Embed Size (px)
Citation preview

© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim 1434-2944/09/510-0542
Internat. Rev. Hydrobiol. 94 2009 5 542–559
DOI: 10.1002/iroh.200811107
SHUCHAN ZHOU1, 2, XIANGFEI HUANG1 and QINGHUA CAI*, 1
1Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, Hubei 430072, P.R. China;e-mail: [email protected]
2Institute of Biodiversity Science, Fudan University, Shanghai 200433, P.R. China;e-mail: [email protected]
Research Paper
Temporal and Spatial Distributions of Rotifers in Xiangxi Bay of the Three Gorges Reservoir, China
key words: Rotifera, temporal and spatial dynamic, new impoundment, longitudinal distribution, Yangtze River
Abstract
From July 2003 to June 2005, investigations of rotifer temporal and spatial distributions were car-ried out in a bay of the Three Gorges Reservoir, Xiangxi Bay, which is the downstream segment of the Xiangxi River and the nearest bay to the Three Gorges Reservoir dam in Hubei Province, China. Thirteen sampling sites were selected. The results revealed a high species diversity, with 76 species, and 14 dominant species; i.e., Polyarthra vulgaris, Keratella cochlearis, Keratella valga, Synchaeta tremula, Synchaeta stylata, Trichocerca lophoessa, Trichocerca pusilla, Brachionus angularis, Brachionus caly-ciflorus, Brachionus forficula forficula, Ascomorpha ovalis, Conochilus unicornis, Ploesoma trunca-tum and Anuraeopsis fissa. After the first year of the reservoir impoundment, the rotifer community was dominated by ten species; one year later it was dominated by eight species. The community in 2003/2004 was dissimilar to that in 2004/2005, which resulted from the succession of the dominant species. The rotifer community exhibited a patchy distribution, with significant heterogeneity observed along the longitudinal axis. All rotifer communities could be divided into three groups, corresponding to the riverine, the transition and the lacustrine zone, respectively.
1. Introduction
Zooplankton heterogeneity and community structure at a range of spatial and temporal scales is an important focus of aquatic ecological research (CLARK et al., 2001). Studies on temporal variations in zooplankton distribution are common, but research on spatial patchiness has received relatively less attention. Zooplankton spatial distributions are as heterogeneous as those of terrestrial and aquatic plants and animals (FERNÁNDEZ-ROSADO and LUCENA, 2001). Now, it is a well-known phenomenon (SEDA and DEVETTER, 2000) and has been shown to be an ecologically important feature of freshwater ecosystems (PINEL-ALLOUL et al., 1999). The structure and composition of organisms and the spatial pattern of a community are of crucial importance for understanding ecosystem functioning (ROSENZWEIG, 1991; ROMARE et al., 2003) because they can affect ecosystem processes, functioning and stability (MAESTRE et al., 2005).
* Corresponding author

Rotifers in Three Gorges Reservoir 543
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
Reservoirs are complex systems, considered as transitions between rivers and lakes. Sev-eral important differences between lakes and reservoirs are observed such as geological age, maximum depth location, shape and water retention time (STRAŠKRABA and TUNDISI, 1999). These differences are mostly pronounced in canyon-shaped reservoirs, which are spa-tially highly heterogeneous because of their relatively short retention times and longitudinal heterogeneity (MAŠÍN et al., 2003). A canyon-shaped reservoir presents three very distinct zones: the riverine zone, in the upper reservoir, which is subject to the influence of either the tributaries or river of origin; the transition zone, downstream from the reservoir, which functions as an intermediate river-lake ecosystem; and the lacustrine zone, located further downstream (MATSUMURA-TUNDISI and TUNDISI, 2005). These zones vary widely in flow and depth, making characterization of the reservoir more difficult than that of most natural lakes (BERNOT et al., 2004).
The Three Gorges Reservoir of China (a newly impounded canyon-shaped reservoir), is a world-famous hydroelectric project (Fig. 1). This reservoir is undergoing a process of
Figure 1. The location of Xiangxi Bay in China (above) and sampling stations (below).

544 S. ZHOU et al.
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
eutrophication, which can strongly affect numbers, standing crops, population dynamics, production and community structure of zooplankton (CAJANDER, 1983). It is reported that man-made lakes present ecological features that lead to the establishment of a very dynamic system in which the plankton communities play an important role (BRANCO et al., 2002). However, zooplankton studies within the Three-Gorge Reservoir are scarce, especially at the initial stages of impoundment. Therefore in this article we reported: 1) the temporal dynam-ics of rotifers in Xiangxi Bay of the Three Gorges Reservoir since impoundment; and 2) the longitudinal and transverse distributions of rotifers in this bay.
2. Materials and Methods
2.1. Study Site Description
The Xiangxi River is an important tributary of the Yangtze River in Hubei province, China. The river, with a catchment area of 3,099 km2, has a length of 94 km, and a natural fall of 1540 m (TANG et al., 2002). With the impoundment of the Three Gorges Reservoir, water depth in the outlet of the Xiangxi River has increased from about 20 to 70 m; water current velocity has decreased from 0.43–0.92 m · s–1 to 0.0020–0.0041 m · s–1. The water body was neutral to slightly alkaline at the time of sampling. The downstream segment (about 25 km) of this river is now called the Xiangxi Bay. It is the biggest bay near the dam of Three Gorges Reservoir in Hubei province, China. There is a depth gradient increasing from the riverine zone to the lacustrine zone (at the mouth of Xiangxi River, about 70 m).
From July 2003 to June 2005, a monthly investigation of the rotifer community was conducted in Xiangxi Bay. Nine sampling sites were selected (Fig. 1), to provide a spatial characterization of the bay. Moreover, to analyze the transverse distribution of rotifers in Xiangxi Bay, two transects (ST 1 and ST 5) with three sites were sampled. These sites followed the flow direction, including the left littoral zone (indicated by L), the middle pelagic zone, and the right littoral zone (indicated by R).
2.2. Sampling Methods
At each site, we sampled three layers: surface water (0.5 m), Secchi disk depth and two times Sec-chi depth with a 10 L modified Schindler-trap sampler. These discrete samples were then pooled and mixed, and subsamples for analyses of chlorophyll a and rotifer community were taken from the pooled samples. For chlorophyll a determinations, 0.6 L of pooled water were filtered through a WHATMAN GF/C glass-filter and analyzed spectrophotometrically after 95% acetone extraction. Then, 1.5 L of pooled water were taken and fixed with 5‰ non-acetic Lugol’s iodine solution for quantification and determination of rotifers. Forty eight hours later, the undisturbed water samples were concentrated to about 50 mL and preserved with 4% formalin.
Rotifers were counted in two Sedgewick-Rafter subsamples. Rotifer densities were determined for discrete species, and identification of species was according to WANG (1961), KOSTE (1978), ZHANG and HUANG (1991), and ZHUGE (1997). Here, dominant species were those whose density amounted to 20% or more of the total rotifer community (HABERMAN, 1983).
2.3. Data Analyses
Since our data did not meet the criteria of normality, we used nonparametric statistics, using SPSS 11.5 for Windows. Nonparametric tests, based on ranks, are alternatives to parametric statistics for test-ing hypotheses about relationships and differences for variables. Friedman ranking test, which could be used for analyzing three or more repeated measurements of ordinal data (SHELDON et al., 1996), was used for analyzing the variation of rotifer density among the sites and months. The Wilcoxon signed ranks test, which was found suitable for analyzing the difference between two correlated ordinal-level measurements, was used for analyzing the difference of data between the two years (HANSSON, 2000).

