Embed Size (px)
Citation preview

Hindawi Publishing CorporationThe Scientific World JournalVolume 2013 Article ID 305081 13 pageshttpdxdoiorg1011552013305081
Research ArticleStudies of the Interaction between Isoimperatorin and HumanSerum Albumin by Multispectroscopic Method Identification ofPossible Binding Site of the Compound Using Esterase Activityof the Protein
Samira Ranjbar1 Yalda Shokoohinia23 Sirous Ghobadi4 Nooshin Bijari1
Saeed Gholamzadeh5 Nastaran Moradi1 Mohammad Reza Ashrafi-Kooshk1
Abbas Aghaei6 and Reza Khodarahmi13
1 Nano Drug Delivery Research Center Kermanshah University of Medical Sciences Kermanshah 6734667149 Iran2Novel Drug Delivery Research Center School of Pharmacy Kermanshah University of Medical Sciences Kermanshah Iran3Department of Pharmacognosy and Biotechnology Faculty of Pharmacy Kermanshah University of Medical SciencesKermanshah 6734667149 Iran
4Department of Biology Faculty of Science Razi University Kermanshah Iran5 Isfahan Pharmaceutical Sciences Research Center School of Pharmacy and Pharmaceutical SciencesIsfahan University of Medical Sciences Isfahan Iran
6Department of Epidemiology Kermanshah University of Medical Sciences Kermanshah Iran
Correspondence should be addressed to Reza Khodarahmi rkhodarahmimbrcacir
Received 13 August 2013 Accepted 10 September 2013
Academic Editors H Gronemeyer and Y Mishina
Copyright copy 2013 Samira Ranjbar et alThis is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Isoimperatorin is one of the main components of Prangos ferulacea as a linear furanocoumarin and used as anti-inflammatoryanalgesic antispasmodic and anticancer drug Human serum albumin (HSA) is a principal extracellular protein with a highconcentration in blood plasma and carrier for many drugs to different molecular targets Since the carrying of drug by HSA mayaffect on its structure and action we decided to investigate the interaction between HSA and isoimperatorin using fluorescenceand UV spectroscopy Fluorescence data indicated that isoimperatorin quenches the intrinsic fluorescence of the HSA via astatic mechanism and hydrophobic interaction play the major role in the drug binding The binding average distance betweenisoimperatorin and Trp 214 of HSA was estimated on the basis of the theory of Forster energy transfer Decrease of proteinsurface hydrophobicity (PSH) was also documented upon isoimperatorin binding Furthermore the synchronous fluorescencespectra show that the microenvironment of the tryptophan residues does not have obvious changes Site marker compettive andfluorescence experiments revealed that the binding of isoimperatorin to HSA occurred at or near site I Finally the binding detailsbetween isoimperatorin and HSA were further confirmed by molecular docking and esterase activity inhibition studies whichrevealed that drug was bound at subdomain IIA
1 Introduction
Interaction between drugs and plasma proteins is an impor-tant pharmacological parameter which has a strong influ-ence on their pharmacodynamic behaviors and affects thedistribution and elimination of drug as well as the struc-ture and physiological action of carrier proteins [1 2]
However problems associated with adsorption distributionmetabolism and elimination (ADME) add considerably tothe complexity and cost of the development of new drugsand are driving the search for techniques to optimise thesecharacteristics at an early stage in the design process Oneof the most important factors affecting the distribution andthe free active concentration of many administered drugs is
2 The Scientific World Journal
OO
O
O
Figure 1 Structure of isoimperatorin
binding affinity for human serum albumin (HSA) HSA isthe most abundant drug carrier protein which is also foundin tissues and bodily secretions throughout the body theextravascular protein comprises 60 of the total albuminHSA is a widely studied protein because its primary structureis well known and its tertiary structure has been determinedby X-ray crystallography [3ndash6] It has an important rolein maintaining the colloidal osmotic pressure in bloodit transports and distributes exogenous and endogenousmolecules and metabolites such as nutrients hormones fattyacids and many diverse drugs [6 7] This heart-shapedprotein which has a net charge of minus15 at neutral pH and anisoelectric point of about 5 consists of 585 amino acid residuesand is composed of three structurally similar domains (III and III) each containing two subdomains (A and B)and being stabilized by 17 disulphide bridges [4ndash8] Crystalstructure analyses have revealed that HSA has binding sitesfor aromatic and heterocyclic ligands within two hydropho-bic pockets in subdomains IIA (commonly referred to asSudlowrsquos site I warfarin-binding site) and IIIA (commonlyreferred to as Sudlowrsquos site II indolebenzodiazepine site)Both hydrophobic and electrostatic interactions play a majorrole in controlling the affinity towards drug binding forsites I and II for site I mainly hydrophobic interactions aredominant whereas for site II a combination of hydrophobichydrogen bonding and electrostatic interactions all play acrucial role The sole tryptophan residue (Trp 214) of HSAis in subdomain IIA (site I) [6ndash8]
Coumarins are widely distributed in nature and arefound in all parts of plants These compounds are especiallycommon in grasses orchids citrus fruits and legumes [9]Being so abundant in nature coumarins make up an impor-tant part of the human diet Based on chemical structurethey can be broadly classified as (a) simple coumarins(b) furanocoumarins of the linear or angular type and(c) pyranocoumarins of the linear or angular type [9ndash11]Isoimperatorin (4-[(3-methyl-2-butenyl) oxy]-7H-furo [32-g] [1] benzopyran-7-one) [12] (Figure 1) [13] is one of themaincomponents of Prangos ferulacea as a linear furanocoumarin[14] The plant has previously shown antispasmodic effects[15 16] and proved to contain coumarins and terpenoids [17ndash19] Isoimperatorin has attracted increasing interest due to itsanti-inflammatory analgesic antispasmodic and anticancer
activities and treatment of Alzheimerrsquos disease [9 10 20]With the increasing importance of isoimperatorin in humanhealth it is necessary to evaluate its effect on biomoleculesfor example proteins as a chemical entity [10 20] Interactionbetween serum albumin and many furocoumarins has beeninvestigated successfully but detailed investigations of theinteraction of HSA with isoimperatorin are not available Soin this study we investigated the effect of isoimperatorin onthe structure and action of HSA bymeans of various spectro-scopic techniquesTheUV-vis spectroscopy and intrinsic andextrinsic fluorometry methods were used for elucidating themode of binding and probable structural alterations of HSAupon drug bindingThe characteristics of the binding that isbinding constant number of binding sites and and nature ofbinding forces as well as the effects of the complexation onthe protein conformation were determined In addition thethermodynamic and Forsterrsquos parameters associated with thebinding process were also calculated
2 Materials and Methods
21 Chemicals The pure warfarin was obtained as a gen-erous gift from Dr S Kashanian (Razi University) 1-Anilinonaphthalene-8-sulfonate (ANS) was purchased fromSigma Chemical Co (St Louis Mo USA)The other reagentswere of analytical grade from Merck (Darmstadt Germany)All of the solutions were prepared in double-distilled waterand all of the experiments were carried out in 50mMphosphate pH 75 as the buffer All of themeasurements weredone in triplicate
22 Extraction Isolation and Identification of IsoimperatorinRoots of Prangos ferulacea L were collected from Yasujin Kohgiluyeh-Boyer-Ahmad province Iran in June 2011The plant was identified at the Botany Department of YasujUniversity Yasuj and compared to the voucher specimen(No 2408) at the Herbarium of School of Pharmacy IsfahanUniversity of Medical Sciences Isfahan Iran
Dried ground roots were extracted with acetone for twodays (5 L times 4) The extract was concentrated to bear aviscous mass which was then put in minus20∘C for two daysand chill filtered in which the filtrate resulted in a solidmass after drying The latter was fractioned by vacuumliquid chromatography (Silica gel MESH 230ndash400) using agradient of heptane ethylacetate (H EtOAc) from 100 0 to0 100 to afford several fractions Third fraction (H EtOAc7 3) rendered a mixture of two coumarins in which furtherpurification via chromatographies (H EtOAc 8 2 to 7 3)resulted in pure isoimperatorin
23 UV-vis Measurements The UV-vis spectra are recordedat room temperature on a Cary Eclips (Varian) spectropho-tometer equipped with 10 cm quartzecells The final drugconcentrations and constant protein concentration were 04ndash8 120583M and 3 120583M respectively The exact concentration ofHSA was determined spectrophotometrically using molarextinction coefficient of 35700Mminus1 cmminus1 [21]
The Scientific World Journal 3
24 Intrinsic Fluorescence Measurements Fluorescence mea-surements were performed on a Cary Eclipse (Varian) spec-trophotometer with jacketed cell holders in which tem-perature was adjusted by an external thermostated watercirculationThe excitation and emission wavelengths were setat 295 nm (to avoid the contribution from tyrosine residues[22]) and 300ndash570 nm respectively with the excitation andemission slit widths of 5 and 10 nm respectively Fluorescenceemission spectra of the HSA were measured in the absenceand presence of the various concentrations of isoimperatorin(02 04 06 08 1 15 2 25 3 35 4 5 6 8 and 10 120583M)Thefinal enzyme concentration was 3 120583M
Synchronous fluorescence spectra of HSA with variousconcentrations of isoimperatorin were obtained (Δ120582 = 15 nmandΔ120582 = 60 nm) with the excitation and emission slit widthsof 5 and 10 nm respectively
241 The Inner Filter Effect The inner filter effect can bea problem for any fluorescencemeasurement especiallywherean absorbing component is being titrated into the cuvetteDuring a fluorescence titration experiment at the wavelengthof excitation or at the wavelengths used to follow an emissionthe absorption of the molecule added resulted in a spuriousdecrease in the observed fluorescence intensity [23] In thesteady-state fluorescence experiments absorption spectrumof isoimperatorin overlapswith the excitation and emission ofHSAThus it is very important to subtract such an effect fromthe raw quenching data The fluorescence data was correctedfor inner filter effect using
119865corr =119865obs antilog (119860em + 119860ex)
2
(1)
where 119865corr and 119865obs are the corrected and observed flu-orescence intensity 119860ex and 119860em are the optical densityof the sample at the excitation and emission wavelengthsrespectively
242 Determination of Quenching Mechanism In order toclarify the mechanism of HSA fluorescence quenching byisoimperatorin the quenching experiments were carried outat 298 303 308 and 313 K where HSA does not undergoany considerable thermal denaturation The decrease influorescence intensity at 120582max was analyzed using the well-known Stern-Volmer equation [24]
119865
0
119865
= 1 + 119870SV [119876] (2)
where 1198650and 119865 are the fluorescence intensities in the absence
and presence of quencher (isoimperatorin) respectively the119870SV is the Stern-Volmer quenching constant and [119876] is theconcentration of the compound isoimperatorin
243 Determination of Association Constant and Number ofBinding Sites The association constant (119870
119887) and number of
binding sites (119899) for binding of small molecules to set of
equivalent sites on amacromolecule can be determined usingthe modified Stern-Volmer equation
log 119865
0minus 119865
119865
= log119870119887+ 119899 log [119876] (3)
where 1198650and 119865 are the fluorescence intensities of HSA in
the absence and presence of the quencher (isoimperatorin)respectively 119870
119887is the association constant and 119899 is the
number of binding sites per HSA [25]The values of 119899 and119870119887
were obtained from the slope and intercept of the modifiedStern-Volmer plot respectively
244 Thermodynamic Analysis of the Binding Process Inorder to explain the nature of the interaction betweenisoimperatorin and HSA the association constant of HSA-isoimperatorin complex was determined at four differenttemperatures (298 303 308 and 313 K)The thermodynamicparameters of binding entropy change (Δ119878∘) and enthalpychange (Δ119867∘) were obtained from vanrsquot Hoff equation
ln 119870119887=
minusΔ119867
∘
119877119879
+
Δ119878
∘
119877
(4)
where119870119887is the association constant at the given temperature
and119877 is the universal gas constantThe values ofΔ119878∘ andΔ119867∘can be obtained from the intercept and slope of the vanrsquot Hoffplot respectively and the free energy change (Δ119866∘) can beestimated by using the Gibbs equation [26]
Δ119866
∘
= Δ119867
∘
minus 119879Δ119878
∘
(5)
25 Energy Transfer between HSA and Isoimperatorin Thedistance between the acceptor (ligand) and the donor (tryp-tophan residues in the protein) 119903 according to Forsterrsquosnonradiation energy transfer theory could be calculated bythe following equation [21]
119864 = 1 minus (
119865
119865
0
) =
119877
6
0
119877
6
0
+ 119903
6
0
(6)
where 119864 is the efficiency of energy transfer between donorand acceptor 119865 and 119865
0are the fluorescence intensities of
donor in the presence and absence of acceptor respectivelyand 119877
0is a critical distance at which the efficiency of energy
transfer is 50 The value of 1198770was calculated using
119877
6
0
= 88 times 10
minus25
119870
2
119873
minus4
Φ119869(7)
where 1198702 is the spatial orientation factor describing thegeometry of the donor and acceptor dipoles 119873 is theaverage refractive index of the mediumΦ is the fluorescencequantum yield of the donor in the absence of acceptor and 119869expresses the degree of spectral overlap between the emissionspectrum of the donor and the absorption spectrum of theacceptor 119869 can be given by
119869 = sum119865 (120582) 120576 (120582) 120582
4
Δ120582 (8)
where 119865(120582) is the fluorescence intensity of the donor atwavelength 120582 and 120576(120582) is the molar absorption coefficient
4 The Scientific World Journal
of the acceptor at 120582 So 119869 can be evaluated by integratingthe overlap between emission spectrum of the donor andabsorption spectrum of the acceptor [27]
The absorption spectrum of isoimperatorin (10 120583M) wasrecorded in the range of 222ndash450 nmThe emission spectrumof HSA (10 120583M) was also recorded in the range of 222ndash450 nm with excitation and emission slit widths of 5 and5 nm respectivelyThe overlap of the absorption spectrum ofisoimperatorin with the fluorescence emission spectrum ofHSA was used to calculate the efficiency of energy transfer
26 Protein Surface Hydrophobicity (PSH) DeterminationThe ANS binding properties of HSA can be investigatedby measuring 119870
app119889
and 119865max of the HSA-ANS complexin the absence and presence of isoimperatorin 119870app
119889
is theapparent dissociation constant of the fluorescent ANS-HSAcomplex and 1119870
app119889
is the binding affinity of ANS tothe protein in the absence and presence of isoimperatorin[28] 119865max is the maximum fluorescence intensity at thesaturated ANS concentration which indicates the numberof surface hydrophobic sites of the protein This methodinvolves titration of the enzyme solutions in the presenceof increasing concentrations of ANS The values of 1119870app
119889
and 119865max can be obtained from the slope and the 119909-interceptof the Scatchard plot respectively In this type of Scatchardplot the fluorescence intensity[ANS]free is drawn versusfluorescence intensity The free ANS concentration must bedetermined via plotting fluorescence intensity versus totalANS concentration In dilute solutions of ANS a linearrelationship exists between fluorescence intensity (119865) andANS concentration (119888) (119865 = 119861119888 where 119861 is the propor-tionality coefficient between fluorescence intensity and ANSconcentration) From a 1mM stock solution ANS was addedto the final concentrations ranging from 0 to 50 120583M Theenzyme concentration was 1 120583M It is assumed that in verydilute ANS solutions (0-1 120583M) all of the ANS molecules arebound to theHSA and thus B (the proportionality coefficientbetween fluorescence intensity and ANS concentration) canbe determined from the slope of the linear portion offluorescence intensity plot versus ANS concentration Foreach of the different total concentrations of ANS used theconcentration of bound ANS molecules was calculated usingthe following equation [ANS]bound = 119865119861 and the free ANSmolecules concentration was calculated from the differencebetween total and bound ANS concentrations ([ANS]free =[ANS]total minus [ANS]bound) [29 30]
The protein surface hydrophobicity index in the absenceand presence of the drug can be calculated by applying thefollowing equation [28]
PSH =
119865max
[HSA] 119870app119889
(9)
Following excitation at 380 nm the fluorescence emissionincrement was recorded at 470 nm until no further increasein fluorescence intensity was observed
27 Competitive Experiments In order to elucidate the bind-ing site of isoimperatorin onHSA we carried out competitive
experiments at the presence of two known drugs (Warfarinas site I marker and Ibuprofen as a drug that preferentiallybinds to site II) [24 31] The concentration of HSA and war-farinibuprofen was stabilized at 10times10minus6M Isoimperatorinwas then gradually added to theHSAHSA-warfarin orHSA-ibuprofen mixtures An excitation wavelength of 295 nm wasselected and flourescence spectra were recorded in the rangeof 300ndash450 nm
Also possible inhibitory effect of isoimperatorin indo-methacin tetracycline ibuprofen aspirin osthole and war-farin on the esterase activity of HSA was evaluated
28 Docking Procedure The AutoDock 405 docking pro-gram was applied to study the interaction between theisoimperatorin and HSA The crystal structure of HSA wasobtained from the Protein Data Bank Of more than 50X-ray crystallographic structures related to human serumalbumin in ProteinData Bank [32] entry with PDB ID1BM0[33] was chosen for dockings because of no missing atomsno cocrystallized ligand and having a reasonably goodresolution (25 A) All the water molecules were removedusing a plain text editor The polar hydrogen partial chargeand solvation parameters for use with the AutoDock 405simulation were applied using AutoDock-Tools The dockingsimulations were carried out with the rigid HSA and aflexible ligand [34] Using AutoGrid tools the grid mapswere generated adequately large to include the binding site(s)of protein as well as significant regions of the surroundingsurfaceThe points of the grids were thus 126times126times126witha grid spacing of 0375 A [35] Afterwards the most suitablestructure for the flexible ligand molecule was optimized bythe rotation of all single bonds in the ligand molecule Thegrid parameter file and the docking parameter file were setup by the AutoDockTools program [36] The binding modeof isoimperatorin was predicted by using the Lamarckiangenetic algorithm search engine implemented in AutoDocksoftware package [37] for the two main drug binding sitesI and II (subdomains IIA and IIIA of HSA resp) [38]Population size was 256 and Cluster analysis was performedon the docked results using a root mean square (RMS)tolerance of 05 A Default settings were used for all otherparameters [35] At the end of the docking simulation theHSA-IIM complex with the most stable energy was adoptedas the most favorable structure of the complex for HSAMolecular graphics were prepared with PyMOL version 099beta06 (httpwwwpymolorg)
3 Results and Discussion
31 UV Absorption Spectra UV absorption spectroscopytechnique can be used to explore the structural changes ofprotein and to investigate protein-ligand complex formationHSA has two main absorption bands that one of them islocated in the range of 260ndash300 nm which is the absorptionband of the aromatic amino acids (Trp Tyr and Phe) [27 39]Figure 2 shows the UV absorption spectra of HSA in theabsence and presence of isoimperatorin As can be seen inFigure 2 HSA has strong absorbance with a peak at 280 nmand the absorbance of HSA increased with the addition of
The Scientific World Journal 5
0
05
1
15
2
25
3
35
4
200 250 300 350Wavelength (nm)
Abso
rban
ce
034
035
036
037
038
039
04
270 274 278 282 286 290
1
7
Figure 2 Absorption spectra of HSA (30 120583M) with variousamounts of isoimperatorin (1ndash7) 0 04 08 2 4 6 and 8 120583MThe absorption of each sample was measured with respect to therespective blank
0
50
100
150
200
250
300
350
400
450
300 350 400 450 500 550Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
1
16
116
Figure 3 Effect of isoimperatorin on fluorescence spectra of HSA(119879 = 298K pH 75) Curve (1) 30 120583MHSA Curves (2ndash16) 30 120583MHSA in the presence of 02 04 06 08 1 15 2 25 3 4 5 6 7 8and 10120583M of isoimperatorin
isoimperatorin In addition the absorbance value of 280 nmof the isoimperatorin at a concentration of 10120583M is very low(lt003) Our results confirmed that the interaction betweenHSA and ligand took place
32 Fluorescence Measurements
321 Intrinsic Fluorescence Measurements The fluorescencespectra of HSA in the absence and presence of differentamounts of Isoimperatorin were recorded in the rangeof 300ndash570 nm upon excitation at 295 nm isoimperatorincauses a concentration dependent quenching of the intrinsicfluorescence of HSA (Figure 3) without changing the emis-sion maximum and shape of the peaks These results indi-cated that there were interactions between isoimperatorinand HSA This interaction was further confirmed by UV-visabsorption technique
0
1
2
3
4
0 2 4 6 8 10 12[Isoimperatorin] (120583M)
F0F
Figure 4 The Stern-Volmer plots for HSA fluorescence quenchingby isoimperatorin at 298 (⧫) 303 (998771) 308 (◼) and 313 (∙) K Thefinal protein concentration in 50mM phosphate pH 75 was 3 120583MBased on ldquoOne-Sample Kolmogorov-Smirnov Testrdquo test distributionwas normal
A gradual decrease in the fluorescence emission intensityof HSA is observed in the presence of increasing concentra-tions of isoimperatorin suggesting the presence of tryptophanresidue of the HSA at or near the isoimperatorin binding siteHSA has one Trp (Trp 214) and its fluorescence is solely dueto the excitation of this residue of HSA located in the bindingdomain-IIA (site I) at the bottom of a 12 A deep crevice [40]
Emerging of a new emission peak at 505 nm uponisoimperatorin addition can be attributed to the bounddrug and radiationless energy transfer between the HSAtryptophan and isoimperatorin [41] The appearance of anisoactinic point at 430 nm might also indicate the existenceof equilibrium between bound and free drug In additionsuch equilibrium may emphasize the formation of the drug-protein complex [42]
322 Fluorescence Quenching Mechanism Quenching canoccur by different mechanisms which is usually classifiedas dynamic quenching and static quenching In dynamicquenching increasing the temperature results in faster dif-fusion and hence larger amounts of collision which causesraising of the quenching constant Opposing to dynamicquenching in static one increasing the temperature weakensthe stability of the formed complex and hence reduces thequenching constant [19]
Fluorescence quenching of HSA upon interaction withisoimperatorin was evaluated at different temperatures (298303 308 and 313 K) and the results are shown in Figure 4As indicated in this figure the Stern-Volmer plots are lin-ear with the slopes increasing with increasing temperature(the linearity in this plot confirmed one-to-one interactionbetween isoimperatorin and HSA) The calculated values of119870SV at different temperatures are listed in Table 1 It can
6 The Scientific World Journal
Table 1 Stern-Volmer quenching constant (119870SV) quenching rate constant (119896119902) association constant (119870119887
) and number of binding sites (119899)for binding of isoimperatorin to HSA in the different temperatures
119879 (K) 10
minus4
times 119870SV(Mminus1)
10
minus12
times 119870
119902
(Mminus1sdotsminus1)119877
a 10
minus5
times 119870
119887
(Mminus1)Number of biding
sites (119899) 119877
a
2981698 plusmn 013 1698 plusmn 013
099621102 plusmn 010 0862 09841
3032019 plusmn 017 2019 plusmn 017
099551149 plusmn 013 0851 09886
3082333 plusmn 016 2333 plusmn 016
099651271 plusmn 022 0844 09950
3132639 plusmn 019 2639 plusmn 019
099811322 plusmn 017 0822 09917
Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870SV values
be found from Table 1 that the value of 119870SV increased withincreasing temperature in the presence of isoimperatorinwhich indicates the involved binding forces are mainlyhydrophobic interactions (endothermic apolar interactionsare strengthened with increasing temperature) Also the rateconstant of the quenching process (119896
119902) which is equal to119870SV
divided by 1205910(with assuming 120591
0= 10
minus8 s)
119896
119902=
119870SV120591
0
(10)
was calculated (Table 1) The results revealed that the valuesof 119896119902were much greater than the maximum diffusion rate
constant of the biomolecule (2 times 1010Mminus1 sminus1) which indi-cated that the fluorescence quenching of HSA was initiatedby complex formation between HSA and isoimperatorinconfirming that the mechanism of quenching is static [19]
323 Determination of Association Constant (119870119887) and Num-
ber of Binding Sites (119899) The values of association constant119870
119887 and the number of binding sites 119899 at different temper-
atures are obtained from the modified Stern-Volmer plots(Figure 5) and listed in Table 1 It can be found from thesedata that the 119899 values at different temperatures are keptaround unity which shows the existence of one binding sitein HSA for isoimperatorin The value of 119870
119887is significant to
understand the distribution of the drug in plasma since theweak binding can lead to a short life time or poor distributionwhile strong binding can decrease the concentration of freedrug in plasma The value of 119870
119887illustrated that there is a
strong binding force between isoimperatorin and HSA Itwas found that the binding constant increased with increasein temperature resulting in an increment of the stabilityof the isoimperatorin-HSA complex it also implies thatisoimperatorin can be tightly stored and carried by HSA inthe body
324 Forces Involved in the Binding Process Consideringthe dependence of binding constant on temperature a ther-modynamic process was considered to be responsible forthe formation of a complex Therefore the thermodynamicparameters dependent on temperatures were analyzed Theacting forces between a small molecule and macromoleculemainly include hydrogen bonds van der Waals forces elec-trostatic forces and hydrophobic interaction forces [23] Inorder to obtain the information about the forces persisting in
minus15
minus13
minus11
minus09
minus07
minus05
minus03
minus01
01
03
05
minus68 minus65 minus62 minus59 minus56 minus53 minus5
log[isoimperatorin]
log[(F0minusF)F]
Figure 5 The modified Stern-Volmer plots for HSA fluorescencequenching by isoimperatorin at 298 (⧫) 303 (998771) 308 (◼) and 313 (∙)K For interpretation of references to color in this figure legend thereader is referred to the web version of this article
the present binding process the thermodynamic parametersof binding were obtained fromVanrsquot Hoff plot followed by theGibbs equation (data not shown) Table 2 shows the values ofΔ119867
∘ andΔ119878∘ obtained for the binding site from the slopes andordinates at the origin of the fitted lines From Table 2 it canbe seen that the negative sign for free energy (Δ119866∘) meansthat the interaction process is spontaneous The positiveenthalpy (Δ119867∘) and entropy (Δ119878∘) values of the interaction ofisoimperatorin and HSA indicate that the binding is mainlyentropy-driven and the enthalpy is unfavorable for it thehydrophobic forces play a major role in the reaction [43]
33 Nonradiative Energy Transfer from HSA to Isoimpera-torin The Forster nonradiative energy transfer (FRET) hasbeen used as a ldquospectroscopic rulerrdquo for measurement ofthe distance between the acceptor (ligand) and the donor(tryptophan residues in the protein) [44] According to thistheory there will be FRET between two molecules if thefollowing conditions were satisfied (a) the donor moleculeemits fluorescence (b) the emission spectrum of the donorhas an enough overlap with the absorption spectrum of
The Scientific World Journal 7
Table 2 Thermodynamic parameters for binding isoimperatorin to HSA
119879 (K) Δ119878
∘ (Jsdotmolminus1sdotKminus1) Δ119867
∘ (kJsdotmolminus1) Δ119866
∘ (kJsdotmolminus1)298 minus2969 plusmn 009
303 17820 plusmn 011 2993 plusmn 013 minus3071 plusmn 007
308 minus3169 plusmn 010
313 minus3299 plusmn 008
0
50
100
150
200
250
300
350
400
450
222 272 322 372 422Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
0
002
004
006
008
01
012
014
Abso
rban
ce
a
b
Figure 6 The overlap of the fluorescence spectrum of HSA (a)and the absorption spectrum of isoimperatorin (b) when [HSA] =[isoimperatorin] = 10 120583MThe enzyme was excited at 295 nm
the acceptor and (c) the donor and acceptor molecules areclose enough with the maximum distance not to exceeding7 nm [45]
There is a good overlapping between the fluorescenceemission spectrum of HSA and absorption spectra of isoim-peratorin (Figure 6) As the fluorescence emission of proteinwas affected by the excitation light around 295 nm thespectrum ranging from 300ndash400 nm was chosen to calculatethe overlapping integral The value of 119869 was calculated to be473 plusmn 005 times 10
minus15 cm3 Lmolminus1 by integrating the overlapspectra for 300ndash400 nm In the present case 1198702 = 23119873 = 136 and Φ = 0118 [21] according to (6) and (7) wecould calculate that 119864 = 045 plusmn 006 119877
0= 322 plusmn 03 nm and
119903 = 29 plusmn 02 nm As mentioned earlier (Section 321) HSAposses one tryptophan residue (Trp 214) and the calculated119903 is actually the average value between the bound drug andTrp 214 Since 119903 is much smaller than 7 nm and 05119877
0lt
119903 lt 15119877
0[41] therefore the energy transfer from HSA
to isoimperatorin occurs with high probability which alsoexplains the efficient quenching of the HSA fluorescence
34 PSH Determination in the Absence and Presence of Isoim-peratorin Several methods exist for measuring the surfacehydrophobicity of proteins that ANS experiment is one ofthem ANS as an extrinsic fluorescent probe is extremelysensitive to polarity of the solvent In aqueous solutions itfluoresces very weakly but upon binding to hydrophobicpatches of proteins its quantum yield increases significantly[29] Thus ANS fluorescence may be applied for monitoringpossible changes in protein surface hydrophobicity induced
0
200
400
600
800
1000
0 10 20 30 40 50
Fluo
resc
ence
inte
nsity
(au
)
[ANS] (120583M)
Figure 7 Binding of ANS to HSA (1120583M) in the absence (∙) andpresence (998771) of 3120583M of isoimperatorin
upon drug binding At a fixed concentration (1 120583M) of HSAand increasing concentrations of ANS (0ndash50120583M) fluores-cence intensity of ANS was measured in the absence andpresence of 3120583M isoimperatorin (Figure 7)
A typical hyperbolic response is observed which demon-strates the saturation nature of ANS binding toHSA It can beseen that the hyperbolic response of ANS fluorescence in thepresence of isoimperatorin is slightly different in comparisonto the absence of the drug The values of 119865max and 119870
app119889
forANS are obtained from the Scatchard plot (Figure 8) andlisted in Table 3 The value of 119870app
119889
for ANS binding in theabsence of drug (09 plusmn 005) was increased in the presenceof 3 120583M isoimperatorin (14 plusmn 007) which shows the weakerbinding of ANS to the HSA-isoimperatorin complex ThenPSH was calculated in the absence and the presence of3 120583M isoimperatorin by using 119865max and119870
app119889
values (Table 3)Decreasing the PSH index of HSA (from 150794 plusmn 1146 to80612 plusmn 007) suggests that the surface hydrophobicity ofHSA is decreased by sim465 upon drug binding
35 Synchronous Fluorescence Spectroscopy Synchronous flu-orescence spectroscopy introduced by Lody has been usedto characterize complex mixtures which can provide finger-prints of complex samples [46] The synchronous fluores-cence spectra can provide information about the molecularenvironment in the vicinity of the chromophore moleculesIn the synchronous fluorescence spectra the sensitivityassociated with fluorescence is maintained while several
8 The Scientific World Journal
Table 3 Surface hydrophobicity parameters for HSA in the absence and the presence of isoimperatorin
Parameters HSA HSA-IIM complex119870
app119889
(120583M) 09 plusmn 005
lowast
14 plusmn 007
lowast
119865max[HSA] 135714 plusmn 85
lowastlowast
112853 plusmn 63
lowastlowast
Surface hydrophobicity index (119865max[HSA] 119870app119889
) 150794 plusmn 1146
lowastlowast
80612 plusmn 007
lowastlowast
Data are expressed as mean plusmn SD of three measurements lowast119875 value lt 00006 lowastlowast119875 value lt 00001
0
50
100
150
200
250
300
350
400
400 500 600 700 800 900 1000Fluorescence intensity (au)
Fmax
Fluo
resc
ence
inte
nsity
(au
)[A
NS]
free
Slope = minus1Kappd
Figure 8 Scatchard plots for titration of HSA with increasingconcentrations of ANS in the absence (∙) and presence (998771) of 3 120583Mof isoimperatorin 119865max and 1119870
app119889
values were obtained from the119909-intercept and slope of this plot For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
advantages are available spectral simplification spectralbandwidth reduction and avoidance of different perturbingeffects The fluorescence spectrum of BSAHSA mainly dueto the Trp residues is sensitive to the microenvironment ofthese chromophoreswith themaximumemissionwavelengthbeing very useful in estimating the hydrophobicity aroundthe tryptophan residues It is well known that the experimenton synchronous fluorescence of HSA will provide the char-acteristic information for the Tyr and Trp residues when thescanning interval Δ120582 is fixed at 15 and 60 nm respectively[47] The shift in the position of fluorescence emissionmaximum corresponds to changes of the polarity aroundthe chromophore molecule A blue shift of 120582max means thatthe amino acid residues are located in a more hydrophobicenvironment and are less exposed to the solvent while a redshift of 120582max implies that the amino acid residues are in apolar environment and are more exposed to the solvent [48]The synchronous fluorescence spectra of interaction betweenisoimperatorin and HSA are presented in Figure 8 It wasshown that the fluorescence intensity of HSA decreased regu-larly along with the addition of isoimperatorin which furtherdemonstrated the occurrence of fluorescence quenching inthe binding process Moreover there is no significant shiftof the maximum emission wavelength with Δ120582 = 15 nm or60 nm (Figures 9(a) and 9(b)) which implies that interaction
of isoimperatorin with HSA does not affect the conformationof the region around the Tyr or Trp residues
As well we can obtain the information about the locationof isoimperatorin binding site from the synchronous fluores-cence dataThe ratios of synchronous fluorescence quenching(RSFQ) were used to determine the binding sites of the drugsto the HSA molecule in drug-HSA complexes The RSFQvalues were calculated from the following equation
RSFQ = 1 minus
119865
119865
0
(11)
where 119865 and 1198650are the synchronous fluorescence intensities
of HSA in the presence and the absence of drug respectivelyIt was obvious from Figure 9(c) that the RSFQ results forΔ120582 = 60 nmwere slightly higher than the corresponding onesfor Δ120582 = 15 nm As illustrated in Figure 9(c) addition ofisoimperatorin resulted in strong fluorescence quenching ofTrp residue (sim47 reduction in fluorescence intensity) con-firming that isoimperatorin reached subdomain IIA whereonly one Trp residue (Trp 214) in HSA is located It wasalso observed from Figure 9(c) that the fluorescence strengthof Tyr residues decreased about sim32 in the presence ofisoimperatorin So the binding of isoimperatorin to HSAquenched flourescence intensity of Tyr and Trp Since Trp214 (the only Trp of HSA) is at site I and Tyr 263 (hasthe strong fluorescence in the case of Tyr residues andmay be one of the residues responsible for the synchronousfluorescence spectrum of HSA when Δ120582 is fixed at 15 nm) isin subdomain IIA (site I) it can be concluded that Trp andTyr residues of HSA may have equal accessibility to bindingsite of isoimperatorin [49] The above results indicated thatisoimperatorin can bind to HSA in the hydrophobic cavity(site I) on subdomain IIA which are in full agreement withquenching competitive binding experiments and theoreticaldocking technique
36 Site Marker Competitive Binding Experiments In orderto further classify drug binding site on HSA competitivebinding experiments have been carried out Warfarin andibuprofen are two markers specific for binding to HSA thatbind to sites I and II which are corresponded to the sub-domains IIA and IIIA respectively Therefore to determinethe location of the isoimperatorin binding site on HSA thecompetitive displacement experimentswere carried out usingwarfarin as a characteristicmarker for site I (Figure 10(a)) andibuprofen for site II (Figure 10(b)) [24 31] In the site markercompetitive experiment isoimperatorin was gradually addedto the solution of HSA HSA-ibuprofen or HSA-warfarincomplex and then fluorescence intensity of system wasrecorded
The Scientific World Journal 9
0
100
200
300
400
500
600
265 270 275 280 285 290 295 300Wavelength (nm)
1
8
Fluo
resc
ence
inte
nsity
(au
)
(a)
Fluo
resc
ence
inte
nsity
(au
)
0
50
100
150
200
250
300
350
400
250 260 270 280 290 300Wavelength (nm)
1
8
(b)
0
10
20
30
40
50
0 05 1 15 2 25 3
RSFQ
()
Isoimperatorin (120583M)
0
02
04
06
08
1
12
005 1 15 2 25 3 35
Isoimperatorin (120583M)
F0
F
(c)
Figure 9 Synchronous fluorescence spectra of interaction between HSA and isoimperatorin (a) at Δ120582 = 15 nm and (b) at Δ120582 = 60 nmConcentration ofHSAwas 30 120583Mwhile concentrations of isoimperatorinwere 0 04 08 12 16 2 25 and 3 120583Mfrom 1 to 8 (c) Comparativeevaluation of isoimperatorin effect on the RSFQ of HSA [HSA] = 3 120583M (◼) Δ120582 = 15 nm and (∙) Δ120582 = 60 nm
As shown in Figure 10(a) with addition of warfarin in theHSA solution the flourescence intensity was slightly higherthan that of without warfarin (and including the red shift)Then after adding the isoimperatorin into the above systemthe flourescence intensity of HSA solution with warfarinheld in equimolar decreased gradually and the intensitywas much lower than that of without warfarin displayingthat the binding of the isoimperatorin to HSA was affectedafter adding warfarin On the contrary in the presence ofibuprofen the flourescence intensity of the HSA-ibuprofencomplex almost had no difference from that recordedwithoutibuprofen under the same conditions (Figure 10(b)) whichindicated that site II marker did not prevent the binding ofisoimperatorin in its usual binding location The next stepprotein intrinsic fluorescence was measured and the relativefluorescence intensities (119865119865
0) versus ligand concentration
([isoimperatorin]) plots are displayed in Figure 10(c) Asshown in Table 4 the binding constant was remarkablydecreased after addition of warfarin while the addition ofibuprofen results in only a small difference These resultsindicate that warfarin can displace the isoimperatorin but
ibuprofen has little effect on the binding of isoimperatorinto HSA In other words these results suggest that isoim-peratorin probably competes with warfarin for binding toHSA [24] Experimental observations were followed up withdocking studies where isoimperatorin was docked to HSAto determine the preferred binding site on the proteinAs described above the 3D crystal structure of HSA is amonomer consisting of three homologous domains whichassemble to form a heart-shaped molecule [4 6] Each ofthe structurally similar 120572-helix domains (IndashIII) has twosubdomains (A andB) with six120572-helices in subdomainA andfour 120572-helices in subdomain BThe crystal structure analysesindicate that the principal regions of ligand binding sitesin HSA are located in hydrophobic cavities in subdomainsIIA and IIIA which are consistent with Sudlowrsquos site I andSudlowrsquos site II respectively Several studies have shown thatHSA is able to bind many ligands in several binding sites[38] In this paper the molecular docking model of theHSAndashisoimperatorin complex was conducted on the basisof the crystal structure of HSA to find the binding positionof isoimperatorin The best docking energy result is shown
10 The Scientific World Journal
0
100
200
300
400
500
600
310 335 360 385 410Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA1
8
(a)
0
100
200
300
400
500
600
700
305 325 345 365 385 405Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA
1
8
(b)
0
05
1
15
0 1 2 3
HSAIbuprofen
Warfarin
Isoimperatorin (120583M)
F0
F
(c)
Figure 10 Effect of isoimperatorin on the quenching of HSA-warfarin complex (a) andHSA-ibuprofen complex (b) fluorescence From 1 to 8[HSA] = [warfarin or iboprufen] = 3 120583M [isoimperatorin] = 0 04 08 12 16 2 25 and 3 120583M All reproduced data shown are representativeof at least 3 independent experiments (c) Fluorescence quenching profiles of HSA (⧫) HSA-ibobrofen (◼) and HSA-warfarin (998771) in thepresence of various concentration of isoimperatorin
in Figure 11 It can be seen that isoimperatorin was situatedwithin subdomain IIA in Sudlowrsquos site I formed by six 120572-helices This figure shows the location of Trp 214 and also thebinding of isoimperatorin in HSA As this fluorescent residueis in subdomain IIA and due to the proximity of the bindingsite of isoimperatorin to the Trp 214 it can be concluded thatisoimperatorin has one reactive site onHSAandprobably thissite is at or near the site I The above docking results are ingood agreement with the displacement experiments results
and revealed that isoimperatorin binds to Sudlowrsquos site I ofHSA
37 Effect of Isoimperatorin on HSA Esterase Activity Pre-liminary studies showed that albumin in addition to itsligand binding capabilities has remarkable promiscuouscatalytic activities toward a broad range of organicmoleculesincluding esters amides phosphates and benzisoxazolesHuman serum albumin had significantly greater esterase-like
The Scientific World Journal 11
Table 4 Binding constants for isoimperatorin-HSA interaction (at 295K) in the absence and presence of the site marker
119870
119887
times 10
minus5
(Mminus1) 119877
a
HSA + isoimperatorin (in the absence ofibuprofen and warfarin) 1110 plusmn 04
lowast 09947
HSA + isoimperatorin + ibuprofen 1099 plusmn 06
lowast 09904HSA + isoimperatorin + warfarin 789 plusmn 03
lowast 09881Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870
119887values lowast119875 value lt 00002
Trp 214
Isoimperatorin
Figure 11 Crystal structure of HSA and the positions of singletryptophan residue (Trp 214) and also the binding site of isoim-peratorin in HSA Protein backbone is shown in the ldquocartoonrdquorepresentation and isoimperatorinand Trp 214 residue in the site Iare shown in the ldquostickrdquo representation The isoimperatorin and Trp214 structures are represented by the red and blue colorsThe crystalstructure of protein was obtained from Protein Data Bank (1BM0)For interpretation of references to color in this figure legend thereader is referred to the web version of this paper
activity than bovine serum albumin and defatted albuminwas more active than untreated albumin [50] InterestinglyHSA exhibits esterase-like activity and hydrolyzes drugshaving an ester group such as aspirin (which can convertaspirin ldquoacetylsalicylic acidrdquo to salicylic acid) Lys199 Lys402Lys519 and Lys545 form the pseudoesterase active site ofHSAthat is located in site I [50] ldquoEsterase-likerdquo activity (hydrolysisof p-nitrophenol esters such as p-NPA) by subdomain III Ahas been also reported and site directed mutagenesis studieshave shown that Arg 410 and Tyr 411 are essential in thisesterase activity However further investigation revealed thatacetylation is not limited to a distinct side chain so that theHSA incubation with high concentrations of pNPAc resultsin the acetylation of 59 Lys 10 Ser 8 Thr 4 Tyr and 1 Asp[51] Because of the above difference here we set out toevaluate the effect of isoimperatorin on the esterase activityof HSA based on our previous work [19] The kinetics ofthe pseudoenzymatic hydrolysis of pNPAc was obtained inthe absence and presence of isoimperatorin and site markers(Figure 12) Accordingly warfarin and indomethacin (as siteI marker) and isoimperatorin inhibited competitively theHSA-catalyzed hydrolysis of pNPAc by binding to site I Incontrast ibuprofen and tetracycline were unable to inhibitesterase activity of HSA The enzyme inhibition results sug-gested that isoimperatorin aspirin warfarin indomethacin
0
005
01
015
02
025
03
035
0 05 1 15 2 25 3Time (min)
HSA TetracyclineIbuprofenAspirin
IndomethacinOstholeWarfarinIsoimperatorin
A400
Figure 12 4-Nitrophenyl acetate esterase activity of HSA in theabsence and presence of tetracycline and ibuprofen (site II) war-farin aspirin indomethacin and osthole (site I) and isoimperatorin(site I) Final concentration of protein pNPA and drugs were 15250 and 250 120583M respectively For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
and osthole binding sites on the HSA molecule are the sameThese results also proposed that the large site I on HSAparticipates effectively to its promiscuous esterase activity
4 Conclusion
The accumulated data on HSA-drug interactions can beused to develop quantitative structure-activity relationshipsfor albumin binding However as stated earlier adsorptiondistribution metabolism and elimination (ADME) is con-trolled by albumin [30] Poor or strong binding to SA isbelieved to affect half-time of drugs [35] and their dosagedepends on their half-life in plasma Thus the bindingaffinity of any drug to SA is one of the major factors thatdetermine the pharmacokinetics halftime and availabilityof the drug in various tissues (see [30] and referencestherein) Further complexities arise in vivo due to interactions
12 The Scientific World Journal
between drugs and endogenous ligands for HSA This isparticularly pertinent for fatty acids which normally occurin serum at levels between 01 and 2mol per mol of HSAand can both compete and cooperate with drugs bindingto the protein In certain disease states these effects areexacerbated as the fatty acid HSA molar ratio may be ashigh as six Other pathological conditions are associatedwith high (micromolar to millimolar) levels of bilirubinhemin or renal toxins (eg 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) indoxyl sulphate) which bindto the protein causing significant drug binding defects
Our major aim in this study was to distinguish thenature of the interaction of isoimperatorin to HSA usingdifferent biophysical techniques This study could be veryvaluable due to importance and presence of isoimperatorinin human diet (food-drug interaction on HSA) The resultsshowed that isoimperatorin quenched the intrinsic fluo-rescence of HSA through dynamic quenching mechanismFurthermore hydrophobic interaction played a major role inthe binding process Using Fluorescence and UV absorptionspectroscopy it has been shown that isoimperatorin inducesprotein structural changes The binding average distancebetween the solo tryptophan of the protein and the boundisoimperatorin was calculated by Forster theory as 290 plusmn02 nmAnalysis of PSHvalues elucidated that isoimperatorinhas induced some compactness in the tertiary structure ofthe enzyme The results of synchronous flourescence onHSA-isoimperatorin system elucidated that the drug binds tothe hydrophobic cavity (site I) on subdomain IIA that wasconfirmed by quenching experiments competitive bindingexperiments and docking technique
Acknowledgment
The authors gratefully acknowledge the Kermanshah Uni-versity of Medical Sciences Research Council for financialsupport of this work
References
[1] Y-J Hu Y Liu T-Q Sun A-M Bai J-Q Lu and Z-B Pi ldquoBinding of anti-inflammatory drug cromolyn sodiumto bovine serum albuminrdquo International Journal of BiologicalMacromolecules vol 39 no 4-5 pp 280ndash285 2006
[2] N Seedher and S Bhatia ldquoReversible binding of celecoxiband valdecoxib with human serum albumin using fluorescencespectroscopic techniquerdquo Pharmacological Research vol 54 no2 pp 77ndash84 2006
[3] H Isogai and N Hirayama ldquoIn silico prediction of interactionsbetween site II on human serum albumin and profen drugsrdquoISRN Pharma vol 2013 Article ID 818364 8 pages 2013
[4] YWangHYuX Shi Z LuoD Lin andMHuang ldquoStructuralmechanism of ring opening reaction of glucose by humanserum albuminrdquo The Journal of Biological Chemistry vol 288pp 15980ndash15987 2013
[5] F Keshavarz M M Alavianmehr and R Yousefi ldquoMolecularinteraction of benzalkonium Ibuprofenate and its discreteingredients with human serum albuminrdquo Physical ChemistryResearch vol 1 no 2 pp 111ndash116 2013
[6] T K Maiti K S Ghosh J Debnath and S Dasgupta ldquoBindingof all-trans retinoic acid to human serum albumin fluores-cence FT-IR and circular dichroism studiesrdquo InternationalJournal of Biological Macromolecules vol 38 no 3ndash5 pp 197ndash202 2006
[7] M-X XieM Long Y Liu C Qin and Y-DWang ldquoCharacter-ization of the interaction between human serum albumin andmorinrdquo Biochimica et Biophysica Acta vol 1760 no 8 pp 1184ndash1191 2006
[8] CD Kanakis P A TarantilisMG Polissiou S Diamantoglouand H A Tajmir-Riahi ldquoAntioxidant flavonoids bind humanserum albuminrdquo Journal of Molecular Structure vol 798 no 1-3 pp 69ndash74 2006
[9] S Marumoto and M Miyazawa ldquoStructure-activity rela-tionships for naturally occurring coumarins as 120573-secretaseinhibitorrdquo Bioorganic and Medicinal Chemistry vol 20 no 2pp 784ndash788 2012
[10] S Marumoto and M Miyazawa ldquoBiotransformation of isoim-peratorin and imperatorin by Glomerella cingulata and 120573-secretase inhibitory activityrdquo Bioorganic and Medicinal Chem-istry vol 18 no 1 pp 455ndash459 2010
[11] S E Sajjadi Y Shokoohinia and S Hemmati ldquoIsolation andidentification of furanocoumarins and a phenylpropanoid fromthe acetone extract and identification of volatile constituentsfrom the essential oil of Peucedanum pastinacifoliumrdquo Chem-istry of Natural Compounds vol 48 no 4 pp 668ndash671 2012
[12] X B Yang J Hou D Liu et al ldquoBiotransformation of isoim-peratorin by Cunninghamella blakesleana AS 3970rdquo Journal ofMolecular Catalysis B vol 88 pp 1ndash6 2013
[13] W Lili S Yehong S Qi et al ldquoIn vitro permeability analy-sis pharmacokinetic and brain distribution study in mice ofimperatorin isoimperatorin and cnidilin in Radix AngelicaeDahuricaerdquo Fitoterapia vol 85 pp 144ndash153 2013
[14] S Wang Q Chen and L He ldquoDevelopment and validationof a gas chromatography-mass spectrometry method for thedetermination of isoimperatorin in rat plasma and tissueapplication to the pharmacokinetic and tissue distributionstudyrdquo Journal of Chromatography B vol 852 no 1-2 pp 473ndash478 2007
[15] H Sadraei Y Shokoohinia S E Sajjadi and M MozafarildquoAntispasmodic effects of Prangos ferulacea acetone extract andits maincomponent osthole on ileum contractionrdquo Research inPharmaceutical Sciences vol 8 pp 137ndash144 2013
[16] H Sadraei Y Shokoohinia S E Sajjadi and B GhadirianldquoAntispasmodic effect of osthole and Prangos ferulacea extracton rat uterus smooth muscle motilityrdquo Research in Pharmaceu-tical Sciences vol 7 pp 141ndash149 2012
[17] S Sajjadi Y Shokoohinia S Gholamzadeh M Behbahani andA Fattahi ldquoAntiviral evaluation of coumarins from Prangosferulacea L (Lindl)rdquo Research in Pharmaceutical Sciences vol7 p 783 2012
