Embed Size (px)
Citation preview

BiologicalJournal of the Linnean Sociep (1995). 56: 505-5 15. With 2 figures
Resistance to thermal stress in adult Drosojhila buzzatii: acclimation and variation among populations
ROBERT A. KREBS’ AND VOLKER LOESCHCKE
Department of Ecology and Genetics, University of Aarhus, Ny Munkegade, 8000 Aarhus C, Denmark
Received 70 Oclober 1994; acceptedforpublication 14 February 1995
Survival after heat stress was examined in adult Drosophila buuatii from seven localities. Males and females were conditioned by a non-lethal high temperature before exposure to a severe thermal stress. Variable times elapsed between conditioning, either as adults or pupae, and exposure to the stress were used, and experimental times to the stress ranged from 2 to 96 hours. Survival after stress varied among populations, and differences generally were consistent across conditioning treatments and across experiments. Resistance to thermal stress was much higher following one conditioning bout 2-4 hours before exposure to a severe stress than when the time elapsed between conditioning and exposure was increased to 24 or more hours. Significantly more adults survived the stress if conditioned 4 days before exposure to the thermal stress, either as adults or as pupae, than if not conditioned. The rank order of resistance roughly followed that predicted from the climatic conditions of the localities of origin.
d lYY.5 The Linnem Society of London
ADDITIONAL KEY WORDS:-acclimation - genetic variation - xuheat shock - temperature resistance.
CONTENTS
Introduction . . . Material and methods . .
Population and time of conditioning variation . Pupal conditioning and adult surviwd . . . . Statistical analyses . . .
Results . . . . Population and time of conditioning variation . . . . Pupal conditioning and adult survival . . . .
Discussion . . . . . . Acknowledgements . . . . . . References . . . . .
506 506 507 508 508 508 508 510 511 513 514
‘Corresponding author current address: Department of Organismal Biology, University of Chicago, 1027 East 57th Street, Chicago IL 60637, U.S.A.
505 0024-4066/95/012505+11$12.00/0 0 1995 The Linnean Society of London

506 R. A. KREBS AND V. LOESCHCKE
INTRODUCTION
Genetic variation for resistance to thermal extremes is present within and among populations of Drosophila (Hosgood & Parsons, 1968; Morrison & Milkman, 1978; Parsons, 1979; Kilias & Alahiotis, 1985; Quintana & Prevosti, 1990a; Loeschcke, Krebs & Barker, 1994; Krebs & Loeschcke, 1994a), and differences in resistance to thermal stress can evolve among lines of Drosophila maintained at different constant conditions in the laboratory (Alahiotis & Stephanou, 1982; Huey, Partridge & Fowler, 199 1; Cavicchi et al., 1995). Further, adult Drosophila that are conditioned by exposure to a mild thermal stress become more resistant than those not conditioned, and resistance is greater for individuals conditioned a greater number of times, although at a decreasing gain in resistance (Loeschcke et al, 1994; Krebs & Loeschcke, 199413).
Despite a greater abundance of analyses on heat induced acclimation in D. mdanogaster Meigen, the evolution of resistance to high temperature stress may be observed better within D. buxatii Patterson & Wheeler, a species in which the ecology is well defined, and high temperatures may affect their distribution in nature (Barker et al., 1984; Krebs & Barker, 1993). Larvae of D. buzatii grow and pupate within necrotic cladodes of Opuntia cactus (rots) (Barker & Mulley, 1976), and may become exposed to high and rapidly changing temperatures under natural conditions (Krebs & Loeschcke, 1994a). Although adult flies may depart from and return to rots, they may be found associated with the cactus at all times of the day as it appears to be the primary location for feeding and mating (Santos et aZ., 1988), and the only suitable sites for oviposition (Barker, 1992). Climatic conditions also vary among localities where cactus and cactophilic flies occur. Therefore, populations collected from localities that are warmer or more variable are predicted to be more heat tolerant than those from cooler or climatically less variable regions, or they may differ in their ability to acclimate to high temperatures.
