Embed Size (px)
Citation preview

Hindawi Publishing CorporationISRN ToxicologyVolume 2013 Article ID 184360 9 pageshttpdxdoiorg1011552013184360
Research ArticleRole of Calcium Channels in Heavy Metal Toxicity
Carla Marchetti
Istituto di Biofisica Consiglio Nazionale delle Ricerche 16149 Genova Italy
Correspondence should be addressed to Carla Marchetti marchettigeibfcnrit
Received 27 November 2012 Accepted 23 December 2012
Academic Editors R W Coppock R Konig and R Mateo
Copyright copy 2013 Carla Marchetti This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
The role of voltage-dependent Ca channels (VDCC) in themembrane permeation of two toxicmetals lead (Pb) and cadmium (Cd)was studied in mammalian cells Both metals interact with Ca-binding sites but while Cd influx appears to occur mainly throughthe same pathways as Ca Pb is also rapidly taken up by different passive transport systems Furthermore I compared the effectof Cd in two Chinese hamster ovary (CHO) cell lines a wild-type and a modified cell line which were permanently transfectedwith an L-type VDCC When cultures were subjected to a brief (30ndash60min) exposure to 50ndash100120583MCd apoptotic features metalaccumulation and death were comparable in both cell lines although in transfected cells the effect of Cd treatment was partiallyprevented by nimodipine (VDCC antagonist) and enhanced by BayK8644 (VDCC agonist)Thus expression of L-type Ca channelsis not sufficient to modify Cd accumulation and sensitivity to a toxicological significant extent and while both Cd and Pb can takeadvantage of VDCC to permeate the membrane these transport proteins are not the only and frequently not the most importantpathways of permeation
1 Introduction
Metals are used by biological systems because of their cat-alytic versatility and some of them namely sodium (Na)potassium (K) calcium (Ca) and magnesium (Mg) areamong the essential nutrients of living cells In contrast othermetals can be fatal to mammalian cells even in tiny amountsA common classification tends to sort these potentially toxicmetals in to two classes those required by living organisms asessential micronutrients and those devoid of any biologicalfunction and thus potentially toxic even at very low con-centration such as cadmium (Cd) chromium (Cr) mercury(Hg) and lead (Pb) The difference between toxic andnontoxic metals is hard to define Even micronutrients suchas cobalt (Co) copper (Cu) iron (Fe) manganese (Mn)molybdenum (Mo) and zinc (Zn) can be detrimental to liv-ing organisms when present in excessive levels and a refinedequilibrium between deficient and toxic concentrations hasto be maintained This is particularly important for veryspecialized tissues such as the brain where metals induceoxidative damage and some of the essential micronutrientssuch as Fe Zn and Cu have been implicated in etiology anddevelopment of different neurological and neurodegenerativediseases [1 2] Less obviously living organism may find
use for nonessential toxic metals in extreme condition Anelegant example of unexpected biological function of Cd hasbeen recently reported in marine diatoms [3]
Because most toxic metals have entered the environmentas a consequence of historical handcraft and industrial pro-cessing specialized cellsmight not have developed the intrin-sic ability to distinguish between physiological and toxic ele-ments For example Cd and Pb represent a threat for mam-malian cells because they are able to replace ormimic essen-tial metals in the first or early step of transport and metab-olism but then are incapable of mediating subsequent vitalfunctions [4 5] The toxic effects are then a consequence ofboth impairment of transport systems and accumulation intothe cell Recent research has gathered an increasing body ofevidence for the interaction of toxic metals with intracellularcomponents that lead to cellular injury and defense butthe mechanisms of transport of these metals and metal-containing species across plasma membranes remain to befully characterized
Cd and Pb mimic Ca and Zn at their specific sites andbind calmodulin [6 7] protein kinase C [8ndash10] and synapticproteins [11] These metal ions antagonize Ca influx throughvoltage-dependent and receptor-operated Ca channels and
2 ISRN Toxicology
these channels are regarded as the major route of their entryinto mammalian cells
Both Cd and Pb are well-known specific blockers ofvoltage-dependent Ca channels (VDCC) but the mechanismof block is quite different While Cd binds to the high affinitysite inside the VDCC pore [12 13] Pb competes with Ca at asite external to the VDCC and the binding is not voltage-dependent [14] Given these differences in the mechanismof interaction permeation of these metals through VDCCis also different VDCCs mediate Cd influx in excitable cells[15ndash18] including mammalian neurons [19] and have beenproposed to participate in Cd uptake also in cells fromnonexcitable tissues [20] In a previouswork ofmy laboratorywe showed that in certain cells Cd permeation occurs mainlythrough VDCC of the L-type [19 21] However in otherstudies the presence of VDCC per se did not seem to enhancesensitivity to Cd for example VDCCs expressing (PC12) andnonexpressing (PC18) cells showed comparable LD
50for Cd
[18] and the role of this pathway in the induction of cell deathappears questionable
The situation is even more complicated for Pb whosechemical basis formimickingCa is not obvious [4] andwhichis known to adapt to structurally diverse binding geometries[22] In cerebellar granule neurons Pb uses at least threepathways of permeation and besides voltage-dependent cal-cium channels (VDCC) and NMDA-activated channels israpidly taken up through passive transport system [23 24]
In this paper I will present experimental evidence con-cerning transport of Pb and Cd across cellular membranesand the relative contribution of VDCC in their uptake
2 Materials and Methods
21 Cell Culture Cerebellar granule neurons were preparedfrom 8-day-old Wistar rats as previously described [19 25]plated on 20mm poly-L-lysine coated glass coverslips andmaintained in basal Eaglersquos culture medium supplementedwith 10 fetal calf serum 100 120583gmL gentamicin and 25mMKCl in a humidified 95 airCO
2atmosphere at 37∘C
Cultures were treated with 10 120583M cytosine arabinoside fromday 1 to minimize the proliferation of nonneuronal cellsExperiments were carried out in cultures between 5 and 13days in vitro
Wild-type Chinese hamster ovary (CHO) cells were ob-tained from American Type Culture Collection CHO stablytransfected with cDNA encoding for several subunits ofvoltage-dependent calcium channels (VDCC) were a gener-ous gift of Franz Hofmann and Norbert Klugbauer (Institutfur Pharmakologie und Toxikologie Munich Germany)The cells used in this study are termed CHOC1205729120573
312057221205754
(abbreviated CHOC120572) to indicate that they express the 1205721119862minus119887
subunit of VDCC as well as 1205733and 120572
21205754 subunits from
smooth muscle [26]Both wild-type CHO and CHOC120572 cells were maintained
in DMEM medium (Sigma Chemical Co St Louis MOUSA) supplemented with 10 bovine serum in a 5 CO
2
humidified atmosphere at 37∘C For CHOC120572 cells the cul-ture medium was routinely supplemented with antibiotic
geneticin G418 20120583grmL For the experiments cells wereplated in 12-multiwell trays at a density of 2 times 105 cellsmL
22Microscopy Measurement of intracellular Cd and Pbwasperformed on monolayer cultures plated on glass coverslipsFor photometric measurements cells were incubated with6 120583M of the cell-permeant Fura2-AM ester form of the dyefor 45min at 37∘C and then washed several times withstandard saline at room temperature as previously described[19 23] Cd and Pb were applied in the absence of Caand treatment was terminated by the addition of themembrane-permeant metal chelator (NNN1015840N1015840-tetrakis-2-pyridyl methyl ethylenediamine TPEN 100 120583m)
For confocal microscopy cells were incubated with Ore-gonGreen 488Bapta-1 a fluorescent indicator whose fluores-cence intensity is increased upon binding Ca Cd or Pb Cellswere imaged using a confocal laser scanning microscopeNikon PCM2000 (Nikon Instr Florence Italy) with an oilimmersion 100 times objective (Na = 13) coupled to a 50120583mconfocal pinhole condition [27]
23 Cd Treatment Cells were subjected to 30ndash60min treat-ments with 50 or 100120583M CdCl
2in culture medium without
serum When specified the medium was supplemented with30mM KCl to depolarize the cell membrane and with thefollowing dihydropyridines (DHPs) BayK 8644 which is anL-type VDCC agonist or nimodipine a channel antagonistStock solutions of nimodipine and BayK (10mM) weremade up in 100 ethanol and diluted in medium to thefinal concentration High KCl alone as well as DHPs (noCd added) had no effect on cell viability and appearanceAfter incubation cells were washed three times with PBScontaining 1mMEDTA and freshmedium containing serumwas replaced Cultures were kept in the incubator for another18ndash24 hours before testing Cell viability in each samplewas assessed by trypan-blue exclusion assay All chemicalsculture media and sera were from Sigma-Aldrich Italy
24 Capacitance and Current Measurements by Patch-ClampTechnique Membrane currents were measured from wild-type CHO and permanently transfected CHOC120572 cells inwhole-cell clamp configuration by a patch-clamp Axopatchamplifier (Molecular Devices Corporation Union City CAUSA) Electrodes were manufactured from borosilicate glasscapillaries (Hilgenberg GmbH Malsfeld Germany) and hadresistance of asymp 4MΩ Voltage stimulation and data acqui-sition were performed by a PC through a Digidata 1440Ainterface and Pclamp-10 software (Molecular Devices) Cur-rents were low-pass filtered at 5 kHz and digitized at 10 kHzCapacitance transients wereminimized by analog compensa-tion and the value obtained by this compensation was takenas an estimate of the cell capacitance All currents traceswere further corrected for leak and residual transients by acomputer generated P4 protocol The holding potential wasminus80mV in all the experiments Current traces were analyzedwith Clampfit-10 and Sigma Plot (Jandel Scientific ErkrathGermany) software
Cells were continuously superfused by gravity flow(10mLmin) with a solution containing (in mM) NaCl 140
ISRN Toxicology 3
KCl 54 CaCl218 Hepes 5 The pH was 74 The internal
(pipette) solution contained (in mM) CsCl 20 CsOH 110Aspartic acid 100 EGTA 5 Hepes 5 with pH adjustedto 73 with Trizma base Calcium currents were recordedin similar external solution containing 130mM NaCl and5mMCaCl
2 Wild-type CHO cells do not possess prominent
voltage-dependent currents and CHOC120572 only contained theL-type VDCC under study therefore there was no needto antagonize the current through other voltage-dependentchannels to resolve Ca currentsThe application of modifierssuch as agonist and antagonist DHPs was accomplished bygravity flow control ion substitution experiments showedthat the external bathwas completely changed in 10 sec whichwas the maximal stimulation rate in these experiments
25 Absorption Spectroscopy Cd Determination Cells wereexposed to Cd in culture medium without added serumAfter treatment cultures were washed three times with PBScontaining 1mM EDTA and harvested immediately Viablecells were counted in triplicate in each sample Cells were thenwashed again three times with the above buffer resuspendedin distilled water and disrupted by sonication (SonopulsUltrasonic Homogenizer Bandelin) for 2 minutes in an icebath The total Cd content of each sample was measured byFlameless Atomic Absorption Spectroscopy (FAAS) using aPerkin Elmer Spectrophotometer (Model 1100 B) equippedwith a graphite furnace (Model HGA 700) The Cd contentwas normalized to the volume occupied by the cells andmolarity was calculated (see results)
26 Statistical Analysis Data are presented as mean plusmn stan-dard error of the mean in at least 3 experiments Statisticalsignificance was evaluated by Student-Newman-Keuls multi-ple comparison test (In Stat GraphPad Software)
3 Results And Discussion
31 Influx of Toxic Metals into Mammalian Cells Influxof toxic divalent metal ions into mammalian cells can bemonitored in cells preloaded with specific fluorescent probesFura2 [17 19 23] or Oregon green These fluorescent indica-tors were developed and are predominantly used for deter-mining intracellular calcium levels but they have relativelyhigh affinity also for other polyvalent ions including Cd andPb
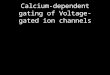
Figure 1 shows how exposure to a solution containing atoxicologically significant concentration of Cd or Pb causesa sizeable increase in Oregon Green fluorescence similar tothat seen following Ca influx [28] This demonstrates fea-sibility of this probe as toxic metal influx indicator In thisexperiment while Cd uptakewas triggered by depolarizationindicating the essential contribution of VDCC (Figure 1(A)(a)-(b)) neurons rapidly accumulate Pb even in the absenceof a specific stimulus (Figure 1(A) (c)-(d))
Figure 1(B) shows real-time measurements of Cd andPb influx in Fura2-loaded neurons [19 23] The influx wasquantified by measuring the fluorescence emission ratio R(E380
E340
) and membranes were depolarized by increasing
concentration of external KCl [25] In the left panel thefluorescent ratio E
340E380
increased negligibly when 50120583MCd was applied in resting condition but the rate of risedRdt was enhanced by gt60 times following depolarizationwith 25KCl In contrast Pb (15ndash50120583M)determined a sizeableincrease of R even in the absence of any specific stimulus thatopens VDCC and dRdt was increased by 5 times from theresting level with 75mMKCl (Figure 1(C)) Fast uptake of Pbin the absence of any specific stimulus appears to be a peculiarfeature of neurons and may be linked to the specific lipidiccomposition of the neuronal membranes while the role ofVDCC is likely to significant but not essential [23]
32 Role of VDCC in Cd-Induced Mortality As VDCC chan-nels and in particular those of L-type [15 19] play a promi-nent role in uptake of Cd by excitable cells the involvement ofthese channels in Cd toxicity was investigated further Tests ofmortality chromatin condensation and DNA fragmentationshowed that a pulsed Cd treatment induces delayed apop-totic cell death in VDCC-containing insulinoma cells andthat nimodipine protects against Cd-induced apoptosis andnecrosis in these cells supporting a specific involvement of L-type VDCC [21] The same treatments were largely harmlessin VDCC-free HeLa cell cultures in which neither death norDNA fragmentation was observed Therefore excitable cellsappear more susceptible than nonexcitable epithelial-likecells to Cd accumulation However it is not clear whether themere presence of VDCC can make cells more vulnerable totoxicmetal injury To answer this question we have comparedthe effects of Cd in two cell types that are virtually identicalexcept for the expression of VDCC in one of the two Thisapproach is made possible by the availability of a CHO cellline that has been permanently transfected with VDCC 120572
1
120573 and 1205722120575 subunits CHOC1205729120573
312057221205754cells [26 29] Promi-
nent voltage-dependent Ca currents were recorded fromthe permanently transfected cell line while such currentsare completely absent in wild-type CHO (Figure 2(a)) Thecurrents are larger with Ba as charge carrier and are sensitiveto DHP modulation (Figure 2(b)) as expected for an L-typeCa channel
Both cell types (wild-type and CHOC120572) were subjectedto a ldquopulse treatmentrdquo with 50 or 100 120583M Cd in serum-freeculture medium and observed after 18ndash24 hour after washCell suffering and mortality was negligible immediately afterthe treatment but it was evident after at least 16 hours Aspreviously observed [21 30 31] Cd accumulates into the cellduring treatment but the toxic mechanism leading to celldeath becomes effective at a later time Figure 3 shows theappearance of the cell monolayer 24 hours after an incubationof 30min with 100120583M Cd Cells exposed to Cd were ofsmaller size than control cells and had an increased tendencyto detach from their neighbors and from the substrate andfloat In addition CHOC120572VDCC-expressing cells frequentlyacquired an irregular shape with shredded edges (Figure 3)The number of viable cells was estimated from the numberof adherent trypan-blue excluding cells and this numberwas normalized to that of adherent viable cells in controlconditions as in our previous works [21] Despite some mor-phological features the dose dependence of Cd sensitivity
4 ISRN Toxicology
08
06
04
02
0
TPEN
TPEN
TPEN
2 min
25 KCl25 KCl
50 Cd 50 Pb15 Pb
75 KCl
Fluo
resc
ence
ratio
Stnd 0 Ca
lowast
lowast
lowast
100
50
0
(A) (B)
(C)
(a) (b)
(c) (d)
25 KCl75 KCl
5 15 50
119889119877119889119905(
minminus1)lowast
103
Figure 1 Uptake of Cd and Pb in cerebellar granule neurons measured by fluorescent dyes (A) Confocal microscopy images of Cd (a b) andPb (c d) uptake by cerebellar granule cells preloaded with the divalent metal-sensitive dye Oregon Green In (a) and (c) neurons were bathedin a physiological saline In (b) they have been superfused with a solution containing 0 Ca 30mM KCl and 100 120583M Cd Cl
2 which caused
the dye fluorescence to increase significantly In (d) external solution contained 0Ca and Pb 15120583M which permeates through the neuronmembrane even in the absence of a depolarizing stimulus and also caused an increase in the dye fluorescence Bar = 10 120583M (B) Real-timerecording of the influx of Cd and Pb in Fura-2 loaded cerebellar granule neuron and the effect of membrane depolarization Cells were treatedwith the metals in nominal Ca-free solution From left to right time course of the fluorescence ratio 119877 = E340E380 following application of50 120583MCd and membrane depolarization (25mM KCl) and application of 15ndash50120583M Pb in resting conditions and increasing depolarizations(25 and 75mM KCl) (C) Summary of the results obtained in basal and in depolarizing solutions (25 and 75mM KCl) with different doses ofPb The time course of R(t) was approximated by a straight line and the slope dRdt was calculated in each case as a measure of Pb influxData are mean plusmn SEM in 4 experiments and lowastindicates significantly different from basal (no depolarization) for each dose of Pb (119875 lt 005)
was very similar in the two cell types (Figure 3) Thereforeby this approach it is not possible to demonstrate a clear roleof VDCC in mediating Cd-induced cell detachment
In subsequent experiments we challenged the cells withCd in the presence of 1120583M DHP modifiers When VDCC-expressing CHOC120572 cells were exposed to Cd in the presence
ISRN Toxicology 5
(a)
Wild type
100 pA
10 ms
(b)
5 mM Ca
5 mM Ba
50 pA
10 ms
Nimo 5 Ba 5 Ca
0 60 120 180 240 300
Curr
ent (
pA)
0
minus100
minus200
(c)
Figure 2 CHOC120572 cells express L-type VDCC (a) Representative microphotographs of monolayer cell cultures of permanently transfectedCHOC120572 cells (left) and wild-type CHO cells (right) Bar = 35 120583M (b) Representative current traces evoked by depolarizing voltage stepsof 40ms duration from minus60 to +80mV from a holding potential of minus80mV in a CHOC120572 cell (left) and a wild-type CHO cell (right) Theexternal solution contained 5mMCaCl
2 This protocol evoked voltage-dependent calcium current in the permanently transfected cell while
no current was present in CHO cell of the wild-type (c) Characterization of the Ca current in CHOC120572 cellThe current was increased bymorethan 50 when the external solution was changed from 5mM CaCl
2to 5mM BaCl
2 and it was reversibly blocked by 10 120583M nimodipine
Current traces evoked by 50ms depolarizing steps from minus80mV (holding potential) to +10mV in the three conditions are shown on the leftThe graph on the right shows the time course of the experiment
of the antagonist (nimodipina) or agonist (BayK 8644) theeffect of the metal was clearly modified with respect to cellsexposed to Cd alone (Figure 4) with a sizeable increase ofmortality in the presence of BayK and protection in the pres-ence of nimodipineDHPmodifierswere largely ineffective inwild-type CHO cells exposed to Cd (not shown) Thereforeit seems that the actual contribution of L-type VDCC to Cduptake is clearly underscored only in the presence of thesemodifiers
Another indicator of apoptosis is cell shrinkage whichcan be quantified by cell capacitance measurements Bothwild-type and CHOC120572 cells showed a decrease in cell capaci-tance 18ndash24 hours afterCd treatment Inwild-type cell capac-itance was 24plusmn2 pF (119899 = 15) in control and 15plusmn 3 pF (119899 = 6)after incubation with 100 120583MCd significantly different fromcontrol with 119875 lt 005 In CHOC120572 cells cell capacitance was35 plusmn 3 pF (119899 = 17) in control and 22 plusmn 2 pF (119899 = 39)after incubation with 100 120583MCd significantly different from
control with 119875 lt 0001 In CHOC120572 cells the effect ofCd treatment was partially prevented by nimodipine thecapacitance was 35plusmn3 pF (119899 = 10) and the difference betweenthis condition and control condition was not significant(Figure 5)This indicates again that the contribution of L-typeVDCC in Cd-induced toxicity is evident only in the presenceof DHPs
Finally we performedmeasurements of total Cd accumu-lation during 60min incubation time by atomic absorptionspectroscopy For these experiments cells were harvestedimmediately after the treatment and the total amount of Cdwas normalized to the cell volume estimated from the single-cell capacitance Assuming that both cell types are approxi-mately spherical and have a specific membrane capacitanceof 1 120583Fcm2 106 wild-type CHO cells comprise a volume of11 120583L while 106 CHOC120572 cells a volume of 195120583L Thisdifference is not negligible and with this method the resultswere considerably more reproducible than using a standard
6 ISRN Toxicology
1
08
06
04
02
01 10 100 1000
Wild type+VDCC
(a) (b)
(c) (d)
(e)
Figure 3 Effect of pulse treatment with Cd in CHOC120572 VDCC-expressing and wild-type CHO cells Microphotographs of control (a b) andCd-treated (c d) cells (a c) VDCC-expressing CHOC120572 cells and (b d) wild-type CHO cells Both cell types were incubated in 100120583M Cdfor 60 minutes in the presence of 30mM KCl Pictures were taken 24 h after wash of the metal Bar = 35 120583MThe graph (e) shows the effectof a 30min pulse treatment with Cd on cell adhesion as a function of concentration Trypan-blue excluding adherent cells were counted24 h after wash of the metal Points are average plusmn sem of 3 experiments in the same condition in CHOC120572 (filled circles) and wild-type CHO(empty circles) and were best fitted to the function NadherNadher(control) = 1(1 + ([Cd]ED50)) where NadherNadher(control) is the number ofadherent cells after Cd treatment normalized to the number of adherent cells in control culture [Cd] is the concentration of Cd and ED
50is
the concentration of Cd that causes detachment from the substrate of 50 of cells The best fit yielded ED50= 40 120583M for CHOC120572 and 43120583M
for wild-type CHOThe two curves are overlapped In contrast with the different appearance the two cell types were similarly affected by Cdtreatment
normalization to the total protein content Data shown inFigure 6 indicate that the difference in Cd accumulation isbarely or not significant when both cell types are incubatedin 100120583M Cd with or without elevated KCl On the otherhand BayK 8644 significantly enhanced and nimodipinesignificantly decreased metal accumulation in depolarizedCHOC120572 cells treated with 50 or 100120583M Cd but not in wild-type CHO cells
Toxicity of both Cd and Pb is mainly due to their abilityto permeate not simply mammalian cell membranes butspecifically tight membrane barriers such as the epitheliallining in the gastrointestinal tract luminal membrane ofproximal tubules in the kidney and the blood-brain barrierIn early work the influx of Pb and Cd into mammaliancells was frequently ascribed to permeation through calciumchannels and in particular L-type DHP-sensitive VDCC [1618 20 32]More recently other types of VDCC different fromthe classical DHP-sensitive L-type have been implicated bothin Cd [19 33] and Pb [23] uptake Moreover Pb has beenshown to use different pathways in several systems [34ndash37]and it is largely independent of VDCC even in neurons[23 24]
Here I have shown that expression of L-type VDCC is notsufficient to modify Cd uptake to a toxicological significantextent at least in the case of a brief intense exposure asthose used in this study Cd accumulation and sensitivitywerecomparable in two cell lines whose only difference was thepresence of L-typeVDCC in one of the two Indeedwild-type
CHO cells which do not possess VDCC are vulnerable to Cdpoisoning and take up Cd during relatively brief exposuresto an extent sufficient to trigger cell death by apoptosisor necrosis similarly to VDCC expressing CHOC120572 cellsAlthough VDCCs play an important role in Cd uptakein CHOC120572 cells as indicated by the sizeable effect of L-type channel DHP modifiers wild-type CHO cells use othertransport proteins with similar efficiency and outcome