[18] S E Sajjadi Y Shokoohinia and S Gholamzadeh ldquoChemicalcomposition of the essential oil of the root of Prangos ferulacea(L) Lindlrdquo Chemija vol 22 pp 178ndash180 2011
[19] N Bijari Y Shokoohinia M-R Ashrafi-Kooshk et al ldquoSpec-troscopic study of interaction between osthole and humanserum albumin identification of possible binding site of thecompoundrdquo Journal of Luminescence vol 143 pp 328ndash336 2013
[20] X Shi M Liu M Zhang et al ldquoIdentification of in vitro and invivo metabolites of isoimperatorin using liquid chromatogra-phymass spectrometryrdquo Food Chemistry vol 141 pp 357ndash3652013
The Scientific World Journal 13
[21] T Chatterjee A Pal S Dey B-K Chatterjee and PChakrabarti ldquoInteraction of virstatin with human serum albu-min spectroscopic analysis andmolecular modelingrdquo Plos Onevol 7 pp 1ndash12 2012
[22] Y-M Chen and L-H Guo ldquoCombined fluorescence and elec-trochemical investigation on the binding interaction betweenorganic acid and human serum albuminrdquo Journal of Environ-mental Sciences vol 21 no 3 pp 373ndash379 2009
[23] J R Lakowicz Principles of Fluorescence Spectroscopy KluwerAcademicPlenum Publishers New York NY USA 2nd edi-tion 1999
[24] F Ding N Li B Han F Liu L Zhang and Y Sun ldquoThe bindingof CI Acid Red 2 to human serum albumin determinationof binding mechanism and binding site using fluorescencespectroscopyrdquo Dyes and Pigments vol 83 no 2 pp 249ndash2572009
[25] M-R Ashrafi Kooshk K Mansouri M Nadi S Khodarahmiand R Khodarahmi ldquoIn vitro anti-cancer activity of nativecurcumin and protein-curcuminrdquo Systems vol 2 no 1 pp 80ndash88 1981
[26] P D Ross and S Subramanian ldquoThermodynamics of proteinassociation reactions forces contributing to stabilityrdquo Biochem-istry vol 20 no 11 pp 3096ndash3102 1981
[27] J Liu J-N Tian J Zhang Z Hu and X Chen ldquoInteractionof magnolol with bovine serum albumin a fluorescence-quenching studyrdquo Analytical and Bioanalytical Chemistry vol376 no 6 pp 864ndash867 2003
[28] A Coi A M Bianucci F Bonomi P Rasmussen G M Muraand M L Ganadu ldquoStructural perturbation of 120572B-crystallinby zinc and temperature related to its chaperone-like activityrdquoInternational Journal of Biological Macromolecules vol 42 no3 pp 229ndash234 2008
[29] M Moller and A Denicola ldquoStudy of protein-ligand bindingby fluorescencerdquoBiochemistry andMolecular Biology Educationvol 30 no 5 pp 309ndash312 2002
[30] R Khodarahmi S A Karimi M R Ashrafi Kooshk S AGhadami S Ghobadi and M Amani ldquoComparative spec-troscopic studies on drug binding characteristics and proteinsurface hydrophobicity of native and modified forms of bovineserum albumin possible relevance to change in protein struc-turefunction upon non-enzymatic glycationrdquo SpectrochimicaActa A vol 89 pp 177ndash186 2012
[31] S-F Ma M Anraku Y Iwao et al ldquoHydrolysis of angiotensinII receptor blocker prodrug olmesartan medoxomil by humanserum albumin and identification of its catalytic active sitesrdquoDrug Metabolism and Disposition vol 33 no 12 pp 1911ndash19192005
[32] H M Berman J Westbrook Z Feng et al ldquoThe protein databankrdquo Nucleic Acids Research vol 28 no 1 pp 235ndash242 2000
[33] S Sugio A Kashima SMochizukiMNoda andKKobayashildquoCrystal structure of human serum albumin at 25 A resolutionrdquoProtein Engineering vol 12 no 6 pp 439ndash446 1999
[34] G M Morris D S Goodsell R Huey and A J OlsonldquoDistributed automated docking of flexible ligands to proteinsparallel applications of AutoDock 24rdquo Journal of Computer-Aided Molecular Design vol 10 no 4 pp 293ndash304 1996
[35] F Faridbod M R Ganjali B Larijani et al ldquoInteraction studyof pioglitazone with albumin by fluorescence spectroscopy andmolecular dockingrdquo Spectrochimica Acta A vol 78 no 1 pp96ndash101 2011
[36] D SGoodsell andA JOlson ldquoAutomated docking of substratesto proteins by simulated annealingrdquo Proteins vol 8 no 3 pp195ndash202 1990
[37] D S Goodsell G M Morris and A J Olson ldquoAutomateddocking of flexible ligands applications of AutoDockrdquo Journalof Molecular Recognition vol 9 no 1 pp 1ndash5 1996
[38] G Sudlow D J Birkett and D N Wade ldquoThe characterizationof two specific drug binding sites on human serum albuminrdquoMolecular Pharmacology vol 11 no 6 pp 824ndash832 1975
[39] Y Yue X Chen J Qin and X Yao ldquoCharacterization of themangiferin-human serum albumin complex by spectroscopicandmolecularmodeling approachesrdquo Journal of Pharmaceuticaland Biomedical Analysis vol 49 no 3 pp 753ndash759 2009
[40] A K Shaw and S K Pal ldquoSpectroscopic studies on the effectof temperature on pH-induced folded states of human serumalbuminrdquo Journal of Photochemistry and Photobiology B vol 90no 1 pp 69ndash77 2008
[41] S Ranjbar S Ghobadi R Khodarahmi and H Nemati ldquoSpec-troscopic characterization of furosemide binding to humancarbonic anhydrase IIrdquo International Journal of BiologicalMacromolecules vol 50 no 4 pp 910ndash917 2012
[42] HNHou Z POi YWOYung F L Liao Y Zhang andY LiyldquoStudies on interaction between vitamin B12 and human serumalbuminrdquo Journal of Pharmaceutical and Biomedical Analysisvol 47 pp 134ndash139 2008
[43] S S Kalanur J Seetharamappa and V K A Kalalbandi ldquoChar-acterization of interaction and the effect of carbamazepine onthe structure of human serum albuminrdquo Journal of Pharmaceu-tical and Biomedical Analysis vol 53 no 3 pp 660ndash666 2010
[44] L Stryer ldquoFluorescence spectroscopy of proteinsrdquo Science vol162 no 3853 pp 526ndash533 1968
[45] B Valeur and J C Brochon New Trends in FluorescenceSpectroscopy Springer Berlin Germany 1999
[46] V Suryawanshi P Anbhule A Gore Sh Patil and G AndKolekar ldquoA spectral deciphering the perturbation of modeltransporter protein (HSA) by antibacterial pyrimidine deriva-tive pharmacokinetic and biophysical insightsrdquo Journal ofPhotochemistry and Photobiology B vol 118 pp 1ndash8 2013
[47] T Yuan AMWeljie andH J Vogel ldquoTryptophan fluorescencequenching by methionine and selenomethionine residues ofcalmodulin orientation of peptide and protein bindingrdquo Bio-chemistry vol 37 no 9 pp 3187ndash3195 1998
[48] TCOrsquoHaver A F Fell G Smith et al ldquoDerivative spectroscopyand its applications in analysisrdquo Analytical Proceedings vol 19no 1 pp 22ndash46 1982
[49] Y-Q Wang B-P Tang H-M Zhang Q-H Zhou and G-C Zhang ldquoStudies on the interaction between imidaclopridand human serum albumin spectroscopic approachrdquo Journal ofPhotochemistry and Photobiology B vol 94 no 3 pp 183ndash1902009
[50] S-J Kim H-W Rhee H-Y Park H-S Kim and J-I HongldquoFluorescent probes designed for detecting human serum albu-min on the basis of its pseudo-esterase activityrdquo Bioorganic ampMedicinal Chemistry Letters vol 23 pp 2093ndash2097 2013
[51] P Ascenzi M Gioia G Fanali M Coletta and M FasanoldquoPseudo-enzymatic hydrolysis of 4-nitrophenyl acetate byhuman serum albumin pH-dependence of rates of individualstepsrdquo Biochemical and Biophysical Research Communicationsvol 424 pp 451ndash455 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 The Scientific World Journal
OO
O
O
Figure 1 Structure of isoimperatorin
binding affinity for human serum albumin (HSA) HSA isthe most abundant drug carrier protein which is also foundin tissues and bodily secretions throughout the body theextravascular protein comprises 60 of the total albuminHSA is a widely studied protein because its primary structureis well known and its tertiary structure has been determinedby X-ray crystallography [3ndash6] It has an important rolein maintaining the colloidal osmotic pressure in bloodit transports and distributes exogenous and endogenousmolecules and metabolites such as nutrients hormones fattyacids and many diverse drugs [6 7] This heart-shapedprotein which has a net charge of minus15 at neutral pH and anisoelectric point of about 5 consists of 585 amino acid residuesand is composed of three structurally similar domains (III and III) each containing two subdomains (A and B)and being stabilized by 17 disulphide bridges [4ndash8] Crystalstructure analyses have revealed that HSA has binding sitesfor aromatic and heterocyclic ligands within two hydropho-bic pockets in subdomains IIA (commonly referred to asSudlowrsquos site I warfarin-binding site) and IIIA (commonlyreferred to as Sudlowrsquos site II indolebenzodiazepine site)Both hydrophobic and electrostatic interactions play a majorrole in controlling the affinity towards drug binding forsites I and II for site I mainly hydrophobic interactions aredominant whereas for site II a combination of hydrophobichydrogen bonding and electrostatic interactions all play acrucial role The sole tryptophan residue (Trp 214) of HSAis in subdomain IIA (site I) [6ndash8]
Coumarins are widely distributed in nature and arefound in all parts of plants These compounds are especiallycommon in grasses orchids citrus fruits and legumes [9]Being so abundant in nature coumarins make up an impor-tant part of the human diet Based on chemical structurethey can be broadly classified as (a) simple coumarins(b) furanocoumarins of the linear or angular type and(c) pyranocoumarins of the linear or angular type [9ndash11]Isoimperatorin (4-[(3-methyl-2-butenyl) oxy]-7H-furo [32-g] [1] benzopyran-7-one) [12] (Figure 1) [13] is one of themaincomponents of Prangos ferulacea as a linear furanocoumarin[14] The plant has previously shown antispasmodic effects[15 16] and proved to contain coumarins and terpenoids [17ndash19] Isoimperatorin has attracted increasing interest due to itsanti-inflammatory analgesic antispasmodic and anticancer
activities and treatment of Alzheimerrsquos disease [9 10 20]With the increasing importance of isoimperatorin in humanhealth it is necessary to evaluate its effect on biomoleculesfor example proteins as a chemical entity [10 20] Interactionbetween serum albumin and many furocoumarins has beeninvestigated successfully but detailed investigations of theinteraction of HSA with isoimperatorin are not available Soin this study we investigated the effect of isoimperatorin onthe structure and action of HSA bymeans of various spectro-scopic techniquesTheUV-vis spectroscopy and intrinsic andextrinsic fluorometry methods were used for elucidating themode of binding and probable structural alterations of HSAupon drug bindingThe characteristics of the binding that isbinding constant number of binding sites and and nature ofbinding forces as well as the effects of the complexation onthe protein conformation were determined In addition thethermodynamic and Forsterrsquos parameters associated with thebinding process were also calculated
2 Materials and Methods
21 Chemicals The pure warfarin was obtained as a gen-erous gift from Dr S Kashanian (Razi University) 1-Anilinonaphthalene-8-sulfonate (ANS) was purchased fromSigma Chemical Co (St Louis Mo USA)The other reagentswere of analytical grade from Merck (Darmstadt Germany)All of the solutions were prepared in double-distilled waterand all of the experiments were carried out in 50mMphosphate pH 75 as the buffer All of themeasurements weredone in triplicate
22 Extraction Isolation and Identification of IsoimperatorinRoots of Prangos ferulacea L were collected from Yasujin Kohgiluyeh-Boyer-Ahmad province Iran in June 2011The plant was identified at the Botany Department of YasujUniversity Yasuj and compared to the voucher specimen(No 2408) at the Herbarium of School of Pharmacy IsfahanUniversity of Medical Sciences Isfahan Iran
Dried ground roots were extracted with acetone for twodays (5 L times 4) The extract was concentrated to bear aviscous mass which was then put in minus20∘C for two daysand chill filtered in which the filtrate resulted in a solidmass after drying The latter was fractioned by vacuumliquid chromatography (Silica gel MESH 230ndash400) using agradient of heptane ethylacetate (H EtOAc) from 100 0 to0 100 to afford several fractions Third fraction (H EtOAc7 3) rendered a mixture of two coumarins in which furtherpurification via chromatographies (H EtOAc 8 2 to 7 3)resulted in pure isoimperatorin
23 UV-vis Measurements The UV-vis spectra are recordedat room temperature on a Cary Eclips (Varian) spectropho-tometer equipped with 10 cm quartzecells The final drugconcentrations and constant protein concentration were 04ndash8 120583M and 3 120583M respectively The exact concentration ofHSA was determined spectrophotometrically using molarextinction coefficient of 35700Mminus1 cmminus1 [21]
The Scientific World Journal 3
24 Intrinsic Fluorescence Measurements Fluorescence mea-surements were performed on a Cary Eclipse (Varian) spec-trophotometer with jacketed cell holders in which tem-perature was adjusted by an external thermostated watercirculationThe excitation and emission wavelengths were setat 295 nm (to avoid the contribution from tyrosine residues[22]) and 300ndash570 nm respectively with the excitation andemission slit widths of 5 and 10 nm respectively Fluorescenceemission spectra of the HSA were measured in the absenceand presence of the various concentrations of isoimperatorin(02 04 06 08 1 15 2 25 3 35 4 5 6 8 and 10 120583M)Thefinal enzyme concentration was 3 120583M
Synchronous fluorescence spectra of HSA with variousconcentrations of isoimperatorin were obtained (Δ120582 = 15 nmandΔ120582 = 60 nm) with the excitation and emission slit widthsof 5 and 10 nm respectively
241 The Inner Filter Effect The inner filter effect can bea problem for any fluorescencemeasurement especiallywherean absorbing component is being titrated into the cuvetteDuring a fluorescence titration experiment at the wavelengthof excitation or at the wavelengths used to follow an emissionthe absorption of the molecule added resulted in a spuriousdecrease in the observed fluorescence intensity [23] In thesteady-state fluorescence experiments absorption spectrumof isoimperatorin overlapswith the excitation and emission ofHSAThus it is very important to subtract such an effect fromthe raw quenching data The fluorescence data was correctedfor inner filter effect using
119865corr =119865obs antilog (119860em + 119860ex)
2
(1)
where 119865corr and 119865obs are the corrected and observed flu-orescence intensity 119860ex and 119860em are the optical densityof the sample at the excitation and emission wavelengthsrespectively
242 Determination of Quenching Mechanism In order toclarify the mechanism of HSA fluorescence quenching byisoimperatorin the quenching experiments were carried outat 298 303 308 and 313 K where HSA does not undergoany considerable thermal denaturation The decrease influorescence intensity at 120582max was analyzed using the well-known Stern-Volmer equation [24]
119865
0
119865
= 1 + 119870SV [119876] (2)
where 1198650and 119865 are the fluorescence intensities in the absence
and presence of quencher (isoimperatorin) respectively the119870SV is the Stern-Volmer quenching constant and [119876] is theconcentration of the compound isoimperatorin
243 Determination of Association Constant and Number ofBinding Sites The association constant (119870
119887) and number of
binding sites (119899) for binding of small molecules to set of
equivalent sites on amacromolecule can be determined usingthe modified Stern-Volmer equation
log 119865
0minus 119865
119865
= log119870119887+ 119899 log [119876] (3)
where 1198650and 119865 are the fluorescence intensities of HSA in
the absence and presence of the quencher (isoimperatorin)respectively 119870
119887is the association constant and 119899 is the
number of binding sites per HSA [25]The values of 119899 and119870119887
were obtained from the slope and intercept of the modifiedStern-Volmer plot respectively
244 Thermodynamic Analysis of the Binding Process Inorder to explain the nature of the interaction betweenisoimperatorin and HSA the association constant of HSA-isoimperatorin complex was determined at four differenttemperatures (298 303 308 and 313 K)The thermodynamicparameters of binding entropy change (Δ119878∘) and enthalpychange (Δ119867∘) were obtained from vanrsquot Hoff equation
ln 119870119887=
minusΔ119867
∘
119877119879
+
Δ119878
∘
119877
(4)
where119870119887is the association constant at the given temperature
and119877 is the universal gas constantThe values ofΔ119878∘ andΔ119867∘can be obtained from the intercept and slope of the vanrsquot Hoffplot respectively and the free energy change (Δ119866∘) can beestimated by using the Gibbs equation [26]
Δ119866
∘
= Δ119867
∘
minus 119879Δ119878
∘
(5)
25 Energy Transfer between HSA and Isoimperatorin Thedistance between the acceptor (ligand) and the donor (tryp-tophan residues in the protein) 119903 according to Forsterrsquosnonradiation energy transfer theory could be calculated bythe following equation [21]
119864 = 1 minus (
119865
119865
0
) =
119877
6
0
119877
6
0
+ 119903
6
0
(6)
where 119864 is the efficiency of energy transfer between donorand acceptor 119865 and 119865
0are the fluorescence intensities of
donor in the presence and absence of acceptor respectivelyand 119877
0is a critical distance at which the efficiency of energy
transfer is 50 The value of 1198770was calculated using
119877
6
0
= 88 times 10
minus25
119870
2
119873
minus4
Φ119869(7)
where 1198702 is the spatial orientation factor describing thegeometry of the donor and acceptor dipoles 119873 is theaverage refractive index of the mediumΦ is the fluorescencequantum yield of the donor in the absence of acceptor and 119869expresses the degree of spectral overlap between the emissionspectrum of the donor and the absorption spectrum of theacceptor 119869 can be given by
119869 = sum119865 (120582) 120576 (120582) 120582
4
Δ120582 (8)
where 119865(120582) is the fluorescence intensity of the donor atwavelength 120582 and 120576(120582) is the molar absorption coefficient
4 The Scientific World Journal
of the acceptor at 120582 So 119869 can be evaluated by integratingthe overlap between emission spectrum of the donor andabsorption spectrum of the acceptor [27]
The absorption spectrum of isoimperatorin (10 120583M) wasrecorded in the range of 222ndash450 nmThe emission spectrumof HSA (10 120583M) was also recorded in the range of 222ndash450 nm with excitation and emission slit widths of 5 and5 nm respectivelyThe overlap of the absorption spectrum ofisoimperatorin with the fluorescence emission spectrum ofHSA was used to calculate the efficiency of energy transfer
26 Protein Surface Hydrophobicity (PSH) DeterminationThe ANS binding properties of HSA can be investigatedby measuring 119870
app119889
and 119865max of the HSA-ANS complexin the absence and presence of isoimperatorin 119870app
119889
is theapparent dissociation constant of the fluorescent ANS-HSAcomplex and 1119870
app119889
is the binding affinity of ANS tothe protein in the absence and presence of isoimperatorin[28] 119865max is the maximum fluorescence intensity at thesaturated ANS concentration which indicates the numberof surface hydrophobic sites of the protein This methodinvolves titration of the enzyme solutions in the presenceof increasing concentrations of ANS The values of 1119870app
119889
and 119865max can be obtained from the slope and the 119909-interceptof the Scatchard plot respectively In this type of Scatchardplot the fluorescence intensity[ANS]free is drawn versusfluorescence intensity The free ANS concentration must bedetermined via plotting fluorescence intensity versus totalANS concentration In dilute solutions of ANS a linearrelationship exists between fluorescence intensity (119865) andANS concentration (119888) (119865 = 119861119888 where 119861 is the propor-tionality coefficient between fluorescence intensity and ANSconcentration) From a 1mM stock solution ANS was addedto the final concentrations ranging from 0 to 50 120583M Theenzyme concentration was 1 120583M It is assumed that in verydilute ANS solutions (0-1 120583M) all of the ANS molecules arebound to theHSA and thus B (the proportionality coefficientbetween fluorescence intensity and ANS concentration) canbe determined from the slope of the linear portion offluorescence intensity plot versus ANS concentration Foreach of the different total concentrations of ANS used theconcentration of bound ANS molecules was calculated usingthe following equation [ANS]bound = 119865119861 and the free ANSmolecules concentration was calculated from the differencebetween total and bound ANS concentrations ([ANS]free =[ANS]total minus [ANS]bound) [29 30]
The protein surface hydrophobicity index in the absenceand presence of the drug can be calculated by applying thefollowing equation [28]
PSH =
119865max
[HSA] 119870app119889
(9)
Following excitation at 380 nm the fluorescence emissionincrement was recorded at 470 nm until no further increasein fluorescence intensity was observed
27 Competitive Experiments In order to elucidate the bind-ing site of isoimperatorin onHSA we carried out competitive
experiments at the presence of two known drugs (Warfarinas site I marker and Ibuprofen as a drug that preferentiallybinds to site II) [24 31] The concentration of HSA and war-farinibuprofen was stabilized at 10times10minus6M Isoimperatorinwas then gradually added to theHSAHSA-warfarin orHSA-ibuprofen mixtures An excitation wavelength of 295 nm wasselected and flourescence spectra were recorded in the rangeof 300ndash450 nm
Also possible inhibitory effect of isoimperatorin indo-methacin tetracycline ibuprofen aspirin osthole and war-farin on the esterase activity of HSA was evaluated
28 Docking Procedure The AutoDock 405 docking pro-gram was applied to study the interaction between theisoimperatorin and HSA The crystal structure of HSA wasobtained from the Protein Data Bank Of more than 50X-ray crystallographic structures related to human serumalbumin in ProteinData Bank [32] entry with PDB ID1BM0[33] was chosen for dockings because of no missing atomsno cocrystallized ligand and having a reasonably goodresolution (25 A) All the water molecules were removedusing a plain text editor The polar hydrogen partial chargeand solvation parameters for use with the AutoDock 405simulation were applied using AutoDock-Tools The dockingsimulations were carried out with the rigid HSA and aflexible ligand [34] Using AutoGrid tools the grid mapswere generated adequately large to include the binding site(s)of protein as well as significant regions of the surroundingsurfaceThe points of the grids were thus 126times126times126witha grid spacing of 0375 A [35] Afterwards the most suitablestructure for the flexible ligand molecule was optimized bythe rotation of all single bonds in the ligand molecule Thegrid parameter file and the docking parameter file were setup by the AutoDockTools program [36] The binding modeof isoimperatorin was predicted by using the Lamarckiangenetic algorithm search engine implemented in AutoDocksoftware package [37] for the two main drug binding sitesI and II (subdomains IIA and IIIA of HSA resp) [38]Population size was 256 and Cluster analysis was performedon the docked results using a root mean square (RMS)tolerance of 05 A Default settings were used for all otherparameters [35] At the end of the docking simulation theHSA-IIM complex with the most stable energy was adoptedas the most favorable structure of the complex for HSAMolecular graphics were prepared with PyMOL version 099beta06 (httpwwwpymolorg)
3 Results and Discussion
31 UV Absorption Spectra UV absorption spectroscopytechnique can be used to explore the structural changes ofprotein and to investigate protein-ligand complex formationHSA has two main absorption bands that one of them islocated in the range of 260ndash300 nm which is the absorptionband of the aromatic amino acids (Trp Tyr and Phe) [27 39]Figure 2 shows the UV absorption spectra of HSA in theabsence and presence of isoimperatorin As can be seen inFigure 2 HSA has strong absorbance with a peak at 280 nmand the absorbance of HSA increased with the addition of
The Scientific World Journal 5
0
05
1
15
2
25
3
35
4
200 250 300 350Wavelength (nm)
Abso
rban
ce
034
035
036
037
038
039
04
270 274 278 282 286 290
1
7
Figure 2 Absorption spectra of HSA (30 120583M) with variousamounts of isoimperatorin (1ndash7) 0 04 08 2 4 6 and 8 120583MThe absorption of each sample was measured with respect to therespective blank
0
50
100
150
200
250
300
350
400
450
300 350 400 450 500 550Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
1
16
116
Figure 3 Effect of isoimperatorin on fluorescence spectra of HSA(119879 = 298K pH 75) Curve (1) 30 120583MHSA Curves (2ndash16) 30 120583MHSA in the presence of 02 04 06 08 1 15 2 25 3 4 5 6 7 8and 10120583M of isoimperatorin
isoimperatorin In addition the absorbance value of 280 nmof the isoimperatorin at a concentration of 10120583M is very low(lt003) Our results confirmed that the interaction betweenHSA and ligand took place
32 Fluorescence Measurements
321 Intrinsic Fluorescence Measurements The fluorescencespectra of HSA in the absence and presence of differentamounts of Isoimperatorin were recorded in the rangeof 300ndash570 nm upon excitation at 295 nm isoimperatorincauses a concentration dependent quenching of the intrinsicfluorescence of HSA (Figure 3) without changing the emis-sion maximum and shape of the peaks These results indi-cated that there were interactions between isoimperatorinand HSA This interaction was further confirmed by UV-visabsorption technique
0
1
2
3
4
0 2 4 6 8 10 12[Isoimperatorin] (120583M)
F0F
Figure 4 The Stern-Volmer plots for HSA fluorescence quenchingby isoimperatorin at 298 (⧫) 303 (998771) 308 (◼) and 313 (∙) K Thefinal protein concentration in 50mM phosphate pH 75 was 3 120583MBased on ldquoOne-Sample Kolmogorov-Smirnov Testrdquo test distributionwas normal
A gradual decrease in the fluorescence emission intensityof HSA is observed in the presence of increasing concentra-tions of isoimperatorin suggesting the presence of tryptophanresidue of the HSA at or near the isoimperatorin binding siteHSA has one Trp (Trp 214) and its fluorescence is solely dueto the excitation of this residue of HSA located in the bindingdomain-IIA (site I) at the bottom of a 12 A deep crevice [40]
Emerging of a new emission peak at 505 nm uponisoimperatorin addition can be attributed to the bounddrug and radiationless energy transfer between the HSAtryptophan and isoimperatorin [41] The appearance of anisoactinic point at 430 nm might also indicate the existenceof equilibrium between bound and free drug In additionsuch equilibrium may emphasize the formation of the drug-protein complex [42]
322 Fluorescence Quenching Mechanism Quenching canoccur by different mechanisms which is usually classifiedas dynamic quenching and static quenching In dynamicquenching increasing the temperature results in faster dif-fusion and hence larger amounts of collision which causesraising of the quenching constant Opposing to dynamicquenching in static one increasing the temperature weakensthe stability of the formed complex and hence reduces thequenching constant [19]
Fluorescence quenching of HSA upon interaction withisoimperatorin was evaluated at different temperatures (298303 308 and 313 K) and the results are shown in Figure 4As indicated in this figure the Stern-Volmer plots are lin-ear with the slopes increasing with increasing temperature(the linearity in this plot confirmed one-to-one interactionbetween isoimperatorin and HSA) The calculated values of119870SV at different temperatures are listed in Table 1 It can
6 The Scientific World Journal
Table 1 Stern-Volmer quenching constant (119870SV) quenching rate constant (119896119902) association constant (119870119887
) and number of binding sites (119899)for binding of isoimperatorin to HSA in the different temperatures
119879 (K) 10
minus4
times 119870SV(Mminus1)
10
minus12
times 119870
119902
(Mminus1sdotsminus1)119877
a 10
minus5
times 119870
119887
(Mminus1)Number of biding
sites (119899) 119877
a
2981698 plusmn 013 1698 plusmn 013
099621102 plusmn 010 0862 09841
3032019 plusmn 017 2019 plusmn 017
099551149 plusmn 013 0851 09886
3082333 plusmn 016 2333 plusmn 016
099651271 plusmn 022 0844 09950
3132639 plusmn 019 2639 plusmn 019
099811322 plusmn 017 0822 09917
Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870SV values
be found from Table 1 that the value of 119870SV increased withincreasing temperature in the presence of isoimperatorinwhich indicates the involved binding forces are mainlyhydrophobic interactions (endothermic apolar interactionsare strengthened with increasing temperature) Also the rateconstant of the quenching process (119896
119902) which is equal to119870SV
divided by 1205910(with assuming 120591
0= 10
minus8 s)
119896
119902=
119870SV120591
0
(10)
was calculated (Table 1) The results revealed that the valuesof 119896119902were much greater than the maximum diffusion rate
constant of the biomolecule (2 times 1010Mminus1 sminus1) which indi-cated that the fluorescence quenching of HSA was initiatedby complex formation between HSA and isoimperatorinconfirming that the mechanism of quenching is static [19]
323 Determination of Association Constant (119870119887) and Num-
ber of Binding Sites (119899) The values of association constant119870
119887 and the number of binding sites 119899 at different temper-
atures are obtained from the modified Stern-Volmer plots(Figure 5) and listed in Table 1 It can be found from thesedata that the 119899 values at different temperatures are keptaround unity which shows the existence of one binding sitein HSA for isoimperatorin The value of 119870
119887is significant to
understand the distribution of the drug in plasma since theweak binding can lead to a short life time or poor distributionwhile strong binding can decrease the concentration of freedrug in plasma The value of 119870
119887illustrated that there is a
strong binding force between isoimperatorin and HSA Itwas found that the binding constant increased with increasein temperature resulting in an increment of the stabilityof the isoimperatorin-HSA complex it also implies thatisoimperatorin can be tightly stored and carried by HSA inthe body
324 Forces Involved in the Binding Process Consideringthe dependence of binding constant on temperature a ther-modynamic process was considered to be responsible forthe formation of a complex Therefore the thermodynamicparameters dependent on temperatures were analyzed Theacting forces between a small molecule and macromoleculemainly include hydrogen bonds van der Waals forces elec-trostatic forces and hydrophobic interaction forces [23] Inorder to obtain the information about the forces persisting in
minus15
minus13
minus11
minus09
minus07
minus05
minus03
minus01
01
03
05
minus68 minus65 minus62 minus59 minus56 minus53 minus5
log[isoimperatorin]
log[(F0minusF)F]
Figure 5 The modified Stern-Volmer plots for HSA fluorescencequenching by isoimperatorin at 298 (⧫) 303 (998771) 308 (◼) and 313 (∙)K For interpretation of references to color in this figure legend thereader is referred to the web version of this article
the present binding process the thermodynamic parametersof binding were obtained fromVanrsquot Hoff plot followed by theGibbs equation (data not shown) Table 2 shows the values ofΔ119867
∘ andΔ119878∘ obtained for the binding site from the slopes andordinates at the origin of the fitted lines From Table 2 it canbe seen that the negative sign for free energy (Δ119866∘) meansthat the interaction process is spontaneous The positiveenthalpy (Δ119867∘) and entropy (Δ119878∘) values of the interaction ofisoimperatorin and HSA indicate that the binding is mainlyentropy-driven and the enthalpy is unfavorable for it thehydrophobic forces play a major role in the reaction [43]
33 Nonradiative Energy Transfer from HSA to Isoimpera-torin The Forster nonradiative energy transfer (FRET) hasbeen used as a ldquospectroscopic rulerrdquo for measurement ofthe distance between the acceptor (ligand) and the donor(tryptophan residues in the protein) [44] According to thistheory there will be FRET between two molecules if thefollowing conditions were satisfied (a) the donor moleculeemits fluorescence (b) the emission spectrum of the donorhas an enough overlap with the absorption spectrum of
The Scientific World Journal 7
Table 2 Thermodynamic parameters for binding isoimperatorin to HSA
119879 (K) Δ119878
∘ (Jsdotmolminus1sdotKminus1) Δ119867
∘ (kJsdotmolminus1) Δ119866
∘ (kJsdotmolminus1)298 minus2969 plusmn 009
303 17820 plusmn 011 2993 plusmn 013 minus3071 plusmn 007
308 minus3169 plusmn 010
313 minus3299 plusmn 008
0
50
100
150
200
250
300
350
400
450
222 272 322 372 422Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
0
002
004
006
008
01
012
014
Abso
rban
ce
a
b
Figure 6 The overlap of the fluorescence spectrum of HSA (a)and the absorption spectrum of isoimperatorin (b) when [HSA] =[isoimperatorin] = 10 120583MThe enzyme was excited at 295 nm
the acceptor and (c) the donor and acceptor molecules areclose enough with the maximum distance not to exceeding7 nm [45]
There is a good overlapping between the fluorescenceemission spectrum of HSA and absorption spectra of isoim-peratorin (Figure 6) As the fluorescence emission of proteinwas affected by the excitation light around 295 nm thespectrum ranging from 300ndash400 nm was chosen to calculatethe overlapping integral The value of 119869 was calculated to be473 plusmn 005 times 10
minus15 cm3 Lmolminus1 by integrating the overlapspectra for 300ndash400 nm In the present case 1198702 = 23119873 = 136 and Φ = 0118 [21] according to (6) and (7) wecould calculate that 119864 = 045 plusmn 006 119877
0= 322 plusmn 03 nm and
119903 = 29 plusmn 02 nm As mentioned earlier (Section 321) HSAposses one tryptophan residue (Trp 214) and the calculated119903 is actually the average value between the bound drug andTrp 214 Since 119903 is much smaller than 7 nm and 05119877
0lt
119903 lt 15119877
0[41] therefore the energy transfer from HSA
to isoimperatorin occurs with high probability which alsoexplains the efficient quenching of the HSA fluorescence
34 PSH Determination in the Absence and Presence of Isoim-peratorin Several methods exist for measuring the surfacehydrophobicity of proteins that ANS experiment is one ofthem ANS as an extrinsic fluorescent probe is extremelysensitive to polarity of the solvent In aqueous solutions itfluoresces very weakly but upon binding to hydrophobicpatches of proteins its quantum yield increases significantly[29] Thus ANS fluorescence may be applied for monitoringpossible changes in protein surface hydrophobicity induced
0
200
400
600
800
1000
0 10 20 30 40 50
Fluo
resc
ence
inte
nsity
(au
)
[ANS] (120583M)
Figure 7 Binding of ANS to HSA (1120583M) in the absence (∙) andpresence (998771) of 3120583M of isoimperatorin
upon drug binding At a fixed concentration (1 120583M) of HSAand increasing concentrations of ANS (0ndash50120583M) fluores-cence intensity of ANS was measured in the absence andpresence of 3120583M isoimperatorin (Figure 7)
A typical hyperbolic response is observed which demon-strates the saturation nature of ANS binding toHSA It can beseen that the hyperbolic response of ANS fluorescence in thepresence of isoimperatorin is slightly different in comparisonto the absence of the drug The values of 119865max and 119870
app119889
forANS are obtained from the Scatchard plot (Figure 8) andlisted in Table 3 The value of 119870app
119889
for ANS binding in theabsence of drug (09 plusmn 005) was increased in the presenceof 3 120583M isoimperatorin (14 plusmn 007) which shows the weakerbinding of ANS to the HSA-isoimperatorin complex ThenPSH was calculated in the absence and the presence of3 120583M isoimperatorin by using 119865max and119870
app119889
values (Table 3)Decreasing the PSH index of HSA (from 150794 plusmn 1146 to80612 plusmn 007) suggests that the surface hydrophobicity ofHSA is decreased by sim465 upon drug binding
35 Synchronous Fluorescence Spectroscopy Synchronous flu-orescence spectroscopy introduced by Lody has been usedto characterize complex mixtures which can provide finger-prints of complex samples [46] The synchronous fluores-cence spectra can provide information about the molecularenvironment in the vicinity of the chromophore moleculesIn the synchronous fluorescence spectra the sensitivityassociated with fluorescence is maintained while several
8 The Scientific World Journal
Table 3 Surface hydrophobicity parameters for HSA in the absence and the presence of isoimperatorin
Parameters HSA HSA-IIM complex119870
app119889
(120583M) 09 plusmn 005
lowast
14 plusmn 007
lowast
119865max[HSA] 135714 plusmn 85
lowastlowast
112853 plusmn 63
lowastlowast
Surface hydrophobicity index (119865max[HSA] 119870app119889
) 150794 plusmn 1146
lowastlowast
80612 plusmn 007
lowastlowast
Data are expressed as mean plusmn SD of three measurements lowast119875 value lt 00006 lowastlowast119875 value lt 00001
0
50
100
150
200
250
300
350
400
400 500 600 700 800 900 1000Fluorescence intensity (au)
Fmax
Fluo
resc
ence
inte
nsity
(au
)[A
NS]
free
Slope = minus1Kappd
Figure 8 Scatchard plots for titration of HSA with increasingconcentrations of ANS in the absence (∙) and presence (998771) of 3 120583Mof isoimperatorin 119865max and 1119870
app119889
values were obtained from the119909-intercept and slope of this plot For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
advantages are available spectral simplification spectralbandwidth reduction and avoidance of different perturbingeffects The fluorescence spectrum of BSAHSA mainly dueto the Trp residues is sensitive to the microenvironment ofthese chromophoreswith themaximumemissionwavelengthbeing very useful in estimating the hydrophobicity aroundthe tryptophan residues It is well known that the experimenton synchronous fluorescence of HSA will provide the char-acteristic information for the Tyr and Trp residues when thescanning interval Δ120582 is fixed at 15 and 60 nm respectively[47] The shift in the position of fluorescence emissionmaximum corresponds to changes of the polarity aroundthe chromophore molecule A blue shift of 120582max means thatthe amino acid residues are located in a more hydrophobicenvironment and are less exposed to the solvent while a redshift of 120582max implies that the amino acid residues are in apolar environment and are more exposed to the solvent [48]The synchronous fluorescence spectra of interaction betweenisoimperatorin and HSA are presented in Figure 8 It wasshown that the fluorescence intensity of HSA decreased regu-larly along with the addition of isoimperatorin which furtherdemonstrated the occurrence of fluorescence quenching inthe binding process Moreover there is no significant shiftof the maximum emission wavelength with Δ120582 = 15 nm or60 nm (Figures 9(a) and 9(b)) which implies that interaction
of isoimperatorin with HSA does not affect the conformationof the region around the Tyr or Trp residues
As well we can obtain the information about the locationof isoimperatorin binding site from the synchronous fluores-cence dataThe ratios of synchronous fluorescence quenching(RSFQ) were used to determine the binding sites of the drugsto the HSA molecule in drug-HSA complexes The RSFQvalues were calculated from the following equation
RSFQ = 1 minus
119865
119865
0
(11)
where 119865 and 1198650are the synchronous fluorescence intensities
of HSA in the presence and the absence of drug respectivelyIt was obvious from Figure 9(c) that the RSFQ results forΔ120582 = 60 nmwere slightly higher than the corresponding onesfor Δ120582 = 15 nm As illustrated in Figure 9(c) addition ofisoimperatorin resulted in strong fluorescence quenching ofTrp residue (sim47 reduction in fluorescence intensity) con-firming that isoimperatorin reached subdomain IIA whereonly one Trp residue (Trp 214) in HSA is located It wasalso observed from Figure 9(c) that the fluorescence strengthof Tyr residues decreased about sim32 in the presence ofisoimperatorin So the binding of isoimperatorin to HSAquenched flourescence intensity of Tyr and Trp Since Trp214 (the only Trp of HSA) is at site I and Tyr 263 (hasthe strong fluorescence in the case of Tyr residues andmay be one of the residues responsible for the synchronousfluorescence spectrum of HSA when Δ120582 is fixed at 15 nm) isin subdomain IIA (site I) it can be concluded that Trp andTyr residues of HSA may have equal accessibility to bindingsite of isoimperatorin [49] The above results indicated thatisoimperatorin can bind to HSA in the hydrophobic cavity(site I) on subdomain IIA which are in full agreement withquenching competitive binding experiments and theoreticaldocking technique
36 Site Marker Competitive Binding Experiments In orderto further classify drug binding site on HSA competitivebinding experiments have been carried out Warfarin andibuprofen are two markers specific for binding to HSA thatbind to sites I and II which are corresponded to the sub-domains IIA and IIIA respectively Therefore to determinethe location of the isoimperatorin binding site on HSA thecompetitive displacement experimentswere carried out usingwarfarin as a characteristicmarker for site I (Figure 10(a)) andibuprofen for site II (Figure 10(b)) [24 31] In the site markercompetitive experiment isoimperatorin was gradually addedto the solution of HSA HSA-ibuprofen or HSA-warfarincomplex and then fluorescence intensity of system wasrecorded
The Scientific World Journal 9
0
100
200
300
400
500
600
265 270 275 280 285 290 295 300Wavelength (nm)
1
8
Fluo
resc
ence
inte
nsity
(au
)
(a)
Fluo
resc
ence
inte
nsity
(au
)
0
50
100
150
200
250
300
350
400
250 260 270 280 290 300Wavelength (nm)
1
8
(b)
0
10
20
30
40
50
0 05 1 15 2 25 3
RSFQ
()
Isoimperatorin (120583M)
0
02
04
06
08
1
12
005 1 15 2 25 3 35
Isoimperatorin (120583M)
F0
F
(c)
Figure 9 Synchronous fluorescence spectra of interaction between HSA and isoimperatorin (a) at Δ120582 = 15 nm and (b) at Δ120582 = 60 nmConcentration ofHSAwas 30 120583Mwhile concentrations of isoimperatorinwere 0 04 08 12 16 2 25 and 3 120583Mfrom 1 to 8 (c) Comparativeevaluation of isoimperatorin effect on the RSFQ of HSA [HSA] = 3 120583M (◼) Δ120582 = 15 nm and (∙) Δ120582 = 60 nm
As shown in Figure 10(a) with addition of warfarin in theHSA solution the flourescence intensity was slightly higherthan that of without warfarin (and including the red shift)Then after adding the isoimperatorin into the above systemthe flourescence intensity of HSA solution with warfarinheld in equimolar decreased gradually and the intensitywas much lower than that of without warfarin displayingthat the binding of the isoimperatorin to HSA was affectedafter adding warfarin On the contrary in the presence ofibuprofen the flourescence intensity of the HSA-ibuprofencomplex almost had no difference from that recordedwithoutibuprofen under the same conditions (Figure 10(b)) whichindicated that site II marker did not prevent the binding ofisoimperatorin in its usual binding location The next stepprotein intrinsic fluorescence was measured and the relativefluorescence intensities (119865119865
0) versus ligand concentration
([isoimperatorin]) plots are displayed in Figure 10(c) Asshown in Table 4 the binding constant was remarkablydecreased after addition of warfarin while the addition ofibuprofen results in only a small difference These resultsindicate that warfarin can displace the isoimperatorin but
ibuprofen has little effect on the binding of isoimperatorinto HSA In other words these results suggest that isoim-peratorin probably competes with warfarin for binding toHSA [24] Experimental observations were followed up withdocking studies where isoimperatorin was docked to HSAto determine the preferred binding site on the proteinAs described above the 3D crystal structure of HSA is amonomer consisting of three homologous domains whichassemble to form a heart-shaped molecule [4 6] Each ofthe structurally similar 120572-helix domains (IndashIII) has twosubdomains (A andB) with six120572-helices in subdomainA andfour 120572-helices in subdomain BThe crystal structure analysesindicate that the principal regions of ligand binding sitesin HSA are located in hydrophobic cavities in subdomainsIIA and IIIA which are consistent with Sudlowrsquos site I andSudlowrsquos site II respectively Several studies have shown thatHSA is able to bind many ligands in several binding sites[38] In this paper the molecular docking model of theHSAndashisoimperatorin complex was conducted on the basisof the crystal structure of HSA to find the binding positionof isoimperatorin The best docking energy result is shown
10 The Scientific World Journal
0
100
200
300
400
500
600
310 335 360 385 410Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA1
8
(a)
0
100
200
300
400
500
600
700
305 325 345 365 385 405Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA
1
8
(b)
0
05
1
15
0 1 2 3
HSAIbuprofen
Warfarin
Isoimperatorin (120583M)
F0
F
(c)
Figure 10 Effect of isoimperatorin on the quenching of HSA-warfarin complex (a) andHSA-ibuprofen complex (b) fluorescence From 1 to 8[HSA] = [warfarin or iboprufen] = 3 120583M [isoimperatorin] = 0 04 08 12 16 2 25 and 3 120583M All reproduced data shown are representativeof at least 3 independent experiments (c) Fluorescence quenching profiles of HSA (⧫) HSA-ibobrofen (◼) and HSA-warfarin (998771) in thepresence of various concentration of isoimperatorin
in Figure 11 It can be seen that isoimperatorin was situatedwithin subdomain IIA in Sudlowrsquos site I formed by six 120572-helices This figure shows the location of Trp 214 and also thebinding of isoimperatorin in HSA As this fluorescent residueis in subdomain IIA and due to the proximity of the bindingsite of isoimperatorin to the Trp 214 it can be concluded thatisoimperatorin has one reactive site onHSAandprobably thissite is at or near the site I The above docking results are ingood agreement with the displacement experiments results
and revealed that isoimperatorin binds to Sudlowrsquos site I ofHSA
37 Effect of Isoimperatorin on HSA Esterase Activity Pre-liminary studies showed that albumin in addition to itsligand binding capabilities has remarkable promiscuouscatalytic activities toward a broad range of organicmoleculesincluding esters amides phosphates and benzisoxazolesHuman serum albumin had significantly greater esterase-like
The Scientific World Journal 11
Table 4 Binding constants for isoimperatorin-HSA interaction (at 295K) in the absence and presence of the site marker
119870
119887
times 10
minus5
(Mminus1) 119877
a
HSA + isoimperatorin (in the absence ofibuprofen and warfarin) 1110 plusmn 04
lowast 09947
HSA + isoimperatorin + ibuprofen 1099 plusmn 06
lowast 09904HSA + isoimperatorin + warfarin 789 plusmn 03
lowast 09881Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870
119887values lowast119875 value lt 00002
Trp 214
Isoimperatorin
Figure 11 Crystal structure of HSA and the positions of singletryptophan residue (Trp 214) and also the binding site of isoim-peratorin in HSA Protein backbone is shown in the ldquocartoonrdquorepresentation and isoimperatorinand Trp 214 residue in the site Iare shown in the ldquostickrdquo representation The isoimperatorin and Trp214 structures are represented by the red and blue colorsThe crystalstructure of protein was obtained from Protein Data Bank (1BM0)For interpretation of references to color in this figure legend thereader is referred to the web version of this paper
activity than bovine serum albumin and defatted albuminwas more active than untreated albumin [50] InterestinglyHSA exhibits esterase-like activity and hydrolyzes drugshaving an ester group such as aspirin (which can convertaspirin ldquoacetylsalicylic acidrdquo to salicylic acid) Lys199 Lys402Lys519 and Lys545 form the pseudoesterase active site ofHSAthat is located in site I [50] ldquoEsterase-likerdquo activity (hydrolysisof p-nitrophenol esters such as p-NPA) by subdomain III Ahas been also reported and site directed mutagenesis studieshave shown that Arg 410 and Tyr 411 are essential in thisesterase activity However further investigation revealed thatacetylation is not limited to a distinct side chain so that theHSA incubation with high concentrations of pNPAc resultsin the acetylation of 59 Lys 10 Ser 8 Thr 4 Tyr and 1 Asp[51] Because of the above difference here we set out toevaluate the effect of isoimperatorin on the esterase activityof HSA based on our previous work [19] The kinetics ofthe pseudoenzymatic hydrolysis of pNPAc was obtained inthe absence and presence of isoimperatorin and site markers(Figure 12) Accordingly warfarin and indomethacin (as siteI marker) and isoimperatorin inhibited competitively theHSA-catalyzed hydrolysis of pNPAc by binding to site I Incontrast ibuprofen and tetracycline were unable to inhibitesterase activity of HSA The enzyme inhibition results sug-gested that isoimperatorin aspirin warfarin indomethacin
0
005
01
015
02
025
03
035
0 05 1 15 2 25 3Time (min)
HSA TetracyclineIbuprofenAspirin
IndomethacinOstholeWarfarinIsoimperatorin
A400
Figure 12 4-Nitrophenyl acetate esterase activity of HSA in theabsence and presence of tetracycline and ibuprofen (site II) war-farin aspirin indomethacin and osthole (site I) and isoimperatorin(site I) Final concentration of protein pNPA and drugs were 15250 and 250 120583M respectively For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
and osthole binding sites on the HSA molecule are the sameThese results also proposed that the large site I on HSAparticipates effectively to its promiscuous esterase activity
4 Conclusion
The accumulated data on HSA-drug interactions can beused to develop quantitative structure-activity relationshipsfor albumin binding However as stated earlier adsorptiondistribution metabolism and elimination (ADME) is con-trolled by albumin [30] Poor or strong binding to SA isbelieved to affect half-time of drugs [35] and their dosagedepends on their half-life in plasma Thus the bindingaffinity of any drug to SA is one of the major factors thatdetermine the pharmacokinetics halftime and availabilityof the drug in various tissues (see [30] and referencestherein) Further complexities arise in vivo due to interactions
12 The Scientific World Journal
between drugs and endogenous ligands for HSA This isparticularly pertinent for fatty acids which normally occurin serum at levels between 01 and 2mol per mol of HSAand can both compete and cooperate with drugs bindingto the protein In certain disease states these effects areexacerbated as the fatty acid HSA molar ratio may be ashigh as six Other pathological conditions are associatedwith high (micromolar to millimolar) levels of bilirubinhemin or renal toxins (eg 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) indoxyl sulphate) which bindto the protein causing significant drug binding defects
Our major aim in this study was to distinguish thenature of the interaction of isoimperatorin to HSA usingdifferent biophysical techniques This study could be veryvaluable due to importance and presence of isoimperatorinin human diet (food-drug interaction on HSA) The resultsshowed that isoimperatorin quenched the intrinsic fluo-rescence of HSA through dynamic quenching mechanismFurthermore hydrophobic interaction played a major role inthe binding process Using Fluorescence and UV absorptionspectroscopy it has been shown that isoimperatorin inducesprotein structural changes The binding average distancebetween the solo tryptophan of the protein and the boundisoimperatorin was calculated by Forster theory as 290 plusmn02 nmAnalysis of PSHvalues elucidated that isoimperatorinhas induced some compactness in the tertiary structure ofthe enzyme The results of synchronous flourescence onHSA-isoimperatorin system elucidated that the drug binds tothe hydrophobic cavity (site I) on subdomain IIA that wasconfirmed by quenching experiments competitive bindingexperiments and docking technique
Acknowledgment
The authors gratefully acknowledge the Kermanshah Uni-versity of Medical Sciences Research Council for financialsupport of this work
References
[1] Y-J Hu Y Liu T-Q Sun A-M Bai J-Q Lu and Z-B Pi ldquoBinding of anti-inflammatory drug cromolyn sodiumto bovine serum albuminrdquo International Journal of BiologicalMacromolecules vol 39 no 4-5 pp 280ndash285 2006
