloid-beta leads toimpaired cellular respiration energy production andmitochon-drial electron chain complex activities in humanneuroblastomacellsrdquo Cellular and Molecular Neurobiology vol 29 no 6-7 pp1063ndash1071 2009
[27] V Rhein X Song A Wiesner et al ldquoAmyloid-120573 and tausynergistically impair the oxidative phosphorylation system intriple transgenic Alzheimerrsquos disease micerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 47 pp 20057ndash20062 2009
[28] R X Santos S C Correia X Wang et al ldquoAlzheimerrsquos diseasediverse aspects ofmitochondrial malfunctioningrdquo InternationalJournal of Clinical and Experimental Pathology vol 3 no 6 pp570ndash581 2010
[29] N R Sims ldquoEnergy metabolism oxidative stress and neuronaldegeneration in Alzheimerrsquos diseaserdquoNeurodegeneration vol 5no 4 pp 435ndash440 1996
[30] H K Anandatheerthavarada G Biswas M-A Robin and NG Avadhani ldquoMitochondrial targeting and a novel transmem-brane arrest of Alzheimerrsquos amyloid precursor protein impairsmitochondrial function in neuronal cellsrdquo The Journal of CellBiology vol 161 no 1 pp 41ndash54 2003
[31] L Devi B M Prabhu D F Galati N G Avadhani and HK Anandatheerthavarada ldquoAccumulation of amyloid precur-sor protein in the mitochondrial import channels of humanAlzheimerrsquos disease brain is associated with mitochondrialdysfunctionrdquo The Journal of Neuroscience vol 26 no 35 pp9057ndash9068 2006
[32] L Devi and H K Anandatheerthavarada ldquoMitochondrialtrafficking of APP and alpha synuclein relevance to mito-chondrial dysfunction in Alzheimerrsquos and Parkinsonrsquos diseasesrdquoBiochimica et Biophysica Acta vol 1802 no 1 pp 11ndash19 2010
[33] M Manczak T S Anekonda E Henson B S Park J Quinnand P H Reddy ldquoMitochondria are a direct site of A120573 accumu-lation in Alzheimerrsquos disease neurons implications for free rad-ical generation and oxidative damage in disease progressionrdquoHuman Molecular Genetics vol 15 no 9 pp 1437ndash1449 2006
[34] P F Pavlov C H Petersen E Glaser and M AnkarcronaldquoMitochondrial accumulation of APP and A120573 significancefor Alzheimer disease pathogenesisrdquo Journal of Cellular andMolecular Medicine vol 13 no 10 pp 4137ndash4145 2009
[35] C Schmidt E Lepsverdize S L Chi et al ldquoAmyloid precursorprotein and amyloid 120573-peptide bind to ATP synthase andregulate its activity at the surface of neural cellsrdquo MolecularPsychiatry vol 13 no 10 pp 953ndash969 2008
[36] K Johar A Priya and M T T Wong-Riley ldquoRegulation ofNa+K+-ATPase by nuclear respiratory factor 1 implication inthe tight coupling of neuronal activity energy generation andenergy consumptionrdquo The Journal of Biological Chemistry vol287 no 48 pp 40381ndash40390 2012
[37] J Astrup P M Sorensen and H R Sorensen ldquoOxygen andglucose consumption related to Na+-K+ transport in caninebrainrdquo Stroke vol 12 no 6 pp 726ndash730 1981
[38] M Erecinska and I A Silver ldquoATP and brain functionrdquo Journalof Cerebral Blood Flow and Metabolism vol 9 no 1 pp 2ndash191989
[39] MMata D J Fink H Gainer et al ldquoActivity-dependent energymetabolism in rat posterior pituitary primarily reflects sodiumpump activityrdquo Journal of Neurochemistry vol 34 no 1 pp 213ndash215 1980
[40] J Kang H-G Lemaire A Unterbeck et al ldquoThe precursor ofAlzheimerrsquos disease amyloid A4 protein resembles a cell-surfacereceptorrdquo Nature vol 325 no 6106 pp 733ndash736 1987
[41] C L Masters and D J Selkoe ldquoBiochemistry of amyloid 120573-protein and amyloid deposits in Alzheimer diseaserdquoCold SpringHarbour Perspectives in Medicine vol 2 no 6 Article IDa006262 2012
[42] A Lai S S Sisodia and I S Trowbridge ldquoCharacterization ofsorting signals in the beta-amyloid precursor protein cytoplas-mic domainrdquo The Journal of Biological Chemistry vol 270 no8 pp 3565ndash3573 1995
[43] A I Placido C M F Pereira A I Duarte et al ldquoThe role ofendoplasmic reticulum in amyloid precursor protein processingand trafficking implications for Alzheimerrsquos diseaserdquo Biochim-ica et Biophysica ActamdashMolecular Basis of Disease vol 1842 no9 pp 1444ndash1453 2014
Oxidative Medicine and Cellular Longevity 13
[44] C Kaether P Skehel and C G Dotti ldquoAxonal membraneproteins are transported in distinct carriers a two-color videomicroscopy study in cultured hippocampal neuronsrdquoMolecularBiology of the Cell vol 11 no 4 pp 1213ndash1224 2000
[45] A Kamal A Almenar-Queralt J F LeBlanc E A Robertsand L S B Goldstein ldquoKinesin-mediated axonal transportof a membrane compartment containing 120573-secretase andpresenilin-1 requires APPrdquo Nature vol 414 no 6864 pp 643ndash648 2001
[46] R G Perez S Soriano J D Hayes et al ldquoMutagenesis identi-fies new signals for 120573-amyloid precursor protein endocytosisturnover and the generation of secreted fragments includingA12057342rdquo The Journal of Biological Chemistry vol 274 no 27 pp18851ndash18856 1999
[47] D G Cook M S Forman J C Sung et al ldquoAlzheimerrsquos A120573(1ndash42) is generated in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo Nature Medicine vol 3 no 9 pp1021ndash1023 1997