It is thus clear that VDCCs are not the main mecha-nism responsible for toxic metal permeation through cellmembranes and tight barriers Rapid passive transport of Pbindependent of VDCC was reported at the brain endothe-lium [34 36 37] and even protein-independent transportof lead has been described [35] In astroglia cell models theuptake of Pb was shown to be driven by store-depletion-activated channels [38] and by two distinct pH-sensitivetransport mechanisms [39] VDCC-independent uptake ofPb by cerebellar granule neurons was inhibited by La [24]and this observation can provide some clue as for the mecha-nism of permeationThe involvement of a La-sensitive store-depletion activated channel as proposed in other systems[36 37 40] is unlikely because Pb influx did not requiredrainage of the stores Another possibility is that Pb2+ entersthe neuron via reverse operation of an exchanger similar (oridentical) to theNandashCa exchanger which is also very sensitiveto La block [41] and whose role in Zn permeation wasdemonstrated in cortical neurons [42] Also this possibilitywas discarded because NandashCa exchanger does not play
ISRN Toxicology 7
4
3
2
1
0Basal Cd K N B
Treatments
lowast
lowastlowast
(a) (b)
(c) (d)
(e)
Figure 4 Effect of dihydropyridines on Cd cytotoxicity in VDCC-expressing CHOC120572 cells Cells were treated for 30min in (a) controlmediumcontaining 30mMKCl (b) 100 120583M Cd+30mM KCl (c) 100 120583M Cd+30mM KCl+1 120583Mnimodipine (d) 100 120583M Cd+30mM KCl+1 120583M BayK8644 Note the partial recovery in shape of the cells treated in the presence of nimodipine and definitive lost of adhesion whenthe treatment was performed in the presence of BayK Bar = 25 120583M The graph (e) represents counts of viable adherent cells in the sameexperiment (average plusmn sem in 3 samples) in control (basal) following for a 30min treatment with 100 120583M Cd (Cd) Cd + 30mM KCl (K)Cd + 30mM KCl and 1 120583M nimodipine (N) and Cd + 30mM KCl and 1 120583M BayK (B) lowast indicates significantly different from control with119875 lt 005 and lowastlowast with 119875 lt 00001
40
30
20
10
0Wild type
ControlCd
Capa
cita
nce (
pf)
lowast
lowastlowast
Cd + nimo
+VDCC
Figure 5 Evidence of Cd-induced apoptosis change in cell capac-itance Cells were incubated with 100120583M Cd for 60 minutes andelectrical measurements were performed 24 h after wash Mem-brane capacitance was estimated from transient compensation (seeSection 24) Bars represent mean plusmn sem Treatment with Cd causedshrinkage of all cell as revealed by reduction of the cell capacitancein both wild-type (119899 = 15 in control and 119899 = 6 with Cd treatment)and CHOC120572 cells (119899 = 17 in control and 119899 = 39with Cd treatment)Nimodipine protected CHOC120572 cells from Cd-induced shrinkage(119899 = 10) lowast indicates significantly different from control with 119875 lt005 and lowastlowast with 119875 lt 0001
a significant role when the Na concentrations are close tothe physiological value as in Esposito et al [24] It is more
probable that Pb is taken up through a La-sensitive ion carriersimilar to the Zn transporter found in brain neurons [43]whose identity and features still wait full definition
Involvement of different transport proteins in Cd trans-port has been described At the epithelial lining of the gas-trointestinal barrier DMT1 the main transporter thatabsorbs Fe in the brush border membrane of the mammalianintestine has been shown to carry also different toxic metals[44ndash46] The role of this transporter in Cd absorption isunderscored by the fact that iron deficiency creates a sig-nificant risk for increased cadmium exposure by increasinggastrointestinal absorption from 5 to as much as 20In general a low level of essential metals favors uptake ofnonessential (toxic) elements that compete for their site andthis stresses the importance of evaluating metal balance incontrast with metal concentration In addition other metal-specific transporters have been identified at the cDNA level[47] and studied in expression systems [48]
4 Conclusion
Ionicmimicry is a useful framework to study themechanismsof metal toxicity Uptake of toxic metals such as Cd and Pbby mammalian cells of different tissues occurs through manydifferent pathways of which VDCCs have frequently beenregarded as prominent Because of the abundance of VDCCexcitable cells may be more susceptible to accumulate Cdand Pb than nonexcitable cells In this chapter I have shownthat although Cd and Pb can permeate through VDCC thisis not the most relevant mechanism of passive uptake ofPb while the mere presence of VDCC is not sufficient tomake cells significantly more vulnerable to Cd injury Otherfactors including different transport systems and expressionof detoxification proteins may be more relevant in the accu-mulation process of these toxic metals
8 ISRN Toxicology
Basal Cd K B NTreatments
Wild type
80
60
40
20
0
lowast
lowast
Figure 6 Total Cd accumulation measured by FAAS in CHOC120572and wild-type CHO cells Both cell types were incubated for 1 hourin medium containing 0 50 or 100 120583M Cd and other modifiersCd determination was normalized to the cell volume (see text)Treatments were as follows control 0 Cd (basal) 60min Cd (Cd)Cd + 30mM KCl (K) Cd + 30mM KCl and 1120583M BayK (B) Cd +30mM KCl and 1120583M nimodipine (N) Depolarization (treatmentK) did not increase significantly the intracellular concentration ofCd (119875 gt 005) In CHOC120572 cells but not in wild-type CHO BayKsignificantly enhanced and nimodipine significantly reduced Cdaccumulation both with 119875 lt 0001 (lowast) with respect to the sametreatments in depolarization Bars represent mean plusmn sem in at least10 experiments in each group
Acknowledgments
The author thanks Ilaria Zanardi and Irena Baranowska-Bosiacka (PUM Szczecin Poland) for help in Cd treatmentexperiments and Elisabetta Morelli for FAAS measurementsThe author is also grateful to Franz Hofmann and NorbertKlugbauer (Institut fur Pharmakologie und ToxikologieMunich Germany) for their permission to use the perma-nently transfected CHO cells and to Maurizio Taglialatelaand Mauro Cataldi (Universita Federico II Naples Italy) forsending him the cells
References
[1] R J Castellani P IMoreira G Liu et al ldquoIron the redox-activecenter of oxidative stress in Alzheimer diseaserdquo NeurochemicalResearch vol 32 no 10 pp 1640ndash1645 2007
[2] D G Smith R Cappai and K J Barnham ldquoThe redox chem-istry of the Alzheimerrsquos disease amyloid 120573 peptiderdquo Biochimicaet Biophysica Acta vol 1768 no 8 pp 1976ndash1990 2007
[3] Y Xu L Feng P D Jeffrey Y Shi and FMMMorel ldquoStructureand metal exchange in the cadmium carbonic anhydrase ofmarine diatomsrdquo Nature vol 452 no 7183 pp 56ndash61 2008
[4] T W Clarkson ldquoMolecular and ionic mimicry of toxic metalsrdquoAnnual Review of Pharmacology and Toxicology vol 33 pp545ndash571 1993
[5] C C Bridges and R K Zalups ldquoMolecular and ionic mimicryand the transport of toxic metalsrdquo Toxicology and Applied
Pharmacology vol 204 no 3 pp 274ndash308 2005[6] H Ouyang and H J Vogel ldquoMetal ion binding to calmodulin
NMRand fluorescence studiesrdquoBioMetals vol 11 no 3 pp 213ndash222 1998
[7] M KernMWisniewski L Cabell andG Audesirk ldquoInorganiclead and calcium interact positively in activation of calmod-ulinrdquo NeuroToxicology vol 21 no 3 pp 353ndash364 2000
[8] J Markovac and GW Goldstein ldquoPicomolar concentrations oflead stimulate brain protein kinase CrdquoNature vol 334 no 6177pp 71ndash73 1988
[9] X Sun X Tian J L Tomsig and J B Suszkiw ldquoAnalysisof differential effects of Pb2+ on protein kinase C isozymesrdquoToxicology and Applied Pharmacology vol 156 no 1 pp 40ndash451999
[10] J L Tomsig and J B Suszkiw ldquoMultisite interactions betweenPb2+ and protein kinase C and its role in norepinephrine releasefrom bovine adrenal chromaffin cellsrdquo Journal of Neurochem-istry vol 64 no 6 pp 2667ndash2673 1995
[11] C M Bouton L P Frelin C E Forde H A Godwin and JPevsner ldquoSynaptotagmin I is amolecular target for leadrdquo Journalof Neurochemistry vol 76 no 6 pp 1724ndash1735 2001
[12] J Yang P T Ellinor W A Sather J F Zhang and R W TsienldquoMolecular determinants of Ca2+ selectivity and ion permeationin L-type Ca2+ channelsrdquoNature vol 366 no 6451 pp 158ndash1611993
[13] MWakamori M Strobeck T Niidome T Teramoto K Imotoand Y Mori ldquoFunctional characterization of ion permeationpathway in the N-type Ca2+ channelrdquo Journal of Neurophysiol-ogy vol 79 no 2 pp 622ndash634 1998
[14] S Peng R K Hajela and W D Atchison ldquoCharacteristics ofblock by Pb2+ of function of human neuronal L- N- and R-type Ca2+ channels transiently expressed in human embryonickidney 293 cellsrdquo Molecular Pharmacology vol 62 no 6 pp1418ndash1430 2002
[15] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium uptake in excitable mammalian cells a microspec-trofluorimetric studyrdquo European Journal of Histochemistry vol41 pp 189ndash190 1997
[16] P M Hinkle P A Kinsella and K C Osterhoudt ldquoCadmiumuptake and toxicity via voltage-sensitive calcium channelsrdquoJournal of Biological Chemistry vol 262 no 34 pp 16333ndash163371987
[17] P M Hinkle E D Shanshala and E J Nelson ldquoMeasurementsof intracellular cadmium with fluorescent dyesmdashfurther evi-dence for the role of calcium channels in cadmium uptakerdquoTheJournal of Biological Chemistry vol 267 pp 25553ndash25559 1992
[18] P M Hinkle and M E Osborne ldquoCadmium toxicity in ratpheochromocytoma cells studies on themechanism of uptakerdquoToxicology and Applied Pharmacology vol 124 no 1 pp 91ndash981994
[19] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium influx in mammalian neuronsrdquo Journal of Neuro-chemistry vol 72 no 5 pp 2154ndash2161 1999
[20] V Souza L Bucio andMCGutierrez-Ruiz ldquoCadmiumuptakeby a human hepatic cell line (WRL-68 cells)rdquo Toxicology vol120 no 3 pp 215ndash220 1997
[21] P Gavazzo E Morelli and C Marchetti ldquoSusceptibility ofinsulinoma cells to cadmiumandmodulation by L-type calciumchannelsrdquo BioMetals vol 18 no 2 pp 131ndash142 2005
[22] M Kirberger and J J Yang ldquoStructural differences betweenPb2+- and Ca2+-binding sites in proteins implications with
ISRN Toxicology 9
respect to toxicityrdquo Journal of Inorganic Biochemistry vol 102no 10 pp 1901ndash1909 2008
[23] MMazzolini S Traverso andCMarchetti ldquoMultiple pathwaysof Pb2+ permeation in rat cerebellar granule neuronesrdquo Journalof Neurochemistry vol 79 no 2 pp 407ndash416 2001
[24] A Esposito M Robello F Pellistri and C Marchetti ldquoTwo-photon analysis of lead accumulation in rat cerebellar granuleneuronsrdquo Neurochemical Research vol 30 no 8 pp 949ndash9542005
[25] C Marchetti C Amico and C Usai ldquoFunctional characteriza-tion of the effect of nimodipine on the calcium current in ratcerebellar granule cellsrdquo Journal of Neurophysiology vol 73 no3 pp 1169ndash1180 1995
[26] A Welling E Bosse A Cavalie et al ldquoStable co-expression ofcalcium channel 1205721120573 and 1205722120575 subunits in a somatic cell linerdquoJournal of Physiology vol 471 pp 749ndash765 1993
[27] A Diaspro S Annunziata M Raimondo P Ramoino and MRobello ldquoA single-pinhole confocal laser scanning microscopefor 3-D imaging of biostructuresrdquo IEEE Engineering inMedicineand Biology Magazine vol 18 no 4 pp 106ndash110 1999
[28] F Pellistri A Cupello A Esposito C Marchetti and MRobello ldquoTwo-photon imaging of calcium accumulation in ratcerebellar granule cellsrdquo NeuroReport vol 15 no 1 pp 83ndash872004
[29] M Cataldi A Secondo A DrsquoAlessio et al ldquoStudies onmaitotoxin-induced intracellular Ca2+ elevation in chinesehamster ovary cells stably transfected with cDNAs encodingfor L-type Ca2+ channel subunitsrdquo Journal of Pharmacology andExperimental Therapeutics vol 290 no 2 pp 725ndash730 1999
[30] A Galan A Troyano N E Vilaboa C Fernandez E DeBlas and P Aller ldquoModulation of the stress response duringapoptosis and necrosis induction in cadmium-treated U-937human promonocytic cellsrdquo Biochimica et Biophysica Acta vol1538 no 1 pp 38ndash46 2001
[31] A H B Poliandri J P Cabilla M O Velardez C C ABodo and B H Duvilanski ldquoCadmium induces apoptosis inanterior pituitary cells that can be reversed by treatment withantioxidantsrdquo Toxicology and Applied Pharmacology vol 190no 1 pp 17ndash24 2003
[32] J L Tomsig and J B Suszkiw ldquoPermeation of Pb2+ throughcalcium channels fura-2 measurements of voltage- anddihydropyridine-sensitive Pb2+ entry in isolated bovinechromaffin cellsrdquo Biochimica et Biophysica Acta vol 1069 no2 pp 197ndash200 1991
[33] EM Leslie J Liu C D Klaassen andM PWaalkes ldquoAcquiredcadmium resistance in metallothionein-III(minusminus) knockoutcells role of the T-type calcium channel Cacn1205721G in cadmiumuptakerdquo Molecular Pharmacology vol 69 no 2 pp 629ndash6392006
[34] R Deane and M W B Bradbury ldquoTransport of lead-203 atthe blood-brain barrier during short cerebrovascular perfusionwith saline in the ratrdquo Journal of Neurochemistry vol 54 no 3pp 905ndash914 1990
[35] R S Diaz and J Monreal ldquoProtein-independent lead perme-ation through myelin lipid liposomesrdquo Molecular Pharmacol-ogy vol 47 no 4 pp 766ndash771 1995
[36] L E Kerper and P M Hinkle ldquoCellular uptake of lead isactivated by depletion of intracellular calcium storesrdquo Journalof Biological Chemistry vol 272 no 13 pp 8346ndash8352 1997
[37] L E Kerper and P M Hinkle ldquoLead uptake in brain capillaryendothelial cells activation by calcium store depletionrdquoToxicol-ogy and Applied Pharmacology vol 146 no 1 pp 127ndash133 1997
[38] M E Legare R Barhoumi E Hebert G R Bratton R CBurghardt and E Tiffany- Castiglioni ldquoAnalysis of Pb2+ entryinto cultured astrogliardquo Toxicological Sciences vol 46 no 1 pp90ndash100 1998
[39] J H Cheong D Bannon L Olivi Y Kim and J BresslerldquoDifferent mechanismsmediate uptake of lead in a rat astroglialcell linerdquoToxicological Sciences vol 77 no 2 pp 334ndash340 2004
[40] T J Evans M R James-Kracke S B Kleiboeker and S WCasteel ldquoLead enters Rcho-1 trophoblastic cells by calciumtransport mechanisms and complexes with cytosolic calcium-binding proteinsrdquo Toxicology and Applied Pharmacology vol186 no 2 pp 77ndash89 2003
[41] G J Kaczorowski R S Slaughter V F King and M LGarcia ldquoInhibitors of sodium-calcium exchange identificationand development of probes of transport activityrdquo Biochimica etBiophysica Acta vol 988 no 2 pp 287ndash302 1989
[42] S L Sensi L M T Canzoniero S P Yu et al ldquoMeasurement ofintracellular free zinc in living cortical neurons routes of entryrdquoJournal of Neuroscience vol 17 no 24 pp 9554ndash9564 1997
[43] R A Colvin ldquoCharacterization of a plasma membrane zinctransporter in rat brainrdquo Neuroscience Letters vol 247 no 2-3pp 147ndash150 1998
[44] V Picard G Govoni N Jabado and P Gross ldquoNramp 2(DCT1DMT1) expressed at the plasma membrane transportsiron and other divalent cations into a calcein-accessible cyto-plasmic poolrdquo Journal of Biological Chemistry vol 275 no 46pp 35738ndash35745 2000
[45] M Okubo K Yamada M Hosoyamada T Shibasaki and HEndou ldquoCadmium transport by human Nramp 2 expressed inXenopus laevis oocytesrdquo Toxicology and Applied Pharmacologyvol 187 no 3 pp 162ndash167 2003
[46] A C Illing A Shawki C L Cunningham and B MackenzieldquoSubstrate profile and metal-ion selectivity of human divalentmetal-ion transporter-1rdquo The Journal of Biological Chemistryvol 287 pp 30485ndash30496 2012
[47] J P Bressler L Olivi J H Cheong Y Kim A Maerten andD Bannon ldquoMetal transporters in intestine and brain theirinvolvement in metal-associated neurotoxicitiesrdquo Human andExperimental Toxicology vol 26 no 3 pp 221ndash229 2007
[48] Z Liu H Li M Soleimani et al ldquoCd2+ versus Zn2+ uptakeby the ZIP8 HCO
3mdashdependent symporter kinetics electro-
genicity and traffickingrdquo Biochemical and Biophysical ResearchCommunications vol 365 no 4 pp 814ndash820 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

2 ISRN Toxicology
these channels are regarded as the major route of their entryinto mammalian cells
Both Cd and Pb are well-known specific blockers ofvoltage-dependent Ca channels (VDCC) but the mechanismof block is quite different While Cd binds to the high affinitysite inside the VDCC pore [12 13] Pb competes with Ca at asite external to the VDCC and the binding is not voltage-dependent [14] Given these differences in the mechanismof interaction permeation of these metals through VDCCis also different VDCCs mediate Cd influx in excitable cells[15ndash18] including mammalian neurons [19] and have beenproposed to participate in Cd uptake also in cells fromnonexcitable tissues [20] In a previouswork ofmy laboratorywe showed that in certain cells Cd permeation occurs mainlythrough VDCC of the L-type [19 21] However in otherstudies the presence of VDCC per se did not seem to enhancesensitivity to Cd for example VDCCs expressing (PC12) andnonexpressing (PC18) cells showed comparable LD
50for Cd
[18] and the role of this pathway in the induction of cell deathappears questionable
The situation is even more complicated for Pb whosechemical basis formimickingCa is not obvious [4] andwhichis known to adapt to structurally diverse binding geometries[22] In cerebellar granule neurons Pb uses at least threepathways of permeation and besides voltage-dependent cal-cium channels (VDCC) and NMDA-activated channels israpidly taken up through passive transport system [23 24]
In this paper I will present experimental evidence con-cerning transport of Pb and Cd across cellular membranesand the relative contribution of VDCC in their uptake
2 Materials and Methods
21 Cell Culture Cerebellar granule neurons were preparedfrom 8-day-old Wistar rats as previously described [19 25]plated on 20mm poly-L-lysine coated glass coverslips andmaintained in basal Eaglersquos culture medium supplementedwith 10 fetal calf serum 100 120583gmL gentamicin and 25mMKCl in a humidified 95 airCO
2atmosphere at 37∘C
Cultures were treated with 10 120583M cytosine arabinoside fromday 1 to minimize the proliferation of nonneuronal cellsExperiments were carried out in cultures between 5 and 13days in vitro
Wild-type Chinese hamster ovary (CHO) cells were ob-tained from American Type Culture Collection CHO stablytransfected with cDNA encoding for several subunits ofvoltage-dependent calcium channels (VDCC) were a gener-ous gift of Franz Hofmann and Norbert Klugbauer (Institutfur Pharmakologie und Toxikologie Munich Germany)The cells used in this study are termed CHOC1205729120573
312057221205754
(abbreviated CHOC120572) to indicate that they express the 1205721119862minus119887
subunit of VDCC as well as 1205733and 120572
21205754 subunits from
smooth muscle [26]Both wild-type CHO and CHOC120572 cells were maintained
in DMEM medium (Sigma Chemical Co St Louis MOUSA) supplemented with 10 bovine serum in a 5 CO
2
humidified atmosphere at 37∘C For CHOC120572 cells the cul-ture medium was routinely supplemented with antibiotic
geneticin G418 20120583grmL For the experiments cells wereplated in 12-multiwell trays at a density of 2 times 105 cellsmL
22Microscopy Measurement of intracellular Cd and Pbwasperformed on monolayer cultures plated on glass coverslipsFor photometric measurements cells were incubated with6 120583M of the cell-permeant Fura2-AM ester form of the dyefor 45min at 37∘C and then washed several times withstandard saline at room temperature as previously described[19 23] Cd and Pb were applied in the absence of Caand treatment was terminated by the addition of themembrane-permeant metal chelator (NNN1015840N1015840-tetrakis-2-pyridyl methyl ethylenediamine TPEN 100 120583m)
For confocal microscopy cells were incubated with Ore-gonGreen 488Bapta-1 a fluorescent indicator whose fluores-cence intensity is increased upon binding Ca Cd or Pb Cellswere imaged using a confocal laser scanning microscopeNikon PCM2000 (Nikon Instr Florence Italy) with an oilimmersion 100 times objective (Na = 13) coupled to a 50120583mconfocal pinhole condition [27]
23 Cd Treatment Cells were subjected to 30ndash60min treat-ments with 50 or 100120583M CdCl
2in culture medium without
serum When specified the medium was supplemented with30mM KCl to depolarize the cell membrane and with thefollowing dihydropyridines (DHPs) BayK 8644 which is anL-type VDCC agonist or nimodipine a channel antagonistStock solutions of nimodipine and BayK (10mM) weremade up in 100 ethanol and diluted in medium to thefinal concentration High KCl alone as well as DHPs (noCd added) had no effect on cell viability and appearanceAfter incubation cells were washed three times with PBScontaining 1mMEDTA and freshmedium containing serumwas replaced Cultures were kept in the incubator for another18ndash24 hours before testing Cell viability in each samplewas assessed by trypan-blue exclusion assay All chemicalsculture media and sera were from Sigma-Aldrich Italy
24 Capacitance and Current Measurements by Patch-ClampTechnique Membrane currents were measured from wild-type CHO and permanently transfected CHOC120572 cells inwhole-cell clamp configuration by a patch-clamp Axopatchamplifier (Molecular Devices Corporation Union City CAUSA) Electrodes were manufactured from borosilicate glasscapillaries (Hilgenberg GmbH Malsfeld Germany) and hadresistance of asymp 4MΩ Voltage stimulation and data acqui-sition were performed by a PC through a Digidata 1440Ainterface and Pclamp-10 software (Molecular Devices) Cur-rents were low-pass filtered at 5 kHz and digitized at 10 kHzCapacitance transients wereminimized by analog compensa-tion and the value obtained by this compensation was takenas an estimate of the cell capacitance All currents traceswere further corrected for leak and residual transients by acomputer generated P4 protocol The holding potential wasminus80mV in all the experiments Current traces were analyzedwith Clampfit-10 and Sigma Plot (Jandel Scientific ErkrathGermany) software
Cells were continuously superfused by gravity flow(10mLmin) with a solution containing (in mM) NaCl 140
ISRN Toxicology 3
KCl 54 CaCl218 Hepes 5 The pH was 74 The internal
(pipette) solution contained (in mM) CsCl 20 CsOH 110Aspartic acid 100 EGTA 5 Hepes 5 with pH adjustedto 73 with Trizma base Calcium currents were recordedin similar external solution containing 130mM NaCl and5mMCaCl
2 Wild-type CHO cells do not possess prominent
voltage-dependent currents and CHOC120572 only contained theL-type VDCC under study therefore there was no needto antagonize the current through other voltage-dependentchannels to resolve Ca currentsThe application of modifierssuch as agonist and antagonist DHPs was accomplished bygravity flow control ion substitution experiments showedthat the external bathwas completely changed in 10 sec whichwas the maximal stimulation rate in these experiments
25 Absorption Spectroscopy Cd Determination Cells wereexposed to Cd in culture medium without added serumAfter treatment cultures were washed three times with PBScontaining 1mM EDTA and harvested immediately Viablecells were counted in triplicate in each sample Cells were thenwashed again three times with the above buffer resuspendedin distilled water and disrupted by sonication (SonopulsUltrasonic Homogenizer Bandelin) for 2 minutes in an icebath The total Cd content of each sample was measured byFlameless Atomic Absorption Spectroscopy (FAAS) using aPerkin Elmer Spectrophotometer (Model 1100 B) equippedwith a graphite furnace (Model HGA 700) The Cd contentwas normalized to the volume occupied by the cells andmolarity was calculated (see results)
26 Statistical Analysis Data are presented as mean plusmn stan-dard error of the mean in at least 3 experiments Statisticalsignificance was evaluated by Student-Newman-Keuls multi-ple comparison test (In Stat GraphPad Software)
3 Results And Discussion
31 Influx of Toxic Metals into Mammalian Cells Influxof toxic divalent metal ions into mammalian cells can bemonitored in cells preloaded with specific fluorescent probesFura2 [17 19 23] or Oregon green These fluorescent indica-tors were developed and are predominantly used for deter-mining intracellular calcium levels but they have relativelyhigh affinity also for other polyvalent ions including Cd andPb
Figure 1 shows how exposure to a solution containing atoxicologically significant concentration of Cd or Pb causesa sizeable increase in Oregon Green fluorescence similar tothat seen following Ca influx [28] This demonstrates fea-sibility of this probe as toxic metal influx indicator In thisexperiment while Cd uptakewas triggered by depolarizationindicating the essential contribution of VDCC (Figure 1(A)(a)-(b)) neurons rapidly accumulate Pb even in the absenceof a specific stimulus (Figure 1(A) (c)-(d))
Figure 1(B) shows real-time measurements of Cd andPb influx in Fura2-loaded neurons [19 23] The influx wasquantified by measuring the fluorescence emission ratio R(E380
E340
) and membranes were depolarized by increasing
concentration of external KCl [25] In the left panel thefluorescent ratio E
340E380
increased negligibly when 50120583MCd was applied in resting condition but the rate of risedRdt was enhanced by gt60 times following depolarizationwith 25KCl In contrast Pb (15ndash50120583M)determined a sizeableincrease of R even in the absence of any specific stimulus thatopens VDCC and dRdt was increased by 5 times from theresting level with 75mMKCl (Figure 1(C)) Fast uptake of Pbin the absence of any specific stimulus appears to be a peculiarfeature of neurons and may be linked to the specific lipidiccomposition of the neuronal membranes while the role ofVDCC is likely to significant but not essential [23]
32 Role of VDCC in Cd-Induced Mortality As VDCC chan-nels and in particular those of L-type [15 19] play a promi-nent role in uptake of Cd by excitable cells the involvement ofthese channels in Cd toxicity was investigated further Tests ofmortality chromatin condensation and DNA fragmentationshowed that a pulsed Cd treatment induces delayed apop-totic cell death in VDCC-containing insulinoma cells andthat nimodipine protects against Cd-induced apoptosis andnecrosis in these cells supporting a specific involvement of L-type VDCC [21] The same treatments were largely harmlessin VDCC-free HeLa cell cultures in which neither death norDNA fragmentation was observed Therefore excitable cellsappear more susceptible than nonexcitable epithelial-likecells to Cd accumulation However it is not clear whether themere presence of VDCC can make cells more vulnerable totoxicmetal injury To answer this question we have comparedthe effects of Cd in two cell types that are virtually identicalexcept for the expression of VDCC in one of the two Thisapproach is made possible by the availability of a CHO cellline that has been permanently transfected with VDCC 120572