[2] N Seedher and S Bhatia ldquoReversible binding of celecoxiband valdecoxib with human serum albumin using fluorescencespectroscopic techniquerdquo Pharmacological Research vol 54 no2 pp 77ndash84 2006
[3] H Isogai and N Hirayama ldquoIn silico prediction of interactionsbetween site II on human serum albumin and profen drugsrdquoISRN Pharma vol 2013 Article ID 818364 8 pages 2013
[4] YWangHYuX Shi Z LuoD Lin andMHuang ldquoStructuralmechanism of ring opening reaction of glucose by humanserum albuminrdquo The Journal of Biological Chemistry vol 288pp 15980ndash15987 2013
[5] F Keshavarz M M Alavianmehr and R Yousefi ldquoMolecularinteraction of benzalkonium Ibuprofenate and its discreteingredients with human serum albuminrdquo Physical ChemistryResearch vol 1 no 2 pp 111ndash116 2013
[6] T K Maiti K S Ghosh J Debnath and S Dasgupta ldquoBindingof all-trans retinoic acid to human serum albumin fluores-cence FT-IR and circular dichroism studiesrdquo InternationalJournal of Biological Macromolecules vol 38 no 3ndash5 pp 197ndash202 2006
[7] M-X XieM Long Y Liu C Qin and Y-DWang ldquoCharacter-ization of the interaction between human serum albumin andmorinrdquo Biochimica et Biophysica Acta vol 1760 no 8 pp 1184ndash1191 2006
[8] CD Kanakis P A TarantilisMG Polissiou S Diamantoglouand H A Tajmir-Riahi ldquoAntioxidant flavonoids bind humanserum albuminrdquo Journal of Molecular Structure vol 798 no 1-3 pp 69ndash74 2006
[9] S Marumoto and M Miyazawa ldquoStructure-activity rela-tionships for naturally occurring coumarins as 120573-secretaseinhibitorrdquo Bioorganic and Medicinal Chemistry vol 20 no 2pp 784ndash788 2012
[10] S Marumoto and M Miyazawa ldquoBiotransformation of isoim-peratorin and imperatorin by Glomerella cingulata and 120573-secretase inhibitory activityrdquo Bioorganic and Medicinal Chem-istry vol 18 no 1 pp 455ndash459 2010
[11] S E Sajjadi Y Shokoohinia and S Hemmati ldquoIsolation andidentification of furanocoumarins and a phenylpropanoid fromthe acetone extract and identification of volatile constituentsfrom the essential oil of Peucedanum pastinacifoliumrdquo Chem-istry of Natural Compounds vol 48 no 4 pp 668ndash671 2012
[12] X B Yang J Hou D Liu et al ldquoBiotransformation of isoim-peratorin by Cunninghamella blakesleana AS 3970rdquo Journal ofMolecular Catalysis B vol 88 pp 1ndash6 2013
[13] W Lili S Yehong S Qi et al ldquoIn vitro permeability analy-sis pharmacokinetic and brain distribution study in mice ofimperatorin isoimperatorin and cnidilin in Radix AngelicaeDahuricaerdquo Fitoterapia vol 85 pp 144ndash153 2013
[14] S Wang Q Chen and L He ldquoDevelopment and validationof a gas chromatography-mass spectrometry method for thedetermination of isoimperatorin in rat plasma and tissueapplication to the pharmacokinetic and tissue distributionstudyrdquo Journal of Chromatography B vol 852 no 1-2 pp 473ndash478 2007
[15] H Sadraei Y Shokoohinia S E Sajjadi and M MozafarildquoAntispasmodic effects of Prangos ferulacea acetone extract andits maincomponent osthole on ileum contractionrdquo Research inPharmaceutical Sciences vol 8 pp 137ndash144 2013
[16] H Sadraei Y Shokoohinia S E Sajjadi and B GhadirianldquoAntispasmodic effect of osthole and Prangos ferulacea extracton rat uterus smooth muscle motilityrdquo Research in Pharmaceu-tical Sciences vol 7 pp 141ndash149 2012
[17] S Sajjadi Y Shokoohinia S Gholamzadeh M Behbahani andA Fattahi ldquoAntiviral evaluation of coumarins from Prangosferulacea L (Lindl)rdquo Research in Pharmaceutical Sciences vol7 p 783 2012
[18] S E Sajjadi Y Shokoohinia and S Gholamzadeh ldquoChemicalcomposition of the essential oil of the root of Prangos ferulacea(L) Lindlrdquo Chemija vol 22 pp 178ndash180 2011
[19] N Bijari Y Shokoohinia M-R Ashrafi-Kooshk et al ldquoSpec-troscopic study of interaction between osthole and humanserum albumin identification of possible binding site of thecompoundrdquo Journal of Luminescence vol 143 pp 328ndash336 2013
[20] X Shi M Liu M Zhang et al ldquoIdentification of in vitro and invivo metabolites of isoimperatorin using liquid chromatogra-phymass spectrometryrdquo Food Chemistry vol 141 pp 357ndash3652013
The Scientific World Journal 13
[21] T Chatterjee A Pal S Dey B-K Chatterjee and PChakrabarti ldquoInteraction of virstatin with human serum albu-min spectroscopic analysis andmolecular modelingrdquo Plos Onevol 7 pp 1ndash12 2012
[22] Y-M Chen and L-H Guo ldquoCombined fluorescence and elec-trochemical investigation on the binding interaction betweenorganic acid and human serum albuminrdquo Journal of Environ-mental Sciences vol 21 no 3 pp 373ndash379 2009
[23] J R Lakowicz Principles of Fluorescence Spectroscopy KluwerAcademicPlenum Publishers New York NY USA 2nd edi-tion 1999
[24] F Ding N Li B Han F Liu L Zhang and Y Sun ldquoThe bindingof CI Acid Red 2 to human serum albumin determinationof binding mechanism and binding site using fluorescencespectroscopyrdquo Dyes and Pigments vol 83 no 2 pp 249ndash2572009
[25] M-R Ashrafi Kooshk K Mansouri M Nadi S Khodarahmiand R Khodarahmi ldquoIn vitro anti-cancer activity of nativecurcumin and protein-curcuminrdquo Systems vol 2 no 1 pp 80ndash88 1981
[26] P D Ross and S Subramanian ldquoThermodynamics of proteinassociation reactions forces contributing to stabilityrdquo Biochem-istry vol 20 no 11 pp 3096ndash3102 1981
[27] J Liu J-N Tian J Zhang Z Hu and X Chen ldquoInteractionof magnolol with bovine serum albumin a fluorescence-quenching studyrdquo Analytical and Bioanalytical Chemistry vol376 no 6 pp 864ndash867 2003
[28] A Coi A M Bianucci F Bonomi P Rasmussen G M Muraand M L Ganadu ldquoStructural perturbation of 120572B-crystallinby zinc and temperature related to its chaperone-like activityrdquoInternational Journal of Biological Macromolecules vol 42 no3 pp 229ndash234 2008
[29] M Moller and A Denicola ldquoStudy of protein-ligand bindingby fluorescencerdquoBiochemistry andMolecular Biology Educationvol 30 no 5 pp 309ndash312 2002
[30] R Khodarahmi S A Karimi M R Ashrafi Kooshk S AGhadami S Ghobadi and M Amani ldquoComparative spec-troscopic studies on drug binding characteristics and proteinsurface hydrophobicity of native and modified forms of bovineserum albumin possible relevance to change in protein struc-turefunction upon non-enzymatic glycationrdquo SpectrochimicaActa A vol 89 pp 177ndash186 2012
[31] S-F Ma M Anraku Y Iwao et al ldquoHydrolysis of angiotensinII receptor blocker prodrug olmesartan medoxomil by humanserum albumin and identification of its catalytic active sitesrdquoDrug Metabolism and Disposition vol 33 no 12 pp 1911ndash19192005
[32] H M Berman J Westbrook Z Feng et al ldquoThe protein databankrdquo Nucleic Acids Research vol 28 no 1 pp 235ndash242 2000
[33] S Sugio A Kashima SMochizukiMNoda andKKobayashildquoCrystal structure of human serum albumin at 25 A resolutionrdquoProtein Engineering vol 12 no 6 pp 439ndash446 1999
[34] G M Morris D S Goodsell R Huey and A J OlsonldquoDistributed automated docking of flexible ligands to proteinsparallel applications of AutoDock 24rdquo Journal of Computer-Aided Molecular Design vol 10 no 4 pp 293ndash304 1996
[35] F Faridbod M R Ganjali B Larijani et al ldquoInteraction studyof pioglitazone with albumin by fluorescence spectroscopy andmolecular dockingrdquo Spectrochimica Acta A vol 78 no 1 pp96ndash101 2011
[36] D SGoodsell andA JOlson ldquoAutomated docking of substratesto proteins by simulated annealingrdquo Proteins vol 8 no 3 pp195ndash202 1990
[37] D S Goodsell G M Morris and A J Olson ldquoAutomateddocking of flexible ligands applications of AutoDockrdquo Journalof Molecular Recognition vol 9 no 1 pp 1ndash5 1996
[38] G Sudlow D J Birkett and D N Wade ldquoThe characterizationof two specific drug binding sites on human serum albuminrdquoMolecular Pharmacology vol 11 no 6 pp 824ndash832 1975
[39] Y Yue X Chen J Qin and X Yao ldquoCharacterization of themangiferin-human serum albumin complex by spectroscopicandmolecularmodeling approachesrdquo Journal of Pharmaceuticaland Biomedical Analysis vol 49 no 3 pp 753ndash759 2009
[40] A K Shaw and S K Pal ldquoSpectroscopic studies on the effectof temperature on pH-induced folded states of human serumalbuminrdquo Journal of Photochemistry and Photobiology B vol 90no 1 pp 69ndash77 2008
[41] S Ranjbar S Ghobadi R Khodarahmi and H Nemati ldquoSpec-troscopic characterization of furosemide binding to humancarbonic anhydrase IIrdquo International Journal of BiologicalMacromolecules vol 50 no 4 pp 910ndash917 2012
[42] HNHou Z POi YWOYung F L Liao Y Zhang andY LiyldquoStudies on interaction between vitamin B12 and human serumalbuminrdquo Journal of Pharmaceutical and Biomedical Analysisvol 47 pp 134ndash139 2008
[43] S S Kalanur J Seetharamappa and V K A Kalalbandi ldquoChar-acterization of interaction and the effect of carbamazepine onthe structure of human serum albuminrdquo Journal of Pharmaceu-tical and Biomedical Analysis vol 53 no 3 pp 660ndash666 2010
[44] L Stryer ldquoFluorescence spectroscopy of proteinsrdquo Science vol162 no 3853 pp 526ndash533 1968
[45] B Valeur and J C Brochon New Trends in FluorescenceSpectroscopy Springer Berlin Germany 1999
[46] V Suryawanshi P Anbhule A Gore Sh Patil and G AndKolekar ldquoA spectral deciphering the perturbation of modeltransporter protein (HSA) by antibacterial pyrimidine deriva-tive pharmacokinetic and biophysical insightsrdquo Journal ofPhotochemistry and Photobiology B vol 118 pp 1ndash8 2013
[47] T Yuan AMWeljie andH J Vogel ldquoTryptophan fluorescencequenching by methionine and selenomethionine residues ofcalmodulin orientation of peptide and protein bindingrdquo Bio-chemistry vol 37 no 9 pp 3187ndash3195 1998
[48] TCOrsquoHaver A F Fell G Smith et al ldquoDerivative spectroscopyand its applications in analysisrdquo Analytical Proceedings vol 19no 1 pp 22ndash46 1982
[49] Y-Q Wang B-P Tang H-M Zhang Q-H Zhou and G-C Zhang ldquoStudies on the interaction between imidaclopridand human serum albumin spectroscopic approachrdquo Journal ofPhotochemistry and Photobiology B vol 94 no 3 pp 183ndash1902009
[50] S-J Kim H-W Rhee H-Y Park H-S Kim and J-I HongldquoFluorescent probes designed for detecting human serum albu-min on the basis of its pseudo-esterase activityrdquo Bioorganic ampMedicinal Chemistry Letters vol 23 pp 2093ndash2097 2013
[51] P Ascenzi M Gioia G Fanali M Coletta and M FasanoldquoPseudo-enzymatic hydrolysis of 4-nitrophenyl acetate byhuman serum albumin pH-dependence of rates of individualstepsrdquo Biochemical and Biophysical Research Communicationsvol 424 pp 451ndash455 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

The Scientific World Journal 3
24 Intrinsic Fluorescence Measurements Fluorescence mea-surements were performed on a Cary Eclipse (Varian) spec-trophotometer with jacketed cell holders in which tem-perature was adjusted by an external thermostated watercirculationThe excitation and emission wavelengths were setat 295 nm (to avoid the contribution from tyrosine residues[22]) and 300ndash570 nm respectively with the excitation andemission slit widths of 5 and 10 nm respectively Fluorescenceemission spectra of the HSA were measured in the absenceand presence of the various concentrations of isoimperatorin(02 04 06 08 1 15 2 25 3 35 4 5 6 8 and 10 120583M)Thefinal enzyme concentration was 3 120583M
Synchronous fluorescence spectra of HSA with variousconcentrations of isoimperatorin were obtained (Δ120582 = 15 nmandΔ120582 = 60 nm) with the excitation and emission slit widthsof 5 and 10 nm respectively
241 The Inner Filter Effect The inner filter effect can bea problem for any fluorescencemeasurement especiallywherean absorbing component is being titrated into the cuvetteDuring a fluorescence titration experiment at the wavelengthof excitation or at the wavelengths used to follow an emissionthe absorption of the molecule added resulted in a spuriousdecrease in the observed fluorescence intensity [23] In thesteady-state fluorescence experiments absorption spectrumof isoimperatorin overlapswith the excitation and emission ofHSAThus it is very important to subtract such an effect fromthe raw quenching data The fluorescence data was correctedfor inner filter effect using
119865corr =119865obs antilog (119860em + 119860ex)
2
(1)
where 119865corr and 119865obs are the corrected and observed flu-orescence intensity 119860ex and 119860em are the optical densityof the sample at the excitation and emission wavelengthsrespectively
242 Determination of Quenching Mechanism In order toclarify the mechanism of HSA fluorescence quenching byisoimperatorin the quenching experiments were carried outat 298 303 308 and 313 K where HSA does not undergoany considerable thermal denaturation The decrease influorescence intensity at 120582max was analyzed using the well-known Stern-Volmer equation [24]
119865
0
119865
= 1 + 119870SV [119876] (2)
where 1198650and 119865 are the fluorescence intensities in the absence
and presence of quencher (isoimperatorin) respectively the119870SV is the Stern-Volmer quenching constant and [119876] is theconcentration of the compound isoimperatorin
243 Determination of Association Constant and Number ofBinding Sites The association constant (119870
119887) and number of
binding sites (119899) for binding of small molecules to set of
equivalent sites on amacromolecule can be determined usingthe modified Stern-Volmer equation
log 119865
0minus 119865
119865
= log119870119887+ 119899 log [119876] (3)
where 1198650and 119865 are the fluorescence intensities of HSA in
the absence and presence of the quencher (isoimperatorin)respectively 119870
119887is the association constant and 119899 is the
number of binding sites per HSA [25]The values of 119899 and119870119887
were obtained from the slope and intercept of the modifiedStern-Volmer plot respectively
244 Thermodynamic Analysis of the Binding Process Inorder to explain the nature of the interaction betweenisoimperatorin and HSA the association constant of HSA-isoimperatorin complex was determined at four differenttemperatures (298 303 308 and 313 K)The thermodynamicparameters of binding entropy change (Δ119878∘) and enthalpychange (Δ119867∘) were obtained from vanrsquot Hoff equation
ln 119870119887=
minusΔ119867
∘
119877119879
+
Δ119878
∘
119877
(4)
where119870119887is the association constant at the given temperature
and119877 is the universal gas constantThe values ofΔ119878∘ andΔ119867∘can be obtained from the intercept and slope of the vanrsquot Hoffplot respectively and the free energy change (Δ119866∘) can beestimated by using the Gibbs equation [26]
Δ119866
∘
= Δ119867
∘
minus 119879Δ119878
∘
(5)
25 Energy Transfer between HSA and Isoimperatorin Thedistance between the acceptor (ligand) and the donor (tryp-tophan residues in the protein) 119903 according to Forsterrsquosnonradiation energy transfer theory could be calculated bythe following equation [21]
119864 = 1 minus (
119865
119865
0
) =
119877
6
0
119877
6
0
+ 119903
6
0
(6)
where 119864 is the efficiency of energy transfer between donorand acceptor 119865 and 119865
0are the fluorescence intensities of
donor in the presence and absence of acceptor respectivelyand 119877
0is a critical distance at which the efficiency of energy
transfer is 50 The value of 1198770was calculated using
119877
6
0
= 88 times 10
minus25
119870
2
119873
minus4
Φ119869(7)
where 1198702 is the spatial orientation factor describing thegeometry of the donor and acceptor dipoles 119873 is theaverage refractive index of the mediumΦ is the fluorescencequantum yield of the donor in the absence of acceptor and 119869expresses the degree of spectral overlap between the emissionspectrum of the donor and the absorption spectrum of theacceptor 119869 can be given by
119869 = sum119865 (120582) 120576 (120582) 120582
4
Δ120582 (8)
where 119865(120582) is the fluorescence intensity of the donor atwavelength 120582 and 120576(120582) is the molar absorption coefficient
4 The Scientific World Journal
of the acceptor at 120582 So 119869 can be evaluated by integratingthe overlap between emission spectrum of the donor andabsorption spectrum of the acceptor [27]
The absorption spectrum of isoimperatorin (10 120583M) wasrecorded in the range of 222ndash450 nmThe emission spectrumof HSA (10 120583M) was also recorded in the range of 222ndash450 nm with excitation and emission slit widths of 5 and5 nm respectivelyThe overlap of the absorption spectrum ofisoimperatorin with the fluorescence emission spectrum ofHSA was used to calculate the efficiency of energy transfer
26 Protein Surface Hydrophobicity (PSH) DeterminationThe ANS binding properties of HSA can be investigatedby measuring 119870
app119889
and 119865max of the HSA-ANS complexin the absence and presence of isoimperatorin 119870app
119889
is theapparent dissociation constant of the fluorescent ANS-HSAcomplex and 1119870
app119889
is the binding affinity of ANS tothe protein in the absence and presence of isoimperatorin[28] 119865max is the maximum fluorescence intensity at thesaturated ANS concentration which indicates the numberof surface hydrophobic sites of the protein This methodinvolves titration of the enzyme solutions in the presenceof increasing concentrations of ANS The values of 1119870app
119889
and 119865max can be obtained from the slope and the 119909-interceptof the Scatchard plot respectively In this type of Scatchardplot the fluorescence intensity[ANS]free is drawn versusfluorescence intensity The free ANS concentration must bedetermined via plotting fluorescence intensity versus totalANS concentration In dilute solutions of ANS a linearrelationship exists between fluorescence intensity (119865) andANS concentration (119888) (119865 = 119861119888 where 119861 is the propor-tionality coefficient between fluorescence intensity and ANSconcentration) From a 1mM stock solution ANS was addedto the final concentrations ranging from 0 to 50 120583M Theenzyme concentration was 1 120583M It is assumed that in verydilute ANS solutions (0-1 120583M) all of the ANS molecules arebound to theHSA and thus B (the proportionality coefficientbetween fluorescence intensity and ANS concentration) canbe determined from the slope of the linear portion offluorescence intensity plot versus ANS concentration Foreach of the different total concentrations of ANS used theconcentration of bound ANS molecules was calculated usingthe following equation [ANS]bound = 119865119861 and the free ANSmolecules concentration was calculated from the differencebetween total and bound ANS concentrations ([ANS]free =[ANS]total minus [ANS]bound) [29 30]
The protein surface hydrophobicity index in the absenceand presence of the drug can be calculated by applying thefollowing equation [28]
PSH =
119865max
[HSA] 119870app119889
(9)
Following excitation at 380 nm the fluorescence emissionincrement was recorded at 470 nm until no further increasein fluorescence intensity was observed
27 Competitive Experiments In order to elucidate the bind-ing site of isoimperatorin onHSA we carried out competitive
experiments at the presence of two known drugs (Warfarinas site I marker and Ibuprofen as a drug that preferentiallybinds to site II) [24 31] The concentration of HSA and war-farinibuprofen was stabilized at 10times10minus6M Isoimperatorinwas then gradually added to theHSAHSA-warfarin orHSA-ibuprofen mixtures An excitation wavelength of 295 nm wasselected and flourescence spectra were recorded in the rangeof 300ndash450 nm
Also possible inhibitory effect of isoimperatorin indo-methacin tetracycline ibuprofen aspirin osthole and war-farin on the esterase activity of HSA was evaluated
28 Docking Procedure The AutoDock 405 docking pro-gram was applied to study the interaction between theisoimperatorin and HSA The crystal structure of HSA wasobtained from the Protein Data Bank Of more than 50X-ray crystallographic structures related to human serumalbumin in ProteinData Bank [32] entry with PDB ID1BM0[33] was chosen for dockings because of no missing atomsno cocrystallized ligand and having a reasonably goodresolution (25 A) All the water molecules were removedusing a plain text editor The polar hydrogen partial chargeand solvation parameters for use with the AutoDock 405simulation were applied using AutoDock-Tools The dockingsimulations were carried out with the rigid HSA and aflexible ligand [34] Using AutoGrid tools the grid mapswere generated adequately large to include the binding site(s)of protein as well as significant regions of the surroundingsurfaceThe points of the grids were thus 126times126times126witha grid spacing of 0375 A [35] Afterwards the most suitablestructure for the flexible ligand molecule was optimized bythe rotation of all single bonds in the ligand molecule Thegrid parameter file and the docking parameter file were setup by the AutoDockTools program [36] The binding modeof isoimperatorin was predicted by using the Lamarckiangenetic algorithm search engine implemented in AutoDocksoftware package [37] for the two main drug binding sitesI and II (subdomains IIA and IIIA of HSA resp) [38]Population size was 256 and Cluster analysis was performedon the docked results using a root mean square (RMS)tolerance of 05 A Default settings were used for all otherparameters [35] At the end of the docking simulation theHSA-IIM complex with the most stable energy was adoptedas the most favorable structure of the complex for HSAMolecular graphics were prepared with PyMOL version 099beta06 (httpwwwpymolorg)
3 Results and Discussion
31 UV Absorption Spectra UV absorption spectroscopytechnique can be used to explore the structural changes ofprotein and to investigate protein-ligand complex formationHSA has two main absorption bands that one of them islocated in the range of 260ndash300 nm which is the absorptionband of the aromatic amino acids (Trp Tyr and Phe) [27 39]Figure 2 shows the UV absorption spectra of HSA in theabsence and presence of isoimperatorin As can be seen inFigure 2 HSA has strong absorbance with a peak at 280 nmand the absorbance of HSA increased with the addition of
The Scientific World Journal 5
0
05
1
15
2
25
3
35
4
200 250 300 350Wavelength (nm)
Abso
rban
ce
034
035
036
037
038
039
04
270 274 278 282 286 290
1
7
Figure 2 Absorption spectra of HSA (30 120583M) with variousamounts of isoimperatorin (1ndash7) 0 04 08 2 4 6 and 8 120583MThe absorption of each sample was measured with respect to therespective blank
0
50
100
150
200
250
300
350
400
450
300 350 400 450 500 550Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
1
16
116
Figure 3 Effect of isoimperatorin on fluorescence spectra of HSA(119879 = 298K pH 75) Curve (1) 30 120583MHSA Curves (2ndash16) 30 120583MHSA in the presence of 02 04 06 08 1 15 2 25 3 4 5 6 7 8and 10120583M of isoimperatorin
isoimperatorin In addition the absorbance value of 280 nmof the isoimperatorin at a concentration of 10120583M is very low(lt003) Our results confirmed that the interaction betweenHSA and ligand took place
32 Fluorescence Measurements
321 Intrinsic Fluorescence Measurements The fluorescencespectra of HSA in the absence and presence of differentamounts of Isoimperatorin were recorded in the rangeof 300ndash570 nm upon excitation at 295 nm isoimperatorincauses a concentration dependent quenching of the intrinsicfluorescence of HSA (Figure 3) without changing the emis-sion maximum and shape of the peaks These results indi-cated that there were interactions between isoimperatorinand HSA This interaction was further confirmed by UV-visabsorption technique
0
1
2
3
4
0 2 4 6 8 10 12[Isoimperatorin] (120583M)
F0F
Figure 4 The Stern-Volmer plots for HSA fluorescence quenchingby isoimperatorin at 298 (⧫) 303 (998771) 308 (◼) and 313 (∙) K Thefinal protein concentration in 50mM phosphate pH 75 was 3 120583MBased on ldquoOne-Sample Kolmogorov-Smirnov Testrdquo test distributionwas normal
A gradual decrease in the fluorescence emission intensityof HSA is observed in the presence of increasing concentra-tions of isoimperatorin suggesting the presence of tryptophanresidue of the HSA at or near the isoimperatorin binding siteHSA has one Trp (Trp 214) and its fluorescence is solely dueto the excitation of this residue of HSA located in the bindingdomain-IIA (site I) at the bottom of a 12 A deep crevice [40]
Emerging of a new emission peak at 505 nm uponisoimperatorin addition can be attributed to the bounddrug and radiationless energy transfer between the HSAtryptophan and isoimperatorin [41] The appearance of anisoactinic point at 430 nm might also indicate the existenceof equilibrium between bound and free drug In additionsuch equilibrium may emphasize the formation of the drug-protein complex [42]
322 Fluorescence Quenching Mechanism Quenching canoccur by different mechanisms which is usually classifiedas dynamic quenching and static quenching In dynamicquenching increasing the temperature results in faster dif-fusion and hence larger amounts of collision which causesraising of the quenching constant Opposing to dynamicquenching in static one increasing the temperature weakensthe stability of the formed complex and hence reduces thequenching constant [19]
Fluorescence quenching of HSA upon interaction withisoimperatorin was evaluated at different temperatures (298303 308 and 313 K) and the results are shown in Figure 4As indicated in this figure the Stern-Volmer plots are lin-ear with the slopes increasing with increasing temperature(the linearity in this plot confirmed one-to-one interactionbetween isoimperatorin and HSA) The calculated values of119870SV at different temperatures are listed in Table 1 It can
6 The Scientific World Journal
Table 1 Stern-Volmer quenching constant (119870SV) quenching rate constant (119896119902) association constant (119870119887
) and number of binding sites (119899)for binding of isoimperatorin to HSA in the different temperatures
119879 (K) 10
minus4
times 119870SV(Mminus1)
10
minus12
times 119870
119902
(Mminus1sdotsminus1)119877
a 10
minus5
times 119870
119887
(Mminus1)Number of biding
sites (119899) 119877
a
2981698 plusmn 013 1698 plusmn 013
099621102 plusmn 010 0862 09841
3032019 plusmn 017 2019 plusmn 017
099551149 plusmn 013 0851 09886
3082333 plusmn 016 2333 plusmn 016
099651271 plusmn 022 0844 09950
3132639 plusmn 019 2639 plusmn 019
099811322 plusmn 017 0822 09917
Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870SV values
be found from Table 1 that the value of 119870SV increased withincreasing temperature in the presence of isoimperatorinwhich indicates the involved binding forces are mainlyhydrophobic interactions (endothermic apolar interactionsare strengthened with increasing temperature) Also the rateconstant of the quenching process (119896
119902) which is equal to119870SV
divided by 1205910(with assuming 120591
0= 10
minus8 s)
119896
119902=
119870SV120591
0
(10)
was calculated (Table 1) The results revealed that the valuesof 119896119902were much greater than the maximum diffusion rate
constant of the biomolecule (2 times 1010Mminus1 sminus1) which indi-cated that the fluorescence quenching of HSA was initiatedby complex formation between HSA and isoimperatorinconfirming that the mechanism of quenching is static [19]
323 Determination of Association Constant (119870119887) and Num-
ber of Binding Sites (119899) The values of association constant119870
119887 and the number of binding sites 119899 at different temper-
atures are obtained from the modified Stern-Volmer plots(Figure 5) and listed in Table 1 It can be found from thesedata that the 119899 values at different temperatures are keptaround unity which shows the existence of one binding sitein HSA for isoimperatorin The value of 119870
119887is significant to
understand the distribution of the drug in plasma since theweak binding can lead to a short life time or poor distributionwhile strong binding can decrease the concentration of freedrug in plasma The value of 119870
119887illustrated that there is a
strong binding force between isoimperatorin and HSA Itwas found that the binding constant increased with increasein temperature resulting in an increment of the stabilityof the isoimperatorin-HSA complex it also implies thatisoimperatorin can be tightly stored and carried by HSA inthe body
324 Forces Involved in the Binding Process Consideringthe dependence of binding constant on temperature a ther-modynamic process was considered to be responsible forthe formation of a complex Therefore the thermodynamicparameters dependent on temperatures were analyzed Theacting forces between a small molecule and macromoleculemainly include hydrogen bonds van der Waals forces elec-trostatic forces and hydrophobic interaction forces [23] Inorder to obtain the information about the forces persisting in
minus15
minus13
minus11
minus09
minus07
minus05
minus03
minus01
01
03
05
minus68 minus65 minus62 minus59 minus56 minus53 minus5
log[isoimperatorin]
log[(F0minusF)F]
Figure 5 The modified Stern-Volmer plots for HSA fluorescencequenching by isoimperatorin at 298 (⧫) 303 (998771) 308 (◼) and 313 (∙)K For interpretation of references to color in this figure legend thereader is referred to the web version of this article
the present binding process the thermodynamic parametersof binding were obtained fromVanrsquot Hoff plot followed by theGibbs equation (data not shown) Table 2 shows the values ofΔ119867
∘ andΔ119878∘ obtained for the binding site from the slopes andordinates at the origin of the fitted lines From Table 2 it canbe seen that the negative sign for free energy (Δ119866∘) meansthat the interaction process is spontaneous The positiveenthalpy (Δ119867∘) and entropy (Δ119878∘) values of the interaction ofisoimperatorin and HSA indicate that the binding is mainlyentropy-driven and the enthalpy is unfavorable for it thehydrophobic forces play a major role in the reaction [43]
33 Nonradiative Energy Transfer from HSA to Isoimpera-torin The Forster nonradiative energy transfer (FRET) hasbeen used as a ldquospectroscopic rulerrdquo for measurement ofthe distance between the acceptor (ligand) and the donor(tryptophan residues in the protein) [44] According to thistheory there will be FRET between two molecules if thefollowing conditions were satisfied (a) the donor moleculeemits fluorescence (b) the emission spectrum of the donorhas an enough overlap with the absorption spectrum of
The Scientific World Journal 7
Table 2 Thermodynamic parameters for binding isoimperatorin to HSA
119879 (K) Δ119878
∘ (Jsdotmolminus1sdotKminus1) Δ119867
∘ (kJsdotmolminus1) Δ119866
∘ (kJsdotmolminus1)298 minus2969 plusmn 009
303 17820 plusmn 011 2993 plusmn 013 minus3071 plusmn 007
308 minus3169 plusmn 010
313 minus3299 plusmn 008
0
50
100
150
200
250
300
350
400
450
222 272 322 372 422Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
0
002
004
006
008
01
012
014
Abso
rban
ce
a
b
Figure 6 The overlap of the fluorescence spectrum of HSA (a)and the absorption spectrum of isoimperatorin (b) when [HSA] =[isoimperatorin] = 10 120583MThe enzyme was excited at 295 nm
the acceptor and (c) the donor and acceptor molecules areclose enough with the maximum distance not to exceeding7 nm [45]
There is a good overlapping between the fluorescenceemission spectrum of HSA and absorption spectra of isoim-peratorin (Figure 6) As the fluorescence emission of proteinwas affected by the excitation light around 295 nm thespectrum ranging from 300ndash400 nm was chosen to calculatethe overlapping integral The value of 119869 was calculated to be473 plusmn 005 times 10
minus15 cm3 Lmolminus1 by integrating the overlapspectra for 300ndash400 nm In the present case 1198702 = 23119873 = 136 and Φ = 0118 [21] according to (6) and (7) wecould calculate that 119864 = 045 plusmn 006 119877
0= 322 plusmn 03 nm and
119903 = 29 plusmn 02 nm As mentioned earlier (Section 321) HSAposses one tryptophan residue (Trp 214) and the calculated119903 is actually the average value between the bound drug andTrp 214 Since 119903 is much smaller than 7 nm and 05119877
0lt
119903 lt 15119877
0[41] therefore the energy transfer from HSA
to isoimperatorin occurs with high probability which alsoexplains the efficient quenching of the HSA fluorescence
34 PSH Determination in the Absence and Presence of Isoim-peratorin Several methods exist for measuring the surfacehydrophobicity of proteins that ANS experiment is one ofthem ANS as an extrinsic fluorescent probe is extremelysensitive to polarity of the solvent In aqueous solutions itfluoresces very weakly but upon binding to hydrophobicpatches of proteins its quantum yield increases significantly[29] Thus ANS fluorescence may be applied for monitoringpossible changes in protein surface hydrophobicity induced
0
200
400
600
800
1000
0 10 20 30 40 50
Fluo
resc
ence
inte
nsity
(au
)
[ANS] (120583M)
Figure 7 Binding of ANS to HSA (1120583M) in the absence (∙) andpresence (998771) of 3120583M of isoimperatorin
upon drug binding At a fixed concentration (1 120583M) of HSAand increasing concentrations of ANS (0ndash50120583M) fluores-cence intensity of ANS was measured in the absence andpresence of 3120583M isoimperatorin (Figure 7)
A typical hyperbolic response is observed which demon-strates the saturation nature of ANS binding toHSA It can beseen that the hyperbolic response of ANS fluorescence in thepresence of isoimperatorin is slightly different in comparisonto the absence of the drug The values of 119865max and 119870
app119889
forANS are obtained from the Scatchard plot (Figure 8) andlisted in Table 3 The value of 119870app
119889
for ANS binding in theabsence of drug (09 plusmn 005) was increased in the presenceof 3 120583M isoimperatorin (14 plusmn 007) which shows the weakerbinding of ANS to the HSA-isoimperatorin complex ThenPSH was calculated in the absence and the presence of3 120583M isoimperatorin by using 119865max and119870
app119889
values (Table 3)Decreasing the PSH index of HSA (from 150794 plusmn 1146 to80612 plusmn 007) suggests that the surface hydrophobicity ofHSA is decreased by sim465 upon drug binding
35 Synchronous Fluorescence Spectroscopy Synchronous flu-orescence spectroscopy introduced by Lody has been usedto characterize complex mixtures which can provide finger-prints of complex samples [46] The synchronous fluores-cence spectra can provide information about the molecularenvironment in the vicinity of the chromophore moleculesIn the synchronous fluorescence spectra the sensitivityassociated with fluorescence is maintained while several
8 The Scientific World Journal
Table 3 Surface hydrophobicity parameters for HSA in the absence and the presence of isoimperatorin
Parameters HSA HSA-IIM complex119870
app119889
(120583M) 09 plusmn 005
lowast
14 plusmn 007
lowast
119865max[HSA] 135714 plusmn 85
lowastlowast
112853 plusmn 63
lowastlowast
Surface hydrophobicity index (119865max[HSA] 119870app119889
) 150794 plusmn 1146
lowastlowast
80612 plusmn 007
lowastlowast
Data are expressed as mean plusmn SD of three measurements lowast119875 value lt 00006 lowastlowast119875 value lt 00001
0
50
100
150
200
250
300
350
400
400 500 600 700 800 900 1000Fluorescence intensity (au)
Fmax
Fluo
resc
ence
inte
nsity
(au
)[A
NS]
free
Slope = minus1Kappd
Figure 8 Scatchard plots for titration of HSA with increasingconcentrations of ANS in the absence (∙) and presence (998771) of 3 120583Mof isoimperatorin 119865max and 1119870
app119889
values were obtained from the119909-intercept and slope of this plot For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
advantages are available spectral simplification spectralbandwidth reduction and avoidance of different perturbingeffects The fluorescence spectrum of BSAHSA mainly dueto the Trp residues is sensitive to the microenvironment ofthese chromophoreswith themaximumemissionwavelengthbeing very useful in estimating the hydrophobicity aroundthe tryptophan residues It is well known that the experimenton synchronous fluorescence of HSA will provide the char-acteristic information for the Tyr and Trp residues when thescanning interval Δ120582 is fixed at 15 and 60 nm respectively[47] The shift in the position of fluorescence emissionmaximum corresponds to changes of the polarity aroundthe chromophore molecule A blue shift of 120582max means thatthe amino acid residues are located in a more hydrophobicenvironment and are less exposed to the solvent while a redshift of 120582max implies that the amino acid residues are in apolar environment and are more exposed to the solvent [48]The synchronous fluorescence spectra of interaction betweenisoimperatorin and HSA are presented in Figure 8 It wasshown that the fluorescence intensity of HSA decreased regu-larly along with the addition of isoimperatorin which furtherdemonstrated the occurrence of fluorescence quenching inthe binding process Moreover there is no significant shiftof the maximum emission wavelength with Δ120582 = 15 nm or60 nm (Figures 9(a) and 9(b)) which implies that interaction
of isoimperatorin with HSA does not affect the conformationof the region around the Tyr or Trp residues
As well we can obtain the information about the locationof isoimperatorin binding site from the synchronous fluores-cence dataThe ratios of synchronous fluorescence quenching(RSFQ) were used to determine the binding sites of the drugsto the HSA molecule in drug-HSA complexes The RSFQvalues were calculated from the following equation
RSFQ = 1 minus
119865
119865
0
(11)
where 119865 and 1198650are the synchronous fluorescence intensities
of HSA in the presence and the absence of drug respectivelyIt was obvious from Figure 9(c) that the RSFQ results forΔ120582 = 60 nmwere slightly higher than the corresponding onesfor Δ120582 = 15 nm As illustrated in Figure 9(c) addition ofisoimperatorin resulted in strong fluorescence quenching ofTrp residue (sim47 reduction in fluorescence intensity) con-firming that isoimperatorin reached subdomain IIA whereonly one Trp residue (Trp 214) in HSA is located It wasalso observed from Figure 9(c) that the fluorescence strengthof Tyr residues decreased about sim32 in the presence ofisoimperatorin So the binding of isoimperatorin to HSAquenched flourescence intensity of Tyr and Trp Since Trp214 (the only Trp of HSA) is at site I and Tyr 263 (hasthe strong fluorescence in the case of Tyr residues andmay be one of the residues responsible for the synchronousfluorescence spectrum of HSA when Δ120582 is fixed at 15 nm) isin subdomain IIA (site I) it can be concluded that Trp andTyr residues of HSA may have equal accessibility to bindingsite of isoimperatorin [49] The above results indicated thatisoimperatorin can bind to HSA in the hydrophobic cavity(site I) on subdomain IIA which are in full agreement withquenching competitive binding experiments and theoreticaldocking technique
36 Site Marker Competitive Binding Experiments In orderto further classify drug binding site on HSA competitivebinding experiments have been carried out Warfarin andibuprofen are two markers specific for binding to HSA thatbind to sites I and II which are corresponded to the sub-domains IIA and IIIA respectively Therefore to determinethe location of the isoimperatorin binding site on HSA thecompetitive displacement experimentswere carried out usingwarfarin as a characteristicmarker for site I (Figure 10(a)) andibuprofen for site II (Figure 10(b)) [24 31] In the site markercompetitive experiment isoimperatorin was gradually addedto the solution of HSA HSA-ibuprofen or HSA-warfarincomplex and then fluorescence intensity of system wasrecorded
The Scientific World Journal 9
0
100
200
300
400
500
600
265 270 275 280 285 290 295 300Wavelength (nm)
1
8
Fluo
resc
ence
inte
nsity
(au
)
(a)
Fluo
resc
ence
inte
nsity
(au
)
0
50
100
150
200
250
300
350
400
250 260 270 280 290 300Wavelength (nm)
1
8
(b)
0
10
20
30
40
50
0 05 1 15 2 25 3
RSFQ
()
Isoimperatorin (120583M)
0
02
04
06
08
1
12
005 1 15 2 25 3 35
Isoimperatorin (120583M)
F0
F
(c)
Figure 9 Synchronous fluorescence spectra of interaction between HSA and isoimperatorin (a) at Δ120582 = 15 nm and (b) at Δ120582 = 60 nmConcentration ofHSAwas 30 120583Mwhile concentrations of isoimperatorinwere 0 04 08 12 16 2 25 and 3 120583Mfrom 1 to 8 (c) Comparativeevaluation of isoimperatorin effect on the RSFQ of HSA [HSA] = 3 120583M (◼) Δ120582 = 15 nm and (∙) Δ120582 = 60 nm
As shown in Figure 10(a) with addition of warfarin in theHSA solution the flourescence intensity was slightly higherthan that of without warfarin (and including the red shift)Then after adding the isoimperatorin into the above systemthe flourescence intensity of HSA solution with warfarinheld in equimolar decreased gradually and the intensitywas much lower than that of without warfarin displayingthat the binding of the isoimperatorin to HSA was affectedafter adding warfarin On the contrary in the presence ofibuprofen the flourescence intensity of the HSA-ibuprofencomplex almost had no difference from that recordedwithoutibuprofen under the same conditions (Figure 10(b)) whichindicated that site II marker did not prevent the binding ofisoimperatorin in its usual binding location The next stepprotein intrinsic fluorescence was measured and the relativefluorescence intensities (119865119865
0) versus ligand concentration
([isoimperatorin]) plots are displayed in Figure 10(c) Asshown in Table 4 the binding constant was remarkablydecreased after addition of warfarin while the addition ofibuprofen results in only a small difference These resultsindicate that warfarin can displace the isoimperatorin but
ibuprofen has little effect on the binding of isoimperatorinto HSA In other words these results suggest that isoim-peratorin probably competes with warfarin for binding toHSA [24] Experimental observations were followed up withdocking studies where isoimperatorin was docked to HSAto determine the preferred binding site on the proteinAs described above the 3D crystal structure of HSA is amonomer consisting of three homologous domains whichassemble to form a heart-shaped molecule [4 6] Each ofthe structurally similar 120572-helix domains (IndashIII) has twosubdomains (A andB) with six120572-helices in subdomainA andfour 120572-helices in subdomain BThe crystal structure analysesindicate that the principal regions of ligand binding sitesin HSA are located in hydrophobic cavities in subdomainsIIA and IIIA which are consistent with Sudlowrsquos site I andSudlowrsquos site II respectively Several studies have shown thatHSA is able to bind many ligands in several binding sites[38] In this paper the molecular docking model of theHSAndashisoimperatorin complex was conducted on the basisof the crystal structure of HSA to find the binding positionof isoimperatorin The best docking energy result is shown
10 The Scientific World Journal
0
100
200
300
400
500
600
310 335 360 385 410Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA1
8
(a)
0
100
200
300
400
500
600
700
305 325 345 365 385 405Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA
1
8
(b)
0
05
1
15
0 1 2 3
HSAIbuprofen
Warfarin
Isoimperatorin (120583M)
F0
F
(c)
Figure 10 Effect of isoimperatorin on the quenching of HSA-warfarin complex (a) andHSA-ibuprofen complex (b) fluorescence From 1 to 8[HSA] = [warfarin or iboprufen] = 3 120583M [isoimperatorin] = 0 04 08 12 16 2 25 and 3 120583M All reproduced data shown are representativeof at least 3 independent experiments (c) Fluorescence quenching profiles of HSA (⧫) HSA-ibobrofen (◼) and HSA-warfarin (998771) in thepresence of various concentration of isoimperatorin
in Figure 11 It can be seen that isoimperatorin was situatedwithin subdomain IIA in Sudlowrsquos site I formed by six 120572-helices This figure shows the location of Trp 214 and also thebinding of isoimperatorin in HSA As this fluorescent residueis in subdomain IIA and due to the proximity of the bindingsite of isoimperatorin to the Trp 214 it can be concluded thatisoimperatorin has one reactive site onHSAandprobably thissite is at or near the site I The above docking results are ingood agreement with the displacement experiments results
and revealed that isoimperatorin binds to Sudlowrsquos site I ofHSA
37 Effect of Isoimperatorin on HSA Esterase Activity Pre-liminary studies showed that albumin in addition to itsligand binding capabilities has remarkable promiscuouscatalytic activities toward a broad range of organicmoleculesincluding esters amides phosphates and benzisoxazolesHuman serum albumin had significantly greater esterase-like
The Scientific World Journal 11
Table 4 Binding constants for isoimperatorin-HSA interaction (at 295K) in the absence and presence of the site marker
119870
119887
times 10
minus5
(Mminus1) 119877
a
HSA + isoimperatorin (in the absence ofibuprofen and warfarin) 1110 plusmn 04
lowast 09947
HSA + isoimperatorin + ibuprofen 1099 plusmn 06
lowast 09904HSA + isoimperatorin + warfarin 789 plusmn 03
lowast 09881Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870
119887values lowast119875 value lt 00002
Trp 214
Isoimperatorin
Figure 11 Crystal structure of HSA and the positions of singletryptophan residue (Trp 214) and also the binding site of isoim-peratorin in HSA Protein backbone is shown in the ldquocartoonrdquorepresentation and isoimperatorinand Trp 214 residue in the site Iare shown in the ldquostickrdquo representation The isoimperatorin and Trp214 structures are represented by the red and blue colorsThe crystalstructure of protein was obtained from Protein Data Bank (1BM0)For interpretation of references to color in this figure legend thereader is referred to the web version of this paper
activity than bovine serum albumin and defatted albuminwas more active than untreated albumin [50] InterestinglyHSA exhibits esterase-like activity and hydrolyzes drugshaving an ester group such as aspirin (which can convertaspirin ldquoacetylsalicylic acidrdquo to salicylic acid) Lys199 Lys402Lys519 and Lys545 form the pseudoesterase active site ofHSAthat is located in site I [50] ldquoEsterase-likerdquo activity (hydrolysisof p-nitrophenol esters such as p-NPA) by subdomain III Ahas been also reported and site directed mutagenesis studieshave shown that Arg 410 and Tyr 411 are essential in thisesterase activity However further investigation revealed thatacetylation is not limited to a distinct side chain so that theHSA incubation with high concentrations of pNPAc resultsin the acetylation of 59 Lys 10 Ser 8 Thr 4 Tyr and 1 Asp[51] Because of the above difference here we set out toevaluate the effect of isoimperatorin on the esterase activityof HSA based on our previous work [19] The kinetics ofthe pseudoenzymatic hydrolysis of pNPAc was obtained inthe absence and presence of isoimperatorin and site markers(Figure 12) Accordingly warfarin and indomethacin (as siteI marker) and isoimperatorin inhibited competitively theHSA-catalyzed hydrolysis of pNPAc by binding to site I Incontrast ibuprofen and tetracycline were unable to inhibitesterase activity of HSA The enzyme inhibition results sug-gested that isoimperatorin aspirin warfarin indomethacin
0
005
01
015
02
025
03
035
0 05 1 15 2 25 3Time (min)
HSA TetracyclineIbuprofenAspirin
IndomethacinOstholeWarfarinIsoimperatorin
A400
Figure 12 4-Nitrophenyl acetate esterase activity of HSA in theabsence and presence of tetracycline and ibuprofen (site II) war-farin aspirin indomethacin and osthole (site I) and isoimperatorin(site I) Final concentration of protein pNPA and drugs were 15250 and 250 120583M respectively For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
and osthole binding sites on the HSA molecule are the sameThese results also proposed that the large site I on HSAparticipates effectively to its promiscuous esterase activity
4 Conclusion
The accumulated data on HSA-drug interactions can beused to develop quantitative structure-activity relationshipsfor albumin binding However as stated earlier adsorptiondistribution metabolism and elimination (ADME) is con-trolled by albumin [30] Poor or strong binding to SA isbelieved to affect half-time of drugs [35] and their dosagedepends on their half-life in plasma Thus the bindingaffinity of any drug to SA is one of the major factors thatdetermine the pharmacokinetics halftime and availabilityof the drug in various tissues (see [30] and referencestherein) Further complexities arise in vivo due to interactions
12 The Scientific World Journal
between drugs and endogenous ligands for HSA This isparticularly pertinent for fatty acids which normally occurin serum at levels between 01 and 2mol per mol of HSAand can both compete and cooperate with drugs bindingto the protein In certain disease states these effects areexacerbated as the fatty acid HSA molar ratio may be ashigh as six Other pathological conditions are associatedwith high (micromolar to millimolar) levels of bilirubinhemin or renal toxins (eg 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) indoxyl sulphate) which bindto the protein causing significant drug binding defects
Our major aim in this study was to distinguish thenature of the interaction of isoimperatorin to HSA usingdifferent biophysical techniques This study could be veryvaluable due to importance and presence of isoimperatorinin human diet (food-drug interaction on HSA) The resultsshowed that isoimperatorin quenched the intrinsic fluo-rescence of HSA through dynamic quenching mechanismFurthermore hydrophobic interaction played a major role inthe binding process Using Fluorescence and UV absorptionspectroscopy it has been shown that isoimperatorin inducesprotein structural changes The binding average distancebetween the solo tryptophan of the protein and the boundisoimperatorin was calculated by Forster theory as 290 plusmn02 nmAnalysis of PSHvalues elucidated that isoimperatorinhas induced some compactness in the tertiary structure ofthe enzyme The results of synchronous flourescence onHSA-isoimperatorin system elucidated that the drug binds tothe hydrophobic cavity (site I) on subdomain IIA that wasconfirmed by quenching experiments competitive bindingexperiments and docking technique
Acknowledgment
The authors gratefully acknowledge the Kermanshah Uni-versity of Medical Sciences Research Council for financialsupport of this work
References
[1] Y-J Hu Y Liu T-Q Sun A-M Bai J-Q Lu and Z-B Pi ldquoBinding of anti-inflammatory drug cromolyn sodiumto bovine serum albuminrdquo International Journal of BiologicalMacromolecules vol 39 no 4-5 pp 280ndash285 2006
[2] N Seedher and S Bhatia ldquoReversible binding of celecoxiband valdecoxib with human serum albumin using fluorescencespectroscopic techniquerdquo Pharmacological Research vol 54 no2 pp 77ndash84 2006
[3] H Isogai and N Hirayama ldquoIn silico prediction of interactionsbetween site II on human serum albumin and profen drugsrdquoISRN Pharma vol 2013 Article ID 818364 8 pages 2013
[4] YWangHYuX Shi Z LuoD Lin andMHuang ldquoStructuralmechanism of ring opening reaction of glucose by humanserum albuminrdquo The Journal of Biological Chemistry vol 288pp 15980ndash15987 2013
[5] F Keshavarz M M Alavianmehr and R Yousefi ldquoMolecularinteraction of benzalkonium Ibuprofenate and its discreteingredients with human serum albuminrdquo Physical ChemistryResearch vol 1 no 2 pp 111ndash116 2013
[6] T K Maiti K S Ghosh J Debnath and S Dasgupta ldquoBindingof all-trans retinoic acid to human serum albumin fluores-cence FT-IR and circular dichroism studiesrdquo InternationalJournal of Biological Macromolecules vol 38 no 3ndash5 pp 197ndash202 2006
[7] M-X XieM Long Y Liu C Qin and Y-DWang ldquoCharacter-ization of the interaction between human serum albumin andmorinrdquo Biochimica et Biophysica Acta vol 1760 no 8 pp 1184ndash1191 2006
[8] CD Kanakis P A TarantilisMG Polissiou S Diamantoglouand H A Tajmir-Riahi ldquoAntioxidant flavonoids bind humanserum albuminrdquo Journal of Molecular Structure vol 798 no 1-3 pp 69ndash74 2006
[9] S Marumoto and M Miyazawa ldquoStructure-activity rela-tionships for naturally occurring coumarins as 120573-secretaseinhibitorrdquo Bioorganic and Medicinal Chemistry vol 20 no 2pp 784ndash788 2012
[10] S Marumoto and M Miyazawa ldquoBiotransformation of isoim-peratorin and imperatorin by Glomerella cingulata and 120573-secretase inhibitory activityrdquo Bioorganic and Medicinal Chem-istry vol 18 no 1 pp 455ndash459 2010