[48] A S C Chyung B D Greenberg D G Cook R W Domsand V M-Y Lee ldquoNovel 120573-secretase cleavage of 120573-amyloidprecursor protein in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo The Journal of Cell Biology vol138 no 3 pp 671ndash680 1997
[49] RW-Y Choy Z Cheng and R Schekman ldquoAmyloid precursorprotein (APP) traffics from the cell surface via endosomesfor amyloid 120573 (A120573) production in the trans-Golgi networkrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 30 pp E2077ndashE2082 2012
[50] S A Small and S Gandy ldquoSorting through the cell biologyof Alzheimerrsquos disease intracellular pathways to pathogenesisrdquoNeuron vol 52 no 1 pp 15ndash31 2006
[51] M Goedert ldquoAlzheimerrsquos and Parkinsonrsquos diseases the prionconcept in relation to assembled A120573 tau and 120572-synucleinrdquoScience vol 349 no 6248 2015
[52] I Benilova E Karran and B De Strooper ldquoThe toxic A120573oligomer and Alzheimerrsquos disease an emperor in need ofclothesrdquo Nature Neuroscience vol 15 no 3 pp 349ndash357 2012
[53] E H Corder A M Saunders W J Strittmatter et al ldquoGenedose of apolipoprotein E type 4 allele and the risk of Alzheimerrsquosdisease in late onset familiesrdquo Science vol 261 no 5123 pp 921ndash923 1993
[54] S Brunholz S Sisodia A Lorenzo C Deyts S Kins and GMorfini ldquoAxonal transport of APP and the spatial regulationof APP cleavage and function in neuronal cellsrdquo ExperimentalBrain Research vol 217 no 3-4 pp 353ndash364 2012
[55] G Thinakaran and E H Koo ldquoAmyloid precursor proteintrafficking processing and functionrdquo The Journal of BiologicalChemistry vol 283 no 44 pp 29615ndash29619 2008
[56] T Saitoh M Sundsmo J-M Roch et al ldquoSecreted formof amyloid beta protein precursor is involved in the growthregulation of fibroblastsrdquo Cell vol 58 no 4 pp 615ndash622 1989
[57] M PMattson B Cheng A R Culwell F S Esch I Lieberburgand R E Rydel ldquoEvidence for excitoprotective and intraneu-ronal calcium-regulating roles for secreted forms of the 120573-amyloid precursor proteinrdquo Neuron vol 10 no 2 pp 243ndash2541993
[58] K J Lee C E H Moussa Y Lee et al ldquoBeta amyloid-independent role of amyloid precursor protein in generationandmaintenance of dendritic spinesrdquoNeuroscience vol 169 no1 pp 344ndash356 2010
[59] H Meziane J-C Dodart C Mathis et al ldquoMemory-enhancingeffects of secreted forms of the 120573-amyloid precursor proteinin normal and amnestic micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no21 pp 12683ndash12688 1998
[60] P R Turner K OrsquoConnor W P Tate and W C AbrahamldquoRoles of amyloid precursor protein and its fragments inregulating neural activity plasticity and memoryrdquo Progress inNeurobiology vol 70 no 1 pp 1ndash32 2003
[61] D Beher L Hesse C L Masters and GMulthaup ldquoRegulationof Amyloid Protein Precursor (APP) binding to collagen andmapping of the binding sites on APP and collagen type Irdquo TheJournal of Biological Chemistry vol 271 no 3 pp 1613ndash16201996
[62] S L Sabo A F Ikin J D Buxbaum and P Greengard ldquoTheamyloid precursor protein and its regulatory protein FE65 ingrowth cones and synapses in vitro and in vivordquo The Journal ofNeuroscience vol 23 no 13 pp 5407ndash5415 2003
[63] D H Small V Nurcombe G Reed et al ldquoA heparin-bindingdomain in the amyloid protein precursor of Alzheimerrsquos diseaseis involved in the regulation of neurite outgrowthrdquoThe Journalof Neuroscience vol 14 no 4 pp 2117ndash2127 1994
[64] T G Williamson S S Mok A Henry et al ldquoSecreted glypicanbinds to the amyloid precursor protein of Alzheimerrsquos disease(APP) and inhibits APP-induced neurite outgrowthrdquoThe Jour-nal of Biological Chemistry vol 271 no 49 pp 31215ndash312211996
[65] T Ondrejcak I Klyubin N-W Hu A E Barry W K Cullenand M J Rowan ldquoAlzheimerrsquos disease amyloid 120573-protein andsynaptic functionrdquo NeuroMolecular Medicine vol 12 no 1 pp13ndash26 2010
[66] D Puzzo L Privitera and A Palmeri ldquoHormetic effect ofamyloid-beta peptide in synaptic plasticity and memoryrdquo Neu-robiology of Aging vol 33 no 7 pp 1484e15ndash1484e24 2012
[67] C M F Pereira ldquoCrosstalk between endoplasmic reticulumstress and protein misfolding in neurodegenerative diseasesrdquoISRN Cell Biology vol 2013 Article ID 256404 22 pages 2013
[68] R-W Shin T C Saido M Maeda and T Kitamoto ldquoNovel 120572-secretase cleavage ofAlzheimerrsquos amyloid120573 precursor protein inthe endoplasmic reticulum of COS7 cellsrdquoNeuroscience Lettersvol 376 no 1 pp 14ndash19 2005
[69] E A Schon and E Area-Gomez ldquoMitochondria-associatedER membranes in Alzheimer diseaserdquo Molecular and CellularNeuroscience vol 55 pp 26ndash36 2013
[70] R Rizzuto P Pinton W Carrington et al ldquoClose contacts withthe endoplasmic reticulum as determinants of mitochondrialCa2+ responsesrdquo Science vol 280 no 5370 pp 1763ndash1766 1998
[71] S E Logue P Cleary S Saveljeva and A Samali ldquoNewdirections in ER-stress induced cell deathrdquoApoptosis vol 18 no5 pp 537ndash546 2013