Rotifers in Three Gorges Reservoir 545
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
Principal component analysis (PCA) was carried out to identify any underlying relationships between zooplankton taxa and temporal and spatial distribution, respectively. PCA has the advantage of reduc-ing the initial number of independent variables to a smaller number of composite variables which can explain a substantial percentage of the total variance (SOUISSI et al., 2000). It was performed by PC-ORD (4.0). The months or sites were variables: the last letter “F” indicated in the first year (from July 2003 to June 2004), while “S” indicated in the second year (from July 2004 to June 2005). The species data were used as cases.
Lastly, cluster analysis, which was carried out by PRIMER v.5 software, was also used to explore the spatial characteristic of the sampling sites. Abundances were Log(x + 1) transformed with no stand-ardization. Bray-Curtis similarity matrices were computed to generate clusters.
3. Results
3.1. Composition and Dominance
Altogether 76 rotifer species were recorded (Table 1), belonging to 20 families and 29 genera. 14 dominant species were observed; i.e., Polyarthra vulgaris (CARLIN, 1943), Keratella cochlearis (GOSSE, 1851), Keratella valga (EHRENBERG, 1834), Synchaeta tremula (O. F. MULLER, 1786), Synchaeta stylata (WIERZEJSKI, 1893), Trichocerca pusilla (LAUTER-BORN, 1898), Trichocerca lophoessa (GOSSE, 1886), Brachionus calyciflorus (PALLAS, 1766), Brachionus angularis (GOSSE, 1851), Brachionus forficula forficula (WIERZEJSKI, 1891), Cono-chilus unicornis (ROUSSELET, 1892), Ascomorpha ovalis (BERGENDAHL, 1892), Ploesoma truncatum (LEVANDER, 1894), and Anuraeopsis fissa (GOSSE, 1851). Polyarthra vulgaris, Keratella sp. and Synchaeta sp. were the prevailing species, whereas the other species were found to be seasonal species.
3.2. Density
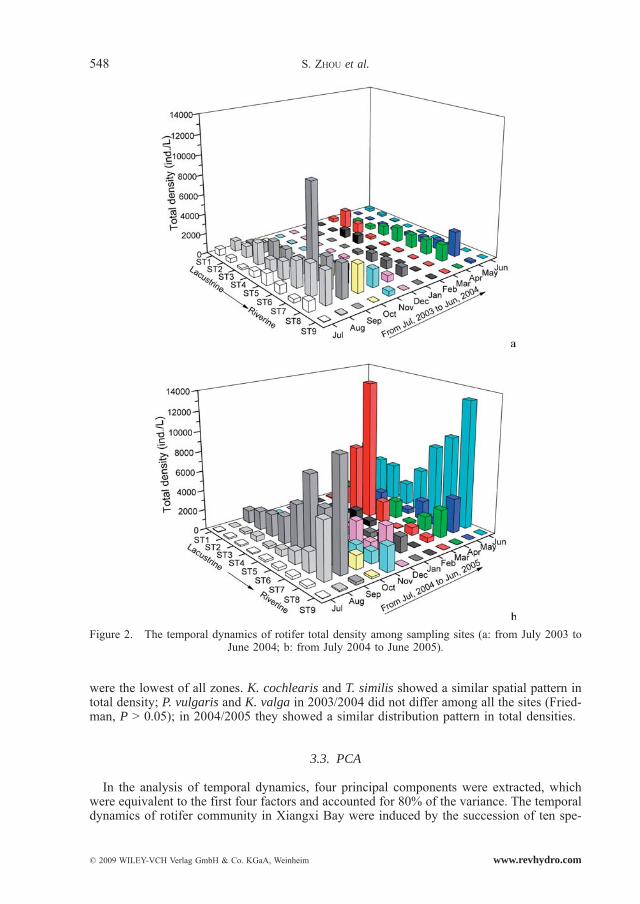