[11] S E Sajjadi Y Shokoohinia and S Hemmati ldquoIsolation andidentification of furanocoumarins and a phenylpropanoid fromthe acetone extract and identification of volatile constituentsfrom the essential oil of Peucedanum pastinacifoliumrdquo Chem-istry of Natural Compounds vol 48 no 4 pp 668ndash671 2012
[12] X B Yang J Hou D Liu et al ldquoBiotransformation of isoim-peratorin by Cunninghamella blakesleana AS 3970rdquo Journal ofMolecular Catalysis B vol 88 pp 1ndash6 2013
[13] W Lili S Yehong S Qi et al ldquoIn vitro permeability analy-sis pharmacokinetic and brain distribution study in mice ofimperatorin isoimperatorin and cnidilin in Radix AngelicaeDahuricaerdquo Fitoterapia vol 85 pp 144ndash153 2013
[14] S Wang Q Chen and L He ldquoDevelopment and validationof a gas chromatography-mass spectrometry method for thedetermination of isoimperatorin in rat plasma and tissueapplication to the pharmacokinetic and tissue distributionstudyrdquo Journal of Chromatography B vol 852 no 1-2 pp 473ndash478 2007
[15] H Sadraei Y Shokoohinia S E Sajjadi and M MozafarildquoAntispasmodic effects of Prangos ferulacea acetone extract andits maincomponent osthole on ileum contractionrdquo Research inPharmaceutical Sciences vol 8 pp 137ndash144 2013
[16] H Sadraei Y Shokoohinia S E Sajjadi and B GhadirianldquoAntispasmodic effect of osthole and Prangos ferulacea extracton rat uterus smooth muscle motilityrdquo Research in Pharmaceu-tical Sciences vol 7 pp 141ndash149 2012
[17] S Sajjadi Y Shokoohinia S Gholamzadeh M Behbahani andA Fattahi ldquoAntiviral evaluation of coumarins from Prangosferulacea L (Lindl)rdquo Research in Pharmaceutical Sciences vol7 p 783 2012
[18] S E Sajjadi Y Shokoohinia and S Gholamzadeh ldquoChemicalcomposition of the essential oil of the root of Prangos ferulacea(L) Lindlrdquo Chemija vol 22 pp 178ndash180 2011
[19] N Bijari Y Shokoohinia M-R Ashrafi-Kooshk et al ldquoSpec-troscopic study of interaction between osthole and humanserum albumin identification of possible binding site of thecompoundrdquo Journal of Luminescence vol 143 pp 328ndash336 2013
[20] X Shi M Liu M Zhang et al ldquoIdentification of in vitro and invivo metabolites of isoimperatorin using liquid chromatogra-phymass spectrometryrdquo Food Chemistry vol 141 pp 357ndash3652013
The Scientific World Journal 13
[21] T Chatterjee A Pal S Dey B-K Chatterjee and PChakrabarti ldquoInteraction of virstatin with human serum albu-min spectroscopic analysis andmolecular modelingrdquo Plos Onevol 7 pp 1ndash12 2012
[22] Y-M Chen and L-H Guo ldquoCombined fluorescence and elec-trochemical investigation on the binding interaction betweenorganic acid and human serum albuminrdquo Journal of Environ-mental Sciences vol 21 no 3 pp 373ndash379 2009
[23] J R Lakowicz Principles of Fluorescence Spectroscopy KluwerAcademicPlenum Publishers New York NY USA 2nd edi-tion 1999
[24] F Ding N Li B Han F Liu L Zhang and Y Sun ldquoThe bindingof CI Acid Red 2 to human serum albumin determinationof binding mechanism and binding site using fluorescencespectroscopyrdquo Dyes and Pigments vol 83 no 2 pp 249ndash2572009
[25] M-R Ashrafi Kooshk K Mansouri M Nadi S Khodarahmiand R Khodarahmi ldquoIn vitro anti-cancer activity of nativecurcumin and protein-curcuminrdquo Systems vol 2 no 1 pp 80ndash88 1981
[26] P D Ross and S Subramanian ldquoThermodynamics of proteinassociation reactions forces contributing to stabilityrdquo Biochem-istry vol 20 no 11 pp 3096ndash3102 1981
[27] J Liu J-N Tian J Zhang Z Hu and X Chen ldquoInteractionof magnolol with bovine serum albumin a fluorescence-quenching studyrdquo Analytical and Bioanalytical Chemistry vol376 no 6 pp 864ndash867 2003
[28] A Coi A M Bianucci F Bonomi P Rasmussen G M Muraand M L Ganadu ldquoStructural perturbation of 120572B-crystallinby zinc and temperature related to its chaperone-like activityrdquoInternational Journal of Biological Macromolecules vol 42 no3 pp 229ndash234 2008
[29] M Moller and A Denicola ldquoStudy of protein-ligand bindingby fluorescencerdquoBiochemistry andMolecular Biology Educationvol 30 no 5 pp 309ndash312 2002
[30] R Khodarahmi S A Karimi M R Ashrafi Kooshk S AGhadami S Ghobadi and M Amani ldquoComparative spec-troscopic studies on drug binding characteristics and proteinsurface hydrophobicity of native and modified forms of bovineserum albumin possible relevance to change in protein struc-turefunction upon non-enzymatic glycationrdquo SpectrochimicaActa A vol 89 pp 177ndash186 2012
[31] S-F Ma M Anraku Y Iwao et al ldquoHydrolysis of angiotensinII receptor blocker prodrug olmesartan medoxomil by humanserum albumin and identification of its catalytic active sitesrdquoDrug Metabolism and Disposition vol 33 no 12 pp 1911ndash19192005
[32] H M Berman J Westbrook Z Feng et al ldquoThe protein databankrdquo Nucleic Acids Research vol 28 no 1 pp 235ndash242 2000
[33] S Sugio A Kashima SMochizukiMNoda andKKobayashildquoCrystal structure of human serum albumin at 25 A resolutionrdquoProtein Engineering vol 12 no 6 pp 439ndash446 1999
[34] G M Morris D S Goodsell R Huey and A J OlsonldquoDistributed automated docking of flexible ligands to proteinsparallel applications of AutoDock 24rdquo Journal of Computer-Aided Molecular Design vol 10 no 4 pp 293ndash304 1996
[35] F Faridbod M R Ganjali B Larijani et al ldquoInteraction studyof pioglitazone with albumin by fluorescence spectroscopy andmolecular dockingrdquo Spectrochimica Acta A vol 78 no 1 pp96ndash101 2011
[36] D SGoodsell andA JOlson ldquoAutomated docking of substratesto proteins by simulated annealingrdquo Proteins vol 8 no 3 pp195ndash202 1990
[37] D S Goodsell G M Morris and A J Olson ldquoAutomateddocking of flexible ligands applications of AutoDockrdquo Journalof Molecular Recognition vol 9 no 1 pp 1ndash5 1996
[38] G Sudlow D J Birkett and D N Wade ldquoThe characterizationof two specific drug binding sites on human serum albuminrdquoMolecular Pharmacology vol 11 no 6 pp 824ndash832 1975
[39] Y Yue X Chen J Qin and X Yao ldquoCharacterization of themangiferin-human serum albumin complex by spectroscopicandmolecularmodeling approachesrdquo Journal of Pharmaceuticaland Biomedical Analysis vol 49 no 3 pp 753ndash759 2009
[40] A K Shaw and S K Pal ldquoSpectroscopic studies on the effectof temperature on pH-induced folded states of human serumalbuminrdquo Journal of Photochemistry and Photobiology B vol 90no 1 pp 69ndash77 2008
[41] S Ranjbar S Ghobadi R Khodarahmi and H Nemati ldquoSpec-troscopic characterization of furosemide binding to humancarbonic anhydrase IIrdquo International Journal of BiologicalMacromolecules vol 50 no 4 pp 910ndash917 2012
[42] HNHou Z POi YWOYung F L Liao Y Zhang andY LiyldquoStudies on interaction between vitamin B12 and human serumalbuminrdquo Journal of Pharmaceutical and Biomedical Analysisvol 47 pp 134ndash139 2008
[43] S S Kalanur J Seetharamappa and V K A Kalalbandi ldquoChar-acterization of interaction and the effect of carbamazepine onthe structure of human serum albuminrdquo Journal of Pharmaceu-tical and Biomedical Analysis vol 53 no 3 pp 660ndash666 2010
[44] L Stryer ldquoFluorescence spectroscopy of proteinsrdquo Science vol162 no 3853 pp 526ndash533 1968
[45] B Valeur and J C Brochon New Trends in FluorescenceSpectroscopy Springer Berlin Germany 1999
[46] V Suryawanshi P Anbhule A Gore Sh Patil and G AndKolekar ldquoA spectral deciphering the perturbation of modeltransporter protein (HSA) by antibacterial pyrimidine deriva-tive pharmacokinetic and biophysical insightsrdquo Journal ofPhotochemistry and Photobiology B vol 118 pp 1ndash8 2013
[47] T Yuan AMWeljie andH J Vogel ldquoTryptophan fluorescencequenching by methionine and selenomethionine residues ofcalmodulin orientation of peptide and protein bindingrdquo Bio-chemistry vol 37 no 9 pp 3187ndash3195 1998
[48] TCOrsquoHaver A F Fell G Smith et al ldquoDerivative spectroscopyand its applications in analysisrdquo Analytical Proceedings vol 19no 1 pp 22ndash46 1982
[49] Y-Q Wang B-P Tang H-M Zhang Q-H Zhou and G-C Zhang ldquoStudies on the interaction between imidaclopridand human serum albumin spectroscopic approachrdquo Journal ofPhotochemistry and Photobiology B vol 94 no 3 pp 183ndash1902009
[50] S-J Kim H-W Rhee H-Y Park H-S Kim and J-I HongldquoFluorescent probes designed for detecting human serum albu-min on the basis of its pseudo-esterase activityrdquo Bioorganic ampMedicinal Chemistry Letters vol 23 pp 2093ndash2097 2013
[51] P Ascenzi M Gioia G Fanali M Coletta and M FasanoldquoPseudo-enzymatic hydrolysis of 4-nitrophenyl acetate byhuman serum albumin pH-dependence of rates of individualstepsrdquo Biochemical and Biophysical Research Communicationsvol 424 pp 451ndash455 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 The Scientific World Journal
of the acceptor at 120582 So 119869 can be evaluated by integratingthe overlap between emission spectrum of the donor andabsorption spectrum of the acceptor [27]
The absorption spectrum of isoimperatorin (10 120583M) wasrecorded in the range of 222ndash450 nmThe emission spectrumof HSA (10 120583M) was also recorded in the range of 222ndash450 nm with excitation and emission slit widths of 5 and5 nm respectivelyThe overlap of the absorption spectrum ofisoimperatorin with the fluorescence emission spectrum ofHSA was used to calculate the efficiency of energy transfer
26 Protein Surface Hydrophobicity (PSH) DeterminationThe ANS binding properties of HSA can be investigatedby measuring 119870
app119889
and 119865max of the HSA-ANS complexin the absence and presence of isoimperatorin 119870app
119889
is theapparent dissociation constant of the fluorescent ANS-HSAcomplex and 1119870
app119889
is the binding affinity of ANS tothe protein in the absence and presence of isoimperatorin[28] 119865max is the maximum fluorescence intensity at thesaturated ANS concentration which indicates the numberof surface hydrophobic sites of the protein This methodinvolves titration of the enzyme solutions in the presenceof increasing concentrations of ANS The values of 1119870app
119889
and 119865max can be obtained from the slope and the 119909-interceptof the Scatchard plot respectively In this type of Scatchardplot the fluorescence intensity[ANS]free is drawn versusfluorescence intensity The free ANS concentration must bedetermined via plotting fluorescence intensity versus totalANS concentration In dilute solutions of ANS a linearrelationship exists between fluorescence intensity (119865) andANS concentration (119888) (119865 = 119861119888 where 119861 is the propor-tionality coefficient between fluorescence intensity and ANSconcentration) From a 1mM stock solution ANS was addedto the final concentrations ranging from 0 to 50 120583M Theenzyme concentration was 1 120583M It is assumed that in verydilute ANS solutions (0-1 120583M) all of the ANS molecules arebound to theHSA and thus B (the proportionality coefficientbetween fluorescence intensity and ANS concentration) canbe determined from the slope of the linear portion offluorescence intensity plot versus ANS concentration Foreach of the different total concentrations of ANS used theconcentration of bound ANS molecules was calculated usingthe following equation [ANS]bound = 119865119861 and the free ANSmolecules concentration was calculated from the differencebetween total and bound ANS concentrations ([ANS]free =[ANS]total minus [ANS]bound) [29 30]
The protein surface hydrophobicity index in the absenceand presence of the drug can be calculated by applying thefollowing equation [28]
PSH =
119865max
[HSA] 119870app119889
(9)
Following excitation at 380 nm the fluorescence emissionincrement was recorded at 470 nm until no further increasein fluorescence intensity was observed
27 Competitive Experiments In order to elucidate the bind-ing site of isoimperatorin onHSA we carried out competitive
experiments at the presence of two known drugs (Warfarinas site I marker and Ibuprofen as a drug that preferentiallybinds to site II) [24 31] The concentration of HSA and war-farinibuprofen was stabilized at 10times10minus6M Isoimperatorinwas then gradually added to theHSAHSA-warfarin orHSA-ibuprofen mixtures An excitation wavelength of 295 nm wasselected and flourescence spectra were recorded in the rangeof 300ndash450 nm
Also possible inhibitory effect of isoimperatorin indo-methacin tetracycline ibuprofen aspirin osthole and war-farin on the esterase activity of HSA was evaluated
28 Docking Procedure The AutoDock 405 docking pro-gram was applied to study the interaction between theisoimperatorin and HSA The crystal structure of HSA wasobtained from the Protein Data Bank Of more than 50X-ray crystallographic structures related to human serumalbumin in ProteinData Bank [32] entry with PDB ID1BM0[33] was chosen for dockings because of no missing atomsno cocrystallized ligand and having a reasonably goodresolution (25 A) All the water molecules were removedusing a plain text editor The polar hydrogen partial chargeand solvation parameters for use with the AutoDock 405simulation were applied using AutoDock-Tools The dockingsimulations were carried out with the rigid HSA and aflexible ligand [34] Using AutoGrid tools the grid mapswere generated adequately large to include the binding site(s)of protein as well as significant regions of the surroundingsurfaceThe points of the grids were thus 126times126times126witha grid spacing of 0375 A [35] Afterwards the most suitablestructure for the flexible ligand molecule was optimized bythe rotation of all single bonds in the ligand molecule Thegrid parameter file and the docking parameter file were setup by the AutoDockTools program [36] The binding modeof isoimperatorin was predicted by using the Lamarckiangenetic algorithm search engine implemented in AutoDocksoftware package [37] for the two main drug binding sitesI and II (subdomains IIA and IIIA of HSA resp) [38]Population size was 256 and Cluster analysis was performedon the docked results using a root mean square (RMS)tolerance of 05 A Default settings were used for all otherparameters [35] At the end of the docking simulation theHSA-IIM complex with the most stable energy was adoptedas the most favorable structure of the complex for HSAMolecular graphics were prepared with PyMOL version 099beta06 (httpwwwpymolorg)
3 Results and Discussion
31 UV Absorption Spectra UV absorption spectroscopytechnique can be used to explore the structural changes ofprotein and to investigate protein-ligand complex formationHSA has two main absorption bands that one of them islocated in the range of 260ndash300 nm which is the absorptionband of the aromatic amino acids (Trp Tyr and Phe) [27 39]Figure 2 shows the UV absorption spectra of HSA in theabsence and presence of isoimperatorin As can be seen inFigure 2 HSA has strong absorbance with a peak at 280 nmand the absorbance of HSA increased with the addition of
The Scientific World Journal 5
0
05
1
15
2
25
3
35
4
200 250 300 350Wavelength (nm)
Abso
rban
ce
034
035
036
037
038
039
04
270 274 278 282 286 290
1
7
Figure 2 Absorption spectra of HSA (30 120583M) with variousamounts of isoimperatorin (1ndash7) 0 04 08 2 4 6 and 8 120583MThe absorption of each sample was measured with respect to therespective blank
0
50
100
150
200
250
300
350
400
450
300 350 400 450 500 550Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
1
16
116
Figure 3 Effect of isoimperatorin on fluorescence spectra of HSA(119879 = 298K pH 75) Curve (1) 30 120583MHSA Curves (2ndash16) 30 120583MHSA in the presence of 02 04 06 08 1 15 2 25 3 4 5 6 7 8and 10120583M of isoimperatorin
isoimperatorin In addition the absorbance value of 280 nmof the isoimperatorin at a concentration of 10120583M is very low(lt003) Our results confirmed that the interaction betweenHSA and ligand took place
32 Fluorescence Measurements
321 Intrinsic Fluorescence Measurements The fluorescencespectra of HSA in the absence and presence of differentamounts of Isoimperatorin were recorded in the rangeof 300ndash570 nm upon excitation at 295 nm isoimperatorincauses a concentration dependent quenching of the intrinsicfluorescence of HSA (Figure 3) without changing the emis-sion maximum and shape of the peaks These results indi-cated that there were interactions between isoimperatorinand HSA This interaction was further confirmed by UV-visabsorption technique
0
1
2
3
4
0 2 4 6 8 10 12[Isoimperatorin] (120583M)
F0F
Figure 4 The Stern-Volmer plots for HSA fluorescence quenchingby isoimperatorin at 298 (⧫) 303 (998771) 308 (◼) and 313 (∙) K Thefinal protein concentration in 50mM phosphate pH 75 was 3 120583MBased on ldquoOne-Sample Kolmogorov-Smirnov Testrdquo test distributionwas normal
A gradual decrease in the fluorescence emission intensityof HSA is observed in the presence of increasing concentra-tions of isoimperatorin suggesting the presence of tryptophanresidue of the HSA at or near the isoimperatorin binding siteHSA has one Trp (Trp 214) and its fluorescence is solely dueto the excitation of this residue of HSA located in the bindingdomain-IIA (site I) at the bottom of a 12 A deep crevice [40]
Emerging of a new emission peak at 505 nm uponisoimperatorin addition can be attributed to the bounddrug and radiationless energy transfer between the HSAtryptophan and isoimperatorin [41] The appearance of anisoactinic point at 430 nm might also indicate the existenceof equilibrium between bound and free drug In additionsuch equilibrium may emphasize the formation of the drug-protein complex [42]
322 Fluorescence Quenching Mechanism Quenching canoccur by different mechanisms which is usually classifiedas dynamic quenching and static quenching In dynamicquenching increasing the temperature results in faster dif-fusion and hence larger amounts of collision which causesraising of the quenching constant Opposing to dynamicquenching in static one increasing the temperature weakensthe stability of the formed complex and hence reduces thequenching constant [19]
Fluorescence quenching of HSA upon interaction withisoimperatorin was evaluated at different temperatures (298303 308 and 313 K) and the results are shown in Figure 4As indicated in this figure the Stern-Volmer plots are lin-ear with the slopes increasing with increasing temperature(the linearity in this plot confirmed one-to-one interactionbetween isoimperatorin and HSA) The calculated values of119870SV at different temperatures are listed in Table 1 It can
6 The Scientific World Journal
Table 1 Stern-Volmer quenching constant (119870SV) quenching rate constant (119896119902) association constant (119870119887
) and number of binding sites (119899)for binding of isoimperatorin to HSA in the different temperatures
119879 (K) 10
minus4
times 119870SV(Mminus1)
10
minus12
times 119870
119902
(Mminus1sdotsminus1)119877
a 10
minus5
times 119870
119887
(Mminus1)Number of biding
sites (119899) 119877
a
2981698 plusmn 013 1698 plusmn 013
099621102 plusmn 010 0862 09841
3032019 plusmn 017 2019 plusmn 017
099551149 plusmn 013 0851 09886
3082333 plusmn 016 2333 plusmn 016
099651271 plusmn 022 0844 09950
3132639 plusmn 019 2639 plusmn 019
099811322 plusmn 017 0822 09917
Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870SV values
be found from Table 1 that the value of 119870SV increased withincreasing temperature in the presence of isoimperatorinwhich indicates the involved binding forces are mainlyhydrophobic interactions (endothermic apolar interactionsare strengthened with increasing temperature) Also the rateconstant of the quenching process (119896
119902) which is equal to119870SV
divided by 1205910(with assuming 120591
0= 10
minus8 s)
119896
119902=
119870SV120591
0
(10)
was calculated (Table 1) The results revealed that the valuesof 119896119902were much greater than the maximum diffusion rate
constant of the biomolecule (2 times 1010Mminus1 sminus1) which indi-cated that the fluorescence quenching of HSA was initiatedby complex formation between HSA and isoimperatorinconfirming that the mechanism of quenching is static [19]
323 Determination of Association Constant (119870119887) and Num-
ber of Binding Sites (119899) The values of association constant119870
119887 and the number of binding sites 119899 at different temper-
atures are obtained from the modified Stern-Volmer plots(Figure 5) and listed in Table 1 It can be found from thesedata that the 119899 values at different temperatures are keptaround unity which shows the existence of one binding sitein HSA for isoimperatorin The value of 119870
119887is significant to
understand the distribution of the drug in plasma since theweak binding can lead to a short life time or poor distributionwhile strong binding can decrease the concentration of freedrug in plasma The value of 119870
119887illustrated that there is a
strong binding force between isoimperatorin and HSA Itwas found that the binding constant increased with increasein temperature resulting in an increment of the stabilityof the isoimperatorin-HSA complex it also implies thatisoimperatorin can be tightly stored and carried by HSA inthe body
324 Forces Involved in the Binding Process Consideringthe dependence of binding constant on temperature a ther-modynamic process was considered to be responsible forthe formation of a complex Therefore the thermodynamicparameters dependent on temperatures were analyzed Theacting forces between a small molecule and macromoleculemainly include hydrogen bonds van der Waals forces elec-trostatic forces and hydrophobic interaction forces [23] Inorder to obtain the information about the forces persisting in
minus15
minus13
minus11
minus09
minus07
minus05
minus03
minus01
01
03
05
minus68 minus65 minus62 minus59 minus56 minus53 minus5
log[isoimperatorin]
log[(F0minusF)F]
Figure 5 The modified Stern-Volmer plots for HSA fluorescencequenching by isoimperatorin at 298 (⧫) 303 (998771) 308 (◼) and 313 (∙)K For interpretation of references to color in this figure legend thereader is referred to the web version of this article
the present binding process the thermodynamic parametersof binding were obtained fromVanrsquot Hoff plot followed by theGibbs equation (data not shown) Table 2 shows the values ofΔ119867
∘ andΔ119878∘ obtained for the binding site from the slopes andordinates at the origin of the fitted lines From Table 2 it canbe seen that the negative sign for free energy (Δ119866∘) meansthat the interaction process is spontaneous The positiveenthalpy (Δ119867∘) and entropy (Δ119878∘) values of the interaction ofisoimperatorin and HSA indicate that the binding is mainlyentropy-driven and the enthalpy is unfavorable for it thehydrophobic forces play a major role in the reaction [43]
33 Nonradiative Energy Transfer from HSA to Isoimpera-torin The Forster nonradiative energy transfer (FRET) hasbeen used as a ldquospectroscopic rulerrdquo for measurement ofthe distance between the acceptor (ligand) and the donor(tryptophan residues in the protein) [44] According to thistheory there will be FRET between two molecules if thefollowing conditions were satisfied (a) the donor moleculeemits fluorescence (b) the emission spectrum of the donorhas an enough overlap with the absorption spectrum of
The Scientific World Journal 7
Table 2 Thermodynamic parameters for binding isoimperatorin to HSA
119879 (K) Δ119878
∘ (Jsdotmolminus1sdotKminus1) Δ119867
∘ (kJsdotmolminus1) Δ119866
∘ (kJsdotmolminus1)298 minus2969 plusmn 009
303 17820 plusmn 011 2993 plusmn 013 minus3071 plusmn 007
308 minus3169 plusmn 010
313 minus3299 plusmn 008
0
50
100
150
200
250
300
350
400
450
222 272 322 372 422Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
0
002
004
006
008
01
012
014
Abso
rban
ce
a
b
Figure 6 The overlap of the fluorescence spectrum of HSA (a)and the absorption spectrum of isoimperatorin (b) when [HSA] =[isoimperatorin] = 10 120583MThe enzyme was excited at 295 nm
the acceptor and (c) the donor and acceptor molecules areclose enough with the maximum distance not to exceeding7 nm [45]
There is a good overlapping between the fluorescenceemission spectrum of HSA and absorption spectra of isoim-peratorin (Figure 6) As the fluorescence emission of proteinwas affected by the excitation light around 295 nm thespectrum ranging from 300ndash400 nm was chosen to calculatethe overlapping integral The value of 119869 was calculated to be473 plusmn 005 times 10
minus15 cm3 Lmolminus1 by integrating the overlapspectra for 300ndash400 nm In the present case 1198702 = 23119873 = 136 and Φ = 0118 [21] according to (6) and (7) wecould calculate that 119864 = 045 plusmn 006 119877
0= 322 plusmn 03 nm and
119903 = 29 plusmn 02 nm As mentioned earlier (Section 321) HSAposses one tryptophan residue (Trp 214) and the calculated119903 is actually the average value between the bound drug andTrp 214 Since 119903 is much smaller than 7 nm and 05119877
0lt
119903 lt 15119877
0[41] therefore the energy transfer from HSA
to isoimperatorin occurs with high probability which alsoexplains the efficient quenching of the HSA fluorescence
34 PSH Determination in the Absence and Presence of Isoim-peratorin Several methods exist for measuring the surfacehydrophobicity of proteins that ANS experiment is one ofthem ANS as an extrinsic fluorescent probe is extremelysensitive to polarity of the solvent In aqueous solutions itfluoresces very weakly but upon binding to hydrophobicpatches of proteins its quantum yield increases significantly[29] Thus ANS fluorescence may be applied for monitoringpossible changes in protein surface hydrophobicity induced
0
200
400
600
800
1000
0 10 20 30 40 50
Fluo
resc
ence
inte
nsity
(au
)
[ANS] (120583M)
Figure 7 Binding of ANS to HSA (1120583M) in the absence (∙) andpresence (998771) of 3120583M of isoimperatorin
upon drug binding At a fixed concentration (1 120583M) of HSAand increasing concentrations of ANS (0ndash50120583M) fluores-cence intensity of ANS was measured in the absence andpresence of 3120583M isoimperatorin (Figure 7)
A typical hyperbolic response is observed which demon-strates the saturation nature of ANS binding toHSA It can beseen that the hyperbolic response of ANS fluorescence in thepresence of isoimperatorin is slightly different in comparisonto the absence of the drug The values of 119865max and 119870
app119889
forANS are obtained from the Scatchard plot (Figure 8) andlisted in Table 3 The value of 119870app
119889
for ANS binding in theabsence of drug (09 plusmn 005) was increased in the presenceof 3 120583M isoimperatorin (14 plusmn 007) which shows the weakerbinding of ANS to the HSA-isoimperatorin complex ThenPSH was calculated in the absence and the presence of3 120583M isoimperatorin by using 119865max and119870
app119889
values (Table 3)Decreasing the PSH index of HSA (from 150794 plusmn 1146 to80612 plusmn 007) suggests that the surface hydrophobicity ofHSA is decreased by sim465 upon drug binding
35 Synchronous Fluorescence Spectroscopy Synchronous flu-orescence spectroscopy introduced by Lody has been usedto characterize complex mixtures which can provide finger-prints of complex samples [46] The synchronous fluores-cence spectra can provide information about the molecularenvironment in the vicinity of the chromophore moleculesIn the synchronous fluorescence spectra the sensitivityassociated with fluorescence is maintained while several
8 The Scientific World Journal
Table 3 Surface hydrophobicity parameters for HSA in the absence and the presence of isoimperatorin
Parameters HSA HSA-IIM complex119870
app119889
(120583M) 09 plusmn 005
lowast
14 plusmn 007
lowast
119865max[HSA] 135714 plusmn 85
lowastlowast
112853 plusmn 63
lowastlowast
Surface hydrophobicity index (119865max[HSA] 119870app119889
) 150794 plusmn 1146
lowastlowast
80612 plusmn 007
lowastlowast
Data are expressed as mean plusmn SD of three measurements lowast119875 value lt 00006 lowastlowast119875 value lt 00001
0
50
100
150
200
250
300
350
400
400 500 600 700 800 900 1000Fluorescence intensity (au)
Fmax
Fluo
resc
ence
inte
nsity
(au
)[A
NS]
free
Slope = minus1Kappd
Figure 8 Scatchard plots for titration of HSA with increasingconcentrations of ANS in the absence (∙) and presence (998771) of 3 120583Mof isoimperatorin 119865max and 1119870
app119889
values were obtained from the119909-intercept and slope of this plot For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
advantages are available spectral simplification spectralbandwidth reduction and avoidance of different perturbingeffects The fluorescence spectrum of BSAHSA mainly dueto the Trp residues is sensitive to the microenvironment ofthese chromophoreswith themaximumemissionwavelengthbeing very useful in estimating the hydrophobicity aroundthe tryptophan residues It is well known that the experimenton synchronous fluorescence of HSA will provide the char-acteristic information for the Tyr and Trp residues when thescanning interval Δ120582 is fixed at 15 and 60 nm respectively[47] The shift in the position of fluorescence emissionmaximum corresponds to changes of the polarity aroundthe chromophore molecule A blue shift of 120582max means thatthe amino acid residues are located in a more hydrophobicenvironment and are less exposed to the solvent while a redshift of 120582max implies that the amino acid residues are in apolar environment and are more exposed to the solvent [48]The synchronous fluorescence spectra of interaction betweenisoimperatorin and HSA are presented in Figure 8 It wasshown that the fluorescence intensity of HSA decreased regu-larly along with the addition of isoimperatorin which furtherdemonstrated the occurrence of fluorescence quenching inthe binding process Moreover there is no significant shiftof the maximum emission wavelength with Δ120582 = 15 nm or60 nm (Figures 9(a) and 9(b)) which implies that interaction
of isoimperatorin with HSA does not affect the conformationof the region around the Tyr or Trp residues
As well we can obtain the information about the locationof isoimperatorin binding site from the synchronous fluores-cence dataThe ratios of synchronous fluorescence quenching(RSFQ) were used to determine the binding sites of the drugsto the HSA molecule in drug-HSA complexes The RSFQvalues were calculated from the following equation
RSFQ = 1 minus
119865
119865
0
(11)
where 119865 and 1198650are the synchronous fluorescence intensities
of HSA in the presence and the absence of drug respectivelyIt was obvious from Figure 9(c) that the RSFQ results forΔ120582 = 60 nmwere slightly higher than the corresponding onesfor Δ120582 = 15 nm As illustrated in Figure 9(c) addition ofisoimperatorin resulted in strong fluorescence quenching ofTrp residue (sim47 reduction in fluorescence intensity) con-firming that isoimperatorin reached subdomain IIA whereonly one Trp residue (Trp 214) in HSA is located It wasalso observed from Figure 9(c) that the fluorescence strengthof Tyr residues decreased about sim32 in the presence ofisoimperatorin So the binding of isoimperatorin to HSAquenched flourescence intensity of Tyr and Trp Since Trp214 (the only Trp of HSA) is at site I and Tyr 263 (hasthe strong fluorescence in the case of Tyr residues andmay be one of the residues responsible for the synchronousfluorescence spectrum of HSA when Δ120582 is fixed at 15 nm) isin subdomain IIA (site I) it can be concluded that Trp andTyr residues of HSA may have equal accessibility to bindingsite of isoimperatorin [49] The above results indicated thatisoimperatorin can bind to HSA in the hydrophobic cavity(site I) on subdomain IIA which are in full agreement withquenching competitive binding experiments and theoreticaldocking technique
36 Site Marker Competitive Binding Experiments In orderto further classify drug binding site on HSA competitivebinding experiments have been carried out Warfarin andibuprofen are two markers specific for binding to HSA thatbind to sites I and II which are corresponded to the sub-domains IIA and IIIA respectively Therefore to determinethe location of the isoimperatorin binding site on HSA thecompetitive displacement experimentswere carried out usingwarfarin as a characteristicmarker for site I (Figure 10(a)) andibuprofen for site II (Figure 10(b)) [24 31] In the site markercompetitive experiment isoimperatorin was gradually addedto the solution of HSA HSA-ibuprofen or HSA-warfarincomplex and then fluorescence intensity of system wasrecorded
The Scientific World Journal 9
0
100
200
300
400
500
600
265 270 275 280 285 290 295 300Wavelength (nm)
1
8
Fluo
resc
ence
inte
nsity
(au
)
(a)
Fluo
resc
ence
inte
nsity
(au
)
0
50
100
150
200
250
300
350
400
250 260 270 280 290 300Wavelength (nm)
1
8
(b)
0
10
20
30
40
50
0 05 1 15 2 25 3
RSFQ
()
Isoimperatorin (120583M)
0
02
04
06
08
1
12
005 1 15 2 25 3 35
Isoimperatorin (120583M)
F0
F
(c)
Figure 9 Synchronous fluorescence spectra of interaction between HSA and isoimperatorin (a) at Δ120582 = 15 nm and (b) at Δ120582 = 60 nmConcentration ofHSAwas 30 120583Mwhile concentrations of isoimperatorinwere 0 04 08 12 16 2 25 and 3 120583Mfrom 1 to 8 (c) Comparativeevaluation of isoimperatorin effect on the RSFQ of HSA [HSA] = 3 120583M (◼) Δ120582 = 15 nm and (∙) Δ120582 = 60 nm
As shown in Figure 10(a) with addition of warfarin in theHSA solution the flourescence intensity was slightly higherthan that of without warfarin (and including the red shift)Then after adding the isoimperatorin into the above systemthe flourescence intensity of HSA solution with warfarinheld in equimolar decreased gradually and the intensitywas much lower than that of without warfarin displayingthat the binding of the isoimperatorin to HSA was affectedafter adding warfarin On the contrary in the presence ofibuprofen the flourescence intensity of the HSA-ibuprofencomplex almost had no difference from that recordedwithoutibuprofen under the same conditions (Figure 10(b)) whichindicated that site II marker did not prevent the binding ofisoimperatorin in its usual binding location The next stepprotein intrinsic fluorescence was measured and the relativefluorescence intensities (119865119865
0) versus ligand concentration
([isoimperatorin]) plots are displayed in Figure 10(c) Asshown in Table 4 the binding constant was remarkablydecreased after addition of warfarin while the addition ofibuprofen results in only a small difference These resultsindicate that warfarin can displace the isoimperatorin but
ibuprofen has little effect on the binding of isoimperatorinto HSA In other words these results suggest that isoim-peratorin probably competes with warfarin for binding toHSA [24] Experimental observations were followed up withdocking studies where isoimperatorin was docked to HSAto determine the preferred binding site on the proteinAs described above the 3D crystal structure of HSA is amonomer consisting of three homologous domains whichassemble to form a heart-shaped molecule [4 6] Each ofthe structurally similar 120572-helix domains (IndashIII) has twosubdomains (A andB) with six120572-helices in subdomainA andfour 120572-helices in subdomain BThe crystal structure analysesindicate that the principal regions of ligand binding sitesin HSA are located in hydrophobic cavities in subdomainsIIA and IIIA which are consistent with Sudlowrsquos site I andSudlowrsquos site II respectively Several studies have shown thatHSA is able to bind many ligands in several binding sites[38] In this paper the molecular docking model of theHSAndashisoimperatorin complex was conducted on the basisof the crystal structure of HSA to find the binding positionof isoimperatorin The best docking energy result is shown
10 The Scientific World Journal
0
100
200
300
400
500
600
310 335 360 385 410Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA1
8
(a)
0
100
200
300
400
500
600
700
305 325 345 365 385 405Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA
1
8
(b)
0
05
1
15
0 1 2 3
HSAIbuprofen
Warfarin
Isoimperatorin (120583M)
F0
F
(c)
Figure 10 Effect of isoimperatorin on the quenching of HSA-warfarin complex (a) andHSA-ibuprofen complex (b) fluorescence From 1 to 8[HSA] = [warfarin or iboprufen] = 3 120583M [isoimperatorin] = 0 04 08 12 16 2 25 and 3 120583M All reproduced data shown are representativeof at least 3 independent experiments (c) Fluorescence quenching profiles of HSA (⧫) HSA-ibobrofen (◼) and HSA-warfarin (998771) in thepresence of various concentration of isoimperatorin
in Figure 11 It can be seen that isoimperatorin was situatedwithin subdomain IIA in Sudlowrsquos site I formed by six 120572-helices This figure shows the location of Trp 214 and also thebinding of isoimperatorin in HSA As this fluorescent residueis in subdomain IIA and due to the proximity of the bindingsite of isoimperatorin to the Trp 214 it can be concluded thatisoimperatorin has one reactive site onHSAandprobably thissite is at or near the site I The above docking results are ingood agreement with the displacement experiments results
and revealed that isoimperatorin binds to Sudlowrsquos site I ofHSA
37 Effect of Isoimperatorin on HSA Esterase Activity Pre-liminary studies showed that albumin in addition to itsligand binding capabilities has remarkable promiscuouscatalytic activities toward a broad range of organicmoleculesincluding esters amides phosphates and benzisoxazolesHuman serum albumin had significantly greater esterase-like
The Scientific World Journal 11
Table 4 Binding constants for isoimperatorin-HSA interaction (at 295K) in the absence and presence of the site marker
119870
119887
times 10
minus5
(Mminus1) 119877
a
HSA + isoimperatorin (in the absence ofibuprofen and warfarin) 1110 plusmn 04
lowast 09947
HSA + isoimperatorin + ibuprofen 1099 plusmn 06
lowast 09904HSA + isoimperatorin + warfarin 789 plusmn 03
lowast 09881Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870
119887values lowast119875 value lt 00002
Trp 214
Isoimperatorin
Figure 11 Crystal structure of HSA and the positions of singletryptophan residue (Trp 214) and also the binding site of isoim-peratorin in HSA Protein backbone is shown in the ldquocartoonrdquorepresentation and isoimperatorinand Trp 214 residue in the site Iare shown in the ldquostickrdquo representation The isoimperatorin and Trp214 structures are represented by the red and blue colorsThe crystalstructure of protein was obtained from Protein Data Bank (1BM0)For interpretation of references to color in this figure legend thereader is referred to the web version of this paper
activity than bovine serum albumin and defatted albuminwas more active than untreated albumin [50] InterestinglyHSA exhibits esterase-like activity and hydrolyzes drugshaving an ester group such as aspirin (which can convertaspirin ldquoacetylsalicylic acidrdquo to salicylic acid) Lys199 Lys402Lys519 and Lys545 form the pseudoesterase active site ofHSAthat is located in site I [50] ldquoEsterase-likerdquo activity (hydrolysisof p-nitrophenol esters such as p-NPA) by subdomain III Ahas been also reported and site directed mutagenesis studieshave shown that Arg 410 and Tyr 411 are essential in thisesterase activity However further investigation revealed thatacetylation is not limited to a distinct side chain so that theHSA incubation with high concentrations of pNPAc resultsin the acetylation of 59 Lys 10 Ser 8 Thr 4 Tyr and 1 Asp[51] Because of the above difference here we set out toevaluate the effect of isoimperatorin on the esterase activityof HSA based on our previous work [19] The kinetics ofthe pseudoenzymatic hydrolysis of pNPAc was obtained inthe absence and presence of isoimperatorin and site markers(Figure 12) Accordingly warfarin and indomethacin (as siteI marker) and isoimperatorin inhibited competitively theHSA-catalyzed hydrolysis of pNPAc by binding to site I Incontrast ibuprofen and tetracycline were unable to inhibitesterase activity of HSA The enzyme inhibition results sug-gested that isoimperatorin aspirin warfarin indomethacin
0
005
01
015
02
025
03
035
0 05 1 15 2 25 3Time (min)
HSA TetracyclineIbuprofenAspirin
IndomethacinOstholeWarfarinIsoimperatorin
A400
Figure 12 4-Nitrophenyl acetate esterase activity of HSA in theabsence and presence of tetracycline and ibuprofen (site II) war-farin aspirin indomethacin and osthole (site I) and isoimperatorin(site I) Final concentration of protein pNPA and drugs were 15250 and 250 120583M respectively For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
and osthole binding sites on the HSA molecule are the sameThese results also proposed that the large site I on HSAparticipates effectively to its promiscuous esterase activity
4 Conclusion
The accumulated data on HSA-drug interactions can beused to develop quantitative structure-activity relationshipsfor albumin binding However as stated earlier adsorptiondistribution metabolism and elimination (ADME) is con-trolled by albumin [30] Poor or strong binding to SA isbelieved to affect half-time of drugs [35] and their dosagedepends on their half-life in plasma Thus the bindingaffinity of any drug to SA is one of the major factors thatdetermine the pharmacokinetics halftime and availabilityof the drug in various tissues (see [30] and referencestherein) Further complexities arise in vivo due to interactions
12 The Scientific World Journal
between drugs and endogenous ligands for HSA This isparticularly pertinent for fatty acids which normally occurin serum at levels between 01 and 2mol per mol of HSAand can both compete and cooperate with drugs bindingto the protein In certain disease states these effects areexacerbated as the fatty acid HSA molar ratio may be ashigh as six Other pathological conditions are associatedwith high (micromolar to millimolar) levels of bilirubinhemin or renal toxins (eg 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) indoxyl sulphate) which bindto the protein causing significant drug binding defects
Our major aim in this study was to distinguish thenature of the interaction of isoimperatorin to HSA usingdifferent biophysical techniques This study could be veryvaluable due to importance and presence of isoimperatorinin human diet (food-drug interaction on HSA) The resultsshowed that isoimperatorin quenched the intrinsic fluo-rescence of HSA through dynamic quenching mechanismFurthermore hydrophobic interaction played a major role inthe binding process Using Fluorescence and UV absorptionspectroscopy it has been shown that isoimperatorin inducesprotein structural changes The binding average distancebetween the solo tryptophan of the protein and the boundisoimperatorin was calculated by Forster theory as 290 plusmn02 nmAnalysis of PSHvalues elucidated that isoimperatorinhas induced some compactness in the tertiary structure ofthe enzyme The results of synchronous flourescence onHSA-isoimperatorin system elucidated that the drug binds tothe hydrophobic cavity (site I) on subdomain IIA that wasconfirmed by quenching experiments competitive bindingexperiments and docking technique
Acknowledgment
The authors gratefully acknowledge the Kermanshah Uni-versity of Medical Sciences Research Council for financialsupport of this work
References
[1] Y-J Hu Y Liu T-Q Sun A-M Bai J-Q Lu and Z-B Pi ldquoBinding of anti-inflammatory drug cromolyn sodiumto bovine serum albuminrdquo International Journal of BiologicalMacromolecules vol 39 no 4-5 pp 280ndash285 2006
[2] N Seedher and S Bhatia ldquoReversible binding of celecoxiband valdecoxib with human serum albumin using fluorescencespectroscopic techniquerdquo Pharmacological Research vol 54 no2 pp 77ndash84 2006
[3] H Isogai and N Hirayama ldquoIn silico prediction of interactionsbetween site II on human serum albumin and profen drugsrdquoISRN Pharma vol 2013 Article ID 818364 8 pages 2013
[4] YWangHYuX Shi Z LuoD Lin andMHuang ldquoStructuralmechanism of ring opening reaction of glucose by humanserum albuminrdquo The Journal of Biological Chemistry vol 288pp 15980ndash15987 2013
[5] F Keshavarz M M Alavianmehr and R Yousefi ldquoMolecularinteraction of benzalkonium Ibuprofenate and its discreteingredients with human serum albuminrdquo Physical ChemistryResearch vol 1 no 2 pp 111ndash116 2013
[6] T K Maiti K S Ghosh J Debnath and S Dasgupta ldquoBindingof all-trans retinoic acid to human serum albumin fluores-cence FT-IR and circular dichroism studiesrdquo InternationalJournal of Biological Macromolecules vol 38 no 3ndash5 pp 197ndash202 2006
[7] M-X XieM Long Y Liu C Qin and Y-DWang ldquoCharacter-ization of the interaction between human serum albumin andmorinrdquo Biochimica et Biophysica Acta vol 1760 no 8 pp 1184ndash1191 2006
[8] CD Kanakis P A TarantilisMG Polissiou S Diamantoglouand H A Tajmir-Riahi ldquoAntioxidant flavonoids bind humanserum albuminrdquo Journal of Molecular Structure vol 798 no 1-3 pp 69ndash74 2006
[9] S Marumoto and M Miyazawa ldquoStructure-activity rela-tionships for naturally occurring coumarins as 120573-secretaseinhibitorrdquo Bioorganic and Medicinal Chemistry vol 20 no 2pp 784ndash788 2012
[10] S Marumoto and M Miyazawa ldquoBiotransformation of isoim-peratorin and imperatorin by Glomerella cingulata and 120573-secretase inhibitory activityrdquo Bioorganic and Medicinal Chem-istry vol 18 no 1 pp 455ndash459 2010
[11] S E Sajjadi Y Shokoohinia and S Hemmati ldquoIsolation andidentification of furanocoumarins and a phenylpropanoid fromthe acetone extract and identification of volatile constituentsfrom the essential oil of Peucedanum pastinacifoliumrdquo Chem-istry of Natural Compounds vol 48 no 4 pp 668ndash671 2012
[12] X B Yang J Hou D Liu et al ldquoBiotransformation of isoim-peratorin by Cunninghamella blakesleana AS 3970rdquo Journal ofMolecular Catalysis B vol 88 pp 1ndash6 2013
[13] W Lili S Yehong S Qi et al ldquoIn vitro permeability analy-sis pharmacokinetic and brain distribution study in mice ofimperatorin isoimperatorin and cnidilin in Radix AngelicaeDahuricaerdquo Fitoterapia vol 85 pp 144ndash153 2013
[14] S Wang Q Chen and L He ldquoDevelopment and validationof a gas chromatography-mass spectrometry method for thedetermination of isoimperatorin in rat plasma and tissueapplication to the pharmacokinetic and tissue distributionstudyrdquo Journal of Chromatography B vol 852 no 1-2 pp 473ndash478 2007
[15] H Sadraei Y Shokoohinia S E Sajjadi and M MozafarildquoAntispasmodic effects of Prangos ferulacea acetone extract andits maincomponent osthole on ileum contractionrdquo Research inPharmaceutical Sciences vol 8 pp 137ndash144 2013
[16] H Sadraei Y Shokoohinia S E Sajjadi and B GhadirianldquoAntispasmodic effect of osthole and Prangos ferulacea extracton rat uterus smooth muscle motilityrdquo Research in Pharmaceu-tical Sciences vol 7 pp 141ndash149 2012
[17] S Sajjadi Y Shokoohinia S Gholamzadeh M Behbahani andA Fattahi ldquoAntiviral evaluation of coumarins from Prangosferulacea L (Lindl)rdquo Research in Pharmaceutical Sciences vol7 p 783 2012
[18] S E Sajjadi Y Shokoohinia and S Gholamzadeh ldquoChemicalcomposition of the essential oil of the root of Prangos ferulacea(L) Lindlrdquo Chemija vol 22 pp 178ndash180 2011
[19] N Bijari Y Shokoohinia M-R Ashrafi-Kooshk et al ldquoSpec-troscopic study of interaction between osthole and humanserum albumin identification of possible binding site of thecompoundrdquo Journal of Luminescence vol 143 pp 328ndash336 2013
[20] X Shi M Liu M Zhang et al ldquoIdentification of in vitro and invivo metabolites of isoimperatorin using liquid chromatogra-phymass spectrometryrdquo Food Chemistry vol 141 pp 357ndash3652013
The Scientific World Journal 13
[21] T Chatterjee A Pal S Dey B-K Chatterjee and PChakrabarti ldquoInteraction of virstatin with human serum albu-min spectroscopic analysis andmolecular modelingrdquo Plos Onevol 7 pp 1ndash12 2012
[22] Y-M Chen and L-H Guo ldquoCombined fluorescence and elec-trochemical investigation on the binding interaction betweenorganic acid and human serum albuminrdquo Journal of Environ-mental Sciences vol 21 no 3 pp 373ndash379 2009
[23] J R Lakowicz Principles of Fluorescence Spectroscopy KluwerAcademicPlenum Publishers New York NY USA 2nd edi-tion 1999
[24] F Ding N Li B Han F Liu L Zhang and Y Sun ldquoThe bindingof CI Acid Red 2 to human serum albumin determinationof binding mechanism and binding site using fluorescencespectroscopyrdquo Dyes and Pigments vol 83 no 2 pp 249ndash2572009
[25] M-R Ashrafi Kooshk K Mansouri M Nadi S Khodarahmiand R Khodarahmi ldquoIn vitro anti-cancer activity of nativecurcumin and protein-curcuminrdquo Systems vol 2 no 1 pp 80ndash88 1981
[26] P D Ross and S Subramanian ldquoThermodynamics of proteinassociation reactions forces contributing to stabilityrdquo Biochem-istry vol 20 no 11 pp 3096ndash3102 1981
[27] J Liu J-N Tian J Zhang Z Hu and X Chen ldquoInteractionof magnolol with bovine serum albumin a fluorescence-quenching studyrdquo Analytical and Bioanalytical Chemistry vol376 no 6 pp 864ndash867 2003
[28] A Coi A M Bianucci F Bonomi P Rasmussen G M Muraand M L Ganadu ldquoStructural perturbation of 120572B-crystallinby zinc and temperature related to its chaperone-like activityrdquoInternational Journal of Biological Macromolecules vol 42 no3 pp 229ndash234 2008
[29] M Moller and A Denicola ldquoStudy of protein-ligand bindingby fluorescencerdquoBiochemistry andMolecular Biology Educationvol 30 no 5 pp 309ndash312 2002
[30] R Khodarahmi S A Karimi M R Ashrafi Kooshk S AGhadami S Ghobadi and M Amani ldquoComparative spec-troscopic studies on drug binding characteristics and proteinsurface hydrophobicity of native and modified forms of bovineserum albumin possible relevance to change in protein struc-turefunction upon non-enzymatic glycationrdquo SpectrochimicaActa A vol 89 pp 177ndash186 2012
[31] S-F Ma M Anraku Y Iwao et al ldquoHydrolysis of angiotensinII receptor blocker prodrug olmesartan medoxomil by humanserum albumin and identification of its catalytic active sitesrdquoDrug Metabolism and Disposition vol 33 no 12 pp 1911ndash19192005
[32] H M Berman J Westbrook Z Feng et al ldquoThe protein databankrdquo Nucleic Acids Research vol 28 no 1 pp 235ndash242 2000
[33] S Sugio A Kashima SMochizukiMNoda andKKobayashildquoCrystal structure of human serum albumin at 25 A resolutionrdquoProtein Engineering vol 12 no 6 pp 439ndash446 1999
[34] G M Morris D S Goodsell R Huey and A J OlsonldquoDistributed automated docking of flexible ligands to proteinsparallel applications of AutoDock 24rdquo Journal of Computer-Aided Molecular Design vol 10 no 4 pp 293ndash304 1996
[35] F Faridbod M R Ganjali B Larijani et al ldquoInteraction studyof pioglitazone with albumin by fluorescence spectroscopy andmolecular dockingrdquo Spectrochimica Acta A vol 78 no 1 pp96ndash101 2011
[36] D SGoodsell andA JOlson ldquoAutomated docking of substratesto proteins by simulated annealingrdquo Proteins vol 8 no 3 pp195ndash202 1990
[37] D S Goodsell G M Morris and A J Olson ldquoAutomateddocking of flexible ligands applications of AutoDockrdquo Journalof Molecular Recognition vol 9 no 1 pp 1ndash5 1996
[38] G Sudlow D J Birkett and D N Wade ldquoThe characterizationof two specific drug binding sites on human serum albuminrdquoMolecular Pharmacology vol 11 no 6 pp 824ndash832 1975
[39] Y Yue X Chen J Qin and X Yao ldquoCharacterization of themangiferin-human serum albumin complex by spectroscopicandmolecularmodeling approachesrdquo Journal of Pharmaceuticaland Biomedical Analysis vol 49 no 3 pp 753ndash759 2009
[40] A K Shaw and S K Pal ldquoSpectroscopic studies on the effectof temperature on pH-induced folded states of human serumalbuminrdquo Journal of Photochemistry and Photobiology B vol 90no 1 pp 69ndash77 2008
[41] S Ranjbar S Ghobadi R Khodarahmi and H Nemati ldquoSpec-troscopic characterization of furosemide binding to humancarbonic anhydrase IIrdquo International Journal of BiologicalMacromolecules vol 50 no 4 pp 910ndash917 2012
[42] HNHou Z POi YWOYung F L Liao Y Zhang andY LiyldquoStudies on interaction between vitamin B12 and human serumalbuminrdquo Journal of Pharmaceutical and Biomedical Analysisvol 47 pp 134ndash139 2008
[43] S S Kalanur J Seetharamappa and V K A Kalalbandi ldquoChar-acterization of interaction and the effect of carbamazepine onthe structure of human serum albuminrdquo Journal of Pharmaceu-tical and Biomedical Analysis vol 53 no 3 pp 660ndash666 2010
[44] L Stryer ldquoFluorescence spectroscopy of proteinsrdquo Science vol162 no 3853 pp 526ndash533 1968