1
120573 and 1205722120575 subunits CHOC1205729120573
312057221205754cells [26 29] Promi-
nent voltage-dependent Ca currents were recorded fromthe permanently transfected cell line while such currentsare completely absent in wild-type CHO (Figure 2(a)) Thecurrents are larger with Ba as charge carrier and are sensitiveto DHP modulation (Figure 2(b)) as expected for an L-typeCa channel
Both cell types (wild-type and CHOC120572) were subjectedto a ldquopulse treatmentrdquo with 50 or 100 120583M Cd in serum-freeculture medium and observed after 18ndash24 hour after washCell suffering and mortality was negligible immediately afterthe treatment but it was evident after at least 16 hours Aspreviously observed [21 30 31] Cd accumulates into the cellduring treatment but the toxic mechanism leading to celldeath becomes effective at a later time Figure 3 shows theappearance of the cell monolayer 24 hours after an incubationof 30min with 100120583M Cd Cells exposed to Cd were ofsmaller size than control cells and had an increased tendencyto detach from their neighbors and from the substrate andfloat In addition CHOC120572VDCC-expressing cells frequentlyacquired an irregular shape with shredded edges (Figure 3)The number of viable cells was estimated from the numberof adherent trypan-blue excluding cells and this numberwas normalized to that of adherent viable cells in controlconditions as in our previous works [21] Despite some mor-phological features the dose dependence of Cd sensitivity
4 ISRN Toxicology
08
06
04
02
0
TPEN
TPEN
TPEN
2 min
25 KCl25 KCl
50 Cd 50 Pb15 Pb
75 KCl
Fluo
resc
ence
ratio
Stnd 0 Ca
lowast
lowast
lowast
100
50
0
(A) (B)
(C)
(a) (b)
(c) (d)
25 KCl75 KCl
5 15 50
119889119877119889119905(
minminus1)lowast
103
Figure 1 Uptake of Cd and Pb in cerebellar granule neurons measured by fluorescent dyes (A) Confocal microscopy images of Cd (a b) andPb (c d) uptake by cerebellar granule cells preloaded with the divalent metal-sensitive dye Oregon Green In (a) and (c) neurons were bathedin a physiological saline In (b) they have been superfused with a solution containing 0 Ca 30mM KCl and 100 120583M Cd Cl
2 which caused
the dye fluorescence to increase significantly In (d) external solution contained 0Ca and Pb 15120583M which permeates through the neuronmembrane even in the absence of a depolarizing stimulus and also caused an increase in the dye fluorescence Bar = 10 120583M (B) Real-timerecording of the influx of Cd and Pb in Fura-2 loaded cerebellar granule neuron and the effect of membrane depolarization Cells were treatedwith the metals in nominal Ca-free solution From left to right time course of the fluorescence ratio 119877 = E340E380 following application of50 120583MCd and membrane depolarization (25mM KCl) and application of 15ndash50120583M Pb in resting conditions and increasing depolarizations(25 and 75mM KCl) (C) Summary of the results obtained in basal and in depolarizing solutions (25 and 75mM KCl) with different doses ofPb The time course of R(t) was approximated by a straight line and the slope dRdt was calculated in each case as a measure of Pb influxData are mean plusmn SEM in 4 experiments and lowastindicates significantly different from basal (no depolarization) for each dose of Pb (119875 lt 005)
was very similar in the two cell types (Figure 3) Thereforeby this approach it is not possible to demonstrate a clear roleof VDCC in mediating Cd-induced cell detachment
In subsequent experiments we challenged the cells withCd in the presence of 1120583M DHP modifiers When VDCC-expressing CHOC120572 cells were exposed to Cd in the presence
ISRN Toxicology 5
(a)
Wild type
100 pA
10 ms
(b)
5 mM Ca
5 mM Ba
50 pA
10 ms
Nimo 5 Ba 5 Ca
0 60 120 180 240 300
Curr
ent (
pA)
0
minus100
minus200
(c)
Figure 2 CHOC120572 cells express L-type VDCC (a) Representative microphotographs of monolayer cell cultures of permanently transfectedCHOC120572 cells (left) and wild-type CHO cells (right) Bar = 35 120583M (b) Representative current traces evoked by depolarizing voltage stepsof 40ms duration from minus60 to +80mV from a holding potential of minus80mV in a CHOC120572 cell (left) and a wild-type CHO cell (right) Theexternal solution contained 5mMCaCl
2 This protocol evoked voltage-dependent calcium current in the permanently transfected cell while
no current was present in CHO cell of the wild-type (c) Characterization of the Ca current in CHOC120572 cellThe current was increased bymorethan 50 when the external solution was changed from 5mM CaCl
2to 5mM BaCl
2 and it was reversibly blocked by 10 120583M nimodipine
Current traces evoked by 50ms depolarizing steps from minus80mV (holding potential) to +10mV in the three conditions are shown on the leftThe graph on the right shows the time course of the experiment
of the antagonist (nimodipina) or agonist (BayK 8644) theeffect of the metal was clearly modified with respect to cellsexposed to Cd alone (Figure 4) with a sizeable increase ofmortality in the presence of BayK and protection in the pres-ence of nimodipineDHPmodifierswere largely ineffective inwild-type CHO cells exposed to Cd (not shown) Thereforeit seems that the actual contribution of L-type VDCC to Cduptake is clearly underscored only in the presence of thesemodifiers
Another indicator of apoptosis is cell shrinkage whichcan be quantified by cell capacitance measurements Bothwild-type and CHOC120572 cells showed a decrease in cell capaci-tance 18ndash24 hours afterCd treatment Inwild-type cell capac-itance was 24plusmn2 pF (119899 = 15) in control and 15plusmn 3 pF (119899 = 6)after incubation with 100 120583MCd significantly different fromcontrol with 119875 lt 005 In CHOC120572 cells cell capacitance was35 plusmn 3 pF (119899 = 17) in control and 22 plusmn 2 pF (119899 = 39)after incubation with 100 120583MCd significantly different from
control with 119875 lt 0001 In CHOC120572 cells the effect ofCd treatment was partially prevented by nimodipine thecapacitance was 35plusmn3 pF (119899 = 10) and the difference betweenthis condition and control condition was not significant(Figure 5)This indicates again that the contribution of L-typeVDCC in Cd-induced toxicity is evident only in the presenceof DHPs
Finally we performedmeasurements of total Cd accumu-lation during 60min incubation time by atomic absorptionspectroscopy For these experiments cells were harvestedimmediately after the treatment and the total amount of Cdwas normalized to the cell volume estimated from the single-cell capacitance Assuming that both cell types are approxi-mately spherical and have a specific membrane capacitanceof 1 120583Fcm2 106 wild-type CHO cells comprise a volume of11 120583L while 106 CHOC120572 cells a volume of 195120583L Thisdifference is not negligible and with this method the resultswere considerably more reproducible than using a standard
6 ISRN Toxicology
1
08
06
04
02
01 10 100 1000
Wild type+VDCC
(a) (b)
(c) (d)
(e)
Figure 3 Effect of pulse treatment with Cd in CHOC120572 VDCC-expressing and wild-type CHO cells Microphotographs of control (a b) andCd-treated (c d) cells (a c) VDCC-expressing CHOC120572 cells and (b d) wild-type CHO cells Both cell types were incubated in 100120583M Cdfor 60 minutes in the presence of 30mM KCl Pictures were taken 24 h after wash of the metal Bar = 35 120583MThe graph (e) shows the effectof a 30min pulse treatment with Cd on cell adhesion as a function of concentration Trypan-blue excluding adherent cells were counted24 h after wash of the metal Points are average plusmn sem of 3 experiments in the same condition in CHOC120572 (filled circles) and wild-type CHO(empty circles) and were best fitted to the function NadherNadher(control) = 1(1 + ([Cd]ED50)) where NadherNadher(control) is the number ofadherent cells after Cd treatment normalized to the number of adherent cells in control culture [Cd] is the concentration of Cd and ED
50is
the concentration of Cd that causes detachment from the substrate of 50 of cells The best fit yielded ED50= 40 120583M for CHOC120572 and 43120583M
for wild-type CHOThe two curves are overlapped In contrast with the different appearance the two cell types were similarly affected by Cdtreatment
normalization to the total protein content Data shown inFigure 6 indicate that the difference in Cd accumulation isbarely or not significant when both cell types are incubatedin 100120583M Cd with or without elevated KCl On the otherhand BayK 8644 significantly enhanced and nimodipinesignificantly decreased metal accumulation in depolarizedCHOC120572 cells treated with 50 or 100120583M Cd but not in wild-type CHO cells
Toxicity of both Cd and Pb is mainly due to their abilityto permeate not simply mammalian cell membranes butspecifically tight membrane barriers such as the epitheliallining in the gastrointestinal tract luminal membrane ofproximal tubules in the kidney and the blood-brain barrierIn early work the influx of Pb and Cd into mammaliancells was frequently ascribed to permeation through calciumchannels and in particular L-type DHP-sensitive VDCC [1618 20 32]More recently other types of VDCC different fromthe classical DHP-sensitive L-type have been implicated bothin Cd [19 33] and Pb [23] uptake Moreover Pb has beenshown to use different pathways in several systems [34ndash37]and it is largely independent of VDCC even in neurons[23 24]
Here I have shown that expression of L-type VDCC is notsufficient to modify Cd uptake to a toxicological significantextent at least in the case of a brief intense exposure asthose used in this study Cd accumulation and sensitivitywerecomparable in two cell lines whose only difference was thepresence of L-typeVDCC in one of the two Indeedwild-type
CHO cells which do not possess VDCC are vulnerable to Cdpoisoning and take up Cd during relatively brief exposuresto an extent sufficient to trigger cell death by apoptosisor necrosis similarly to VDCC expressing CHOC120572 cellsAlthough VDCCs play an important role in Cd uptakein CHOC120572 cells as indicated by the sizeable effect of L-type channel DHP modifiers wild-type CHO cells use othertransport proteins with similar efficiency and outcome
It is thus clear that VDCCs are not the main mecha-nism responsible for toxic metal permeation through cellmembranes and tight barriers Rapid passive transport of Pbindependent of VDCC was reported at the brain endothe-lium [34 36 37] and even protein-independent transportof lead has been described [35] In astroglia cell models theuptake of Pb was shown to be driven by store-depletion-activated channels [38] and by two distinct pH-sensitivetransport mechanisms [39] VDCC-independent uptake ofPb by cerebellar granule neurons was inhibited by La [24]and this observation can provide some clue as for the mecha-nism of permeationThe involvement of a La-sensitive store-depletion activated channel as proposed in other systems[36 37 40] is unlikely because Pb influx did not requiredrainage of the stores Another possibility is that Pb2+ entersthe neuron via reverse operation of an exchanger similar (oridentical) to theNandashCa exchanger which is also very sensitiveto La block [41] and whose role in Zn permeation wasdemonstrated in cortical neurons [42] Also this possibilitywas discarded because NandashCa exchanger does not play
ISRN Toxicology 7
4
3
2
1
0Basal Cd K N B
Treatments
lowast
lowastlowast
(a) (b)
(c) (d)
(e)
Figure 4 Effect of dihydropyridines on Cd cytotoxicity in VDCC-expressing CHOC120572 cells Cells were treated for 30min in (a) controlmediumcontaining 30mMKCl (b) 100 120583M Cd+30mM KCl (c) 100 120583M Cd+30mM KCl+1 120583Mnimodipine (d) 100 120583M Cd+30mM KCl+1 120583M BayK8644 Note the partial recovery in shape of the cells treated in the presence of nimodipine and definitive lost of adhesion whenthe treatment was performed in the presence of BayK Bar = 25 120583M The graph (e) represents counts of viable adherent cells in the sameexperiment (average plusmn sem in 3 samples) in control (basal) following for a 30min treatment with 100 120583M Cd (Cd) Cd + 30mM KCl (K)Cd + 30mM KCl and 1 120583M nimodipine (N) and Cd + 30mM KCl and 1 120583M BayK (B) lowast indicates significantly different from control with119875 lt 005 and lowastlowast with 119875 lt 00001
40
30
20
10
0Wild type
ControlCd
Capa
cita
nce (
pf)
lowast
lowastlowast
Cd + nimo
+VDCC
Figure 5 Evidence of Cd-induced apoptosis change in cell capac-itance Cells were incubated with 100120583M Cd for 60 minutes andelectrical measurements were performed 24 h after wash Mem-brane capacitance was estimated from transient compensation (seeSection 24) Bars represent mean plusmn sem Treatment with Cd causedshrinkage of all cell as revealed by reduction of the cell capacitancein both wild-type (119899 = 15 in control and 119899 = 6 with Cd treatment)and CHOC120572 cells (119899 = 17 in control and 119899 = 39with Cd treatment)Nimodipine protected CHOC120572 cells from Cd-induced shrinkage(119899 = 10) lowast indicates significantly different from control with 119875 lt005 and lowastlowast with 119875 lt 0001
a significant role when the Na concentrations are close tothe physiological value as in Esposito et al [24] It is more
probable that Pb is taken up through a La-sensitive ion carriersimilar to the Zn transporter found in brain neurons [43]whose identity and features still wait full definition
Involvement of different transport proteins in Cd trans-port has been described At the epithelial lining of the gas-trointestinal barrier DMT1 the main transporter thatabsorbs Fe in the brush border membrane of the mammalianintestine has been shown to carry also different toxic metals[44ndash46] The role of this transporter in Cd absorption isunderscored by the fact that iron deficiency creates a sig-nificant risk for increased cadmium exposure by increasinggastrointestinal absorption from 5 to as much as 20In general a low level of essential metals favors uptake ofnonessential (toxic) elements that compete for their site andthis stresses the importance of evaluating metal balance incontrast with metal concentration In addition other metal-specific transporters have been identified at the cDNA level[47] and studied in expression systems [48]
4 Conclusion
Ionicmimicry is a useful framework to study themechanismsof metal toxicity Uptake of toxic metals such as Cd and Pbby mammalian cells of different tissues occurs through manydifferent pathways of which VDCCs have frequently beenregarded as prominent Because of the abundance of VDCCexcitable cells may be more susceptible to accumulate Cdand Pb than nonexcitable cells In this chapter I have shownthat although Cd and Pb can permeate through VDCC thisis not the most relevant mechanism of passive uptake ofPb while the mere presence of VDCC is not sufficient tomake cells significantly more vulnerable to Cd injury Otherfactors including different transport systems and expressionof detoxification proteins may be more relevant in the accu-mulation process of these toxic metals
8 ISRN Toxicology
Basal Cd K B NTreatments
Wild type
80
60
40
20
0
lowast
lowast
Figure 6 Total Cd accumulation measured by FAAS in CHOC120572and wild-type CHO cells Both cell types were incubated for 1 hourin medium containing 0 50 or 100 120583M Cd and other modifiersCd determination was normalized to the cell volume (see text)Treatments were as follows control 0 Cd (basal) 60min Cd (Cd)Cd + 30mM KCl (K) Cd + 30mM KCl and 1120583M BayK (B) Cd +30mM KCl and 1120583M nimodipine (N) Depolarization (treatmentK) did not increase significantly the intracellular concentration ofCd (119875 gt 005) In CHOC120572 cells but not in wild-type CHO BayKsignificantly enhanced and nimodipine significantly reduced Cdaccumulation both with 119875 lt 0001 (lowast) with respect to the sametreatments in depolarization Bars represent mean plusmn sem in at least10 experiments in each group
Acknowledgments
The author thanks Ilaria Zanardi and Irena Baranowska-Bosiacka (PUM Szczecin Poland) for help in Cd treatmentexperiments and Elisabetta Morelli for FAAS measurementsThe author is also grateful to Franz Hofmann and NorbertKlugbauer (Institut fur Pharmakologie und ToxikologieMunich Germany) for their permission to use the perma-nently transfected CHO cells and to Maurizio Taglialatelaand Mauro Cataldi (Universita Federico II Naples Italy) forsending him the cells
References
[1] R J Castellani P IMoreira G Liu et al ldquoIron the redox-activecenter of oxidative stress in Alzheimer diseaserdquo NeurochemicalResearch vol 32 no 10 pp 1640ndash1645 2007
[2] D G Smith R Cappai and K J Barnham ldquoThe redox chem-istry of the Alzheimerrsquos disease amyloid 120573 peptiderdquo Biochimicaet Biophysica Acta vol 1768 no 8 pp 1976ndash1990 2007
[3] Y Xu L Feng P D Jeffrey Y Shi and FMMMorel ldquoStructureand metal exchange in the cadmium carbonic anhydrase ofmarine diatomsrdquo Nature vol 452 no 7183 pp 56ndash61 2008
[4] T W Clarkson ldquoMolecular and ionic mimicry of toxic metalsrdquoAnnual Review of Pharmacology and Toxicology vol 33 pp545ndash571 1993
[5] C C Bridges and R K Zalups ldquoMolecular and ionic mimicryand the transport of toxic metalsrdquo Toxicology and Applied
Pharmacology vol 204 no 3 pp 274ndash308 2005[6] H Ouyang and H J Vogel ldquoMetal ion binding to calmodulin
NMRand fluorescence studiesrdquoBioMetals vol 11 no 3 pp 213ndash222 1998
[7] M KernMWisniewski L Cabell andG Audesirk ldquoInorganiclead and calcium interact positively in activation of calmod-ulinrdquo NeuroToxicology vol 21 no 3 pp 353ndash364 2000
[8] J Markovac and GW Goldstein ldquoPicomolar concentrations oflead stimulate brain protein kinase CrdquoNature vol 334 no 6177pp 71ndash73 1988
[9] X Sun X Tian J L Tomsig and J B Suszkiw ldquoAnalysisof differential effects of Pb2+ on protein kinase C isozymesrdquoToxicology and Applied Pharmacology vol 156 no 1 pp 40ndash451999
[10] J L Tomsig and J B Suszkiw ldquoMultisite interactions betweenPb2+ and protein kinase C and its role in norepinephrine releasefrom bovine adrenal chromaffin cellsrdquo Journal of Neurochem-istry vol 64 no 6 pp 2667ndash2673 1995
[11] C M Bouton L P Frelin C E Forde H A Godwin and JPevsner ldquoSynaptotagmin I is amolecular target for leadrdquo Journalof Neurochemistry vol 76 no 6 pp 1724ndash1735 2001
[12] J Yang P T Ellinor W A Sather J F Zhang and R W TsienldquoMolecular determinants of Ca2+ selectivity and ion permeationin L-type Ca2+ channelsrdquoNature vol 366 no 6451 pp 158ndash1611993
[13] MWakamori M Strobeck T Niidome T Teramoto K Imotoand Y Mori ldquoFunctional characterization of ion permeationpathway in the N-type Ca2+ channelrdquo Journal of Neurophysiol-ogy vol 79 no 2 pp 622ndash634 1998
[14] S Peng R K Hajela and W D Atchison ldquoCharacteristics ofblock by Pb2+ of function of human neuronal L- N- and R-type Ca2+ channels transiently expressed in human embryonickidney 293 cellsrdquo Molecular Pharmacology vol 62 no 6 pp1418ndash1430 2002
[15] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium uptake in excitable mammalian cells a microspec-trofluorimetric studyrdquo European Journal of Histochemistry vol41 pp 189ndash190 1997
[16] P M Hinkle P A Kinsella and K C Osterhoudt ldquoCadmiumuptake and toxicity via voltage-sensitive calcium channelsrdquoJournal of Biological Chemistry vol 262 no 34 pp 16333ndash163371987
[17] P M Hinkle E D Shanshala and E J Nelson ldquoMeasurementsof intracellular cadmium with fluorescent dyesmdashfurther evi-dence for the role of calcium channels in cadmium uptakerdquoTheJournal of Biological Chemistry vol 267 pp 25553ndash25559 1992
[18] P M Hinkle and M E Osborne ldquoCadmium toxicity in ratpheochromocytoma cells studies on themechanism of uptakerdquoToxicology and Applied Pharmacology vol 124 no 1 pp 91ndash981994
[19] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium influx in mammalian neuronsrdquo Journal of Neuro-chemistry vol 72 no 5 pp 2154ndash2161 1999
[20] V Souza L Bucio andMCGutierrez-Ruiz ldquoCadmiumuptakeby a human hepatic cell line (WRL-68 cells)rdquo Toxicology vol120 no 3 pp 215ndash220 1997
[21] P Gavazzo E Morelli and C Marchetti ldquoSusceptibility ofinsulinoma cells to cadmiumandmodulation by L-type calciumchannelsrdquo BioMetals vol 18 no 2 pp 131ndash142 2005
[22] M Kirberger and J J Yang ldquoStructural differences betweenPb2+- and Ca2+-binding sites in proteins implications with
ISRN Toxicology 9
respect to toxicityrdquo Journal of Inorganic Biochemistry vol 102no 10 pp 1901ndash1909 2008
[23] MMazzolini S Traverso andCMarchetti ldquoMultiple pathwaysof Pb2+ permeation in rat cerebellar granule neuronesrdquo Journalof Neurochemistry vol 79 no 2 pp 407ndash416 2001
[24] A Esposito M Robello F Pellistri and C Marchetti ldquoTwo-photon analysis of lead accumulation in rat cerebellar granuleneuronsrdquo Neurochemical Research vol 30 no 8 pp 949ndash9542005
[25] C Marchetti C Amico and C Usai ldquoFunctional characteriza-tion of the effect of nimodipine on the calcium current in ratcerebellar granule cellsrdquo Journal of Neurophysiology vol 73 no3 pp 1169ndash1180 1995
[26] A Welling E Bosse A Cavalie et al ldquoStable co-expression ofcalcium channel 1205721120573 and 1205722120575 subunits in a somatic cell linerdquoJournal of Physiology vol 471 pp 749ndash765 1993
[27] A Diaspro S Annunziata M Raimondo P Ramoino and MRobello ldquoA single-pinhole confocal laser scanning microscopefor 3-D imaging of biostructuresrdquo IEEE Engineering inMedicineand Biology Magazine vol 18 no 4 pp 106ndash110 1999
[28] F Pellistri A Cupello A Esposito C Marchetti and MRobello ldquoTwo-photon imaging of calcium accumulation in ratcerebellar granule cellsrdquo NeuroReport vol 15 no 1 pp 83ndash872004
[29] M Cataldi A Secondo A DrsquoAlessio et al ldquoStudies onmaitotoxin-induced intracellular Ca2+ elevation in chinesehamster ovary cells stably transfected with cDNAs encodingfor L-type Ca2+ channel subunitsrdquo Journal of Pharmacology andExperimental Therapeutics vol 290 no 2 pp 725ndash730 1999
[30] A Galan A Troyano N E Vilaboa C Fernandez E DeBlas and P Aller ldquoModulation of the stress response duringapoptosis and necrosis induction in cadmium-treated U-937human promonocytic cellsrdquo Biochimica et Biophysica Acta vol1538 no 1 pp 38ndash46 2001
[31] A H B Poliandri J P Cabilla M O Velardez C C ABodo and B H Duvilanski ldquoCadmium induces apoptosis inanterior pituitary cells that can be reversed by treatment withantioxidantsrdquo Toxicology and Applied Pharmacology vol 190no 1 pp 17ndash24 2003
[32] J L Tomsig and J B Suszkiw ldquoPermeation of Pb2+ throughcalcium channels fura-2 measurements of voltage- anddihydropyridine-sensitive Pb2+ entry in isolated bovinechromaffin cellsrdquo Biochimica et Biophysica Acta vol 1069 no2 pp 197ndash200 1991
[33] EM Leslie J Liu C D Klaassen andM PWaalkes ldquoAcquiredcadmium resistance in metallothionein-III(minusminus) knockoutcells role of the T-type calcium channel Cacn1205721G in cadmiumuptakerdquo Molecular Pharmacology vol 69 no 2 pp 629ndash6392006
[34] R Deane and M W B Bradbury ldquoTransport of lead-203 atthe blood-brain barrier during short cerebrovascular perfusionwith saline in the ratrdquo Journal of Neurochemistry vol 54 no 3pp 905ndash914 1990
[35] R S Diaz and J Monreal ldquoProtein-independent lead perme-ation through myelin lipid liposomesrdquo Molecular Pharmacol-ogy vol 47 no 4 pp 766ndash771 1995
[36] L E Kerper and P M Hinkle ldquoCellular uptake of lead isactivated by depletion of intracellular calcium storesrdquo Journalof Biological Chemistry vol 272 no 13 pp 8346ndash8352 1997
[37] L E Kerper and P M Hinkle ldquoLead uptake in brain capillaryendothelial cells activation by calcium store depletionrdquoToxicol-ogy and Applied Pharmacology vol 146 no 1 pp 127ndash133 1997
[38] M E Legare R Barhoumi E Hebert G R Bratton R CBurghardt and E Tiffany- Castiglioni ldquoAnalysis of Pb2+ entryinto cultured astrogliardquo Toxicological Sciences vol 46 no 1 pp90ndash100 1998
[39] J H Cheong D Bannon L Olivi Y Kim and J BresslerldquoDifferent mechanismsmediate uptake of lead in a rat astroglialcell linerdquoToxicological Sciences vol 77 no 2 pp 334ndash340 2004
[40] T J Evans M R James-Kracke S B Kleiboeker and S WCasteel ldquoLead enters Rcho-1 trophoblastic cells by calciumtransport mechanisms and complexes with cytosolic calcium-binding proteinsrdquo Toxicology and Applied Pharmacology vol186 no 2 pp 77ndash89 2003
[41] G J Kaczorowski R S Slaughter V F King and M LGarcia ldquoInhibitors of sodium-calcium exchange identificationand development of probes of transport activityrdquo Biochimica etBiophysica Acta vol 988 no 2 pp 287ndash302 1989
[42] S L Sensi L M T Canzoniero S P Yu et al ldquoMeasurement ofintracellular free zinc in living cortical neurons routes of entryrdquoJournal of Neuroscience vol 17 no 24 pp 9554ndash9564 1997
[43] R A Colvin ldquoCharacterization of a plasma membrane zinctransporter in rat brainrdquo Neuroscience Letters vol 247 no 2-3pp 147ndash150 1998
[44] V Picard G Govoni N Jabado and P Gross ldquoNramp 2(DCT1DMT1) expressed at the plasma membrane transportsiron and other divalent cations into a calcein-accessible cyto-plasmic poolrdquo Journal of Biological Chemistry vol 275 no 46pp 35738ndash35745 2000