[72] J P Decuypere KWelkenhuyzen T Luyten et al ldquoIns(145)P3receptor-mediated BI-1 Ca2+ signaling and autophagy induc-tion are interrelatedrdquo Autophagy vol 7 pp 1472ndash1489 2011
[73] K S Vetrivel and G Thinakaran ldquoMembrane rafts in Alz-heimerrsquos disease beta-amyloid productionrdquo Biochimica et Bio-physica Acta vol 1801 no 8 pp 860ndash867 2010
[74] H Urra E Dufey F Lisbona D Rojas-Rivera and C HetzldquoWhen ER stress reaches a dead endrdquo Biochimica et BiophysicaActa (BBA) Molecular Cell Research vol 1833 no 12 pp 3507ndash3517 2013
14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 13
[44] C Kaether P Skehel and C G Dotti ldquoAxonal membraneproteins are transported in distinct carriers a two-color videomicroscopy study in cultured hippocampal neuronsrdquoMolecularBiology of the Cell vol 11 no 4 pp 1213ndash1224 2000
[45] A Kamal A Almenar-Queralt J F LeBlanc E A Robertsand L S B Goldstein ldquoKinesin-mediated axonal transportof a membrane compartment containing 120573-secretase andpresenilin-1 requires APPrdquo Nature vol 414 no 6864 pp 643ndash648 2001
[46] R G Perez S Soriano J D Hayes et al ldquoMutagenesis identi-fies new signals for 120573-amyloid precursor protein endocytosisturnover and the generation of secreted fragments includingA12057342rdquo The Journal of Biological Chemistry vol 274 no 27 pp18851ndash18856 1999
[47] D G Cook M S Forman J C Sung et al ldquoAlzheimerrsquos A120573(1ndash42) is generated in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo Nature Medicine vol 3 no 9 pp1021ndash1023 1997
[48] A S C Chyung B D Greenberg D G Cook R W Domsand V M-Y Lee ldquoNovel 120573-secretase cleavage of 120573-amyloidprecursor protein in the endoplasmic reticulumintermediatecompartment of NT2N cellsrdquo The Journal of Cell Biology vol138 no 3 pp 671ndash680 1997
[49] RW-Y Choy Z Cheng and R Schekman ldquoAmyloid precursorprotein (APP) traffics from the cell surface via endosomesfor amyloid 120573 (A120573) production in the trans-Golgi networkrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 30 pp E2077ndashE2082 2012
[50] S A Small and S Gandy ldquoSorting through the cell biologyof Alzheimerrsquos disease intracellular pathways to pathogenesisrdquoNeuron vol 52 no 1 pp 15ndash31 2006
[51] M Goedert ldquoAlzheimerrsquos and Parkinsonrsquos diseases the prionconcept in relation to assembled A120573 tau and 120572-synucleinrdquoScience vol 349 no 6248 2015
[52] I Benilova E Karran and B De Strooper ldquoThe toxic A120573oligomer and Alzheimerrsquos disease an emperor in need ofclothesrdquo Nature Neuroscience vol 15 no 3 pp 349ndash357 2012
[53] E H Corder A M Saunders W J Strittmatter et al ldquoGenedose of apolipoprotein E type 4 allele and the risk of Alzheimerrsquosdisease in late onset familiesrdquo Science vol 261 no 5123 pp 921ndash923 1993
[54] S Brunholz S Sisodia A Lorenzo C Deyts S Kins and GMorfini ldquoAxonal transport of APP and the spatial regulationof APP cleavage and function in neuronal cellsrdquo ExperimentalBrain Research vol 217 no 3-4 pp 353ndash364 2012
[55] G Thinakaran and E H Koo ldquoAmyloid precursor proteintrafficking processing and functionrdquo The Journal of BiologicalChemistry vol 283 no 44 pp 29615ndash29619 2008
[56] T Saitoh M Sundsmo J-M Roch et al ldquoSecreted formof amyloid beta protein precursor is involved in the growthregulation of fibroblastsrdquo Cell vol 58 no 4 pp 615ndash622 1989
[57] M PMattson B Cheng A R Culwell F S Esch I Lieberburgand R E Rydel ldquoEvidence for excitoprotective and intraneu-ronal calcium-regulating roles for secreted forms of the 120573-amyloid precursor proteinrdquo Neuron vol 10 no 2 pp 243ndash2541993
[58] K J Lee C E H Moussa Y Lee et al ldquoBeta amyloid-independent role of amyloid precursor protein in generationandmaintenance of dendritic spinesrdquoNeuroscience vol 169 no1 pp 344ndash356 2010
[59] H Meziane J-C Dodart C Mathis et al ldquoMemory-enhancingeffects of secreted forms of the 120573-amyloid precursor proteinin normal and amnestic micerdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 95 no21 pp 12683ndash12688 1998
[60] P R Turner K OrsquoConnor W P Tate and W C AbrahamldquoRoles of amyloid precursor protein and its fragments inregulating neural activity plasticity and memoryrdquo Progress inNeurobiology vol 70 no 1 pp 1ndash32 2003
[61] D Beher L Hesse C L Masters and GMulthaup ldquoRegulationof Amyloid Protein Precursor (APP) binding to collagen andmapping of the binding sites on APP and collagen type Irdquo TheJournal of Biological Chemistry vol 271 no 3 pp 1613ndash16201996
[62] S L Sabo A F Ikin J D Buxbaum and P Greengard ldquoTheamyloid precursor protein and its regulatory protein FE65 ingrowth cones and synapses in vitro and in vivordquo The Journal ofNeuroscience vol 23 no 13 pp 5407ndash5415 2003
[63] D H Small V Nurcombe G Reed et al ldquoA heparin-bindingdomain in the amyloid protein precursor of Alzheimerrsquos diseaseis involved in the regulation of neurite outgrowthrdquoThe Journalof Neuroscience vol 14 no 4 pp 2117ndash2127 1994