[45] B Valeur and J C Brochon New Trends in FluorescenceSpectroscopy Springer Berlin Germany 1999
[46] V Suryawanshi P Anbhule A Gore Sh Patil and G AndKolekar ldquoA spectral deciphering the perturbation of modeltransporter protein (HSA) by antibacterial pyrimidine deriva-tive pharmacokinetic and biophysical insightsrdquo Journal ofPhotochemistry and Photobiology B vol 118 pp 1ndash8 2013
[47] T Yuan AMWeljie andH J Vogel ldquoTryptophan fluorescencequenching by methionine and selenomethionine residues ofcalmodulin orientation of peptide and protein bindingrdquo Bio-chemistry vol 37 no 9 pp 3187ndash3195 1998
[48] TCOrsquoHaver A F Fell G Smith et al ldquoDerivative spectroscopyand its applications in analysisrdquo Analytical Proceedings vol 19no 1 pp 22ndash46 1982
[49] Y-Q Wang B-P Tang H-M Zhang Q-H Zhou and G-C Zhang ldquoStudies on the interaction between imidaclopridand human serum albumin spectroscopic approachrdquo Journal ofPhotochemistry and Photobiology B vol 94 no 3 pp 183ndash1902009
[50] S-J Kim H-W Rhee H-Y Park H-S Kim and J-I HongldquoFluorescent probes designed for detecting human serum albu-min on the basis of its pseudo-esterase activityrdquo Bioorganic ampMedicinal Chemistry Letters vol 23 pp 2093ndash2097 2013
[51] P Ascenzi M Gioia G Fanali M Coletta and M FasanoldquoPseudo-enzymatic hydrolysis of 4-nitrophenyl acetate byhuman serum albumin pH-dependence of rates of individualstepsrdquo Biochemical and Biophysical Research Communicationsvol 424 pp 451ndash455 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
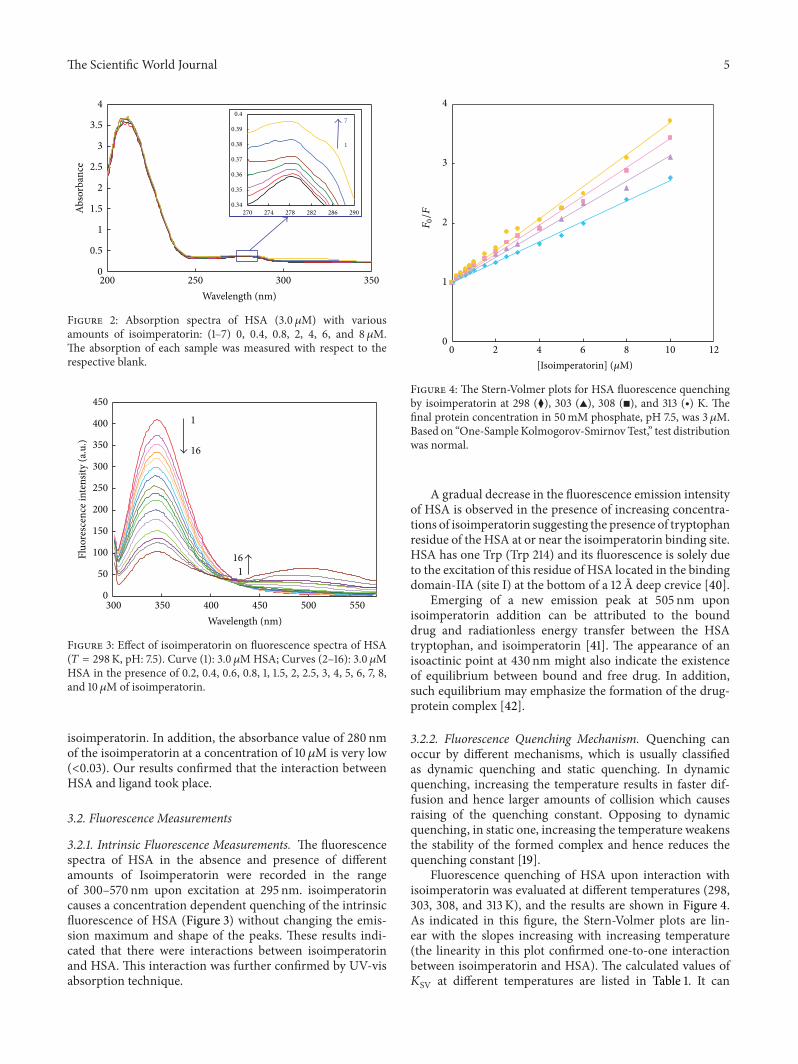
The Scientific World Journal 5
0
05
1
15
2
25
3
35
4
200 250 300 350Wavelength (nm)
Abso
rban
ce
034
035
036
037
038
039
04
270 274 278 282 286 290
1
7
Figure 2 Absorption spectra of HSA (30 120583M) with variousamounts of isoimperatorin (1ndash7) 0 04 08 2 4 6 and 8 120583MThe absorption of each sample was measured with respect to therespective blank
0
50
100
150
200
250
300
350
400
450
300 350 400 450 500 550Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
1
16
116
Figure 3 Effect of isoimperatorin on fluorescence spectra of HSA(119879 = 298K pH 75) Curve (1) 30 120583MHSA Curves (2ndash16) 30 120583MHSA in the presence of 02 04 06 08 1 15 2 25 3 4 5 6 7 8and 10120583M of isoimperatorin
isoimperatorin In addition the absorbance value of 280 nmof the isoimperatorin at a concentration of 10120583M is very low(lt003) Our results confirmed that the interaction betweenHSA and ligand took place
32 Fluorescence Measurements
321 Intrinsic Fluorescence Measurements The fluorescencespectra of HSA in the absence and presence of differentamounts of Isoimperatorin were recorded in the rangeof 300ndash570 nm upon excitation at 295 nm isoimperatorincauses a concentration dependent quenching of the intrinsicfluorescence of HSA (Figure 3) without changing the emis-sion maximum and shape of the peaks These results indi-cated that there were interactions between isoimperatorinand HSA This interaction was further confirmed by UV-visabsorption technique
0
1
2
3
4
0 2 4 6 8 10 12[Isoimperatorin] (120583M)
F0F
Figure 4 The Stern-Volmer plots for HSA fluorescence quenchingby isoimperatorin at 298 (⧫) 303 (998771) 308 (◼) and 313 (∙) K Thefinal protein concentration in 50mM phosphate pH 75 was 3 120583MBased on ldquoOne-Sample Kolmogorov-Smirnov Testrdquo test distributionwas normal
A gradual decrease in the fluorescence emission intensityof HSA is observed in the presence of increasing concentra-tions of isoimperatorin suggesting the presence of tryptophanresidue of the HSA at or near the isoimperatorin binding siteHSA has one Trp (Trp 214) and its fluorescence is solely dueto the excitation of this residue of HSA located in the bindingdomain-IIA (site I) at the bottom of a 12 A deep crevice [40]
Emerging of a new emission peak at 505 nm uponisoimperatorin addition can be attributed to the bounddrug and radiationless energy transfer between the HSAtryptophan and isoimperatorin [41] The appearance of anisoactinic point at 430 nm might also indicate the existenceof equilibrium between bound and free drug In additionsuch equilibrium may emphasize the formation of the drug-protein complex [42]
322 Fluorescence Quenching Mechanism Quenching canoccur by different mechanisms which is usually classifiedas dynamic quenching and static quenching In dynamicquenching increasing the temperature results in faster dif-fusion and hence larger amounts of collision which causesraising of the quenching constant Opposing to dynamicquenching in static one increasing the temperature weakensthe stability of the formed complex and hence reduces thequenching constant [19]
Fluorescence quenching of HSA upon interaction withisoimperatorin was evaluated at different temperatures (298303 308 and 313 K) and the results are shown in Figure 4As indicated in this figure the Stern-Volmer plots are lin-ear with the slopes increasing with increasing temperature(the linearity in this plot confirmed one-to-one interactionbetween isoimperatorin and HSA) The calculated values of119870SV at different temperatures are listed in Table 1 It can
6 The Scientific World Journal
Table 1 Stern-Volmer quenching constant (119870SV) quenching rate constant (119896119902) association constant (119870119887
) and number of binding sites (119899)for binding of isoimperatorin to HSA in the different temperatures
119879 (K) 10
minus4
times 119870SV(Mminus1)
10
minus12
times 119870
119902
(Mminus1sdotsminus1)119877
a 10
minus5
times 119870
119887
(Mminus1)Number of biding
sites (119899) 119877
a
2981698 plusmn 013 1698 plusmn 013
099621102 plusmn 010 0862 09841
3032019 plusmn 017 2019 plusmn 017
099551149 plusmn 013 0851 09886
3082333 plusmn 016 2333 plusmn 016
099651271 plusmn 022 0844 09950
3132639 plusmn 019 2639 plusmn 019
099811322 plusmn 017 0822 09917
Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870SV values
be found from Table 1 that the value of 119870SV increased withincreasing temperature in the presence of isoimperatorinwhich indicates the involved binding forces are mainlyhydrophobic interactions (endothermic apolar interactionsare strengthened with increasing temperature) Also the rateconstant of the quenching process (119896
119902) which is equal to119870SV
divided by 1205910(with assuming 120591
0= 10
minus8 s)
119896
119902=
119870SV120591
0
(10)
was calculated (Table 1) The results revealed that the valuesof 119896119902were much greater than the maximum diffusion rate
constant of the biomolecule (2 times 1010Mminus1 sminus1) which indi-cated that the fluorescence quenching of HSA was initiatedby complex formation between HSA and isoimperatorinconfirming that the mechanism of quenching is static [19]
323 Determination of Association Constant (119870119887) and Num-
ber of Binding Sites (119899) The values of association constant119870
119887 and the number of binding sites 119899 at different temper-
atures are obtained from the modified Stern-Volmer plots(Figure 5) and listed in Table 1 It can be found from thesedata that the 119899 values at different temperatures are keptaround unity which shows the existence of one binding sitein HSA for isoimperatorin The value of 119870
119887is significant to
understand the distribution of the drug in plasma since theweak binding can lead to a short life time or poor distributionwhile strong binding can decrease the concentration of freedrug in plasma The value of 119870
119887illustrated that there is a
strong binding force between isoimperatorin and HSA Itwas found that the binding constant increased with increasein temperature resulting in an increment of the stabilityof the isoimperatorin-HSA complex it also implies thatisoimperatorin can be tightly stored and carried by HSA inthe body
324 Forces Involved in the Binding Process Consideringthe dependence of binding constant on temperature a ther-modynamic process was considered to be responsible forthe formation of a complex Therefore the thermodynamicparameters dependent on temperatures were analyzed Theacting forces between a small molecule and macromoleculemainly include hydrogen bonds van der Waals forces elec-trostatic forces and hydrophobic interaction forces [23] Inorder to obtain the information about the forces persisting in
minus15
minus13
minus11
minus09
minus07
minus05
minus03
minus01
01
03
05
minus68 minus65 minus62 minus59 minus56 minus53 minus5
log[isoimperatorin]
log[(F0minusF)F]
Figure 5 The modified Stern-Volmer plots for HSA fluorescencequenching by isoimperatorin at 298 (⧫) 303 (998771) 308 (◼) and 313 (∙)K For interpretation of references to color in this figure legend thereader is referred to the web version of this article
the present binding process the thermodynamic parametersof binding were obtained fromVanrsquot Hoff plot followed by theGibbs equation (data not shown) Table 2 shows the values ofΔ119867
∘ andΔ119878∘ obtained for the binding site from the slopes andordinates at the origin of the fitted lines From Table 2 it canbe seen that the negative sign for free energy (Δ119866∘) meansthat the interaction process is spontaneous The positiveenthalpy (Δ119867∘) and entropy (Δ119878∘) values of the interaction ofisoimperatorin and HSA indicate that the binding is mainlyentropy-driven and the enthalpy is unfavorable for it thehydrophobic forces play a major role in the reaction [43]
33 Nonradiative Energy Transfer from HSA to Isoimpera-torin The Forster nonradiative energy transfer (FRET) hasbeen used as a ldquospectroscopic rulerrdquo for measurement ofthe distance between the acceptor (ligand) and the donor(tryptophan residues in the protein) [44] According to thistheory there will be FRET between two molecules if thefollowing conditions were satisfied (a) the donor moleculeemits fluorescence (b) the emission spectrum of the donorhas an enough overlap with the absorption spectrum of
The Scientific World Journal 7
Table 2 Thermodynamic parameters for binding isoimperatorin to HSA
119879 (K) Δ119878
∘ (Jsdotmolminus1sdotKminus1) Δ119867
∘ (kJsdotmolminus1) Δ119866
∘ (kJsdotmolminus1)298 minus2969 plusmn 009
303 17820 plusmn 011 2993 plusmn 013 minus3071 plusmn 007
308 minus3169 plusmn 010
313 minus3299 plusmn 008
0
50
100
150
200
250
300
350
400
450
222 272 322 372 422Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
0
002
004
006
008
01
012
014
Abso
rban
ce
a
b
Figure 6 The overlap of the fluorescence spectrum of HSA (a)and the absorption spectrum of isoimperatorin (b) when [HSA] =[isoimperatorin] = 10 120583MThe enzyme was excited at 295 nm
the acceptor and (c) the donor and acceptor molecules areclose enough with the maximum distance not to exceeding7 nm [45]
There is a good overlapping between the fluorescenceemission spectrum of HSA and absorption spectra of isoim-peratorin (Figure 6) As the fluorescence emission of proteinwas affected by the excitation light around 295 nm thespectrum ranging from 300ndash400 nm was chosen to calculatethe overlapping integral The value of 119869 was calculated to be473 plusmn 005 times 10
minus15 cm3 Lmolminus1 by integrating the overlapspectra for 300ndash400 nm In the present case 1198702 = 23119873 = 136 and Φ = 0118 [21] according to (6) and (7) wecould calculate that 119864 = 045 plusmn 006 119877
0= 322 plusmn 03 nm and
119903 = 29 plusmn 02 nm As mentioned earlier (Section 321) HSAposses one tryptophan residue (Trp 214) and the calculated119903 is actually the average value between the bound drug andTrp 214 Since 119903 is much smaller than 7 nm and 05119877
0lt
119903 lt 15119877
0[41] therefore the energy transfer from HSA
to isoimperatorin occurs with high probability which alsoexplains the efficient quenching of the HSA fluorescence
34 PSH Determination in the Absence and Presence of Isoim-peratorin Several methods exist for measuring the surfacehydrophobicity of proteins that ANS experiment is one ofthem ANS as an extrinsic fluorescent probe is extremelysensitive to polarity of the solvent In aqueous solutions itfluoresces very weakly but upon binding to hydrophobicpatches of proteins its quantum yield increases significantly[29] Thus ANS fluorescence may be applied for monitoringpossible changes in protein surface hydrophobicity induced
0
200
400
600
800
1000
0 10 20 30 40 50
Fluo
resc
ence
inte
nsity
(au
)
[ANS] (120583M)
Figure 7 Binding of ANS to HSA (1120583M) in the absence (∙) andpresence (998771) of 3120583M of isoimperatorin
upon drug binding At a fixed concentration (1 120583M) of HSAand increasing concentrations of ANS (0ndash50120583M) fluores-cence intensity of ANS was measured in the absence andpresence of 3120583M isoimperatorin (Figure 7)
A typical hyperbolic response is observed which demon-strates the saturation nature of ANS binding toHSA It can beseen that the hyperbolic response of ANS fluorescence in thepresence of isoimperatorin is slightly different in comparisonto the absence of the drug The values of 119865max and 119870
app119889
forANS are obtained from the Scatchard plot (Figure 8) andlisted in Table 3 The value of 119870app
119889
for ANS binding in theabsence of drug (09 plusmn 005) was increased in the presenceof 3 120583M isoimperatorin (14 plusmn 007) which shows the weakerbinding of ANS to the HSA-isoimperatorin complex ThenPSH was calculated in the absence and the presence of3 120583M isoimperatorin by using 119865max and119870
app119889
values (Table 3)Decreasing the PSH index of HSA (from 150794 plusmn 1146 to80612 plusmn 007) suggests that the surface hydrophobicity ofHSA is decreased by sim465 upon drug binding
35 Synchronous Fluorescence Spectroscopy Synchronous flu-orescence spectroscopy introduced by Lody has been usedto characterize complex mixtures which can provide finger-prints of complex samples [46] The synchronous fluores-cence spectra can provide information about the molecularenvironment in the vicinity of the chromophore moleculesIn the synchronous fluorescence spectra the sensitivityassociated with fluorescence is maintained while several
8 The Scientific World Journal
Table 3 Surface hydrophobicity parameters for HSA in the absence and the presence of isoimperatorin
Parameters HSA HSA-IIM complex119870
app119889
(120583M) 09 plusmn 005
lowast
14 plusmn 007
lowast
119865max[HSA] 135714 plusmn 85
lowastlowast
112853 plusmn 63
lowastlowast
Surface hydrophobicity index (119865max[HSA] 119870app119889
) 150794 plusmn 1146
lowastlowast
80612 plusmn 007
lowastlowast
Data are expressed as mean plusmn SD of three measurements lowast119875 value lt 00006 lowastlowast119875 value lt 00001
0
50
100
150
200
250
300
350
400
400 500 600 700 800 900 1000Fluorescence intensity (au)
Fmax
Fluo
resc
ence
inte
nsity
(au
)[A
NS]
free
Slope = minus1Kappd
Figure 8 Scatchard plots for titration of HSA with increasingconcentrations of ANS in the absence (∙) and presence (998771) of 3 120583Mof isoimperatorin 119865max and 1119870
app119889
values were obtained from the119909-intercept and slope of this plot For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
advantages are available spectral simplification spectralbandwidth reduction and avoidance of different perturbingeffects The fluorescence spectrum of BSAHSA mainly dueto the Trp residues is sensitive to the microenvironment ofthese chromophoreswith themaximumemissionwavelengthbeing very useful in estimating the hydrophobicity aroundthe tryptophan residues It is well known that the experimenton synchronous fluorescence of HSA will provide the char-acteristic information for the Tyr and Trp residues when thescanning interval Δ120582 is fixed at 15 and 60 nm respectively[47] The shift in the position of fluorescence emissionmaximum corresponds to changes of the polarity aroundthe chromophore molecule A blue shift of 120582max means thatthe amino acid residues are located in a more hydrophobicenvironment and are less exposed to the solvent while a redshift of 120582max implies that the amino acid residues are in apolar environment and are more exposed to the solvent [48]The synchronous fluorescence spectra of interaction betweenisoimperatorin and HSA are presented in Figure 8 It wasshown that the fluorescence intensity of HSA decreased regu-larly along with the addition of isoimperatorin which furtherdemonstrated the occurrence of fluorescence quenching inthe binding process Moreover there is no significant shiftof the maximum emission wavelength with Δ120582 = 15 nm or60 nm (Figures 9(a) and 9(b)) which implies that interaction
of isoimperatorin with HSA does not affect the conformationof the region around the Tyr or Trp residues
As well we can obtain the information about the locationof isoimperatorin binding site from the synchronous fluores-cence dataThe ratios of synchronous fluorescence quenching(RSFQ) were used to determine the binding sites of the drugsto the HSA molecule in drug-HSA complexes The RSFQvalues were calculated from the following equation
RSFQ = 1 minus
119865
119865
0
(11)
where 119865 and 1198650are the synchronous fluorescence intensities
of HSA in the presence and the absence of drug respectivelyIt was obvious from Figure 9(c) that the RSFQ results forΔ120582 = 60 nmwere slightly higher than the corresponding onesfor Δ120582 = 15 nm As illustrated in Figure 9(c) addition ofisoimperatorin resulted in strong fluorescence quenching ofTrp residue (sim47 reduction in fluorescence intensity) con-firming that isoimperatorin reached subdomain IIA whereonly one Trp residue (Trp 214) in HSA is located It wasalso observed from Figure 9(c) that the fluorescence strengthof Tyr residues decreased about sim32 in the presence ofisoimperatorin So the binding of isoimperatorin to HSAquenched flourescence intensity of Tyr and Trp Since Trp214 (the only Trp of HSA) is at site I and Tyr 263 (hasthe strong fluorescence in the case of Tyr residues andmay be one of the residues responsible for the synchronousfluorescence spectrum of HSA when Δ120582 is fixed at 15 nm) isin subdomain IIA (site I) it can be concluded that Trp andTyr residues of HSA may have equal accessibility to bindingsite of isoimperatorin [49] The above results indicated thatisoimperatorin can bind to HSA in the hydrophobic cavity(site I) on subdomain IIA which are in full agreement withquenching competitive binding experiments and theoreticaldocking technique
36 Site Marker Competitive Binding Experiments In orderto further classify drug binding site on HSA competitivebinding experiments have been carried out Warfarin andibuprofen are two markers specific for binding to HSA thatbind to sites I and II which are corresponded to the sub-domains IIA and IIIA respectively Therefore to determinethe location of the isoimperatorin binding site on HSA thecompetitive displacement experimentswere carried out usingwarfarin as a characteristicmarker for site I (Figure 10(a)) andibuprofen for site II (Figure 10(b)) [24 31] In the site markercompetitive experiment isoimperatorin was gradually addedto the solution of HSA HSA-ibuprofen or HSA-warfarincomplex and then fluorescence intensity of system wasrecorded
The Scientific World Journal 9
0
100
200
300
400
500
600
265 270 275 280 285 290 295 300Wavelength (nm)
1
8
Fluo
resc
ence
inte
nsity
(au
)
(a)
Fluo
resc
ence
inte
nsity
(au
)
0
50
100
150
200
250
300
350
400
250 260 270 280 290 300Wavelength (nm)
1
8
(b)
0
10
20
30
40
50
0 05 1 15 2 25 3
RSFQ
()
Isoimperatorin (120583M)
0
02
04
06
08
1
12
005 1 15 2 25 3 35
Isoimperatorin (120583M)
F0
F
(c)
Figure 9 Synchronous fluorescence spectra of interaction between HSA and isoimperatorin (a) at Δ120582 = 15 nm and (b) at Δ120582 = 60 nmConcentration ofHSAwas 30 120583Mwhile concentrations of isoimperatorinwere 0 04 08 12 16 2 25 and 3 120583Mfrom 1 to 8 (c) Comparativeevaluation of isoimperatorin effect on the RSFQ of HSA [HSA] = 3 120583M (◼) Δ120582 = 15 nm and (∙) Δ120582 = 60 nm
As shown in Figure 10(a) with addition of warfarin in theHSA solution the flourescence intensity was slightly higherthan that of without warfarin (and including the red shift)Then after adding the isoimperatorin into the above systemthe flourescence intensity of HSA solution with warfarinheld in equimolar decreased gradually and the intensitywas much lower than that of without warfarin displayingthat the binding of the isoimperatorin to HSA was affectedafter adding warfarin On the contrary in the presence ofibuprofen the flourescence intensity of the HSA-ibuprofencomplex almost had no difference from that recordedwithoutibuprofen under the same conditions (Figure 10(b)) whichindicated that site II marker did not prevent the binding ofisoimperatorin in its usual binding location The next stepprotein intrinsic fluorescence was measured and the relativefluorescence intensities (119865119865
0) versus ligand concentration
([isoimperatorin]) plots are displayed in Figure 10(c) Asshown in Table 4 the binding constant was remarkablydecreased after addition of warfarin while the addition ofibuprofen results in only a small difference These resultsindicate that warfarin can displace the isoimperatorin but
ibuprofen has little effect on the binding of isoimperatorinto HSA In other words these results suggest that isoim-peratorin probably competes with warfarin for binding toHSA [24] Experimental observations were followed up withdocking studies where isoimperatorin was docked to HSAto determine the preferred binding site on the proteinAs described above the 3D crystal structure of HSA is amonomer consisting of three homologous domains whichassemble to form a heart-shaped molecule [4 6] Each ofthe structurally similar 120572-helix domains (IndashIII) has twosubdomains (A andB) with six120572-helices in subdomainA andfour 120572-helices in subdomain BThe crystal structure analysesindicate that the principal regions of ligand binding sitesin HSA are located in hydrophobic cavities in subdomainsIIA and IIIA which are consistent with Sudlowrsquos site I andSudlowrsquos site II respectively Several studies have shown thatHSA is able to bind many ligands in several binding sites[38] In this paper the molecular docking model of theHSAndashisoimperatorin complex was conducted on the basisof the crystal structure of HSA to find the binding positionof isoimperatorin The best docking energy result is shown
10 The Scientific World Journal
0
100
200
300
400
500
600
310 335 360 385 410Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA1
8
(a)
0
100
200
300
400
500
600
700
305 325 345 365 385 405Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA
1
8
(b)
0
05
1
15
0 1 2 3
HSAIbuprofen
Warfarin
Isoimperatorin (120583M)
F0
F
(c)
Figure 10 Effect of isoimperatorin on the quenching of HSA-warfarin complex (a) andHSA-ibuprofen complex (b) fluorescence From 1 to 8[HSA] = [warfarin or iboprufen] = 3 120583M [isoimperatorin] = 0 04 08 12 16 2 25 and 3 120583M All reproduced data shown are representativeof at least 3 independent experiments (c) Fluorescence quenching profiles of HSA (⧫) HSA-ibobrofen (◼) and HSA-warfarin (998771) in thepresence of various concentration of isoimperatorin
in Figure 11 It can be seen that isoimperatorin was situatedwithin subdomain IIA in Sudlowrsquos site I formed by six 120572-helices This figure shows the location of Trp 214 and also thebinding of isoimperatorin in HSA As this fluorescent residueis in subdomain IIA and due to the proximity of the bindingsite of isoimperatorin to the Trp 214 it can be concluded thatisoimperatorin has one reactive site onHSAandprobably thissite is at or near the site I The above docking results are ingood agreement with the displacement experiments results
and revealed that isoimperatorin binds to Sudlowrsquos site I ofHSA
37 Effect of Isoimperatorin on HSA Esterase Activity Pre-liminary studies showed that albumin in addition to itsligand binding capabilities has remarkable promiscuouscatalytic activities toward a broad range of organicmoleculesincluding esters amides phosphates and benzisoxazolesHuman serum albumin had significantly greater esterase-like
The Scientific World Journal 11
Table 4 Binding constants for isoimperatorin-HSA interaction (at 295K) in the absence and presence of the site marker
119870
119887
times 10
minus5
(Mminus1) 119877
a
HSA + isoimperatorin (in the absence ofibuprofen and warfarin) 1110 plusmn 04
lowast 09947
HSA + isoimperatorin + ibuprofen 1099 plusmn 06
lowast 09904HSA + isoimperatorin + warfarin 789 plusmn 03
lowast 09881Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870
119887values lowast119875 value lt 00002
Trp 214
Isoimperatorin
Figure 11 Crystal structure of HSA and the positions of singletryptophan residue (Trp 214) and also the binding site of isoim-peratorin in HSA Protein backbone is shown in the ldquocartoonrdquorepresentation and isoimperatorinand Trp 214 residue in the site Iare shown in the ldquostickrdquo representation The isoimperatorin and Trp214 structures are represented by the red and blue colorsThe crystalstructure of protein was obtained from Protein Data Bank (1BM0)For interpretation of references to color in this figure legend thereader is referred to the web version of this paper
activity than bovine serum albumin and defatted albuminwas more active than untreated albumin [50] InterestinglyHSA exhibits esterase-like activity and hydrolyzes drugshaving an ester group such as aspirin (which can convertaspirin ldquoacetylsalicylic acidrdquo to salicylic acid) Lys199 Lys402Lys519 and Lys545 form the pseudoesterase active site ofHSAthat is located in site I [50] ldquoEsterase-likerdquo activity (hydrolysisof p-nitrophenol esters such as p-NPA) by subdomain III Ahas been also reported and site directed mutagenesis studieshave shown that Arg 410 and Tyr 411 are essential in thisesterase activity However further investigation revealed thatacetylation is not limited to a distinct side chain so that theHSA incubation with high concentrations of pNPAc resultsin the acetylation of 59 Lys 10 Ser 8 Thr 4 Tyr and 1 Asp[51] Because of the above difference here we set out toevaluate the effect of isoimperatorin on the esterase activityof HSA based on our previous work [19] The kinetics ofthe pseudoenzymatic hydrolysis of pNPAc was obtained inthe absence and presence of isoimperatorin and site markers(Figure 12) Accordingly warfarin and indomethacin (as siteI marker) and isoimperatorin inhibited competitively theHSA-catalyzed hydrolysis of pNPAc by binding to site I Incontrast ibuprofen and tetracycline were unable to inhibitesterase activity of HSA The enzyme inhibition results sug-gested that isoimperatorin aspirin warfarin indomethacin
0
005
01
015
02
025
03
035
0 05 1 15 2 25 3Time (min)
HSA TetracyclineIbuprofenAspirin
IndomethacinOstholeWarfarinIsoimperatorin
A400
Figure 12 4-Nitrophenyl acetate esterase activity of HSA in theabsence and presence of tetracycline and ibuprofen (site II) war-farin aspirin indomethacin and osthole (site I) and isoimperatorin(site I) Final concentration of protein pNPA and drugs were 15250 and 250 120583M respectively For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
and osthole binding sites on the HSA molecule are the sameThese results also proposed that the large site I on HSAparticipates effectively to its promiscuous esterase activity
4 Conclusion
The accumulated data on HSA-drug interactions can beused to develop quantitative structure-activity relationshipsfor albumin binding However as stated earlier adsorptiondistribution metabolism and elimination (ADME) is con-trolled by albumin [30] Poor or strong binding to SA isbelieved to affect half-time of drugs [35] and their dosagedepends on their half-life in plasma Thus the bindingaffinity of any drug to SA is one of the major factors thatdetermine the pharmacokinetics halftime and availabilityof the drug in various tissues (see [30] and referencestherein) Further complexities arise in vivo due to interactions
12 The Scientific World Journal
between drugs and endogenous ligands for HSA This isparticularly pertinent for fatty acids which normally occurin serum at levels between 01 and 2mol per mol of HSAand can both compete and cooperate with drugs bindingto the protein In certain disease states these effects areexacerbated as the fatty acid HSA molar ratio may be ashigh as six Other pathological conditions are associatedwith high (micromolar to millimolar) levels of bilirubinhemin or renal toxins (eg 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) indoxyl sulphate) which bindto the protein causing significant drug binding defects
Our major aim in this study was to distinguish thenature of the interaction of isoimperatorin to HSA usingdifferent biophysical techniques This study could be veryvaluable due to importance and presence of isoimperatorinin human diet (food-drug interaction on HSA) The resultsshowed that isoimperatorin quenched the intrinsic fluo-rescence of HSA through dynamic quenching mechanismFurthermore hydrophobic interaction played a major role inthe binding process Using Fluorescence and UV absorptionspectroscopy it has been shown that isoimperatorin inducesprotein structural changes The binding average distancebetween the solo tryptophan of the protein and the boundisoimperatorin was calculated by Forster theory as 290 plusmn02 nmAnalysis of PSHvalues elucidated that isoimperatorinhas induced some compactness in the tertiary structure ofthe enzyme The results of synchronous flourescence onHSA-isoimperatorin system elucidated that the drug binds tothe hydrophobic cavity (site I) on subdomain IIA that wasconfirmed by quenching experiments competitive bindingexperiments and docking technique
Acknowledgment
The authors gratefully acknowledge the Kermanshah Uni-versity of Medical Sciences Research Council for financialsupport of this work
References
[1] Y-J Hu Y Liu T-Q Sun A-M Bai J-Q Lu and Z-B Pi ldquoBinding of anti-inflammatory drug cromolyn sodiumto bovine serum albuminrdquo International Journal of BiologicalMacromolecules vol 39 no 4-5 pp 280ndash285 2006
[2] N Seedher and S Bhatia ldquoReversible binding of celecoxiband valdecoxib with human serum albumin using fluorescencespectroscopic techniquerdquo Pharmacological Research vol 54 no2 pp 77ndash84 2006
[3] H Isogai and N Hirayama ldquoIn silico prediction of interactionsbetween site II on human serum albumin and profen drugsrdquoISRN Pharma vol 2013 Article ID 818364 8 pages 2013
[4] YWangHYuX Shi Z LuoD Lin andMHuang ldquoStructuralmechanism of ring opening reaction of glucose by humanserum albuminrdquo The Journal of Biological Chemistry vol 288pp 15980ndash15987 2013
[5] F Keshavarz M M Alavianmehr and R Yousefi ldquoMolecularinteraction of benzalkonium Ibuprofenate and its discreteingredients with human serum albuminrdquo Physical ChemistryResearch vol 1 no 2 pp 111ndash116 2013
[6] T K Maiti K S Ghosh J Debnath and S Dasgupta ldquoBindingof all-trans retinoic acid to human serum albumin fluores-cence FT-IR and circular dichroism studiesrdquo InternationalJournal of Biological Macromolecules vol 38 no 3ndash5 pp 197ndash202 2006
[7] M-X XieM Long Y Liu C Qin and Y-DWang ldquoCharacter-ization of the interaction between human serum albumin andmorinrdquo Biochimica et Biophysica Acta vol 1760 no 8 pp 1184ndash1191 2006
[8] CD Kanakis P A TarantilisMG Polissiou S Diamantoglouand H A Tajmir-Riahi ldquoAntioxidant flavonoids bind humanserum albuminrdquo Journal of Molecular Structure vol 798 no 1-3 pp 69ndash74 2006
[9] S Marumoto and M Miyazawa ldquoStructure-activity rela-tionships for naturally occurring coumarins as 120573-secretaseinhibitorrdquo Bioorganic and Medicinal Chemistry vol 20 no 2pp 784ndash788 2012
[10] S Marumoto and M Miyazawa ldquoBiotransformation of isoim-peratorin and imperatorin by Glomerella cingulata and 120573-secretase inhibitory activityrdquo Bioorganic and Medicinal Chem-istry vol 18 no 1 pp 455ndash459 2010
[11] S E Sajjadi Y Shokoohinia and S Hemmati ldquoIsolation andidentification of furanocoumarins and a phenylpropanoid fromthe acetone extract and identification of volatile constituentsfrom the essential oil of Peucedanum pastinacifoliumrdquo Chem-istry of Natural Compounds vol 48 no 4 pp 668ndash671 2012
[12] X B Yang J Hou D Liu et al ldquoBiotransformation of isoim-peratorin by Cunninghamella blakesleana AS 3970rdquo Journal ofMolecular Catalysis B vol 88 pp 1ndash6 2013
[13] W Lili S Yehong S Qi et al ldquoIn vitro permeability analy-sis pharmacokinetic and brain distribution study in mice ofimperatorin isoimperatorin and cnidilin in Radix AngelicaeDahuricaerdquo Fitoterapia vol 85 pp 144ndash153 2013
[14] S Wang Q Chen and L He ldquoDevelopment and validationof a gas chromatography-mass spectrometry method for thedetermination of isoimperatorin in rat plasma and tissueapplication to the pharmacokinetic and tissue distributionstudyrdquo Journal of Chromatography B vol 852 no 1-2 pp 473ndash478 2007
[15] H Sadraei Y Shokoohinia S E Sajjadi and M MozafarildquoAntispasmodic effects of Prangos ferulacea acetone extract andits maincomponent osthole on ileum contractionrdquo Research inPharmaceutical Sciences vol 8 pp 137ndash144 2013
[16] H Sadraei Y Shokoohinia S E Sajjadi and B GhadirianldquoAntispasmodic effect of osthole and Prangos ferulacea extracton rat uterus smooth muscle motilityrdquo Research in Pharmaceu-tical Sciences vol 7 pp 141ndash149 2012
[17] S Sajjadi Y Shokoohinia S Gholamzadeh M Behbahani andA Fattahi ldquoAntiviral evaluation of coumarins from Prangosferulacea L (Lindl)rdquo Research in Pharmaceutical Sciences vol7 p 783 2012
[18] S E Sajjadi Y Shokoohinia and S Gholamzadeh ldquoChemicalcomposition of the essential oil of the root of Prangos ferulacea(L) Lindlrdquo Chemija vol 22 pp 178ndash180 2011
[19] N Bijari Y Shokoohinia M-R Ashrafi-Kooshk et al ldquoSpec-troscopic study of interaction between osthole and humanserum albumin identification of possible binding site of thecompoundrdquo Journal of Luminescence vol 143 pp 328ndash336 2013
[20] X Shi M Liu M Zhang et al ldquoIdentification of in vitro and invivo metabolites of isoimperatorin using liquid chromatogra-phymass spectrometryrdquo Food Chemistry vol 141 pp 357ndash3652013
The Scientific World Journal 13
[21] T Chatterjee A Pal S Dey B-K Chatterjee and PChakrabarti ldquoInteraction of virstatin with human serum albu-min spectroscopic analysis andmolecular modelingrdquo Plos Onevol 7 pp 1ndash12 2012
[22] Y-M Chen and L-H Guo ldquoCombined fluorescence and elec-trochemical investigation on the binding interaction betweenorganic acid and human serum albuminrdquo Journal of Environ-mental Sciences vol 21 no 3 pp 373ndash379 2009
[23] J R Lakowicz Principles of Fluorescence Spectroscopy KluwerAcademicPlenum Publishers New York NY USA 2nd edi-tion 1999
[24] F Ding N Li B Han F Liu L Zhang and Y Sun ldquoThe bindingof CI Acid Red 2 to human serum albumin determinationof binding mechanism and binding site using fluorescencespectroscopyrdquo Dyes and Pigments vol 83 no 2 pp 249ndash2572009
[25] M-R Ashrafi Kooshk K Mansouri M Nadi S Khodarahmiand R Khodarahmi ldquoIn vitro anti-cancer activity of nativecurcumin and protein-curcuminrdquo Systems vol 2 no 1 pp 80ndash88 1981
[26] P D Ross and S Subramanian ldquoThermodynamics of proteinassociation reactions forces contributing to stabilityrdquo Biochem-istry vol 20 no 11 pp 3096ndash3102 1981
[27] J Liu J-N Tian J Zhang Z Hu and X Chen ldquoInteractionof magnolol with bovine serum albumin a fluorescence-quenching studyrdquo Analytical and Bioanalytical Chemistry vol376 no 6 pp 864ndash867 2003
[28] A Coi A M Bianucci F Bonomi P Rasmussen G M Muraand M L Ganadu ldquoStructural perturbation of 120572B-crystallinby zinc and temperature related to its chaperone-like activityrdquoInternational Journal of Biological Macromolecules vol 42 no3 pp 229ndash234 2008
[29] M Moller and A Denicola ldquoStudy of protein-ligand bindingby fluorescencerdquoBiochemistry andMolecular Biology Educationvol 30 no 5 pp 309ndash312 2002
[30] R Khodarahmi S A Karimi M R Ashrafi Kooshk S AGhadami S Ghobadi and M Amani ldquoComparative spec-troscopic studies on drug binding characteristics and proteinsurface hydrophobicity of native and modified forms of bovineserum albumin possible relevance to change in protein struc-turefunction upon non-enzymatic glycationrdquo SpectrochimicaActa A vol 89 pp 177ndash186 2012
[31] S-F Ma M Anraku Y Iwao et al ldquoHydrolysis of angiotensinII receptor blocker prodrug olmesartan medoxomil by humanserum albumin and identification of its catalytic active sitesrdquoDrug Metabolism and Disposition vol 33 no 12 pp 1911ndash19192005
[32] H M Berman J Westbrook Z Feng et al ldquoThe protein databankrdquo Nucleic Acids Research vol 28 no 1 pp 235ndash242 2000
[33] S Sugio A Kashima SMochizukiMNoda andKKobayashildquoCrystal structure of human serum albumin at 25 A resolutionrdquoProtein Engineering vol 12 no 6 pp 439ndash446 1999
[34] G M Morris D S Goodsell R Huey and A J OlsonldquoDistributed automated docking of flexible ligands to proteinsparallel applications of AutoDock 24rdquo Journal of Computer-Aided Molecular Design vol 10 no 4 pp 293ndash304 1996
[35] F Faridbod M R Ganjali B Larijani et al ldquoInteraction studyof pioglitazone with albumin by fluorescence spectroscopy andmolecular dockingrdquo Spectrochimica Acta A vol 78 no 1 pp96ndash101 2011
[36] D SGoodsell andA JOlson ldquoAutomated docking of substratesto proteins by simulated annealingrdquo Proteins vol 8 no 3 pp195ndash202 1990
[37] D S Goodsell G M Morris and A J Olson ldquoAutomateddocking of flexible ligands applications of AutoDockrdquo Journalof Molecular Recognition vol 9 no 1 pp 1ndash5 1996
[38] G Sudlow D J Birkett and D N Wade ldquoThe characterizationof two specific drug binding sites on human serum albuminrdquoMolecular Pharmacology vol 11 no 6 pp 824ndash832 1975
[39] Y Yue X Chen J Qin and X Yao ldquoCharacterization of themangiferin-human serum albumin complex by spectroscopicandmolecularmodeling approachesrdquo Journal of Pharmaceuticaland Biomedical Analysis vol 49 no 3 pp 753ndash759 2009
[40] A K Shaw and S K Pal ldquoSpectroscopic studies on the effectof temperature on pH-induced folded states of human serumalbuminrdquo Journal of Photochemistry and Photobiology B vol 90no 1 pp 69ndash77 2008
[41] S Ranjbar S Ghobadi R Khodarahmi and H Nemati ldquoSpec-troscopic characterization of furosemide binding to humancarbonic anhydrase IIrdquo International Journal of BiologicalMacromolecules vol 50 no 4 pp 910ndash917 2012
[42] HNHou Z POi YWOYung F L Liao Y Zhang andY LiyldquoStudies on interaction between vitamin B12 and human serumalbuminrdquo Journal of Pharmaceutical and Biomedical Analysisvol 47 pp 134ndash139 2008
[43] S S Kalanur J Seetharamappa and V K A Kalalbandi ldquoChar-acterization of interaction and the effect of carbamazepine onthe structure of human serum albuminrdquo Journal of Pharmaceu-tical and Biomedical Analysis vol 53 no 3 pp 660ndash666 2010
[44] L Stryer ldquoFluorescence spectroscopy of proteinsrdquo Science vol162 no 3853 pp 526ndash533 1968
[45] B Valeur and J C Brochon New Trends in FluorescenceSpectroscopy Springer Berlin Germany 1999
[46] V Suryawanshi P Anbhule A Gore Sh Patil and G AndKolekar ldquoA spectral deciphering the perturbation of modeltransporter protein (HSA) by antibacterial pyrimidine deriva-tive pharmacokinetic and biophysical insightsrdquo Journal ofPhotochemistry and Photobiology B vol 118 pp 1ndash8 2013
[47] T Yuan AMWeljie andH J Vogel ldquoTryptophan fluorescencequenching by methionine and selenomethionine residues ofcalmodulin orientation of peptide and protein bindingrdquo Bio-chemistry vol 37 no 9 pp 3187ndash3195 1998
[48] TCOrsquoHaver A F Fell G Smith et al ldquoDerivative spectroscopyand its applications in analysisrdquo Analytical Proceedings vol 19no 1 pp 22ndash46 1982
[49] Y-Q Wang B-P Tang H-M Zhang Q-H Zhou and G-C Zhang ldquoStudies on the interaction between imidaclopridand human serum albumin spectroscopic approachrdquo Journal ofPhotochemistry and Photobiology B vol 94 no 3 pp 183ndash1902009
[50] S-J Kim H-W Rhee H-Y Park H-S Kim and J-I HongldquoFluorescent probes designed for detecting human serum albu-min on the basis of its pseudo-esterase activityrdquo Bioorganic ampMedicinal Chemistry Letters vol 23 pp 2093ndash2097 2013
[51] P Ascenzi M Gioia G Fanali M Coletta and M FasanoldquoPseudo-enzymatic hydrolysis of 4-nitrophenyl acetate byhuman serum albumin pH-dependence of rates of individualstepsrdquo Biochemical and Biophysical Research Communicationsvol 424 pp 451ndash455 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 The Scientific World Journal
Table 1 Stern-Volmer quenching constant (119870SV) quenching rate constant (119896119902) association constant (119870119887
) and number of binding sites (119899)for binding of isoimperatorin to HSA in the different temperatures
119879 (K) 10
minus4
times 119870SV(Mminus1)
10
minus12
times 119870
119902
(Mminus1sdotsminus1)119877
a 10
minus5
times 119870
119887
(Mminus1)Number of biding
sites (119899) 119877
a
2981698 plusmn 013 1698 plusmn 013
099621102 plusmn 010 0862 09841
3032019 plusmn 017 2019 plusmn 017
099551149 plusmn 013 0851 09886
3082333 plusmn 016 2333 plusmn 016
099651271 plusmn 022 0844 09950
3132639 plusmn 019 2639 plusmn 019
099811322 plusmn 017 0822 09917
Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870SV values
be found from Table 1 that the value of 119870SV increased withincreasing temperature in the presence of isoimperatorinwhich indicates the involved binding forces are mainlyhydrophobic interactions (endothermic apolar interactionsare strengthened with increasing temperature) Also the rateconstant of the quenching process (119896
119902) which is equal to119870SV
divided by 1205910(with assuming 120591
0= 10
minus8 s)
119896
119902=
119870SV120591
0
(10)
was calculated (Table 1) The results revealed that the valuesof 119896119902were much greater than the maximum diffusion rate
constant of the biomolecule (2 times 1010Mminus1 sminus1) which indi-cated that the fluorescence quenching of HSA was initiatedby complex formation between HSA and isoimperatorinconfirming that the mechanism of quenching is static [19]
323 Determination of Association Constant (119870119887) and Num-
ber of Binding Sites (119899) The values of association constant119870
119887 and the number of binding sites 119899 at different temper-
atures are obtained from the modified Stern-Volmer plots(Figure 5) and listed in Table 1 It can be found from thesedata that the 119899 values at different temperatures are keptaround unity which shows the existence of one binding sitein HSA for isoimperatorin The value of 119870
119887is significant to
understand the distribution of the drug in plasma since theweak binding can lead to a short life time or poor distributionwhile strong binding can decrease the concentration of freedrug in plasma The value of 119870
119887illustrated that there is a
strong binding force between isoimperatorin and HSA Itwas found that the binding constant increased with increasein temperature resulting in an increment of the stabilityof the isoimperatorin-HSA complex it also implies thatisoimperatorin can be tightly stored and carried by HSA inthe body
324 Forces Involved in the Binding Process Consideringthe dependence of binding constant on temperature a ther-modynamic process was considered to be responsible forthe formation of a complex Therefore the thermodynamicparameters dependent on temperatures were analyzed Theacting forces between a small molecule and macromoleculemainly include hydrogen bonds van der Waals forces elec-trostatic forces and hydrophobic interaction forces [23] Inorder to obtain the information about the forces persisting in
minus15
minus13
minus11
minus09
minus07
minus05
minus03
minus01
01
03
05
minus68 minus65 minus62 minus59 minus56 minus53 minus5
log[isoimperatorin]
log[(F0minusF)F]
Figure 5 The modified Stern-Volmer plots for HSA fluorescencequenching by isoimperatorin at 298 (⧫) 303 (998771) 308 (◼) and 313 (∙)K For interpretation of references to color in this figure legend thereader is referred to the web version of this article
the present binding process the thermodynamic parametersof binding were obtained fromVanrsquot Hoff plot followed by theGibbs equation (data not shown) Table 2 shows the values ofΔ119867
∘ andΔ119878∘ obtained for the binding site from the slopes andordinates at the origin of the fitted lines From Table 2 it canbe seen that the negative sign for free energy (Δ119866∘) meansthat the interaction process is spontaneous The positiveenthalpy (Δ119867∘) and entropy (Δ119878∘) values of the interaction ofisoimperatorin and HSA indicate that the binding is mainlyentropy-driven and the enthalpy is unfavorable for it thehydrophobic forces play a major role in the reaction [43]
33 Nonradiative Energy Transfer from HSA to Isoimpera-torin The Forster nonradiative energy transfer (FRET) hasbeen used as a ldquospectroscopic rulerrdquo for measurement ofthe distance between the acceptor (ligand) and the donor(tryptophan residues in the protein) [44] According to thistheory there will be FRET between two molecules if thefollowing conditions were satisfied (a) the donor moleculeemits fluorescence (b) the emission spectrum of the donorhas an enough overlap with the absorption spectrum of
The Scientific World Journal 7
Table 2 Thermodynamic parameters for binding isoimperatorin to HSA
119879 (K) Δ119878
∘ (Jsdotmolminus1sdotKminus1) Δ119867
∘ (kJsdotmolminus1) Δ119866
∘ (kJsdotmolminus1)298 minus2969 plusmn 009
303 17820 plusmn 011 2993 plusmn 013 minus3071 plusmn 007
308 minus3169 plusmn 010
313 minus3299 plusmn 008
0
50
100
150
200
250
300
350
400
450
222 272 322 372 422Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
0
002
004
006
008
01
012
014
Abso
rban
ce
a
b
Figure 6 The overlap of the fluorescence spectrum of HSA (a)and the absorption spectrum of isoimperatorin (b) when [HSA] =[isoimperatorin] = 10 120583MThe enzyme was excited at 295 nm
the acceptor and (c) the donor and acceptor molecules areclose enough with the maximum distance not to exceeding7 nm [45]
There is a good overlapping between the fluorescenceemission spectrum of HSA and absorption spectra of isoim-peratorin (Figure 6) As the fluorescence emission of proteinwas affected by the excitation light around 295 nm thespectrum ranging from 300ndash400 nm was chosen to calculatethe overlapping integral The value of 119869 was calculated to be473 plusmn 005 times 10
minus15 cm3 Lmolminus1 by integrating the overlapspectra for 300ndash400 nm In the present case 1198702 = 23119873 = 136 and Φ = 0118 [21] according to (6) and (7) wecould calculate that 119864 = 045 plusmn 006 119877
0= 322 plusmn 03 nm and
119903 = 29 plusmn 02 nm As mentioned earlier (Section 321) HSAposses one tryptophan residue (Trp 214) and the calculated119903 is actually the average value between the bound drug andTrp 214 Since 119903 is much smaller than 7 nm and 05119877
0lt
119903 lt 15119877
0[41] therefore the energy transfer from HSA
to isoimperatorin occurs with high probability which alsoexplains the efficient quenching of the HSA fluorescence
34 PSH Determination in the Absence and Presence of Isoim-peratorin Several methods exist for measuring the surfacehydrophobicity of proteins that ANS experiment is one ofthem ANS as an extrinsic fluorescent probe is extremelysensitive to polarity of the solvent In aqueous solutions itfluoresces very weakly but upon binding to hydrophobicpatches of proteins its quantum yield increases significantly[29] Thus ANS fluorescence may be applied for monitoringpossible changes in protein surface hydrophobicity induced
0
200
400
600
800
1000
0 10 20 30 40 50
Fluo
resc
ence
inte
nsity
(au
)
[ANS] (120583M)
Figure 7 Binding of ANS to HSA (1120583M) in the absence (∙) andpresence (998771) of 3120583M of isoimperatorin
upon drug binding At a fixed concentration (1 120583M) of HSAand increasing concentrations of ANS (0ndash50120583M) fluores-cence intensity of ANS was measured in the absence andpresence of 3120583M isoimperatorin (Figure 7)
A typical hyperbolic response is observed which demon-strates the saturation nature of ANS binding toHSA It can beseen that the hyperbolic response of ANS fluorescence in thepresence of isoimperatorin is slightly different in comparisonto the absence of the drug The values of 119865max and 119870
app119889
forANS are obtained from the Scatchard plot (Figure 8) andlisted in Table 3 The value of 119870app
119889
for ANS binding in theabsence of drug (09 plusmn 005) was increased in the presenceof 3 120583M isoimperatorin (14 plusmn 007) which shows the weakerbinding of ANS to the HSA-isoimperatorin complex ThenPSH was calculated in the absence and the presence of3 120583M isoimperatorin by using 119865max and119870
app119889
values (Table 3)Decreasing the PSH index of HSA (from 150794 plusmn 1146 to80612 plusmn 007) suggests that the surface hydrophobicity ofHSA is decreased by sim465 upon drug binding
35 Synchronous Fluorescence Spectroscopy Synchronous flu-orescence spectroscopy introduced by Lody has been usedto characterize complex mixtures which can provide finger-prints of complex samples [46] The synchronous fluores-cence spectra can provide information about the molecularenvironment in the vicinity of the chromophore moleculesIn the synchronous fluorescence spectra the sensitivityassociated with fluorescence is maintained while several
8 The Scientific World Journal
Table 3 Surface hydrophobicity parameters for HSA in the absence and the presence of isoimperatorin
Parameters HSA HSA-IIM complex119870
app119889
(120583M) 09 plusmn 005
lowast
14 plusmn 007
lowast
119865max[HSA] 135714 plusmn 85
lowastlowast
112853 plusmn 63
lowastlowast
Surface hydrophobicity index (119865max[HSA] 119870app119889
) 150794 plusmn 1146
lowastlowast
80612 plusmn 007
lowastlowast
Data are expressed as mean plusmn SD of three measurements lowast119875 value lt 00006 lowastlowast119875 value lt 00001
0
50
100
150
200
250
300
350
400
400 500 600 700 800 900 1000Fluorescence intensity (au)
Fmax
Fluo
resc
ence
inte
nsity
(au
)[A
NS]
free
Slope = minus1Kappd
Figure 8 Scatchard plots for titration of HSA with increasingconcentrations of ANS in the absence (∙) and presence (998771) of 3 120583Mof isoimperatorin 119865max and 1119870
app119889
values were obtained from the119909-intercept and slope of this plot For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
advantages are available spectral simplification spectralbandwidth reduction and avoidance of different perturbingeffects The fluorescence spectrum of BSAHSA mainly dueto the Trp residues is sensitive to the microenvironment ofthese chromophoreswith themaximumemissionwavelengthbeing very useful in estimating the hydrophobicity aroundthe tryptophan residues It is well known that the experimenton synchronous fluorescence of HSA will provide the char-acteristic information for the Tyr and Trp residues when thescanning interval Δ120582 is fixed at 15 and 60 nm respectively[47] The shift in the position of fluorescence emissionmaximum corresponds to changes of the polarity aroundthe chromophore molecule A blue shift of 120582max means thatthe amino acid residues are located in a more hydrophobicenvironment and are less exposed to the solvent while a redshift of 120582max implies that the amino acid residues are in apolar environment and are more exposed to the solvent [48]The synchronous fluorescence spectra of interaction betweenisoimperatorin and HSA are presented in Figure 8 It wasshown that the fluorescence intensity of HSA decreased regu-larly along with the addition of isoimperatorin which furtherdemonstrated the occurrence of fluorescence quenching inthe binding process Moreover there is no significant shiftof the maximum emission wavelength with Δ120582 = 15 nm or60 nm (Figures 9(a) and 9(b)) which implies that interaction
of isoimperatorin with HSA does not affect the conformationof the region around the Tyr or Trp residues
As well we can obtain the information about the locationof isoimperatorin binding site from the synchronous fluores-cence dataThe ratios of synchronous fluorescence quenching(RSFQ) were used to determine the binding sites of the drugsto the HSA molecule in drug-HSA complexes The RSFQvalues were calculated from the following equation