From July 2003 to June 2004, the maximum mean density, 2459 ind./L, occurred in September, while the minimum mean density, 78 ind./L, was observed in December. From July 2004 to June 2005, the maximum mean density, 5506 ind./L, occurred in June, and the minimum mean density, 186 ind./L, occurred in July. Temporal dynamics of differences in densities were significant (Friedman test, P < 0.01) (Fig. 2). Rotifer densities peaked in spring, summer and autumn and decreased to a minimum value in winter. Densities in the second year were significantly higher than those in the first year (Wilcoxon Signed Rank test, P < 0.05).
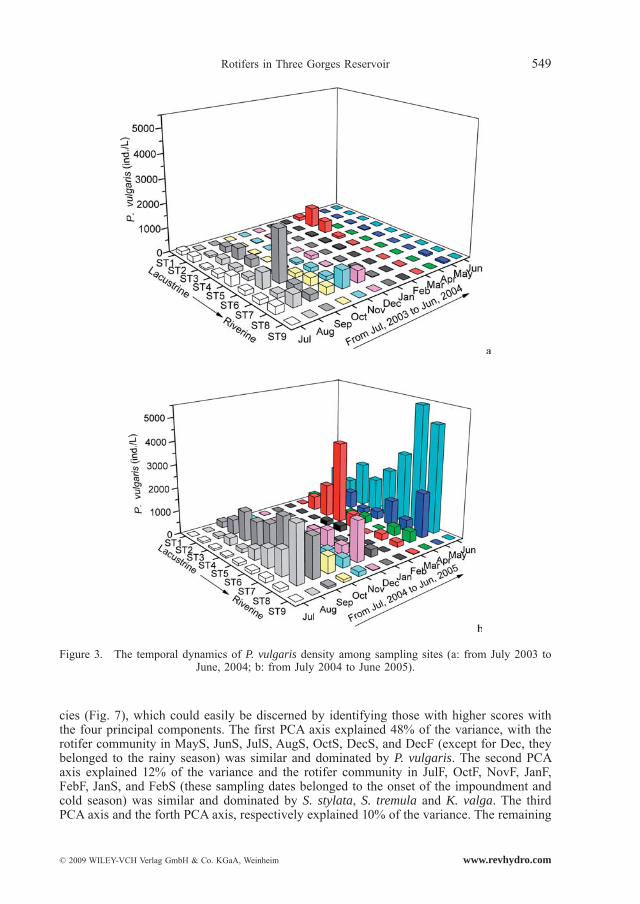
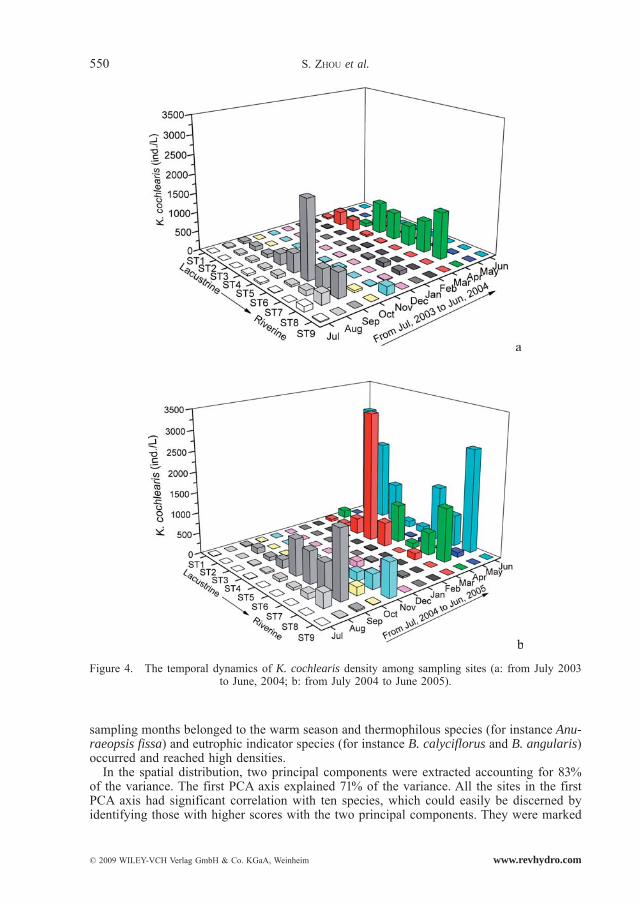
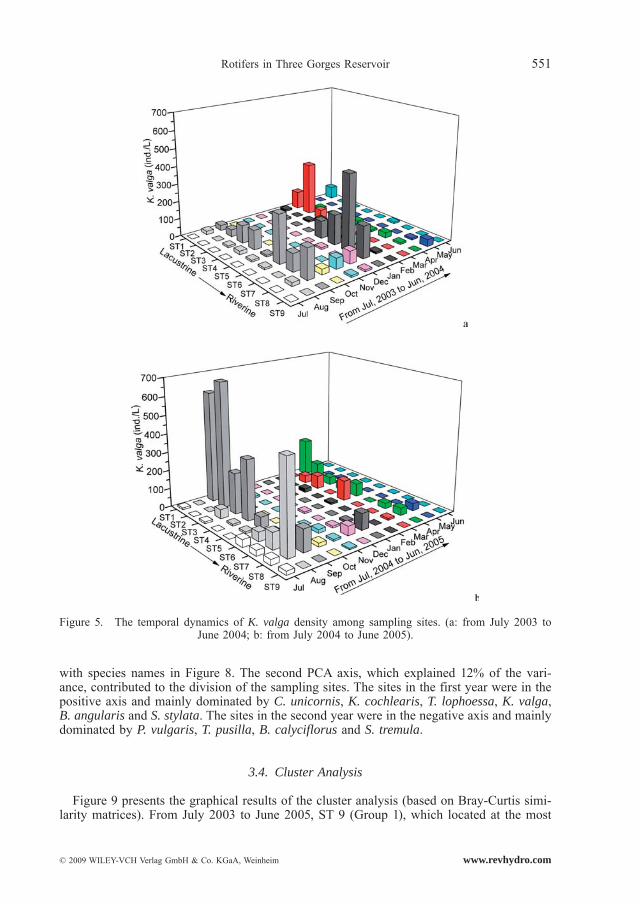
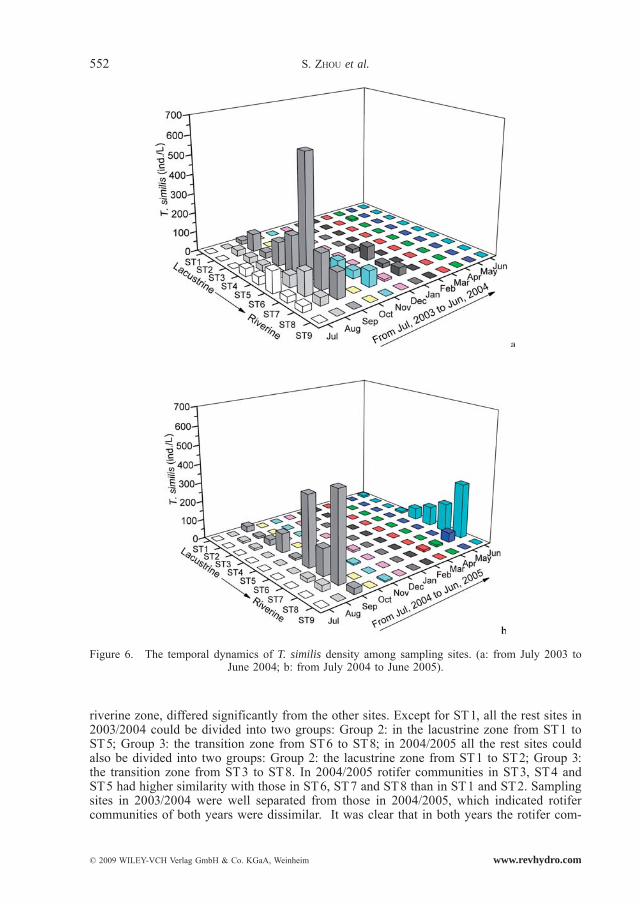
Polyarthra vulgaris, K. cochlearis, K. valga and Trichocerca similis occurred all year and had a high abundance (Fig. 3, Fig. 4, Fig. 5 and Fig. 6). Monthly fluctuations of the four species were apparent (Friedman, P < 0.01); density peaks were observed in March, April, June, August and September. Densities of P. vulgaris and K. cochlearis in 2004/2005 were significantly higher than those in 2003/2004 (Wilcoxon Signed Rank test, P < 0.001), whereas the densities of T. similis showed an opposite trend (Wilcoxon Singed Rank Test, P < 0.05). For K. valga, there was no significant difference between the two years (Wilcoxon Singed Rank Test, P > 0.05).
Significant heterogeneity of rotifer community along the longitudinal axis was observed. Based on densities, the rotifer community could be divided into three zones: the riverine zone, the transition zone and the lacustrine zone. There were no statistical differences of densities within the lacustrine zone (from ST 1 to ST 4) and within the transition zone (from ST 5 to ST 8) (Friedman, P > 0.05); but densities in the transition zone were significantly higher than in the lacustrine zone (Friedman, P < 0.05). Densities in the riverine zone (ST9)