[64] T G Williamson S S Mok A Henry et al ldquoSecreted glypicanbinds to the amyloid precursor protein of Alzheimerrsquos disease(APP) and inhibits APP-induced neurite outgrowthrdquoThe Jour-nal of Biological Chemistry vol 271 no 49 pp 31215ndash312211996
[65] T Ondrejcak I Klyubin N-W Hu A E Barry W K Cullenand M J Rowan ldquoAlzheimerrsquos disease amyloid 120573-protein andsynaptic functionrdquo NeuroMolecular Medicine vol 12 no 1 pp13ndash26 2010
[66] D Puzzo L Privitera and A Palmeri ldquoHormetic effect ofamyloid-beta peptide in synaptic plasticity and memoryrdquo Neu-robiology of Aging vol 33 no 7 pp 1484e15ndash1484e24 2012
[67] C M F Pereira ldquoCrosstalk between endoplasmic reticulumstress and protein misfolding in neurodegenerative diseasesrdquoISRN Cell Biology vol 2013 Article ID 256404 22 pages 2013
[68] R-W Shin T C Saido M Maeda and T Kitamoto ldquoNovel 120572-secretase cleavage ofAlzheimerrsquos amyloid120573 precursor protein inthe endoplasmic reticulum of COS7 cellsrdquoNeuroscience Lettersvol 376 no 1 pp 14ndash19 2005
[69] E A Schon and E Area-Gomez ldquoMitochondria-associatedER membranes in Alzheimer diseaserdquo Molecular and CellularNeuroscience vol 55 pp 26ndash36 2013
[70] R Rizzuto P Pinton W Carrington et al ldquoClose contacts withthe endoplasmic reticulum as determinants of mitochondrialCa2+ responsesrdquo Science vol 280 no 5370 pp 1763ndash1766 1998
[71] S E Logue P Cleary S Saveljeva and A Samali ldquoNewdirections in ER-stress induced cell deathrdquoApoptosis vol 18 no5 pp 537ndash546 2013
[72] J P Decuypere KWelkenhuyzen T Luyten et al ldquoIns(145)P3receptor-mediated BI-1 Ca2+ signaling and autophagy induc-tion are interrelatedrdquo Autophagy vol 7 pp 1472ndash1489 2011
[73] K S Vetrivel and G Thinakaran ldquoMembrane rafts in Alz-heimerrsquos disease beta-amyloid productionrdquo Biochimica et Bio-physica Acta vol 1801 no 8 pp 860ndash867 2010
[74] H Urra E Dufey F Lisbona D Rojas-Rivera and C HetzldquoWhen ER stress reaches a dead endrdquo Biochimica et BiophysicaActa (BBA) Molecular Cell Research vol 1833 no 12 pp 3507ndash3517 2013
14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

14 Oxidative Medicine and Cellular Longevity
[75] G Li M Mongillo K-T Chin et al ldquoRole of ERO1-120572-mediatedstimulation of inositol 145-triphosphate receptor activity inendoplasmic reticulum stress-induced apoptosisrdquo The Journalof Cell Biology vol 186 no 6 pp 783ndash792 2009
[76] K Yamamoto R Fujii Y Toyofuku et al ldquoThe KDEL receptormediates a retrieval mechanism that contributes to qualitycontrol at the endoplasmic reticulumrdquoThe EMBO Journal vol20 no 12 pp 3082ndash3091 2001
[77] TKatayamaK ImaizumiN Sato et al ldquoPresenilin-1mutationsdownregulate the signalling pathway of the unfolded-proteinresponserdquo Nature Cell Biology vol 1 no 8 pp 479ndash485 1999
[78] C Ballatore VM-Y Lee and J Q Trojanowski ldquoTau-mediatedneurodegeneration in Alzheimerrsquos disease and related disor-dersrdquo Nature Reviews Neuroscience vol 8 no 9 pp 663ndash6722007
[79] B Frost R L Jacks and M I Diamond ldquoPropagation of Taumisfolding from the outside to the inside of a cellrdquoThe Journalof Biological Chemistry vol 284 no 19 pp 12845ndash12852 2009
[80] T Nonaka S T Watanabe T Iwatsubo and M HasegawaldquoSeeded aggregation and toxicity of 120572-synuclein and tau cellu-lar models of neurodegenerative diseasesrdquo Journal of BiologicalChemistry vol 285 no 45 pp 34885ndash34898 2010
[81] J F Crary J Q Trojanowski J A Schneider et al ldquoPrimary age-related tauopathy (PART) a commonpathology associatedwithhuman agingrdquo Acta Neuropathologica vol 128 no 6 pp 755ndash766 2014
[82] C R Jack Jr D S KnopmanW J Jagust et al ldquoTracking patho-physiological processes in Alzheimerrsquos disease an updatedhypothetical model of dynamic biomarkersrdquo The Lancet Neu-rology vol 12 no 2 pp 207ndash216 2013
[83] A Serrano-Pozo M P Frosch E Masliah and B T HymanldquoNeuropathological alterations in Alzheimer diseaserdquo ColdSpring Harbor Perspectives in Medicine vol 1 no 1 pp 1ndash232011
[84] E Ferreiro and C M F Pereira ldquoEndoplasmic reticulum stressa new playER in tauopathiesrdquo Journal of Pathology vol 226 no5 pp 687ndash692 2012
[85] J J M Hoozemans and W Scheper ldquoEndoplasmic reticulumthe unfolded protein response is tangled in neurodegenerationrdquoInternational Journal of Biochemistry amp Cell Biology vol 44 no8 pp 1295ndash1298 2012
[86] J J M Hoozemans E S van Haastert D A T Nijholt A JM Rozemuller P Eikelenboom andW Scheper ldquoThe unfoldedprotein response is activated in pretangle neurons in alzheimerrsquosdisease hippocampusrdquo The American Journal of Pathology vol174 no 4 pp 1241ndash1251 2009
[87] Y-S Ho X Yang J C-F Lau et al ldquoEndoplasmic reticulumstress induces tau pathology and forms a vicious cycle implica-tion in Alzheimerrsquos disease pathogenesisrdquo Journal of AlzheimerrsquosDisease vol 28 no 4 pp 839ndash854 2012
