Embed Size (px)
Citation preview

425RESPUESTA DE RAPACES A UNA PRESA INTRODUCIDARevista Chilena de Historia Natural78: 425-439, 2005
Respuesta dietaria de tres rapaces frente a una presaintroducida en Patagonia
Dietary response of three raptor species to an introduced prey in Patagonia
ANA L. MONSERRAT1*, MARTÍN C. FUNES2 & ANDRÉS J. NOVARO3
1Laboratorio de Geología Marina y Dinámica de Costas, Departamento de Geología, Facultad de Ciencias Exactas yNaturales, Universidad de Buenos Aires, Pabellón II, Ciudad Universitaria, C1428EHA, Buenos Aires, Argentina
2Departamento de Fauna Terrestre, Centro de Ecología Aplicada del Neuquén,Casilla de Correos 7, Junín de los Andes, 8371, Argentina
3CONICET - Wildlife Conservation Society, Centro de Ecología Aplicada del Neuquén,Casilla de Correos 7, Junín de los Andes, 8371, Argentina
*Corresponding author: e-mail: [email protected]
RESUMEN
Los depredadores de marcada respuesta funcional pueden poseer la capacidad de estabilizar las poblaciones desus presas, lo cual es destacable en el caso de presas introducidas, que suelen causar impactos negativos en losecosistemas nativos y las actividades agropecuarias. La situación actual de la Patagonia es crítica respecto a laintroducción de especies. Estudios previos indican que los depredadores nativos exhiben un cambio general ensus dietas debido a la introducción de presas exóticas que están reemplazando a sus presas originales.Determinamos los hábitos alimenticios del águila (Geranoaetus melanoluecus), el búho magallánico (Bubomagellanicus) y el aguilucho común (Buteo polyosoma), tres de las principales aves depredadoras de la liebreeuropea (Lepus europaeus), introducida en el noroeste de Patagonia. Para esto analizamos 321, 115 y 78egagrópilas de águila, búho y aguilucho respectivamente. Evaluamos las respuestas funcionales a la densidad deliebre europea y comparamos las dietas entre sitios diferentes. Las principales presas del aguilucho y del búhofueron los roedores sigmodontinos y en segundo lugar los tuco-tucos (Ctenomys spp.). El águila consumióprincipalmente liebres y en segundo lugar tuco-tucos, presentando una respuesta funcional significativa para lapresa introducida. La dieta de este rapaz fue diferente entre sitios con alta y baja densidad de liebres. Para losotros dos rapaces no encontramos una respuesta funcional significativa para la liebre europea, pero la dieta delaguilucho varió en función del sitio geográfico. Concluimos que tanto el águila mora como el aguilucho sealimentan de manera generalista, siendo el búho el único de los tres rapaces con tendencia especialista sobrepequeños roedores. Por otra parte, la variación de la dieta del águila en función de la densidad de liebre europeale brinda el potencial de contribuir a la regulación de las poblaciones de la misma.
Palabras clave: respuesta funcional, rapaces, águila mora, búho, aguilucho, liebre europea, Patagonia.
ABSTRACT
Predators with strong functional responses may be able to help stabilize prey population levels. This is worthnoting mainly in the case of introduced prey, whose impact on native ecosystems often involves significanteconomic and environmental damage. The current Patagonian situation is critical with respect to introducedspecies. Previous studies have shown that native vertebrate predators are able to change their feeding patterns,switching from native to exotic prey. We assessed the food habits of the grey buzzard-eagle (Geranoaetusmelanoleucus), the Magellanic horned owl (Bubo magellanicus), and the red-backed hawk (Buteo polyosoma), thethree most important avian predators of the introduced European hare (Lepus europaeus) in north-westernPatagonia. We analysed 321, 115 and 78 eagle, owl, and hawk pellets respectively. We evaluated their functionalresponses to hare densities and compared their diets between different sites. Sigmodontine rodents were the hawk’sand owl’s main prey, followed by the tuco-tuco (Ctenomys spp.). The eagle consumed mainly European hares andthen tuco-tucos, showing a significant functional response to the introduced prey. This raptor also had differentdiets in sites with high and low hare density. The other two raptors did not show a significant functional responseto hare densities. However, the hawk’s diet varied according to geographic location. We conclude that eagles andhawks are generalist predators, whereas the owl tends to specialize on small mammals. The changes in the eagle’sdiet in relation to European hare densities provide the potential to contribute to regulate the hare population.
Keywords: functional response, raptors, grey buzzard-eagle, red -backed hawk, Magellanic owl, Europeanhare, Patagonia.

426 MONSERRAT ET AL.
INTRODUCCIÓN
Los depredadores pueden responder a cambiosen la disponibilidad de sus presas a través devariaciones en sus dietas (respuesta funcional)o en sus números poblacionales (respuestanumérica). La respuesta funcional de undepredador mide su tasa de consumo en funciónde la disponibilidad de la presa. Teóricamentetoda curva de respuesta funcional alcanza unnivel de saturación a altas densidades de presas.Esto ocurre cuando el depredador se sacia, peroesta respuesta puede presentar diferentesformas, reconociéndose tres tipos principalesde respuesta funcional: lineal, convexa ysigmoidea (Holling 1959). Una condiciónnecesaria, pero no suficiente, para que undepredador pueda regular una población depresas es que la pendiente de la curva derespuesta funcional sea positiva (Solomon1949). Las evidencias empíricas sugieren quelos depredadores con marcada respuestafuncional, especialmente aquellos que puedenconsumir presas alternativas con facilidad y porlo tanto suelen presentar respuesta sigmoidea,poseen mayor capacidad de estabilizar laspoblaciones de sus presas (Erlinge et al. 1984,Schmitz 1995, Norrdahl & Korpimäki 2000).
Los antecedentes en aves rapaces conrespuestas funcionales son numerosos (Lindén& Wikman 1983, Korpimäki & Norrdahl 1989,1991, Salamolard et al. 2000), pero no se haencontrado un patrón general de respuesta.Poco se conoce, por otro lado, sobreinteracciones entre depredadores y presasvertebrados cuando uno o ambos han sidointroducidos y por lo tanto no han tenido unahistoria evolutiva común. El conocimiento de ladinámica de las interacciones predador-presaadquiere relevancia económica y deconservación cuando uno de ellos es exótico,pues suelen perjudicar las actividadesproductivas humanas y el estado de losecosistemas. Tal es el caso, por ejemplo, deroedores y lagomorfos que han invadidoecosistemas productivos en todo el mundo(Davis et al. 2003, Wood & Chung Gait Fee2003).
La situación actual de la Patagonia es críticarespecto a la introducción de especies. Se haencontrado, por ejemplo, que los depredadoresnativos exhiben un cambio general en susdietas debido a la introducción de presas
exóticas que están reemplazando a sus presasoriginales (Franklin et al. 1999, Novaro et al.2000). Particularmente, la liebre europea(Lepus europaeus), que fue introducida a finesdel siglo diecinueve con propósitos de caza(Grigera & Rapoport 1983), presenta elevadasdensidades, especialmente en los ambientesmás productivos de la Patagonia. La liebreeuropea tiene un importante impacto negativosobre los hábitats y las actividadesagropecuarias, por lo que es relevanteinvestigar la posibilidad de su regulación pordepredadores naturales.