Here we provide an expansion of earlier analyses of variation within and among populations of D. buzatii (Loeschcke et al, 1994; Krebs & Loeschcke, 1994a) using acclimation treatments that have been identified in D. melanogaster to induce large changes in the type and concentration of proteins produced (DiDomenico, Bugaisky & Lindquist, 1982a; 1982b). We examine variation in resistance to heat shock among seven populations to answer the following questions: Does the time elapsed between conditioning flies to high temperature and exposure to heat stress alter resistance to stress, and if so, what is the temporal pattern of induction and decay of resistance? Are resistance patterns similar for all populations ? Further, what effect might conditioning flies as pupae have on stress resistance of adults?
MATERIAL AND METHODS
Mass populations of D. buzzatii were prepared from flies collected from seven localities. Collections were made at four Australian localities (obtained April, 1991, with the northern flies collected by J. S. F. Barker with either R. Krebs or V. Loeschcke): from Oxford Downs, OD, (Lat. 21”5O’S) and

HEAT SHOCK RESPONSE IN ADULTDROSOPHLLA 507
Dixalea, DX, (Lat. 23”56’S) in Queensland, from Metz Gorge, ME, (30”21’S) in New South Wales and from Bulla, BU, near Melbourne, in Victoria (Lat. 38”OO’S, collected by Neil Murray). The Argentina population, ARG, (collected March, 1991) was obtained near El Chanar, in Tucuman province (Lat. 26”48’S, collected by Andre LaChance for J. S. F. Barker), the Canary Island population, CI, (28”10’N), collected November, 1992, we obtained from southwestern Teneriffe, and that from Spain, near Cordoba, COR, (Lat. 37”53’N) was collected by Jean David, also in November, 1992. Most populations were derived from isofemale lines, each derived from large numbers of offspring from a single female collected in nature (10 each except for Argentina, with seven lines only, and the Canary Islands with 20 lines). The population from Cordoba was derived from pooled offspring of more than ten females. The Australian and Argentinean lines had been maintained on a cactus supplemented medium (Starmer & Barker, 1986) until September, 1992. In Denmark, all populations were maintained on instant Drosophila medium (Carolina Biological Supply). Repeated experimentation on oviposition preference in D. buuatii (Barker, 1992) and stress resistance in other Drosophila species (Parsons, 1980a) indicate these maintenance techniques preserve most of the genetic variation at the population level, although some genetic variation within lines may be lost since collection.
Population and time of conditioning variation
Adults from six populations, Canary Islands (CI), Oxford Downs (OD), Dixalea (DX), Metz Gorge (ME), Bulla (BU) and Argentina (ARG), were exposed to heat stress when 6-7-days-old. Emerging adults were collected and sorted to sex under ether anaesthesia and held in groups of c. 20 per vial on a yeast-sucrose-agar medium with added live yeast. All flies were transferred to fresh vials every 3 days, and for conditioning or for exposure to the thermal stress. Flies conditioned more than 24 hours before exposure to heat stress were placed in fresh yeasted vials 24 h after conditioning. In the first experiment, individuals were conditioned 24, 48, 72 or 96 h before exposure to heat stress. In the second experiment, the population from Cordoba (COR) was added, adults were exposed at 4 days of age, and individuals were conditioned 2, 4 or 24 h before exposure to heat stress, or they were not conditioned.
The conditioning temperature applied for 75 min to adults in both experiments was 38.0% (incubator temperature). Immediately before conditioning, stoppers to the vials were removed, moistened, and replaced, and vials were inverted. Thereby, close to saturated humidity within vials was reached. The conditioning treatment approaches the temperature at which this species becomes immobile, and appears to be a mild stress based on a behavioural change. Flies move to the base of the chamber, which is the coolest part of the vial, but the treatment used here caused no mortality.
The stress treatment used was 415°C for a duration of 100 min in the first experiment (also in inverted vials with moistened stoppers), which was determined from earlier analyses as a treatment that consistently caused 20- 50% mortality when acclimation was performed 24 h before exposure to the stress (Krebs & Loeschcke, 1994a). The stress first immobilizes the flies,

508 R. A. KREBS AND V. LOESCHCKE
called ‘knockdown’ (Huey et al., 1992), and continued exposure kills. The stress duration was raised to 110 min to increase mortality in the second experiment because the acclimation treatment was performed closer in time to the stress exposure. Data were the proportion of flies surviving in each vial, which was scored as those that could walk 24 h after exposure to allow time for recovery of mobility, although flight is not regained.