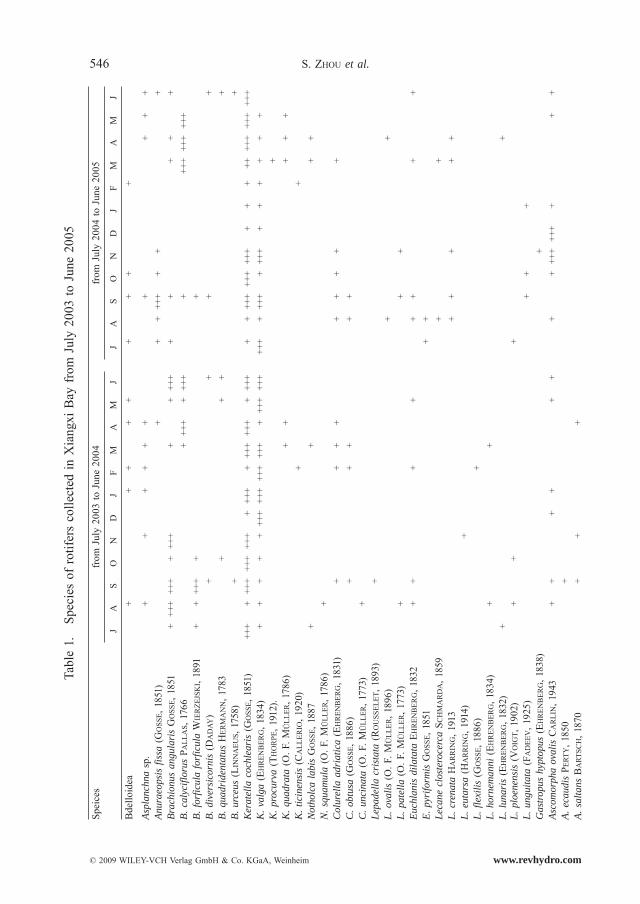
546 S. ZHOU et al.
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
Spei
ces
from
Jul
y 20
03 to
Jun
e 20
04fr
om J
uly
2004
to J
une
2005
JA
SO
ND
JF
MA
MJ
JA
SO
ND
JF
MA
MJ
Bde
lloid
ea+
++
++
++
++
Aspl
anch
na s
p.+
++
++
++
++
+An
urae
opsi
s fis
sa (
GO
SSE,
185
1)+
++
+++
++
+Br
achi
onus
ang
ular
is G
OSS
E, 1
851
+++
+++
++
+++
++
+++
++
++
+B.
cal
ycifl
orus
PA
LLA
S, 1
766
+++
++
+++
+++
+++
+++
+B.
forf
icul
a fo
rfic
ula
WIE
RZE
JSK
I, 18
91+
+++
++
+B.
div
ersi
corn
is (
DA
DAY
) +
++
+B.
qua
drid
enta
tus
HER
MA
NN
, 178
3+
++
+B.
urc
eus
(LIN
NA
EUS,
175
8)
++
Ker
atel
la c
ochl
eari
s (G
OSS
E, 1
851)
+++
+++
+++
+++
++
+++
+++
+++
++
+++
++
+++
+++
+++
++
+++
+++
+++
+++
K. v
alga
(EH
REN
BER
G, 1
834)
++
++
+++
+++
+++
+++
++
+++
+++
+++
+++
++
+++
++
++
++
K. p
rocu
rva
(TH
OR
PE, 1
912)
. +
K. q
uadr
ata
(O. F
. MÜ
LLER
, 178
6)
++
++
+K
. tic
inen
sis
(CA
LLER
IO, 1
920)
+
+N
otho
lca
labi
s G
OSS
E, 1
887
++
++
N. s
quam
ula
(O. F
. MÜ
LLER
, 178
6)
+C
olur
ella
adr
iatic
a (E
HR
ENB
ERG
, 183
1)
++
++
++
++
+C
. obt
usa
(GO
SSE,
188
6)
++
++
+C
. unc
inat
a (O
. F. M
ÜLL
ER, 1
773)
+Le
pade
lla c
rist
ata
(RO
USS
ELET
, 189
3)
+L.
ova
lis (
O. F
. MÜ
LLER
, 189
6)+
+L.
pat
ella
(O
. F. M
ÜLL
ER, 1
773)
+
++
Euch
lani
s di
lata
ta E
HR
ENB
ERG
, 183
2 +
++
++
++
+E.
pyr
iform
is G
OSS
E, 1
851
++
Leca
ne c
lost
eroc
erca
SC
HM
AR
DA
, 185
9+
+L.
cre
nata
HA
RR
ING
, 191
3 +
++
++
L. e
utar
sa (
HA
RR
ING
, 191
4)+
L. fl
exili
s (G
OSS
E, 1
886)
+
L. h
orne
man
ni (
EHR
ENB
ERG
, 183
4)+
+L.
luna
ris
(EH
REN
BER
G, 1
832)
+
+L.
plo
enen
sis
(VO
IGT,
190
2)
++
+L.
ung
uita
ta (
FAD
EEV
, 192
5)+
++
Gas
tropu
s hy
ptop
us (
EHR
ENB
ERG
, 183
8)
+As
com
orph
a ov
alis
CA
RLI
N, 1
943
++
++
++
++
+++
+++
++
+A.
eca
udis
PER
TY, 1
850
+A.
sal
tans
BA
RTSC
H, 1
870
++
+
Tabl
e 1.
Sp
ecie
s of
rot
ifers
col
lect
ed in
Xia
ngxi
Bay
fro
m J
uly
2003
to J
une
2005
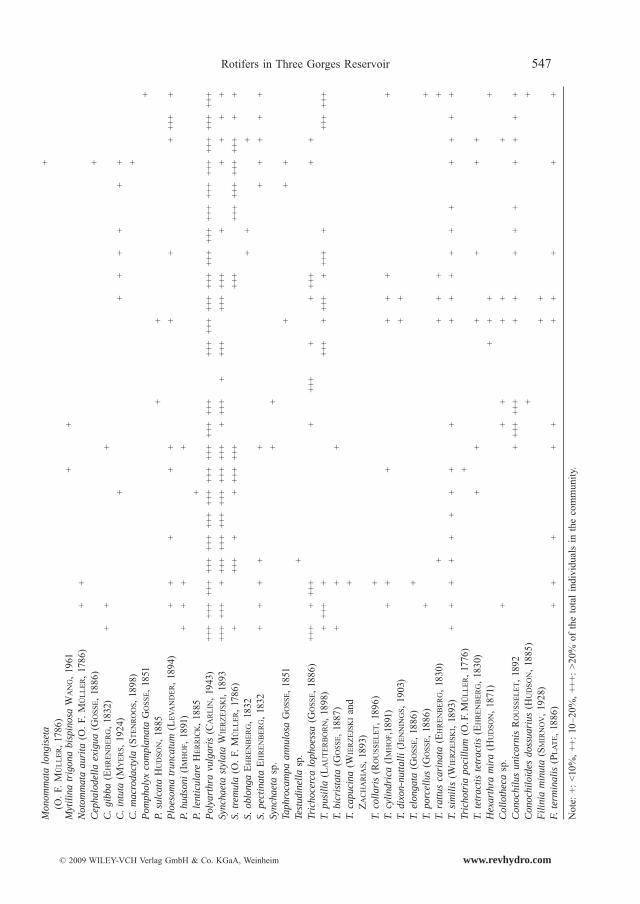
Rotifers in Three Gorges Reservoir 547
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
Mon
omm
ata
long
iset
a
(O. F
. MÜ
LLER
, 178
6)
+
Myt
ilina
trig
ona
bisp
inos
a W
AN
G, 1
961
++
Not
omm
ata
auri
ta (
O. F
. MÜ
LLER
, 178
6)
++
Cep
halo
della
exi
gua
(GO
SSE,
188
6)
+C
. gib
ba (
EHR
ENB
ERG
, 183
2)+
++
C. i
ntut
a (M
YER
S, 1
924)
++
++
++
+C
. mac
roda
ctyl
a (S
TEN
RO
OS,
189
8)+
Pom
phol
yx c
ompl
anat
a G
OSS
E, 1
851
+P.
sul
cata
HU
DSO
N, 1
885
++
Ploe
som
a tr
unca
tum
(LE
VAN
DER
, 189
4)
++
++
++
++
+++
+P.
hud
soni
(IM
HO
F, 1
891)
+
++
+P.
lent
icul
are
HER
RIC
K, 1
885
+Po
lyar
thra
vul
gari
s (C
AR
LIN
, 194
3)++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+++
+Sy
ncha
eta
styl
ata
WIE
RZE
JSK
I, 18
93
+++
+++
+++
+++
+++
+++
+++
+++
++
+++
+++
+++
+++
++
++
++
S. tr
emul
a (O
. F. M
ÜLL
ER, 1
786)
+++
++
+++
+++
+++
+++
+++
+++
+++
++
+S.
obl
onga
EH
REN
BER
G, 1
832
++
+S.
pec
tinat
a EH
REN
BER
G, 1
832
++
++
++
++
++
Sync
haet
a sp
.+
+Ta
phro
cam
pa a
nnul
osa
GO
SSE,
185
1 +
++
Test
udin
ella
sp.