En este trabajo analizamos la respuestadietaria de tres rapaces nativas, el águila(Geranoaetus melanoleucus) , el búhomagallánico (Bubo magellanicus) y elaguilucho (Buteo polyosoma), ante la liebreeuropea introducida en el noroeste de laPatagonia. Estas rapaces son las tres principalesaves depredadoras que se presumen capaces decazar liebres europeas en la región (Jiménez &Jaksic 1990, Jiménez & Jaksic 1991, Pavez etal. 1992, Hiraldo et al. 1995, Donázar et al.1997). A través de este estudio pretendemosprofundizar el conocimiento sobre elcomportamiento alimenticio de estos tresrapaces, analizando sus dietas en diferentessitios y evaluando si presentan respuestasfuncionales en el consumo de una presaintroducida.
MATERIALES Y MÉTODOS
Área de estudio
El área de estudio se localiza en el noroeste dela Patagonia, provincia de Neuquén, Argentina(40º S, 71º O; Fig. 1), en el Distrito Occidentalde la Provincia Fitogeográfica Patagónica(Cabrera 1976). El clima es seco y frío, conheladas durante todo el año. La temperaturamedia anual es de 11 ºC y la precipitaciónmedia anual es de 280 a 1.000 mm, seconcentra durante el invierno y se incrementade este a oeste.
El área de estudio abarca una superficie de3.000 km2, con 12 sitios de muestreo en el surde Neuquén y un sitio para el búho 330 km alnorte (Fig. 1). Los sitios del sur de Neuquénque se encuentran al este en general son losmás áridos e incluyen a Calcatre, Mesa, La

427RESPUESTA DE RAPACES A UNA PRESA INTRODUCIDA
Fig. 1: Arriba: ubicación del área de estudio dentro de la provincia de Neuquén, Argentina, y enSudamérica. Abajo: detalle de los sitios de estudio en el sur de Neuquén.Above: study area location in Neuquén Province, Argentina, and in Southamerica. Below: detail of the study sites insouthern Neuquén.

428 MONSERRAT ET AL.
Papay, San Ignacio, Los Remolinos, valle deCatan Lil y La Dalmacia. La vegetacióndominante es una estepa arbustivo graminosacon Mullinum spinosum (neneo) y Senecio spp.como arbustos más comunes y diferentesespecies de Stipa spp. y Poa spp. comogramíneas predominantes. En esta zona se críaganado ovino y vacuno. Calcatre y Mesa sonsitios con suaves lomadas y mesetasatravesadas por cañadones que bajan al ríoCatan Lil . La Papay, San Ignacio y LaDalmacia se ubican sobre amplias mesetas a900-1.200 m de altitud, de las cuales sedesprenden cañadones hacia vallescircundantes. Los Remolinos también selocaliza sobre una meseta elevada pero tienemayor proporción de “mallines” o zonashúmedas con densa vegetación herbácea deCortadeira araucana, Juncus spp. y Carex spp.Valle de Catan Lil es el más bajo y abrigado delos sitios orientales, con extensos mallines conalta productividad primaria.
Los sitios más occidentales son Malleo,Riego Faldeo, Mallín Grande, Pinar Collún Coy Mendaña y están ubicados en el ecotono entrela estepa y el bosque subantártico, conpredominio de ganadería vacuna. Malleo sesitúa en el valle del río del mismo nombre, es elsitio con precipitaciones más abundantes y suvegetación de ecotono está caracterizada por lapresencia de araucarias (Araucaria araucana),habitualmente usadas como perchas por losrapaces. Riego Faldeo, Mallín Grande y PinarCollún Co son sitios de lomadas suaves conextensos pastizales y mallines y manchones dearbustos y árboles como michay (Berberisspp.), chacay (Chacaya trinervis), maitén(Maytenus boaria) y ñire (Nothofagusantarctica). Mendaña es el sitio más seco entrelos occidentales, con 700 mm de precipitaciónanual y una menor densidad de árboles yarbustos nativos. Por último, el sitio Tromen seencuentra en el norte de Neuquén a unos 1.500m de altitud en las laderas del volcán delmismo nombre, con predominio de vegetaciónherbácea y ganadería ovina y caprina.
Análisis de las dietas
Determinamos los hábitos alimenticios de losrapaces a través del análisis de egagrópilas. Larecolección de muestras se realizó de maneraoportunista en nidos o perchas entre septiembre
de 1993 y marzo de 2002, en 13 sitios (véaseÁrea de estudio); 321 egagrópilas (516 ítemes)correspondieron al águila, 115 (390 ítemes) albúho y 78 (193 ítemes) al aguilucho. Dado quelas muestras se disgregan con el tiempo,principalmente durante los meses húmedos(otoño e invierno), consideramos que la edad delas egagrópilas no era mayor a seis meses (Marti1987) y solo incluimos las muestras bienconservadas en el análisis. Una vez colectadas,las muestras se secaron en una estufa, y sedisectaron utilizando las técnicas estándares(Marti 1987). Como especímenes de referenciausamos muestras colectadas en el área de estudio(Centro de Ecología Aplicada del Neuquén,CEAN) y colecciones de museo (MuseoArgentino de Ciencias Naturales BernardinoRivadavia, de Buenos Aires). Identificamos a lospequeños mamíferos utilizando clavestaxonómicas de pelos (Chehébar & Martín 1989)y huesos (Pearson 1995). Calculamos el mínimonúmero de individuos a partir de los pares óseosencontrados en las egagrópilas (mandíbulas ypelvis). Para la cuantificación de artrópodoscontamos el número de cabezas encontradas enla muestra, mientras que para aves y reptilesconsideramos un solo individuo en la muestra encaso de encontrar plumas o escamas durante elanálisis. La nomenclatura sigue a Pearson (1995)para mamíferos y a Dickinson (2003) para aves.
Con el objeto de describir la dieta de lostres rapaces calculamos un índice de diversidaddietario, utilizando la fórmula de Levins(1968):
Bsta = (Bobs – Bmín) / (Bmax – Bmin),
donde Bobs = 1/ Σpi2, pi es la proporción del
taxón i en la dieta de un predador, Bmin es elmínimo posible de diversidad (i.e., 1) y Bmax esel máximo posible (i.e., número total de presasdiferentes consumidas por el depredador). Esteíndice estandarizado permite realizarcomparaciones entre las especies. Solo para eláguila realizamos una regresión lineal entre elvalor obtenido del índice de diversidad dietarioy la densidad de liebres en el sitio (véasesección Análisis de datos).