Pupal conditioning and adult survival
Pupae from all seven populations were collected in flasks prepared with instant Drosophila medium on which c. 20 pairs of adults had oviposited for two days. Eight days later 2 x 5 cm pieces of filter paper were folded lengthwise and placed within the flasks, three per flask, to provide attractive surfaces for pupation. These papers were removed 24 h later and were placed in vials containing agar medium, with about 20 pupae per vial. After another 24 h, half of the pupae were conditioned by exposure to 38°C (incubator temperature) for 60 min. As for adults, stoppers were moistened to increase humidity for the conditioning treatment, but there was no need to invert vials. Adults were collected from these pupae without anaesthesia (eclosion proportion greater than 95%), and were held for one day before exposing them to heat stress (41.5”C, for 100 min) 96 hrs after conditioning, as above. Males and females were not separated during exposure.
Statistical analyses
For the three experiments, a general linear models procedure (SAS institute, 1989) was used to test significance of effects, primarily those of locality, sex (except where pupae were conditioned), and conditioning treatment, and interactions among these effects. AU effects were treated as fixed. For main effects where more than two treatments were administered, the Tukey’s multiple comparison test was used to clarify differences among groups.
Block effects were common in the analyses possibly due to day-to-day variation in the temperature or perhaps the heat-up time within incubators, either of which would affect the stress level imposed on individuals for any particular block of replicates. However, interactions between block and other main effects were minimal, and where present could be identified generally as statistical effects of the change in variance among blocks when means are proportions, i.e. variance is at a maximum for a mean of 0.5, and decreases symmetrically towards one and zero. Therefore, block interactions are not expected to be zero unless all block means are equal. This problem was reduced by use of the arcsine-square-root transformation in analyses of variance. Nevertheless, while block effects were partitioned in all analyses of variance, interactions with block were left as a component of the error variance.
RESULTS
Population and time of conditioning variation
Sign&cant variation for survival of D. buzz&ii after heat stress was present among the six localities (Fig. lA,B), and, as observed previously, survival of
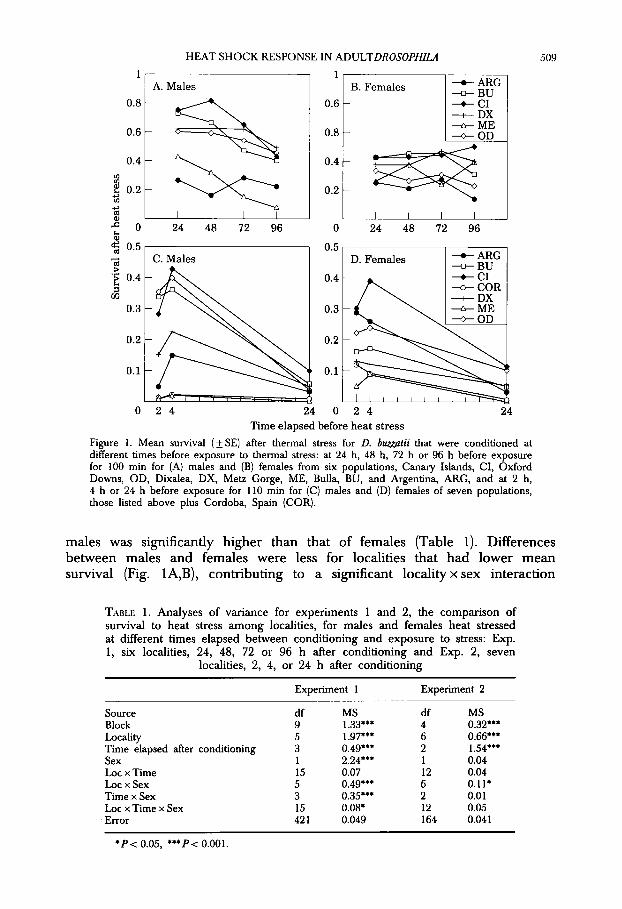
HEAT SHOCK RESPONSE IN ADULTDROSOPHILA 509
1 A. Males
24 48 ‘72 96
C. Males
0.6
0.8
0.4
0.2
0
0.5
0.4
0.3
0.2
0.1
24 24 0
B. Females -ARG -BU - CI -+-DX -ME +OD
_I- 24 48 72 96
D. Females -ARG -BU - CI - COR -i-DX
24 Time elapsed before heat stress
Figure 1. Mean survival (+ SE) after thermal stress for D. b-hi that were conditioned at different times before exposure to thermal stress: at 24 h, 48 h, 72 h or 96 h before exposure for 100 min for (A) males and (B) females from six populations, Canary Islands, CI, Oxford Downs, OD, Dixalea, DX, Metz Gorge, ME, Bulla, BU, and Argentina, ARG, and at 2 h, 4 h or 24 h before exposure for 110 min for (C) males and (D) females of seven populations, those listed above plus Cordoba, Spain (COR).