+Tr
icho
cerc
a lo
phoe
ssa
(GO
SSE,
188
6)
+++
+++
++
+++
++
+++
++
T. p
usill
a (L
AU
TER
BO
RN
, 189
8)+
+++
+++
++
+++
+++
++
+++
+++
T. b
icri
stat
a (G
OSS
E, 1
887)
++
+T.
cap
ucin
a (W
IER
ZEJS
KI a
nd
ZA
CH
AR
IAS,
189
3)
+
T. c
olla
ris
(RO
USS
ELET
, 189
6)
+T.
cyl
indr
ica
(IM
HO
F,18
91)
++
++
++
+T.
dix
on-n
utta
lli (
JEN
NIN
GS,
190
3)+
+T.
elo
ngat
a (G
OSS
E, 1
886)
+
T. p
orce
llus
(GO
SSE,
188
6)+
+T.
rat
tus
cari
nata
(EH
REN
BER
G, 1
830)
+
++
++
T. s
imili
s (W
IER
ZEJS
KI,
1893
)+
++
++
++
++
++
++
++
++
++
Tric
hotr
ia p
ocill
um (
O. F
. MÜ
LLER
, 177
6)+
T. te
trac
tis te
trac
tis (
EHR
ENB
ERG
, 183
0)+
++
++
+H
exar
thra
mir
a (H
UD
SON
, 187
1)+
++
+C
ollo
thec
a sp
.+
++
++
+C
onoc
hilu
s un
icor
nis
RO
USS
ELET
, 189
2 +
+++
+++
++
++
++
++
+C
onoc
hilo
ides
dos
suar
ius
(HU
DSO
N, 1
885)
+
+Fi
linia
min
uta
(SM
IRN
OV
, 192
8)+
+F.
term
inal
is (
PLAT
E, 1
886)
+
++
++
++
++
+
Not
e: +
: <10
%, +
+: 1
0–20
%, +
++: >
20%
of
the
tota
l ind
ivid
uals
in th
e co
mm
unity
.
548 S. ZHOU et al.
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
were the lowest of all zones. K. cochlearis and T. similis showed a similar spatial pattern in total density; P. vulgaris and K. valga in 2003/2004 did not differ among all the sites (Fried-man, P > 0.05); in 2004/2005 they showed a similar distribution pattern in total densities.
3.3. PCA
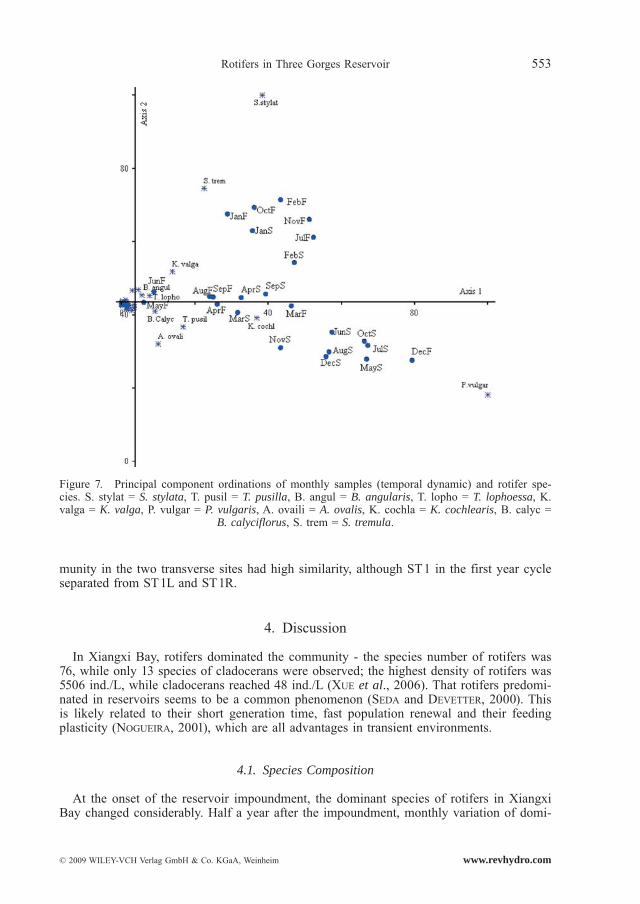
In the analysis of temporal dynamics, four principal components were extracted, which were equivalent to the first four factors and accounted for 80% of the variance. The temporal dynamics of rotifer community in Xiangxi Bay were induced by the succession of ten spe-
Figure 2. The temporal dynamics of rotifer total density among sampling sites (a: from July 2003 to June 2004; b: from July 2004 to June 2005).
Rotifers in Three Gorges Reservoir 549
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
cies (Fig. 7), which could easily be discerned by identifying those with higher scores with the four principal components. The first PCA axis explained 48% of the variance, with the rotifer community in MayS, JunS, JulS, AugS, OctS, DecS, and DecF (except for Dec, they belonged to the rainy season) was similar and dominated by P. vulgaris. The second PCA axis explained 12% of the variance and the rotifer community in JulF, OctF, NovF, JanF, FebF, JanS, and FebS (these sampling dates belonged to the onset of the impoundment and cold season) was similar and dominated by S. stylata, S. tremula and K. valga. The third PCA axis and the forth PCA axis, respectively explained 10% of the variance. The remaining
Figure 3. The temporal dynamics of P. vulgaris density among sampling sites (a: from July 2003 to June, 2004; b: from July 2004 to June 2005).
550 S. ZHOU et al.
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
sampling months belonged to the warm season and thermophilous species (for instance Anu-raeopsis fissa) and eutrophic indicator species (for instance B. calyciflorus and B. angularis) occurred and reached high densities.