Análisis de datos
Analizamos la respuesta funcional mediante elestudio de la relación entre el porcentaje de

429RESPUESTA DE RAPACES A UNA PRESA INTRODUCIDA
ocurrencia de liebres en la dieta de cada rapaz,como un estimador de la tasa de ataque deldepredador (Lindén & Wikman 1983), y ladisponibilidad de liebres en el sitio. Esta últimafue estimada a partir de la densidad de liebresobtenida de un estudio simultáneo sobre ladinámica poblacional de la liebre en la región(Novaro & Funes, datos no publicados) en elcual se estimaron densidades utilizando losmétodos de conteo de heces (Novaro et al.1992) y transectas lineales (Novaro et al.2000). En los casos en que no contamos conestimaciones del mismo sitio, extrapolamosdensidades de sitios cercanos con hábitatsimilares. La densidad de conejo europeo(Oryctolagus cuniculus) en el área de estudiofue mucho menor que la de liebre (Novaro yFunes, observación personal), por lo que seutilizó como disponibilidad solo a la densidadde la última, asumiendo que todos los ítemespresa en la dieta correspondieron a liebre. Paradetectar la respuesta funcional hicimos unanálisis de regresión lineal simple (Sokal &Rohlf 1995). Para determinar el tipo derespuesta funcional evaluamos el ajuste de lostres modelos teóricos: lineal, convexo ysigmoideo, a la proporción de liebresconsumida por águilas para cada densidad deliebres utilizando el programa STATISTICA(StatSoft, Inc. 2001). No realizamos este ajustepara las otras dos especies de rapaces debido alescaso número de sitios muestreados.
Estimamos la influencia de la liebre en ladieta del águila comparando la composición dela dieta (utilizando frecuencia de los ítemes)entre grupos de sit ios con diferentedisponibilidad de liebre. Para esto hicimos unaprueba de independencia con el estadístico G, aposteriori del análisis de respuesta funcionalantes mencionado. Realizamos un análisissimilar para el búho y el aguilucho,comparando sus dietas entre sitios, utilizandoun criterio de comparación geográfico. Para elbúho comparamos la distribución defrecuencias de los ítemes de los dos sitios al surcon el sitio del norte (Tromen), el cual estáexpuesto a condiciones climáticas másrigurosas y por lo tanto es menos productivo.Para el aguilucho comparamos la dieta en losdos sitios del oeste con la de los dos sitios deleste, pretendiendo abarcar en esta comparaciónel efecto del gradiente de precipitación de laregión.
Cuando las diferencias entre los sitiosresultaron significativas, evaluamos lacontribución a la variación de los ítemesprincipales con una prueba de χ2 para muestrasindependientes con partición de grados delibertad y manteniendo una frecuencia mínimade cinco observaciones por celda (Siegel &Castellan 1995).
Para el águila también realizamos unaregresión a posteriori entre el índice de diversidaddietario y la densidad de liebres en cada sitio(Sokal & Rohlf 1995). No realizamos este análisiscon las otras dos especies de rapaces, porqueconsideramos que el tamaño de la muestra erademasiado pequeño, tomando en cuenta que laliebre no fue la presa principal para ninguna delas dos aves. Los niveles de significanciautilizados siempre fueron iguales al 5 %.
RESULTADOS
Dieta de los rapaces
En los tres casos estudiados, los mamíferos fueronlas presas más frecuentes en las dietas. Losmamíferos nativos constituyeron la mayor parte dela dieta del búho y del aguilucho, siendo losratones sigmodontinos la presa más importantepara cada una de ellas (72 y 42 %,respectivamente). La principal presa del águila fuela liebre europea, la que conformó el 35 % del totalde ítemes consumidos. La segunda presavertebrada en orden de importancia por sufrecuencia en la dieta, en los tres casos, fue el tuco-tuco (Ctenomys spp.), roedor de tamaño mediano.Solamente el águila consumió carroña,principalmente ganado menor. El ítem aves estuvopresente en la dieta del águila y del aguilucho, perono en la dieta del búho (Tablas 1, 2 y 3).
El índice de diversidad obtenido siemprefue menor a 0,25 en todos los casos.Considerando todas las presas diferentesconsumidas agrupadas al máximo nivel deresolución taxonómica logrado (sin discriminarentre diferentes invertebrados), el aguiluchopresentó la dieta más diversa (Bsta = 0,24)mientras que el búho exhibió la más selectiva(Bsta = 0,18) y el águila una dieta de diversidadintermedia (Bsta = 0,19). Un patrón diferentesurgió al considerarse solamente las presasvertebradas para el cálculo de este índice, conel búho presentando la dieta más diversa,
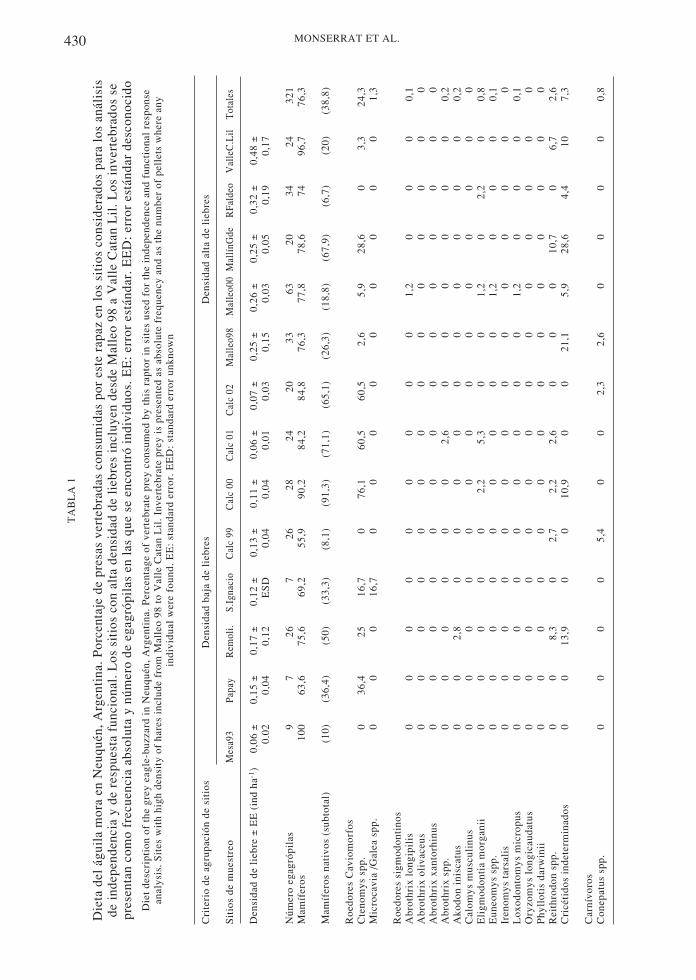
430 MONSERRAT ET AL.
TA
BL
A 1
Die
ta d
el á
guil
a m
ora
en N
euqu
én, A
rgen
tina
. Por
cent
aje
de p
resa
s ve
rteb
rada
s co
nsum
idas
por
est
e ra
paz
en l
os s
itio
s co
nsid
erad
os p
ara
los
anál
isis
de i
ndep
ende
ncia
y d
e re
spue
sta
func
iona
l. L
os s
itio
s co
n al
ta d
ensi
dad
de l
iebr
es i
nclu
yen
desd
e M
alle
o 98
a V
alle
Cat
an L
il. L
os i
nver
tebr
ados
se
pres
enta
n co
mo
frec
uenc
ia a
bsol
uta
y nú
mer
o de
ega
gróp
ilas
en
las
que
se e
ncon
tró
indi
vidu
os. E
E:
erro
r es
tánd
ar. E
ED
: er
ror
está
ndar
des
cono
cido
Die
t de
scri
ptio
n of
the
gre
y ea
gle-
buzz
ard
in N
euqu
én,
Arg
enti
na.