[88] Z-Q Fu Y Yang J Song et al ldquoLiCl attenuates thapsigargin-induced tau hyperphosphorylation by inhibiting GSK-3120573 invivo and in vitrordquo Journal of Alzheimerrsquos Disease vol 21 no 4pp 1107ndash1117 2010
[89] B Schenk F Fernandez and C J Waechter ldquoThe ins(die) andout(side) of dolichyl phosphate biosynthesis and recycling inthe endoplasmic reticulumrdquo Glycobiology vol 11 pp 61Rndash67R2001
[90] A Helenius and M Aebi ldquoIntracellular functions of N-linkedglycansrdquo Science vol 291 no 5512 pp 2364ndash2369 2001
[91] J Hitomi T Katayama Y Eguchi et al ldquoInvolvement ofcaspase-4 in endoplasmic reticulum stress-induced apoptosisand A120573-induced cell deathrdquoThe Journal of Cell Biology vol 165no 3 pp 347ndash356 2004
[92] N Morishima K Nakanishi H Takenouchi T Shibata andY Yasuhiko ldquoAn endoplasmic reticulum stress-specific caspasecascade in apoptosis Cytochrome c-independent activation ofcaspase-9 by caspase-12rdquo The Journal of Biological Chemistryvol 277 no 37 pp 34287ndash34294 2002
[93] D Rodriguez D Rojas-Rivera and C Hetz ldquoIntegrating stresssignals at the endoplasmic reticulum the BCL-2 protein familyrheostatrdquoBiochimica et BiophysicaActa vol 1813 no 4 pp 564ndash574 2011
[94] S Y Gilady M Bui E M Lynes et al ldquoEro1120572 requires oxidizingand normoxic conditions to localize to the mitochondria-associated membrane (MAM)rdquo Cell Stress and Chaperones vol15 no 5 pp 619ndash629 2010
[95] SWang andW S El-Deiry ldquoCytochrome c a crosslink betweenthe mitochondria and the endoplasmic reticulum in calcium-dependent apoptosisrdquo Cancer Biology andTherapy vol 3 no 1pp 44ndash46 2004
[96] I Tabas and D Ron ldquoIntegrating the mechanisms of apoptosisinduced by endoplasmic reticulum stressrdquo Nature Cell Biologyvol 13 no 3 pp 184ndash190 2011
[97] T Yoneda K Imaizumi K Oono et al ldquoActivation of Caspase-12 an endoplastic reticulum (ER) resident caspase throughtumor necrosis factor receptor-associated factor 2-dependentmechanism in response to the ER stressrdquo The Journal ofBiological Chemistry vol 276 no 17 pp 13935ndash13940 2001
[98] A Ebneth G Drewes E-M Mandelkow and E MandelkowldquoPhosphorylation ofMAP2c andMAP4byMARKkinases leadsto the destabilization of microtubules in cellsrdquo Cell Motility andthe Cytoskeleton vol 44 no 3 pp 209ndash224 1999
[99] J Gotz L M Ittner M Fandrich and N Schonrock ldquoIstau aggregation toxic or protective a sensible question in theabsence of sensitive methodsrdquo Journal of Alzheimerrsquos Diseasevol 14 no 4 pp 423ndash429 2008
[100] L M Ittner Y D Ke F Delerue et al ldquoDendritic function oftau mediates amyloid-120573 toxicity in Alzheimerrsquos disease mousemodelsrdquo Cell vol 142 no 3 pp 387ndash397 2010
[101] K Stamer R Vogel E Thies E Mandelkow and E-MMandelkow ldquoTau blocks traffic of organelles neurofilamentsand APP vesicles in neurons and enhances oxidative stressrdquoTheJournal of Cell Biology vol 156 no 6 pp 1051ndash1063 2002
[102] C S Casley L Canevari J M Land J B Clark and M ASharpe ldquo120573-Amyloid inhibits integrated mitochondrial respira-tion and key enzyme activitiesrdquo Journal of Neurochemistry vol80 no 1 pp 91ndash100 2002
[103] C S Casley J M LandM A Sharpe J B ClarkM R Duchenand L Canevari ldquo120573-Amyloid fragment 25ndash35 causes mitochon-drial dysfunction in primary cortical neuronsrdquo Neurobiology ofDisease vol 10 no 3 pp 258ndash267 2002
[104] S Sorbi E D Bird and J P Blass ldquoDecreased pyruvatedehydrogenase complex activity in Huntington and Alzheimerbrainrdquo Annals of Neurology vol 13 no 1 pp 72ndash78 1983
[105] K Takahashi T Niidome A Akaike T Kihara and H Sugi-moto ldquoAmyloid precursor protein promotes endoplasmic retic-ulum stress-induced cell death via CEBP homologous protein-mediated pathwayrdquo Journal of Neurochemistry vol 109 no 5pp 1324ndash1337 2009
Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 15
[106] D Kogel C G Concannon T Muller et al ldquoThe APP intracel-lular domain (AICD) potentiates ER stress-induced apoptosisrdquoNeurobiology of Aging vol 33 no 9 pp 2200ndash2209 2012
[107] C A Hansson Petersen N Alikhani H Behbahani et alldquoThe amyloid 120573-peptide is imported into mitochondria viathe TOM import machinery and localized to mitochondrialcristaerdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 35 pp 13145ndash13150 2008
[108] H K Anandatheerthavarada N B V Sepuri G Biswas and NG Avadhani ldquoAn unusual TOM20TOM22 bypass mechanismfor the mitochondrial targeting of cytochrome P450 proteinscontaining n-terminal chimeric signalsrdquoThe Journal of Biologi-cal Chemistry vol 283 no 28 pp 19769ndash19780 2008
[109] M-A Robin H K Anandatheerthavarada G Biswas et alldquoBimodal targeting of microsomal CYP2E1 to mitochondriathrough activation of an N-terminal chimeric signal by cAMP-mediated phosphorylationrdquoThe Journal of Biological Chemistryvol 277 no 43 pp 40583ndash40593 2002
[110] J Zha H Harada E Yang J Jockel and S J Korsmeyer ldquoSerinephosphorylation of death agonist BAD in response to survivalfactor results in binding to 14-3-3 not BCL-X(L)rdquo Cell vol 87no 4 pp 619ndash628 1996