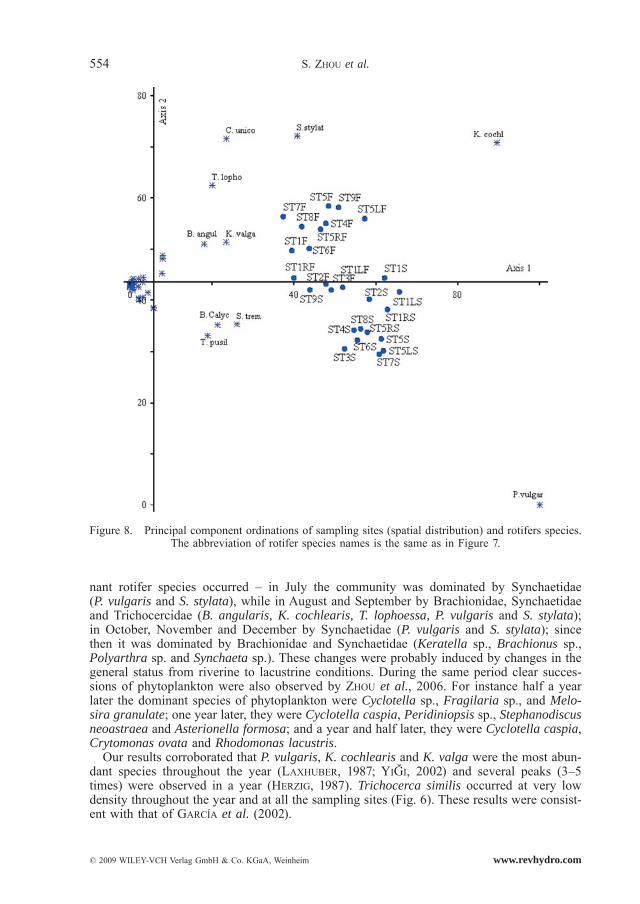
In the spatial distribution, two principal components were extracted accounting for 83% of the variance. The first PCA axis explained 71% of the variance. All the sites in the first PCA axis had significant correlation with ten species, which could easily be discerned by identifying those with higher scores with the two principal components. They were marked
Figure 4. The temporal dynamics of K. cochlearis density among sampling sites (a: from July 2003 to June, 2004; b: from July 2004 to June 2005).
Rotifers in Three Gorges Reservoir 551
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
with species names in Figure 8. The second PCA axis, which explained 12% of the vari-ance, contributed to the division of the sampling sites. The sites in the first year were in the positive axis and mainly dominated by C. unicornis, K. cochlearis, T. lophoessa, K. valga, B. angularis and S. stylata. The sites in the second year were in the negative axis and mainly dominated by P. vulgaris, T. pusilla, B. calyciflorus and S. tremula.
3.4. Cluster Analysis
Figure 9 presents the graphical results of the cluster analysis (based on Bray-Curtis simi-larity matrices). From July 2003 to June 2005, ST 9 (Group 1), which located at the most
Figure 5. The temporal dynamics of K. valga density among sampling sites. (a: from July 2003 to June 2004; b: from July 2004 to June 2005).
552 S. ZHOU et al.
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
riverine zone, differed significantly from the other sites. Except for ST 1, all the rest sites in 2003/2004 could be divided into two groups: Group 2: in the lacustrine zone from ST 1 to ST 5; Group 3: the transition zone from ST 6 to ST 8; in 2004/2005 all the rest sites could also be divided into two groups: Group 2: the lacustrine zone from ST 1 to ST 2; Group 3: the transition zone from ST 3 to ST 8. In 2004/2005 rotifer communities in ST 3, ST 4 and ST 5 had higher similarity with those in ST 6, ST 7 and ST 8 than in ST 1 and ST 2. Sampling sites in 2003/2004 were well separated from those in 2004/2005, which indicated rotifer communities of both years were dissimilar. It was clear that in both years the rotifer com-
Figure 6. The temporal dynamics of T. similis density among sampling sites. (a: from July 2003 to June 2004; b: from July 2004 to June 2005).
Rotifers in Three Gorges Reservoir 553
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
Figure 7. Principal component ordinations of monthly samples (temporal dynamic) and rotifer spe-cies. S. stylat = S. stylata, T. pusil = T. pusilla, B. angul = B. angularis, T. lopho = T. lophoessa, K. valga = K. valga, P. vulgar = P. vulgaris, A. ovaili = A. ovalis, K. cochla = K. cochlearis, B. calyc =
B. calyciflorus, S. trem = S. tremula.
munity in the two transverse sites had high similarity, although ST 1 in the first year cycle separated from ST 1L and ST 1R.
4. Discussion
In Xiangxi Bay, rotifers dominated the community - the species number of rotifers was 76, while only 13 species of cladocerans were observed; the highest density of rotifers was 5506 ind./L, while cladocerans reached 48 ind./L (XUE et al., 2006). That rotifers predomi-nated in reservoirs seems to be a common phenomenon (SEDA and DEVETTER, 2000). This is likely related to their short generation time, fast population renewal and their feeding plasticity (NOGUEIRA, 2001), which are all advantages in transient environments.
4.1. Species Composition
At the onset of the reservoir impoundment, the dominant species of rotifers in Xiangxi Bay changed considerably. Half a year after the impoundment, monthly variation of domi-
554 S. ZHOU et al.
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
nant rotifer species occurred – in July the community was dominated by Synchaetidae (P. vulgaris and S. stylata), while in August and September by Brachionidae, Synchaetidae and Trichocercidae (B. angularis, K. cochlearis, T. lophoessa, P. vulgaris and S. stylata); in October, November and December by Synchaetidae (P. vulgaris and S. stylata); since then it was dominated by Brachionidae and Synchaetidae (Keratella sp., Brachionus sp., Polyarthra sp. and Synchaeta sp.). These changes were probably induced by changes in the general status from riverine to lacustrine conditions. During the same period clear succes-sions of phytoplankton were also observed by ZHOU et al., 2006. For instance half a year later the dominant species of phytoplankton were Cyclotella sp., Fragilaria sp., and Melo-sira granulate; one year later, they were Cyclotella caspia, Peridiniopsis sp., Stephanodiscus neoastraea and Asterionella formosa; and a year and half later, they were Cyclotella caspia, Crytomonas ovata and Rhodomonas lacustris.
Our results corroborated that P. vulgaris, K. cochlearis and K. valga were the most abun-dant species throughout the year (LAXHUBER, 1987; YIĞI, 2002) and several peaks (3–5 times) were observed in a year (HERZIG, 1987). Trichocerca similis occurred at very low density throughout the year and at all the sampling sites (Fig. 6). These results were consist-ent with that of GARCÍA et al. (2002).
Figure 8. Principal component ordinations of sampling sites (spatial distribution) and rotifers species. The abbreviation of rotifer species names is the same as in Figure 7.

Rotifers in Three Gorges Reservoir 555
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
Except for the most abundant species, some species in Xiangxi Bay occurred in special months. For instance, B. angularis, which is referred to as a warm stenothermal species (BERZINS and PEJLER, 1989) and A. fissa and Hexarthra mira, which occurred in warm months (HABERMAN, 1995; FERNÁNDEZ-ROSADO and LUCENA, 2001), were found in high water temperature months (from March to September). Notholca labis, a cold-stenothermal species, was observed in winter.
4.2. Temporal Variation
Similar with studies of FERNÁDEZ-ROSADO and LUCENA (2001) and ROMARE et al. (2005), rotifer species composition and population density in Xiangxi Bay changed significant-ly with time (Fig. 2 and Fig. 7). The community in 2003/2004 was dissimilar to that in 2004/2005 (see Fig. 8), and mostly caused by the succession of dominant species. Cluster analysis results (Fig. 9) also confirmed that rotifer community structure in the second year was significantly different from that in the first year.