Per
cent
age
of v
erte
brat
e pr
ey c
onsu
med
by
this
rap
tor
in s
ites
use
d fo
r th
e in
depe
nden
ce a
nd f
unct
iona
l re
spon
sean
alys
is.
Sit
es w
ith
high
den
sity
of
hare
s in
clud
e fr
om M
alle
o 98
to
Val
le C
atan
Lil
. In
vert
ebra
te p
rey
is p
rese
nted
as
abso
lute
fre
quen
cy a
nd a
s th
e nu
mbe
r of
pel
lets
whe
re a
nyin
divi
dual
wer
e fo
und.
EE
: st
anda
rd e
rror
. E
ED
: st
anda
rd e
rror
unk
now
n
Cri
teri
o de
agr
upac
ión
de s
itio
sD
ensi
dad
baja
de
lieb
res
Den
sida
d al
ta d
e li
ebre
s
Sit
ios
de m
uest
reo
Mes
a93
Papa
yR
emol
i.S.
Igna
cio
Cal
c 99
Cal
c 00
Cal
c 01
Cal
c 02
Mal
leo9
8M
alle
o00
Mal
línG
deR
Fald
eoV
alle
C.L
ilT
otal
es
Den
sida
d de
lie
bre ±
EE
(in
d ha
-1)
0,06
±0,
15 ±
0,17
±0,
12 ±
0,13
±0,
11 ±
0,06
±0,
07 ±
0,25
±0,
26 ±
0,25
±0,
32 ±
0,48
±0.
020,
040,
12E
SD
0,04
0,04
0,01
0,03
0,15
0,03
0,05
0,19
0,17
Núm
ero
egag
rópi
las
97
267
2628
2420
3363
2034
2432
1M
amíf
eros
100
63,6
75,6
69,2
55,9
90,2
84,2
84,8
76,3
77,8
78,6
7496
,776
,3
Mam
ífer
os n
ativ
os (
subt
otal
)(1
0)(3
6,4)
(50)
(33,
3)(8
,1)
(91,
3)(7
1,1)
(65,
1)(2
6,3)
(18,
8)(6
7,9)
(6,7
)(2
0)(3
8,8)
Roe
dore
s C
a vio
mor
fos
Cte
nom
ys s
pp.
036
,425
16,7
076
,160
,560
,52,
65,
928
,60
3,3
24,3
Mic
roc a
via
/Ga l
e a s
pp.
00
016
,70
00
00
00
00
1,3
Roe
dore
s si
gmod
onti
nos
Abr
othr
ix l
ongi
pili
s0
00
00
00
00
1,2
00
00,
1A
brot
hrix
oli
vac e
us0
00
00
00
00
00
00
0A
brot
hrix
xa n
torh
inus
00
00
00
00
00
00
00
Abr
othr
ix s
pp.
00
00
00
2,6
00
00
00
0,2
Ako
don
inis
c atu
s0
02,
80
00
00
00
00
00,
2C
a lom
ys m
usc u
linu
s0
00
00
00
00
00
00
0E
ligm
odon
tia
mor
gani
i0
00
00
2,2
5,3
00
1,2
02,
20
0,8
Eun
eom
ys s
pp.
00
00
00
00
01,
20
00
0,1
Ire n
omys
ta r
sali
s0
00
00
00
00
00
00
0L
oxod
onto
mys
mic
ropu
s0
00
00
00
00
1,2
00
00,
1O
ryzo
mys
lon
gic a
uda t
us0
00
00
00
00
00
00
0P
hyll
otis
da r
win
ii0
00
00
00
00
00
00
0R
e ith
rodo
n sp
p.0
08,
30
2,7
2,2
2,6
00
010
,70
6,7
2,6
Cri
c éti
dos
inde
term
inad
os0
013
,90
010
,90
021
,15,
928
,64,
410
7,3
Ca r
nívo
ros
Con
epa t
us s
pp.
00
00
5,4
00
2,3
2,6
00
00
0,8

431RESPUESTA DE RAPACES A UNA PRESA INTRODUCIDA
Cri
teri
o de
agr
upac
ión
de s
itio
sD
ensi
dad
baja
de
lieb
res
Den
sida
d al
ta d
e li
ebre
s
Sit
ios
de m
uest
reo
Mes
a93
Papa
yR
emol
i.S.
Igna
cio
Cal
c 99
Cal
c 00
Cal
c 01
Cal
c 02
Mal
leo9
8M
alle
o00
Mal
línG
deR
Fald
eoV
alle
C.L
ilT
otal
es
Ede
ntad
osC
haet
ophr
actu
s /
Zae
dyus
spp
.10
00
00
00
2,3
01,
20
00
1
Car
roña
Lam
a gu
anic
oe0
00
00
00
00
1,2
00
00,
1
Mam
ífer
os in
trod
ucid
os (
subt
otal
)(9
0)(2
7,3)
(36,
1)(4
1,7)
(81,
1)(8
,7)
(13,
2)(2
5,6)
(50)
(63,
5)(1
0,7)
(75,
6)(7
6,7)
(46,
2)
Roe
dore
sR
attu
s sp
p.0
00
00
00
2,3
03,
50
00
0,5
Lag
omor
fos
L.
euro
paeu
s/ O
. cu
nicu
lus
3027
,325
41,7
37,8
8,7
7,9
20,9
5057
,610
,775
,663
,335
,1
Car
roña
Ovi
s /
Cap
ra60
08,
30
43,2
05,
32,
30
2,4
00
13,3
10,4
Cer
vus
elap
hus
00
2,8
00
00
00
00
00
0,2
Roe
dore
s in
dete
rmin
ados
00
2,8
00
00
00
1,2
00
00,
3M
amíf
e ros
ind
e te r
min
ados
00
00
5,4
00
2,3
2,6
00
00
0,8
Rep
tile
sO
fidi
os0
27,3
00
00
13,2
70
00
4,4
04
Sau
rios
00
2,8
00
00
02,
60
00
00,
4R
epti
les
inde
term
inad
os0
00
00
02,
60
07,
10
4,4
01,
1
Ave
s0
9,1
8,3
255,
40
00
18,4
9,4
21,4
8,9
3,3
8,4
Ve r
tebr
a dos
ind
e te r
min
ados
00
00
00
00
00
00
00
Núm
e ro
de v
e rte
bra d
os10
1136
1237
4638
4338
8528
4530
459
Inve
rte b
rado
sE
gagr
ópil
a s c
on i
nve r
tebr
a dos
50
41
144
02
05
03
341
Col
e ópt
e ros
10
51
200
03
02
01
033
Art
rópo
dos
inde
term
inad
os6
00
02
50
00
30
44
24
Núm
e ro
de í
tem
e s t
ota l
e s17
1141
1359
5138
4638
9028
5034
516
Am
plitu
d de
la d
ieta
(va
lor
c alc
ula d
o0,
197
0,08
4pa
ra e
l gru
po d
e si
tios)
TA
BL
A 1
(co
ntin
uaci
ón)