[111] C A Hansson S Frykman and M R Farmery ldquoNicastrinpresenilin APH-1 and PEN-2 form active gamma-secretasecomplexes in mitochondriardquo The Journal of Biological Chem-istry vol 279 pp 51654ndash51660 2004
[112] D Narendra A Tanaka D-F Suen and R J Youle ldquoParkin isrecruited selectively to impaired mitochondria and promotestheir autophagyrdquoThe Journal of Cell Biology vol 183 no 5 pp795ndash803 2008
[113] A Nechushtan C L Smith Y-T Hsu and R J Youle ldquoCon-formation of the Bax Cminusterminus regulates subcellular locationand cell deathrdquoThe EMBO Journal vol 18 no 9 pp 2330ndash23411999
[114] M Suzuki R J Youle and N Tjandra ldquoStructure of bax coreg-ulation of dimer formation and intracellular localizationrdquo Cellvol 103 no 4 pp 645ndash654 2000
[115] B Y Sheng Y Niu H Zhou et al ldquoThemitochondrial functionwas impaired in APP knockout mouse embryo fibroblast cellsrdquoChinese Science Bulletin vol 54 no 10 pp 1725ndash1731 2009
[116] K Chandrasekaran K Hatanpaa D R Brady J Stoll and SI Rapoport ldquoDown regulation of oxidative phosphorylation inAlzheimer disease loss of cytochrome oxidase subunit mRNAin the hippocampus and entorhinal cortexrdquo Brain Research vol796 no 1-2 pp 13ndash19 1998
[117] I Maurer S Zierz and H-J Moller ldquoA selective defect ofcytochrome c oxidase is present in brain of Alzheimer diseasepatientsrdquo Neurobiology of Aging vol 21 no 3 pp 455ndash4622000
[118] W D Parker Jr ldquoCytochrome oxidase deficiency in Alzheimerrsquosdiseaserdquo Annals of the New York Academy of Sciences vol 640pp 59ndash64 1991
[119] W D Parker Jr N J Mahr C M Filley et al ldquoReduced plateletcytochrome c oxidase activity in Alzheimerrsquos diseaserdquo Neurol-ogy vol 44 no 6 pp 1086ndash1090 1994
[120] H K Anandatheerthavarada and L Devi ldquoMitochondrialtranslocation of amyloid precursor protein and its cleaved prod-ucts relevance to mitochondrial dysfunction in AlzheimerrsquosdiseaserdquoReviews in the Neurosciences vol 18 no 5 pp 343ndash3542007
[121] H-J Park S-S Kim Y-M Seong et al ldquo120573-Amyloid precursorprotein is a direct cleavage target of HtrA2 serine proteaseimplications for the physiological function of HtrA2 in themitochondriardquoThe Journal of Biological Chemistry vol 281 no45 pp 34277ndash34287 2006
[122] X Wang B Su S L Siedlak et al ldquoAmyloid-120573 overproductioncauses abnormal mitochondrial dynamics via differential mod-ulation of mitochondrial fissionfusion proteinsrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 105 no 49 pp 19318ndash19323 2008
[123] D C David S Hauptmann I Scherping et al ldquoProteomicand functional analyses reveal a mitochondrial dysfunction inP301L tau transgenic micerdquoThe Journal of Biological Chemistryvol 280 no 25 pp 23802ndash23814 2005
[124] D C David L M Ittner P Gehrig et al ldquo120573-Amyloid treat-ment of two complementary P301L tau-expressing Alzheimerrsquosdisease models reveals similar deregulated cellular processesrdquoProteomics vol 6 no 24 pp 6566ndash6577 2006
[125] S M Cardoso M T Proenca S Santos I Santana and CR Oliveira ldquoCytochrome coxidase is decreased in Alzheimerrsquosdisease plateletsrdquo Neurobiology of Aging vol 25 pp 105ndash1102004
[126] S J Kish C Bergeron A Rajput et al ldquoBrain cytochromeoxidase in Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol59 no 2 pp 776ndash779 1992
[127] P J Crouch R Blake J A Duce et al ldquoCopper-dependent inhi-bition of human cytochrome c oxidase by a dimeric conformerof amyloid-1205731ndash42rdquo The Journal of Neuroscience vol 25 no 3pp 672ndash679 2005
[128] H Du L Guo F Fang et al ldquoCyclophilin D deficiency attenu-ates mitochondrial and neuronal perturbation and ameliorateslearning and memory in Alzheimerrsquos diseaserdquoNature Medicinevol 14 no 10 pp 1097ndash1105 2008
[129] J W Lustbader M Cirilli C Lin et al ldquoABAD directly links A120573to mitochondrial toxicity in Alzheimerrsquos diseaserdquo Science vol304 no 5669 pp 448ndash452 2004
[130] D Sirk Z Zhu J SWadia et al ldquoChronic exposure to sub-lethalbeta-amyloid (A120573) inhibits the import of nuclear-encodedproteins to mitochondria in differentiated PC12 cellsrdquo Journalof Neurochemistry vol 103 no 5 pp 1989ndash2003 2007
[131] S Melov P A Adlard K Morten et al ldquoMitochondrial oxida-tive stress causes hyperphosphorylation of taurdquo PLoS ONE vol2 no 6 article e536 2007
[132] P Bubber VHaroutunian G Fisch J P Blass andG E GibsonldquoMitochondrial abnormalities in Alzheimer brain mechanisticimplicationsrdquo Annals of Neurology vol 57 no 5 pp 695ndash7032005
[133] H Du and S S Yan ldquoMitochondrial permeability transitionpore in Alzheimerrsquos disease cyclophilin D and amyloid betardquoBiochimica et Biophysica Acta (BBA)Molecular Basis of Diseasevol 1802 no 1 pp 198ndash204 2010