RSFQ = 1 minus
119865
119865
0
(11)
where 119865 and 1198650are the synchronous fluorescence intensities
of HSA in the presence and the absence of drug respectivelyIt was obvious from Figure 9(c) that the RSFQ results forΔ120582 = 60 nmwere slightly higher than the corresponding onesfor Δ120582 = 15 nm As illustrated in Figure 9(c) addition ofisoimperatorin resulted in strong fluorescence quenching ofTrp residue (sim47 reduction in fluorescence intensity) con-firming that isoimperatorin reached subdomain IIA whereonly one Trp residue (Trp 214) in HSA is located It wasalso observed from Figure 9(c) that the fluorescence strengthof Tyr residues decreased about sim32 in the presence ofisoimperatorin So the binding of isoimperatorin to HSAquenched flourescence intensity of Tyr and Trp Since Trp214 (the only Trp of HSA) is at site I and Tyr 263 (hasthe strong fluorescence in the case of Tyr residues andmay be one of the residues responsible for the synchronousfluorescence spectrum of HSA when Δ120582 is fixed at 15 nm) isin subdomain IIA (site I) it can be concluded that Trp andTyr residues of HSA may have equal accessibility to bindingsite of isoimperatorin [49] The above results indicated thatisoimperatorin can bind to HSA in the hydrophobic cavity(site I) on subdomain IIA which are in full agreement withquenching competitive binding experiments and theoreticaldocking technique
36 Site Marker Competitive Binding Experiments In orderto further classify drug binding site on HSA competitivebinding experiments have been carried out Warfarin andibuprofen are two markers specific for binding to HSA thatbind to sites I and II which are corresponded to the sub-domains IIA and IIIA respectively Therefore to determinethe location of the isoimperatorin binding site on HSA thecompetitive displacement experimentswere carried out usingwarfarin as a characteristicmarker for site I (Figure 10(a)) andibuprofen for site II (Figure 10(b)) [24 31] In the site markercompetitive experiment isoimperatorin was gradually addedto the solution of HSA HSA-ibuprofen or HSA-warfarincomplex and then fluorescence intensity of system wasrecorded
The Scientific World Journal 9
0
100
200
300
400
500
600
265 270 275 280 285 290 295 300Wavelength (nm)
1
8
Fluo
resc
ence
inte
nsity
(au
)
(a)
Fluo
resc
ence
inte
nsity
(au
)
0
50
100
150
200
250
300
350
400
250 260 270 280 290 300Wavelength (nm)
1
8
(b)
0
10
20
30
40
50
0 05 1 15 2 25 3
RSFQ
()
Isoimperatorin (120583M)
0
02
04
06
08
1
12
005 1 15 2 25 3 35
Isoimperatorin (120583M)
F0
F
(c)
Figure 9 Synchronous fluorescence spectra of interaction between HSA and isoimperatorin (a) at Δ120582 = 15 nm and (b) at Δ120582 = 60 nmConcentration ofHSAwas 30 120583Mwhile concentrations of isoimperatorinwere 0 04 08 12 16 2 25 and 3 120583Mfrom 1 to 8 (c) Comparativeevaluation of isoimperatorin effect on the RSFQ of HSA [HSA] = 3 120583M (◼) Δ120582 = 15 nm and (∙) Δ120582 = 60 nm
As shown in Figure 10(a) with addition of warfarin in theHSA solution the flourescence intensity was slightly higherthan that of without warfarin (and including the red shift)Then after adding the isoimperatorin into the above systemthe flourescence intensity of HSA solution with warfarinheld in equimolar decreased gradually and the intensitywas much lower than that of without warfarin displayingthat the binding of the isoimperatorin to HSA was affectedafter adding warfarin On the contrary in the presence ofibuprofen the flourescence intensity of the HSA-ibuprofencomplex almost had no difference from that recordedwithoutibuprofen under the same conditions (Figure 10(b)) whichindicated that site II marker did not prevent the binding ofisoimperatorin in its usual binding location The next stepprotein intrinsic fluorescence was measured and the relativefluorescence intensities (119865119865
0) versus ligand concentration
([isoimperatorin]) plots are displayed in Figure 10(c) Asshown in Table 4 the binding constant was remarkablydecreased after addition of warfarin while the addition ofibuprofen results in only a small difference These resultsindicate that warfarin can displace the isoimperatorin but
ibuprofen has little effect on the binding of isoimperatorinto HSA In other words these results suggest that isoim-peratorin probably competes with warfarin for binding toHSA [24] Experimental observations were followed up withdocking studies where isoimperatorin was docked to HSAto determine the preferred binding site on the proteinAs described above the 3D crystal structure of HSA is amonomer consisting of three homologous domains whichassemble to form a heart-shaped molecule [4 6] Each ofthe structurally similar 120572-helix domains (IndashIII) has twosubdomains (A andB) with six120572-helices in subdomainA andfour 120572-helices in subdomain BThe crystal structure analysesindicate that the principal regions of ligand binding sitesin HSA are located in hydrophobic cavities in subdomainsIIA and IIIA which are consistent with Sudlowrsquos site I andSudlowrsquos site II respectively Several studies have shown thatHSA is able to bind many ligands in several binding sites[38] In this paper the molecular docking model of theHSAndashisoimperatorin complex was conducted on the basisof the crystal structure of HSA to find the binding positionof isoimperatorin The best docking energy result is shown
10 The Scientific World Journal
0
100
200
300
400
500
600
310 335 360 385 410Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA1
8
(a)
0
100
200
300
400
500
600
700
305 325 345 365 385 405Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA
1
8
(b)
0
05
1
15
0 1 2 3
HSAIbuprofen
Warfarin
Isoimperatorin (120583M)
F0
F
(c)
Figure 10 Effect of isoimperatorin on the quenching of HSA-warfarin complex (a) andHSA-ibuprofen complex (b) fluorescence From 1 to 8[HSA] = [warfarin or iboprufen] = 3 120583M [isoimperatorin] = 0 04 08 12 16 2 25 and 3 120583M All reproduced data shown are representativeof at least 3 independent experiments (c) Fluorescence quenching profiles of HSA (⧫) HSA-ibobrofen (◼) and HSA-warfarin (998771) in thepresence of various concentration of isoimperatorin
in Figure 11 It can be seen that isoimperatorin was situatedwithin subdomain IIA in Sudlowrsquos site I formed by six 120572-helices This figure shows the location of Trp 214 and also thebinding of isoimperatorin in HSA As this fluorescent residueis in subdomain IIA and due to the proximity of the bindingsite of isoimperatorin to the Trp 214 it can be concluded thatisoimperatorin has one reactive site onHSAandprobably thissite is at or near the site I The above docking results are ingood agreement with the displacement experiments results
and revealed that isoimperatorin binds to Sudlowrsquos site I ofHSA
37 Effect of Isoimperatorin on HSA Esterase Activity Pre-liminary studies showed that albumin in addition to itsligand binding capabilities has remarkable promiscuouscatalytic activities toward a broad range of organicmoleculesincluding esters amides phosphates and benzisoxazolesHuman serum albumin had significantly greater esterase-like
The Scientific World Journal 11
Table 4 Binding constants for isoimperatorin-HSA interaction (at 295K) in the absence and presence of the site marker
119870
119887
times 10
minus5
(Mminus1) 119877
a
HSA + isoimperatorin (in the absence ofibuprofen and warfarin) 1110 plusmn 04
lowast 09947
HSA + isoimperatorin + ibuprofen 1099 plusmn 06
lowast 09904HSA + isoimperatorin + warfarin 789 plusmn 03
lowast 09881Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870
119887values lowast119875 value lt 00002
Trp 214
Isoimperatorin
Figure 11 Crystal structure of HSA and the positions of singletryptophan residue (Trp 214) and also the binding site of isoim-peratorin in HSA Protein backbone is shown in the ldquocartoonrdquorepresentation and isoimperatorinand Trp 214 residue in the site Iare shown in the ldquostickrdquo representation The isoimperatorin and Trp214 structures are represented by the red and blue colorsThe crystalstructure of protein was obtained from Protein Data Bank (1BM0)For interpretation of references to color in this figure legend thereader is referred to the web version of this paper
activity than bovine serum albumin and defatted albuminwas more active than untreated albumin [50] InterestinglyHSA exhibits esterase-like activity and hydrolyzes drugshaving an ester group such as aspirin (which can convertaspirin ldquoacetylsalicylic acidrdquo to salicylic acid) Lys199 Lys402Lys519 and Lys545 form the pseudoesterase active site ofHSAthat is located in site I [50] ldquoEsterase-likerdquo activity (hydrolysisof p-nitrophenol esters such as p-NPA) by subdomain III Ahas been also reported and site directed mutagenesis studieshave shown that Arg 410 and Tyr 411 are essential in thisesterase activity However further investigation revealed thatacetylation is not limited to a distinct side chain so that theHSA incubation with high concentrations of pNPAc resultsin the acetylation of 59 Lys 10 Ser 8 Thr 4 Tyr and 1 Asp[51] Because of the above difference here we set out toevaluate the effect of isoimperatorin on the esterase activityof HSA based on our previous work [19] The kinetics ofthe pseudoenzymatic hydrolysis of pNPAc was obtained inthe absence and presence of isoimperatorin and site markers(Figure 12) Accordingly warfarin and indomethacin (as siteI marker) and isoimperatorin inhibited competitively theHSA-catalyzed hydrolysis of pNPAc by binding to site I Incontrast ibuprofen and tetracycline were unable to inhibitesterase activity of HSA The enzyme inhibition results sug-gested that isoimperatorin aspirin warfarin indomethacin
0
005
01
015
02
025
03
035
0 05 1 15 2 25 3Time (min)
HSA TetracyclineIbuprofenAspirin
IndomethacinOstholeWarfarinIsoimperatorin
A400
Figure 12 4-Nitrophenyl acetate esterase activity of HSA in theabsence and presence of tetracycline and ibuprofen (site II) war-farin aspirin indomethacin and osthole (site I) and isoimperatorin(site I) Final concentration of protein pNPA and drugs were 15250 and 250 120583M respectively For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
and osthole binding sites on the HSA molecule are the sameThese results also proposed that the large site I on HSAparticipates effectively to its promiscuous esterase activity
4 Conclusion
The accumulated data on HSA-drug interactions can beused to develop quantitative structure-activity relationshipsfor albumin binding However as stated earlier adsorptiondistribution metabolism and elimination (ADME) is con-trolled by albumin [30] Poor or strong binding to SA isbelieved to affect half-time of drugs [35] and their dosagedepends on their half-life in plasma Thus the bindingaffinity of any drug to SA is one of the major factors thatdetermine the pharmacokinetics halftime and availabilityof the drug in various tissues (see [30] and referencestherein) Further complexities arise in vivo due to interactions
12 The Scientific World Journal
between drugs and endogenous ligands for HSA This isparticularly pertinent for fatty acids which normally occurin serum at levels between 01 and 2mol per mol of HSAand can both compete and cooperate with drugs bindingto the protein In certain disease states these effects areexacerbated as the fatty acid HSA molar ratio may be ashigh as six Other pathological conditions are associatedwith high (micromolar to millimolar) levels of bilirubinhemin or renal toxins (eg 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) indoxyl sulphate) which bindto the protein causing significant drug binding defects
Our major aim in this study was to distinguish thenature of the interaction of isoimperatorin to HSA usingdifferent biophysical techniques This study could be veryvaluable due to importance and presence of isoimperatorinin human diet (food-drug interaction on HSA) The resultsshowed that isoimperatorin quenched the intrinsic fluo-rescence of HSA through dynamic quenching mechanismFurthermore hydrophobic interaction played a major role inthe binding process Using Fluorescence and UV absorptionspectroscopy it has been shown that isoimperatorin inducesprotein structural changes The binding average distancebetween the solo tryptophan of the protein and the boundisoimperatorin was calculated by Forster theory as 290 plusmn02 nmAnalysis of PSHvalues elucidated that isoimperatorinhas induced some compactness in the tertiary structure ofthe enzyme The results of synchronous flourescence onHSA-isoimperatorin system elucidated that the drug binds tothe hydrophobic cavity (site I) on subdomain IIA that wasconfirmed by quenching experiments competitive bindingexperiments and docking technique
Acknowledgment
The authors gratefully acknowledge the Kermanshah Uni-versity of Medical Sciences Research Council for financialsupport of this work
References
[1] Y-J Hu Y Liu T-Q Sun A-M Bai J-Q Lu and Z-B Pi ldquoBinding of anti-inflammatory drug cromolyn sodiumto bovine serum albuminrdquo International Journal of BiologicalMacromolecules vol 39 no 4-5 pp 280ndash285 2006
[2] N Seedher and S Bhatia ldquoReversible binding of celecoxiband valdecoxib with human serum albumin using fluorescencespectroscopic techniquerdquo Pharmacological Research vol 54 no2 pp 77ndash84 2006
[3] H Isogai and N Hirayama ldquoIn silico prediction of interactionsbetween site II on human serum albumin and profen drugsrdquoISRN Pharma vol 2013 Article ID 818364 8 pages 2013
[4] YWangHYuX Shi Z LuoD Lin andMHuang ldquoStructuralmechanism of ring opening reaction of glucose by humanserum albuminrdquo The Journal of Biological Chemistry vol 288pp 15980ndash15987 2013
[5] F Keshavarz M M Alavianmehr and R Yousefi ldquoMolecularinteraction of benzalkonium Ibuprofenate and its discreteingredients with human serum albuminrdquo Physical ChemistryResearch vol 1 no 2 pp 111ndash116 2013
[6] T K Maiti K S Ghosh J Debnath and S Dasgupta ldquoBindingof all-trans retinoic acid to human serum albumin fluores-cence FT-IR and circular dichroism studiesrdquo InternationalJournal of Biological Macromolecules vol 38 no 3ndash5 pp 197ndash202 2006
[7] M-X XieM Long Y Liu C Qin and Y-DWang ldquoCharacter-ization of the interaction between human serum albumin andmorinrdquo Biochimica et Biophysica Acta vol 1760 no 8 pp 1184ndash1191 2006
[8] CD Kanakis P A TarantilisMG Polissiou S Diamantoglouand H A Tajmir-Riahi ldquoAntioxidant flavonoids bind humanserum albuminrdquo Journal of Molecular Structure vol 798 no 1-3 pp 69ndash74 2006
[9] S Marumoto and M Miyazawa ldquoStructure-activity rela-tionships for naturally occurring coumarins as 120573-secretaseinhibitorrdquo Bioorganic and Medicinal Chemistry vol 20 no 2pp 784ndash788 2012
[10] S Marumoto and M Miyazawa ldquoBiotransformation of isoim-peratorin and imperatorin by Glomerella cingulata and 120573-secretase inhibitory activityrdquo Bioorganic and Medicinal Chem-istry vol 18 no 1 pp 455ndash459 2010
[11] S E Sajjadi Y Shokoohinia and S Hemmati ldquoIsolation andidentification of furanocoumarins and a phenylpropanoid fromthe acetone extract and identification of volatile constituentsfrom the essential oil of Peucedanum pastinacifoliumrdquo Chem-istry of Natural Compounds vol 48 no 4 pp 668ndash671 2012
[12] X B Yang J Hou D Liu et al ldquoBiotransformation of isoim-peratorin by Cunninghamella blakesleana AS 3970rdquo Journal ofMolecular Catalysis B vol 88 pp 1ndash6 2013
[13] W Lili S Yehong S Qi et al ldquoIn vitro permeability analy-sis pharmacokinetic and brain distribution study in mice ofimperatorin isoimperatorin and cnidilin in Radix AngelicaeDahuricaerdquo Fitoterapia vol 85 pp 144ndash153 2013
[14] S Wang Q Chen and L He ldquoDevelopment and validationof a gas chromatography-mass spectrometry method for thedetermination of isoimperatorin in rat plasma and tissueapplication to the pharmacokinetic and tissue distributionstudyrdquo Journal of Chromatography B vol 852 no 1-2 pp 473ndash478 2007
[15] H Sadraei Y Shokoohinia S E Sajjadi and M MozafarildquoAntispasmodic effects of Prangos ferulacea acetone extract andits maincomponent osthole on ileum contractionrdquo Research inPharmaceutical Sciences vol 8 pp 137ndash144 2013
[16] H Sadraei Y Shokoohinia S E Sajjadi and B GhadirianldquoAntispasmodic effect of osthole and Prangos ferulacea extracton rat uterus smooth muscle motilityrdquo Research in Pharmaceu-tical Sciences vol 7 pp 141ndash149 2012
[17] S Sajjadi Y Shokoohinia S Gholamzadeh M Behbahani andA Fattahi ldquoAntiviral evaluation of coumarins from Prangosferulacea L (Lindl)rdquo Research in Pharmaceutical Sciences vol7 p 783 2012
[18] S E Sajjadi Y Shokoohinia and S Gholamzadeh ldquoChemicalcomposition of the essential oil of the root of Prangos ferulacea(L) Lindlrdquo Chemija vol 22 pp 178ndash180 2011
[19] N Bijari Y Shokoohinia M-R Ashrafi-Kooshk et al ldquoSpec-troscopic study of interaction between osthole and humanserum albumin identification of possible binding site of thecompoundrdquo Journal of Luminescence vol 143 pp 328ndash336 2013
[20] X Shi M Liu M Zhang et al ldquoIdentification of in vitro and invivo metabolites of isoimperatorin using liquid chromatogra-phymass spectrometryrdquo Food Chemistry vol 141 pp 357ndash3652013
The Scientific World Journal 13
[21] T Chatterjee A Pal S Dey B-K Chatterjee and PChakrabarti ldquoInteraction of virstatin with human serum albu-min spectroscopic analysis andmolecular modelingrdquo Plos Onevol 7 pp 1ndash12 2012
[22] Y-M Chen and L-H Guo ldquoCombined fluorescence and elec-trochemical investigation on the binding interaction betweenorganic acid and human serum albuminrdquo Journal of Environ-mental Sciences vol 21 no 3 pp 373ndash379 2009
[23] J R Lakowicz Principles of Fluorescence Spectroscopy KluwerAcademicPlenum Publishers New York NY USA 2nd edi-tion 1999
[24] F Ding N Li B Han F Liu L Zhang and Y Sun ldquoThe bindingof CI Acid Red 2 to human serum albumin determinationof binding mechanism and binding site using fluorescencespectroscopyrdquo Dyes and Pigments vol 83 no 2 pp 249ndash2572009
[25] M-R Ashrafi Kooshk K Mansouri M Nadi S Khodarahmiand R Khodarahmi ldquoIn vitro anti-cancer activity of nativecurcumin and protein-curcuminrdquo Systems vol 2 no 1 pp 80ndash88 1981
[26] P D Ross and S Subramanian ldquoThermodynamics of proteinassociation reactions forces contributing to stabilityrdquo Biochem-istry vol 20 no 11 pp 3096ndash3102 1981
[27] J Liu J-N Tian J Zhang Z Hu and X Chen ldquoInteractionof magnolol with bovine serum albumin a fluorescence-quenching studyrdquo Analytical and Bioanalytical Chemistry vol376 no 6 pp 864ndash867 2003
[28] A Coi A M Bianucci F Bonomi P Rasmussen G M Muraand M L Ganadu ldquoStructural perturbation of 120572B-crystallinby zinc and temperature related to its chaperone-like activityrdquoInternational Journal of Biological Macromolecules vol 42 no3 pp 229ndash234 2008
[29] M Moller and A Denicola ldquoStudy of protein-ligand bindingby fluorescencerdquoBiochemistry andMolecular Biology Educationvol 30 no 5 pp 309ndash312 2002
[30] R Khodarahmi S A Karimi M R Ashrafi Kooshk S AGhadami S Ghobadi and M Amani ldquoComparative spec-troscopic studies on drug binding characteristics and proteinsurface hydrophobicity of native and modified forms of bovineserum albumin possible relevance to change in protein struc-turefunction upon non-enzymatic glycationrdquo SpectrochimicaActa A vol 89 pp 177ndash186 2012
[31] S-F Ma M Anraku Y Iwao et al ldquoHydrolysis of angiotensinII receptor blocker prodrug olmesartan medoxomil by humanserum albumin and identification of its catalytic active sitesrdquoDrug Metabolism and Disposition vol 33 no 12 pp 1911ndash19192005
[32] H M Berman J Westbrook Z Feng et al ldquoThe protein databankrdquo Nucleic Acids Research vol 28 no 1 pp 235ndash242 2000
[33] S Sugio A Kashima SMochizukiMNoda andKKobayashildquoCrystal structure of human serum albumin at 25 A resolutionrdquoProtein Engineering vol 12 no 6 pp 439ndash446 1999
[34] G M Morris D S Goodsell R Huey and A J OlsonldquoDistributed automated docking of flexible ligands to proteinsparallel applications of AutoDock 24rdquo Journal of Computer-Aided Molecular Design vol 10 no 4 pp 293ndash304 1996
[35] F Faridbod M R Ganjali B Larijani et al ldquoInteraction studyof pioglitazone with albumin by fluorescence spectroscopy andmolecular dockingrdquo Spectrochimica Acta A vol 78 no 1 pp96ndash101 2011
[36] D SGoodsell andA JOlson ldquoAutomated docking of substratesto proteins by simulated annealingrdquo Proteins vol 8 no 3 pp195ndash202 1990
[37] D S Goodsell G M Morris and A J Olson ldquoAutomateddocking of flexible ligands applications of AutoDockrdquo Journalof Molecular Recognition vol 9 no 1 pp 1ndash5 1996
[38] G Sudlow D J Birkett and D N Wade ldquoThe characterizationof two specific drug binding sites on human serum albuminrdquoMolecular Pharmacology vol 11 no 6 pp 824ndash832 1975
[39] Y Yue X Chen J Qin and X Yao ldquoCharacterization of themangiferin-human serum albumin complex by spectroscopicandmolecularmodeling approachesrdquo Journal of Pharmaceuticaland Biomedical Analysis vol 49 no 3 pp 753ndash759 2009
[40] A K Shaw and S K Pal ldquoSpectroscopic studies on the effectof temperature on pH-induced folded states of human serumalbuminrdquo Journal of Photochemistry and Photobiology B vol 90no 1 pp 69ndash77 2008
[41] S Ranjbar S Ghobadi R Khodarahmi and H Nemati ldquoSpec-troscopic characterization of furosemide binding to humancarbonic anhydrase IIrdquo International Journal of BiologicalMacromolecules vol 50 no 4 pp 910ndash917 2012
[42] HNHou Z POi YWOYung F L Liao Y Zhang andY LiyldquoStudies on interaction between vitamin B12 and human serumalbuminrdquo Journal of Pharmaceutical and Biomedical Analysisvol 47 pp 134ndash139 2008
[43] S S Kalanur J Seetharamappa and V K A Kalalbandi ldquoChar-acterization of interaction and the effect of carbamazepine onthe structure of human serum albuminrdquo Journal of Pharmaceu-tical and Biomedical Analysis vol 53 no 3 pp 660ndash666 2010
[44] L Stryer ldquoFluorescence spectroscopy of proteinsrdquo Science vol162 no 3853 pp 526ndash533 1968
[45] B Valeur and J C Brochon New Trends in FluorescenceSpectroscopy Springer Berlin Germany 1999
[46] V Suryawanshi P Anbhule A Gore Sh Patil and G AndKolekar ldquoA spectral deciphering the perturbation of modeltransporter protein (HSA) by antibacterial pyrimidine deriva-tive pharmacokinetic and biophysical insightsrdquo Journal ofPhotochemistry and Photobiology B vol 118 pp 1ndash8 2013
[47] T Yuan AMWeljie andH J Vogel ldquoTryptophan fluorescencequenching by methionine and selenomethionine residues ofcalmodulin orientation of peptide and protein bindingrdquo Bio-chemistry vol 37 no 9 pp 3187ndash3195 1998
[48] TCOrsquoHaver A F Fell G Smith et al ldquoDerivative spectroscopyand its applications in analysisrdquo Analytical Proceedings vol 19no 1 pp 22ndash46 1982
[49] Y-Q Wang B-P Tang H-M Zhang Q-H Zhou and G-C Zhang ldquoStudies on the interaction between imidaclopridand human serum albumin spectroscopic approachrdquo Journal ofPhotochemistry and Photobiology B vol 94 no 3 pp 183ndash1902009
[50] S-J Kim H-W Rhee H-Y Park H-S Kim and J-I HongldquoFluorescent probes designed for detecting human serum albu-min on the basis of its pseudo-esterase activityrdquo Bioorganic ampMedicinal Chemistry Letters vol 23 pp 2093ndash2097 2013
[51] P Ascenzi M Gioia G Fanali M Coletta and M FasanoldquoPseudo-enzymatic hydrolysis of 4-nitrophenyl acetate byhuman serum albumin pH-dependence of rates of individualstepsrdquo Biochemical and Biophysical Research Communicationsvol 424 pp 451ndash455 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
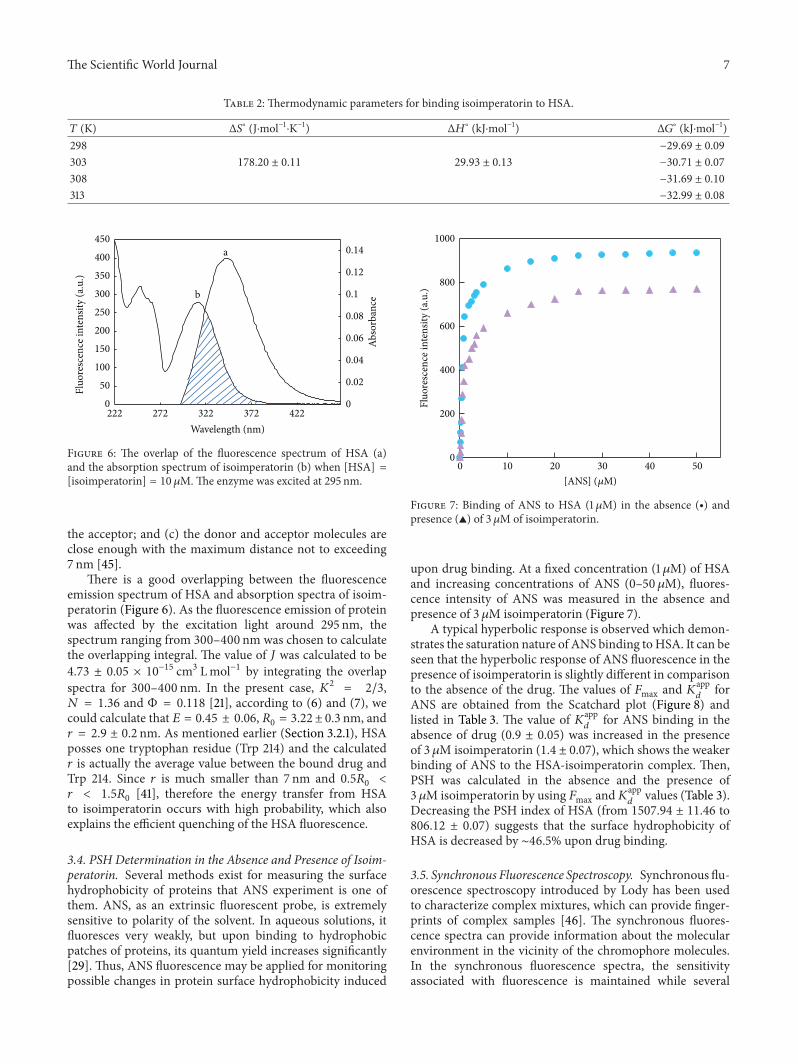
The Scientific World Journal 7
Table 2 Thermodynamic parameters for binding isoimperatorin to HSA
119879 (K) Δ119878
∘ (Jsdotmolminus1sdotKminus1) Δ119867
∘ (kJsdotmolminus1) Δ119866
∘ (kJsdotmolminus1)298 minus2969 plusmn 009
303 17820 plusmn 011 2993 plusmn 013 minus3071 plusmn 007
308 minus3169 plusmn 010
313 minus3299 plusmn 008
0
50
100
150
200
250
300
350
400
450
222 272 322 372 422Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
0
002
004
006
008
01
012
014
Abso
rban
ce
a
b
Figure 6 The overlap of the fluorescence spectrum of HSA (a)and the absorption spectrum of isoimperatorin (b) when [HSA] =[isoimperatorin] = 10 120583MThe enzyme was excited at 295 nm
the acceptor and (c) the donor and acceptor molecules areclose enough with the maximum distance not to exceeding7 nm [45]
There is a good overlapping between the fluorescenceemission spectrum of HSA and absorption spectra of isoim-peratorin (Figure 6) As the fluorescence emission of proteinwas affected by the excitation light around 295 nm thespectrum ranging from 300ndash400 nm was chosen to calculatethe overlapping integral The value of 119869 was calculated to be473 plusmn 005 times 10
minus15 cm3 Lmolminus1 by integrating the overlapspectra for 300ndash400 nm In the present case 1198702 = 23119873 = 136 and Φ = 0118 [21] according to (6) and (7) wecould calculate that 119864 = 045 plusmn 006 119877
0= 322 plusmn 03 nm and
119903 = 29 plusmn 02 nm As mentioned earlier (Section 321) HSAposses one tryptophan residue (Trp 214) and the calculated119903 is actually the average value between the bound drug andTrp 214 Since 119903 is much smaller than 7 nm and 05119877
0lt
119903 lt 15119877
0[41] therefore the energy transfer from HSA
to isoimperatorin occurs with high probability which alsoexplains the efficient quenching of the HSA fluorescence
34 PSH Determination in the Absence and Presence of Isoim-peratorin Several methods exist for measuring the surfacehydrophobicity of proteins that ANS experiment is one ofthem ANS as an extrinsic fluorescent probe is extremelysensitive to polarity of the solvent In aqueous solutions itfluoresces very weakly but upon binding to hydrophobicpatches of proteins its quantum yield increases significantly[29] Thus ANS fluorescence may be applied for monitoringpossible changes in protein surface hydrophobicity induced
0
200
400
600
800
1000
0 10 20 30 40 50
Fluo
resc
ence
inte
nsity
(au
)
[ANS] (120583M)
Figure 7 Binding of ANS to HSA (1120583M) in the absence (∙) andpresence (998771) of 3120583M of isoimperatorin
upon drug binding At a fixed concentration (1 120583M) of HSAand increasing concentrations of ANS (0ndash50120583M) fluores-cence intensity of ANS was measured in the absence andpresence of 3120583M isoimperatorin (Figure 7)
A typical hyperbolic response is observed which demon-strates the saturation nature of ANS binding toHSA It can beseen that the hyperbolic response of ANS fluorescence in thepresence of isoimperatorin is slightly different in comparisonto the absence of the drug The values of 119865max and 119870
app119889
forANS are obtained from the Scatchard plot (Figure 8) andlisted in Table 3 The value of 119870app
119889
for ANS binding in theabsence of drug (09 plusmn 005) was increased in the presenceof 3 120583M isoimperatorin (14 plusmn 007) which shows the weakerbinding of ANS to the HSA-isoimperatorin complex ThenPSH was calculated in the absence and the presence of3 120583M isoimperatorin by using 119865max and119870
app119889
values (Table 3)Decreasing the PSH index of HSA (from 150794 plusmn 1146 to80612 plusmn 007) suggests that the surface hydrophobicity ofHSA is decreased by sim465 upon drug binding
35 Synchronous Fluorescence Spectroscopy Synchronous flu-orescence spectroscopy introduced by Lody has been usedto characterize complex mixtures which can provide finger-prints of complex samples [46] The synchronous fluores-cence spectra can provide information about the molecularenvironment in the vicinity of the chromophore moleculesIn the synchronous fluorescence spectra the sensitivityassociated with fluorescence is maintained while several
8 The Scientific World Journal
Table 3 Surface hydrophobicity parameters for HSA in the absence and the presence of isoimperatorin
Parameters HSA HSA-IIM complex119870
app119889
(120583M) 09 plusmn 005
lowast
14 plusmn 007
lowast
119865max[HSA] 135714 plusmn 85
lowastlowast
112853 plusmn 63
lowastlowast
Surface hydrophobicity index (119865max[HSA] 119870app119889
) 150794 plusmn 1146
lowastlowast
80612 plusmn 007
lowastlowast
Data are expressed as mean plusmn SD of three measurements lowast119875 value lt 00006 lowastlowast119875 value lt 00001
0
50
100
150
200
250
300
350
400
400 500 600 700 800 900 1000Fluorescence intensity (au)
Fmax
Fluo
resc
ence
inte
nsity
(au
)[A
NS]
free
Slope = minus1Kappd
Figure 8 Scatchard plots for titration of HSA with increasingconcentrations of ANS in the absence (∙) and presence (998771) of 3 120583Mof isoimperatorin 119865max and 1119870
app119889
values were obtained from the119909-intercept and slope of this plot For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
advantages are available spectral simplification spectralbandwidth reduction and avoidance of different perturbingeffects The fluorescence spectrum of BSAHSA mainly dueto the Trp residues is sensitive to the microenvironment ofthese chromophoreswith themaximumemissionwavelengthbeing very useful in estimating the hydrophobicity aroundthe tryptophan residues It is well known that the experimenton synchronous fluorescence of HSA will provide the char-acteristic information for the Tyr and Trp residues when thescanning interval Δ120582 is fixed at 15 and 60 nm respectively[47] The shift in the position of fluorescence emissionmaximum corresponds to changes of the polarity aroundthe chromophore molecule A blue shift of 120582max means thatthe amino acid residues are located in a more hydrophobicenvironment and are less exposed to the solvent while a redshift of 120582max implies that the amino acid residues are in apolar environment and are more exposed to the solvent [48]The synchronous fluorescence spectra of interaction betweenisoimperatorin and HSA are presented in Figure 8 It wasshown that the fluorescence intensity of HSA decreased regu-larly along with the addition of isoimperatorin which furtherdemonstrated the occurrence of fluorescence quenching inthe binding process Moreover there is no significant shiftof the maximum emission wavelength with Δ120582 = 15 nm or60 nm (Figures 9(a) and 9(b)) which implies that interaction
of isoimperatorin with HSA does not affect the conformationof the region around the Tyr or Trp residues
As well we can obtain the information about the locationof isoimperatorin binding site from the synchronous fluores-cence dataThe ratios of synchronous fluorescence quenching(RSFQ) were used to determine the binding sites of the drugsto the HSA molecule in drug-HSA complexes The RSFQvalues were calculated from the following equation
RSFQ = 1 minus
119865
119865
0
(11)
where 119865 and 1198650are the synchronous fluorescence intensities
of HSA in the presence and the absence of drug respectivelyIt was obvious from Figure 9(c) that the RSFQ results forΔ120582 = 60 nmwere slightly higher than the corresponding onesfor Δ120582 = 15 nm As illustrated in Figure 9(c) addition ofisoimperatorin resulted in strong fluorescence quenching ofTrp residue (sim47 reduction in fluorescence intensity) con-firming that isoimperatorin reached subdomain IIA whereonly one Trp residue (Trp 214) in HSA is located It wasalso observed from Figure 9(c) that the fluorescence strengthof Tyr residues decreased about sim32 in the presence ofisoimperatorin So the binding of isoimperatorin to HSAquenched flourescence intensity of Tyr and Trp Since Trp214 (the only Trp of HSA) is at site I and Tyr 263 (hasthe strong fluorescence in the case of Tyr residues andmay be one of the residues responsible for the synchronousfluorescence spectrum of HSA when Δ120582 is fixed at 15 nm) isin subdomain IIA (site I) it can be concluded that Trp andTyr residues of HSA may have equal accessibility to bindingsite of isoimperatorin [49] The above results indicated thatisoimperatorin can bind to HSA in the hydrophobic cavity(site I) on subdomain IIA which are in full agreement withquenching competitive binding experiments and theoreticaldocking technique
36 Site Marker Competitive Binding Experiments In orderto further classify drug binding site on HSA competitivebinding experiments have been carried out Warfarin andibuprofen are two markers specific for binding to HSA thatbind to sites I and II which are corresponded to the sub-domains IIA and IIIA respectively Therefore to determinethe location of the isoimperatorin binding site on HSA thecompetitive displacement experimentswere carried out usingwarfarin as a characteristicmarker for site I (Figure 10(a)) andibuprofen for site II (Figure 10(b)) [24 31] In the site markercompetitive experiment isoimperatorin was gradually addedto the solution of HSA HSA-ibuprofen or HSA-warfarincomplex and then fluorescence intensity of system wasrecorded
The Scientific World Journal 9
0
100
200
300
400
500
600
265 270 275 280 285 290 295 300Wavelength (nm)
1
8
Fluo
resc
ence
inte
nsity
(au
)
(a)
Fluo
resc
ence
inte
nsity
(au
)
0
50
100
150
200
250
300
350
400
250 260 270 280 290 300Wavelength (nm)
1
8
(b)
0
10
20
30
40
50
0 05 1 15 2 25 3
RSFQ
()
Isoimperatorin (120583M)
0
02
04
06
08
1
12
005 1 15 2 25 3 35
Isoimperatorin (120583M)
F0
F
(c)
Figure 9 Synchronous fluorescence spectra of interaction between HSA and isoimperatorin (a) at Δ120582 = 15 nm and (b) at Δ120582 = 60 nmConcentration ofHSAwas 30 120583Mwhile concentrations of isoimperatorinwere 0 04 08 12 16 2 25 and 3 120583Mfrom 1 to 8 (c) Comparativeevaluation of isoimperatorin effect on the RSFQ of HSA [HSA] = 3 120583M (◼) Δ120582 = 15 nm and (∙) Δ120582 = 60 nm
As shown in Figure 10(a) with addition of warfarin in theHSA solution the flourescence intensity was slightly higherthan that of without warfarin (and including the red shift)Then after adding the isoimperatorin into the above systemthe flourescence intensity of HSA solution with warfarinheld in equimolar decreased gradually and the intensitywas much lower than that of without warfarin displayingthat the binding of the isoimperatorin to HSA was affectedafter adding warfarin On the contrary in the presence ofibuprofen the flourescence intensity of the HSA-ibuprofencomplex almost had no difference from that recordedwithoutibuprofen under the same conditions (Figure 10(b)) whichindicated that site II marker did not prevent the binding ofisoimperatorin in its usual binding location The next stepprotein intrinsic fluorescence was measured and the relativefluorescence intensities (119865119865
0) versus ligand concentration
([isoimperatorin]) plots are displayed in Figure 10(c) Asshown in Table 4 the binding constant was remarkablydecreased after addition of warfarin while the addition ofibuprofen results in only a small difference These resultsindicate that warfarin can displace the isoimperatorin but
ibuprofen has little effect on the binding of isoimperatorinto HSA In other words these results suggest that isoim-peratorin probably competes with warfarin for binding toHSA [24] Experimental observations were followed up withdocking studies where isoimperatorin was docked to HSAto determine the preferred binding site on the proteinAs described above the 3D crystal structure of HSA is amonomer consisting of three homologous domains whichassemble to form a heart-shaped molecule [4 6] Each ofthe structurally similar 120572-helix domains (IndashIII) has twosubdomains (A andB) with six120572-helices in subdomainA andfour 120572-helices in subdomain BThe crystal structure analysesindicate that the principal regions of ligand binding sitesin HSA are located in hydrophobic cavities in subdomainsIIA and IIIA which are consistent with Sudlowrsquos site I andSudlowrsquos site II respectively Several studies have shown thatHSA is able to bind many ligands in several binding sites[38] In this paper the molecular docking model of theHSAndashisoimperatorin complex was conducted on the basisof the crystal structure of HSA to find the binding positionof isoimperatorin The best docking energy result is shown
10 The Scientific World Journal
0
100
200
300
400
500
600
310 335 360 385 410Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA1
8
(a)
0
100
200
300
400
500
600
700
305 325 345 365 385 405Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA
1
8
(b)
0
05
1
15
0 1 2 3
HSAIbuprofen
Warfarin
Isoimperatorin (120583M)
F0
F
(c)
Figure 10 Effect of isoimperatorin on the quenching of HSA-warfarin complex (a) andHSA-ibuprofen complex (b) fluorescence From 1 to 8[HSA] = [warfarin or iboprufen] = 3 120583M [isoimperatorin] = 0 04 08 12 16 2 25 and 3 120583M All reproduced data shown are representativeof at least 3 independent experiments (c) Fluorescence quenching profiles of HSA (⧫) HSA-ibobrofen (◼) and HSA-warfarin (998771) in thepresence of various concentration of isoimperatorin
in Figure 11 It can be seen that isoimperatorin was situatedwithin subdomain IIA in Sudlowrsquos site I formed by six 120572-helices This figure shows the location of Trp 214 and also thebinding of isoimperatorin in HSA As this fluorescent residueis in subdomain IIA and due to the proximity of the bindingsite of isoimperatorin to the Trp 214 it can be concluded thatisoimperatorin has one reactive site onHSAandprobably thissite is at or near the site I The above docking results are ingood agreement with the displacement experiments results
and revealed that isoimperatorin binds to Sudlowrsquos site I ofHSA
37 Effect of Isoimperatorin on HSA Esterase Activity Pre-liminary studies showed that albumin in addition to itsligand binding capabilities has remarkable promiscuouscatalytic activities toward a broad range of organicmoleculesincluding esters amides phosphates and benzisoxazolesHuman serum albumin had significantly greater esterase-like
The Scientific World Journal 11
Table 4 Binding constants for isoimperatorin-HSA interaction (at 295K) in the absence and presence of the site marker
119870
119887
times 10
minus5
(Mminus1) 119877
a
HSA + isoimperatorin (in the absence ofibuprofen and warfarin) 1110 plusmn 04
lowast 09947
HSA + isoimperatorin + ibuprofen 1099 plusmn 06
lowast 09904HSA + isoimperatorin + warfarin 789 plusmn 03
lowast 09881Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870
119887values lowast119875 value lt 00002
Trp 214
Isoimperatorin
Figure 11 Crystal structure of HSA and the positions of singletryptophan residue (Trp 214) and also the binding site of isoim-peratorin in HSA Protein backbone is shown in the ldquocartoonrdquorepresentation and isoimperatorinand Trp 214 residue in the site Iare shown in the ldquostickrdquo representation The isoimperatorin and Trp214 structures are represented by the red and blue colorsThe crystalstructure of protein was obtained from Protein Data Bank (1BM0)For interpretation of references to color in this figure legend thereader is referred to the web version of this paper
activity than bovine serum albumin and defatted albuminwas more active than untreated albumin [50] InterestinglyHSA exhibits esterase-like activity and hydrolyzes drugshaving an ester group such as aspirin (which can convertaspirin ldquoacetylsalicylic acidrdquo to salicylic acid) Lys199 Lys402Lys519 and Lys545 form the pseudoesterase active site ofHSAthat is located in site I [50] ldquoEsterase-likerdquo activity (hydrolysisof p-nitrophenol esters such as p-NPA) by subdomain III Ahas been also reported and site directed mutagenesis studieshave shown that Arg 410 and Tyr 411 are essential in thisesterase activity However further investigation revealed thatacetylation is not limited to a distinct side chain so that theHSA incubation with high concentrations of pNPAc resultsin the acetylation of 59 Lys 10 Ser 8 Thr 4 Tyr and 1 Asp[51] Because of the above difference here we set out toevaluate the effect of isoimperatorin on the esterase activityof HSA based on our previous work [19] The kinetics ofthe pseudoenzymatic hydrolysis of pNPAc was obtained inthe absence and presence of isoimperatorin and site markers(Figure 12) Accordingly warfarin and indomethacin (as siteI marker) and isoimperatorin inhibited competitively theHSA-catalyzed hydrolysis of pNPAc by binding to site I Incontrast ibuprofen and tetracycline were unable to inhibitesterase activity of HSA The enzyme inhibition results sug-gested that isoimperatorin aspirin warfarin indomethacin
0
005
01
015
02
025
03
035
0 05 1 15 2 25 3Time (min)
HSA TetracyclineIbuprofenAspirin
IndomethacinOstholeWarfarinIsoimperatorin
A400
Figure 12 4-Nitrophenyl acetate esterase activity of HSA in theabsence and presence of tetracycline and ibuprofen (site II) war-farin aspirin indomethacin and osthole (site I) and isoimperatorin(site I) Final concentration of protein pNPA and drugs were 15250 and 250 120583M respectively For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
and osthole binding sites on the HSA molecule are the sameThese results also proposed that the large site I on HSAparticipates effectively to its promiscuous esterase activity
4 Conclusion
The accumulated data on HSA-drug interactions can beused to develop quantitative structure-activity relationshipsfor albumin binding However as stated earlier adsorptiondistribution metabolism and elimination (ADME) is con-trolled by albumin [30] Poor or strong binding to SA isbelieved to affect half-time of drugs [35] and their dosagedepends on their half-life in plasma Thus the bindingaffinity of any drug to SA is one of the major factors thatdetermine the pharmacokinetics halftime and availabilityof the drug in various tissues (see [30] and referencestherein) Further complexities arise in vivo due to interactions
12 The Scientific World Journal
between drugs and endogenous ligands for HSA This isparticularly pertinent for fatty acids which normally occurin serum at levels between 01 and 2mol per mol of HSAand can both compete and cooperate with drugs bindingto the protein In certain disease states these effects areexacerbated as the fatty acid HSA molar ratio may be ashigh as six Other pathological conditions are associatedwith high (micromolar to millimolar) levels of bilirubinhemin or renal toxins (eg 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) indoxyl sulphate) which bindto the protein causing significant drug binding defects
Our major aim in this study was to distinguish thenature of the interaction of isoimperatorin to HSA usingdifferent biophysical techniques This study could be veryvaluable due to importance and presence of isoimperatorinin human diet (food-drug interaction on HSA) The resultsshowed that isoimperatorin quenched the intrinsic fluo-rescence of HSA through dynamic quenching mechanismFurthermore hydrophobic interaction played a major role inthe binding process Using Fluorescence and UV absorptionspectroscopy it has been shown that isoimperatorin inducesprotein structural changes The binding average distancebetween the solo tryptophan of the protein and the boundisoimperatorin was calculated by Forster theory as 290 plusmn02 nmAnalysis of PSHvalues elucidated that isoimperatorinhas induced some compactness in the tertiary structure ofthe enzyme The results of synchronous flourescence onHSA-isoimperatorin system elucidated that the drug binds tothe hydrophobic cavity (site I) on subdomain IIA that wasconfirmed by quenching experiments competitive bindingexperiments and docking technique
Acknowledgment
The authors gratefully acknowledge the Kermanshah Uni-versity of Medical Sciences Research Council for financialsupport of this work
References
[1] Y-J Hu Y Liu T-Q Sun A-M Bai J-Q Lu and Z-B Pi ldquoBinding of anti-inflammatory drug cromolyn sodiumto bovine serum albuminrdquo International Journal of BiologicalMacromolecules vol 39 no 4-5 pp 280ndash285 2006
[2] N Seedher and S Bhatia ldquoReversible binding of celecoxiband valdecoxib with human serum albumin using fluorescencespectroscopic techniquerdquo Pharmacological Research vol 54 no2 pp 77ndash84 2006
[3] H Isogai and N Hirayama ldquoIn silico prediction of interactionsbetween site II on human serum albumin and profen drugsrdquoISRN Pharma vol 2013 Article ID 818364 8 pages 2013
[4] YWangHYuX Shi Z LuoD Lin andMHuang ldquoStructuralmechanism of ring opening reaction of glucose by humanserum albuminrdquo The Journal of Biological Chemistry vol 288pp 15980ndash15987 2013
[5] F Keshavarz M M Alavianmehr and R Yousefi ldquoMolecularinteraction of benzalkonium Ibuprofenate and its discreteingredients with human serum albuminrdquo Physical ChemistryResearch vol 1 no 2 pp 111ndash116 2013
[6] T K Maiti K S Ghosh J Debnath and S Dasgupta ldquoBindingof all-trans retinoic acid to human serum albumin fluores-cence FT-IR and circular dichroism studiesrdquo InternationalJournal of Biological Macromolecules vol 38 no 3ndash5 pp 197ndash202 2006
[7] M-X XieM Long Y Liu C Qin and Y-DWang ldquoCharacter-ization of the interaction between human serum albumin andmorinrdquo Biochimica et Biophysica Acta vol 1760 no 8 pp 1184ndash1191 2006
[8] CD Kanakis P A TarantilisMG Polissiou S Diamantoglouand H A Tajmir-Riahi ldquoAntioxidant flavonoids bind humanserum albuminrdquo Journal of Molecular Structure vol 798 no 1-3 pp 69ndash74 2006
[9] S Marumoto and M Miyazawa ldquoStructure-activity rela-tionships for naturally occurring coumarins as 120573-secretaseinhibitorrdquo Bioorganic and Medicinal Chemistry vol 20 no 2pp 784ndash788 2012
[10] S Marumoto and M Miyazawa ldquoBiotransformation of isoim-peratorin and imperatorin by Glomerella cingulata and 120573-secretase inhibitory activityrdquo Bioorganic and Medicinal Chem-istry vol 18 no 1 pp 455ndash459 2010
[11] S E Sajjadi Y Shokoohinia and S Hemmati ldquoIsolation andidentification of furanocoumarins and a phenylpropanoid fromthe acetone extract and identification of volatile constituentsfrom the essential oil of Peucedanum pastinacifoliumrdquo Chem-istry of Natural Compounds vol 48 no 4 pp 668ndash671 2012
[12] X B Yang J Hou D Liu et al ldquoBiotransformation of isoim-peratorin by Cunninghamella blakesleana AS 3970rdquo Journal ofMolecular Catalysis B vol 88 pp 1ndash6 2013
[13] W Lili S Yehong S Qi et al ldquoIn vitro permeability analy-sis pharmacokinetic and brain distribution study in mice ofimperatorin isoimperatorin and cnidilin in Radix AngelicaeDahuricaerdquo Fitoterapia vol 85 pp 144ndash153 2013
[14] S Wang Q Chen and L He ldquoDevelopment and validationof a gas chromatography-mass spectrometry method for thedetermination of isoimperatorin in rat plasma and tissueapplication to the pharmacokinetic and tissue distributionstudyrdquo Journal of Chromatography B vol 852 no 1-2 pp 473ndash478 2007
[15] H Sadraei Y Shokoohinia S E Sajjadi and M MozafarildquoAntispasmodic effects of Prangos ferulacea acetone extract andits maincomponent osthole on ileum contractionrdquo Research inPharmaceutical Sciences vol 8 pp 137ndash144 2013
[16] H Sadraei Y Shokoohinia S E Sajjadi and B GhadirianldquoAntispasmodic effect of osthole and Prangos ferulacea extracton rat uterus smooth muscle motilityrdquo Research in Pharmaceu-tical Sciences vol 7 pp 141ndash149 2012
[17] S Sajjadi Y Shokoohinia S Gholamzadeh M Behbahani andA Fattahi ldquoAntiviral evaluation of coumarins from Prangosferulacea L (Lindl)rdquo Research in Pharmaceutical Sciences vol7 p 783 2012
[18] S E Sajjadi Y Shokoohinia and S Gholamzadeh ldquoChemicalcomposition of the essential oil of the root of Prangos ferulacea(L) Lindlrdquo Chemija vol 22 pp 178ndash180 2011
[19] N Bijari Y Shokoohinia M-R Ashrafi-Kooshk et al ldquoSpec-troscopic study of interaction between osthole and humanserum albumin identification of possible binding site of thecompoundrdquo Journal of Luminescence vol 143 pp 328ndash336 2013
[20] X Shi M Liu M Zhang et al ldquoIdentification of in vitro and invivo metabolites of isoimperatorin using liquid chromatogra-phymass spectrometryrdquo Food Chemistry vol 141 pp 357ndash3652013
The Scientific World Journal 13
[21] T Chatterjee A Pal S Dey B-K Chatterjee and PChakrabarti ldquoInteraction of virstatin with human serum albu-min spectroscopic analysis andmolecular modelingrdquo Plos Onevol 7 pp 1ndash12 2012
[22] Y-M Chen and L-H Guo ldquoCombined fluorescence and elec-trochemical investigation on the binding interaction betweenorganic acid and human serum albuminrdquo Journal of Environ-mental Sciences vol 21 no 3 pp 373ndash379 2009
[23] J R Lakowicz Principles of Fluorescence Spectroscopy KluwerAcademicPlenum Publishers New York NY USA 2nd edi-tion 1999
[24] F Ding N Li B Han F Liu L Zhang and Y Sun ldquoThe bindingof CI Acid Red 2 to human serum albumin determinationof binding mechanism and binding site using fluorescencespectroscopyrdquo Dyes and Pigments vol 83 no 2 pp 249ndash2572009
[25] M-R Ashrafi Kooshk K Mansouri M Nadi S Khodarahmiand R Khodarahmi ldquoIn vitro anti-cancer activity of nativecurcumin and protein-curcuminrdquo Systems vol 2 no 1 pp 80ndash88 1981
[26] P D Ross and S Subramanian ldquoThermodynamics of proteinassociation reactions forces contributing to stabilityrdquo Biochem-istry vol 20 no 11 pp 3096ndash3102 1981
[27] J Liu J-N Tian J Zhang Z Hu and X Chen ldquoInteractionof magnolol with bovine serum albumin a fluorescence-quenching studyrdquo Analytical and Bioanalytical Chemistry vol376 no 6 pp 864ndash867 2003
[28] A Coi A M Bianucci F Bonomi P Rasmussen G M Muraand M L Ganadu ldquoStructural perturbation of 120572B-crystallinby zinc and temperature related to its chaperone-like activityrdquoInternational Journal of Biological Macromolecules vol 42 no3 pp 229ndash234 2008
[29] M Moller and A Denicola ldquoStudy of protein-ligand bindingby fluorescencerdquoBiochemistry andMolecular Biology Educationvol 30 no 5 pp 309ndash312 2002
[30] R Khodarahmi S A Karimi M R Ashrafi Kooshk S AGhadami S Ghobadi and M Amani ldquoComparative spec-troscopic studies on drug binding characteristics and proteinsurface hydrophobicity of native and modified forms of bovineserum albumin possible relevance to change in protein struc-turefunction upon non-enzymatic glycationrdquo SpectrochimicaActa A vol 89 pp 177ndash186 2012
[31] S-F Ma M Anraku Y Iwao et al ldquoHydrolysis of angiotensinII receptor blocker prodrug olmesartan medoxomil by humanserum albumin and identification of its catalytic active sitesrdquoDrug Metabolism and Disposition vol 33 no 12 pp 1911ndash19192005
[32] H M Berman J Westbrook Z Feng et al ldquoThe protein databankrdquo Nucleic Acids Research vol 28 no 1 pp 235ndash242 2000
[33] S Sugio A Kashima SMochizukiMNoda andKKobayashildquoCrystal structure of human serum albumin at 25 A resolutionrdquoProtein Engineering vol 12 no 6 pp 439ndash446 1999
[34] G M Morris D S Goodsell R Huey and A J OlsonldquoDistributed automated docking of flexible ligands to proteinsparallel applications of AutoDock 24rdquo Journal of Computer-Aided Molecular Design vol 10 no 4 pp 293ndash304 1996
[35] F Faridbod M R Ganjali B Larijani et al ldquoInteraction studyof pioglitazone with albumin by fluorescence spectroscopy andmolecular dockingrdquo Spectrochimica Acta A vol 78 no 1 pp96ndash101 2011
[36] D SGoodsell andA JOlson ldquoAutomated docking of substratesto proteins by simulated annealingrdquo Proteins vol 8 no 3 pp195ndash202 1990
[37] D S Goodsell G M Morris and A J Olson ldquoAutomateddocking of flexible ligands applications of AutoDockrdquo Journalof Molecular Recognition vol 9 no 1 pp 1ndash5 1996
[38] G Sudlow D J Birkett and D N Wade ldquoThe characterizationof two specific drug binding sites on human serum albuminrdquoMolecular Pharmacology vol 11 no 6 pp 824ndash832 1975
[39] Y Yue X Chen J Qin and X Yao ldquoCharacterization of themangiferin-human serum albumin complex by spectroscopicandmolecularmodeling approachesrdquo Journal of Pharmaceuticaland Biomedical Analysis vol 49 no 3 pp 753ndash759 2009
[40] A K Shaw and S K Pal ldquoSpectroscopic studies on the effectof temperature on pH-induced folded states of human serumalbuminrdquo Journal of Photochemistry and Photobiology B vol 90no 1 pp 69ndash77 2008
[41] S Ranjbar S Ghobadi R Khodarahmi and H Nemati ldquoSpec-troscopic characterization of furosemide binding to humancarbonic anhydrase IIrdquo International Journal of BiologicalMacromolecules vol 50 no 4 pp 910ndash917 2012