males was significantly higher than that of females (Table 1). Differences between males and females were less for localities that had lower mean survival (Fig. lA,B), contributing to a significant locality x sex interaction
TABLE 1. Analyses of variance for experiments 1 and 2, the comparison of survival to heat stress among localities, for males and females heat stressed at different times elapsed between conditioning and exposure to stress: Exp. 1, six localities, 24, 48, 72 or 96 h after conditioning and Exp. 2, seven
localities, 2, 4, or 24 h after conditioning
Experiment 1 Experiment 2
Source df MS df MS Block 9 1.33*** 4 0.32*** Locality 5 1.97*** 6 0.66*** Time elapsed after conditioning 3 0.49*** 2 1.54*** Sex 1 2.24*** 1 0.04 Lot x Time 15 0.07 12 0.04 ILK x Sex 5 0.49*** 6 0.11* Time x Sex 3 0.35*** 2 0.01 Lot x Time x Sex 15 0.08; 12 0.05 Error 421 0.049 164 0.041
*p< 0.05, ***p-c 0.001.

510 R. A. KREBS AND V. LOESCHCKE
(Table 1). The time elapsed between conditioning and exposure to severe stress significantly affected survival (Table l), but this difference was due almost entirely to the decline in survival of males with increased time between conditioning and heat stress (Fig. 1A). The time elapsed between conditioning and heat stress had little effect on survival of females (Fig. lB), with the time x sex interaction significant (Table 1). From individual ANOVAs for each sex (not shown), male survival with conditioning 24 h before exposure to stress differed from that at 72 h before, and survival for conditioning times 24, 48 and 72 h differed significantly from 96 h (Tukey’s test, PC 0.05). Therefore, the difference between males and females for resistance to heat stress decreased with increased time between conditioning and heat stress, with survival of males not significantly different from that of females when flies were conditioned 96 h before exposure to stress.
Using a greater stress than in the first experiment, 110 min at 41.5”C compared with 100 min, survival of D. bumtii was much lower (Fig. 1). Almost all flies that were not conditioned died (10 survivors out of c. 1700). Therefore, the analysis of variance (Table 1) and presentation of means (Fig. lC,D) used only data from the three groups of flies that were conditioned with different times elapsed before exposure to stress: 2, 4 or 24 h. The main effects, locality, sex and time elapsed between conditioning and stress were significant (Table 1). As before, survival of males exceeded that of females except for the populations where survival was lowest, the Argentina, Cordoba and Metz Gorge localities (Fig. lC,D), which contributed to the significant locality x sex interaction (Table 1).
Survival was higher both for males and females (Fig. lC, D) that were conditioned 4 h before exposure to heat stress than for those conditioned only 2 h before exposure, but both of these short times elapsed between conditioning and stress led to significantly higher survival than conditioning 24 h before exposure (Tukey’s test, P-C 0.05). As before, the interaction between locality of origin and the time elapsed before conditioning was not significant (Table l), but the effect of the time at which conditioning was done was similar in males and females (time x sex interaction, ns).