432 MONSERRAT ET AL.
TABLA 2
Dieta del aguilucho común (Buteo polyosoma) en Neuquén, Argentina. Porcentaje de presas verte-bradas consumidas por este rapaz en los sitios considerados para los análisis de independencia y derespuesta funcional. Los invertebrados se presentan como frecuencia absoluta y número de egagró-
pilas en las que se encontró individuos. EE: error estándar. EED: error estándar desconocidoDiet description of the red-backed hawk in Neuquén, Argentina. Percentage of vertebrate prey consumed by this raptor insites used for the independence and functional response analysis. Invertebrate prey is presented as absolute frequency and
as the number of pellets where any individual were found. EE: standard error. EED: standard error unknown
Criterio de agrupación de sitios Oeste Este
Sitios de muestreo Valle C. Lil 3 Mendaña Dalmacia S. Ignacio 2 Totales
Densidad de liebre ± EE (ind ha-1) 0,8 ± 0,14 0,25 ± 0,17 0,29 ± 0,06 0,12 ± ESDNúmero egagrópilas 24 21 13 20 78Mamíferos 90,9 72,7 84,6 81,3 82,4
Mamíferos nativos (subtotal) (81,8) (68,2) (84,6) (65,6) (75,1)
Roedores caviomorfosCtenomys spp. 51,5 15,9 46,2 18,8 33,1Microcavia /Galea spp. 0 0 0 0 0
Roedores sigmodontinosAbrothrix longipilis 0 0 0 0 0Abrothrix olivaceus 0 0 0 0 0Abrothrix xantorhinus 0 0 0 0 0Abrothrix spp. 6,1 0 0 6,3 3,1Akodon iniscatus 0 0 0 0 0Calomys musculinus 3 0 0 0 0,8Eligmodontia morganii 3 6,8 15,4 0 6,3Euneomys spp. 0 0 0 0 0Irenomys tarsalis 0 0 0 0 0Loxodontomys micropus 0 0 0 0 0Oryzomys longicaudatus 0 0 0 0 0Phyllotis darwinii 3 0 0 0 0,8Reithrodon spp. 0 4,5 15,4 3,1 5,8Cricétidos indeterminados 15,2 40,9 7,7 37,5 25,3
CarnívorosConepatus spp. 0 0 0 0 0
EdentadosChaetophractus / Zaedyus spp. 0 0 0 0 0
CarroñaLama guanicoe 0 0 0 0 0
Mamíferos introducidos (subtotal) (9,1) (2,3) (0) (9,4) (5,2)
RoedoresRattus spp. 0 0 0 6,3 1,6
LagomorfosLepus europaeus/ Oryctolagus cuniculus 9,1 2,3 0 3,1 3,6
CarroñaOvis / Capra 0 0 0 0 0Cervus elaphus 0 0 0 0 0
Roedores indeterminados 0 2,3 0 6,3 2,1Mamíferos indeterminados 0 0 0 0 0

433RESPUESTA DE RAPACES A UNA PRESA INTRODUCIDA
Criterio de agrupación de sitios Oeste Este
Sitios de muestreo Valle C. Lil 3 Mendaña Dalmacia S. Ignacio 2 Totales
ReptilesOfidios 0 2,3 11,5 0 3,5Saurios 3 4,5 3,8 3,1 3,6Reptiles indeterminados 0 18,2 0 15,6 8,5
Aves 6,1 2,3 0 0 2,1
Vertebrados indeterminados 0 0 0 0 0Número de vertebrados 33 44 26 32 135
InvertebradosEgagrópilas con invertebrados 7 7 8 5 27Coleópteros 2 11 3 5 21Artrópodos indeterminados 7 1 28 1 37Número de ítemes totales 42 56 57 38 193Amplitud de la dieta 0,276 0,193(valor calculado para el grupode sitios)
TABLA 2 (continuación)
TABLA 3
Dieta del búho magallánico (Bubo magellanicus) en Neuquén, Argentina. Porcentaje de presasvertebradas consumidas por este rapaz en los sitios considerados para los análisis de independencia
y de respuesta funcional. Los invertebrados se presentan como frecuencia absoluta y número deegagrópilas en las que se encontró individuos. EE: error estándar
Diet description of the Magellanic horned owl in Neuquén, Argentina. Percentage of vertebrate prey consumed by thisraptor in sites used for the independence and functional response analysis. Invertebrate prey is presented as absolute
frequency and as the number of pellets where any individual were found. EE: standard error
Criterio de agrupación de sitios Sur Norte
Sitios de muestreo Pinar Collún Co Valle C. Lil 2 Tromen Totales
Densidad de liebre ± EE (ind ha-1) 0,14 ± 0,08 0,58 ± 0,11 0,22 ± 0,11Número egagrópilas 26 15 74 115Mamíferos 100 97,4 98,1 98,5
Mamíferos nativos (subtotal) (96,4) (92,3) (87,3) (92)
Roedores caviomorfosCtenomys spp. 12,7 25,6 19,7 19,4Microcavia /Galea spp. 0 0 0,6 0,2
Roedores sigmodontinosAbrothrix longipilis 1,8 0 4,5 2,1Abrothrix olivaceus 5,5 0 1,9 2,5Abrothrix xantorhinus 0 0 0,6 0,2Abrothrix spp. 0 0 2,5 0,8Akodon iniscatus 1,8 0 0,6 0,8Calomys musculinus 3,6 7,7 0,6 4Eligmodontia morganii 12,7 7,7 18,5 13Euneomys spp. 0 0 5,1 1,7Irenomys tarsalis 1,8 0 0 0,6Loxodontomys micropus 9,1 0 0,6 3,2Oryzomys longicaudatus 1,8 10,3 0 4Phyllotis darwinii 1,8 12,8 0 4,9Reithrodon spp. 16,4 12,8 7 12,1Cricétidos indeterminados 27,3 15,4 24,8 22,5
CarnívorosConepatus spp. 0 0 0 0

434 MONSERRAT ET AL.
Criterio de agrupación de sitios Sur Norte
Sitios de muestreo Pinar Collún Co Valle C. Lil 2 Tromen Totales
EdentadosChaetophractus / Zaedyus spp. 0 0 0 0
CarroñaLama guanicoe 0 0 0 0
Mamíferos introducidos (subtotal) (0) (2,6) (7) (3,2)
RoedoresRattus spp. 0 0 2,5 0,8
LagomorfosL. europaeus/ O. cuniculus 0 2,6 4,5 2,3
CarroñaOvis / Capra 0 0 0 0Cervus elaphus 0 0 0 0
Roedores indeterminados 3,6 2,6 3,8 3,3Mamíferos indeterminados 0 0 0 0
ReptilesOfidios 0 0 0,6 0,2Saurios 0 2,6 0,6 1,1Reptiles indeterminados 0 0 0 0
Aves 0 0 0 0
Vertebrados indeterminados 0 0 0,6 0,2Número de vertebrados 55 39 157 251
InvertebradosEgagrópilas con invertebrados 0 3 135 130Coleópteros 0 3 90 93Artrópodos indeterminados 0 1 45 46Número de ítemes totales 55 43 292 390Amplitud de la dieta (valor calculado 0,363 0,104para el grupo de sitios)
seguido por el aguilucho y luego por el águila(Bsta = 0,33, 0,24 y 0,15, respectivamente).