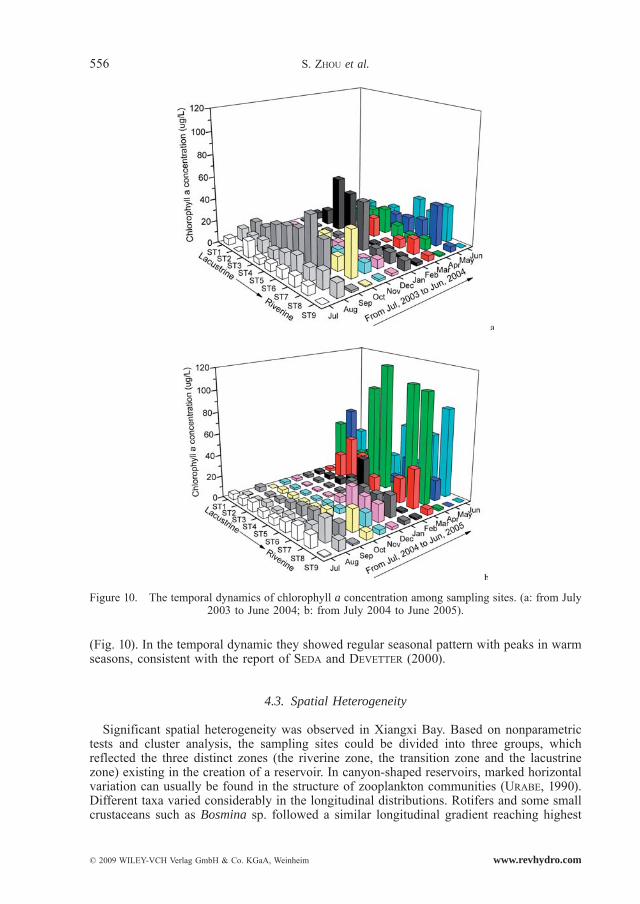
Our results corroborate the findings that reservoir development can result in the increase of zooplankton community biomass and a rapid shift in species composition (CAMPBELL et al., 1998). Rotifer densities in the second year were significantly higher than those in the first year. This was due to a combination of enhanced phytoplankton populations and longer water retention times of the impoundment compared with the original system. After the cre-ation of the Three Gorges Reservoir, water retention times were longer (the current velocity decreased from 0.43–0.92 m · s–1 to 0.0020–0.0041 m.s–1); which benefited the development of rotifers (ZHOU et al., 2008); as the food resource of zooplankton (ZHOU et al., 2007), phy-toplankton (as indicated by chlorophyll a) reached a high level (2.63–92.27 μg/L) (Fig. 10) and was significantly higher in the second year (a mean of 15.40 μg/L) compared to that in the first year (a mean of 11.20 μg/L) (Wilcoxon Signed Rank Test, P < 0.05), which showed an opposite pattern against STRAšKRABOVÁ et al. (2005). The different pattern could be explained by serious diatom blooms occurred in the second year in March and April
ST9FST9SST1RSST2SST1SST1LSST3SST4SST5LSST7SST5SST5RSST6SST8SST1FST6FST7FST8FST4FST5FST5LFST5RFST1RFST2FST1LFST3F
00108060402
Similarity (%)
Figure 9. The cluster analysis of the sampling sites, based on Bray-Curtis similarity matrices.
556 S. ZHOU et al.
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
(Fig. 10). In the temporal dynamic they showed regular seasonal pattern with peaks in warm seasons, consistent with the report of SEDA and DEVETTER (2000).
4.3. Spatial Heterogeneity
Significant spatial heterogeneity was observed in Xiangxi Bay. Based on nonparametric tests and cluster analysis, the sampling sites could be divided into three groups, which reflected the three distinct zones (the riverine zone, the transition zone and the lacustrine zone) existing in the creation of a reservoir. In canyon-shaped reservoirs, marked horizontal variation can usually be found in the structure of zooplankton communities (URABE, 1990). Different taxa varied considerably in the longitudinal distributions. Rotifers and some small crustaceans such as Bosmina sp. followed a similar longitudinal gradient reaching highest
Figure 10. The temporal dynamics of chlorophyll a concentration among sampling sites. (a: from July 2003 to June 2004; b: from July 2004 to June 2005).

Rotifers in Three Gorges Reservoir 557
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
densities in the transition zone and lower in the lacustrine zone (URABE and MURANO, 1986; URABE, 1989). Copepods, on the other hand, were more abundant in the lacustrine zone (ZANATA and ESPÍNDOLA, 2002). The longitudinal distribution of rotifers in Xiangxi Bay was similar with the research of ZANATA and ESPÍNDOLA (2002) and densities were the highest in the transition zone, followed by the lacustrine and then in the riverine zone. Rotifer density was greater in the transition zone, which was often observed by other authors (SEDA and DEVETTER, 2000; ZANATA and ESPÍNDOLA, 2002). This may be due to the transition zone hav-ing a lower current velocity than the riverine zone and supporting autochthonous organisms instead of allochthonous ones. The densities of rotifers in the lacustrine zone were lower than those in the transition zone. This was likely due to the more abundant large crustaceans in the lacustrine zone (Daphnia sp. and Diaphanosoma sp.), which are thought to be effective competitors or predators of rotifer community.
From the cluster results, rotifer communities in transects of Xiangxi Bay were evenly distributed, and all the transverse sites were grouped together. No significant heterogeneity in the two transects can be explained by reservoir topography, water movements and lack of plants along the shores (FERNÁDEZ-ROSADO and LUCENA, 2001). Spatial heterogeneity in Xiangxi Bay occurred mainly in the longitudinal direction. All rotifer communities could be divided into three groups, corresponding to the riverine, the transition and the lacustrine zone, respectively.
5. Acknowledgements
Special thanks are expressed to two anonymous reviewers and R. RAZAVI of Queen’s University and N. WU of Kiel University for their constructive comments on the manuscript. We thank D. LI, R. LIU, M. SHAO, G. ZHOU and H. TANG for their assistances with samples collection. Foundation item: the National Natural Science Foundation of China (30330140, 40671197) and the Key Project of Knowledge Innovation Program of the CAS (KZCX2-YW-427).
6. References
BERNOT, R. J., W. K. DODDS, M. C. QUIST and C. S. GUY, 2004: Spatial and temporal variability of zooplankton in a great plains reservoir. – Hydrobiologia 525: 101–112.
BERZINS, B. and B. PEJLER, 1989: Rotifer occurrence in relation to temperature. – Hydrobiologia 175: 223–231.
BRANCO, C. W. C., M. I. A. ROCHA, G. F. S. PINTO, G. A. GÔMARA and R. D. FILIPPO, 2002: Limnologi-cal features of Funil Reservoir (R.J., Brazil) and indicator properties of rotifers and cladocerans of the zooplankton community. – Lakes Reserv. Res. Manage. 7: 87–92.
CAJANDER, V. R., 1983: Production of planktonic Rotatoria in Ormajärvi, an eutrophicated lake in south-ern Finland. – Hydrobiologia 104: 329–333.
CAMPBELL, C. E., R. KNOECHEL and D. COPEMAN, 1998: Evaluation of factors related to increased zooplankton biomass and altered species composition following impoundment of a Newfoundland reservoir. – Can. J. Fish. Aquat. Sci. 55: 230–238.
CLARK, D. R., K. V. AZEM and G. C. HAYS, 2001: Zooplankton abundance and community structure over a 4000 km transect in the north-east Atlantic. – J. Plankton Res. 23: 365–372.
FERNÁDEZ-ROSADO, M. J. and J. LUCENA, 2001: Space-time heterogeneities of the zooplankton distribu-tion in La Concepción reservoir (Istán Málaga; Spain). – Hydrobiologia 455: 157–170.