Pupal conditioning and adult survival
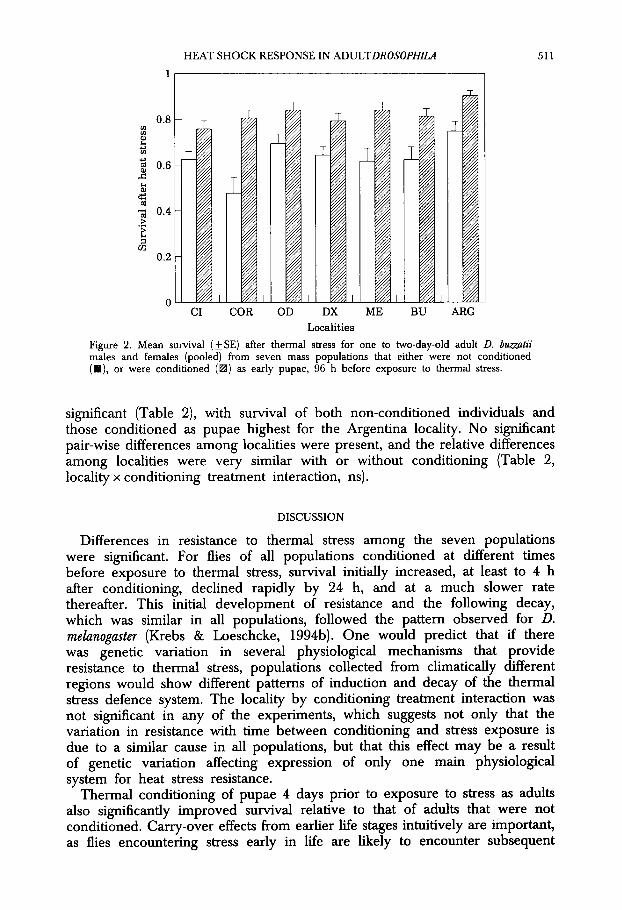
Survival of adult D. bumtii that were conditioned as pupae 96 hrs before exposure to stress was significantly higher than that for adults that were not conditioned (Table 2, Fig. 2). V ariation among the seven localities was
T,w,e 2. Analysis of variance for the comparison of survival of D. bwtii to heat stress among seven localities for flies conditioned or not as pupae and heat stressed as adults
96 h later
Source df MS Block 1 0.84;;; LOCdity 6 0.12*+ Conditioning trt 1 3.01*** LocxCond trt 6 0.03 Error 196 0.040
**p< 0.01, l **p< 0.001.
HEAT SHOCK RESPONSE IN ADULTDROSOPHLol
1
I 0.8 7
COR OD DX Localities
ME BU
511
Figure 2. Mean survival (*SE) after thermal stress for one to hvo-day-old adult D. buzz&ii males and females (pooled) from seven mass populations that either were not conditioned (W), or were conditioned (m) as early pupae, 96 h before exposure to thermal stress.
significant (Table 2), with survival of both non-conditioned individuals and those conditioned as pupae highest for the Argentina locality. No significant pair-wise differences among localities were present, and the relative differences among localities were very similar with or without conditioning (Table 2, locality x conditioning treatment interaction, ns).
DISCUSSION
Differences in resistance to thermal stress among the seven populations were significant. For flies of all populations conditioned at different times before exposure to thermal stress, survival initially increased, at least to 4 h after conditioning, declined rapidly by 24 h, and at a much slower rate thereafter. This initial development of resistance and the following decay, which was similar in all populations, followed the pattern observed for D. melanogaster (Krebs & Loeschcke, 199413). One would predict that if there was genetic variation in several physiological mechanisms that provide resistance to thermal stress, populations collected from climatically different regions would show different patterns of induction and decay of the thermal stress defence system. The locality by conditioning treatment interaction was not significant in any of the experiments, which suggests not only that the variation in resistance with time between conditioning and stress exposure is due to a similar cause in all populations, but that this effect may be a result of genetic variation affecting expression of only one main physiological system for heat stress resistance.
Thermal conditioning of pupae 4 days prior to exposure to stress as adults also significantly improved survival relative to that of adults that were not conditioned. Carry-over effects from earlier life stages intuitively are important, as flies encountering stress early in life are likely to encounter subsequent

512 R. A. KREBS AND V. LOESCHCKE
stresses. Quantitative differences in survival to heat stress between flies conditioned or not as pupae are similar to those reported for adults (Loeschcke et aZ., 1994) suggesting that the decay in resistance is similar in adults and pupae. This result also is predicted from the hypothesis that a single physiological system largely is responsible for resistance to thermal stress in adult Drosophila.