El índice sugirió que el águila expande sudieta en los sitios en donde la densidad de liebrees baja, y el aguilucho hace lo mismo en lossitios al oeste. Sin embargo, la regresión linealentre el índice y la densidad de liebres en cadasitio para el águila resultó ser no significativa,tanto al considerar todos los ítemes de su dietacomo al incluir solo las presas vertebradas (P =0,53, R2 = 0,04, mismo valor en ambos casos).
Respuestas funcionales
El águila es el único de los tres rapaces quepresentó un incremento significativo en elconsumo de liebres con relación a densidadescrecientes de liebre (n = 13, R2 = 0,50, P < 0,01).
Las respuestas funcionales del aguilucho (n = 4,R2 = 0,8036, P = 0,10), así como la del búho (n =3, R2 = 0,07, P = 0,83) no fueron detectadas eneste estudio. El águila presentó una saturaciónen su respuesta funcional a partir de unadensidad aproximada de 0,3 liebres por hectárea,con una frecuencia de 65-75 % de liebre (Fig. 2).El ajuste de los tres modelos no nos permitedefinir cuál modelo explica mejor los datos yaque, excepto por la asíntota del modelosigmoideo, ninguno de los parámetros estimadosdifirió significativamente de cero (Tabla 4).
Cambios de dieta entre sitios
El águila cambió sus hábitos alimenticiosdependiendo del sitio (G = 129,9, P < 0,0001),consumiendo diferencialmente todos los ítemes
TABLA 3 (continuación)

435RESPUESTA DE RAPACES A UNA PRESA INTRODUCIDA
Fig. 2: Respuesta funcional de tres especies de aves rapaces frente a la liebre europea en Neuquén,noroeste de Patagonia, Argentina. (A) Respuesta del águila. La ecuación de la regresión y R2 semuestra dentro del área de gráfico. (B) Respuesta del aguilucho. (C) Respuesta del búho.Functional response of three raptor species to the European hare in Neuquén, northwestern Patagonia, Argentina. (A) Greyeagle-buzzard response. Regression equation and R2 are shown inside the graphic area. (B) Red-backed hawk response. (C)Magellanic horned-owl response

436 MONSERRAT ET AL.
considerados para el análisis (P < 0,005): tuco-tuco, liebre, roedores sigmodontinos, aves,carroña (ganado menor) y reptiles. Losartrópodos podrían haber sido incidentalmenteingeridos, por lo cual no fueron incluidos en elanálisis, dado que podrían enmascarar lospatrones de los demás ítemes. La mayor partede la varianza fue explicada por el mayorconsumo de liebres en sitios donde la misma esmás abundante, como se observa en losresultados arrojados por la partición de la tablade contingencia (X2 = 57,3). En los sitios debaja densidad de liebre, el águila consumióprincipalmente tuco-tucos entre las presasnativas (X2 = 38), y también más carroña (X2 =14,9).
También el aguilucho varió la composiciónde su dieta entre sitios (G = 18,3, P < 0,005).Consideramos los mismos grupos de ítemes queen el caso del águila, excepto por la carroñaque no fue considerada por estar ausente en ladieta de este rapaz. La proporción de roedoresnativos explicó la mayor parte de la variaciónentre sitios. El análisis de partición de gradosde libertad demostró que el aguilucho consumemayor cantidad de tuco-tucos en el este, ymenor cantidad de roedores sigmodontinos (X2
= 20,24, P < 0,005). Las proporciones del restode los ítems no explicaron una cantidadsignificativa de variación de la dieta entre sitios(X2 = 0,003, P > 0,05).
En el caso del búho, no hubo diferenciassignificativas entre la dieta en los sitios al nortey al sur (G = 3,27, P > 0,05). Los ítemes que
consideramos para este análisis fueron tuco-tucos, roedores sigmodontinos, liebre y reptiles.La frecuencia de ocurrencia del resto de losítemes no fue suficiente como para incluirlos enel análisis.
DISCUSIÓN
Dieta de las rapaces
Actualmente la principal presa del águila en elárea de estudio es la liebre europea. Estudiosprevios en la región muestran este mismocomportamiento, reportando siempre unlagomorfo como presa principal de este rapaz(Pavez et al. 1992, Hiraldo et al. 1995). Elcambio de consumo de presas nativas aintroducidas no es exclusivo de las aves rapacesen la región (Franklin et al. 1999, Novaro et al.2000). Novaro et al. (2000) encontraron unpatrón similar entre los carnívoros terrestres dela zona, con lo que es evidente el impacto delas especies introducidas en las comunidadesdel noroeste de Patagonia.
El águila es el rapaz que presenta el mayornúmero de ítemes presa diferentes y a la vez elmenor índice de diversidad de presasvertebradas, lo cual sugiere que podría tenermayor capacidad para cazar diferentes presas,pero que concentra su dieta en ciertos ítemes,aparentemente los más redituables desde elpunto de vista energético. Tanto el aguiluchocomo el búho son capaces de consumir liebres
TABLA 4
Parámetros de los distintos modelos ajustados a los datos de respuesta funcional del águila enNeuquén, Argentina. Se presentan en negrita los valores que se esperaban significativos. El valor
con asterisco presenta aquel que efectivamente fue significativoDifferent models parameters adjusted to the functional response data of the grey eagle-buzzard in Neuquén, Argentina. Thevalues presented in bold are those expected to be significant. The value with the asterisk presents the one that indeed was
significant
Modelo sigmoideo Modelo convexo Modelo lineal (piecewise)
Asíntota Pendiente Intersección Pendiente Constante Ordenada Pendiente Inflexión Pendienteal origen inicial del plateau
Valor estimado 75,62 5,25 -8,22 136,62 0,5 13,45 104,31 100,07 -76,6Error estándar 28,89 3,57 5,26 122,84 0,68 10,6 64,22 55,74 136,65Estadístico t 2,62 1,47 -1,56 1,11 0,73 1,27 1,62 1,8 -0,56Valor de P 0,03* 0,17 0,15 0,29 0,48 0,24 0,14 0,11 0,59Grados de libertad 10 11 9Valor de R 0,72 0,7 0,79% varianza explicada 51,54 49,38 61,83

437RESPUESTA DE RAPACES A UNA PRESA INTRODUCIDA
europeas, pero en ambos casos sus presasprincipales fueron pequeños roedores nativos.Pese a que la liebre es una presa más redituableen cuanto a su peso, la mayor proporción detuco-tuco en la dieta del aguilucho y del búho,posiblemente se deba a la dificultad de estosrapaces para cazar liebres debido a su pequeñotamaño corporal. Si los roedores pequeños sonmás abundantes que los roedores de medianotamaño como los tuco-tucos, esto explicaría elmayor consumo de los primeros por ambosrapaces según, la teoría de forrajeo óptimo(Schoener 1971).