[45] M Okubo K Yamada M Hosoyamada T Shibasaki and HEndou ldquoCadmium transport by human Nramp 2 expressed inXenopus laevis oocytesrdquo Toxicology and Applied Pharmacologyvol 187 no 3 pp 162ndash167 2003
[46] A C Illing A Shawki C L Cunningham and B MackenzieldquoSubstrate profile and metal-ion selectivity of human divalentmetal-ion transporter-1rdquo The Journal of Biological Chemistryvol 287 pp 30485ndash30496 2012
[47] J P Bressler L Olivi J H Cheong Y Kim A Maerten andD Bannon ldquoMetal transporters in intestine and brain theirinvolvement in metal-associated neurotoxicitiesrdquo Human andExperimental Toxicology vol 26 no 3 pp 221ndash229 2007
[48] Z Liu H Li M Soleimani et al ldquoCd2+ versus Zn2+ uptakeby the ZIP8 HCO
3mdashdependent symporter kinetics electro-
genicity and traffickingrdquo Biochemical and Biophysical ResearchCommunications vol 365 no 4 pp 814ndash820 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

ISRN Toxicology 3
KCl 54 CaCl218 Hepes 5 The pH was 74 The internal
(pipette) solution contained (in mM) CsCl 20 CsOH 110Aspartic acid 100 EGTA 5 Hepes 5 with pH adjustedto 73 with Trizma base Calcium currents were recordedin similar external solution containing 130mM NaCl and5mMCaCl
2 Wild-type CHO cells do not possess prominent
voltage-dependent currents and CHOC120572 only contained theL-type VDCC under study therefore there was no needto antagonize the current through other voltage-dependentchannels to resolve Ca currentsThe application of modifierssuch as agonist and antagonist DHPs was accomplished bygravity flow control ion substitution experiments showedthat the external bathwas completely changed in 10 sec whichwas the maximal stimulation rate in these experiments
25 Absorption Spectroscopy Cd Determination Cells wereexposed to Cd in culture medium without added serumAfter treatment cultures were washed three times with PBScontaining 1mM EDTA and harvested immediately Viablecells were counted in triplicate in each sample Cells were thenwashed again three times with the above buffer resuspendedin distilled water and disrupted by sonication (SonopulsUltrasonic Homogenizer Bandelin) for 2 minutes in an icebath The total Cd content of each sample was measured byFlameless Atomic Absorption Spectroscopy (FAAS) using aPerkin Elmer Spectrophotometer (Model 1100 B) equippedwith a graphite furnace (Model HGA 700) The Cd contentwas normalized to the volume occupied by the cells andmolarity was calculated (see results)
26 Statistical Analysis Data are presented as mean plusmn stan-dard error of the mean in at least 3 experiments Statisticalsignificance was evaluated by Student-Newman-Keuls multi-ple comparison test (In Stat GraphPad Software)
3 Results And Discussion
31 Influx of Toxic Metals into Mammalian Cells Influxof toxic divalent metal ions into mammalian cells can bemonitored in cells preloaded with specific fluorescent probesFura2 [17 19 23] or Oregon green These fluorescent indica-tors were developed and are predominantly used for deter-mining intracellular calcium levels but they have relativelyhigh affinity also for other polyvalent ions including Cd andPb
Figure 1 shows how exposure to a solution containing atoxicologically significant concentration of Cd or Pb causesa sizeable increase in Oregon Green fluorescence similar tothat seen following Ca influx [28] This demonstrates fea-sibility of this probe as toxic metal influx indicator In thisexperiment while Cd uptakewas triggered by depolarizationindicating the essential contribution of VDCC (Figure 1(A)(a)-(b)) neurons rapidly accumulate Pb even in the absenceof a specific stimulus (Figure 1(A) (c)-(d))
Figure 1(B) shows real-time measurements of Cd andPb influx in Fura2-loaded neurons [19 23] The influx wasquantified by measuring the fluorescence emission ratio R(E380
E340
) and membranes were depolarized by increasing
concentration of external KCl [25] In the left panel thefluorescent ratio E
340E380
increased negligibly when 50120583MCd was applied in resting condition but the rate of risedRdt was enhanced by gt60 times following depolarizationwith 25KCl In contrast Pb (15ndash50120583M)determined a sizeableincrease of R even in the absence of any specific stimulus thatopens VDCC and dRdt was increased by 5 times from theresting level with 75mMKCl (Figure 1(C)) Fast uptake of Pbin the absence of any specific stimulus appears to be a peculiarfeature of neurons and may be linked to the specific lipidiccomposition of the neuronal membranes while the role ofVDCC is likely to significant but not essential [23]
32 Role of VDCC in Cd-Induced Mortality As VDCC chan-nels and in particular those of L-type [15 19] play a promi-nent role in uptake of Cd by excitable cells the involvement ofthese channels in Cd toxicity was investigated further Tests ofmortality chromatin condensation and DNA fragmentationshowed that a pulsed Cd treatment induces delayed apop-totic cell death in VDCC-containing insulinoma cells andthat nimodipine protects against Cd-induced apoptosis andnecrosis in these cells supporting a specific involvement of L-type VDCC [21] The same treatments were largely harmlessin VDCC-free HeLa cell cultures in which neither death norDNA fragmentation was observed Therefore excitable cellsappear more susceptible than nonexcitable epithelial-likecells to Cd accumulation However it is not clear whether themere presence of VDCC can make cells more vulnerable totoxicmetal injury To answer this question we have comparedthe effects of Cd in two cell types that are virtually identicalexcept for the expression of VDCC in one of the two Thisapproach is made possible by the availability of a CHO cellline that has been permanently transfected with VDCC 120572
1
120573 and 1205722120575 subunits CHOC1205729120573
312057221205754cells [26 29] Promi-
nent voltage-dependent Ca currents were recorded fromthe permanently transfected cell line while such currentsare completely absent in wild-type CHO (Figure 2(a)) Thecurrents are larger with Ba as charge carrier and are sensitiveto DHP modulation (Figure 2(b)) as expected for an L-typeCa channel
Both cell types (wild-type and CHOC120572) were subjectedto a ldquopulse treatmentrdquo with 50 or 100 120583M Cd in serum-freeculture medium and observed after 18ndash24 hour after washCell suffering and mortality was negligible immediately afterthe treatment but it was evident after at least 16 hours Aspreviously observed [21 30 31] Cd accumulates into the cellduring treatment but the toxic mechanism leading to celldeath becomes effective at a later time Figure 3 shows theappearance of the cell monolayer 24 hours after an incubationof 30min with 100120583M Cd Cells exposed to Cd were ofsmaller size than control cells and had an increased tendencyto detach from their neighbors and from the substrate andfloat In addition CHOC120572VDCC-expressing cells frequentlyacquired an irregular shape with shredded edges (Figure 3)The number of viable cells was estimated from the numberof adherent trypan-blue excluding cells and this numberwas normalized to that of adherent viable cells in controlconditions as in our previous works [21] Despite some mor-phological features the dose dependence of Cd sensitivity
4 ISRN Toxicology
08
06
04
02
0
TPEN
TPEN
TPEN
2 min
25 KCl25 KCl
50 Cd 50 Pb15 Pb
75 KCl
Fluo
resc
ence
ratio
Stnd 0 Ca
lowast
lowast
lowast
100
50
0
(A) (B)
(C)
(a) (b)
(c) (d)
25 KCl75 KCl
5 15 50
119889119877119889119905(
minminus1)lowast
103
Figure 1 Uptake of Cd and Pb in cerebellar granule neurons measured by fluorescent dyes (A) Confocal microscopy images of Cd (a b) andPb (c d) uptake by cerebellar granule cells preloaded with the divalent metal-sensitive dye Oregon Green In (a) and (c) neurons were bathedin a physiological saline In (b) they have been superfused with a solution containing 0 Ca 30mM KCl and 100 120583M Cd Cl
2 which caused
the dye fluorescence to increase significantly In (d) external solution contained 0Ca and Pb 15120583M which permeates through the neuronmembrane even in the absence of a depolarizing stimulus and also caused an increase in the dye fluorescence Bar = 10 120583M (B) Real-timerecording of the influx of Cd and Pb in Fura-2 loaded cerebellar granule neuron and the effect of membrane depolarization Cells were treatedwith the metals in nominal Ca-free solution From left to right time course of the fluorescence ratio 119877 = E340E380 following application of50 120583MCd and membrane depolarization (25mM KCl) and application of 15ndash50120583M Pb in resting conditions and increasing depolarizations(25 and 75mM KCl) (C) Summary of the results obtained in basal and in depolarizing solutions (25 and 75mM KCl) with different doses ofPb The time course of R(t) was approximated by a straight line and the slope dRdt was calculated in each case as a measure of Pb influxData are mean plusmn SEM in 4 experiments and lowastindicates significantly different from basal (no depolarization) for each dose of Pb (119875 lt 005)
was very similar in the two cell types (Figure 3) Thereforeby this approach it is not possible to demonstrate a clear roleof VDCC in mediating Cd-induced cell detachment
In subsequent experiments we challenged the cells withCd in the presence of 1120583M DHP modifiers When VDCC-expressing CHOC120572 cells were exposed to Cd in the presence
ISRN Toxicology 5
(a)
Wild type
100 pA
10 ms
(b)
5 mM Ca
5 mM Ba
50 pA
10 ms
Nimo 5 Ba 5 Ca
0 60 120 180 240 300
Curr
ent (
pA)
0
minus100
minus200
(c)
Figure 2 CHOC120572 cells express L-type VDCC (a) Representative microphotographs of monolayer cell cultures of permanently transfectedCHOC120572 cells (left) and wild-type CHO cells (right) Bar = 35 120583M (b) Representative current traces evoked by depolarizing voltage stepsof 40ms duration from minus60 to +80mV from a holding potential of minus80mV in a CHOC120572 cell (left) and a wild-type CHO cell (right) Theexternal solution contained 5mMCaCl
2 This protocol evoked voltage-dependent calcium current in the permanently transfected cell while
no current was present in CHO cell of the wild-type (c) Characterization of the Ca current in CHOC120572 cellThe current was increased bymorethan 50 when the external solution was changed from 5mM CaCl
2to 5mM BaCl
2 and it was reversibly blocked by 10 120583M nimodipine
Current traces evoked by 50ms depolarizing steps from minus80mV (holding potential) to +10mV in the three conditions are shown on the leftThe graph on the right shows the time course of the experiment
of the antagonist (nimodipina) or agonist (BayK 8644) theeffect of the metal was clearly modified with respect to cellsexposed to Cd alone (Figure 4) with a sizeable increase ofmortality in the presence of BayK and protection in the pres-ence of nimodipineDHPmodifierswere largely ineffective inwild-type CHO cells exposed to Cd (not shown) Thereforeit seems that the actual contribution of L-type VDCC to Cduptake is clearly underscored only in the presence of thesemodifiers
Another indicator of apoptosis is cell shrinkage whichcan be quantified by cell capacitance measurements Bothwild-type and CHOC120572 cells showed a decrease in cell capaci-tance 18ndash24 hours afterCd treatment Inwild-type cell capac-itance was 24plusmn2 pF (119899 = 15) in control and 15plusmn 3 pF (119899 = 6)after incubation with 100 120583MCd significantly different fromcontrol with 119875 lt 005 In CHOC120572 cells cell capacitance was35 plusmn 3 pF (119899 = 17) in control and 22 plusmn 2 pF (119899 = 39)after incubation with 100 120583MCd significantly different from
control with 119875 lt 0001 In CHOC120572 cells the effect ofCd treatment was partially prevented by nimodipine thecapacitance was 35plusmn3 pF (119899 = 10) and the difference betweenthis condition and control condition was not significant(Figure 5)This indicates again that the contribution of L-typeVDCC in Cd-induced toxicity is evident only in the presenceof DHPs
Finally we performedmeasurements of total Cd accumu-lation during 60min incubation time by atomic absorptionspectroscopy For these experiments cells were harvestedimmediately after the treatment and the total amount of Cdwas normalized to the cell volume estimated from the single-cell capacitance Assuming that both cell types are approxi-mately spherical and have a specific membrane capacitanceof 1 120583Fcm2 106 wild-type CHO cells comprise a volume of11 120583L while 106 CHOC120572 cells a volume of 195120583L Thisdifference is not negligible and with this method the resultswere considerably more reproducible than using a standard
6 ISRN Toxicology
1
08
06
04
02
01 10 100 1000
Wild type+VDCC
(a) (b)
(c) (d)
(e)
Figure 3 Effect of pulse treatment with Cd in CHOC120572 VDCC-expressing and wild-type CHO cells Microphotographs of control (a b) andCd-treated (c d) cells (a c) VDCC-expressing CHOC120572 cells and (b d) wild-type CHO cells Both cell types were incubated in 100120583M Cdfor 60 minutes in the presence of 30mM KCl Pictures were taken 24 h after wash of the metal Bar = 35 120583MThe graph (e) shows the effectof a 30min pulse treatment with Cd on cell adhesion as a function of concentration Trypan-blue excluding adherent cells were counted24 h after wash of the metal Points are average plusmn sem of 3 experiments in the same condition in CHOC120572 (filled circles) and wild-type CHO(empty circles) and were best fitted to the function NadherNadher(control) = 1(1 + ([Cd]ED50)) where NadherNadher(control) is the number ofadherent cells after Cd treatment normalized to the number of adherent cells in control culture [Cd] is the concentration of Cd and ED
50is
the concentration of Cd that causes detachment from the substrate of 50 of cells The best fit yielded ED50= 40 120583M for CHOC120572 and 43120583M
for wild-type CHOThe two curves are overlapped In contrast with the different appearance the two cell types were similarly affected by Cdtreatment
normalization to the total protein content Data shown inFigure 6 indicate that the difference in Cd accumulation isbarely or not significant when both cell types are incubatedin 100120583M Cd with or without elevated KCl On the otherhand BayK 8644 significantly enhanced and nimodipinesignificantly decreased metal accumulation in depolarizedCHOC120572 cells treated with 50 or 100120583M Cd but not in wild-type CHO cells
Toxicity of both Cd and Pb is mainly due to their abilityto permeate not simply mammalian cell membranes butspecifically tight membrane barriers such as the epitheliallining in the gastrointestinal tract luminal membrane ofproximal tubules in the kidney and the blood-brain barrierIn early work the influx of Pb and Cd into mammaliancells was frequently ascribed to permeation through calciumchannels and in particular L-type DHP-sensitive VDCC [1618 20 32]More recently other types of VDCC different fromthe classical DHP-sensitive L-type have been implicated bothin Cd [19 33] and Pb [23] uptake Moreover Pb has beenshown to use different pathways in several systems [34ndash37]and it is largely independent of VDCC even in neurons[23 24]
Here I have shown that expression of L-type VDCC is notsufficient to modify Cd uptake to a toxicological significantextent at least in the case of a brief intense exposure asthose used in this study Cd accumulation and sensitivitywerecomparable in two cell lines whose only difference was thepresence of L-typeVDCC in one of the two Indeedwild-type
CHO cells which do not possess VDCC are vulnerable to Cdpoisoning and take up Cd during relatively brief exposuresto an extent sufficient to trigger cell death by apoptosisor necrosis similarly to VDCC expressing CHOC120572 cellsAlthough VDCCs play an important role in Cd uptakein CHOC120572 cells as indicated by the sizeable effect of L-type channel DHP modifiers wild-type CHO cells use othertransport proteins with similar efficiency and outcome
It is thus clear that VDCCs are not the main mecha-nism responsible for toxic metal permeation through cellmembranes and tight barriers Rapid passive transport of Pbindependent of VDCC was reported at the brain endothe-lium [34 36 37] and even protein-independent transportof lead has been described [35] In astroglia cell models theuptake of Pb was shown to be driven by store-depletion-activated channels [38] and by two distinct pH-sensitivetransport mechanisms [39] VDCC-independent uptake ofPb by cerebellar granule neurons was inhibited by La [24]and this observation can provide some clue as for the mecha-nism of permeationThe involvement of a La-sensitive store-depletion activated channel as proposed in other systems[36 37 40] is unlikely because Pb influx did not requiredrainage of the stores Another possibility is that Pb2+ entersthe neuron via reverse operation of an exchanger similar (oridentical) to theNandashCa exchanger which is also very sensitiveto La block [41] and whose role in Zn permeation wasdemonstrated in cortical neurons [42] Also this possibilitywas discarded because NandashCa exchanger does not play
ISRN Toxicology 7
4
3
2
1
0Basal Cd K N B
Treatments
lowast
lowastlowast
(a) (b)
(c) (d)
(e)
Figure 4 Effect of dihydropyridines on Cd cytotoxicity in VDCC-expressing CHOC120572 cells Cells were treated for 30min in (a) controlmediumcontaining 30mMKCl (b) 100 120583M Cd+30mM KCl (c) 100 120583M Cd+30mM KCl+1 120583Mnimodipine (d) 100 120583M Cd+30mM KCl+1 120583M BayK8644 Note the partial recovery in shape of the cells treated in the presence of nimodipine and definitive lost of adhesion whenthe treatment was performed in the presence of BayK Bar = 25 120583M The graph (e) represents counts of viable adherent cells in the sameexperiment (average plusmn sem in 3 samples) in control (basal) following for a 30min treatment with 100 120583M Cd (Cd) Cd + 30mM KCl (K)Cd + 30mM KCl and 1 120583M nimodipine (N) and Cd + 30mM KCl and 1 120583M BayK (B) lowast indicates significantly different from control with119875 lt 005 and lowastlowast with 119875 lt 00001
40
30
20
10
0Wild type
ControlCd
Capa
cita
nce (
pf)
lowast
lowastlowast
Cd + nimo
+VDCC
Figure 5 Evidence of Cd-induced apoptosis change in cell capac-itance Cells were incubated with 100120583M Cd for 60 minutes andelectrical measurements were performed 24 h after wash Mem-brane capacitance was estimated from transient compensation (seeSection 24) Bars represent mean plusmn sem Treatment with Cd causedshrinkage of all cell as revealed by reduction of the cell capacitancein both wild-type (119899 = 15 in control and 119899 = 6 with Cd treatment)and CHOC120572 cells (119899 = 17 in control and 119899 = 39with Cd treatment)Nimodipine protected CHOC120572 cells from Cd-induced shrinkage(119899 = 10) lowast indicates significantly different from control with 119875 lt005 and lowastlowast with 119875 lt 0001
a significant role when the Na concentrations are close tothe physiological value as in Esposito et al [24] It is more
probable that Pb is taken up through a La-sensitive ion carriersimilar to the Zn transporter found in brain neurons [43]whose identity and features still wait full definition
Involvement of different transport proteins in Cd trans-port has been described At the epithelial lining of the gas-trointestinal barrier DMT1 the main transporter thatabsorbs Fe in the brush border membrane of the mammalianintestine has been shown to carry also different toxic metals[44ndash46] The role of this transporter in Cd absorption isunderscored by the fact that iron deficiency creates a sig-nificant risk for increased cadmium exposure by increasinggastrointestinal absorption from 5 to as much as 20In general a low level of essential metals favors uptake ofnonessential (toxic) elements that compete for their site andthis stresses the importance of evaluating metal balance incontrast with metal concentration In addition other metal-specific transporters have been identified at the cDNA level[47] and studied in expression systems [48]
4 Conclusion
Ionicmimicry is a useful framework to study themechanismsof metal toxicity Uptake of toxic metals such as Cd and Pbby mammalian cells of different tissues occurs through manydifferent pathways of which VDCCs have frequently beenregarded as prominent Because of the abundance of VDCCexcitable cells may be more susceptible to accumulate Cdand Pb than nonexcitable cells In this chapter I have shownthat although Cd and Pb can permeate through VDCC thisis not the most relevant mechanism of passive uptake ofPb while the mere presence of VDCC is not sufficient tomake cells significantly more vulnerable to Cd injury Otherfactors including different transport systems and expressionof detoxification proteins may be more relevant in the accu-mulation process of these toxic metals
8 ISRN Toxicology
Basal Cd K B NTreatments
Wild type
80
60
40
20
0
lowast
lowast
Figure 6 Total Cd accumulation measured by FAAS in CHOC120572and wild-type CHO cells Both cell types were incubated for 1 hourin medium containing 0 50 or 100 120583M Cd and other modifiersCd determination was normalized to the cell volume (see text)Treatments were as follows control 0 Cd (basal) 60min Cd (Cd)Cd + 30mM KCl (K) Cd + 30mM KCl and 1120583M BayK (B) Cd +30mM KCl and 1120583M nimodipine (N) Depolarization (treatmentK) did not increase significantly the intracellular concentration ofCd (119875 gt 005) In CHOC120572 cells but not in wild-type CHO BayKsignificantly enhanced and nimodipine significantly reduced Cdaccumulation both with 119875 lt 0001 (lowast) with respect to the sametreatments in depolarization Bars represent mean plusmn sem in at least10 experiments in each group
Acknowledgments
The author thanks Ilaria Zanardi and Irena Baranowska-Bosiacka (PUM Szczecin Poland) for help in Cd treatmentexperiments and Elisabetta Morelli for FAAS measurementsThe author is also grateful to Franz Hofmann and NorbertKlugbauer (Institut fur Pharmakologie und ToxikologieMunich Germany) for their permission to use the perma-nently transfected CHO cells and to Maurizio Taglialatelaand Mauro Cataldi (Universita Federico II Naples Italy) forsending him the cells
References
[1] R J Castellani P IMoreira G Liu et al ldquoIron the redox-activecenter of oxidative stress in Alzheimer diseaserdquo NeurochemicalResearch vol 32 no 10 pp 1640ndash1645 2007
[2] D G Smith R Cappai and K J Barnham ldquoThe redox chem-istry of the Alzheimerrsquos disease amyloid 120573 peptiderdquo Biochimicaet Biophysica Acta vol 1768 no 8 pp 1976ndash1990 2007
[3] Y Xu L Feng P D Jeffrey Y Shi and FMMMorel ldquoStructureand metal exchange in the cadmium carbonic anhydrase ofmarine diatomsrdquo Nature vol 452 no 7183 pp 56ndash61 2008
[4] T W Clarkson ldquoMolecular and ionic mimicry of toxic metalsrdquoAnnual Review of Pharmacology and Toxicology vol 33 pp545ndash571 1993
[5] C C Bridges and R K Zalups ldquoMolecular and ionic mimicryand the transport of toxic metalsrdquo Toxicology and Applied
Pharmacology vol 204 no 3 pp 274ndash308 2005[6] H Ouyang and H J Vogel ldquoMetal ion binding to calmodulin
NMRand fluorescence studiesrdquoBioMetals vol 11 no 3 pp 213ndash222 1998
[7] M KernMWisniewski L Cabell andG Audesirk ldquoInorganiclead and calcium interact positively in activation of calmod-ulinrdquo NeuroToxicology vol 21 no 3 pp 353ndash364 2000
[8] J Markovac and GW Goldstein ldquoPicomolar concentrations oflead stimulate brain protein kinase CrdquoNature vol 334 no 6177pp 71ndash73 1988
[9] X Sun X Tian J L Tomsig and J B Suszkiw ldquoAnalysisof differential effects of Pb2+ on protein kinase C isozymesrdquoToxicology and Applied Pharmacology vol 156 no 1 pp 40ndash451999
[10] J L Tomsig and J B Suszkiw ldquoMultisite interactions betweenPb2+ and protein kinase C and its role in norepinephrine releasefrom bovine adrenal chromaffin cellsrdquo Journal of Neurochem-istry vol 64 no 6 pp 2667ndash2673 1995
[11] C M Bouton L P Frelin C E Forde H A Godwin and JPevsner ldquoSynaptotagmin I is amolecular target for leadrdquo Journalof Neurochemistry vol 76 no 6 pp 1724ndash1735 2001
[12] J Yang P T Ellinor W A Sather J F Zhang and R W TsienldquoMolecular determinants of Ca2+ selectivity and ion permeationin L-type Ca2+ channelsrdquoNature vol 366 no 6451 pp 158ndash1611993
[13] MWakamori M Strobeck T Niidome T Teramoto K Imotoand Y Mori ldquoFunctional characterization of ion permeationpathway in the N-type Ca2+ channelrdquo Journal of Neurophysiol-ogy vol 79 no 2 pp 622ndash634 1998
[14] S Peng R K Hajela and W D Atchison ldquoCharacteristics ofblock by Pb2+ of function of human neuronal L- N- and R-type Ca2+ channels transiently expressed in human embryonickidney 293 cellsrdquo Molecular Pharmacology vol 62 no 6 pp1418ndash1430 2002
[15] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium uptake in excitable mammalian cells a microspec-trofluorimetric studyrdquo European Journal of Histochemistry vol41 pp 189ndash190 1997
[16] P M Hinkle P A Kinsella and K C Osterhoudt ldquoCadmiumuptake and toxicity via voltage-sensitive calcium channelsrdquoJournal of Biological Chemistry vol 262 no 34 pp 16333ndash163371987
[17] P M Hinkle E D Shanshala and E J Nelson ldquoMeasurementsof intracellular cadmium with fluorescent dyesmdashfurther evi-dence for the role of calcium channels in cadmium uptakerdquoTheJournal of Biological Chemistry vol 267 pp 25553ndash25559 1992
[18] P M Hinkle and M E Osborne ldquoCadmium toxicity in ratpheochromocytoma cells studies on themechanism of uptakerdquoToxicology and Applied Pharmacology vol 124 no 1 pp 91ndash981994
[19] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium influx in mammalian neuronsrdquo Journal of Neuro-chemistry vol 72 no 5 pp 2154ndash2161 1999
[20] V Souza L Bucio andMCGutierrez-Ruiz ldquoCadmiumuptakeby a human hepatic cell line (WRL-68 cells)rdquo Toxicology vol120 no 3 pp 215ndash220 1997
[21] P Gavazzo E Morelli and C Marchetti ldquoSusceptibility ofinsulinoma cells to cadmiumandmodulation by L-type calciumchannelsrdquo BioMetals vol 18 no 2 pp 131ndash142 2005