[42] HNHou Z POi YWOYung F L Liao Y Zhang andY LiyldquoStudies on interaction between vitamin B12 and human serumalbuminrdquo Journal of Pharmaceutical and Biomedical Analysisvol 47 pp 134ndash139 2008
[43] S S Kalanur J Seetharamappa and V K A Kalalbandi ldquoChar-acterization of interaction and the effect of carbamazepine onthe structure of human serum albuminrdquo Journal of Pharmaceu-tical and Biomedical Analysis vol 53 no 3 pp 660ndash666 2010
[44] L Stryer ldquoFluorescence spectroscopy of proteinsrdquo Science vol162 no 3853 pp 526ndash533 1968
[45] B Valeur and J C Brochon New Trends in FluorescenceSpectroscopy Springer Berlin Germany 1999
[46] V Suryawanshi P Anbhule A Gore Sh Patil and G AndKolekar ldquoA spectral deciphering the perturbation of modeltransporter protein (HSA) by antibacterial pyrimidine deriva-tive pharmacokinetic and biophysical insightsrdquo Journal ofPhotochemistry and Photobiology B vol 118 pp 1ndash8 2013
[47] T Yuan AMWeljie andH J Vogel ldquoTryptophan fluorescencequenching by methionine and selenomethionine residues ofcalmodulin orientation of peptide and protein bindingrdquo Bio-chemistry vol 37 no 9 pp 3187ndash3195 1998
[48] TCOrsquoHaver A F Fell G Smith et al ldquoDerivative spectroscopyand its applications in analysisrdquo Analytical Proceedings vol 19no 1 pp 22ndash46 1982
[49] Y-Q Wang B-P Tang H-M Zhang Q-H Zhou and G-C Zhang ldquoStudies on the interaction between imidaclopridand human serum albumin spectroscopic approachrdquo Journal ofPhotochemistry and Photobiology B vol 94 no 3 pp 183ndash1902009
[50] S-J Kim H-W Rhee H-Y Park H-S Kim and J-I HongldquoFluorescent probes designed for detecting human serum albu-min on the basis of its pseudo-esterase activityrdquo Bioorganic ampMedicinal Chemistry Letters vol 23 pp 2093ndash2097 2013
[51] P Ascenzi M Gioia G Fanali M Coletta and M FasanoldquoPseudo-enzymatic hydrolysis of 4-nitrophenyl acetate byhuman serum albumin pH-dependence of rates of individualstepsrdquo Biochemical and Biophysical Research Communicationsvol 424 pp 451ndash455 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 The Scientific World Journal
Table 3 Surface hydrophobicity parameters for HSA in the absence and the presence of isoimperatorin
Parameters HSA HSA-IIM complex119870
app119889
(120583M) 09 plusmn 005
lowast
14 plusmn 007
lowast
119865max[HSA] 135714 plusmn 85
lowastlowast
112853 plusmn 63
lowastlowast
Surface hydrophobicity index (119865max[HSA] 119870app119889
) 150794 plusmn 1146
lowastlowast
80612 plusmn 007
lowastlowast
Data are expressed as mean plusmn SD of three measurements lowast119875 value lt 00006 lowastlowast119875 value lt 00001
0
50
100
150
200
250
300
350
400
400 500 600 700 800 900 1000Fluorescence intensity (au)
Fmax
Fluo
resc
ence
inte
nsity
(au
)[A
NS]
free
Slope = minus1Kappd
Figure 8 Scatchard plots for titration of HSA with increasingconcentrations of ANS in the absence (∙) and presence (998771) of 3 120583Mof isoimperatorin 119865max and 1119870
app119889
values were obtained from the119909-intercept and slope of this plot For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
advantages are available spectral simplification spectralbandwidth reduction and avoidance of different perturbingeffects The fluorescence spectrum of BSAHSA mainly dueto the Trp residues is sensitive to the microenvironment ofthese chromophoreswith themaximumemissionwavelengthbeing very useful in estimating the hydrophobicity aroundthe tryptophan residues It is well known that the experimenton synchronous fluorescence of HSA will provide the char-acteristic information for the Tyr and Trp residues when thescanning interval Δ120582 is fixed at 15 and 60 nm respectively[47] The shift in the position of fluorescence emissionmaximum corresponds to changes of the polarity aroundthe chromophore molecule A blue shift of 120582max means thatthe amino acid residues are located in a more hydrophobicenvironment and are less exposed to the solvent while a redshift of 120582max implies that the amino acid residues are in apolar environment and are more exposed to the solvent [48]The synchronous fluorescence spectra of interaction betweenisoimperatorin and HSA are presented in Figure 8 It wasshown that the fluorescence intensity of HSA decreased regu-larly along with the addition of isoimperatorin which furtherdemonstrated the occurrence of fluorescence quenching inthe binding process Moreover there is no significant shiftof the maximum emission wavelength with Δ120582 = 15 nm or60 nm (Figures 9(a) and 9(b)) which implies that interaction
of isoimperatorin with HSA does not affect the conformationof the region around the Tyr or Trp residues
As well we can obtain the information about the locationof isoimperatorin binding site from the synchronous fluores-cence dataThe ratios of synchronous fluorescence quenching(RSFQ) were used to determine the binding sites of the drugsto the HSA molecule in drug-HSA complexes The RSFQvalues were calculated from the following equation
RSFQ = 1 minus
119865
119865
0
(11)
where 119865 and 1198650are the synchronous fluorescence intensities
of HSA in the presence and the absence of drug respectivelyIt was obvious from Figure 9(c) that the RSFQ results forΔ120582 = 60 nmwere slightly higher than the corresponding onesfor Δ120582 = 15 nm As illustrated in Figure 9(c) addition ofisoimperatorin resulted in strong fluorescence quenching ofTrp residue (sim47 reduction in fluorescence intensity) con-firming that isoimperatorin reached subdomain IIA whereonly one Trp residue (Trp 214) in HSA is located It wasalso observed from Figure 9(c) that the fluorescence strengthof Tyr residues decreased about sim32 in the presence ofisoimperatorin So the binding of isoimperatorin to HSAquenched flourescence intensity of Tyr and Trp Since Trp214 (the only Trp of HSA) is at site I and Tyr 263 (hasthe strong fluorescence in the case of Tyr residues andmay be one of the residues responsible for the synchronousfluorescence spectrum of HSA when Δ120582 is fixed at 15 nm) isin subdomain IIA (site I) it can be concluded that Trp andTyr residues of HSA may have equal accessibility to bindingsite of isoimperatorin [49] The above results indicated thatisoimperatorin can bind to HSA in the hydrophobic cavity(site I) on subdomain IIA which are in full agreement withquenching competitive binding experiments and theoreticaldocking technique
36 Site Marker Competitive Binding Experiments In orderto further classify drug binding site on HSA competitivebinding experiments have been carried out Warfarin andibuprofen are two markers specific for binding to HSA thatbind to sites I and II which are corresponded to the sub-domains IIA and IIIA respectively Therefore to determinethe location of the isoimperatorin binding site on HSA thecompetitive displacement experimentswere carried out usingwarfarin as a characteristicmarker for site I (Figure 10(a)) andibuprofen for site II (Figure 10(b)) [24 31] In the site markercompetitive experiment isoimperatorin was gradually addedto the solution of HSA HSA-ibuprofen or HSA-warfarincomplex and then fluorescence intensity of system wasrecorded
The Scientific World Journal 9
0
100
200
300
400
500
600
265 270 275 280 285 290 295 300Wavelength (nm)
1
8
Fluo
resc
ence
inte
nsity
(au
)
(a)
Fluo
resc
ence
inte
nsity
(au
)
0
50
100
150
200
250
300
350
400
250 260 270 280 290 300Wavelength (nm)
1
8
(b)
0
10
20
30
40
50
0 05 1 15 2 25 3
RSFQ
()
Isoimperatorin (120583M)
0
02
04
06
08
1
12
005 1 15 2 25 3 35
Isoimperatorin (120583M)
F0
F
(c)
Figure 9 Synchronous fluorescence spectra of interaction between HSA and isoimperatorin (a) at Δ120582 = 15 nm and (b) at Δ120582 = 60 nmConcentration ofHSAwas 30 120583Mwhile concentrations of isoimperatorinwere 0 04 08 12 16 2 25 and 3 120583Mfrom 1 to 8 (c) Comparativeevaluation of isoimperatorin effect on the RSFQ of HSA [HSA] = 3 120583M (◼) Δ120582 = 15 nm and (∙) Δ120582 = 60 nm
As shown in Figure 10(a) with addition of warfarin in theHSA solution the flourescence intensity was slightly higherthan that of without warfarin (and including the red shift)Then after adding the isoimperatorin into the above systemthe flourescence intensity of HSA solution with warfarinheld in equimolar decreased gradually and the intensitywas much lower than that of without warfarin displayingthat the binding of the isoimperatorin to HSA was affectedafter adding warfarin On the contrary in the presence ofibuprofen the flourescence intensity of the HSA-ibuprofencomplex almost had no difference from that recordedwithoutibuprofen under the same conditions (Figure 10(b)) whichindicated that site II marker did not prevent the binding ofisoimperatorin in its usual binding location The next stepprotein intrinsic fluorescence was measured and the relativefluorescence intensities (119865119865
0) versus ligand concentration
([isoimperatorin]) plots are displayed in Figure 10(c) Asshown in Table 4 the binding constant was remarkablydecreased after addition of warfarin while the addition ofibuprofen results in only a small difference These resultsindicate that warfarin can displace the isoimperatorin but
ibuprofen has little effect on the binding of isoimperatorinto HSA In other words these results suggest that isoim-peratorin probably competes with warfarin for binding toHSA [24] Experimental observations were followed up withdocking studies where isoimperatorin was docked to HSAto determine the preferred binding site on the proteinAs described above the 3D crystal structure of HSA is amonomer consisting of three homologous domains whichassemble to form a heart-shaped molecule [4 6] Each ofthe structurally similar 120572-helix domains (IndashIII) has twosubdomains (A andB) with six120572-helices in subdomainA andfour 120572-helices in subdomain BThe crystal structure analysesindicate that the principal regions of ligand binding sitesin HSA are located in hydrophobic cavities in subdomainsIIA and IIIA which are consistent with Sudlowrsquos site I andSudlowrsquos site II respectively Several studies have shown thatHSA is able to bind many ligands in several binding sites[38] In this paper the molecular docking model of theHSAndashisoimperatorin complex was conducted on the basisof the crystal structure of HSA to find the binding positionof isoimperatorin The best docking energy result is shown
10 The Scientific World Journal
0
100
200
300
400
500
600
310 335 360 385 410Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA1
8
(a)
0
100
200
300
400
500
600
700
305 325 345 365 385 405Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA
1
8
(b)
0
05
1
15
0 1 2 3
HSAIbuprofen
Warfarin
Isoimperatorin (120583M)
F0
F
(c)
Figure 10 Effect of isoimperatorin on the quenching of HSA-warfarin complex (a) andHSA-ibuprofen complex (b) fluorescence From 1 to 8[HSA] = [warfarin or iboprufen] = 3 120583M [isoimperatorin] = 0 04 08 12 16 2 25 and 3 120583M All reproduced data shown are representativeof at least 3 independent experiments (c) Fluorescence quenching profiles of HSA (⧫) HSA-ibobrofen (◼) and HSA-warfarin (998771) in thepresence of various concentration of isoimperatorin
in Figure 11 It can be seen that isoimperatorin was situatedwithin subdomain IIA in Sudlowrsquos site I formed by six 120572-helices This figure shows the location of Trp 214 and also thebinding of isoimperatorin in HSA As this fluorescent residueis in subdomain IIA and due to the proximity of the bindingsite of isoimperatorin to the Trp 214 it can be concluded thatisoimperatorin has one reactive site onHSAandprobably thissite is at or near the site I The above docking results are ingood agreement with the displacement experiments results
and revealed that isoimperatorin binds to Sudlowrsquos site I ofHSA
37 Effect of Isoimperatorin on HSA Esterase Activity Pre-liminary studies showed that albumin in addition to itsligand binding capabilities has remarkable promiscuouscatalytic activities toward a broad range of organicmoleculesincluding esters amides phosphates and benzisoxazolesHuman serum albumin had significantly greater esterase-like
The Scientific World Journal 11
Table 4 Binding constants for isoimperatorin-HSA interaction (at 295K) in the absence and presence of the site marker
119870
119887
times 10
minus5
(Mminus1) 119877
a
HSA + isoimperatorin (in the absence ofibuprofen and warfarin) 1110 plusmn 04
lowast 09947
HSA + isoimperatorin + ibuprofen 1099 plusmn 06
lowast 09904HSA + isoimperatorin + warfarin 789 plusmn 03
lowast 09881Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870
119887values lowast119875 value lt 00002
Trp 214
Isoimperatorin
Figure 11 Crystal structure of HSA and the positions of singletryptophan residue (Trp 214) and also the binding site of isoim-peratorin in HSA Protein backbone is shown in the ldquocartoonrdquorepresentation and isoimperatorinand Trp 214 residue in the site Iare shown in the ldquostickrdquo representation The isoimperatorin and Trp214 structures are represented by the red and blue colorsThe crystalstructure of protein was obtained from Protein Data Bank (1BM0)For interpretation of references to color in this figure legend thereader is referred to the web version of this paper
activity than bovine serum albumin and defatted albuminwas more active than untreated albumin [50] InterestinglyHSA exhibits esterase-like activity and hydrolyzes drugshaving an ester group such as aspirin (which can convertaspirin ldquoacetylsalicylic acidrdquo to salicylic acid) Lys199 Lys402Lys519 and Lys545 form the pseudoesterase active site ofHSAthat is located in site I [50] ldquoEsterase-likerdquo activity (hydrolysisof p-nitrophenol esters such as p-NPA) by subdomain III Ahas been also reported and site directed mutagenesis studieshave shown that Arg 410 and Tyr 411 are essential in thisesterase activity However further investigation revealed thatacetylation is not limited to a distinct side chain so that theHSA incubation with high concentrations of pNPAc resultsin the acetylation of 59 Lys 10 Ser 8 Thr 4 Tyr and 1 Asp[51] Because of the above difference here we set out toevaluate the effect of isoimperatorin on the esterase activityof HSA based on our previous work [19] The kinetics ofthe pseudoenzymatic hydrolysis of pNPAc was obtained inthe absence and presence of isoimperatorin and site markers(Figure 12) Accordingly warfarin and indomethacin (as siteI marker) and isoimperatorin inhibited competitively theHSA-catalyzed hydrolysis of pNPAc by binding to site I Incontrast ibuprofen and tetracycline were unable to inhibitesterase activity of HSA The enzyme inhibition results sug-gested that isoimperatorin aspirin warfarin indomethacin
0
005
01
015
02
025
03
035
0 05 1 15 2 25 3Time (min)
HSA TetracyclineIbuprofenAspirin
IndomethacinOstholeWarfarinIsoimperatorin
A400
Figure 12 4-Nitrophenyl acetate esterase activity of HSA in theabsence and presence of tetracycline and ibuprofen (site II) war-farin aspirin indomethacin and osthole (site I) and isoimperatorin(site I) Final concentration of protein pNPA and drugs were 15250 and 250 120583M respectively For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
and osthole binding sites on the HSA molecule are the sameThese results also proposed that the large site I on HSAparticipates effectively to its promiscuous esterase activity
4 Conclusion
The accumulated data on HSA-drug interactions can beused to develop quantitative structure-activity relationshipsfor albumin binding However as stated earlier adsorptiondistribution metabolism and elimination (ADME) is con-trolled by albumin [30] Poor or strong binding to SA isbelieved to affect half-time of drugs [35] and their dosagedepends on their half-life in plasma Thus the bindingaffinity of any drug to SA is one of the major factors thatdetermine the pharmacokinetics halftime and availabilityof the drug in various tissues (see [30] and referencestherein) Further complexities arise in vivo due to interactions
12 The Scientific World Journal
between drugs and endogenous ligands for HSA This isparticularly pertinent for fatty acids which normally occurin serum at levels between 01 and 2mol per mol of HSAand can both compete and cooperate with drugs bindingto the protein In certain disease states these effects areexacerbated as the fatty acid HSA molar ratio may be ashigh as six Other pathological conditions are associatedwith high (micromolar to millimolar) levels of bilirubinhemin or renal toxins (eg 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) indoxyl sulphate) which bindto the protein causing significant drug binding defects
Our major aim in this study was to distinguish thenature of the interaction of isoimperatorin to HSA usingdifferent biophysical techniques This study could be veryvaluable due to importance and presence of isoimperatorinin human diet (food-drug interaction on HSA) The resultsshowed that isoimperatorin quenched the intrinsic fluo-rescence of HSA through dynamic quenching mechanismFurthermore hydrophobic interaction played a major role inthe binding process Using Fluorescence and UV absorptionspectroscopy it has been shown that isoimperatorin inducesprotein structural changes The binding average distancebetween the solo tryptophan of the protein and the boundisoimperatorin was calculated by Forster theory as 290 plusmn02 nmAnalysis of PSHvalues elucidated that isoimperatorinhas induced some compactness in the tertiary structure ofthe enzyme The results of synchronous flourescence onHSA-isoimperatorin system elucidated that the drug binds tothe hydrophobic cavity (site I) on subdomain IIA that wasconfirmed by quenching experiments competitive bindingexperiments and docking technique
Acknowledgment
The authors gratefully acknowledge the Kermanshah Uni-versity of Medical Sciences Research Council for financialsupport of this work
References
[1] Y-J Hu Y Liu T-Q Sun A-M Bai J-Q Lu and Z-B Pi ldquoBinding of anti-inflammatory drug cromolyn sodiumto bovine serum albuminrdquo International Journal of BiologicalMacromolecules vol 39 no 4-5 pp 280ndash285 2006
[2] N Seedher and S Bhatia ldquoReversible binding of celecoxiband valdecoxib with human serum albumin using fluorescencespectroscopic techniquerdquo Pharmacological Research vol 54 no2 pp 77ndash84 2006
[3] H Isogai and N Hirayama ldquoIn silico prediction of interactionsbetween site II on human serum albumin and profen drugsrdquoISRN Pharma vol 2013 Article ID 818364 8 pages 2013
[4] YWangHYuX Shi Z LuoD Lin andMHuang ldquoStructuralmechanism of ring opening reaction of glucose by humanserum albuminrdquo The Journal of Biological Chemistry vol 288pp 15980ndash15987 2013
[5] F Keshavarz M M Alavianmehr and R Yousefi ldquoMolecularinteraction of benzalkonium Ibuprofenate and its discreteingredients with human serum albuminrdquo Physical ChemistryResearch vol 1 no 2 pp 111ndash116 2013
[6] T K Maiti K S Ghosh J Debnath and S Dasgupta ldquoBindingof all-trans retinoic acid to human serum albumin fluores-cence FT-IR and circular dichroism studiesrdquo InternationalJournal of Biological Macromolecules vol 38 no 3ndash5 pp 197ndash202 2006
[7] M-X XieM Long Y Liu C Qin and Y-DWang ldquoCharacter-ization of the interaction between human serum albumin andmorinrdquo Biochimica et Biophysica Acta vol 1760 no 8 pp 1184ndash1191 2006
[8] CD Kanakis P A TarantilisMG Polissiou S Diamantoglouand H A Tajmir-Riahi ldquoAntioxidant flavonoids bind humanserum albuminrdquo Journal of Molecular Structure vol 798 no 1-3 pp 69ndash74 2006
[9] S Marumoto and M Miyazawa ldquoStructure-activity rela-tionships for naturally occurring coumarins as 120573-secretaseinhibitorrdquo Bioorganic and Medicinal Chemistry vol 20 no 2pp 784ndash788 2012
[10] S Marumoto and M Miyazawa ldquoBiotransformation of isoim-peratorin and imperatorin by Glomerella cingulata and 120573-secretase inhibitory activityrdquo Bioorganic and Medicinal Chem-istry vol 18 no 1 pp 455ndash459 2010
[11] S E Sajjadi Y Shokoohinia and S Hemmati ldquoIsolation andidentification of furanocoumarins and a phenylpropanoid fromthe acetone extract and identification of volatile constituentsfrom the essential oil of Peucedanum pastinacifoliumrdquo Chem-istry of Natural Compounds vol 48 no 4 pp 668ndash671 2012
[12] X B Yang J Hou D Liu et al ldquoBiotransformation of isoim-peratorin by Cunninghamella blakesleana AS 3970rdquo Journal ofMolecular Catalysis B vol 88 pp 1ndash6 2013
[13] W Lili S Yehong S Qi et al ldquoIn vitro permeability analy-sis pharmacokinetic and brain distribution study in mice ofimperatorin isoimperatorin and cnidilin in Radix AngelicaeDahuricaerdquo Fitoterapia vol 85 pp 144ndash153 2013
[14] S Wang Q Chen and L He ldquoDevelopment and validationof a gas chromatography-mass spectrometry method for thedetermination of isoimperatorin in rat plasma and tissueapplication to the pharmacokinetic and tissue distributionstudyrdquo Journal of Chromatography B vol 852 no 1-2 pp 473ndash478 2007
[15] H Sadraei Y Shokoohinia S E Sajjadi and M MozafarildquoAntispasmodic effects of Prangos ferulacea acetone extract andits maincomponent osthole on ileum contractionrdquo Research inPharmaceutical Sciences vol 8 pp 137ndash144 2013
[16] H Sadraei Y Shokoohinia S E Sajjadi and B GhadirianldquoAntispasmodic effect of osthole and Prangos ferulacea extracton rat uterus smooth muscle motilityrdquo Research in Pharmaceu-tical Sciences vol 7 pp 141ndash149 2012
[17] S Sajjadi Y Shokoohinia S Gholamzadeh M Behbahani andA Fattahi ldquoAntiviral evaluation of coumarins from Prangosferulacea L (Lindl)rdquo Research in Pharmaceutical Sciences vol7 p 783 2012
[18] S E Sajjadi Y Shokoohinia and S Gholamzadeh ldquoChemicalcomposition of the essential oil of the root of Prangos ferulacea(L) Lindlrdquo Chemija vol 22 pp 178ndash180 2011
[19] N Bijari Y Shokoohinia M-R Ashrafi-Kooshk et al ldquoSpec-troscopic study of interaction between osthole and humanserum albumin identification of possible binding site of thecompoundrdquo Journal of Luminescence vol 143 pp 328ndash336 2013
[20] X Shi M Liu M Zhang et al ldquoIdentification of in vitro and invivo metabolites of isoimperatorin using liquid chromatogra-phymass spectrometryrdquo Food Chemistry vol 141 pp 357ndash3652013
The Scientific World Journal 13
[21] T Chatterjee A Pal S Dey B-K Chatterjee and PChakrabarti ldquoInteraction of virstatin with human serum albu-min spectroscopic analysis andmolecular modelingrdquo Plos Onevol 7 pp 1ndash12 2012
[22] Y-M Chen and L-H Guo ldquoCombined fluorescence and elec-trochemical investigation on the binding interaction betweenorganic acid and human serum albuminrdquo Journal of Environ-mental Sciences vol 21 no 3 pp 373ndash379 2009
[23] J R Lakowicz Principles of Fluorescence Spectroscopy KluwerAcademicPlenum Publishers New York NY USA 2nd edi-tion 1999
[24] F Ding N Li B Han F Liu L Zhang and Y Sun ldquoThe bindingof CI Acid Red 2 to human serum albumin determinationof binding mechanism and binding site using fluorescencespectroscopyrdquo Dyes and Pigments vol 83 no 2 pp 249ndash2572009
[25] M-R Ashrafi Kooshk K Mansouri M Nadi S Khodarahmiand R Khodarahmi ldquoIn vitro anti-cancer activity of nativecurcumin and protein-curcuminrdquo Systems vol 2 no 1 pp 80ndash88 1981
[26] P D Ross and S Subramanian ldquoThermodynamics of proteinassociation reactions forces contributing to stabilityrdquo Biochem-istry vol 20 no 11 pp 3096ndash3102 1981
[27] J Liu J-N Tian J Zhang Z Hu and X Chen ldquoInteractionof magnolol with bovine serum albumin a fluorescence-quenching studyrdquo Analytical and Bioanalytical Chemistry vol376 no 6 pp 864ndash867 2003
[28] A Coi A M Bianucci F Bonomi P Rasmussen G M Muraand M L Ganadu ldquoStructural perturbation of 120572B-crystallinby zinc and temperature related to its chaperone-like activityrdquoInternational Journal of Biological Macromolecules vol 42 no3 pp 229ndash234 2008
[29] M Moller and A Denicola ldquoStudy of protein-ligand bindingby fluorescencerdquoBiochemistry andMolecular Biology Educationvol 30 no 5 pp 309ndash312 2002
[30] R Khodarahmi S A Karimi M R Ashrafi Kooshk S AGhadami S Ghobadi and M Amani ldquoComparative spec-troscopic studies on drug binding characteristics and proteinsurface hydrophobicity of native and modified forms of bovineserum albumin possible relevance to change in protein struc-turefunction upon non-enzymatic glycationrdquo SpectrochimicaActa A vol 89 pp 177ndash186 2012
[31] S-F Ma M Anraku Y Iwao et al ldquoHydrolysis of angiotensinII receptor blocker prodrug olmesartan medoxomil by humanserum albumin and identification of its catalytic active sitesrdquoDrug Metabolism and Disposition vol 33 no 12 pp 1911ndash19192005
[32] H M Berman J Westbrook Z Feng et al ldquoThe protein databankrdquo Nucleic Acids Research vol 28 no 1 pp 235ndash242 2000
[33] S Sugio A Kashima SMochizukiMNoda andKKobayashildquoCrystal structure of human serum albumin at 25 A resolutionrdquoProtein Engineering vol 12 no 6 pp 439ndash446 1999
[34] G M Morris D S Goodsell R Huey and A J OlsonldquoDistributed automated docking of flexible ligands to proteinsparallel applications of AutoDock 24rdquo Journal of Computer-Aided Molecular Design vol 10 no 4 pp 293ndash304 1996
[35] F Faridbod M R Ganjali B Larijani et al ldquoInteraction studyof pioglitazone with albumin by fluorescence spectroscopy andmolecular dockingrdquo Spectrochimica Acta A vol 78 no 1 pp96ndash101 2011
[36] D SGoodsell andA JOlson ldquoAutomated docking of substratesto proteins by simulated annealingrdquo Proteins vol 8 no 3 pp195ndash202 1990
[37] D S Goodsell G M Morris and A J Olson ldquoAutomateddocking of flexible ligands applications of AutoDockrdquo Journalof Molecular Recognition vol 9 no 1 pp 1ndash5 1996
[38] G Sudlow D J Birkett and D N Wade ldquoThe characterizationof two specific drug binding sites on human serum albuminrdquoMolecular Pharmacology vol 11 no 6 pp 824ndash832 1975
[39] Y Yue X Chen J Qin and X Yao ldquoCharacterization of themangiferin-human serum albumin complex by spectroscopicandmolecularmodeling approachesrdquo Journal of Pharmaceuticaland Biomedical Analysis vol 49 no 3 pp 753ndash759 2009
[40] A K Shaw and S K Pal ldquoSpectroscopic studies on the effectof temperature on pH-induced folded states of human serumalbuminrdquo Journal of Photochemistry and Photobiology B vol 90no 1 pp 69ndash77 2008
[41] S Ranjbar S Ghobadi R Khodarahmi and H Nemati ldquoSpec-troscopic characterization of furosemide binding to humancarbonic anhydrase IIrdquo International Journal of BiologicalMacromolecules vol 50 no 4 pp 910ndash917 2012
[42] HNHou Z POi YWOYung F L Liao Y Zhang andY LiyldquoStudies on interaction between vitamin B12 and human serumalbuminrdquo Journal of Pharmaceutical and Biomedical Analysisvol 47 pp 134ndash139 2008
[43] S S Kalanur J Seetharamappa and V K A Kalalbandi ldquoChar-acterization of interaction and the effect of carbamazepine onthe structure of human serum albuminrdquo Journal of Pharmaceu-tical and Biomedical Analysis vol 53 no 3 pp 660ndash666 2010
[44] L Stryer ldquoFluorescence spectroscopy of proteinsrdquo Science vol162 no 3853 pp 526ndash533 1968
[45] B Valeur and J C Brochon New Trends in FluorescenceSpectroscopy Springer Berlin Germany 1999
[46] V Suryawanshi P Anbhule A Gore Sh Patil and G AndKolekar ldquoA spectral deciphering the perturbation of modeltransporter protein (HSA) by antibacterial pyrimidine deriva-tive pharmacokinetic and biophysical insightsrdquo Journal ofPhotochemistry and Photobiology B vol 118 pp 1ndash8 2013
[47] T Yuan AMWeljie andH J Vogel ldquoTryptophan fluorescencequenching by methionine and selenomethionine residues ofcalmodulin orientation of peptide and protein bindingrdquo Bio-chemistry vol 37 no 9 pp 3187ndash3195 1998
[48] TCOrsquoHaver A F Fell G Smith et al ldquoDerivative spectroscopyand its applications in analysisrdquo Analytical Proceedings vol 19no 1 pp 22ndash46 1982
[49] Y-Q Wang B-P Tang H-M Zhang Q-H Zhou and G-C Zhang ldquoStudies on the interaction between imidaclopridand human serum albumin spectroscopic approachrdquo Journal ofPhotochemistry and Photobiology B vol 94 no 3 pp 183ndash1902009
[50] S-J Kim H-W Rhee H-Y Park H-S Kim and J-I HongldquoFluorescent probes designed for detecting human serum albu-min on the basis of its pseudo-esterase activityrdquo Bioorganic ampMedicinal Chemistry Letters vol 23 pp 2093ndash2097 2013
[51] P Ascenzi M Gioia G Fanali M Coletta and M FasanoldquoPseudo-enzymatic hydrolysis of 4-nitrophenyl acetate byhuman serum albumin pH-dependence of rates of individualstepsrdquo Biochemical and Biophysical Research Communicationsvol 424 pp 451ndash455 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
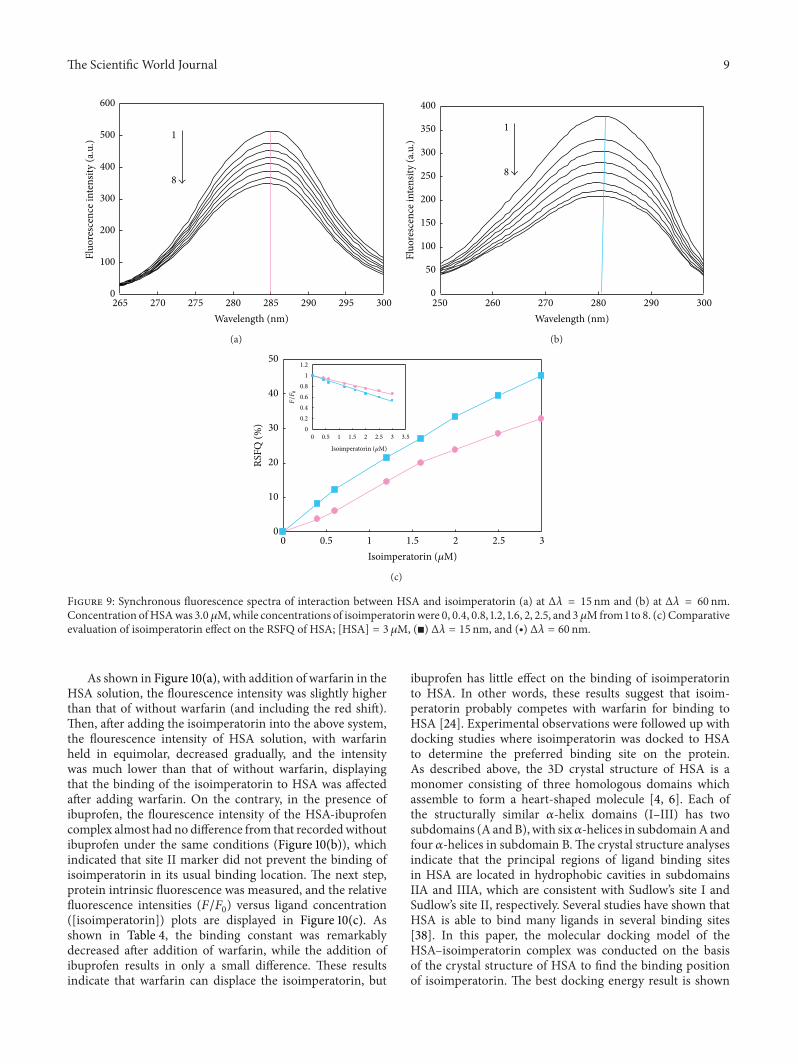
The Scientific World Journal 9
0
100
200
300
400
500
600
265 270 275 280 285 290 295 300Wavelength (nm)
1
8
Fluo
resc
ence
inte
nsity
(au
)
(a)
Fluo
resc
ence
inte
nsity
(au
)
0
50
100
150
200
250
300
350
400
250 260 270 280 290 300Wavelength (nm)
1
8
(b)
0
10
20
30
40
50
0 05 1 15 2 25 3
RSFQ
()
Isoimperatorin (120583M)
0
02
04
06
08
1
12
005 1 15 2 25 3 35
Isoimperatorin (120583M)
F0
F
(c)
Figure 9 Synchronous fluorescence spectra of interaction between HSA and isoimperatorin (a) at Δ120582 = 15 nm and (b) at Δ120582 = 60 nmConcentration ofHSAwas 30 120583Mwhile concentrations of isoimperatorinwere 0 04 08 12 16 2 25 and 3 120583Mfrom 1 to 8 (c) Comparativeevaluation of isoimperatorin effect on the RSFQ of HSA [HSA] = 3 120583M (◼) Δ120582 = 15 nm and (∙) Δ120582 = 60 nm
As shown in Figure 10(a) with addition of warfarin in theHSA solution the flourescence intensity was slightly higherthan that of without warfarin (and including the red shift)Then after adding the isoimperatorin into the above systemthe flourescence intensity of HSA solution with warfarinheld in equimolar decreased gradually and the intensitywas much lower than that of without warfarin displayingthat the binding of the isoimperatorin to HSA was affectedafter adding warfarin On the contrary in the presence ofibuprofen the flourescence intensity of the HSA-ibuprofencomplex almost had no difference from that recordedwithoutibuprofen under the same conditions (Figure 10(b)) whichindicated that site II marker did not prevent the binding ofisoimperatorin in its usual binding location The next stepprotein intrinsic fluorescence was measured and the relativefluorescence intensities (119865119865
0) versus ligand concentration
([isoimperatorin]) plots are displayed in Figure 10(c) Asshown in Table 4 the binding constant was remarkablydecreased after addition of warfarin while the addition ofibuprofen results in only a small difference These resultsindicate that warfarin can displace the isoimperatorin but
ibuprofen has little effect on the binding of isoimperatorinto HSA In other words these results suggest that isoim-peratorin probably competes with warfarin for binding toHSA [24] Experimental observations were followed up withdocking studies where isoimperatorin was docked to HSAto determine the preferred binding site on the proteinAs described above the 3D crystal structure of HSA is amonomer consisting of three homologous domains whichassemble to form a heart-shaped molecule [4 6] Each ofthe structurally similar 120572-helix domains (IndashIII) has twosubdomains (A andB) with six120572-helices in subdomainA andfour 120572-helices in subdomain BThe crystal structure analysesindicate that the principal regions of ligand binding sitesin HSA are located in hydrophobic cavities in subdomainsIIA and IIIA which are consistent with Sudlowrsquos site I andSudlowrsquos site II respectively Several studies have shown thatHSA is able to bind many ligands in several binding sites[38] In this paper the molecular docking model of theHSAndashisoimperatorin complex was conducted on the basisof the crystal structure of HSA to find the binding positionof isoimperatorin The best docking energy result is shown
10 The Scientific World Journal
0
100
200
300
400
500
600
310 335 360 385 410Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA1
8
(a)
0
100
200
300
400
500
600
700
305 325 345 365 385 405Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA
1
8
(b)
0
05
1
15
0 1 2 3
HSAIbuprofen
Warfarin
Isoimperatorin (120583M)
F0
F
(c)
Figure 10 Effect of isoimperatorin on the quenching of HSA-warfarin complex (a) andHSA-ibuprofen complex (b) fluorescence From 1 to 8[HSA] = [warfarin or iboprufen] = 3 120583M [isoimperatorin] = 0 04 08 12 16 2 25 and 3 120583M All reproduced data shown are representativeof at least 3 independent experiments (c) Fluorescence quenching profiles of HSA (⧫) HSA-ibobrofen (◼) and HSA-warfarin (998771) in thepresence of various concentration of isoimperatorin
in Figure 11 It can be seen that isoimperatorin was situatedwithin subdomain IIA in Sudlowrsquos site I formed by six 120572-helices This figure shows the location of Trp 214 and also thebinding of isoimperatorin in HSA As this fluorescent residueis in subdomain IIA and due to the proximity of the bindingsite of isoimperatorin to the Trp 214 it can be concluded thatisoimperatorin has one reactive site onHSAandprobably thissite is at or near the site I The above docking results are ingood agreement with the displacement experiments results
and revealed that isoimperatorin binds to Sudlowrsquos site I ofHSA
37 Effect of Isoimperatorin on HSA Esterase Activity Pre-liminary studies showed that albumin in addition to itsligand binding capabilities has remarkable promiscuouscatalytic activities toward a broad range of organicmoleculesincluding esters amides phosphates and benzisoxazolesHuman serum albumin had significantly greater esterase-like
The Scientific World Journal 11
Table 4 Binding constants for isoimperatorin-HSA interaction (at 295K) in the absence and presence of the site marker
119870
119887
times 10
minus5
(Mminus1) 119877
a
HSA + isoimperatorin (in the absence ofibuprofen and warfarin) 1110 plusmn 04
lowast 09947
HSA + isoimperatorin + ibuprofen 1099 plusmn 06
lowast 09904HSA + isoimperatorin + warfarin 789 plusmn 03
lowast 09881Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870
119887values lowast119875 value lt 00002
Trp 214
Isoimperatorin
Figure 11 Crystal structure of HSA and the positions of singletryptophan residue (Trp 214) and also the binding site of isoim-peratorin in HSA Protein backbone is shown in the ldquocartoonrdquorepresentation and isoimperatorinand Trp 214 residue in the site Iare shown in the ldquostickrdquo representation The isoimperatorin and Trp214 structures are represented by the red and blue colorsThe crystalstructure of protein was obtained from Protein Data Bank (1BM0)For interpretation of references to color in this figure legend thereader is referred to the web version of this paper
activity than bovine serum albumin and defatted albuminwas more active than untreated albumin [50] InterestinglyHSA exhibits esterase-like activity and hydrolyzes drugshaving an ester group such as aspirin (which can convertaspirin ldquoacetylsalicylic acidrdquo to salicylic acid) Lys199 Lys402Lys519 and Lys545 form the pseudoesterase active site ofHSAthat is located in site I [50] ldquoEsterase-likerdquo activity (hydrolysisof p-nitrophenol esters such as p-NPA) by subdomain III Ahas been also reported and site directed mutagenesis studieshave shown that Arg 410 and Tyr 411 are essential in thisesterase activity However further investigation revealed thatacetylation is not limited to a distinct side chain so that theHSA incubation with high concentrations of pNPAc resultsin the acetylation of 59 Lys 10 Ser 8 Thr 4 Tyr and 1 Asp[51] Because of the above difference here we set out toevaluate the effect of isoimperatorin on the esterase activityof HSA based on our previous work [19] The kinetics ofthe pseudoenzymatic hydrolysis of pNPAc was obtained inthe absence and presence of isoimperatorin and site markers(Figure 12) Accordingly warfarin and indomethacin (as siteI marker) and isoimperatorin inhibited competitively theHSA-catalyzed hydrolysis of pNPAc by binding to site I Incontrast ibuprofen and tetracycline were unable to inhibitesterase activity of HSA The enzyme inhibition results sug-gested that isoimperatorin aspirin warfarin indomethacin
0
005
01
015
02
025
03
035
0 05 1 15 2 25 3Time (min)
HSA TetracyclineIbuprofenAspirin
IndomethacinOstholeWarfarinIsoimperatorin
A400
Figure 12 4-Nitrophenyl acetate esterase activity of HSA in theabsence and presence of tetracycline and ibuprofen (site II) war-farin aspirin indomethacin and osthole (site I) and isoimperatorin(site I) Final concentration of protein pNPA and drugs were 15250 and 250 120583M respectively For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
and osthole binding sites on the HSA molecule are the sameThese results also proposed that the large site I on HSAparticipates effectively to its promiscuous esterase activity
4 Conclusion
The accumulated data on HSA-drug interactions can beused to develop quantitative structure-activity relationshipsfor albumin binding However as stated earlier adsorptiondistribution metabolism and elimination (ADME) is con-trolled by albumin [30] Poor or strong binding to SA isbelieved to affect half-time of drugs [35] and their dosagedepends on their half-life in plasma Thus the bindingaffinity of any drug to SA is one of the major factors thatdetermine the pharmacokinetics halftime and availabilityof the drug in various tissues (see [30] and referencestherein) Further complexities arise in vivo due to interactions
12 The Scientific World Journal
between drugs and endogenous ligands for HSA This isparticularly pertinent for fatty acids which normally occurin serum at levels between 01 and 2mol per mol of HSAand can both compete and cooperate with drugs bindingto the protein In certain disease states these effects areexacerbated as the fatty acid HSA molar ratio may be ashigh as six Other pathological conditions are associatedwith high (micromolar to millimolar) levels of bilirubinhemin or renal toxins (eg 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) indoxyl sulphate) which bindto the protein causing significant drug binding defects
Our major aim in this study was to distinguish thenature of the interaction of isoimperatorin to HSA usingdifferent biophysical techniques This study could be veryvaluable due to importance and presence of isoimperatorinin human diet (food-drug interaction on HSA) The resultsshowed that isoimperatorin quenched the intrinsic fluo-rescence of HSA through dynamic quenching mechanismFurthermore hydrophobic interaction played a major role inthe binding process Using Fluorescence and UV absorptionspectroscopy it has been shown that isoimperatorin inducesprotein structural changes The binding average distancebetween the solo tryptophan of the protein and the boundisoimperatorin was calculated by Forster theory as 290 plusmn02 nmAnalysis of PSHvalues elucidated that isoimperatorinhas induced some compactness in the tertiary structure ofthe enzyme The results of synchronous flourescence onHSA-isoimperatorin system elucidated that the drug binds tothe hydrophobic cavity (site I) on subdomain IIA that wasconfirmed by quenching experiments competitive bindingexperiments and docking technique
Acknowledgment
The authors gratefully acknowledge the Kermanshah Uni-versity of Medical Sciences Research Council for financialsupport of this work
References
[1] Y-J Hu Y Liu T-Q Sun A-M Bai J-Q Lu and Z-B Pi ldquoBinding of anti-inflammatory drug cromolyn sodiumto bovine serum albuminrdquo International Journal of BiologicalMacromolecules vol 39 no 4-5 pp 280ndash285 2006
[2] N Seedher and S Bhatia ldquoReversible binding of celecoxiband valdecoxib with human serum albumin using fluorescencespectroscopic techniquerdquo Pharmacological Research vol 54 no2 pp 77ndash84 2006
[3] H Isogai and N Hirayama ldquoIn silico prediction of interactionsbetween site II on human serum albumin and profen drugsrdquoISRN Pharma vol 2013 Article ID 818364 8 pages 2013
[4] YWangHYuX Shi Z LuoD Lin andMHuang ldquoStructuralmechanism of ring opening reaction of glucose by humanserum albuminrdquo The Journal of Biological Chemistry vol 288pp 15980ndash15987 2013
[5] F Keshavarz M M Alavianmehr and R Yousefi ldquoMolecularinteraction of benzalkonium Ibuprofenate and its discreteingredients with human serum albuminrdquo Physical ChemistryResearch vol 1 no 2 pp 111ndash116 2013
[6] T K Maiti K S Ghosh J Debnath and S Dasgupta ldquoBindingof all-trans retinoic acid to human serum albumin fluores-cence FT-IR and circular dichroism studiesrdquo InternationalJournal of Biological Macromolecules vol 38 no 3ndash5 pp 197ndash202 2006
[7] M-X XieM Long Y Liu C Qin and Y-DWang ldquoCharacter-ization of the interaction between human serum albumin andmorinrdquo Biochimica et Biophysica Acta vol 1760 no 8 pp 1184ndash1191 2006
[8] CD Kanakis P A TarantilisMG Polissiou S Diamantoglouand H A Tajmir-Riahi ldquoAntioxidant flavonoids bind humanserum albuminrdquo Journal of Molecular Structure vol 798 no 1-3 pp 69ndash74 2006
[9] S Marumoto and M Miyazawa ldquoStructure-activity rela-tionships for naturally occurring coumarins as 120573-secretaseinhibitorrdquo Bioorganic and Medicinal Chemistry vol 20 no 2pp 784ndash788 2012
[10] S Marumoto and M Miyazawa ldquoBiotransformation of isoim-peratorin and imperatorin by Glomerella cingulata and 120573-secretase inhibitory activityrdquo Bioorganic and Medicinal Chem-istry vol 18 no 1 pp 455ndash459 2010
[11] S E Sajjadi Y Shokoohinia and S Hemmati ldquoIsolation andidentification of furanocoumarins and a phenylpropanoid fromthe acetone extract and identification of volatile constituentsfrom the essential oil of Peucedanum pastinacifoliumrdquo Chem-istry of Natural Compounds vol 48 no 4 pp 668ndash671 2012
[12] X B Yang J Hou D Liu et al ldquoBiotransformation of isoim-peratorin by Cunninghamella blakesleana AS 3970rdquo Journal ofMolecular Catalysis B vol 88 pp 1ndash6 2013
[13] W Lili S Yehong S Qi et al ldquoIn vitro permeability analy-sis pharmacokinetic and brain distribution study in mice ofimperatorin isoimperatorin and cnidilin in Radix AngelicaeDahuricaerdquo Fitoterapia vol 85 pp 144ndash153 2013
[14] S Wang Q Chen and L He ldquoDevelopment and validationof a gas chromatography-mass spectrometry method for thedetermination of isoimperatorin in rat plasma and tissueapplication to the pharmacokinetic and tissue distributionstudyrdquo Journal of Chromatography B vol 852 no 1-2 pp 473ndash478 2007
[15] H Sadraei Y Shokoohinia S E Sajjadi and M MozafarildquoAntispasmodic effects of Prangos ferulacea acetone extract andits maincomponent osthole on ileum contractionrdquo Research inPharmaceutical Sciences vol 8 pp 137ndash144 2013
[16] H Sadraei Y Shokoohinia S E Sajjadi and B GhadirianldquoAntispasmodic effect of osthole and Prangos ferulacea extracton rat uterus smooth muscle motilityrdquo Research in Pharmaceu-tical Sciences vol 7 pp 141ndash149 2012
[17] S Sajjadi Y Shokoohinia S Gholamzadeh M Behbahani andA Fattahi ldquoAntiviral evaluation of coumarins from Prangosferulacea L (Lindl)rdquo Research in Pharmaceutical Sciences vol7 p 783 2012
[18] S E Sajjadi Y Shokoohinia and S Gholamzadeh ldquoChemicalcomposition of the essential oil of the root of Prangos ferulacea(L) Lindlrdquo Chemija vol 22 pp 178ndash180 2011
[19] N Bijari Y Shokoohinia M-R Ashrafi-Kooshk et al ldquoSpec-troscopic study of interaction between osthole and humanserum albumin identification of possible binding site of thecompoundrdquo Journal of Luminescence vol 143 pp 328ndash336 2013
[20] X Shi M Liu M Zhang et al ldquoIdentification of in vitro and invivo metabolites of isoimperatorin using liquid chromatogra-phymass spectrometryrdquo Food Chemistry vol 141 pp 357ndash3652013
The Scientific World Journal 13
[21] T Chatterjee A Pal S Dey B-K Chatterjee and PChakrabarti ldquoInteraction of virstatin with human serum albu-min spectroscopic analysis andmolecular modelingrdquo Plos Onevol 7 pp 1ndash12 2012
[22] Y-M Chen and L-H Guo ldquoCombined fluorescence and elec-trochemical investigation on the binding interaction betweenorganic acid and human serum albuminrdquo Journal of Environ-mental Sciences vol 21 no 3 pp 373ndash379 2009
[23] J R Lakowicz Principles of Fluorescence Spectroscopy KluwerAcademicPlenum Publishers New York NY USA 2nd edi-tion 1999
[24] F Ding N Li B Han F Liu L Zhang and Y Sun ldquoThe bindingof CI Acid Red 2 to human serum albumin determinationof binding mechanism and binding site using fluorescencespectroscopyrdquo Dyes and Pigments vol 83 no 2 pp 249ndash2572009
[25] M-R Ashrafi Kooshk K Mansouri M Nadi S Khodarahmiand R Khodarahmi ldquoIn vitro anti-cancer activity of nativecurcumin and protein-curcuminrdquo Systems vol 2 no 1 pp 80ndash88 1981
[26] P D Ross and S Subramanian ldquoThermodynamics of proteinassociation reactions forces contributing to stabilityrdquo Biochem-istry vol 20 no 11 pp 3096ndash3102 1981
[27] J Liu J-N Tian J Zhang Z Hu and X Chen ldquoInteractionof magnolol with bovine serum albumin a fluorescence-quenching studyrdquo Analytical and Bioanalytical Chemistry vol376 no 6 pp 864ndash867 2003
[28] A Coi A M Bianucci F Bonomi P Rasmussen G M Muraand M L Ganadu ldquoStructural perturbation of 120572B-crystallinby zinc and temperature related to its chaperone-like activityrdquoInternational Journal of Biological Macromolecules vol 42 no3 pp 229ndash234 2008
[29] M Moller and A Denicola ldquoStudy of protein-ligand bindingby fluorescencerdquoBiochemistry andMolecular Biology Educationvol 30 no 5 pp 309ndash312 2002
[30] R Khodarahmi S A Karimi M R Ashrafi Kooshk S AGhadami S Ghobadi and M Amani ldquoComparative spec-troscopic studies on drug binding characteristics and proteinsurface hydrophobicity of native and modified forms of bovineserum albumin possible relevance to change in protein struc-turefunction upon non-enzymatic glycationrdquo SpectrochimicaActa A vol 89 pp 177ndash186 2012
[31] S-F Ma M Anraku Y Iwao et al ldquoHydrolysis of angiotensinII receptor blocker prodrug olmesartan medoxomil by humanserum albumin and identification of its catalytic active sitesrdquoDrug Metabolism and Disposition vol 33 no 12 pp 1911ndash19192005
[32] H M Berman J Westbrook Z Feng et al ldquoThe protein databankrdquo Nucleic Acids Research vol 28 no 1 pp 235ndash242 2000
[33] S Sugio A Kashima SMochizukiMNoda andKKobayashildquoCrystal structure of human serum albumin at 25 A resolutionrdquoProtein Engineering vol 12 no 6 pp 439ndash446 1999
[34] G M Morris D S Goodsell R Huey and A J OlsonldquoDistributed automated docking of flexible ligands to proteinsparallel applications of AutoDock 24rdquo Journal of Computer-Aided Molecular Design vol 10 no 4 pp 293ndash304 1996
[35] F Faridbod M R Ganjali B Larijani et al ldquoInteraction studyof pioglitazone with albumin by fluorescence spectroscopy andmolecular dockingrdquo Spectrochimica Acta A vol 78 no 1 pp96ndash101 2011
[36] D SGoodsell andA JOlson ldquoAutomated docking of substratesto proteins by simulated annealingrdquo Proteins vol 8 no 3 pp195ndash202 1990
[37] D S Goodsell G M Morris and A J Olson ldquoAutomateddocking of flexible ligands applications of AutoDockrdquo Journalof Molecular Recognition vol 9 no 1 pp 1ndash5 1996
[38] G Sudlow D J Birkett and D N Wade ldquoThe characterizationof two specific drug binding sites on human serum albuminrdquoMolecular Pharmacology vol 11 no 6 pp 824ndash832 1975
[39] Y Yue X Chen J Qin and X Yao ldquoCharacterization of themangiferin-human serum albumin complex by spectroscopicandmolecularmodeling approachesrdquo Journal of Pharmaceuticaland Biomedical Analysis vol 49 no 3 pp 753ndash759 2009
[40] A K Shaw and S K Pal ldquoSpectroscopic studies on the effectof temperature on pH-induced folded states of human serumalbuminrdquo Journal of Photochemistry and Photobiology B vol 90no 1 pp 69ndash77 2008
[41] S Ranjbar S Ghobadi R Khodarahmi and H Nemati ldquoSpec-troscopic characterization of furosemide binding to humancarbonic anhydrase IIrdquo International Journal of BiologicalMacromolecules vol 50 no 4 pp 910ndash917 2012
[42] HNHou Z POi YWOYung F L Liao Y Zhang andY LiyldquoStudies on interaction between vitamin B12 and human serumalbuminrdquo Journal of Pharmaceutical and Biomedical Analysisvol 47 pp 134ndash139 2008
[43] S S Kalanur J Seetharamappa and V K A Kalalbandi ldquoChar-acterization of interaction and the effect of carbamazepine onthe structure of human serum albuminrdquo Journal of Pharmaceu-tical and Biomedical Analysis vol 53 no 3 pp 660ndash666 2010
[44] L Stryer ldquoFluorescence spectroscopy of proteinsrdquo Science vol162 no 3853 pp 526ndash533 1968
[45] B Valeur and J C Brochon New Trends in FluorescenceSpectroscopy Springer Berlin Germany 1999
[46] V Suryawanshi P Anbhule A Gore Sh Patil and G AndKolekar ldquoA spectral deciphering the perturbation of modeltransporter protein (HSA) by antibacterial pyrimidine deriva-tive pharmacokinetic and biophysical insightsrdquo Journal ofPhotochemistry and Photobiology B vol 118 pp 1ndash8 2013
[47] T Yuan AMWeljie andH J Vogel ldquoTryptophan fluorescencequenching by methionine and selenomethionine residues ofcalmodulin orientation of peptide and protein bindingrdquo Bio-chemistry vol 37 no 9 pp 3187ndash3195 1998
[48] TCOrsquoHaver A F Fell G Smith et al ldquoDerivative spectroscopyand its applications in analysisrdquo Analytical Proceedings vol 19no 1 pp 22ndash46 1982
[49] Y-Q Wang B-P Tang H-M Zhang Q-H Zhou and G-C Zhang ldquoStudies on the interaction between imidaclopridand human serum albumin spectroscopic approachrdquo Journal ofPhotochemistry and Photobiology B vol 94 no 3 pp 183ndash1902009
[50] S-J Kim H-W Rhee H-Y Park H-S Kim and J-I HongldquoFluorescent probes designed for detecting human serum albu-min on the basis of its pseudo-esterase activityrdquo Bioorganic ampMedicinal Chemistry Letters vol 23 pp 2093ndash2097 2013
[51] P Ascenzi M Gioia G Fanali M Coletta and M FasanoldquoPseudo-enzymatic hydrolysis of 4-nitrophenyl acetate byhuman serum albumin pH-dependence of rates of individualstepsrdquo Biochemical and Biophysical Research Communicationsvol 424 pp 451ndash455 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
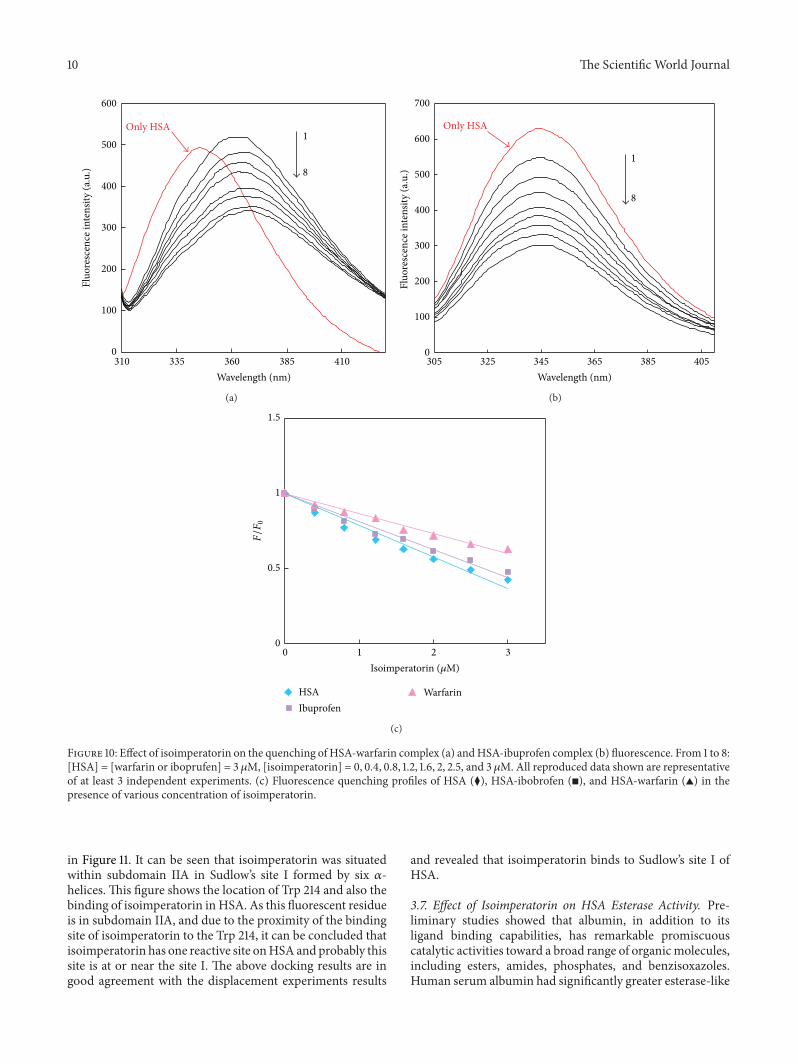
10 The Scientific World Journal
0
100
200
300
400
500
600
310 335 360 385 410Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA1
8
(a)
0
100
200
300
400
500
600
700
305 325 345 365 385 405Wavelength (nm)
Fluo
resc
ence
inte
nsity
(au
)
Only HSA
1
8
(b)
0
05
1
15
0 1 2 3
HSAIbuprofen
Warfarin
Isoimperatorin (120583M)
F0
F
(c)
Figure 10 Effect of isoimperatorin on the quenching of HSA-warfarin complex (a) andHSA-ibuprofen complex (b) fluorescence From 1 to 8[HSA] = [warfarin or iboprufen] = 3 120583M [isoimperatorin] = 0 04 08 12 16 2 25 and 3 120583M All reproduced data shown are representativeof at least 3 independent experiments (c) Fluorescence quenching profiles of HSA (⧫) HSA-ibobrofen (◼) and HSA-warfarin (998771) in thepresence of various concentration of isoimperatorin
in Figure 11 It can be seen that isoimperatorin was situatedwithin subdomain IIA in Sudlowrsquos site I formed by six 120572-helices This figure shows the location of Trp 214 and also thebinding of isoimperatorin in HSA As this fluorescent residueis in subdomain IIA and due to the proximity of the bindingsite of isoimperatorin to the Trp 214 it can be concluded thatisoimperatorin has one reactive site onHSAandprobably thissite is at or near the site I The above docking results are ingood agreement with the displacement experiments results
and revealed that isoimperatorin binds to Sudlowrsquos site I ofHSA
37 Effect of Isoimperatorin on HSA Esterase Activity Pre-liminary studies showed that albumin in addition to itsligand binding capabilities has remarkable promiscuouscatalytic activities toward a broad range of organicmoleculesincluding esters amides phosphates and benzisoxazolesHuman serum albumin had significantly greater esterase-like
The Scientific World Journal 11
Table 4 Binding constants for isoimperatorin-HSA interaction (at 295K) in the absence and presence of the site marker
119870
119887
times 10
minus5
(Mminus1) 119877
a