One possible system for the control of thermal resistance is the regulation of the production of heat shock proteins (hsps). The phenotypic expression of increased heat shock resistance is associated with the synthesis of a small group of proteins that are rapidly induced following exposure to temperatures lO”C-15°C above those optimal for growth, while most preexisting protein synthesis is repressed. Normal protein synthesis is gradually reactivated after the stress is removed (Lindquist, 1986). A direct link between higher concentrations of these proteins and increased resistance to thermal extremes has been supported in three ways: (1) heat shock protein concentrations in D. melanogaster selected for increased resistance to thermal stress appeared higher than that of lines selected for susceptibility to heat shock (Stephanou et cd, 1983), (2) 1’ mes of D. melanogaster that express hsp70 from a heterologous promotor at 25°C can survive direct exposure to high temperatures without a conditioning treatment (Solomon et al, 1991), and (3) ermbryos of D. melanogaster strains that have 12 extra copies of the hsp70 gene obtain resistance to heat stress much faster than those of wild type strains (Welte et al., 1993). The correlation between the conditions inducing acclimation to heat stress and those inducing production of the heat shock proteins is also very high (Ashburner, 1982; Parse11 & Lindquist, 1994). The pattern and speed of acclimation, occurring within 2 h and increasing up to at least 4 h (Fig. I), is similar to the pattern of expression of mRNA for hsp70 in D. melunogaster (DiDomenico et al., 1982a; 198213). Finally, the half life of heat shock proteins is known to be very long (Landry et aZ., 1982) and the shape of the decay curve for some heat shock proteins (DeRocher et al., 1991), which is similar to that for decay in resistance to stress (Fig. l), shows that heat shock proteins persist for a long time after cells are returned to non- stress conditions. The possibility that genetic variation in resistance to thermal stress is affected by quantitative or qualitative differences in the expression of heat shock proteins therefore should not be ignored.
Hoffmann & Parsons (1991) proposed that resistance to thermal stress should be related to the environment of origin. The significant variation among populations of D. buxatii for survival to heat stress roughly followed environmental variation, as was suggested for three other Australian populations (Loeschcke et aZ., 1994). Greater thermal extremes in Melbourne, Australia than in tropical Townsville were proposed as the cause of D. melanogaster and D. simulans Sturtevant from this locality being more resistant to several stress types (Parsons, 1977, 1979, 1980b). For D. buzzztii, the host plants both on the Canary Islands and at Bulla (near Melbourne, Australia), which were the most resistant to heat stress, are shrubby prickly pear species fully exposed to the sun. Necrotic cactus pads may reach very high temperatures all year round on the Canary Islands (Krebs & Loeschcke, 1994a). At the subtropical localities, Dixalea and Oxford Downs, rots of the tree pear, Opuntia tomentosa Salm-Dyck, which grows within open- woodlands, are

HEAT SHOCK RESPONSE IN ADULTDROSOPHlLA 513
utilized. Therefore, while average air temperatures recorded may be warmer at these sites than, for example, in Melbourne, the conditions to which flies are exposed may be more mild. Resistance to thermal stress of D. buzz&ii from these sites was intermediate. Of the least resistant populations, a tree pear (0. jicus-indicu) also is utilized at the Cordoba site, which is at a more northern latitude comparable to Melbourne, while the Metz Gorge site, although containing a shrub pear, is at a high altitude and in a canyon exposed much less to direct sunlight. Information on the Argentina site is insufficient to include these flies in the comparison. An analysis of many more populations is necessary to clearly describe relationships between the environment and stress resistance.
An alternative cause of population variation could be changes within populations with maintenance in the laboratory. However, the rank order for resistance remained roughly consistent during more than one year of maintenance of lines beginning with preliminary experiments and one set of results published previously (Krebs & Loeschcke, 1994a), and for two follow- up experiments (Krebs, unpublished). The one exception to consistency was for resistance of flies from the Argentina locality, which increased from early experiments to become intermediate. The cause of the change is unknown as maintenance of all populations was identical. After the shift, relative resistance remained consistently intermediate.
Large differences between males and females for survival after thermal stress are clear from the results here and in Loeschcke et al. (1994), but their presence is difficult to explain. In D. melanogaster survival of females, on average, exceeds that of males (Huey et aZ., 1991; Krebs & Loeschcke, 1994c, Cavicchi et al., 1995), although for field collected D. melanogaster and D. simuluns, survival of males was higher than that of females (Jenkins & Hoffmann, 1994). The cause of the significant population by sex interaction in the analyses of variance may be in part a sex by stress level effect (Dahlgaard, Krebs & Loeschcke, 1995). As the level of stress is increased, the relative advantage of males declines. Larger differences between males and females occurred when survival of males was high, and survival of females exceeded that of males when mean survival was very low, which explains the greater similarity of means among populations for females than for males (Fig. 1). An analogous sex difference is observed in D. subobscura, where survival after thermal stress is higher for males in young flies, but for females in old flies, which are less stress resistant (Quintana & Prevosti, 1990b).