[22] M Kirberger and J J Yang ldquoStructural differences betweenPb2+- and Ca2+-binding sites in proteins implications with
ISRN Toxicology 9
respect to toxicityrdquo Journal of Inorganic Biochemistry vol 102no 10 pp 1901ndash1909 2008
[23] MMazzolini S Traverso andCMarchetti ldquoMultiple pathwaysof Pb2+ permeation in rat cerebellar granule neuronesrdquo Journalof Neurochemistry vol 79 no 2 pp 407ndash416 2001
[24] A Esposito M Robello F Pellistri and C Marchetti ldquoTwo-photon analysis of lead accumulation in rat cerebellar granuleneuronsrdquo Neurochemical Research vol 30 no 8 pp 949ndash9542005
[25] C Marchetti C Amico and C Usai ldquoFunctional characteriza-tion of the effect of nimodipine on the calcium current in ratcerebellar granule cellsrdquo Journal of Neurophysiology vol 73 no3 pp 1169ndash1180 1995
[26] A Welling E Bosse A Cavalie et al ldquoStable co-expression ofcalcium channel 1205721120573 and 1205722120575 subunits in a somatic cell linerdquoJournal of Physiology vol 471 pp 749ndash765 1993
[27] A Diaspro S Annunziata M Raimondo P Ramoino and MRobello ldquoA single-pinhole confocal laser scanning microscopefor 3-D imaging of biostructuresrdquo IEEE Engineering inMedicineand Biology Magazine vol 18 no 4 pp 106ndash110 1999
[28] F Pellistri A Cupello A Esposito C Marchetti and MRobello ldquoTwo-photon imaging of calcium accumulation in ratcerebellar granule cellsrdquo NeuroReport vol 15 no 1 pp 83ndash872004
[29] M Cataldi A Secondo A DrsquoAlessio et al ldquoStudies onmaitotoxin-induced intracellular Ca2+ elevation in chinesehamster ovary cells stably transfected with cDNAs encodingfor L-type Ca2+ channel subunitsrdquo Journal of Pharmacology andExperimental Therapeutics vol 290 no 2 pp 725ndash730 1999
[30] A Galan A Troyano N E Vilaboa C Fernandez E DeBlas and P Aller ldquoModulation of the stress response duringapoptosis and necrosis induction in cadmium-treated U-937human promonocytic cellsrdquo Biochimica et Biophysica Acta vol1538 no 1 pp 38ndash46 2001
[31] A H B Poliandri J P Cabilla M O Velardez C C ABodo and B H Duvilanski ldquoCadmium induces apoptosis inanterior pituitary cells that can be reversed by treatment withantioxidantsrdquo Toxicology and Applied Pharmacology vol 190no 1 pp 17ndash24 2003
[32] J L Tomsig and J B Suszkiw ldquoPermeation of Pb2+ throughcalcium channels fura-2 measurements of voltage- anddihydropyridine-sensitive Pb2+ entry in isolated bovinechromaffin cellsrdquo Biochimica et Biophysica Acta vol 1069 no2 pp 197ndash200 1991
[33] EM Leslie J Liu C D Klaassen andM PWaalkes ldquoAcquiredcadmium resistance in metallothionein-III(minusminus) knockoutcells role of the T-type calcium channel Cacn1205721G in cadmiumuptakerdquo Molecular Pharmacology vol 69 no 2 pp 629ndash6392006
[34] R Deane and M W B Bradbury ldquoTransport of lead-203 atthe blood-brain barrier during short cerebrovascular perfusionwith saline in the ratrdquo Journal of Neurochemistry vol 54 no 3pp 905ndash914 1990
[35] R S Diaz and J Monreal ldquoProtein-independent lead perme-ation through myelin lipid liposomesrdquo Molecular Pharmacol-ogy vol 47 no 4 pp 766ndash771 1995
[36] L E Kerper and P M Hinkle ldquoCellular uptake of lead isactivated by depletion of intracellular calcium storesrdquo Journalof Biological Chemistry vol 272 no 13 pp 8346ndash8352 1997
[37] L E Kerper and P M Hinkle ldquoLead uptake in brain capillaryendothelial cells activation by calcium store depletionrdquoToxicol-ogy and Applied Pharmacology vol 146 no 1 pp 127ndash133 1997
[38] M E Legare R Barhoumi E Hebert G R Bratton R CBurghardt and E Tiffany- Castiglioni ldquoAnalysis of Pb2+ entryinto cultured astrogliardquo Toxicological Sciences vol 46 no 1 pp90ndash100 1998
[39] J H Cheong D Bannon L Olivi Y Kim and J BresslerldquoDifferent mechanismsmediate uptake of lead in a rat astroglialcell linerdquoToxicological Sciences vol 77 no 2 pp 334ndash340 2004
[40] T J Evans M R James-Kracke S B Kleiboeker and S WCasteel ldquoLead enters Rcho-1 trophoblastic cells by calciumtransport mechanisms and complexes with cytosolic calcium-binding proteinsrdquo Toxicology and Applied Pharmacology vol186 no 2 pp 77ndash89 2003
[41] G J Kaczorowski R S Slaughter V F King and M LGarcia ldquoInhibitors of sodium-calcium exchange identificationand development of probes of transport activityrdquo Biochimica etBiophysica Acta vol 988 no 2 pp 287ndash302 1989
[42] S L Sensi L M T Canzoniero S P Yu et al ldquoMeasurement ofintracellular free zinc in living cortical neurons routes of entryrdquoJournal of Neuroscience vol 17 no 24 pp 9554ndash9564 1997
[43] R A Colvin ldquoCharacterization of a plasma membrane zinctransporter in rat brainrdquo Neuroscience Letters vol 247 no 2-3pp 147ndash150 1998
[44] V Picard G Govoni N Jabado and P Gross ldquoNramp 2(DCT1DMT1) expressed at the plasma membrane transportsiron and other divalent cations into a calcein-accessible cyto-plasmic poolrdquo Journal of Biological Chemistry vol 275 no 46pp 35738ndash35745 2000
[45] M Okubo K Yamada M Hosoyamada T Shibasaki and HEndou ldquoCadmium transport by human Nramp 2 expressed inXenopus laevis oocytesrdquo Toxicology and Applied Pharmacologyvol 187 no 3 pp 162ndash167 2003
[46] A C Illing A Shawki C L Cunningham and B MackenzieldquoSubstrate profile and metal-ion selectivity of human divalentmetal-ion transporter-1rdquo The Journal of Biological Chemistryvol 287 pp 30485ndash30496 2012
[47] J P Bressler L Olivi J H Cheong Y Kim A Maerten andD Bannon ldquoMetal transporters in intestine and brain theirinvolvement in metal-associated neurotoxicitiesrdquo Human andExperimental Toxicology vol 26 no 3 pp 221ndash229 2007
[48] Z Liu H Li M Soleimani et al ldquoCd2+ versus Zn2+ uptakeby the ZIP8 HCO
3mdashdependent symporter kinetics electro-
genicity and traffickingrdquo Biochemical and Biophysical ResearchCommunications vol 365 no 4 pp 814ndash820 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
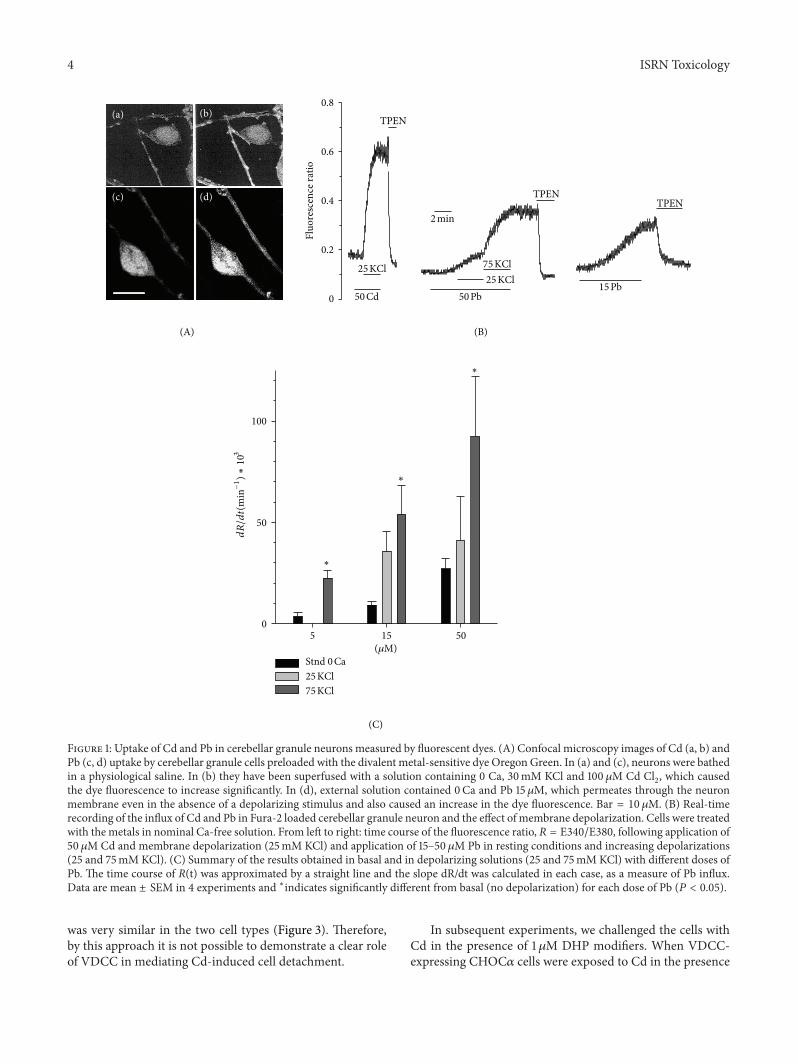
4 ISRN Toxicology
08
06
04
02
0
TPEN
TPEN
TPEN
2 min
25 KCl25 KCl
50 Cd 50 Pb15 Pb
75 KCl
Fluo
resc
ence
ratio
Stnd 0 Ca
lowast
lowast
lowast
100
50
0
(A) (B)
(C)
(a) (b)
(c) (d)
25 KCl75 KCl
5 15 50
119889119877119889119905(
minminus1)lowast
103
Figure 1 Uptake of Cd and Pb in cerebellar granule neurons measured by fluorescent dyes (A) Confocal microscopy images of Cd (a b) andPb (c d) uptake by cerebellar granule cells preloaded with the divalent metal-sensitive dye Oregon Green In (a) and (c) neurons were bathedin a physiological saline In (b) they have been superfused with a solution containing 0 Ca 30mM KCl and 100 120583M Cd Cl
2 which caused
the dye fluorescence to increase significantly In (d) external solution contained 0Ca and Pb 15120583M which permeates through the neuronmembrane even in the absence of a depolarizing stimulus and also caused an increase in the dye fluorescence Bar = 10 120583M (B) Real-timerecording of the influx of Cd and Pb in Fura-2 loaded cerebellar granule neuron and the effect of membrane depolarization Cells were treatedwith the metals in nominal Ca-free solution From left to right time course of the fluorescence ratio 119877 = E340E380 following application of50 120583MCd and membrane depolarization (25mM KCl) and application of 15ndash50120583M Pb in resting conditions and increasing depolarizations(25 and 75mM KCl) (C) Summary of the results obtained in basal and in depolarizing solutions (25 and 75mM KCl) with different doses ofPb The time course of R(t) was approximated by a straight line and the slope dRdt was calculated in each case as a measure of Pb influxData are mean plusmn SEM in 4 experiments and lowastindicates significantly different from basal (no depolarization) for each dose of Pb (119875 lt 005)
was very similar in the two cell types (Figure 3) Thereforeby this approach it is not possible to demonstrate a clear roleof VDCC in mediating Cd-induced cell detachment
In subsequent experiments we challenged the cells withCd in the presence of 1120583M DHP modifiers When VDCC-expressing CHOC120572 cells were exposed to Cd in the presence
ISRN Toxicology 5
(a)
Wild type
100 pA
10 ms
(b)
5 mM Ca
5 mM Ba
50 pA
10 ms
Nimo 5 Ba 5 Ca
0 60 120 180 240 300
Curr
ent (
pA)
0
minus100
minus200
(c)
Figure 2 CHOC120572 cells express L-type VDCC (a) Representative microphotographs of monolayer cell cultures of permanently transfectedCHOC120572 cells (left) and wild-type CHO cells (right) Bar = 35 120583M (b) Representative current traces evoked by depolarizing voltage stepsof 40ms duration from minus60 to +80mV from a holding potential of minus80mV in a CHOC120572 cell (left) and a wild-type CHO cell (right) Theexternal solution contained 5mMCaCl
2 This protocol evoked voltage-dependent calcium current in the permanently transfected cell while
no current was present in CHO cell of the wild-type (c) Characterization of the Ca current in CHOC120572 cellThe current was increased bymorethan 50 when the external solution was changed from 5mM CaCl
2to 5mM BaCl
2 and it was reversibly blocked by 10 120583M nimodipine
Current traces evoked by 50ms depolarizing steps from minus80mV (holding potential) to +10mV in the three conditions are shown on the leftThe graph on the right shows the time course of the experiment
of the antagonist (nimodipina) or agonist (BayK 8644) theeffect of the metal was clearly modified with respect to cellsexposed to Cd alone (Figure 4) with a sizeable increase ofmortality in the presence of BayK and protection in the pres-ence of nimodipineDHPmodifierswere largely ineffective inwild-type CHO cells exposed to Cd (not shown) Thereforeit seems that the actual contribution of L-type VDCC to Cduptake is clearly underscored only in the presence of thesemodifiers
Another indicator of apoptosis is cell shrinkage whichcan be quantified by cell capacitance measurements Bothwild-type and CHOC120572 cells showed a decrease in cell capaci-tance 18ndash24 hours afterCd treatment Inwild-type cell capac-itance was 24plusmn2 pF (119899 = 15) in control and 15plusmn 3 pF (119899 = 6)after incubation with 100 120583MCd significantly different fromcontrol with 119875 lt 005 In CHOC120572 cells cell capacitance was35 plusmn 3 pF (119899 = 17) in control and 22 plusmn 2 pF (119899 = 39)after incubation with 100 120583MCd significantly different from
control with 119875 lt 0001 In CHOC120572 cells the effect ofCd treatment was partially prevented by nimodipine thecapacitance was 35plusmn3 pF (119899 = 10) and the difference betweenthis condition and control condition was not significant(Figure 5)This indicates again that the contribution of L-typeVDCC in Cd-induced toxicity is evident only in the presenceof DHPs
Finally we performedmeasurements of total Cd accumu-lation during 60min incubation time by atomic absorptionspectroscopy For these experiments cells were harvestedimmediately after the treatment and the total amount of Cdwas normalized to the cell volume estimated from the single-cell capacitance Assuming that both cell types are approxi-mately spherical and have a specific membrane capacitanceof 1 120583Fcm2 106 wild-type CHO cells comprise a volume of11 120583L while 106 CHOC120572 cells a volume of 195120583L Thisdifference is not negligible and with this method the resultswere considerably more reproducible than using a standard
6 ISRN Toxicology
1
08
06
04
02
01 10 100 1000
Wild type+VDCC
(a) (b)
(c) (d)
(e)
Figure 3 Effect of pulse treatment with Cd in CHOC120572 VDCC-expressing and wild-type CHO cells Microphotographs of control (a b) andCd-treated (c d) cells (a c) VDCC-expressing CHOC120572 cells and (b d) wild-type CHO cells Both cell types were incubated in 100120583M Cdfor 60 minutes in the presence of 30mM KCl Pictures were taken 24 h after wash of the metal Bar = 35 120583MThe graph (e) shows the effectof a 30min pulse treatment with Cd on cell adhesion as a function of concentration Trypan-blue excluding adherent cells were counted24 h after wash of the metal Points are average plusmn sem of 3 experiments in the same condition in CHOC120572 (filled circles) and wild-type CHO(empty circles) and were best fitted to the function NadherNadher(control) = 1(1 + ([Cd]ED50)) where NadherNadher(control) is the number ofadherent cells after Cd treatment normalized to the number of adherent cells in control culture [Cd] is the concentration of Cd and ED
50is
the concentration of Cd that causes detachment from the substrate of 50 of cells The best fit yielded ED50= 40 120583M for CHOC120572 and 43120583M
for wild-type CHOThe two curves are overlapped In contrast with the different appearance the two cell types were similarly affected by Cdtreatment
normalization to the total protein content Data shown inFigure 6 indicate that the difference in Cd accumulation isbarely or not significant when both cell types are incubatedin 100120583M Cd with or without elevated KCl On the otherhand BayK 8644 significantly enhanced and nimodipinesignificantly decreased metal accumulation in depolarizedCHOC120572 cells treated with 50 or 100120583M Cd but not in wild-type CHO cells
Toxicity of both Cd and Pb is mainly due to their abilityto permeate not simply mammalian cell membranes butspecifically tight membrane barriers such as the epitheliallining in the gastrointestinal tract luminal membrane ofproximal tubules in the kidney and the blood-brain barrierIn early work the influx of Pb and Cd into mammaliancells was frequently ascribed to permeation through calciumchannels and in particular L-type DHP-sensitive VDCC [1618 20 32]More recently other types of VDCC different fromthe classical DHP-sensitive L-type have been implicated bothin Cd [19 33] and Pb [23] uptake Moreover Pb has beenshown to use different pathways in several systems [34ndash37]and it is largely independent of VDCC even in neurons[23 24]
Here I have shown that expression of L-type VDCC is notsufficient to modify Cd uptake to a toxicological significantextent at least in the case of a brief intense exposure asthose used in this study Cd accumulation and sensitivitywerecomparable in two cell lines whose only difference was thepresence of L-typeVDCC in one of the two Indeedwild-type
CHO cells which do not possess VDCC are vulnerable to Cdpoisoning and take up Cd during relatively brief exposuresto an extent sufficient to trigger cell death by apoptosisor necrosis similarly to VDCC expressing CHOC120572 cellsAlthough VDCCs play an important role in Cd uptakein CHOC120572 cells as indicated by the sizeable effect of L-type channel DHP modifiers wild-type CHO cells use othertransport proteins with similar efficiency and outcome
It is thus clear that VDCCs are not the main mecha-nism responsible for toxic metal permeation through cellmembranes and tight barriers Rapid passive transport of Pbindependent of VDCC was reported at the brain endothe-lium [34 36 37] and even protein-independent transportof lead has been described [35] In astroglia cell models theuptake of Pb was shown to be driven by store-depletion-activated channels [38] and by two distinct pH-sensitivetransport mechanisms [39] VDCC-independent uptake ofPb by cerebellar granule neurons was inhibited by La [24]and this observation can provide some clue as for the mecha-nism of permeationThe involvement of a La-sensitive store-depletion activated channel as proposed in other systems[36 37 40] is unlikely because Pb influx did not requiredrainage of the stores Another possibility is that Pb2+ entersthe neuron via reverse operation of an exchanger similar (oridentical) to theNandashCa exchanger which is also very sensitiveto La block [41] and whose role in Zn permeation wasdemonstrated in cortical neurons [42] Also this possibilitywas discarded because NandashCa exchanger does not play
ISRN Toxicology 7
4
3
2
1
0Basal Cd K N B
Treatments
lowast
lowastlowast
(a) (b)
(c) (d)
(e)
Figure 4 Effect of dihydropyridines on Cd cytotoxicity in VDCC-expressing CHOC120572 cells Cells were treated for 30min in (a) controlmediumcontaining 30mMKCl (b) 100 120583M Cd+30mM KCl (c) 100 120583M Cd+30mM KCl+1 120583Mnimodipine (d) 100 120583M Cd+30mM KCl+1 120583M BayK8644 Note the partial recovery in shape of the cells treated in the presence of nimodipine and definitive lost of adhesion whenthe treatment was performed in the presence of BayK Bar = 25 120583M The graph (e) represents counts of viable adherent cells in the sameexperiment (average plusmn sem in 3 samples) in control (basal) following for a 30min treatment with 100 120583M Cd (Cd) Cd + 30mM KCl (K)Cd + 30mM KCl and 1 120583M nimodipine (N) and Cd + 30mM KCl and 1 120583M BayK (B) lowast indicates significantly different from control with119875 lt 005 and lowastlowast with 119875 lt 00001
40
30
20
10
0Wild type
ControlCd
Capa
cita
nce (
pf)
lowast
lowastlowast
Cd + nimo
+VDCC
Figure 5 Evidence of Cd-induced apoptosis change in cell capac-itance Cells were incubated with 100120583M Cd for 60 minutes andelectrical measurements were performed 24 h after wash Mem-brane capacitance was estimated from transient compensation (seeSection 24) Bars represent mean plusmn sem Treatment with Cd causedshrinkage of all cell as revealed by reduction of the cell capacitancein both wild-type (119899 = 15 in control and 119899 = 6 with Cd treatment)and CHOC120572 cells (119899 = 17 in control and 119899 = 39with Cd treatment)Nimodipine protected CHOC120572 cells from Cd-induced shrinkage(119899 = 10) lowast indicates significantly different from control with 119875 lt005 and lowastlowast with 119875 lt 0001
a significant role when the Na concentrations are close tothe physiological value as in Esposito et al [24] It is more
probable that Pb is taken up through a La-sensitive ion carriersimilar to the Zn transporter found in brain neurons [43]whose identity and features still wait full definition
Involvement of different transport proteins in Cd trans-port has been described At the epithelial lining of the gas-trointestinal barrier DMT1 the main transporter thatabsorbs Fe in the brush border membrane of the mammalianintestine has been shown to carry also different toxic metals[44ndash46] The role of this transporter in Cd absorption isunderscored by the fact that iron deficiency creates a sig-nificant risk for increased cadmium exposure by increasinggastrointestinal absorption from 5 to as much as 20In general a low level of essential metals favors uptake ofnonessential (toxic) elements that compete for their site andthis stresses the importance of evaluating metal balance incontrast with metal concentration In addition other metal-specific transporters have been identified at the cDNA level[47] and studied in expression systems [48]
4 Conclusion
Ionicmimicry is a useful framework to study themechanismsof metal toxicity Uptake of toxic metals such as Cd and Pbby mammalian cells of different tissues occurs through manydifferent pathways of which VDCCs have frequently beenregarded as prominent Because of the abundance of VDCCexcitable cells may be more susceptible to accumulate Cdand Pb than nonexcitable cells In this chapter I have shownthat although Cd and Pb can permeate through VDCC thisis not the most relevant mechanism of passive uptake ofPb while the mere presence of VDCC is not sufficient tomake cells significantly more vulnerable to Cd injury Otherfactors including different transport systems and expressionof detoxification proteins may be more relevant in the accu-mulation process of these toxic metals
8 ISRN Toxicology
Basal Cd K B NTreatments
Wild type
80
60
40
20
0
lowast
lowast
Figure 6 Total Cd accumulation measured by FAAS in CHOC120572and wild-type CHO cells Both cell types were incubated for 1 hourin medium containing 0 50 or 100 120583M Cd and other modifiersCd determination was normalized to the cell volume (see text)Treatments were as follows control 0 Cd (basal) 60min Cd (Cd)Cd + 30mM KCl (K) Cd + 30mM KCl and 1120583M BayK (B) Cd +30mM KCl and 1120583M nimodipine (N) Depolarization (treatmentK) did not increase significantly the intracellular concentration ofCd (119875 gt 005) In CHOC120572 cells but not in wild-type CHO BayKsignificantly enhanced and nimodipine significantly reduced Cdaccumulation both with 119875 lt 0001 (lowast) with respect to the sametreatments in depolarization Bars represent mean plusmn sem in at least10 experiments in each group
Acknowledgments
The author thanks Ilaria Zanardi and Irena Baranowska-Bosiacka (PUM Szczecin Poland) for help in Cd treatmentexperiments and Elisabetta Morelli for FAAS measurementsThe author is also grateful to Franz Hofmann and NorbertKlugbauer (Institut fur Pharmakologie und ToxikologieMunich Germany) for their permission to use the perma-nently transfected CHO cells and to Maurizio Taglialatelaand Mauro Cataldi (Universita Federico II Naples Italy) forsending him the cells
References
[1] R J Castellani P IMoreira G Liu et al ldquoIron the redox-activecenter of oxidative stress in Alzheimer diseaserdquo NeurochemicalResearch vol 32 no 10 pp 1640ndash1645 2007
[2] D G Smith R Cappai and K J Barnham ldquoThe redox chem-istry of the Alzheimerrsquos disease amyloid 120573 peptiderdquo Biochimicaet Biophysica Acta vol 1768 no 8 pp 1976ndash1990 2007
[3] Y Xu L Feng P D Jeffrey Y Shi and FMMMorel ldquoStructureand metal exchange in the cadmium carbonic anhydrase ofmarine diatomsrdquo Nature vol 452 no 7183 pp 56ndash61 2008
[4] T W Clarkson ldquoMolecular and ionic mimicry of toxic metalsrdquoAnnual Review of Pharmacology and Toxicology vol 33 pp545ndash571 1993
[5] C C Bridges and R K Zalups ldquoMolecular and ionic mimicryand the transport of toxic metalsrdquo Toxicology and Applied
Pharmacology vol 204 no 3 pp 274ndash308 2005[6] H Ouyang and H J Vogel ldquoMetal ion binding to calmodulin
NMRand fluorescence studiesrdquoBioMetals vol 11 no 3 pp 213ndash222 1998
[7] M KernMWisniewski L Cabell andG Audesirk ldquoInorganiclead and calcium interact positively in activation of calmod-ulinrdquo NeuroToxicology vol 21 no 3 pp 353ndash364 2000
[8] J Markovac and GW Goldstein ldquoPicomolar concentrations oflead stimulate brain protein kinase CrdquoNature vol 334 no 6177pp 71ndash73 1988
[9] X Sun X Tian J L Tomsig and J B Suszkiw ldquoAnalysisof differential effects of Pb2+ on protein kinase C isozymesrdquoToxicology and Applied Pharmacology vol 156 no 1 pp 40ndash451999
[10] J L Tomsig and J B Suszkiw ldquoMultisite interactions betweenPb2+ and protein kinase C and its role in norepinephrine releasefrom bovine adrenal chromaffin cellsrdquo Journal of Neurochem-istry vol 64 no 6 pp 2667ndash2673 1995
[11] C M Bouton L P Frelin C E Forde H A Godwin and JPevsner ldquoSynaptotagmin I is amolecular target for leadrdquo Journalof Neurochemistry vol 76 no 6 pp 1724ndash1735 2001
[12] J Yang P T Ellinor W A Sather J F Zhang and R W TsienldquoMolecular determinants of Ca2+ selectivity and ion permeationin L-type Ca2+ channelsrdquoNature vol 366 no 6451 pp 158ndash1611993
[13] MWakamori M Strobeck T Niidome T Teramoto K Imotoand Y Mori ldquoFunctional characterization of ion permeationpathway in the N-type Ca2+ channelrdquo Journal of Neurophysiol-ogy vol 79 no 2 pp 622ndash634 1998
[14] S Peng R K Hajela and W D Atchison ldquoCharacteristics ofblock by Pb2+ of function of human neuronal L- N- and R-type Ca2+ channels transiently expressed in human embryonickidney 293 cellsrdquo Molecular Pharmacology vol 62 no 6 pp1418ndash1430 2002
[15] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium uptake in excitable mammalian cells a microspec-trofluorimetric studyrdquo European Journal of Histochemistry vol41 pp 189ndash190 1997
[16] P M Hinkle P A Kinsella and K C Osterhoudt ldquoCadmiumuptake and toxicity via voltage-sensitive calcium channelsrdquoJournal of Biological Chemistry vol 262 no 34 pp 16333ndash163371987
[17] P M Hinkle E D Shanshala and E J Nelson ldquoMeasurementsof intracellular cadmium with fluorescent dyesmdashfurther evi-dence for the role of calcium channels in cadmium uptakerdquoTheJournal of Biological Chemistry vol 267 pp 25553ndash25559 1992
[18] P M Hinkle and M E Osborne ldquoCadmium toxicity in ratpheochromocytoma cells studies on themechanism of uptakerdquoToxicology and Applied Pharmacology vol 124 no 1 pp 91ndash981994
[19] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium influx in mammalian neuronsrdquo Journal of Neuro-chemistry vol 72 no 5 pp 2154ndash2161 1999
[20] V Souza L Bucio andMCGutierrez-Ruiz ldquoCadmiumuptakeby a human hepatic cell line (WRL-68 cells)rdquo Toxicology vol120 no 3 pp 215ndash220 1997
[21] P Gavazzo E Morelli and C Marchetti ldquoSusceptibility ofinsulinoma cells to cadmiumandmodulation by L-type calciumchannelsrdquo BioMetals vol 18 no 2 pp 131ndash142 2005
[22] M Kirberger and J J Yang ldquoStructural differences betweenPb2+- and Ca2+-binding sites in proteins implications with
ISRN Toxicology 9
respect to toxicityrdquo Journal of Inorganic Biochemistry vol 102no 10 pp 1901ndash1909 2008
[23] MMazzolini S Traverso andCMarchetti ldquoMultiple pathwaysof Pb2+ permeation in rat cerebellar granule neuronesrdquo Journalof Neurochemistry vol 79 no 2 pp 407ndash416 2001
[24] A Esposito M Robello F Pellistri and C Marchetti ldquoTwo-photon analysis of lead accumulation in rat cerebellar granuleneuronsrdquo Neurochemical Research vol 30 no 8 pp 949ndash9542005
[25] C Marchetti C Amico and C Usai ldquoFunctional characteriza-tion of the effect of nimodipine on the calcium current in ratcerebellar granule cellsrdquo Journal of Neurophysiology vol 73 no3 pp 1169ndash1180 1995
[26] A Welling E Bosse A Cavalie et al ldquoStable co-expression ofcalcium channel 1205721120573 and 1205722120575 subunits in a somatic cell linerdquoJournal of Physiology vol 471 pp 749ndash765 1993