GARCÍA, P. R., S. NANDINI, S. S. S. SARMA, E. R. VALDERRAMA, I. CUESTA and M. D. HURTADO, 2002: Seasonal variations of zooplankton abundance in the freshwater reservoir Valle de Bravo (Mexico). – Hydrobiologia 467: 99–108.
HABERMAN, J., 1983: Comparative analysis of plankton rotifer biomass in large Estonian lakes. – Hydro biologia 104: 293–296.

558 S. ZHOU et al.
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
HABERMAN, J., 1995: Dominant rotifer of Võrtsjärv (Estonia). – Hydrobiologia 313/314: 313–317.HANSSON, L. A., 2000: Synergistic effects of food chain dynamics and induced behavioral responses in
aquatic ecosystems. – Ecology 81: 842–851.HERZIG, A., 1987: The analysis of planktonic rotifer populations: A plea for long-term investigations. –
Hydrobiologia 147: 163–180.KOSTE, W., 1978: Rotatoria. Die Rädertiere Mitteleuropas (Überordnung Monogononta), Bestimmungs-
werk begründet von Max Voigt. Vol. 2. Gebrüder Borntraeger, Stuttgart. LAXHUBER, R., 1987: Abundance and distribution of pelagic rotifers in a cold, deep oligotrophic alpine
lake (Königssee). – Hydrobiologia 147: 189–196.MAESTRE, F. T., A. ESCUDERO, I. MARTINEZ, C. GUERRERO and A. RUBIO, 2005: Does spatial pattern
matter to ecosystem functioning? Insights from biological soil crusts. – Functional Ecology 19: 566–573.
MAŠÍN, M., J. JEZBERA, J. NEDOMA, V. STRAŠKRABOVÁ, J. HEIGZLAR and K. ŠIMEK, 2003: Changes in bacterial community composition and microbial activities along the longitudinal axis of two canyon-shaped reservoirs with different inflow loading. – Hydrobiologia 504: 99–113.
MATSUMURA-TUNDISI, T. and J. G. TUNDISI, 2005: Plankton richness in a eutrophic reservoir (Barra Bonita Reservoir, SP, Brazil). – Hydrobiologia 542: 367–378.
NOGUEIRA, M. G., 2001: Zooplankton composition, dominance and abundance as indicators of envi-ronmental compartmentalization in Jurumirim Reservoir (Paranapanema River), São Paulo, Brazil. – Hydrobiologia 455: 1–18.
PINEL-ALLOUL, B., C. GUAY, N. ANGELI, P. LEGENDRE, P. DUTILLEUL, G. BALVAY, D. GERDEAUX and J. GUILLARD, 1999: Large-scale spatial heterogeneity of macrozooplankton in Lake of Geneva. – Can. J. Fish Aquat. Sci. 56: 1437–1451.
ROMARE, P., S. BERG, T. LAURIDSEN and E. JEPPESEN, 2003: Spatial and temporal distribution of fish and zooplankton in a shallow lake. – Freshw. Biol. 48: 1353–1362.
ROMARE, P., D. E. SCHINDLER, M. D. SCHEUERELL, J. M. SCHEUERELL, A. H. LITT and J. H. SHEPHERD, 2005: Variation in spatial and temporal gradients in zooplankton spring development: the effect of climatic factors. – Freshw. Biol. 50: 1007–1021.
ROSENZWEIG, M. L., 1991: Habitat selection and population interactions: the search for mechanisms. – American Naturalist 137: 5–28.
SEDA, J. and M. DEVETTER, 2000: Zooplankton community structure along a trophic gradient in a canyon-shaped dam reservoir. – J. Plankton Res. 22: 1829–1840.
SHELDON, M. R., M. J. FILLGAW and W. D. THOMPSON, 1996: The use and interpretation of the fried-man test in the analysis of ordinal-scale data in repeated measures designs. – Physiotherapy Research International 1: 221–228.
SOUISSI, S., O. D. YAHIA-KÉFI and M. N. D. YAHIA, 2000: Spatial characterization of nutrient dynam-ics in the Bay of Tunis (south-western Mediterranean) using multivariate analyses: consequences for phyto- and zooplankton distribution. – J. Plankton Res. 22: 2039–2059.
STRAŠKRABA, M. and J. G. TUNDISI, 1999: Guidelines of lake management Volume 9 Reservoir water quality management. – International Lake Environment Committee, pp. 4–5.
STRAšKRABOVá, V., K. ŠIMEK and J. VRBA, 2005: Long-term development of reservoir ecosystems-changes in pelagic food webs and their microbial component. – Limnetica 24: 9–20.
TANG, T., Q. H. CAI, R. Q. LIU, D. F. LI and Z. C. XIE, 2002: Distribution of Epilithic algae in the Xiangxi Rive System and their relationships with environmental factors. – J. Freshw. Ecol. 17: 345–352.
URABE, J. and M. MURANO, 1986: Seasonal and horizontal variations in the zooplankton community of Ogochi Reservoir, Tokyo. – Bulletin of Plankton Society of Japan 33: 101–112.
URABE, J., 1989: Relative importance of temporal and spatial heterogeneity in the zooplankton com-munity of an artificial reservoir. – Hydrobiologia 184: 1–6.
URABE, J., 1990: Stable horizontal variation in the zooplankton community structure of a reservoir maintained by predation and competition. – Limno. Oceanogr. 35: 1703–1717.
WANG, J. J., 1961: The freshwater rotifer of China. Science Press, Beijing (in Chinese).XUE, J. Z., X. Q. HAN, Q. H. CAI and J. K. LIU, 2006: Composition and vertical distribution of cladocer-
ans in Xiangxi Bay of Three-Gorge Reservoir. – Acta Hydrobiol. Sinica 30: 120–122 (in Chinese with English abstract).
YIGI, T. S., 2002: Seasonal Fluctuation in the Rotifer Fauna of Kesikk.pr. Dam Lake (Ankara, Turkey). – Turk. J. Zool. 26: 341–348.

Rotifers in Three Gorges Reservoir 559
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.revhydro.com
ZANATA, L. H. and E. L. G. ESPÍNDOLA, 2002: Longitudinal processes in Salto Grande Reservoir (Ameri-cana, SP, Brazil) and its influence in the formation of compartment system. – Braz. J. Biol. 62: 347–361.
ZHANG, Z. S. and X. F. HUANG, 1991: Methods for study on freshwater plankton. Science Press, Beijing (in Chinese).
ZHOU, G. J., Q. J. KUANG, Z. Y. HU and Q. H. CAI, 2006: Study on the succession of algae and the trend of water-bloom occurred in Xiangxi Bay. – Acta Hydrobiol. Sinica 30: 42–46 (in Chinese with English abstract).
ZHOU, S. C., X. F. HUANG and Q. H. CAI, 2007: Vertical distribution and migration of planktonic rotifers in Xiangxi Bay of the Three Gorges Reservoir, China. – J. Freshw. Ecol. 22: 441–449.
ZHOU, S. C., T. TANG, N. C. WU, X. C. FU and Q. H. CAI, 2008: Impact of a small dam on riverine zooplankton. – Internat. Rev. Hydrobiol. 93: 297–311.
ZHUGE, Y., 1997: Studies on taxonomy and distribution of rotifera in typical zones of China. Ph. D. Dis-sertation, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, P.R. China (in Chinese with English abstract).
Manuscript received September 8th, 2008; revised May 29th, 2009; accepted June 17th, 2009