La dieta del aguilucho muestra que este esel depredador más generalista de los tresestudiados. El aguilucho posee una dieta menosdiversa en los sitios del este del área de estudio,donde posiblemente el tuco-tuco es másabundante. El águila presenta la mismatendencia en su comportamiento alimentario,con una dieta menos diversa en los sitios dondela liebre es más abundante, aunque el resultadono significativo de la regresión lineal sugiereque este patrón puede ser no lineal.
Respuestas funcionales
Pavez et al. (1992) e Hiraldo et al. (1995)sugirieron la existencia de una respuestafuncional en el águila con respecto al conejo yla liebre europea, respectivamente, cuandodetectaron variaciones temporales en lafrecuencia de dichos lagomorfos en la dieta deláguila. Nuestro estudio es el primero encuantificar dicha respuesta funcional. Nodetectamos respuestas funcionales respecto a laliebre europea en el comportamientoalimentario del aguilucho y del búho. Aunquenuestro número de muestras en ambos casos esdemasiado pequeño para realizar conclusionesdefinitivas, los resultados sugieren que ladisponibilidad de libres puede no tener unefecto importante en la dieta de estos rapacesdebido a su talla. Detectamos como principalpresa del búho a los roedores sigmodontinos aligual que Trejo & Grigera (1998). Sin embargo,no podemos descartar que este resultado reflejesimplemente el consumo por un depredadoroportunista de las presas más abundantes, puesno poseemos datos de disponibilidad de estosroedores en todos los sitios de estudio.
Numerosos estudios previos en aves rapacesencontraron que la curva de respuesta funcional
llega a un techo o plateau, indicando lasaturación del depredador. Sin embargo,diferentes autores han ajustado diversosmodelos a la respuesta funcional de águilas,correspondiendo en algunos casos al tipoconvexo, típico de depredadores especialistas(Restani et al. 2000, Salamolard et al. 2000).No obstante, una respuesta funcional sigmoideapuede verse enmascarada cuando la densidadde la presa principal cambia más abruptamenteque la de la presa alternativa, obteniéndose unarespuesta del tipo convexo, con lo que lasconsecuencias de la depredación en lapoblación de presas no deberían ser inferidasúnicamente a partir de la curva de respuestaante la presa principal (Murdoch 1973).Aunque encontramos una respuesta funcionalsignificativa para el águila, no podemos decidirsobre la base de nuestros resultados, quémodelo de respuesta explica mejor elcomportamiento del águila, posiblementedebido a un insuficiente número de sitios demuestreo. Jaksic et al. (1992) no encontraronuna respuesta funcional significativa en avesdepredadoras estudiadas en Chile, pero sí unafuerte respuesta numérica ante las fluctuacionesde pequeños mamíferos. En nuestro caso noevaluamos la respuesta numérica de losrapaces, pero cabe mencionar que el águilaparece ser fuertemente territorial (Sánchez-Aldao et al . 2001), siendo esta unacaracterística que limitaría una respuestanumérica por migración como la reportada porJaksic et al. (1992). No obstante, es posible queeste carácter territorial de las águilas serestrinja a las estaciones reproductivas. Si eltamaño poblacional de este rapaz se mantienerelativamente constante a través del año, sereforzaría el efecto de depredación del águilasobre la población de liebres. Es posible, porotro lado, que la depredación por parte deláguila contribuya a la regulación de lapoblación de liebre europea al sumarse alconsumo de liebres que realizan los mamíferosdepredadores de la zona, como el zorro culpeo(Pseudalopex culpaeus), el zorro chilla (P.griseus) y el puma (Puma concolor) (Novaro etal. 2000).
Comparación de la dieta entre sitios
La dieta del águila varía marcadamente enfunción de la densidad de liebres de cada sitio.

438 MONSERRAT ET AL.
Esta respuesta implica un comportamientogeneralista, por el cual el consumo de cadapresa depende no solo de su disponibilidad sinotambién de la disponibilidad de liebres. Estemecanismo suele estar ligado a un tipo derespuesta funcional sigmoidea frente a la presaprincipal, posibili tando la regulaciónpoblacional de la misma por depredación(Murdoch 1973, Schmitz 1995).
El aguilucho mostró también uncomportamiento alimentario generalista, condiferencias en su dieta entre los sitios orientalesy occidentales del área de estudio. Esta variaciónes explicada por el consumo diferencial deroedores nativos, siendo en el oeste su principalpresa los sigmodontinos, pero presentando en eleste un mayor consumo de tuco-tucos. En lazona oriental del área de estudio, los sueloshabitualmente son más arenosos y menosconsolidados y por ende más favorables paraespecies fosoriales como los tuco-tucos, cuyascuevas son observadas con mayor frecuencia(Funes y Novaro, observación personal). Por lotanto, el aguilucho podría estar depredando mástuco- tucos donde estos son más abundantes,dado que es una presa más redituableenergéticamente que los sigmodontinos, debidoa su mayor tamaño. Por otra parte, no seobservaron diferencias marcadas en la dieta delbúho entre sitios. No podemos concluir sobre labase de este resultado que exista uncomportamiento especialista de este depredadorporque no poseemos datos de disponibilidad desus principales presas. Sin embargo, es muypoco probable que los ensambles de presasdisponibles para el búho sean los mismos en lossitios comparados dado que son ambientesseparados geográficamente y notablementediferentes.
AGRADECIMIENTOS
Agradecemos al departamento de FaunaTerrestre del CEAN (Centro de EcologíaAplicada del Neuquén) por brindar materiales detrabajo y el laboratorio para el análisis de lasegagrópilas, y en particular a Guillermo SánchezAldao, Oscar Pailacura y Obdulio Monsalvo porla recolección de egagrópilas. Agradecemos aSusan Walker, quien confeccionó los mapas delárea de estudio y asistió con los análisisestadísticos, y a los dos revisores anónimos que
brindaron sugerencias para mejorar elmanuscrito. También agradecemos el espaciocedido en el Museo Argentino de CienciasNaturales Bernardino Rivadavia. Las tareas decampo fueron subvencionadas por el CEAN, laSecretaría de Agricultura, Ganadería, Pesca yAlimentación de la Nación y la WildlifeConservation Society.