[27] A Diaspro S Annunziata M Raimondo P Ramoino and MRobello ldquoA single-pinhole confocal laser scanning microscopefor 3-D imaging of biostructuresrdquo IEEE Engineering inMedicineand Biology Magazine vol 18 no 4 pp 106ndash110 1999
[28] F Pellistri A Cupello A Esposito C Marchetti and MRobello ldquoTwo-photon imaging of calcium accumulation in ratcerebellar granule cellsrdquo NeuroReport vol 15 no 1 pp 83ndash872004
[29] M Cataldi A Secondo A DrsquoAlessio et al ldquoStudies onmaitotoxin-induced intracellular Ca2+ elevation in chinesehamster ovary cells stably transfected with cDNAs encodingfor L-type Ca2+ channel subunitsrdquo Journal of Pharmacology andExperimental Therapeutics vol 290 no 2 pp 725ndash730 1999
[30] A Galan A Troyano N E Vilaboa C Fernandez E DeBlas and P Aller ldquoModulation of the stress response duringapoptosis and necrosis induction in cadmium-treated U-937human promonocytic cellsrdquo Biochimica et Biophysica Acta vol1538 no 1 pp 38ndash46 2001
[31] A H B Poliandri J P Cabilla M O Velardez C C ABodo and B H Duvilanski ldquoCadmium induces apoptosis inanterior pituitary cells that can be reversed by treatment withantioxidantsrdquo Toxicology and Applied Pharmacology vol 190no 1 pp 17ndash24 2003
[32] J L Tomsig and J B Suszkiw ldquoPermeation of Pb2+ throughcalcium channels fura-2 measurements of voltage- anddihydropyridine-sensitive Pb2+ entry in isolated bovinechromaffin cellsrdquo Biochimica et Biophysica Acta vol 1069 no2 pp 197ndash200 1991
[33] EM Leslie J Liu C D Klaassen andM PWaalkes ldquoAcquiredcadmium resistance in metallothionein-III(minusminus) knockoutcells role of the T-type calcium channel Cacn1205721G in cadmiumuptakerdquo Molecular Pharmacology vol 69 no 2 pp 629ndash6392006
[34] R Deane and M W B Bradbury ldquoTransport of lead-203 atthe blood-brain barrier during short cerebrovascular perfusionwith saline in the ratrdquo Journal of Neurochemistry vol 54 no 3pp 905ndash914 1990
[35] R S Diaz and J Monreal ldquoProtein-independent lead perme-ation through myelin lipid liposomesrdquo Molecular Pharmacol-ogy vol 47 no 4 pp 766ndash771 1995
[36] L E Kerper and P M Hinkle ldquoCellular uptake of lead isactivated by depletion of intracellular calcium storesrdquo Journalof Biological Chemistry vol 272 no 13 pp 8346ndash8352 1997
[37] L E Kerper and P M Hinkle ldquoLead uptake in brain capillaryendothelial cells activation by calcium store depletionrdquoToxicol-ogy and Applied Pharmacology vol 146 no 1 pp 127ndash133 1997
[38] M E Legare R Barhoumi E Hebert G R Bratton R CBurghardt and E Tiffany- Castiglioni ldquoAnalysis of Pb2+ entryinto cultured astrogliardquo Toxicological Sciences vol 46 no 1 pp90ndash100 1998
[39] J H Cheong D Bannon L Olivi Y Kim and J BresslerldquoDifferent mechanismsmediate uptake of lead in a rat astroglialcell linerdquoToxicological Sciences vol 77 no 2 pp 334ndash340 2004
[40] T J Evans M R James-Kracke S B Kleiboeker and S WCasteel ldquoLead enters Rcho-1 trophoblastic cells by calciumtransport mechanisms and complexes with cytosolic calcium-binding proteinsrdquo Toxicology and Applied Pharmacology vol186 no 2 pp 77ndash89 2003
[41] G J Kaczorowski R S Slaughter V F King and M LGarcia ldquoInhibitors of sodium-calcium exchange identificationand development of probes of transport activityrdquo Biochimica etBiophysica Acta vol 988 no 2 pp 287ndash302 1989
[42] S L Sensi L M T Canzoniero S P Yu et al ldquoMeasurement ofintracellular free zinc in living cortical neurons routes of entryrdquoJournal of Neuroscience vol 17 no 24 pp 9554ndash9564 1997
[43] R A Colvin ldquoCharacterization of a plasma membrane zinctransporter in rat brainrdquo Neuroscience Letters vol 247 no 2-3pp 147ndash150 1998
[44] V Picard G Govoni N Jabado and P Gross ldquoNramp 2(DCT1DMT1) expressed at the plasma membrane transportsiron and other divalent cations into a calcein-accessible cyto-plasmic poolrdquo Journal of Biological Chemistry vol 275 no 46pp 35738ndash35745 2000
[45] M Okubo K Yamada M Hosoyamada T Shibasaki and HEndou ldquoCadmium transport by human Nramp 2 expressed inXenopus laevis oocytesrdquo Toxicology and Applied Pharmacologyvol 187 no 3 pp 162ndash167 2003
[46] A C Illing A Shawki C L Cunningham and B MackenzieldquoSubstrate profile and metal-ion selectivity of human divalentmetal-ion transporter-1rdquo The Journal of Biological Chemistryvol 287 pp 30485ndash30496 2012
[47] J P Bressler L Olivi J H Cheong Y Kim A Maerten andD Bannon ldquoMetal transporters in intestine and brain theirinvolvement in metal-associated neurotoxicitiesrdquo Human andExperimental Toxicology vol 26 no 3 pp 221ndash229 2007
[48] Z Liu H Li M Soleimani et al ldquoCd2+ versus Zn2+ uptakeby the ZIP8 HCO
3mdashdependent symporter kinetics electro-
genicity and traffickingrdquo Biochemical and Biophysical ResearchCommunications vol 365 no 4 pp 814ndash820 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

ISRN Toxicology 5
(a)
Wild type
100 pA
10 ms
(b)
5 mM Ca
5 mM Ba
50 pA
10 ms
Nimo 5 Ba 5 Ca
0 60 120 180 240 300
Curr
ent (
pA)
0
minus100
minus200
(c)
Figure 2 CHOC120572 cells express L-type VDCC (a) Representative microphotographs of monolayer cell cultures of permanently transfectedCHOC120572 cells (left) and wild-type CHO cells (right) Bar = 35 120583M (b) Representative current traces evoked by depolarizing voltage stepsof 40ms duration from minus60 to +80mV from a holding potential of minus80mV in a CHOC120572 cell (left) and a wild-type CHO cell (right) Theexternal solution contained 5mMCaCl
2 This protocol evoked voltage-dependent calcium current in the permanently transfected cell while
no current was present in CHO cell of the wild-type (c) Characterization of the Ca current in CHOC120572 cellThe current was increased bymorethan 50 when the external solution was changed from 5mM CaCl
2to 5mM BaCl
2 and it was reversibly blocked by 10 120583M nimodipine
Current traces evoked by 50ms depolarizing steps from minus80mV (holding potential) to +10mV in the three conditions are shown on the leftThe graph on the right shows the time course of the experiment
of the antagonist (nimodipina) or agonist (BayK 8644) theeffect of the metal was clearly modified with respect to cellsexposed to Cd alone (Figure 4) with a sizeable increase ofmortality in the presence of BayK and protection in the pres-ence of nimodipineDHPmodifierswere largely ineffective inwild-type CHO cells exposed to Cd (not shown) Thereforeit seems that the actual contribution of L-type VDCC to Cduptake is clearly underscored only in the presence of thesemodifiers
Another indicator of apoptosis is cell shrinkage whichcan be quantified by cell capacitance measurements Bothwild-type and CHOC120572 cells showed a decrease in cell capaci-tance 18ndash24 hours afterCd treatment Inwild-type cell capac-itance was 24plusmn2 pF (119899 = 15) in control and 15plusmn 3 pF (119899 = 6)after incubation with 100 120583MCd significantly different fromcontrol with 119875 lt 005 In CHOC120572 cells cell capacitance was35 plusmn 3 pF (119899 = 17) in control and 22 plusmn 2 pF (119899 = 39)after incubation with 100 120583MCd significantly different from
control with 119875 lt 0001 In CHOC120572 cells the effect ofCd treatment was partially prevented by nimodipine thecapacitance was 35plusmn3 pF (119899 = 10) and the difference betweenthis condition and control condition was not significant(Figure 5)This indicates again that the contribution of L-typeVDCC in Cd-induced toxicity is evident only in the presenceof DHPs
Finally we performedmeasurements of total Cd accumu-lation during 60min incubation time by atomic absorptionspectroscopy For these experiments cells were harvestedimmediately after the treatment and the total amount of Cdwas normalized to the cell volume estimated from the single-cell capacitance Assuming that both cell types are approxi-mately spherical and have a specific membrane capacitanceof 1 120583Fcm2 106 wild-type CHO cells comprise a volume of11 120583L while 106 CHOC120572 cells a volume of 195120583L Thisdifference is not negligible and with this method the resultswere considerably more reproducible than using a standard
6 ISRN Toxicology
1
08
06
04
02
01 10 100 1000
Wild type+VDCC
(a) (b)
(c) (d)
(e)
Figure 3 Effect of pulse treatment with Cd in CHOC120572 VDCC-expressing and wild-type CHO cells Microphotographs of control (a b) andCd-treated (c d) cells (a c) VDCC-expressing CHOC120572 cells and (b d) wild-type CHO cells Both cell types were incubated in 100120583M Cdfor 60 minutes in the presence of 30mM KCl Pictures were taken 24 h after wash of the metal Bar = 35 120583MThe graph (e) shows the effectof a 30min pulse treatment with Cd on cell adhesion as a function of concentration Trypan-blue excluding adherent cells were counted24 h after wash of the metal Points are average plusmn sem of 3 experiments in the same condition in CHOC120572 (filled circles) and wild-type CHO(empty circles) and were best fitted to the function NadherNadher(control) = 1(1 + ([Cd]ED50)) where NadherNadher(control) is the number ofadherent cells after Cd treatment normalized to the number of adherent cells in control culture [Cd] is the concentration of Cd and ED
50is
the concentration of Cd that causes detachment from the substrate of 50 of cells The best fit yielded ED50= 40 120583M for CHOC120572 and 43120583M
for wild-type CHOThe two curves are overlapped In contrast with the different appearance the two cell types were similarly affected by Cdtreatment
normalization to the total protein content Data shown inFigure 6 indicate that the difference in Cd accumulation isbarely or not significant when both cell types are incubatedin 100120583M Cd with or without elevated KCl On the otherhand BayK 8644 significantly enhanced and nimodipinesignificantly decreased metal accumulation in depolarizedCHOC120572 cells treated with 50 or 100120583M Cd but not in wild-type CHO cells
Toxicity of both Cd and Pb is mainly due to their abilityto permeate not simply mammalian cell membranes butspecifically tight membrane barriers such as the epitheliallining in the gastrointestinal tract luminal membrane ofproximal tubules in the kidney and the blood-brain barrierIn early work the influx of Pb and Cd into mammaliancells was frequently ascribed to permeation through calciumchannels and in particular L-type DHP-sensitive VDCC [1618 20 32]More recently other types of VDCC different fromthe classical DHP-sensitive L-type have been implicated bothin Cd [19 33] and Pb [23] uptake Moreover Pb has beenshown to use different pathways in several systems [34ndash37]and it is largely independent of VDCC even in neurons[23 24]
Here I have shown that expression of L-type VDCC is notsufficient to modify Cd uptake to a toxicological significantextent at least in the case of a brief intense exposure asthose used in this study Cd accumulation and sensitivitywerecomparable in two cell lines whose only difference was thepresence of L-typeVDCC in one of the two Indeedwild-type
CHO cells which do not possess VDCC are vulnerable to Cdpoisoning and take up Cd during relatively brief exposuresto an extent sufficient to trigger cell death by apoptosisor necrosis similarly to VDCC expressing CHOC120572 cellsAlthough VDCCs play an important role in Cd uptakein CHOC120572 cells as indicated by the sizeable effect of L-type channel DHP modifiers wild-type CHO cells use othertransport proteins with similar efficiency and outcome
It is thus clear that VDCCs are not the main mecha-nism responsible for toxic metal permeation through cellmembranes and tight barriers Rapid passive transport of Pbindependent of VDCC was reported at the brain endothe-lium [34 36 37] and even protein-independent transportof lead has been described [35] In astroglia cell models theuptake of Pb was shown to be driven by store-depletion-activated channels [38] and by two distinct pH-sensitivetransport mechanisms [39] VDCC-independent uptake ofPb by cerebellar granule neurons was inhibited by La [24]and this observation can provide some clue as for the mecha-nism of permeationThe involvement of a La-sensitive store-depletion activated channel as proposed in other systems[36 37 40] is unlikely because Pb influx did not requiredrainage of the stores Another possibility is that Pb2+ entersthe neuron via reverse operation of an exchanger similar (oridentical) to theNandashCa exchanger which is also very sensitiveto La block [41] and whose role in Zn permeation wasdemonstrated in cortical neurons [42] Also this possibilitywas discarded because NandashCa exchanger does not play
ISRN Toxicology 7
4
3
2
1
0Basal Cd K N B
Treatments
lowast
lowastlowast
(a) (b)
(c) (d)
(e)
Figure 4 Effect of dihydropyridines on Cd cytotoxicity in VDCC-expressing CHOC120572 cells Cells were treated for 30min in (a) controlmediumcontaining 30mMKCl (b) 100 120583M Cd+30mM KCl (c) 100 120583M Cd+30mM KCl+1 120583Mnimodipine (d) 100 120583M Cd+30mM KCl+1 120583M BayK8644 Note the partial recovery in shape of the cells treated in the presence of nimodipine and definitive lost of adhesion whenthe treatment was performed in the presence of BayK Bar = 25 120583M The graph (e) represents counts of viable adherent cells in the sameexperiment (average plusmn sem in 3 samples) in control (basal) following for a 30min treatment with 100 120583M Cd (Cd) Cd + 30mM KCl (K)Cd + 30mM KCl and 1 120583M nimodipine (N) and Cd + 30mM KCl and 1 120583M BayK (B) lowast indicates significantly different from control with119875 lt 005 and lowastlowast with 119875 lt 00001
40
30
20
10
0Wild type
ControlCd
Capa
cita
nce (
pf)
lowast
lowastlowast
Cd + nimo
+VDCC
Figure 5 Evidence of Cd-induced apoptosis change in cell capac-itance Cells were incubated with 100120583M Cd for 60 minutes andelectrical measurements were performed 24 h after wash Mem-brane capacitance was estimated from transient compensation (seeSection 24) Bars represent mean plusmn sem Treatment with Cd causedshrinkage of all cell as revealed by reduction of the cell capacitancein both wild-type (119899 = 15 in control and 119899 = 6 with Cd treatment)and CHOC120572 cells (119899 = 17 in control and 119899 = 39with Cd treatment)Nimodipine protected CHOC120572 cells from Cd-induced shrinkage(119899 = 10) lowast indicates significantly different from control with 119875 lt005 and lowastlowast with 119875 lt 0001
a significant role when the Na concentrations are close tothe physiological value as in Esposito et al [24] It is more
probable that Pb is taken up through a La-sensitive ion carriersimilar to the Zn transporter found in brain neurons [43]whose identity and features still wait full definition
Involvement of different transport proteins in Cd trans-port has been described At the epithelial lining of the gas-trointestinal barrier DMT1 the main transporter thatabsorbs Fe in the brush border membrane of the mammalianintestine has been shown to carry also different toxic metals[44ndash46] The role of this transporter in Cd absorption isunderscored by the fact that iron deficiency creates a sig-nificant risk for increased cadmium exposure by increasinggastrointestinal absorption from 5 to as much as 20In general a low level of essential metals favors uptake ofnonessential (toxic) elements that compete for their site andthis stresses the importance of evaluating metal balance incontrast with metal concentration In addition other metal-specific transporters have been identified at the cDNA level[47] and studied in expression systems [48]
4 Conclusion
Ionicmimicry is a useful framework to study themechanismsof metal toxicity Uptake of toxic metals such as Cd and Pbby mammalian cells of different tissues occurs through manydifferent pathways of which VDCCs have frequently beenregarded as prominent Because of the abundance of VDCCexcitable cells may be more susceptible to accumulate Cdand Pb than nonexcitable cells In this chapter I have shownthat although Cd and Pb can permeate through VDCC thisis not the most relevant mechanism of passive uptake ofPb while the mere presence of VDCC is not sufficient tomake cells significantly more vulnerable to Cd injury Otherfactors including different transport systems and expressionof detoxification proteins may be more relevant in the accu-mulation process of these toxic metals
8 ISRN Toxicology
Basal Cd K B NTreatments
Wild type
80
60
40
20
0
lowast
lowast
Figure 6 Total Cd accumulation measured by FAAS in CHOC120572and wild-type CHO cells Both cell types were incubated for 1 hourin medium containing 0 50 or 100 120583M Cd and other modifiersCd determination was normalized to the cell volume (see text)Treatments were as follows control 0 Cd (basal) 60min Cd (Cd)Cd + 30mM KCl (K) Cd + 30mM KCl and 1120583M BayK (B) Cd +30mM KCl and 1120583M nimodipine (N) Depolarization (treatmentK) did not increase significantly the intracellular concentration ofCd (119875 gt 005) In CHOC120572 cells but not in wild-type CHO BayKsignificantly enhanced and nimodipine significantly reduced Cdaccumulation both with 119875 lt 0001 (lowast) with respect to the sametreatments in depolarization Bars represent mean plusmn sem in at least10 experiments in each group
Acknowledgments
The author thanks Ilaria Zanardi and Irena Baranowska-Bosiacka (PUM Szczecin Poland) for help in Cd treatmentexperiments and Elisabetta Morelli for FAAS measurementsThe author is also grateful to Franz Hofmann and NorbertKlugbauer (Institut fur Pharmakologie und ToxikologieMunich Germany) for their permission to use the perma-nently transfected CHO cells and to Maurizio Taglialatelaand Mauro Cataldi (Universita Federico II Naples Italy) forsending him the cells
References
[1] R J Castellani P IMoreira G Liu et al ldquoIron the redox-activecenter of oxidative stress in Alzheimer diseaserdquo NeurochemicalResearch vol 32 no 10 pp 1640ndash1645 2007
[2] D G Smith R Cappai and K J Barnham ldquoThe redox chem-istry of the Alzheimerrsquos disease amyloid 120573 peptiderdquo Biochimicaet Biophysica Acta vol 1768 no 8 pp 1976ndash1990 2007
[3] Y Xu L Feng P D Jeffrey Y Shi and FMMMorel ldquoStructureand metal exchange in the cadmium carbonic anhydrase ofmarine diatomsrdquo Nature vol 452 no 7183 pp 56ndash61 2008
[4] T W Clarkson ldquoMolecular and ionic mimicry of toxic metalsrdquoAnnual Review of Pharmacology and Toxicology vol 33 pp545ndash571 1993
[5] C C Bridges and R K Zalups ldquoMolecular and ionic mimicryand the transport of toxic metalsrdquo Toxicology and Applied
Pharmacology vol 204 no 3 pp 274ndash308 2005[6] H Ouyang and H J Vogel ldquoMetal ion binding to calmodulin
NMRand fluorescence studiesrdquoBioMetals vol 11 no 3 pp 213ndash222 1998
[7] M KernMWisniewski L Cabell andG Audesirk ldquoInorganiclead and calcium interact positively in activation of calmod-ulinrdquo NeuroToxicology vol 21 no 3 pp 353ndash364 2000
[8] J Markovac and GW Goldstein ldquoPicomolar concentrations oflead stimulate brain protein kinase CrdquoNature vol 334 no 6177pp 71ndash73 1988
[9] X Sun X Tian J L Tomsig and J B Suszkiw ldquoAnalysisof differential effects of Pb2+ on protein kinase C isozymesrdquoToxicology and Applied Pharmacology vol 156 no 1 pp 40ndash451999
[10] J L Tomsig and J B Suszkiw ldquoMultisite interactions betweenPb2+ and protein kinase C and its role in norepinephrine releasefrom bovine adrenal chromaffin cellsrdquo Journal of Neurochem-istry vol 64 no 6 pp 2667ndash2673 1995
[11] C M Bouton L P Frelin C E Forde H A Godwin and JPevsner ldquoSynaptotagmin I is amolecular target for leadrdquo Journalof Neurochemistry vol 76 no 6 pp 1724ndash1735 2001
[12] J Yang P T Ellinor W A Sather J F Zhang and R W TsienldquoMolecular determinants of Ca2+ selectivity and ion permeationin L-type Ca2+ channelsrdquoNature vol 366 no 6451 pp 158ndash1611993
[13] MWakamori M Strobeck T Niidome T Teramoto K Imotoand Y Mori ldquoFunctional characterization of ion permeationpathway in the N-type Ca2+ channelrdquo Journal of Neurophysiol-ogy vol 79 no 2 pp 622ndash634 1998
[14] S Peng R K Hajela and W D Atchison ldquoCharacteristics ofblock by Pb2+ of function of human neuronal L- N- and R-type Ca2+ channels transiently expressed in human embryonickidney 293 cellsrdquo Molecular Pharmacology vol 62 no 6 pp1418ndash1430 2002
[15] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium uptake in excitable mammalian cells a microspec-trofluorimetric studyrdquo European Journal of Histochemistry vol41 pp 189ndash190 1997
[16] P M Hinkle P A Kinsella and K C Osterhoudt ldquoCadmiumuptake and toxicity via voltage-sensitive calcium channelsrdquoJournal of Biological Chemistry vol 262 no 34 pp 16333ndash163371987
[17] P M Hinkle E D Shanshala and E J Nelson ldquoMeasurementsof intracellular cadmium with fluorescent dyesmdashfurther evi-dence for the role of calcium channels in cadmium uptakerdquoTheJournal of Biological Chemistry vol 267 pp 25553ndash25559 1992
[18] P M Hinkle and M E Osborne ldquoCadmium toxicity in ratpheochromocytoma cells studies on themechanism of uptakerdquoToxicology and Applied Pharmacology vol 124 no 1 pp 91ndash981994
[19] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium influx in mammalian neuronsrdquo Journal of Neuro-chemistry vol 72 no 5 pp 2154ndash2161 1999
[20] V Souza L Bucio andMCGutierrez-Ruiz ldquoCadmiumuptakeby a human hepatic cell line (WRL-68 cells)rdquo Toxicology vol120 no 3 pp 215ndash220 1997
[21] P Gavazzo E Morelli and C Marchetti ldquoSusceptibility ofinsulinoma cells to cadmiumandmodulation by L-type calciumchannelsrdquo BioMetals vol 18 no 2 pp 131ndash142 2005
[22] M Kirberger and J J Yang ldquoStructural differences betweenPb2+- and Ca2+-binding sites in proteins implications with
ISRN Toxicology 9
respect to toxicityrdquo Journal of Inorganic Biochemistry vol 102no 10 pp 1901ndash1909 2008
[23] MMazzolini S Traverso andCMarchetti ldquoMultiple pathwaysof Pb2+ permeation in rat cerebellar granule neuronesrdquo Journalof Neurochemistry vol 79 no 2 pp 407ndash416 2001
[24] A Esposito M Robello F Pellistri and C Marchetti ldquoTwo-photon analysis of lead accumulation in rat cerebellar granuleneuronsrdquo Neurochemical Research vol 30 no 8 pp 949ndash9542005
[25] C Marchetti C Amico and C Usai ldquoFunctional characteriza-tion of the effect of nimodipine on the calcium current in ratcerebellar granule cellsrdquo Journal of Neurophysiology vol 73 no3 pp 1169ndash1180 1995
[26] A Welling E Bosse A Cavalie et al ldquoStable co-expression ofcalcium channel 1205721120573 and 1205722120575 subunits in a somatic cell linerdquoJournal of Physiology vol 471 pp 749ndash765 1993
[27] A Diaspro S Annunziata M Raimondo P Ramoino and MRobello ldquoA single-pinhole confocal laser scanning microscopefor 3-D imaging of biostructuresrdquo IEEE Engineering inMedicineand Biology Magazine vol 18 no 4 pp 106ndash110 1999
[28] F Pellistri A Cupello A Esposito C Marchetti and MRobello ldquoTwo-photon imaging of calcium accumulation in ratcerebellar granule cellsrdquo NeuroReport vol 15 no 1 pp 83ndash872004
[29] M Cataldi A Secondo A DrsquoAlessio et al ldquoStudies onmaitotoxin-induced intracellular Ca2+ elevation in chinesehamster ovary cells stably transfected with cDNAs encodingfor L-type Ca2+ channel subunitsrdquo Journal of Pharmacology andExperimental Therapeutics vol 290 no 2 pp 725ndash730 1999
[30] A Galan A Troyano N E Vilaboa C Fernandez E DeBlas and P Aller ldquoModulation of the stress response duringapoptosis and necrosis induction in cadmium-treated U-937human promonocytic cellsrdquo Biochimica et Biophysica Acta vol1538 no 1 pp 38ndash46 2001
[31] A H B Poliandri J P Cabilla M O Velardez C C ABodo and B H Duvilanski ldquoCadmium induces apoptosis inanterior pituitary cells that can be reversed by treatment withantioxidantsrdquo Toxicology and Applied Pharmacology vol 190no 1 pp 17ndash24 2003
[32] J L Tomsig and J B Suszkiw ldquoPermeation of Pb2+ throughcalcium channels fura-2 measurements of voltage- anddihydropyridine-sensitive Pb2+ entry in isolated bovinechromaffin cellsrdquo Biochimica et Biophysica Acta vol 1069 no2 pp 197ndash200 1991
[33] EM Leslie J Liu C D Klaassen andM PWaalkes ldquoAcquiredcadmium resistance in metallothionein-III(minusminus) knockoutcells role of the T-type calcium channel Cacn1205721G in cadmiumuptakerdquo Molecular Pharmacology vol 69 no 2 pp 629ndash6392006
[34] R Deane and M W B Bradbury ldquoTransport of lead-203 atthe blood-brain barrier during short cerebrovascular perfusionwith saline in the ratrdquo Journal of Neurochemistry vol 54 no 3pp 905ndash914 1990
[35] R S Diaz and J Monreal ldquoProtein-independent lead perme-ation through myelin lipid liposomesrdquo Molecular Pharmacol-ogy vol 47 no 4 pp 766ndash771 1995
[36] L E Kerper and P M Hinkle ldquoCellular uptake of lead isactivated by depletion of intracellular calcium storesrdquo Journalof Biological Chemistry vol 272 no 13 pp 8346ndash8352 1997
[37] L E Kerper and P M Hinkle ldquoLead uptake in brain capillaryendothelial cells activation by calcium store depletionrdquoToxicol-ogy and Applied Pharmacology vol 146 no 1 pp 127ndash133 1997
[38] M E Legare R Barhoumi E Hebert G R Bratton R CBurghardt and E Tiffany- Castiglioni ldquoAnalysis of Pb2+ entryinto cultured astrogliardquo Toxicological Sciences vol 46 no 1 pp90ndash100 1998
[39] J H Cheong D Bannon L Olivi Y Kim and J BresslerldquoDifferent mechanismsmediate uptake of lead in a rat astroglialcell linerdquoToxicological Sciences vol 77 no 2 pp 334ndash340 2004
[40] T J Evans M R James-Kracke S B Kleiboeker and S WCasteel ldquoLead enters Rcho-1 trophoblastic cells by calciumtransport mechanisms and complexes with cytosolic calcium-binding proteinsrdquo Toxicology and Applied Pharmacology vol186 no 2 pp 77ndash89 2003
[41] G J Kaczorowski R S Slaughter V F King and M LGarcia ldquoInhibitors of sodium-calcium exchange identificationand development of probes of transport activityrdquo Biochimica etBiophysica Acta vol 988 no 2 pp 287ndash302 1989