[134] J Yao M Taylor F Davey et al ldquoInteraction of amyloidbinding alcohol dehydrogenaseA120573 mediates up-regulation ofperoxiredoxin II in the brains of Alzheimerrsquos disease patientsand a transgenic Alzheimerrsquos disease mouse modelrdquo Molecularand Cellular Neuroscience vol 35 no 2 pp 377ndash382 2007
[135] H Du L Guo W Zhang M Rydzewska and S Yan ldquoCyclo-philin D deficiency improves mitochondrial function andlearningmemory in aging Alzheimer disease mouse modelrdquoNeurobiology of Aging vol 32 no 3 pp 398ndash406 2011
16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

16 Oxidative Medicine and Cellular Longevity
[136] D-H Cho T Nakamura J Fang et al ldquoS-nitrosylation of Drp1mediates120573-amyloid-relatedmitochondrial fission andneuronalinjuryrdquo Science vol 324 no 5923 pp 102ndash105 2009
[137] X Wang B Su H Fujioka and X Zhu ldquoDynamin-like protein1 reduction underlies mitochondrial morphology and distri-bution abnormalities in fibroblasts from sporadic Alzheimerrsquosdisease patientsrdquo American Journal of Pathology vol 173 no 2pp 470ndash482 2008
[138] P Fernandez-Vizarra A P Fernandez S Castro-Blanco et alldquoIntra- and extracellular A120573 and PHF in clinically evaluatedcases of Alzheimerrsquos diseaserdquoHistology and Histopathology vol19 no 3 pp 823ndash844 2004
[139] M Escobar-Henriques and T Langer ldquoMitochondrial shapingcutsrdquo Biochimica et Biophysica Acta (BBA) Molecular CellResearch vol 1763 no 5-6 pp 422ndash429 2006
[140] A Neutzner R J Youle and M Karbowski ldquoOuter mito-chondrial membrane protein degradation by the proteasomerdquoNovartis Foundation Symposium vol 287 pp 4ndash14 2007
[141] I V Kurochkin ldquoInsulin-degrading enzyme embarking onamyloid destructionrdquoTrends in Biochemical Sciences vol 26 no7 pp 421ndash425 2001
[142] D J Selkoe ldquoClearing the brainrsquos amyloid cobwebsrdquoNeuron vol32 no 2 pp 177ndash180 2001
[143] R E Tanzi R D Moir and S L Wagner ldquoClearance of Alz-heimerrsquos A120573 peptide the many roads to perditionrdquoNeuron vol43 no 5 pp 605ndash608 2004
[144] A Falkevall N Alikhani S Bhushan et al ldquoDegradation ofthe amyloid120573-protein by the novelmitochondrial peptidasomePrePrdquo The Journal of Biological Chemistry vol 281 no 39 pp29096ndash29104 2006
[145] K A Johnson S Bhushan A Stahl et al ldquoThe closed structureof presequence protease PreP forms a unique 10 000 A3 chamberfor proteolysisrdquoTheEMBO Journal vol 25 no 9 pp 1977ndash19862006
[146] P Moberg A Stah S Bhushan et al ldquoCharacterization ofa novel zinc metalloprotease involved in degrading targetingpeptides in mitochondria and chloroplastsrdquo The Plant Journalvol 36 no 5 pp 616ndash628 2003
[147] N Mzhavia Y L Berman Y Qian L Yan and L A DevildquoCloning expression and characterization of human met-alloprotease 1 a novel member of the pitrilysin family ofmetalloendoproteasesrdquo DNA and Cell Biology vol 18 no 5 pp369ndash380 1999
[148] A Stahl P Moberg J Ytterberg et al ldquoIsolation and identi-fication of a novel mitochondrial metalloprotease (PreP) thatdegrades targeting presequences in plantsrdquo The Journal ofBiological Chemistry vol 277 no 44 pp 41931ndash41939 2002
[149] A Stahl S Nilsson P Lundberg et al ldquoTwo novel target-ing peptide degrading proteases PrePs in mitochondria andchloroplasts so similar and still differentrdquo Journal of MolecularBiology vol 349 no 4 pp 847ndash860 2005
[150] M Challa S Malladi B J Pellock et al ldquoDrosophila Omia mitochondrial-localized IAP antagonist and proapoptoticserine proteaserdquo The EMBO Journal vol 26 no 13 pp 3144ndash3156 2007
[151] H J Huttunen S Y Guenette C Peach et al ldquoHtrA2 regu-lates 120573-amyloid precursor protein (APP) metabolism throughendoplasmic reticulum-associated degradationrdquoThe Journal ofBiological Chemistry vol 282 no 38 pp 28285ndash28295 2007
[152] D L Vaux and J Silke ldquoMammalianmitochondrial IAP bindingproteinsrdquo Biochemical and Biophysical Research Communica-tions vol 304 no 3 pp 499ndash504 2003
[153] L Vande Walle M Lamkanfi and P Vandenabeele ldquoThemitochondrial serine protease HtrA2Omi an overviewrdquo CellDeath and Differentiation vol 15 no 3 pp 453ndash460 2008
[154] H Plun-Favreau K Klupsch N Moisoi et al ldquoThe mito-chondrial protease HtrA2 is regulated by Parkinsonrsquos disease-associated kinase PINK1rdquo Nature Cell Biology vol 9 no 11 pp1243ndash1252 2007
[155] D Harman ldquoAging a theory based on free radical and radiationchemistryrdquo Journal of Gerontology vol 11 no 3 pp 298ndash3001956
[156] A Trifunovic A Hansson A Wredenberg et al ldquoSomaticmtDNA mutations cause aging phenotypes without affectingreactive oxygen species productionrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 102 no50 pp 17993ndash17998 2005
[157] M Vermulst J H Bielas G C Kujoth et al ldquoMitochondrialpoint mutations do not limit the natural lifespan of micerdquoNature Genetics vol 39 no 4 pp 540ndash543 2007
[158] S E Schriner N J Linford G M Martin et al ldquoExtensionof murine life span by overexpression of catalase targeted tomitochondriardquo Science vol 308 no 5730 pp 1909ndash1911 2005