LITERATURA CITADA
CABRERA AL (1976) Regiones fitogeográficas argenti-nas. Enciclopedia Argentina de agricultura y Jardi-nería. Segunda Edición, Tomo II, Fase 1 ACME,Buenos Aires, Argentina. 85 pp.
CHEHÉBAR C & S MARTÍN (1989) Guía para el recono-cimiento microscópico de los pelos de los mamífe-ros de la Patagonia. Doñana, Acta Vertebrata (Espa-ña) 16: 247-291.
DAVIS SA, H LEIRS, R P PECH, Z ZHANG & NCSTENSETH (2003) On the economic benefit ofpredicting rodent outbreaks in agricultural systems.Crop Protection 23: 305-314.
DICKINSON E. ed (2003) The Howard and Moore com-plete checklist of the birds of the world. PrincetonUniversity Press, Princeton, New Jersey, USA.1056 pp.
DONÁZAR JA, A TRAVAINI, O CEBALLOS, MDELIBES & F HIRALDO (1997) Food habits of thegreat horned owl in northwestern ArgentinePatagonia: the role of introduced lagomorphs.Journal of Raptor Research (USA)31: 364-369.
ERLINGE S, G GORANSSON, G HOGSTEDT, GJANSSON, O LIBERG, J LOMAN, IN NILSSON,T VON SCHANTZ & M SYLVÉN (1984) Canvertebrate predators regulate their prey? AmericanNaturalist 123: 125-133.
FRANKLIN WL, WE JOHNSON, RJ SARNO & JAIRIARTE (1999) Ecology of the Patagonia pumaFelis concolor patagonica in southern Chile.Biological Conservation 90: 33-40.
GRIGERA DE & EH RAPOPORT (1983) Status anddistribution of the European hare in South America.Journal of Mammalogy 64: 163-166.
HIRALDO F, JA DONÁZAR, O CEBALLOS, A TRAVAINI,J BUSTAMANTE & MC FUNES (1995) Breedingbiology of the grey eagle-buzzard population inPatagonia. Wilson Bulletin 107: 675-685.
HOLLING CS (1959) Some characteristics of simple typesof predation and parasitism. Canadian Entomologist91: 385-398.
JAKSIC FM, JE JIMÉNEZ, SA CASTRO & PFEINSINGER (1992) Numerical and functionalresponse of predators to a long-term decline inmammalian prey at semi-arid Neotropical site.Oecologia 89: 90-101.
JIMÉNEZ JE & FM JAKSIC (1990) Historia natural deGeranoaetus melanoleucus. Hornero 13: 97-110.
JIMÉNEZ JE & FM JAKSIC (1991) Behavioural ecologyof red-backed hawks in central Chile. WilsonBulletin 103: 132-137.
KORPIMÄKI E & K NORRDAHL (1989) Predation ofTengmalm’s owls: numerical responses, functionalresponses and dampening impact on populationfluctuations of microtines. Oikos 54:154-164.

439RESPUESTA DE RAPACES A UNA PRESA INTRODUCIDA
KORPIMÄKI E & K NORRDAHL (1991) Numerical andfunctional responses of kestrels, Short-eared owls,and long-eared owls to voles densities. Ecology 72:814-826.
LEVINS R (1968) Evolution in changing environments.Princeton University Press, Princeton, New Jersey,USA. 120 pp.
LINDÉN H & M WIKMAN (1983) Goshawk predation ontetraonids: availability of prey and diet of thepredator in the breeding season. Journal of AnimalEcology 52: 953-968.
MARTI CD (1987) Raptor food habits studies. NationalWildlife Federation, Raptor management techniquesmanual: 67-80. Washington, District of Columbia,USA.
MURDOCH WW (1973) The functional response ofpredators. Journal of Applied Ecology 10: 335-342.
NORRDAHL K & E KORPIMÄKI (2000) Do predatorsl imit the abundance of al ternative prey?Experiments with vole-eating avian and mammalianpredators. Oikos 91: 528-540.
NOVARO AJ, MC FUNES & RS WALKER (2000)Ecological extinction of native prey of a carnivoreassemblage in Argentine Patagonia. BiologicalConservation 92: 25-33.
NOVARO AJ, AF CAPURRO, A TRAVAINI, MCFUNES & JE RABINOVICH (1992) Pellet-countsampling based on spatial distribution: a cases tudy o f t he European ha re in Pa tagon ia .Ecología Austral (Argentina) 2: 11-18.
PAVEZ EF, CA GONZÁLEZ & JE JIMÉNEZ (1992) Dietshifts of Black–chested Eagles (Geranoaetusmelanoleucus) from native prey to EuropeanRabbits in Chile. Journal of Raptor Research (USA)26: 27-32.
PEARSON OP (1995) Annotated keys for identifying smallmammals living in or near Nahuel Huapi NationalPark or Lanín National Park, southern Argentina.Mastozoología Neotropical (Argentina) 2: 99-148.
RESTANI M, AR HARMATA & EM MADDEN (2000)Numerical and functional responses of migrant baldeagles exploiting a seasonally concentrated foodsource. Condor 102: 561-568.
SALAMOLARD M, A BUTET, A LEROUX & VBRETAGNOLLE (2000) Responses of an avianpredator to variations in prey density at a temperatelatitude. Ecology 81: 2428-2441.
SÁNCHEZ-ALDAO G, MC FUNES, OB MONSALVO,MB BONGIORNO & O PAILACURA (2001) Eláguila mora en el sudeste del DepartamentoHuiliches, Provincia del Neuquén. Departamento deFauna Silvestre Terrestre, Centro de Ecología Apli-cada del Neuquén, Argentina. 11 pp.
SCHMITZ OJ (1995) Functional responses of optimalconsumers and the potential for regulation ofresource populations. Wildlife Research 22: 101-113.
SCHOENER TW (1971) Theory of feeding strategies.Annual Review of Ecology and Systematics 11:396-404.
SIEGEL S & NJ CASTELLAN (1995) Estadística noparamétrica: aplicada a las ciencias de la conduc-ta. Cuarta edición. Editorial Trillas, México. 437pp.
SOKAL RR & FJ ROHLF (1995) Biometry: the principlesand practice of statistics in biological research.Third edition. WH Freeman and Company, NewYork, New York, USA. 850 pp.
SOLOMON ME (1949) The natural control of animalpopulations. Journal of Animal Ecology 18: 1-35.
STATSOFT, INC (2001) STATISTICA (data analysis soft-ware system), version 6. www.statsoft.com.
TREJO A & D GRIGERA (1998) Food habits of the greathorned owl (Bubo virginianus) in a Patagoniansteppe in Argentina. Journal of Raptor Research(USA) 32: 306-311.
WOOD BJ & CHUNG GAIT FEE (2003) A critical reviewof the development of rat control in Malaysianagriculture since the 1960s. Crop Protection 22:445-461.
Editor Asociado: Cristian EstadesRecibido el 11 de agosto de 2004; aceptado el 6 de enero de 2005





