[42] S L Sensi L M T Canzoniero S P Yu et al ldquoMeasurement ofintracellular free zinc in living cortical neurons routes of entryrdquoJournal of Neuroscience vol 17 no 24 pp 9554ndash9564 1997
[43] R A Colvin ldquoCharacterization of a plasma membrane zinctransporter in rat brainrdquo Neuroscience Letters vol 247 no 2-3pp 147ndash150 1998
[44] V Picard G Govoni N Jabado and P Gross ldquoNramp 2(DCT1DMT1) expressed at the plasma membrane transportsiron and other divalent cations into a calcein-accessible cyto-plasmic poolrdquo Journal of Biological Chemistry vol 275 no 46pp 35738ndash35745 2000
[45] M Okubo K Yamada M Hosoyamada T Shibasaki and HEndou ldquoCadmium transport by human Nramp 2 expressed inXenopus laevis oocytesrdquo Toxicology and Applied Pharmacologyvol 187 no 3 pp 162ndash167 2003
[46] A C Illing A Shawki C L Cunningham and B MackenzieldquoSubstrate profile and metal-ion selectivity of human divalentmetal-ion transporter-1rdquo The Journal of Biological Chemistryvol 287 pp 30485ndash30496 2012
[47] J P Bressler L Olivi J H Cheong Y Kim A Maerten andD Bannon ldquoMetal transporters in intestine and brain theirinvolvement in metal-associated neurotoxicitiesrdquo Human andExperimental Toxicology vol 26 no 3 pp 221ndash229 2007
[48] Z Liu H Li M Soleimani et al ldquoCd2+ versus Zn2+ uptakeby the ZIP8 HCO
3mdashdependent symporter kinetics electro-
genicity and traffickingrdquo Biochemical and Biophysical ResearchCommunications vol 365 no 4 pp 814ndash820 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
6 ISRN Toxicology
1
08
06
04
02
01 10 100 1000
Wild type+VDCC
(a) (b)
(c) (d)
(e)
Figure 3 Effect of pulse treatment with Cd in CHOC120572 VDCC-expressing and wild-type CHO cells Microphotographs of control (a b) andCd-treated (c d) cells (a c) VDCC-expressing CHOC120572 cells and (b d) wild-type CHO cells Both cell types were incubated in 100120583M Cdfor 60 minutes in the presence of 30mM KCl Pictures were taken 24 h after wash of the metal Bar = 35 120583MThe graph (e) shows the effectof a 30min pulse treatment with Cd on cell adhesion as a function of concentration Trypan-blue excluding adherent cells were counted24 h after wash of the metal Points are average plusmn sem of 3 experiments in the same condition in CHOC120572 (filled circles) and wild-type CHO(empty circles) and were best fitted to the function NadherNadher(control) = 1(1 + ([Cd]ED50)) where NadherNadher(control) is the number ofadherent cells after Cd treatment normalized to the number of adherent cells in control culture [Cd] is the concentration of Cd and ED
50is
the concentration of Cd that causes detachment from the substrate of 50 of cells The best fit yielded ED50= 40 120583M for CHOC120572 and 43120583M
for wild-type CHOThe two curves are overlapped In contrast with the different appearance the two cell types were similarly affected by Cdtreatment
normalization to the total protein content Data shown inFigure 6 indicate that the difference in Cd accumulation isbarely or not significant when both cell types are incubatedin 100120583M Cd with or without elevated KCl On the otherhand BayK 8644 significantly enhanced and nimodipinesignificantly decreased metal accumulation in depolarizedCHOC120572 cells treated with 50 or 100120583M Cd but not in wild-type CHO cells
Toxicity of both Cd and Pb is mainly due to their abilityto permeate not simply mammalian cell membranes butspecifically tight membrane barriers such as the epitheliallining in the gastrointestinal tract luminal membrane ofproximal tubules in the kidney and the blood-brain barrierIn early work the influx of Pb and Cd into mammaliancells was frequently ascribed to permeation through calciumchannels and in particular L-type DHP-sensitive VDCC [1618 20 32]More recently other types of VDCC different fromthe classical DHP-sensitive L-type have been implicated bothin Cd [19 33] and Pb [23] uptake Moreover Pb has beenshown to use different pathways in several systems [34ndash37]and it is largely independent of VDCC even in neurons[23 24]
Here I have shown that expression of L-type VDCC is notsufficient to modify Cd uptake to a toxicological significantextent at least in the case of a brief intense exposure asthose used in this study Cd accumulation and sensitivitywerecomparable in two cell lines whose only difference was thepresence of L-typeVDCC in one of the two Indeedwild-type
CHO cells which do not possess VDCC are vulnerable to Cdpoisoning and take up Cd during relatively brief exposuresto an extent sufficient to trigger cell death by apoptosisor necrosis similarly to VDCC expressing CHOC120572 cellsAlthough VDCCs play an important role in Cd uptakein CHOC120572 cells as indicated by the sizeable effect of L-type channel DHP modifiers wild-type CHO cells use othertransport proteins with similar efficiency and outcome
It is thus clear that VDCCs are not the main mecha-nism responsible for toxic metal permeation through cellmembranes and tight barriers Rapid passive transport of Pbindependent of VDCC was reported at the brain endothe-lium [34 36 37] and even protein-independent transportof lead has been described [35] In astroglia cell models theuptake of Pb was shown to be driven by store-depletion-activated channels [38] and by two distinct pH-sensitivetransport mechanisms [39] VDCC-independent uptake ofPb by cerebellar granule neurons was inhibited by La [24]and this observation can provide some clue as for the mecha-nism of permeationThe involvement of a La-sensitive store-depletion activated channel as proposed in other systems[36 37 40] is unlikely because Pb influx did not requiredrainage of the stores Another possibility is that Pb2+ entersthe neuron via reverse operation of an exchanger similar (oridentical) to theNandashCa exchanger which is also very sensitiveto La block [41] and whose role in Zn permeation wasdemonstrated in cortical neurons [42] Also this possibilitywas discarded because NandashCa exchanger does not play
ISRN Toxicology 7
4
3
2
1
0Basal Cd K N B
Treatments
lowast
lowastlowast
(a) (b)
(c) (d)
(e)
Figure 4 Effect of dihydropyridines on Cd cytotoxicity in VDCC-expressing CHOC120572 cells Cells were treated for 30min in (a) controlmediumcontaining 30mMKCl (b) 100 120583M Cd+30mM KCl (c) 100 120583M Cd+30mM KCl+1 120583Mnimodipine (d) 100 120583M Cd+30mM KCl+1 120583M BayK8644 Note the partial recovery in shape of the cells treated in the presence of nimodipine and definitive lost of adhesion whenthe treatment was performed in the presence of BayK Bar = 25 120583M The graph (e) represents counts of viable adherent cells in the sameexperiment (average plusmn sem in 3 samples) in control (basal) following for a 30min treatment with 100 120583M Cd (Cd) Cd + 30mM KCl (K)Cd + 30mM KCl and 1 120583M nimodipine (N) and Cd + 30mM KCl and 1 120583M BayK (B) lowast indicates significantly different from control with119875 lt 005 and lowastlowast with 119875 lt 00001
40
30
20
10
0Wild type
ControlCd
Capa
cita
nce (
pf)
lowast
lowastlowast
Cd + nimo
+VDCC
Figure 5 Evidence of Cd-induced apoptosis change in cell capac-itance Cells were incubated with 100120583M Cd for 60 minutes andelectrical measurements were performed 24 h after wash Mem-brane capacitance was estimated from transient compensation (seeSection 24) Bars represent mean plusmn sem Treatment with Cd causedshrinkage of all cell as revealed by reduction of the cell capacitancein both wild-type (119899 = 15 in control and 119899 = 6 with Cd treatment)and CHOC120572 cells (119899 = 17 in control and 119899 = 39with Cd treatment)Nimodipine protected CHOC120572 cells from Cd-induced shrinkage(119899 = 10) lowast indicates significantly different from control with 119875 lt005 and lowastlowast with 119875 lt 0001
a significant role when the Na concentrations are close tothe physiological value as in Esposito et al [24] It is more
probable that Pb is taken up through a La-sensitive ion carriersimilar to the Zn transporter found in brain neurons [43]whose identity and features still wait full definition
Involvement of different transport proteins in Cd trans-port has been described At the epithelial lining of the gas-trointestinal barrier DMT1 the main transporter thatabsorbs Fe in the brush border membrane of the mammalianintestine has been shown to carry also different toxic metals[44ndash46] The role of this transporter in Cd absorption isunderscored by the fact that iron deficiency creates a sig-nificant risk for increased cadmium exposure by increasinggastrointestinal absorption from 5 to as much as 20In general a low level of essential metals favors uptake ofnonessential (toxic) elements that compete for their site andthis stresses the importance of evaluating metal balance incontrast with metal concentration In addition other metal-specific transporters have been identified at the cDNA level[47] and studied in expression systems [48]
4 Conclusion
Ionicmimicry is a useful framework to study themechanismsof metal toxicity Uptake of toxic metals such as Cd and Pbby mammalian cells of different tissues occurs through manydifferent pathways of which VDCCs have frequently beenregarded as prominent Because of the abundance of VDCCexcitable cells may be more susceptible to accumulate Cdand Pb than nonexcitable cells In this chapter I have shownthat although Cd and Pb can permeate through VDCC thisis not the most relevant mechanism of passive uptake ofPb while the mere presence of VDCC is not sufficient tomake cells significantly more vulnerable to Cd injury Otherfactors including different transport systems and expressionof detoxification proteins may be more relevant in the accu-mulation process of these toxic metals
8 ISRN Toxicology
Basal Cd K B NTreatments
Wild type
80
60
40
20
0
lowast
lowast
Figure 6 Total Cd accumulation measured by FAAS in CHOC120572and wild-type CHO cells Both cell types were incubated for 1 hourin medium containing 0 50 or 100 120583M Cd and other modifiersCd determination was normalized to the cell volume (see text)Treatments were as follows control 0 Cd (basal) 60min Cd (Cd)Cd + 30mM KCl (K) Cd + 30mM KCl and 1120583M BayK (B) Cd +30mM KCl and 1120583M nimodipine (N) Depolarization (treatmentK) did not increase significantly the intracellular concentration ofCd (119875 gt 005) In CHOC120572 cells but not in wild-type CHO BayKsignificantly enhanced and nimodipine significantly reduced Cdaccumulation both with 119875 lt 0001 (lowast) with respect to the sametreatments in depolarization Bars represent mean plusmn sem in at least10 experiments in each group
Acknowledgments
The author thanks Ilaria Zanardi and Irena Baranowska-Bosiacka (PUM Szczecin Poland) for help in Cd treatmentexperiments and Elisabetta Morelli for FAAS measurementsThe author is also grateful to Franz Hofmann and NorbertKlugbauer (Institut fur Pharmakologie und ToxikologieMunich Germany) for their permission to use the perma-nently transfected CHO cells and to Maurizio Taglialatelaand Mauro Cataldi (Universita Federico II Naples Italy) forsending him the cells
References
[1] R J Castellani P IMoreira G Liu et al ldquoIron the redox-activecenter of oxidative stress in Alzheimer diseaserdquo NeurochemicalResearch vol 32 no 10 pp 1640ndash1645 2007
[2] D G Smith R Cappai and K J Barnham ldquoThe redox chem-istry of the Alzheimerrsquos disease amyloid 120573 peptiderdquo Biochimicaet Biophysica Acta vol 1768 no 8 pp 1976ndash1990 2007
[3] Y Xu L Feng P D Jeffrey Y Shi and FMMMorel ldquoStructureand metal exchange in the cadmium carbonic anhydrase ofmarine diatomsrdquo Nature vol 452 no 7183 pp 56ndash61 2008
[4] T W Clarkson ldquoMolecular and ionic mimicry of toxic metalsrdquoAnnual Review of Pharmacology and Toxicology vol 33 pp545ndash571 1993
[5] C C Bridges and R K Zalups ldquoMolecular and ionic mimicryand the transport of toxic metalsrdquo Toxicology and Applied
Pharmacology vol 204 no 3 pp 274ndash308 2005[6] H Ouyang and H J Vogel ldquoMetal ion binding to calmodulin
NMRand fluorescence studiesrdquoBioMetals vol 11 no 3 pp 213ndash222 1998
[7] M KernMWisniewski L Cabell andG Audesirk ldquoInorganiclead and calcium interact positively in activation of calmod-ulinrdquo NeuroToxicology vol 21 no 3 pp 353ndash364 2000
[8] J Markovac and GW Goldstein ldquoPicomolar concentrations oflead stimulate brain protein kinase CrdquoNature vol 334 no 6177pp 71ndash73 1988
[9] X Sun X Tian J L Tomsig and J B Suszkiw ldquoAnalysisof differential effects of Pb2+ on protein kinase C isozymesrdquoToxicology and Applied Pharmacology vol 156 no 1 pp 40ndash451999
[10] J L Tomsig and J B Suszkiw ldquoMultisite interactions betweenPb2+ and protein kinase C and its role in norepinephrine releasefrom bovine adrenal chromaffin cellsrdquo Journal of Neurochem-istry vol 64 no 6 pp 2667ndash2673 1995
[11] C M Bouton L P Frelin C E Forde H A Godwin and JPevsner ldquoSynaptotagmin I is amolecular target for leadrdquo Journalof Neurochemistry vol 76 no 6 pp 1724ndash1735 2001
[12] J Yang P T Ellinor W A Sather J F Zhang and R W TsienldquoMolecular determinants of Ca2+ selectivity and ion permeationin L-type Ca2+ channelsrdquoNature vol 366 no 6451 pp 158ndash1611993
[13] MWakamori M Strobeck T Niidome T Teramoto K Imotoand Y Mori ldquoFunctional characterization of ion permeationpathway in the N-type Ca2+ channelrdquo Journal of Neurophysiol-ogy vol 79 no 2 pp 622ndash634 1998
[14] S Peng R K Hajela and W D Atchison ldquoCharacteristics ofblock by Pb2+ of function of human neuronal L- N- and R-type Ca2+ channels transiently expressed in human embryonickidney 293 cellsrdquo Molecular Pharmacology vol 62 no 6 pp1418ndash1430 2002
[15] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium uptake in excitable mammalian cells a microspec-trofluorimetric studyrdquo European Journal of Histochemistry vol41 pp 189ndash190 1997
[16] P M Hinkle P A Kinsella and K C Osterhoudt ldquoCadmiumuptake and toxicity via voltage-sensitive calcium channelsrdquoJournal of Biological Chemistry vol 262 no 34 pp 16333ndash163371987
[17] P M Hinkle E D Shanshala and E J Nelson ldquoMeasurementsof intracellular cadmium with fluorescent dyesmdashfurther evi-dence for the role of calcium channels in cadmium uptakerdquoTheJournal of Biological Chemistry vol 267 pp 25553ndash25559 1992
[18] P M Hinkle and M E Osborne ldquoCadmium toxicity in ratpheochromocytoma cells studies on themechanism of uptakerdquoToxicology and Applied Pharmacology vol 124 no 1 pp 91ndash981994
[19] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium influx in mammalian neuronsrdquo Journal of Neuro-chemistry vol 72 no 5 pp 2154ndash2161 1999
[20] V Souza L Bucio andMCGutierrez-Ruiz ldquoCadmiumuptakeby a human hepatic cell line (WRL-68 cells)rdquo Toxicology vol120 no 3 pp 215ndash220 1997
[21] P Gavazzo E Morelli and C Marchetti ldquoSusceptibility ofinsulinoma cells to cadmiumandmodulation by L-type calciumchannelsrdquo BioMetals vol 18 no 2 pp 131ndash142 2005
[22] M Kirberger and J J Yang ldquoStructural differences betweenPb2+- and Ca2+-binding sites in proteins implications with
ISRN Toxicology 9
respect to toxicityrdquo Journal of Inorganic Biochemistry vol 102no 10 pp 1901ndash1909 2008
[23] MMazzolini S Traverso andCMarchetti ldquoMultiple pathwaysof Pb2+ permeation in rat cerebellar granule neuronesrdquo Journalof Neurochemistry vol 79 no 2 pp 407ndash416 2001
[24] A Esposito M Robello F Pellistri and C Marchetti ldquoTwo-photon analysis of lead accumulation in rat cerebellar granuleneuronsrdquo Neurochemical Research vol 30 no 8 pp 949ndash9542005
[25] C Marchetti C Amico and C Usai ldquoFunctional characteriza-tion of the effect of nimodipine on the calcium current in ratcerebellar granule cellsrdquo Journal of Neurophysiology vol 73 no3 pp 1169ndash1180 1995
[26] A Welling E Bosse A Cavalie et al ldquoStable co-expression ofcalcium channel 1205721120573 and 1205722120575 subunits in a somatic cell linerdquoJournal of Physiology vol 471 pp 749ndash765 1993
[27] A Diaspro S Annunziata M Raimondo P Ramoino and MRobello ldquoA single-pinhole confocal laser scanning microscopefor 3-D imaging of biostructuresrdquo IEEE Engineering inMedicineand Biology Magazine vol 18 no 4 pp 106ndash110 1999
[28] F Pellistri A Cupello A Esposito C Marchetti and MRobello ldquoTwo-photon imaging of calcium accumulation in ratcerebellar granule cellsrdquo NeuroReport vol 15 no 1 pp 83ndash872004
[29] M Cataldi A Secondo A DrsquoAlessio et al ldquoStudies onmaitotoxin-induced intracellular Ca2+ elevation in chinesehamster ovary cells stably transfected with cDNAs encodingfor L-type Ca2+ channel subunitsrdquo Journal of Pharmacology andExperimental Therapeutics vol 290 no 2 pp 725ndash730 1999
[30] A Galan A Troyano N E Vilaboa C Fernandez E DeBlas and P Aller ldquoModulation of the stress response duringapoptosis and necrosis induction in cadmium-treated U-937human promonocytic cellsrdquo Biochimica et Biophysica Acta vol1538 no 1 pp 38ndash46 2001
[31] A H B Poliandri J P Cabilla M O Velardez C C ABodo and B H Duvilanski ldquoCadmium induces apoptosis inanterior pituitary cells that can be reversed by treatment withantioxidantsrdquo Toxicology and Applied Pharmacology vol 190no 1 pp 17ndash24 2003
[32] J L Tomsig and J B Suszkiw ldquoPermeation of Pb2+ throughcalcium channels fura-2 measurements of voltage- anddihydropyridine-sensitive Pb2+ entry in isolated bovinechromaffin cellsrdquo Biochimica et Biophysica Acta vol 1069 no2 pp 197ndash200 1991
[33] EM Leslie J Liu C D Klaassen andM PWaalkes ldquoAcquiredcadmium resistance in metallothionein-III(minusminus) knockoutcells role of the T-type calcium channel Cacn1205721G in cadmiumuptakerdquo Molecular Pharmacology vol 69 no 2 pp 629ndash6392006
[34] R Deane and M W B Bradbury ldquoTransport of lead-203 atthe blood-brain barrier during short cerebrovascular perfusionwith saline in the ratrdquo Journal of Neurochemistry vol 54 no 3pp 905ndash914 1990
[35] R S Diaz and J Monreal ldquoProtein-independent lead perme-ation through myelin lipid liposomesrdquo Molecular Pharmacol-ogy vol 47 no 4 pp 766ndash771 1995
[36] L E Kerper and P M Hinkle ldquoCellular uptake of lead isactivated by depletion of intracellular calcium storesrdquo Journalof Biological Chemistry vol 272 no 13 pp 8346ndash8352 1997
[37] L E Kerper and P M Hinkle ldquoLead uptake in brain capillaryendothelial cells activation by calcium store depletionrdquoToxicol-ogy and Applied Pharmacology vol 146 no 1 pp 127ndash133 1997
[38] M E Legare R Barhoumi E Hebert G R Bratton R CBurghardt and E Tiffany- Castiglioni ldquoAnalysis of Pb2+ entryinto cultured astrogliardquo Toxicological Sciences vol 46 no 1 pp90ndash100 1998
[39] J H Cheong D Bannon L Olivi Y Kim and J BresslerldquoDifferent mechanismsmediate uptake of lead in a rat astroglialcell linerdquoToxicological Sciences vol 77 no 2 pp 334ndash340 2004
[40] T J Evans M R James-Kracke S B Kleiboeker and S WCasteel ldquoLead enters Rcho-1 trophoblastic cells by calciumtransport mechanisms and complexes with cytosolic calcium-binding proteinsrdquo Toxicology and Applied Pharmacology vol186 no 2 pp 77ndash89 2003
[41] G J Kaczorowski R S Slaughter V F King and M LGarcia ldquoInhibitors of sodium-calcium exchange identificationand development of probes of transport activityrdquo Biochimica etBiophysica Acta vol 988 no 2 pp 287ndash302 1989
[42] S L Sensi L M T Canzoniero S P Yu et al ldquoMeasurement ofintracellular free zinc in living cortical neurons routes of entryrdquoJournal of Neuroscience vol 17 no 24 pp 9554ndash9564 1997
[43] R A Colvin ldquoCharacterization of a plasma membrane zinctransporter in rat brainrdquo Neuroscience Letters vol 247 no 2-3pp 147ndash150 1998
[44] V Picard G Govoni N Jabado and P Gross ldquoNramp 2(DCT1DMT1) expressed at the plasma membrane transportsiron and other divalent cations into a calcein-accessible cyto-plasmic poolrdquo Journal of Biological Chemistry vol 275 no 46pp 35738ndash35745 2000
[45] M Okubo K Yamada M Hosoyamada T Shibasaki and HEndou ldquoCadmium transport by human Nramp 2 expressed inXenopus laevis oocytesrdquo Toxicology and Applied Pharmacologyvol 187 no 3 pp 162ndash167 2003
[46] A C Illing A Shawki C L Cunningham and B MackenzieldquoSubstrate profile and metal-ion selectivity of human divalentmetal-ion transporter-1rdquo The Journal of Biological Chemistryvol 287 pp 30485ndash30496 2012
[47] J P Bressler L Olivi J H Cheong Y Kim A Maerten andD Bannon ldquoMetal transporters in intestine and brain theirinvolvement in metal-associated neurotoxicitiesrdquo Human andExperimental Toxicology vol 26 no 3 pp 221ndash229 2007
[48] Z Liu H Li M Soleimani et al ldquoCd2+ versus Zn2+ uptakeby the ZIP8 HCO
3mdashdependent symporter kinetics electro-
genicity and traffickingrdquo Biochemical and Biophysical ResearchCommunications vol 365 no 4 pp 814ndash820 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

ISRN Toxicology 7
4
3
2
1
0Basal Cd K N B
Treatments
lowast
lowastlowast
(a) (b)
(c) (d)
(e)
Figure 4 Effect of dihydropyridines on Cd cytotoxicity in VDCC-expressing CHOC120572 cells Cells were treated for 30min in (a) controlmediumcontaining 30mMKCl (b) 100 120583M Cd+30mM KCl (c) 100 120583M Cd+30mM KCl+1 120583Mnimodipine (d) 100 120583M Cd+30mM KCl+1 120583M BayK8644 Note the partial recovery in shape of the cells treated in the presence of nimodipine and definitive lost of adhesion whenthe treatment was performed in the presence of BayK Bar = 25 120583M The graph (e) represents counts of viable adherent cells in the sameexperiment (average plusmn sem in 3 samples) in control (basal) following for a 30min treatment with 100 120583M Cd (Cd) Cd + 30mM KCl (K)Cd + 30mM KCl and 1 120583M nimodipine (N) and Cd + 30mM KCl and 1 120583M BayK (B) lowast indicates significantly different from control with119875 lt 005 and lowastlowast with 119875 lt 00001
40
30
20
10
0Wild type
ControlCd
Capa
cita
nce (
pf)
lowast
lowastlowast
Cd + nimo
+VDCC
Figure 5 Evidence of Cd-induced apoptosis change in cell capac-itance Cells were incubated with 100120583M Cd for 60 minutes andelectrical measurements were performed 24 h after wash Mem-brane capacitance was estimated from transient compensation (seeSection 24) Bars represent mean plusmn sem Treatment with Cd causedshrinkage of all cell as revealed by reduction of the cell capacitancein both wild-type (119899 = 15 in control and 119899 = 6 with Cd treatment)and CHOC120572 cells (119899 = 17 in control and 119899 = 39with Cd treatment)Nimodipine protected CHOC120572 cells from Cd-induced shrinkage(119899 = 10) lowast indicates significantly different from control with 119875 lt005 and lowastlowast with 119875 lt 0001
a significant role when the Na concentrations are close tothe physiological value as in Esposito et al [24] It is more
probable that Pb is taken up through a La-sensitive ion carriersimilar to the Zn transporter found in brain neurons [43]whose identity and features still wait full definition
Involvement of different transport proteins in Cd trans-port has been described At the epithelial lining of the gas-trointestinal barrier DMT1 the main transporter thatabsorbs Fe in the brush border membrane of the mammalianintestine has been shown to carry also different toxic metals[44ndash46] The role of this transporter in Cd absorption isunderscored by the fact that iron deficiency creates a sig-nificant risk for increased cadmium exposure by increasinggastrointestinal absorption from 5 to as much as 20In general a low level of essential metals favors uptake ofnonessential (toxic) elements that compete for their site andthis stresses the importance of evaluating metal balance incontrast with metal concentration In addition other metal-specific transporters have been identified at the cDNA level[47] and studied in expression systems [48]
4 Conclusion
Ionicmimicry is a useful framework to study themechanismsof metal toxicity Uptake of toxic metals such as Cd and Pbby mammalian cells of different tissues occurs through manydifferent pathways of which VDCCs have frequently beenregarded as prominent Because of the abundance of VDCCexcitable cells may be more susceptible to accumulate Cdand Pb than nonexcitable cells In this chapter I have shownthat although Cd and Pb can permeate through VDCC thisis not the most relevant mechanism of passive uptake ofPb while the mere presence of VDCC is not sufficient tomake cells significantly more vulnerable to Cd injury Otherfactors including different transport systems and expressionof detoxification proteins may be more relevant in the accu-mulation process of these toxic metals
8 ISRN Toxicology
Basal Cd K B NTreatments
Wild type
80
60
40
20
0
lowast
lowast
Figure 6 Total Cd accumulation measured by FAAS in CHOC120572and wild-type CHO cells Both cell types were incubated for 1 hourin medium containing 0 50 or 100 120583M Cd and other modifiersCd determination was normalized to the cell volume (see text)Treatments were as follows control 0 Cd (basal) 60min Cd (Cd)Cd + 30mM KCl (K) Cd + 30mM KCl and 1120583M BayK (B) Cd +30mM KCl and 1120583M nimodipine (N) Depolarization (treatmentK) did not increase significantly the intracellular concentration ofCd (119875 gt 005) In CHOC120572 cells but not in wild-type CHO BayKsignificantly enhanced and nimodipine significantly reduced Cdaccumulation both with 119875 lt 0001 (lowast) with respect to the sametreatments in depolarization Bars represent mean plusmn sem in at least10 experiments in each group
Acknowledgments
The author thanks Ilaria Zanardi and Irena Baranowska-Bosiacka (PUM Szczecin Poland) for help in Cd treatmentexperiments and Elisabetta Morelli for FAAS measurementsThe author is also grateful to Franz Hofmann and NorbertKlugbauer (Institut fur Pharmakologie und ToxikologieMunich Germany) for their permission to use the perma-nently transfected CHO cells and to Maurizio Taglialatelaand Mauro Cataldi (Universita Federico II Naples Italy) forsending him the cells