[159] J Liu DWKillilea and BNAmes ldquoAge-associatedmitochon-drial oxidative decay improvement of carnitine acetyltrans-ferase substrate-binding affinity and activity in brain by feedingold rats acetyl-L-carnitine andor R-120572-lipoic acidrdquo Proceedingsof the National Academy of Sciences of the United States ofAmerica vol 99 no 4 pp 1876ndash1881 2002
[160] A Navarro C Gomez M-J Sanchez-Pino et al ldquoVitaminE at high doses improves survival neurological performanceand brain mitochondrial function in aging male micerdquo TheAmerican Journal of PhysiologymdashRegulatory Integrative andComparative Physiology vol 289 no 5 pp R1392ndashR1399 2005
[161] L-J Yan and R S Sohal ldquoMitochondrial adenine nucleotidetranslocase is modified oxidatively during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 12896ndash12901 1998
[162] A Bobba G Amadoro V A Petragallo P Calissano andA Atlante ldquoDissecting the molecular mechanism by whichNH2htau and A1205731-42 peptides impair mitochondrial ANT-1
in Alzheimer diseaserdquo Biochimica et Biophysica Acta (BBA)mdashBioenergetics vol 1827 no 7 pp 848ndash860 2013
[163] I Parentini G Cavallini A Donati Z Gori and E BergaminildquoAccumulation of dolichol in older tissues satisfies the proposedcriteria to be qualified a biomarker of agingrdquo The Journals ofGerontology Series A Biological Sciences and Medical Sciencesvol 60 no 1 pp 39ndash43 2005
[164] C Edlund M Soderberg and K Kristensson ldquoIsoprenoids inaging and neurodegenerationrdquo Neurochemistry Internationalvol 25 no 1 pp 35ndash38 1994
[165] P Navas J M Villalba and R de Cabo ldquoThe importance ofplasma membrane coenzyme Q in aging and stress responsesrdquoMitochondrion vol 7 supplement pp S34ndashS40 2007
[166] H Kraskiewicz andU Fitzgerald ldquoIterfERingwith endoplasmicreticulum stressrdquoTrends in Pharmacological Sciences vol 33 no2 pp 53ndash63 2012
[167] A Ricobaraza M Cuadrado-Tejedor S Marco I Perez-Otanoand A Garcıa-Osta ldquoPhenylbutyrate rescues dendritic spineloss associated with memory deficits in a mouse model ofAlzheimer diseaserdquoHippocampus vol 22 no 5 pp 1040ndash10502012
Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 17
[168] A F Nunes J D Amaral A C Lo et al ldquoTUDCA a bile acidattenuates amyloid precursor protein processing and amyloid-120573 deposition in APPPS1micerdquoMolecular Neurobiology vol 45no 3 pp 440ndash445 2012
[169] B Gong L-Y Zhang C-P Pang D S-C Lam and G H-FYam ldquoTrimethylamine N-oxide alleviates the severe aggrega-tion and ER stress caused by G98R 120572A-crystallinrdquo MolecularVision vol 15 pp 2829ndash2840 2009
[170] M Boyce K F Bryant C Jousse et al ldquoA selective inhibitor ofeIF2120572 dephosphorylation protects cells from ER stressrdquo Sciencevol 307 no 5711 pp 935ndash939 2005
[171] P Jiang M Gan A S Ebrahim W-L Lin H L Melrose andS-H C Yen ldquoER stress response plays an important role inaggregation of 120572-synucleinrdquo Molecular Neurodegeneration vol5 no 1 article 56 2010
[172] D Y Lee K-S LeeH J Lee et al ldquoActivation of PERK signalingattenuates A120573-mediated ER stressrdquo PLoS ONE vol 5 no 5Article ID e10489 2010
[173] Y Inokuchi Y Nakajima M Shimazawa et al ldquoEffect ofan inducer of BiP a molecular chaperone on endoplasmicreticulum (ER) stress-induced retinal cell deathrdquo InvestigativeOphthalmology amp Visual Science vol 50 no 1 pp 334ndash3442009
[174] K Takano Y Kitao Y Tabata et al ldquoA dibenzoylmethanederivative protects dopaminergic neurons against both oxida-tive stress and endoplasmic reticulum stressrdquo American Journalof PhysiologymdashCell Physiology vol 293 no 6 pp C1884ndashC18942007
[175] I Yoshida A Monji K-I Tashiro K-I Nakamura R Inoueand S Kanba ldquoDepletion of intracellular Ca2+ store itselfmay bea major factor in thapsigargin-induced ER stress and apoptosisin PC12 cellsrdquo Neurochemistry International vol 48 no 8 pp696ndash702 2006
[176] H Miura K Takano Y Kitao et al ldquoA carbazole derivativeprotects cells against endoplasmic reticulum (ER) stress andglutathione depletionrdquo Journal of Pharmacological Sciences vol108 no 2 pp 164ndash171 2008
[177] E Kojro P Fuger C Prinzen et al ldquoStatins and the squa-lene synthase inhibitor zaragozic acid stimulate the non-amyloidogenic pathway by suppression of cholesterol synthe-sisrdquo Journal of Alzheimerrsquos Disease vol 20 pp 1215ndash1231 2010
[178] G Marwarha B Dasari and O Ghribi ldquoEndoplasmic retic-ulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cellstreated with the oxysterol 27-hydroxycholesterolrdquo Cellular Sig-nalling vol 24 no 2 pp 484ndash492 2012
[179] H Wang A Megill K He A Kirkwood and H-K Lee ldquoCon-sequences of inhibiting amyloid precursor protein processingenzymes on synaptic function and plasticityrdquo Neural Plasticityvol 2012 Article ID 272374 24 pages 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom





















![Military Resistance 8K14 Killing Them Softly[1]](https://img.pdfslide.us/doc/110x75/577d34ba1a28ab3a6b8eb478/military-resistance-8k14-killing-them-softly1.jpg)