In summary, all analysed populations of D. buzatii were similar for the activation and decay of resistance to thermal stress over a variety of conditioning treatments. The acclimation response prepares an organism for a variety of stress types, although its expression is most pronounced for thermal stress. The activation and regulation of this genetic system therefore must be a critical part of the way Drosophila and other organisms respond physiologically to thermal change in nature.
ACKNOWLEDGEMENTS
We thank Camilla Hakansson, Doth Andersen and especially Jennifer Krebs for their assistance with data collection, Jesper Dahlgaard for many

514 R. A. KREBS AND V. LOESCHCKE
useful discussions and, Ary Hoffmann, Stuart Barker, Peter Parsons and an anonymous reviewer for comments on the manuscript. This research was supported by grants to V.L. from the Danish Natural Science Research Council (No. 11-9639-2 and 11-9719-1) and the Carlsberg Foundation (No. 93-0280-30).
REFERENCES
Alahioti~ SN, Stephanou, G. 1982. Temperature adaption of Drosophila populations. The heat shock protein system. Comparative Biochemistry and Physiology 73B: 529-533.
Ashburner, M. 1982. The effect of heat shock and other stress on gene activity: an introduction. In: Schlesinger, MJ, Ashbumer M, Tissieres A, eds. Heat Shock Cold Spring Harbor: Cold Spring Harbor Laboratory Press, l-9.
Barker JSF. 1992. Genetic variation in cactophilic Drosophila for oviposition on natural yeast substrates. Evolution 46: 1070-1083.
Barker JSF, Mulley JC. 1976. Isozyme variation in natural populations of Drosophila buwtii. Evolution 30: 2 13-233.
Barker JSF, East PD, Phaff HJ, Miranda M. 1984. The ecology of the yeast tlora in necrotic Opuntia cacti & of associated Drosophila in Australia. Microbial Ecology 10: 379-399.
Cavlccbl S, Guerra D, La Terre V., Huey RB. 1995. Chromosomal analysis of heat-shock tolerance in Drosophila melanogast~ evolving at different temperatures in the laboratory. Evolution 49: 676-684.
Dahlgaard J, Krebs RA, Loeschcke V. 1995. Heat-shock tolerance and inbreeding in Drosophila buzz&ii. Heredity 74: 157-163.
DeRocher AE, Helm KW, Lauzon LM, Vierling E. 1991. Expression of a conserved family of cytoplasmic low molecular weight heat shock proteins during heat-stress and recovery. Plant Physiology 96: 1038-1047.
DiDomenico BJ, Bugaisky GE, Lindquist S. 1982a. The heat shock response is self-regulated at both the transcriptional and posttranscriptional levels. Cell 31: 593-603.
DiDomenico BJ, Bugaisky GE, Lindquist S. 1982b. Heat shock and recovery are mediated by different translational mechanisms. fioceedings of the National Academy of Sciences (USA) 79: 6181- 6185.
Hotiann AA, Parsons PA. 1991. Evolutionary Genetics and Environmental Stress. Oxford: Oxford Science Publications.
Hosgood SMW, Parsons PA. 1968. Polymorphism in natural populations of Drosophila for the ability to withstand temperature shocks. Experientia 24: 727-728.
Huey RB, Partridge L, Fowler K. 1991. Thermal sensitivity of Drosophila melanogaster responds rapidly to laboratory natural selection. Evolution 45: 751-756.
Huey RB, Grill WD, Kingsolver JG, Weber KE. 1992. A method for rapid measurement of heat or cold resistance of small insects. Functional Ecology 6: 489-494.
Jenkins NL, Hofhann AA. 1994. Genetic and maternal variation for heat resistance in Drosophila from the field. Genetics 137: 783-789.
KiIla~ G, AlaMotls SN. 1985. Indirect thermal selection in Drosophila melanogaster and adaptive consequences. Theoretical and Applied Genetics 69: 645-650.