HSA + isoimperatorin (in the absence ofibuprofen and warfarin) 1110 plusmn 04
lowast 09947
HSA + isoimperatorin + ibuprofen 1099 plusmn 06
lowast 09904HSA + isoimperatorin + warfarin 789 plusmn 03
lowast 09881Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870
119887values lowast119875 value lt 00002
Trp 214
Isoimperatorin
Figure 11 Crystal structure of HSA and the positions of singletryptophan residue (Trp 214) and also the binding site of isoim-peratorin in HSA Protein backbone is shown in the ldquocartoonrdquorepresentation and isoimperatorinand Trp 214 residue in the site Iare shown in the ldquostickrdquo representation The isoimperatorin and Trp214 structures are represented by the red and blue colorsThe crystalstructure of protein was obtained from Protein Data Bank (1BM0)For interpretation of references to color in this figure legend thereader is referred to the web version of this paper
activity than bovine serum albumin and defatted albuminwas more active than untreated albumin [50] InterestinglyHSA exhibits esterase-like activity and hydrolyzes drugshaving an ester group such as aspirin (which can convertaspirin ldquoacetylsalicylic acidrdquo to salicylic acid) Lys199 Lys402Lys519 and Lys545 form the pseudoesterase active site ofHSAthat is located in site I [50] ldquoEsterase-likerdquo activity (hydrolysisof p-nitrophenol esters such as p-NPA) by subdomain III Ahas been also reported and site directed mutagenesis studieshave shown that Arg 410 and Tyr 411 are essential in thisesterase activity However further investigation revealed thatacetylation is not limited to a distinct side chain so that theHSA incubation with high concentrations of pNPAc resultsin the acetylation of 59 Lys 10 Ser 8 Thr 4 Tyr and 1 Asp[51] Because of the above difference here we set out toevaluate the effect of isoimperatorin on the esterase activityof HSA based on our previous work [19] The kinetics ofthe pseudoenzymatic hydrolysis of pNPAc was obtained inthe absence and presence of isoimperatorin and site markers(Figure 12) Accordingly warfarin and indomethacin (as siteI marker) and isoimperatorin inhibited competitively theHSA-catalyzed hydrolysis of pNPAc by binding to site I Incontrast ibuprofen and tetracycline were unable to inhibitesterase activity of HSA The enzyme inhibition results sug-gested that isoimperatorin aspirin warfarin indomethacin
0
005
01
015
02
025
03
035
0 05 1 15 2 25 3Time (min)
HSA TetracyclineIbuprofenAspirin
IndomethacinOstholeWarfarinIsoimperatorin
A400
Figure 12 4-Nitrophenyl acetate esterase activity of HSA in theabsence and presence of tetracycline and ibuprofen (site II) war-farin aspirin indomethacin and osthole (site I) and isoimperatorin(site I) Final concentration of protein pNPA and drugs were 15250 and 250 120583M respectively For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
and osthole binding sites on the HSA molecule are the sameThese results also proposed that the large site I on HSAparticipates effectively to its promiscuous esterase activity
4 Conclusion
The accumulated data on HSA-drug interactions can beused to develop quantitative structure-activity relationshipsfor albumin binding However as stated earlier adsorptiondistribution metabolism and elimination (ADME) is con-trolled by albumin [30] Poor or strong binding to SA isbelieved to affect half-time of drugs [35] and their dosagedepends on their half-life in plasma Thus the bindingaffinity of any drug to SA is one of the major factors thatdetermine the pharmacokinetics halftime and availabilityof the drug in various tissues (see [30] and referencestherein) Further complexities arise in vivo due to interactions
12 The Scientific World Journal
between drugs and endogenous ligands for HSA This isparticularly pertinent for fatty acids which normally occurin serum at levels between 01 and 2mol per mol of HSAand can both compete and cooperate with drugs bindingto the protein In certain disease states these effects areexacerbated as the fatty acid HSA molar ratio may be ashigh as six Other pathological conditions are associatedwith high (micromolar to millimolar) levels of bilirubinhemin or renal toxins (eg 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) indoxyl sulphate) which bindto the protein causing significant drug binding defects
Our major aim in this study was to distinguish thenature of the interaction of isoimperatorin to HSA usingdifferent biophysical techniques This study could be veryvaluable due to importance and presence of isoimperatorinin human diet (food-drug interaction on HSA) The resultsshowed that isoimperatorin quenched the intrinsic fluo-rescence of HSA through dynamic quenching mechanismFurthermore hydrophobic interaction played a major role inthe binding process Using Fluorescence and UV absorptionspectroscopy it has been shown that isoimperatorin inducesprotein structural changes The binding average distancebetween the solo tryptophan of the protein and the boundisoimperatorin was calculated by Forster theory as 290 plusmn02 nmAnalysis of PSHvalues elucidated that isoimperatorinhas induced some compactness in the tertiary structure ofthe enzyme The results of synchronous flourescence onHSA-isoimperatorin system elucidated that the drug binds tothe hydrophobic cavity (site I) on subdomain IIA that wasconfirmed by quenching experiments competitive bindingexperiments and docking technique
Acknowledgment
The authors gratefully acknowledge the Kermanshah Uni-versity of Medical Sciences Research Council for financialsupport of this work
References
[1] Y-J Hu Y Liu T-Q Sun A-M Bai J-Q Lu and Z-B Pi ldquoBinding of anti-inflammatory drug cromolyn sodiumto bovine serum albuminrdquo International Journal of BiologicalMacromolecules vol 39 no 4-5 pp 280ndash285 2006
[2] N Seedher and S Bhatia ldquoReversible binding of celecoxiband valdecoxib with human serum albumin using fluorescencespectroscopic techniquerdquo Pharmacological Research vol 54 no2 pp 77ndash84 2006
[3] H Isogai and N Hirayama ldquoIn silico prediction of interactionsbetween site II on human serum albumin and profen drugsrdquoISRN Pharma vol 2013 Article ID 818364 8 pages 2013
[4] YWangHYuX Shi Z LuoD Lin andMHuang ldquoStructuralmechanism of ring opening reaction of glucose by humanserum albuminrdquo The Journal of Biological Chemistry vol 288pp 15980ndash15987 2013
[5] F Keshavarz M M Alavianmehr and R Yousefi ldquoMolecularinteraction of benzalkonium Ibuprofenate and its discreteingredients with human serum albuminrdquo Physical ChemistryResearch vol 1 no 2 pp 111ndash116 2013
[6] T K Maiti K S Ghosh J Debnath and S Dasgupta ldquoBindingof all-trans retinoic acid to human serum albumin fluores-cence FT-IR and circular dichroism studiesrdquo InternationalJournal of Biological Macromolecules vol 38 no 3ndash5 pp 197ndash202 2006
[7] M-X XieM Long Y Liu C Qin and Y-DWang ldquoCharacter-ization of the interaction between human serum albumin andmorinrdquo Biochimica et Biophysica Acta vol 1760 no 8 pp 1184ndash1191 2006
[8] CD Kanakis P A TarantilisMG Polissiou S Diamantoglouand H A Tajmir-Riahi ldquoAntioxidant flavonoids bind humanserum albuminrdquo Journal of Molecular Structure vol 798 no 1-3 pp 69ndash74 2006
[9] S Marumoto and M Miyazawa ldquoStructure-activity rela-tionships for naturally occurring coumarins as 120573-secretaseinhibitorrdquo Bioorganic and Medicinal Chemistry vol 20 no 2pp 784ndash788 2012
[10] S Marumoto and M Miyazawa ldquoBiotransformation of isoim-peratorin and imperatorin by Glomerella cingulata and 120573-secretase inhibitory activityrdquo Bioorganic and Medicinal Chem-istry vol 18 no 1 pp 455ndash459 2010
[11] S E Sajjadi Y Shokoohinia and S Hemmati ldquoIsolation andidentification of furanocoumarins and a phenylpropanoid fromthe acetone extract and identification of volatile constituentsfrom the essential oil of Peucedanum pastinacifoliumrdquo Chem-istry of Natural Compounds vol 48 no 4 pp 668ndash671 2012
[12] X B Yang J Hou D Liu et al ldquoBiotransformation of isoim-peratorin by Cunninghamella blakesleana AS 3970rdquo Journal ofMolecular Catalysis B vol 88 pp 1ndash6 2013
[13] W Lili S Yehong S Qi et al ldquoIn vitro permeability analy-sis pharmacokinetic and brain distribution study in mice ofimperatorin isoimperatorin and cnidilin in Radix AngelicaeDahuricaerdquo Fitoterapia vol 85 pp 144ndash153 2013
[14] S Wang Q Chen and L He ldquoDevelopment and validationof a gas chromatography-mass spectrometry method for thedetermination of isoimperatorin in rat plasma and tissueapplication to the pharmacokinetic and tissue distributionstudyrdquo Journal of Chromatography B vol 852 no 1-2 pp 473ndash478 2007
[15] H Sadraei Y Shokoohinia S E Sajjadi and M MozafarildquoAntispasmodic effects of Prangos ferulacea acetone extract andits maincomponent osthole on ileum contractionrdquo Research inPharmaceutical Sciences vol 8 pp 137ndash144 2013
[16] H Sadraei Y Shokoohinia S E Sajjadi and B GhadirianldquoAntispasmodic effect of osthole and Prangos ferulacea extracton rat uterus smooth muscle motilityrdquo Research in Pharmaceu-tical Sciences vol 7 pp 141ndash149 2012
[17] S Sajjadi Y Shokoohinia S Gholamzadeh M Behbahani andA Fattahi ldquoAntiviral evaluation of coumarins from Prangosferulacea L (Lindl)rdquo Research in Pharmaceutical Sciences vol7 p 783 2012
[18] S E Sajjadi Y Shokoohinia and S Gholamzadeh ldquoChemicalcomposition of the essential oil of the root of Prangos ferulacea(L) Lindlrdquo Chemija vol 22 pp 178ndash180 2011
[19] N Bijari Y Shokoohinia M-R Ashrafi-Kooshk et al ldquoSpec-troscopic study of interaction between osthole and humanserum albumin identification of possible binding site of thecompoundrdquo Journal of Luminescence vol 143 pp 328ndash336 2013
[20] X Shi M Liu M Zhang et al ldquoIdentification of in vitro and invivo metabolites of isoimperatorin using liquid chromatogra-phymass spectrometryrdquo Food Chemistry vol 141 pp 357ndash3652013
The Scientific World Journal 13
[21] T Chatterjee A Pal S Dey B-K Chatterjee and PChakrabarti ldquoInteraction of virstatin with human serum albu-min spectroscopic analysis andmolecular modelingrdquo Plos Onevol 7 pp 1ndash12 2012
[22] Y-M Chen and L-H Guo ldquoCombined fluorescence and elec-trochemical investigation on the binding interaction betweenorganic acid and human serum albuminrdquo Journal of Environ-mental Sciences vol 21 no 3 pp 373ndash379 2009
[23] J R Lakowicz Principles of Fluorescence Spectroscopy KluwerAcademicPlenum Publishers New York NY USA 2nd edi-tion 1999
[24] F Ding N Li B Han F Liu L Zhang and Y Sun ldquoThe bindingof CI Acid Red 2 to human serum albumin determinationof binding mechanism and binding site using fluorescencespectroscopyrdquo Dyes and Pigments vol 83 no 2 pp 249ndash2572009
[25] M-R Ashrafi Kooshk K Mansouri M Nadi S Khodarahmiand R Khodarahmi ldquoIn vitro anti-cancer activity of nativecurcumin and protein-curcuminrdquo Systems vol 2 no 1 pp 80ndash88 1981
[26] P D Ross and S Subramanian ldquoThermodynamics of proteinassociation reactions forces contributing to stabilityrdquo Biochem-istry vol 20 no 11 pp 3096ndash3102 1981
[27] J Liu J-N Tian J Zhang Z Hu and X Chen ldquoInteractionof magnolol with bovine serum albumin a fluorescence-quenching studyrdquo Analytical and Bioanalytical Chemistry vol376 no 6 pp 864ndash867 2003
[28] A Coi A M Bianucci F Bonomi P Rasmussen G M Muraand M L Ganadu ldquoStructural perturbation of 120572B-crystallinby zinc and temperature related to its chaperone-like activityrdquoInternational Journal of Biological Macromolecules vol 42 no3 pp 229ndash234 2008
[29] M Moller and A Denicola ldquoStudy of protein-ligand bindingby fluorescencerdquoBiochemistry andMolecular Biology Educationvol 30 no 5 pp 309ndash312 2002
[30] R Khodarahmi S A Karimi M R Ashrafi Kooshk S AGhadami S Ghobadi and M Amani ldquoComparative spec-troscopic studies on drug binding characteristics and proteinsurface hydrophobicity of native and modified forms of bovineserum albumin possible relevance to change in protein struc-turefunction upon non-enzymatic glycationrdquo SpectrochimicaActa A vol 89 pp 177ndash186 2012
[31] S-F Ma M Anraku Y Iwao et al ldquoHydrolysis of angiotensinII receptor blocker prodrug olmesartan medoxomil by humanserum albumin and identification of its catalytic active sitesrdquoDrug Metabolism and Disposition vol 33 no 12 pp 1911ndash19192005
[32] H M Berman J Westbrook Z Feng et al ldquoThe protein databankrdquo Nucleic Acids Research vol 28 no 1 pp 235ndash242 2000
[33] S Sugio A Kashima SMochizukiMNoda andKKobayashildquoCrystal structure of human serum albumin at 25 A resolutionrdquoProtein Engineering vol 12 no 6 pp 439ndash446 1999
[34] G M Morris D S Goodsell R Huey and A J OlsonldquoDistributed automated docking of flexible ligands to proteinsparallel applications of AutoDock 24rdquo Journal of Computer-Aided Molecular Design vol 10 no 4 pp 293ndash304 1996
[35] F Faridbod M R Ganjali B Larijani et al ldquoInteraction studyof pioglitazone with albumin by fluorescence spectroscopy andmolecular dockingrdquo Spectrochimica Acta A vol 78 no 1 pp96ndash101 2011
[36] D SGoodsell andA JOlson ldquoAutomated docking of substratesto proteins by simulated annealingrdquo Proteins vol 8 no 3 pp195ndash202 1990
[37] D S Goodsell G M Morris and A J Olson ldquoAutomateddocking of flexible ligands applications of AutoDockrdquo Journalof Molecular Recognition vol 9 no 1 pp 1ndash5 1996
[38] G Sudlow D J Birkett and D N Wade ldquoThe characterizationof two specific drug binding sites on human serum albuminrdquoMolecular Pharmacology vol 11 no 6 pp 824ndash832 1975
[39] Y Yue X Chen J Qin and X Yao ldquoCharacterization of themangiferin-human serum albumin complex by spectroscopicandmolecularmodeling approachesrdquo Journal of Pharmaceuticaland Biomedical Analysis vol 49 no 3 pp 753ndash759 2009
[40] A K Shaw and S K Pal ldquoSpectroscopic studies on the effectof temperature on pH-induced folded states of human serumalbuminrdquo Journal of Photochemistry and Photobiology B vol 90no 1 pp 69ndash77 2008
[41] S Ranjbar S Ghobadi R Khodarahmi and H Nemati ldquoSpec-troscopic characterization of furosemide binding to humancarbonic anhydrase IIrdquo International Journal of BiologicalMacromolecules vol 50 no 4 pp 910ndash917 2012
[42] HNHou Z POi YWOYung F L Liao Y Zhang andY LiyldquoStudies on interaction between vitamin B12 and human serumalbuminrdquo Journal of Pharmaceutical and Biomedical Analysisvol 47 pp 134ndash139 2008
[43] S S Kalanur J Seetharamappa and V K A Kalalbandi ldquoChar-acterization of interaction and the effect of carbamazepine onthe structure of human serum albuminrdquo Journal of Pharmaceu-tical and Biomedical Analysis vol 53 no 3 pp 660ndash666 2010
[44] L Stryer ldquoFluorescence spectroscopy of proteinsrdquo Science vol162 no 3853 pp 526ndash533 1968
[45] B Valeur and J C Brochon New Trends in FluorescenceSpectroscopy Springer Berlin Germany 1999
[46] V Suryawanshi P Anbhule A Gore Sh Patil and G AndKolekar ldquoA spectral deciphering the perturbation of modeltransporter protein (HSA) by antibacterial pyrimidine deriva-tive pharmacokinetic and biophysical insightsrdquo Journal ofPhotochemistry and Photobiology B vol 118 pp 1ndash8 2013
[47] T Yuan AMWeljie andH J Vogel ldquoTryptophan fluorescencequenching by methionine and selenomethionine residues ofcalmodulin orientation of peptide and protein bindingrdquo Bio-chemistry vol 37 no 9 pp 3187ndash3195 1998
[48] TCOrsquoHaver A F Fell G Smith et al ldquoDerivative spectroscopyand its applications in analysisrdquo Analytical Proceedings vol 19no 1 pp 22ndash46 1982
[49] Y-Q Wang B-P Tang H-M Zhang Q-H Zhou and G-C Zhang ldquoStudies on the interaction between imidaclopridand human serum albumin spectroscopic approachrdquo Journal ofPhotochemistry and Photobiology B vol 94 no 3 pp 183ndash1902009
[50] S-J Kim H-W Rhee H-Y Park H-S Kim and J-I HongldquoFluorescent probes designed for detecting human serum albu-min on the basis of its pseudo-esterase activityrdquo Bioorganic ampMedicinal Chemistry Letters vol 23 pp 2093ndash2097 2013
[51] P Ascenzi M Gioia G Fanali M Coletta and M FasanoldquoPseudo-enzymatic hydrolysis of 4-nitrophenyl acetate byhuman serum albumin pH-dependence of rates of individualstepsrdquo Biochemical and Biophysical Research Communicationsvol 424 pp 451ndash455 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
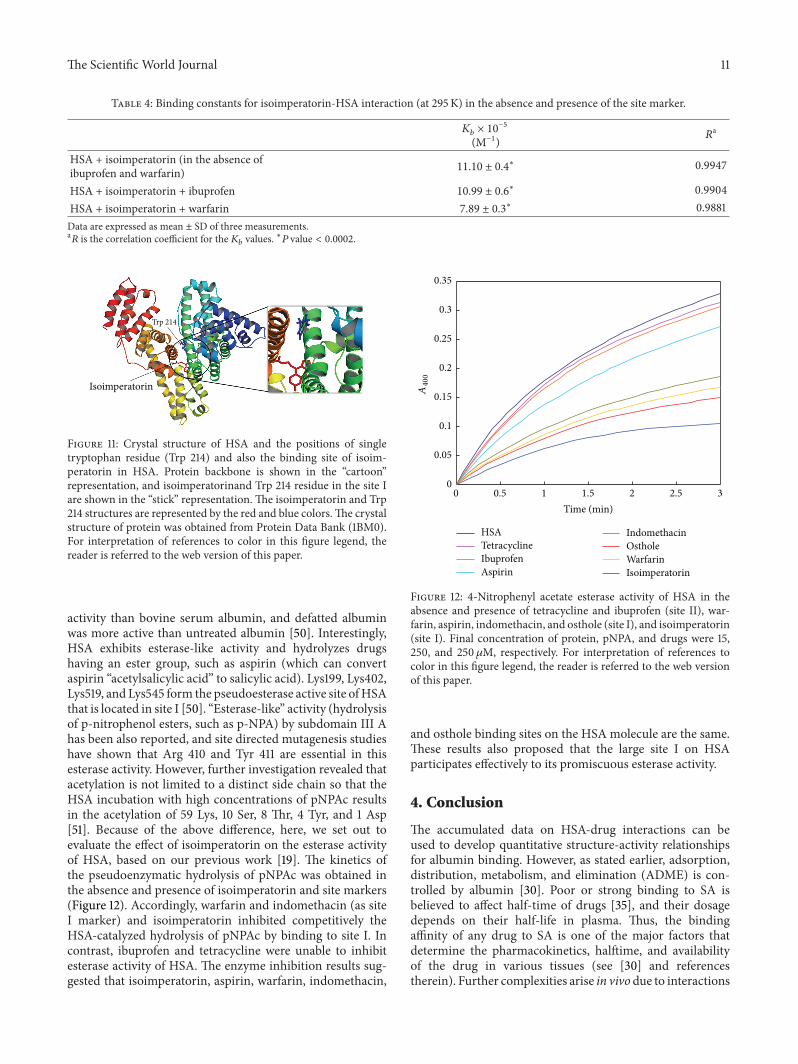
The Scientific World Journal 11
Table 4 Binding constants for isoimperatorin-HSA interaction (at 295K) in the absence and presence of the site marker
119870
119887
times 10
minus5
(Mminus1) 119877
a
HSA + isoimperatorin (in the absence ofibuprofen and warfarin) 1110 plusmn 04
lowast 09947
HSA + isoimperatorin + ibuprofen 1099 plusmn 06
lowast 09904HSA + isoimperatorin + warfarin 789 plusmn 03
lowast 09881Data are expressed as mean plusmn SD of three measurementsa119877 is the correlation coefficient for the119870
119887values lowast119875 value lt 00002
Trp 214
Isoimperatorin
Figure 11 Crystal structure of HSA and the positions of singletryptophan residue (Trp 214) and also the binding site of isoim-peratorin in HSA Protein backbone is shown in the ldquocartoonrdquorepresentation and isoimperatorinand Trp 214 residue in the site Iare shown in the ldquostickrdquo representation The isoimperatorin and Trp214 structures are represented by the red and blue colorsThe crystalstructure of protein was obtained from Protein Data Bank (1BM0)For interpretation of references to color in this figure legend thereader is referred to the web version of this paper
activity than bovine serum albumin and defatted albuminwas more active than untreated albumin [50] InterestinglyHSA exhibits esterase-like activity and hydrolyzes drugshaving an ester group such as aspirin (which can convertaspirin ldquoacetylsalicylic acidrdquo to salicylic acid) Lys199 Lys402Lys519 and Lys545 form the pseudoesterase active site ofHSAthat is located in site I [50] ldquoEsterase-likerdquo activity (hydrolysisof p-nitrophenol esters such as p-NPA) by subdomain III Ahas been also reported and site directed mutagenesis studieshave shown that Arg 410 and Tyr 411 are essential in thisesterase activity However further investigation revealed thatacetylation is not limited to a distinct side chain so that theHSA incubation with high concentrations of pNPAc resultsin the acetylation of 59 Lys 10 Ser 8 Thr 4 Tyr and 1 Asp[51] Because of the above difference here we set out toevaluate the effect of isoimperatorin on the esterase activityof HSA based on our previous work [19] The kinetics ofthe pseudoenzymatic hydrolysis of pNPAc was obtained inthe absence and presence of isoimperatorin and site markers(Figure 12) Accordingly warfarin and indomethacin (as siteI marker) and isoimperatorin inhibited competitively theHSA-catalyzed hydrolysis of pNPAc by binding to site I Incontrast ibuprofen and tetracycline were unable to inhibitesterase activity of HSA The enzyme inhibition results sug-gested that isoimperatorin aspirin warfarin indomethacin
0
005
01
015
02
025
03
035
0 05 1 15 2 25 3Time (min)
HSA TetracyclineIbuprofenAspirin
IndomethacinOstholeWarfarinIsoimperatorin
A400
Figure 12 4-Nitrophenyl acetate esterase activity of HSA in theabsence and presence of tetracycline and ibuprofen (site II) war-farin aspirin indomethacin and osthole (site I) and isoimperatorin(site I) Final concentration of protein pNPA and drugs were 15250 and 250 120583M respectively For interpretation of references tocolor in this figure legend the reader is referred to the web versionof this paper
and osthole binding sites on the HSA molecule are the sameThese results also proposed that the large site I on HSAparticipates effectively to its promiscuous esterase activity
4 Conclusion
The accumulated data on HSA-drug interactions can beused to develop quantitative structure-activity relationshipsfor albumin binding However as stated earlier adsorptiondistribution metabolism and elimination (ADME) is con-trolled by albumin [30] Poor or strong binding to SA isbelieved to affect half-time of drugs [35] and their dosagedepends on their half-life in plasma Thus the bindingaffinity of any drug to SA is one of the major factors thatdetermine the pharmacokinetics halftime and availabilityof the drug in various tissues (see [30] and referencestherein) Further complexities arise in vivo due to interactions
12 The Scientific World Journal
between drugs and endogenous ligands for HSA This isparticularly pertinent for fatty acids which normally occurin serum at levels between 01 and 2mol per mol of HSAand can both compete and cooperate with drugs bindingto the protein In certain disease states these effects areexacerbated as the fatty acid HSA molar ratio may be ashigh as six Other pathological conditions are associatedwith high (micromolar to millimolar) levels of bilirubinhemin or renal toxins (eg 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) indoxyl sulphate) which bindto the protein causing significant drug binding defects
Our major aim in this study was to distinguish thenature of the interaction of isoimperatorin to HSA usingdifferent biophysical techniques This study could be veryvaluable due to importance and presence of isoimperatorinin human diet (food-drug interaction on HSA) The resultsshowed that isoimperatorin quenched the intrinsic fluo-rescence of HSA through dynamic quenching mechanismFurthermore hydrophobic interaction played a major role inthe binding process Using Fluorescence and UV absorptionspectroscopy it has been shown that isoimperatorin inducesprotein structural changes The binding average distancebetween the solo tryptophan of the protein and the boundisoimperatorin was calculated by Forster theory as 290 plusmn02 nmAnalysis of PSHvalues elucidated that isoimperatorinhas induced some compactness in the tertiary structure ofthe enzyme The results of synchronous flourescence onHSA-isoimperatorin system elucidated that the drug binds tothe hydrophobic cavity (site I) on subdomain IIA that wasconfirmed by quenching experiments competitive bindingexperiments and docking technique
Acknowledgment
The authors gratefully acknowledge the Kermanshah Uni-versity of Medical Sciences Research Council for financialsupport of this work
References
[1] Y-J Hu Y Liu T-Q Sun A-M Bai J-Q Lu and Z-B Pi ldquoBinding of anti-inflammatory drug cromolyn sodiumto bovine serum albuminrdquo International Journal of BiologicalMacromolecules vol 39 no 4-5 pp 280ndash285 2006
[2] N Seedher and S Bhatia ldquoReversible binding of celecoxiband valdecoxib with human serum albumin using fluorescencespectroscopic techniquerdquo Pharmacological Research vol 54 no2 pp 77ndash84 2006
[3] H Isogai and N Hirayama ldquoIn silico prediction of interactionsbetween site II on human serum albumin and profen drugsrdquoISRN Pharma vol 2013 Article ID 818364 8 pages 2013
[4] YWangHYuX Shi Z LuoD Lin andMHuang ldquoStructuralmechanism of ring opening reaction of glucose by humanserum albuminrdquo The Journal of Biological Chemistry vol 288pp 15980ndash15987 2013
[5] F Keshavarz M M Alavianmehr and R Yousefi ldquoMolecularinteraction of benzalkonium Ibuprofenate and its discreteingredients with human serum albuminrdquo Physical ChemistryResearch vol 1 no 2 pp 111ndash116 2013
[6] T K Maiti K S Ghosh J Debnath and S Dasgupta ldquoBindingof all-trans retinoic acid to human serum albumin fluores-cence FT-IR and circular dichroism studiesrdquo InternationalJournal of Biological Macromolecules vol 38 no 3ndash5 pp 197ndash202 2006
[7] M-X XieM Long Y Liu C Qin and Y-DWang ldquoCharacter-ization of the interaction between human serum albumin andmorinrdquo Biochimica et Biophysica Acta vol 1760 no 8 pp 1184ndash1191 2006
[8] CD Kanakis P A TarantilisMG Polissiou S Diamantoglouand H A Tajmir-Riahi ldquoAntioxidant flavonoids bind humanserum albuminrdquo Journal of Molecular Structure vol 798 no 1-3 pp 69ndash74 2006
[9] S Marumoto and M Miyazawa ldquoStructure-activity rela-tionships for naturally occurring coumarins as 120573-secretaseinhibitorrdquo Bioorganic and Medicinal Chemistry vol 20 no 2pp 784ndash788 2012
[10] S Marumoto and M Miyazawa ldquoBiotransformation of isoim-peratorin and imperatorin by Glomerella cingulata and 120573-secretase inhibitory activityrdquo Bioorganic and Medicinal Chem-istry vol 18 no 1 pp 455ndash459 2010
[11] S E Sajjadi Y Shokoohinia and S Hemmati ldquoIsolation andidentification of furanocoumarins and a phenylpropanoid fromthe acetone extract and identification of volatile constituentsfrom the essential oil of Peucedanum pastinacifoliumrdquo Chem-istry of Natural Compounds vol 48 no 4 pp 668ndash671 2012
[12] X B Yang J Hou D Liu et al ldquoBiotransformation of isoim-peratorin by Cunninghamella blakesleana AS 3970rdquo Journal ofMolecular Catalysis B vol 88 pp 1ndash6 2013
[13] W Lili S Yehong S Qi et al ldquoIn vitro permeability analy-sis pharmacokinetic and brain distribution study in mice ofimperatorin isoimperatorin and cnidilin in Radix AngelicaeDahuricaerdquo Fitoterapia vol 85 pp 144ndash153 2013
[14] S Wang Q Chen and L He ldquoDevelopment and validationof a gas chromatography-mass spectrometry method for thedetermination of isoimperatorin in rat plasma and tissueapplication to the pharmacokinetic and tissue distributionstudyrdquo Journal of Chromatography B vol 852 no 1-2 pp 473ndash478 2007
[15] H Sadraei Y Shokoohinia S E Sajjadi and M MozafarildquoAntispasmodic effects of Prangos ferulacea acetone extract andits maincomponent osthole on ileum contractionrdquo Research inPharmaceutical Sciences vol 8 pp 137ndash144 2013
[16] H Sadraei Y Shokoohinia S E Sajjadi and B GhadirianldquoAntispasmodic effect of osthole and Prangos ferulacea extracton rat uterus smooth muscle motilityrdquo Research in Pharmaceu-tical Sciences vol 7 pp 141ndash149 2012
[17] S Sajjadi Y Shokoohinia S Gholamzadeh M Behbahani andA Fattahi ldquoAntiviral evaluation of coumarins from Prangosferulacea L (Lindl)rdquo Research in Pharmaceutical Sciences vol7 p 783 2012
[18] S E Sajjadi Y Shokoohinia and S Gholamzadeh ldquoChemicalcomposition of the essential oil of the root of Prangos ferulacea(L) Lindlrdquo Chemija vol 22 pp 178ndash180 2011
[19] N Bijari Y Shokoohinia M-R Ashrafi-Kooshk et al ldquoSpec-troscopic study of interaction between osthole and humanserum albumin identification of possible binding site of thecompoundrdquo Journal of Luminescence vol 143 pp 328ndash336 2013
[20] X Shi M Liu M Zhang et al ldquoIdentification of in vitro and invivo metabolites of isoimperatorin using liquid chromatogra-phymass spectrometryrdquo Food Chemistry vol 141 pp 357ndash3652013
The Scientific World Journal 13
[21] T Chatterjee A Pal S Dey B-K Chatterjee and PChakrabarti ldquoInteraction of virstatin with human serum albu-min spectroscopic analysis andmolecular modelingrdquo Plos Onevol 7 pp 1ndash12 2012
[22] Y-M Chen and L-H Guo ldquoCombined fluorescence and elec-trochemical investigation on the binding interaction betweenorganic acid and human serum albuminrdquo Journal of Environ-mental Sciences vol 21 no 3 pp 373ndash379 2009
[23] J R Lakowicz Principles of Fluorescence Spectroscopy KluwerAcademicPlenum Publishers New York NY USA 2nd edi-tion 1999
[24] F Ding N Li B Han F Liu L Zhang and Y Sun ldquoThe bindingof CI Acid Red 2 to human serum albumin determinationof binding mechanism and binding site using fluorescencespectroscopyrdquo Dyes and Pigments vol 83 no 2 pp 249ndash2572009
[25] M-R Ashrafi Kooshk K Mansouri M Nadi S Khodarahmiand R Khodarahmi ldquoIn vitro anti-cancer activity of nativecurcumin and protein-curcuminrdquo Systems vol 2 no 1 pp 80ndash88 1981
[26] P D Ross and S Subramanian ldquoThermodynamics of proteinassociation reactions forces contributing to stabilityrdquo Biochem-istry vol 20 no 11 pp 3096ndash3102 1981
[27] J Liu J-N Tian J Zhang Z Hu and X Chen ldquoInteractionof magnolol with bovine serum albumin a fluorescence-quenching studyrdquo Analytical and Bioanalytical Chemistry vol376 no 6 pp 864ndash867 2003
[28] A Coi A M Bianucci F Bonomi P Rasmussen G M Muraand M L Ganadu ldquoStructural perturbation of 120572B-crystallinby zinc and temperature related to its chaperone-like activityrdquoInternational Journal of Biological Macromolecules vol 42 no3 pp 229ndash234 2008
[29] M Moller and A Denicola ldquoStudy of protein-ligand bindingby fluorescencerdquoBiochemistry andMolecular Biology Educationvol 30 no 5 pp 309ndash312 2002
[30] R Khodarahmi S A Karimi M R Ashrafi Kooshk S AGhadami S Ghobadi and M Amani ldquoComparative spec-troscopic studies on drug binding characteristics and proteinsurface hydrophobicity of native and modified forms of bovineserum albumin possible relevance to change in protein struc-turefunction upon non-enzymatic glycationrdquo SpectrochimicaActa A vol 89 pp 177ndash186 2012
[31] S-F Ma M Anraku Y Iwao et al ldquoHydrolysis of angiotensinII receptor blocker prodrug olmesartan medoxomil by humanserum albumin and identification of its catalytic active sitesrdquoDrug Metabolism and Disposition vol 33 no 12 pp 1911ndash19192005
[32] H M Berman J Westbrook Z Feng et al ldquoThe protein databankrdquo Nucleic Acids Research vol 28 no 1 pp 235ndash242 2000
[33] S Sugio A Kashima SMochizukiMNoda andKKobayashildquoCrystal structure of human serum albumin at 25 A resolutionrdquoProtein Engineering vol 12 no 6 pp 439ndash446 1999
[34] G M Morris D S Goodsell R Huey and A J OlsonldquoDistributed automated docking of flexible ligands to proteinsparallel applications of AutoDock 24rdquo Journal of Computer-Aided Molecular Design vol 10 no 4 pp 293ndash304 1996
[35] F Faridbod M R Ganjali B Larijani et al ldquoInteraction studyof pioglitazone with albumin by fluorescence spectroscopy andmolecular dockingrdquo Spectrochimica Acta A vol 78 no 1 pp96ndash101 2011
[36] D SGoodsell andA JOlson ldquoAutomated docking of substratesto proteins by simulated annealingrdquo Proteins vol 8 no 3 pp195ndash202 1990
[37] D S Goodsell G M Morris and A J Olson ldquoAutomateddocking of flexible ligands applications of AutoDockrdquo Journalof Molecular Recognition vol 9 no 1 pp 1ndash5 1996
[38] G Sudlow D J Birkett and D N Wade ldquoThe characterizationof two specific drug binding sites on human serum albuminrdquoMolecular Pharmacology vol 11 no 6 pp 824ndash832 1975
[39] Y Yue X Chen J Qin and X Yao ldquoCharacterization of themangiferin-human serum albumin complex by spectroscopicandmolecularmodeling approachesrdquo Journal of Pharmaceuticaland Biomedical Analysis vol 49 no 3 pp 753ndash759 2009
[40] A K Shaw and S K Pal ldquoSpectroscopic studies on the effectof temperature on pH-induced folded states of human serumalbuminrdquo Journal of Photochemistry and Photobiology B vol 90no 1 pp 69ndash77 2008
[41] S Ranjbar S Ghobadi R Khodarahmi and H Nemati ldquoSpec-troscopic characterization of furosemide binding to humancarbonic anhydrase IIrdquo International Journal of BiologicalMacromolecules vol 50 no 4 pp 910ndash917 2012
[42] HNHou Z POi YWOYung F L Liao Y Zhang andY LiyldquoStudies on interaction between vitamin B12 and human serumalbuminrdquo Journal of Pharmaceutical and Biomedical Analysisvol 47 pp 134ndash139 2008
[43] S S Kalanur J Seetharamappa and V K A Kalalbandi ldquoChar-acterization of interaction and the effect of carbamazepine onthe structure of human serum albuminrdquo Journal of Pharmaceu-tical and Biomedical Analysis vol 53 no 3 pp 660ndash666 2010
[44] L Stryer ldquoFluorescence spectroscopy of proteinsrdquo Science vol162 no 3853 pp 526ndash533 1968
[45] B Valeur and J C Brochon New Trends in FluorescenceSpectroscopy Springer Berlin Germany 1999
[46] V Suryawanshi P Anbhule A Gore Sh Patil and G AndKolekar ldquoA spectral deciphering the perturbation of modeltransporter protein (HSA) by antibacterial pyrimidine deriva-tive pharmacokinetic and biophysical insightsrdquo Journal ofPhotochemistry and Photobiology B vol 118 pp 1ndash8 2013
[47] T Yuan AMWeljie andH J Vogel ldquoTryptophan fluorescencequenching by methionine and selenomethionine residues ofcalmodulin orientation of peptide and protein bindingrdquo Bio-chemistry vol 37 no 9 pp 3187ndash3195 1998
[48] TCOrsquoHaver A F Fell G Smith et al ldquoDerivative spectroscopyand its applications in analysisrdquo Analytical Proceedings vol 19no 1 pp 22ndash46 1982
[49] Y-Q Wang B-P Tang H-M Zhang Q-H Zhou and G-C Zhang ldquoStudies on the interaction between imidaclopridand human serum albumin spectroscopic approachrdquo Journal ofPhotochemistry and Photobiology B vol 94 no 3 pp 183ndash1902009
[50] S-J Kim H-W Rhee H-Y Park H-S Kim and J-I HongldquoFluorescent probes designed for detecting human serum albu-min on the basis of its pseudo-esterase activityrdquo Bioorganic ampMedicinal Chemistry Letters vol 23 pp 2093ndash2097 2013
[51] P Ascenzi M Gioia G Fanali M Coletta and M FasanoldquoPseudo-enzymatic hydrolysis of 4-nitrophenyl acetate byhuman serum albumin pH-dependence of rates of individualstepsrdquo Biochemical and Biophysical Research Communicationsvol 424 pp 451ndash455 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

12 The Scientific World Journal
between drugs and endogenous ligands for HSA This isparticularly pertinent for fatty acids which normally occurin serum at levels between 01 and 2mol per mol of HSAand can both compete and cooperate with drugs bindingto the protein In certain disease states these effects areexacerbated as the fatty acid HSA molar ratio may be ashigh as six Other pathological conditions are associatedwith high (micromolar to millimolar) levels of bilirubinhemin or renal toxins (eg 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) indoxyl sulphate) which bindto the protein causing significant drug binding defects
Our major aim in this study was to distinguish thenature of the interaction of isoimperatorin to HSA usingdifferent biophysical techniques This study could be veryvaluable due to importance and presence of isoimperatorinin human diet (food-drug interaction on HSA) The resultsshowed that isoimperatorin quenched the intrinsic fluo-rescence of HSA through dynamic quenching mechanismFurthermore hydrophobic interaction played a major role inthe binding process Using Fluorescence and UV absorptionspectroscopy it has been shown that isoimperatorin inducesprotein structural changes The binding average distancebetween the solo tryptophan of the protein and the boundisoimperatorin was calculated by Forster theory as 290 plusmn02 nmAnalysis of PSHvalues elucidated that isoimperatorinhas induced some compactness in the tertiary structure ofthe enzyme The results of synchronous flourescence onHSA-isoimperatorin system elucidated that the drug binds tothe hydrophobic cavity (site I) on subdomain IIA that wasconfirmed by quenching experiments competitive bindingexperiments and docking technique
Acknowledgment
The authors gratefully acknowledge the Kermanshah Uni-versity of Medical Sciences Research Council for financialsupport of this work
References
[1] Y-J Hu Y Liu T-Q Sun A-M Bai J-Q Lu and Z-B Pi ldquoBinding of anti-inflammatory drug cromolyn sodiumto bovine serum albuminrdquo International Journal of BiologicalMacromolecules vol 39 no 4-5 pp 280ndash285 2006
[2] N Seedher and S Bhatia ldquoReversible binding of celecoxiband valdecoxib with human serum albumin using fluorescencespectroscopic techniquerdquo Pharmacological Research vol 54 no2 pp 77ndash84 2006
[3] H Isogai and N Hirayama ldquoIn silico prediction of interactionsbetween site II on human serum albumin and profen drugsrdquoISRN Pharma vol 2013 Article ID 818364 8 pages 2013
[4] YWangHYuX Shi Z LuoD Lin andMHuang ldquoStructuralmechanism of ring opening reaction of glucose by humanserum albuminrdquo The Journal of Biological Chemistry vol 288pp 15980ndash15987 2013
[5] F Keshavarz M M Alavianmehr and R Yousefi ldquoMolecularinteraction of benzalkonium Ibuprofenate and its discreteingredients with human serum albuminrdquo Physical ChemistryResearch vol 1 no 2 pp 111ndash116 2013
[6] T K Maiti K S Ghosh J Debnath and S Dasgupta ldquoBindingof all-trans retinoic acid to human serum albumin fluores-cence FT-IR and circular dichroism studiesrdquo InternationalJournal of Biological Macromolecules vol 38 no 3ndash5 pp 197ndash202 2006
[7] M-X XieM Long Y Liu C Qin and Y-DWang ldquoCharacter-ization of the interaction between human serum albumin andmorinrdquo Biochimica et Biophysica Acta vol 1760 no 8 pp 1184ndash1191 2006
[8] CD Kanakis P A TarantilisMG Polissiou S Diamantoglouand H A Tajmir-Riahi ldquoAntioxidant flavonoids bind humanserum albuminrdquo Journal of Molecular Structure vol 798 no 1-3 pp 69ndash74 2006
[9] S Marumoto and M Miyazawa ldquoStructure-activity rela-tionships for naturally occurring coumarins as 120573-secretaseinhibitorrdquo Bioorganic and Medicinal Chemistry vol 20 no 2pp 784ndash788 2012
[10] S Marumoto and M Miyazawa ldquoBiotransformation of isoim-peratorin and imperatorin by Glomerella cingulata and 120573-secretase inhibitory activityrdquo Bioorganic and Medicinal Chem-istry vol 18 no 1 pp 455ndash459 2010
[11] S E Sajjadi Y Shokoohinia and S Hemmati ldquoIsolation andidentification of furanocoumarins and a phenylpropanoid fromthe acetone extract and identification of volatile constituentsfrom the essential oil of Peucedanum pastinacifoliumrdquo Chem-istry of Natural Compounds vol 48 no 4 pp 668ndash671 2012
[12] X B Yang J Hou D Liu et al ldquoBiotransformation of isoim-peratorin by Cunninghamella blakesleana AS 3970rdquo Journal ofMolecular Catalysis B vol 88 pp 1ndash6 2013
[13] W Lili S Yehong S Qi et al ldquoIn vitro permeability analy-sis pharmacokinetic and brain distribution study in mice ofimperatorin isoimperatorin and cnidilin in Radix AngelicaeDahuricaerdquo Fitoterapia vol 85 pp 144ndash153 2013
[14] S Wang Q Chen and L He ldquoDevelopment and validationof a gas chromatography-mass spectrometry method for thedetermination of isoimperatorin in rat plasma and tissueapplication to the pharmacokinetic and tissue distributionstudyrdquo Journal of Chromatography B vol 852 no 1-2 pp 473ndash478 2007
[15] H Sadraei Y Shokoohinia S E Sajjadi and M MozafarildquoAntispasmodic effects of Prangos ferulacea acetone extract andits maincomponent osthole on ileum contractionrdquo Research inPharmaceutical Sciences vol 8 pp 137ndash144 2013
[16] H Sadraei Y Shokoohinia S E Sajjadi and B GhadirianldquoAntispasmodic effect of osthole and Prangos ferulacea extracton rat uterus smooth muscle motilityrdquo Research in Pharmaceu-tical Sciences vol 7 pp 141ndash149 2012
[17] S Sajjadi Y Shokoohinia S Gholamzadeh M Behbahani andA Fattahi ldquoAntiviral evaluation of coumarins from Prangosferulacea L (Lindl)rdquo Research in Pharmaceutical Sciences vol7 p 783 2012
[18] S E Sajjadi Y Shokoohinia and S Gholamzadeh ldquoChemicalcomposition of the essential oil of the root of Prangos ferulacea(L) Lindlrdquo Chemija vol 22 pp 178ndash180 2011
[19] N Bijari Y Shokoohinia M-R Ashrafi-Kooshk et al ldquoSpec-troscopic study of interaction between osthole and humanserum albumin identification of possible binding site of thecompoundrdquo Journal of Luminescence vol 143 pp 328ndash336 2013
[20] X Shi M Liu M Zhang et al ldquoIdentification of in vitro and invivo metabolites of isoimperatorin using liquid chromatogra-phymass spectrometryrdquo Food Chemistry vol 141 pp 357ndash3652013
The Scientific World Journal 13
[21] T Chatterjee A Pal S Dey B-K Chatterjee and PChakrabarti ldquoInteraction of virstatin with human serum albu-min spectroscopic analysis andmolecular modelingrdquo Plos Onevol 7 pp 1ndash12 2012
[22] Y-M Chen and L-H Guo ldquoCombined fluorescence and elec-trochemical investigation on the binding interaction betweenorganic acid and human serum albuminrdquo Journal of Environ-mental Sciences vol 21 no 3 pp 373ndash379 2009
[23] J R Lakowicz Principles of Fluorescence Spectroscopy KluwerAcademicPlenum Publishers New York NY USA 2nd edi-tion 1999
[24] F Ding N Li B Han F Liu L Zhang and Y Sun ldquoThe bindingof CI Acid Red 2 to human serum albumin determinationof binding mechanism and binding site using fluorescencespectroscopyrdquo Dyes and Pigments vol 83 no 2 pp 249ndash2572009
[25] M-R Ashrafi Kooshk K Mansouri M Nadi S Khodarahmiand R Khodarahmi ldquoIn vitro anti-cancer activity of nativecurcumin and protein-curcuminrdquo Systems vol 2 no 1 pp 80ndash88 1981
[26] P D Ross and S Subramanian ldquoThermodynamics of proteinassociation reactions forces contributing to stabilityrdquo Biochem-istry vol 20 no 11 pp 3096ndash3102 1981
[27] J Liu J-N Tian J Zhang Z Hu and X Chen ldquoInteractionof magnolol with bovine serum albumin a fluorescence-quenching studyrdquo Analytical and Bioanalytical Chemistry vol376 no 6 pp 864ndash867 2003
[28] A Coi A M Bianucci F Bonomi P Rasmussen G M Muraand M L Ganadu ldquoStructural perturbation of 120572B-crystallinby zinc and temperature related to its chaperone-like activityrdquoInternational Journal of Biological Macromolecules vol 42 no3 pp 229ndash234 2008
[29] M Moller and A Denicola ldquoStudy of protein-ligand bindingby fluorescencerdquoBiochemistry andMolecular Biology Educationvol 30 no 5 pp 309ndash312 2002
[30] R Khodarahmi S A Karimi M R Ashrafi Kooshk S AGhadami S Ghobadi and M Amani ldquoComparative spec-troscopic studies on drug binding characteristics and proteinsurface hydrophobicity of native and modified forms of bovineserum albumin possible relevance to change in protein struc-turefunction upon non-enzymatic glycationrdquo SpectrochimicaActa A vol 89 pp 177ndash186 2012
[31] S-F Ma M Anraku Y Iwao et al ldquoHydrolysis of angiotensinII receptor blocker prodrug olmesartan medoxomil by humanserum albumin and identification of its catalytic active sitesrdquoDrug Metabolism and Disposition vol 33 no 12 pp 1911ndash19192005
[32] H M Berman J Westbrook Z Feng et al ldquoThe protein databankrdquo Nucleic Acids Research vol 28 no 1 pp 235ndash242 2000
[33] S Sugio A Kashima SMochizukiMNoda andKKobayashildquoCrystal structure of human serum albumin at 25 A resolutionrdquoProtein Engineering vol 12 no 6 pp 439ndash446 1999
[34] G M Morris D S Goodsell R Huey and A J OlsonldquoDistributed automated docking of flexible ligands to proteinsparallel applications of AutoDock 24rdquo Journal of Computer-Aided Molecular Design vol 10 no 4 pp 293ndash304 1996
[35] F Faridbod M R Ganjali B Larijani et al ldquoInteraction studyof pioglitazone with albumin by fluorescence spectroscopy andmolecular dockingrdquo Spectrochimica Acta A vol 78 no 1 pp96ndash101 2011
[36] D SGoodsell andA JOlson ldquoAutomated docking of substratesto proteins by simulated annealingrdquo Proteins vol 8 no 3 pp195ndash202 1990
[37] D S Goodsell G M Morris and A J Olson ldquoAutomateddocking of flexible ligands applications of AutoDockrdquo Journalof Molecular Recognition vol 9 no 1 pp 1ndash5 1996
[38] G Sudlow D J Birkett and D N Wade ldquoThe characterizationof two specific drug binding sites on human serum albuminrdquoMolecular Pharmacology vol 11 no 6 pp 824ndash832 1975
[39] Y Yue X Chen J Qin and X Yao ldquoCharacterization of themangiferin-human serum albumin complex by spectroscopicandmolecularmodeling approachesrdquo Journal of Pharmaceuticaland Biomedical Analysis vol 49 no 3 pp 753ndash759 2009
[40] A K Shaw and S K Pal ldquoSpectroscopic studies on the effectof temperature on pH-induced folded states of human serumalbuminrdquo Journal of Photochemistry and Photobiology B vol 90no 1 pp 69ndash77 2008
[41] S Ranjbar S Ghobadi R Khodarahmi and H Nemati ldquoSpec-troscopic characterization of furosemide binding to humancarbonic anhydrase IIrdquo International Journal of BiologicalMacromolecules vol 50 no 4 pp 910ndash917 2012
[42] HNHou Z POi YWOYung F L Liao Y Zhang andY LiyldquoStudies on interaction between vitamin B12 and human serumalbuminrdquo Journal of Pharmaceutical and Biomedical Analysisvol 47 pp 134ndash139 2008
[43] S S Kalanur J Seetharamappa and V K A Kalalbandi ldquoChar-acterization of interaction and the effect of carbamazepine onthe structure of human serum albuminrdquo Journal of Pharmaceu-tical and Biomedical Analysis vol 53 no 3 pp 660ndash666 2010
[44] L Stryer ldquoFluorescence spectroscopy of proteinsrdquo Science vol162 no 3853 pp 526ndash533 1968
[45] B Valeur and J C Brochon New Trends in FluorescenceSpectroscopy Springer Berlin Germany 1999
[46] V Suryawanshi P Anbhule A Gore Sh Patil and G AndKolekar ldquoA spectral deciphering the perturbation of modeltransporter protein (HSA) by antibacterial pyrimidine deriva-tive pharmacokinetic and biophysical insightsrdquo Journal ofPhotochemistry and Photobiology B vol 118 pp 1ndash8 2013
[47] T Yuan AMWeljie andH J Vogel ldquoTryptophan fluorescencequenching by methionine and selenomethionine residues ofcalmodulin orientation of peptide and protein bindingrdquo Bio-chemistry vol 37 no 9 pp 3187ndash3195 1998
[48] TCOrsquoHaver A F Fell G Smith et al ldquoDerivative spectroscopyand its applications in analysisrdquo Analytical Proceedings vol 19no 1 pp 22ndash46 1982
[49] Y-Q Wang B-P Tang H-M Zhang Q-H Zhou and G-C Zhang ldquoStudies on the interaction between imidaclopridand human serum albumin spectroscopic approachrdquo Journal ofPhotochemistry and Photobiology B vol 94 no 3 pp 183ndash1902009
[50] S-J Kim H-W Rhee H-Y Park H-S Kim and J-I HongldquoFluorescent probes designed for detecting human serum albu-min on the basis of its pseudo-esterase activityrdquo Bioorganic ampMedicinal Chemistry Letters vol 23 pp 2093ndash2097 2013
[51] P Ascenzi M Gioia G Fanali M Coletta and M FasanoldquoPseudo-enzymatic hydrolysis of 4-nitrophenyl acetate byhuman serum albumin pH-dependence of rates of individualstepsrdquo Biochemical and Biophysical Research Communicationsvol 424 pp 451ndash455 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

The Scientific World Journal 13
[21] T Chatterjee A Pal S Dey B-K Chatterjee and PChakrabarti ldquoInteraction of virstatin with human serum albu-min spectroscopic analysis andmolecular modelingrdquo Plos Onevol 7 pp 1ndash12 2012
[22] Y-M Chen and L-H Guo ldquoCombined fluorescence and elec-trochemical investigation on the binding interaction betweenorganic acid and human serum albuminrdquo Journal of Environ-mental Sciences vol 21 no 3 pp 373ndash379 2009
[23] J R Lakowicz Principles of Fluorescence Spectroscopy KluwerAcademicPlenum Publishers New York NY USA 2nd edi-tion 1999
[24] F Ding N Li B Han F Liu L Zhang and Y Sun ldquoThe bindingof CI Acid Red 2 to human serum albumin determinationof binding mechanism and binding site using fluorescencespectroscopyrdquo Dyes and Pigments vol 83 no 2 pp 249ndash2572009
[25] M-R Ashrafi Kooshk K Mansouri M Nadi S Khodarahmiand R Khodarahmi ldquoIn vitro anti-cancer activity of nativecurcumin and protein-curcuminrdquo Systems vol 2 no 1 pp 80ndash88 1981
[26] P D Ross and S Subramanian ldquoThermodynamics of proteinassociation reactions forces contributing to stabilityrdquo Biochem-istry vol 20 no 11 pp 3096ndash3102 1981
[27] J Liu J-N Tian J Zhang Z Hu and X Chen ldquoInteractionof magnolol with bovine serum albumin a fluorescence-quenching studyrdquo Analytical and Bioanalytical Chemistry vol376 no 6 pp 864ndash867 2003
[28] A Coi A M Bianucci F Bonomi P Rasmussen G M Muraand M L Ganadu ldquoStructural perturbation of 120572B-crystallinby zinc and temperature related to its chaperone-like activityrdquoInternational Journal of Biological Macromolecules vol 42 no3 pp 229ndash234 2008
[29] M Moller and A Denicola ldquoStudy of protein-ligand bindingby fluorescencerdquoBiochemistry andMolecular Biology Educationvol 30 no 5 pp 309ndash312 2002
[30] R Khodarahmi S A Karimi M R Ashrafi Kooshk S AGhadami S Ghobadi and M Amani ldquoComparative spec-troscopic studies on drug binding characteristics and proteinsurface hydrophobicity of native and modified forms of bovineserum albumin possible relevance to change in protein struc-turefunction upon non-enzymatic glycationrdquo SpectrochimicaActa A vol 89 pp 177ndash186 2012
[31] S-F Ma M Anraku Y Iwao et al ldquoHydrolysis of angiotensinII receptor blocker prodrug olmesartan medoxomil by humanserum albumin and identification of its catalytic active sitesrdquoDrug Metabolism and Disposition vol 33 no 12 pp 1911ndash19192005
[32] H M Berman J Westbrook Z Feng et al ldquoThe protein databankrdquo Nucleic Acids Research vol 28 no 1 pp 235ndash242 2000
[33] S Sugio A Kashima SMochizukiMNoda andKKobayashildquoCrystal structure of human serum albumin at 25 A resolutionrdquoProtein Engineering vol 12 no 6 pp 439ndash446 1999
[34] G M Morris D S Goodsell R Huey and A J OlsonldquoDistributed automated docking of flexible ligands to proteinsparallel applications of AutoDock 24rdquo Journal of Computer-Aided Molecular Design vol 10 no 4 pp 293ndash304 1996
[35] F Faridbod M R Ganjali B Larijani et al ldquoInteraction studyof pioglitazone with albumin by fluorescence spectroscopy andmolecular dockingrdquo Spectrochimica Acta A vol 78 no 1 pp96ndash101 2011
[36] D SGoodsell andA JOlson ldquoAutomated docking of substratesto proteins by simulated annealingrdquo Proteins vol 8 no 3 pp195ndash202 1990
[37] D S Goodsell G M Morris and A J Olson ldquoAutomateddocking of flexible ligands applications of AutoDockrdquo Journalof Molecular Recognition vol 9 no 1 pp 1ndash5 1996
[38] G Sudlow D J Birkett and D N Wade ldquoThe characterizationof two specific drug binding sites on human serum albuminrdquoMolecular Pharmacology vol 11 no 6 pp 824ndash832 1975
[39] Y Yue X Chen J Qin and X Yao ldquoCharacterization of themangiferin-human serum albumin complex by spectroscopicandmolecularmodeling approachesrdquo Journal of Pharmaceuticaland Biomedical Analysis vol 49 no 3 pp 753ndash759 2009
[40] A K Shaw and S K Pal ldquoSpectroscopic studies on the effectof temperature on pH-induced folded states of human serumalbuminrdquo Journal of Photochemistry and Photobiology B vol 90no 1 pp 69ndash77 2008
[41] S Ranjbar S Ghobadi R Khodarahmi and H Nemati ldquoSpec-troscopic characterization of furosemide binding to humancarbonic anhydrase IIrdquo International Journal of BiologicalMacromolecules vol 50 no 4 pp 910ndash917 2012
[42] HNHou Z POi YWOYung F L Liao Y Zhang andY LiyldquoStudies on interaction between vitamin B12 and human serumalbuminrdquo Journal of Pharmaceutical and Biomedical Analysisvol 47 pp 134ndash139 2008
[43] S S Kalanur J Seetharamappa and V K A Kalalbandi ldquoChar-acterization of interaction and the effect of carbamazepine onthe structure of human serum albuminrdquo Journal of Pharmaceu-tical and Biomedical Analysis vol 53 no 3 pp 660ndash666 2010
[44] L Stryer ldquoFluorescence spectroscopy of proteinsrdquo Science vol162 no 3853 pp 526ndash533 1968
[45] B Valeur and J C Brochon New Trends in FluorescenceSpectroscopy Springer Berlin Germany 1999
[46] V Suryawanshi P Anbhule A Gore Sh Patil and G AndKolekar ldquoA spectral deciphering the perturbation of modeltransporter protein (HSA) by antibacterial pyrimidine deriva-tive pharmacokinetic and biophysical insightsrdquo Journal ofPhotochemistry and Photobiology B vol 118 pp 1ndash8 2013
[47] T Yuan AMWeljie andH J Vogel ldquoTryptophan fluorescencequenching by methionine and selenomethionine residues ofcalmodulin orientation of peptide and protein bindingrdquo Bio-chemistry vol 37 no 9 pp 3187ndash3195 1998
[48] TCOrsquoHaver A F Fell G Smith et al ldquoDerivative spectroscopyand its applications in analysisrdquo Analytical Proceedings vol 19no 1 pp 22ndash46 1982
[49] Y-Q Wang B-P Tang H-M Zhang Q-H Zhou and G-C Zhang ldquoStudies on the interaction between imidaclopridand human serum albumin spectroscopic approachrdquo Journal ofPhotochemistry and Photobiology B vol 94 no 3 pp 183ndash1902009
[50] S-J Kim H-W Rhee H-Y Park H-S Kim and J-I HongldquoFluorescent probes designed for detecting human serum albu-min on the basis of its pseudo-esterase activityrdquo Bioorganic ampMedicinal Chemistry Letters vol 23 pp 2093ndash2097 2013
[51] P Ascenzi M Gioia G Fanali M Coletta and M FasanoldquoPseudo-enzymatic hydrolysis of 4-nitrophenyl acetate byhuman serum albumin pH-dependence of rates of individualstepsrdquo Biochemical and Biophysical Research Communicationsvol 424 pp 451ndash455 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology





