Krebs RA, Barker JSF. 1993. Coexistence of ecologically similar colonising species. II. Population differentiation in Drosophila aldrichi and D. bmtii for competitive effects and responses at different temperatures, and allozyme variation in D. aldrichi Jounal of Evolutionary Biology 6: 281-298.
Krebs RA, Loescbcke V. 1994a Response to environmental change: genetic variation and fitness in Drosophila bwtii following temperature stress. In: Loeschcke V, Tomiuk J, Jain SK, eds. Conservation Genetics. Basel: Birkhauser, 309-32 1.
Krebs RA, Loeschcke V. 199413. Costs and benefits of activation of the heat shock response in Drosophila melanogaster. Functional EEoloby 8: 730-737.
Krebs RA, Loeschcke V. 1994c Effects of exposure to short-term thermal extremes on fitness components in Drosophila melanogaster. Journal of Evolutionary Biology 7: 39-49.
Landry J, Bemier D, Chr&tien P, Nicole LM, Tanguay RM, Marceau N. 1982. Synthesis & degradation of heat shock proteins during development and decay of thermotolerance. Cancer Research 42: 2457-2461.
Lindquist S. 1986. The heat-shock response. Annual Beviezo of Biochemistry 55: 1151-1191. Loeschcke V, Krebs RA, Barker JSF. 1994. Genetic variation for resistance and acclimation to high
temperature stress in Drosophila buzatii. Biological Journal of the Linnean Society 52: 83-92. Morrison WW, Milkman R. 1978. Modification of heat resistance in Drosophila by selection. Nature
273: 49-50. Parsell DA, Lindquist S. 1994. Heat shock proteins and stress tolerance. In: Morimoto RI, Tissieres

HEAT SHOCK RESPONSE IN ADULTDROSOPHLLA 515
A, Georgopoulos C, eds. The Biology of Heat Shock Proteins and Molecular Chaperones. Cold Spring Harbor: Cold Spring Harbor Laboratory Press, 457-494.
Parsons PA. 1977. Resistance to cold temperature stress in populations of Drosophila melanogaster and D. simularu. Australian Journal of Zoology 25: 693-698.
Parsons PA. 1979. Resistance of the sibling species Drosophila melasogaster and D. simulanr to high temperatures in relation to humidity: evolutionary implications. Evolution 33: 131-136.
Parsons PA. 1980a Isofemale strains and evolutionary strategies in natural populations. Evolutionary Biology 13: 175-217.
Parsons PA. 1980b. Parallel climatic races for tolerances to high temperature-desiccation stress in hvo Drosophila species. Journal of Biogeography 7: 97-101.
Qnintana A, F’revosti A. 199Oa. Genetic and environmental factors in the resistance of Drosophila subobscura adults to high temperature shock 2. Modification of heat resistance by indirect selection. Theoretical and Applied Genetics 80: 847-851.
[email protected] A, Prevosti A. 1990b. Genetic and environmental factors in the resistance of Drosophila subobscura adults to high temperature shock 1. Breeding temperature and crowding. Theoretical and Applied Genetics 79: 103-107.
SAS Institute Inc 1989. SAS/Stat Users Guide, Gary, N.C. Santos M, Ruiz A, Barbadilla A, Quazada-DiazJE, Hasson E, Fontdevila A. 1988. The evolutionary
history of Drosophila buwtii XIV. Larger flies mate more often in nature. Heredity 61: 255-262. Solomon JM, Rossi JM, Golic K, McGarry T, Lindquist S. 1991. Changes in Hsp70 alter
thermotolerance and heat-shock regulation in Drosophila. New Biologkt 3: 1106-1120. Starmer WT, Barker JSF. 1986. Ecological genetics of the Adh-7 locus of Drosophila buzatii. Biological
Journal of the Linnean Society 28: 373-385. Stephanou G, Alahiotis SN, Christodoulou C, Marmaras VJ. 1983. Adaptation of Drosophila to
temperature: Heat-shock proteins and survival in Drosophila melanogaster. Developmental Genetics 3: 299-308.
Welte MA, Tetrault, JM, Dellavalle RP, Lindquist S. 1993. A new method for manipulating transgenes: Engineering heat tolerance in a complex multicellular organism. Current Biology 3: 842- 853.





