References
[1] R J Castellani P IMoreira G Liu et al ldquoIron the redox-activecenter of oxidative stress in Alzheimer diseaserdquo NeurochemicalResearch vol 32 no 10 pp 1640ndash1645 2007
[2] D G Smith R Cappai and K J Barnham ldquoThe redox chem-istry of the Alzheimerrsquos disease amyloid 120573 peptiderdquo Biochimicaet Biophysica Acta vol 1768 no 8 pp 1976ndash1990 2007
[3] Y Xu L Feng P D Jeffrey Y Shi and FMMMorel ldquoStructureand metal exchange in the cadmium carbonic anhydrase ofmarine diatomsrdquo Nature vol 452 no 7183 pp 56ndash61 2008
[4] T W Clarkson ldquoMolecular and ionic mimicry of toxic metalsrdquoAnnual Review of Pharmacology and Toxicology vol 33 pp545ndash571 1993
[5] C C Bridges and R K Zalups ldquoMolecular and ionic mimicryand the transport of toxic metalsrdquo Toxicology and Applied
Pharmacology vol 204 no 3 pp 274ndash308 2005[6] H Ouyang and H J Vogel ldquoMetal ion binding to calmodulin
NMRand fluorescence studiesrdquoBioMetals vol 11 no 3 pp 213ndash222 1998
[7] M KernMWisniewski L Cabell andG Audesirk ldquoInorganiclead and calcium interact positively in activation of calmod-ulinrdquo NeuroToxicology vol 21 no 3 pp 353ndash364 2000
[8] J Markovac and GW Goldstein ldquoPicomolar concentrations oflead stimulate brain protein kinase CrdquoNature vol 334 no 6177pp 71ndash73 1988
[9] X Sun X Tian J L Tomsig and J B Suszkiw ldquoAnalysisof differential effects of Pb2+ on protein kinase C isozymesrdquoToxicology and Applied Pharmacology vol 156 no 1 pp 40ndash451999
[10] J L Tomsig and J B Suszkiw ldquoMultisite interactions betweenPb2+ and protein kinase C and its role in norepinephrine releasefrom bovine adrenal chromaffin cellsrdquo Journal of Neurochem-istry vol 64 no 6 pp 2667ndash2673 1995
[11] C M Bouton L P Frelin C E Forde H A Godwin and JPevsner ldquoSynaptotagmin I is amolecular target for leadrdquo Journalof Neurochemistry vol 76 no 6 pp 1724ndash1735 2001
[12] J Yang P T Ellinor W A Sather J F Zhang and R W TsienldquoMolecular determinants of Ca2+ selectivity and ion permeationin L-type Ca2+ channelsrdquoNature vol 366 no 6451 pp 158ndash1611993
[13] MWakamori M Strobeck T Niidome T Teramoto K Imotoand Y Mori ldquoFunctional characterization of ion permeationpathway in the N-type Ca2+ channelrdquo Journal of Neurophysiol-ogy vol 79 no 2 pp 622ndash634 1998
[14] S Peng R K Hajela and W D Atchison ldquoCharacteristics ofblock by Pb2+ of function of human neuronal L- N- and R-type Ca2+ channels transiently expressed in human embryonickidney 293 cellsrdquo Molecular Pharmacology vol 62 no 6 pp1418ndash1430 2002
[15] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium uptake in excitable mammalian cells a microspec-trofluorimetric studyrdquo European Journal of Histochemistry vol41 pp 189ndash190 1997
[16] P M Hinkle P A Kinsella and K C Osterhoudt ldquoCadmiumuptake and toxicity via voltage-sensitive calcium channelsrdquoJournal of Biological Chemistry vol 262 no 34 pp 16333ndash163371987
[17] P M Hinkle E D Shanshala and E J Nelson ldquoMeasurementsof intracellular cadmium with fluorescent dyesmdashfurther evi-dence for the role of calcium channels in cadmium uptakerdquoTheJournal of Biological Chemistry vol 267 pp 25553ndash25559 1992
[18] P M Hinkle and M E Osborne ldquoCadmium toxicity in ratpheochromocytoma cells studies on themechanism of uptakerdquoToxicology and Applied Pharmacology vol 124 no 1 pp 91ndash981994
[19] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium influx in mammalian neuronsrdquo Journal of Neuro-chemistry vol 72 no 5 pp 2154ndash2161 1999
[20] V Souza L Bucio andMCGutierrez-Ruiz ldquoCadmiumuptakeby a human hepatic cell line (WRL-68 cells)rdquo Toxicology vol120 no 3 pp 215ndash220 1997
[21] P Gavazzo E Morelli and C Marchetti ldquoSusceptibility ofinsulinoma cells to cadmiumandmodulation by L-type calciumchannelsrdquo BioMetals vol 18 no 2 pp 131ndash142 2005
[22] M Kirberger and J J Yang ldquoStructural differences betweenPb2+- and Ca2+-binding sites in proteins implications with
ISRN Toxicology 9
respect to toxicityrdquo Journal of Inorganic Biochemistry vol 102no 10 pp 1901ndash1909 2008
[23] MMazzolini S Traverso andCMarchetti ldquoMultiple pathwaysof Pb2+ permeation in rat cerebellar granule neuronesrdquo Journalof Neurochemistry vol 79 no 2 pp 407ndash416 2001
[24] A Esposito M Robello F Pellistri and C Marchetti ldquoTwo-photon analysis of lead accumulation in rat cerebellar granuleneuronsrdquo Neurochemical Research vol 30 no 8 pp 949ndash9542005
[25] C Marchetti C Amico and C Usai ldquoFunctional characteriza-tion of the effect of nimodipine on the calcium current in ratcerebellar granule cellsrdquo Journal of Neurophysiology vol 73 no3 pp 1169ndash1180 1995
[26] A Welling E Bosse A Cavalie et al ldquoStable co-expression ofcalcium channel 1205721120573 and 1205722120575 subunits in a somatic cell linerdquoJournal of Physiology vol 471 pp 749ndash765 1993
[27] A Diaspro S Annunziata M Raimondo P Ramoino and MRobello ldquoA single-pinhole confocal laser scanning microscopefor 3-D imaging of biostructuresrdquo IEEE Engineering inMedicineand Biology Magazine vol 18 no 4 pp 106ndash110 1999
[28] F Pellistri A Cupello A Esposito C Marchetti and MRobello ldquoTwo-photon imaging of calcium accumulation in ratcerebellar granule cellsrdquo NeuroReport vol 15 no 1 pp 83ndash872004
[29] M Cataldi A Secondo A DrsquoAlessio et al ldquoStudies onmaitotoxin-induced intracellular Ca2+ elevation in chinesehamster ovary cells stably transfected with cDNAs encodingfor L-type Ca2+ channel subunitsrdquo Journal of Pharmacology andExperimental Therapeutics vol 290 no 2 pp 725ndash730 1999
[30] A Galan A Troyano N E Vilaboa C Fernandez E DeBlas and P Aller ldquoModulation of the stress response duringapoptosis and necrosis induction in cadmium-treated U-937human promonocytic cellsrdquo Biochimica et Biophysica Acta vol1538 no 1 pp 38ndash46 2001
[31] A H B Poliandri J P Cabilla M O Velardez C C ABodo and B H Duvilanski ldquoCadmium induces apoptosis inanterior pituitary cells that can be reversed by treatment withantioxidantsrdquo Toxicology and Applied Pharmacology vol 190no 1 pp 17ndash24 2003
[32] J L Tomsig and J B Suszkiw ldquoPermeation of Pb2+ throughcalcium channels fura-2 measurements of voltage- anddihydropyridine-sensitive Pb2+ entry in isolated bovinechromaffin cellsrdquo Biochimica et Biophysica Acta vol 1069 no2 pp 197ndash200 1991
[33] EM Leslie J Liu C D Klaassen andM PWaalkes ldquoAcquiredcadmium resistance in metallothionein-III(minusminus) knockoutcells role of the T-type calcium channel Cacn1205721G in cadmiumuptakerdquo Molecular Pharmacology vol 69 no 2 pp 629ndash6392006
[34] R Deane and M W B Bradbury ldquoTransport of lead-203 atthe blood-brain barrier during short cerebrovascular perfusionwith saline in the ratrdquo Journal of Neurochemistry vol 54 no 3pp 905ndash914 1990
[35] R S Diaz and J Monreal ldquoProtein-independent lead perme-ation through myelin lipid liposomesrdquo Molecular Pharmacol-ogy vol 47 no 4 pp 766ndash771 1995
[36] L E Kerper and P M Hinkle ldquoCellular uptake of lead isactivated by depletion of intracellular calcium storesrdquo Journalof Biological Chemistry vol 272 no 13 pp 8346ndash8352 1997
[37] L E Kerper and P M Hinkle ldquoLead uptake in brain capillaryendothelial cells activation by calcium store depletionrdquoToxicol-ogy and Applied Pharmacology vol 146 no 1 pp 127ndash133 1997
[38] M E Legare R Barhoumi E Hebert G R Bratton R CBurghardt and E Tiffany- Castiglioni ldquoAnalysis of Pb2+ entryinto cultured astrogliardquo Toxicological Sciences vol 46 no 1 pp90ndash100 1998
[39] J H Cheong D Bannon L Olivi Y Kim and J BresslerldquoDifferent mechanismsmediate uptake of lead in a rat astroglialcell linerdquoToxicological Sciences vol 77 no 2 pp 334ndash340 2004
[40] T J Evans M R James-Kracke S B Kleiboeker and S WCasteel ldquoLead enters Rcho-1 trophoblastic cells by calciumtransport mechanisms and complexes with cytosolic calcium-binding proteinsrdquo Toxicology and Applied Pharmacology vol186 no 2 pp 77ndash89 2003
[41] G J Kaczorowski R S Slaughter V F King and M LGarcia ldquoInhibitors of sodium-calcium exchange identificationand development of probes of transport activityrdquo Biochimica etBiophysica Acta vol 988 no 2 pp 287ndash302 1989
[42] S L Sensi L M T Canzoniero S P Yu et al ldquoMeasurement ofintracellular free zinc in living cortical neurons routes of entryrdquoJournal of Neuroscience vol 17 no 24 pp 9554ndash9564 1997
[43] R A Colvin ldquoCharacterization of a plasma membrane zinctransporter in rat brainrdquo Neuroscience Letters vol 247 no 2-3pp 147ndash150 1998
[44] V Picard G Govoni N Jabado and P Gross ldquoNramp 2(DCT1DMT1) expressed at the plasma membrane transportsiron and other divalent cations into a calcein-accessible cyto-plasmic poolrdquo Journal of Biological Chemistry vol 275 no 46pp 35738ndash35745 2000
[45] M Okubo K Yamada M Hosoyamada T Shibasaki and HEndou ldquoCadmium transport by human Nramp 2 expressed inXenopus laevis oocytesrdquo Toxicology and Applied Pharmacologyvol 187 no 3 pp 162ndash167 2003
[46] A C Illing A Shawki C L Cunningham and B MackenzieldquoSubstrate profile and metal-ion selectivity of human divalentmetal-ion transporter-1rdquo The Journal of Biological Chemistryvol 287 pp 30485ndash30496 2012
[47] J P Bressler L Olivi J H Cheong Y Kim A Maerten andD Bannon ldquoMetal transporters in intestine and brain theirinvolvement in metal-associated neurotoxicitiesrdquo Human andExperimental Toxicology vol 26 no 3 pp 221ndash229 2007
[48] Z Liu H Li M Soleimani et al ldquoCd2+ versus Zn2+ uptakeby the ZIP8 HCO
3mdashdependent symporter kinetics electro-
genicity and traffickingrdquo Biochemical and Biophysical ResearchCommunications vol 365 no 4 pp 814ndash820 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

8 ISRN Toxicology
Basal Cd K B NTreatments
Wild type
80
60
40
20
0
lowast
lowast
Figure 6 Total Cd accumulation measured by FAAS in CHOC120572and wild-type CHO cells Both cell types were incubated for 1 hourin medium containing 0 50 or 100 120583M Cd and other modifiersCd determination was normalized to the cell volume (see text)Treatments were as follows control 0 Cd (basal) 60min Cd (Cd)Cd + 30mM KCl (K) Cd + 30mM KCl and 1120583M BayK (B) Cd +30mM KCl and 1120583M nimodipine (N) Depolarization (treatmentK) did not increase significantly the intracellular concentration ofCd (119875 gt 005) In CHOC120572 cells but not in wild-type CHO BayKsignificantly enhanced and nimodipine significantly reduced Cdaccumulation both with 119875 lt 0001 (lowast) with respect to the sametreatments in depolarization Bars represent mean plusmn sem in at least10 experiments in each group
Acknowledgments
The author thanks Ilaria Zanardi and Irena Baranowska-Bosiacka (PUM Szczecin Poland) for help in Cd treatmentexperiments and Elisabetta Morelli for FAAS measurementsThe author is also grateful to Franz Hofmann and NorbertKlugbauer (Institut fur Pharmakologie und ToxikologieMunich Germany) for their permission to use the perma-nently transfected CHO cells and to Maurizio Taglialatelaand Mauro Cataldi (Universita Federico II Naples Italy) forsending him the cells
References
[1] R J Castellani P IMoreira G Liu et al ldquoIron the redox-activecenter of oxidative stress in Alzheimer diseaserdquo NeurochemicalResearch vol 32 no 10 pp 1640ndash1645 2007
[2] D G Smith R Cappai and K J Barnham ldquoThe redox chem-istry of the Alzheimerrsquos disease amyloid 120573 peptiderdquo Biochimicaet Biophysica Acta vol 1768 no 8 pp 1976ndash1990 2007
[3] Y Xu L Feng P D Jeffrey Y Shi and FMMMorel ldquoStructureand metal exchange in the cadmium carbonic anhydrase ofmarine diatomsrdquo Nature vol 452 no 7183 pp 56ndash61 2008
[4] T W Clarkson ldquoMolecular and ionic mimicry of toxic metalsrdquoAnnual Review of Pharmacology and Toxicology vol 33 pp545ndash571 1993
[5] C C Bridges and R K Zalups ldquoMolecular and ionic mimicryand the transport of toxic metalsrdquo Toxicology and Applied
Pharmacology vol 204 no 3 pp 274ndash308 2005[6] H Ouyang and H J Vogel ldquoMetal ion binding to calmodulin
NMRand fluorescence studiesrdquoBioMetals vol 11 no 3 pp 213ndash222 1998
[7] M KernMWisniewski L Cabell andG Audesirk ldquoInorganiclead and calcium interact positively in activation of calmod-ulinrdquo NeuroToxicology vol 21 no 3 pp 353ndash364 2000
[8] J Markovac and GW Goldstein ldquoPicomolar concentrations oflead stimulate brain protein kinase CrdquoNature vol 334 no 6177pp 71ndash73 1988
[9] X Sun X Tian J L Tomsig and J B Suszkiw ldquoAnalysisof differential effects of Pb2+ on protein kinase C isozymesrdquoToxicology and Applied Pharmacology vol 156 no 1 pp 40ndash451999
[10] J L Tomsig and J B Suszkiw ldquoMultisite interactions betweenPb2+ and protein kinase C and its role in norepinephrine releasefrom bovine adrenal chromaffin cellsrdquo Journal of Neurochem-istry vol 64 no 6 pp 2667ndash2673 1995
[11] C M Bouton L P Frelin C E Forde H A Godwin and JPevsner ldquoSynaptotagmin I is amolecular target for leadrdquo Journalof Neurochemistry vol 76 no 6 pp 1724ndash1735 2001
[12] J Yang P T Ellinor W A Sather J F Zhang and R W TsienldquoMolecular determinants of Ca2+ selectivity and ion permeationin L-type Ca2+ channelsrdquoNature vol 366 no 6451 pp 158ndash1611993
[13] MWakamori M Strobeck T Niidome T Teramoto K Imotoand Y Mori ldquoFunctional characterization of ion permeationpathway in the N-type Ca2+ channelrdquo Journal of Neurophysiol-ogy vol 79 no 2 pp 622ndash634 1998
[14] S Peng R K Hajela and W D Atchison ldquoCharacteristics ofblock by Pb2+ of function of human neuronal L- N- and R-type Ca2+ channels transiently expressed in human embryonickidney 293 cellsrdquo Molecular Pharmacology vol 62 no 6 pp1418ndash1430 2002
[15] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium uptake in excitable mammalian cells a microspec-trofluorimetric studyrdquo European Journal of Histochemistry vol41 pp 189ndash190 1997
[16] P M Hinkle P A Kinsella and K C Osterhoudt ldquoCadmiumuptake and toxicity via voltage-sensitive calcium channelsrdquoJournal of Biological Chemistry vol 262 no 34 pp 16333ndash163371987
[17] P M Hinkle E D Shanshala and E J Nelson ldquoMeasurementsof intracellular cadmium with fluorescent dyesmdashfurther evi-dence for the role of calcium channels in cadmium uptakerdquoTheJournal of Biological Chemistry vol 267 pp 25553ndash25559 1992
[18] P M Hinkle and M E Osborne ldquoCadmium toxicity in ratpheochromocytoma cells studies on themechanism of uptakerdquoToxicology and Applied Pharmacology vol 124 no 1 pp 91ndash981994
[19] CUsai A Barberis LMoccagatta andCMarchetti ldquoPathwaysof cadmium influx in mammalian neuronsrdquo Journal of Neuro-chemistry vol 72 no 5 pp 2154ndash2161 1999
[20] V Souza L Bucio andMCGutierrez-Ruiz ldquoCadmiumuptakeby a human hepatic cell line (WRL-68 cells)rdquo Toxicology vol120 no 3 pp 215ndash220 1997
[21] P Gavazzo E Morelli and C Marchetti ldquoSusceptibility ofinsulinoma cells to cadmiumandmodulation by L-type calciumchannelsrdquo BioMetals vol 18 no 2 pp 131ndash142 2005
[22] M Kirberger and J J Yang ldquoStructural differences betweenPb2+- and Ca2+-binding sites in proteins implications with
ISRN Toxicology 9
respect to toxicityrdquo Journal of Inorganic Biochemistry vol 102no 10 pp 1901ndash1909 2008
[23] MMazzolini S Traverso andCMarchetti ldquoMultiple pathwaysof Pb2+ permeation in rat cerebellar granule neuronesrdquo Journalof Neurochemistry vol 79 no 2 pp 407ndash416 2001
[24] A Esposito M Robello F Pellistri and C Marchetti ldquoTwo-photon analysis of lead accumulation in rat cerebellar granuleneuronsrdquo Neurochemical Research vol 30 no 8 pp 949ndash9542005
[25] C Marchetti C Amico and C Usai ldquoFunctional characteriza-tion of the effect of nimodipine on the calcium current in ratcerebellar granule cellsrdquo Journal of Neurophysiology vol 73 no3 pp 1169ndash1180 1995
[26] A Welling E Bosse A Cavalie et al ldquoStable co-expression ofcalcium channel 1205721120573 and 1205722120575 subunits in a somatic cell linerdquoJournal of Physiology vol 471 pp 749ndash765 1993
[27] A Diaspro S Annunziata M Raimondo P Ramoino and MRobello ldquoA single-pinhole confocal laser scanning microscopefor 3-D imaging of biostructuresrdquo IEEE Engineering inMedicineand Biology Magazine vol 18 no 4 pp 106ndash110 1999
[28] F Pellistri A Cupello A Esposito C Marchetti and MRobello ldquoTwo-photon imaging of calcium accumulation in ratcerebellar granule cellsrdquo NeuroReport vol 15 no 1 pp 83ndash872004
[29] M Cataldi A Secondo A DrsquoAlessio et al ldquoStudies onmaitotoxin-induced intracellular Ca2+ elevation in chinesehamster ovary cells stably transfected with cDNAs encodingfor L-type Ca2+ channel subunitsrdquo Journal of Pharmacology andExperimental Therapeutics vol 290 no 2 pp 725ndash730 1999
[30] A Galan A Troyano N E Vilaboa C Fernandez E DeBlas and P Aller ldquoModulation of the stress response duringapoptosis and necrosis induction in cadmium-treated U-937human promonocytic cellsrdquo Biochimica et Biophysica Acta vol1538 no 1 pp 38ndash46 2001
[31] A H B Poliandri J P Cabilla M O Velardez C C ABodo and B H Duvilanski ldquoCadmium induces apoptosis inanterior pituitary cells that can be reversed by treatment withantioxidantsrdquo Toxicology and Applied Pharmacology vol 190no 1 pp 17ndash24 2003
[32] J L Tomsig and J B Suszkiw ldquoPermeation of Pb2+ throughcalcium channels fura-2 measurements of voltage- anddihydropyridine-sensitive Pb2+ entry in isolated bovinechromaffin cellsrdquo Biochimica et Biophysica Acta vol 1069 no2 pp 197ndash200 1991
[33] EM Leslie J Liu C D Klaassen andM PWaalkes ldquoAcquiredcadmium resistance in metallothionein-III(minusminus) knockoutcells role of the T-type calcium channel Cacn1205721G in cadmiumuptakerdquo Molecular Pharmacology vol 69 no 2 pp 629ndash6392006
[34] R Deane and M W B Bradbury ldquoTransport of lead-203 atthe blood-brain barrier during short cerebrovascular perfusionwith saline in the ratrdquo Journal of Neurochemistry vol 54 no 3pp 905ndash914 1990
[35] R S Diaz and J Monreal ldquoProtein-independent lead perme-ation through myelin lipid liposomesrdquo Molecular Pharmacol-ogy vol 47 no 4 pp 766ndash771 1995
[36] L E Kerper and P M Hinkle ldquoCellular uptake of lead isactivated by depletion of intracellular calcium storesrdquo Journalof Biological Chemistry vol 272 no 13 pp 8346ndash8352 1997
[37] L E Kerper and P M Hinkle ldquoLead uptake in brain capillaryendothelial cells activation by calcium store depletionrdquoToxicol-ogy and Applied Pharmacology vol 146 no 1 pp 127ndash133 1997
[38] M E Legare R Barhoumi E Hebert G R Bratton R CBurghardt and E Tiffany- Castiglioni ldquoAnalysis of Pb2+ entryinto cultured astrogliardquo Toxicological Sciences vol 46 no 1 pp90ndash100 1998
[39] J H Cheong D Bannon L Olivi Y Kim and J BresslerldquoDifferent mechanismsmediate uptake of lead in a rat astroglialcell linerdquoToxicological Sciences vol 77 no 2 pp 334ndash340 2004
[40] T J Evans M R James-Kracke S B Kleiboeker and S WCasteel ldquoLead enters Rcho-1 trophoblastic cells by calciumtransport mechanisms and complexes with cytosolic calcium-binding proteinsrdquo Toxicology and Applied Pharmacology vol186 no 2 pp 77ndash89 2003
[41] G J Kaczorowski R S Slaughter V F King and M LGarcia ldquoInhibitors of sodium-calcium exchange identificationand development of probes of transport activityrdquo Biochimica etBiophysica Acta vol 988 no 2 pp 287ndash302 1989
[42] S L Sensi L M T Canzoniero S P Yu et al ldquoMeasurement ofintracellular free zinc in living cortical neurons routes of entryrdquoJournal of Neuroscience vol 17 no 24 pp 9554ndash9564 1997
[43] R A Colvin ldquoCharacterization of a plasma membrane zinctransporter in rat brainrdquo Neuroscience Letters vol 247 no 2-3pp 147ndash150 1998
[44] V Picard G Govoni N Jabado and P Gross ldquoNramp 2(DCT1DMT1) expressed at the plasma membrane transportsiron and other divalent cations into a calcein-accessible cyto-plasmic poolrdquo Journal of Biological Chemistry vol 275 no 46pp 35738ndash35745 2000
[45] M Okubo K Yamada M Hosoyamada T Shibasaki and HEndou ldquoCadmium transport by human Nramp 2 expressed inXenopus laevis oocytesrdquo Toxicology and Applied Pharmacologyvol 187 no 3 pp 162ndash167 2003
[46] A C Illing A Shawki C L Cunningham and B MackenzieldquoSubstrate profile and metal-ion selectivity of human divalentmetal-ion transporter-1rdquo The Journal of Biological Chemistryvol 287 pp 30485ndash30496 2012
[47] J P Bressler L Olivi J H Cheong Y Kim A Maerten andD Bannon ldquoMetal transporters in intestine and brain theirinvolvement in metal-associated neurotoxicitiesrdquo Human andExperimental Toxicology vol 26 no 3 pp 221ndash229 2007
[48] Z Liu H Li M Soleimani et al ldquoCd2+ versus Zn2+ uptakeby the ZIP8 HCO
3mdashdependent symporter kinetics electro-
genicity and traffickingrdquo Biochemical and Biophysical ResearchCommunications vol 365 no 4 pp 814ndash820 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

ISRN Toxicology 9
respect to toxicityrdquo Journal of Inorganic Biochemistry vol 102no 10 pp 1901ndash1909 2008
[23] MMazzolini S Traverso andCMarchetti ldquoMultiple pathwaysof Pb2+ permeation in rat cerebellar granule neuronesrdquo Journalof Neurochemistry vol 79 no 2 pp 407ndash416 2001
[24] A Esposito M Robello F Pellistri and C Marchetti ldquoTwo-photon analysis of lead accumulation in rat cerebellar granuleneuronsrdquo Neurochemical Research vol 30 no 8 pp 949ndash9542005
[25] C Marchetti C Amico and C Usai ldquoFunctional characteriza-tion of the effect of nimodipine on the calcium current in ratcerebellar granule cellsrdquo Journal of Neurophysiology vol 73 no3 pp 1169ndash1180 1995
[26] A Welling E Bosse A Cavalie et al ldquoStable co-expression ofcalcium channel 1205721120573 and 1205722120575 subunits in a somatic cell linerdquoJournal of Physiology vol 471 pp 749ndash765 1993
[27] A Diaspro S Annunziata M Raimondo P Ramoino and MRobello ldquoA single-pinhole confocal laser scanning microscopefor 3-D imaging of biostructuresrdquo IEEE Engineering inMedicineand Biology Magazine vol 18 no 4 pp 106ndash110 1999
[28] F Pellistri A Cupello A Esposito C Marchetti and MRobello ldquoTwo-photon imaging of calcium accumulation in ratcerebellar granule cellsrdquo NeuroReport vol 15 no 1 pp 83ndash872004
[29] M Cataldi A Secondo A DrsquoAlessio et al ldquoStudies onmaitotoxin-induced intracellular Ca2+ elevation in chinesehamster ovary cells stably transfected with cDNAs encodingfor L-type Ca2+ channel subunitsrdquo Journal of Pharmacology andExperimental Therapeutics vol 290 no 2 pp 725ndash730 1999
[30] A Galan A Troyano N E Vilaboa C Fernandez E DeBlas and P Aller ldquoModulation of the stress response duringapoptosis and necrosis induction in cadmium-treated U-937human promonocytic cellsrdquo Biochimica et Biophysica Acta vol1538 no 1 pp 38ndash46 2001
[31] A H B Poliandri J P Cabilla M O Velardez C C ABodo and B H Duvilanski ldquoCadmium induces apoptosis inanterior pituitary cells that can be reversed by treatment withantioxidantsrdquo Toxicology and Applied Pharmacology vol 190no 1 pp 17ndash24 2003
[32] J L Tomsig and J B Suszkiw ldquoPermeation of Pb2+ throughcalcium channels fura-2 measurements of voltage- anddihydropyridine-sensitive Pb2+ entry in isolated bovinechromaffin cellsrdquo Biochimica et Biophysica Acta vol 1069 no2 pp 197ndash200 1991
[33] EM Leslie J Liu C D Klaassen andM PWaalkes ldquoAcquiredcadmium resistance in metallothionein-III(minusminus) knockoutcells role of the T-type calcium channel Cacn1205721G in cadmiumuptakerdquo Molecular Pharmacology vol 69 no 2 pp 629ndash6392006
[34] R Deane and M W B Bradbury ldquoTransport of lead-203 atthe blood-brain barrier during short cerebrovascular perfusionwith saline in the ratrdquo Journal of Neurochemistry vol 54 no 3pp 905ndash914 1990
[35] R S Diaz and J Monreal ldquoProtein-independent lead perme-ation through myelin lipid liposomesrdquo Molecular Pharmacol-ogy vol 47 no 4 pp 766ndash771 1995
[36] L E Kerper and P M Hinkle ldquoCellular uptake of lead isactivated by depletion of intracellular calcium storesrdquo Journalof Biological Chemistry vol 272 no 13 pp 8346ndash8352 1997
[37] L E Kerper and P M Hinkle ldquoLead uptake in brain capillaryendothelial cells activation by calcium store depletionrdquoToxicol-ogy and Applied Pharmacology vol 146 no 1 pp 127ndash133 1997
[38] M E Legare R Barhoumi E Hebert G R Bratton R CBurghardt and E Tiffany- Castiglioni ldquoAnalysis of Pb2+ entryinto cultured astrogliardquo Toxicological Sciences vol 46 no 1 pp90ndash100 1998
[39] J H Cheong D Bannon L Olivi Y Kim and J BresslerldquoDifferent mechanismsmediate uptake of lead in a rat astroglialcell linerdquoToxicological Sciences vol 77 no 2 pp 334ndash340 2004
[40] T J Evans M R James-Kracke S B Kleiboeker and S WCasteel ldquoLead enters Rcho-1 trophoblastic cells by calciumtransport mechanisms and complexes with cytosolic calcium-binding proteinsrdquo Toxicology and Applied Pharmacology vol186 no 2 pp 77ndash89 2003
[41] G J Kaczorowski R S Slaughter V F King and M LGarcia ldquoInhibitors of sodium-calcium exchange identificationand development of probes of transport activityrdquo Biochimica etBiophysica Acta vol 988 no 2 pp 287ndash302 1989
[42] S L Sensi L M T Canzoniero S P Yu et al ldquoMeasurement ofintracellular free zinc in living cortical neurons routes of entryrdquoJournal of Neuroscience vol 17 no 24 pp 9554ndash9564 1997
[43] R A Colvin ldquoCharacterization of a plasma membrane zinctransporter in rat brainrdquo Neuroscience Letters vol 247 no 2-3pp 147ndash150 1998
[44] V Picard G Govoni N Jabado and P Gross ldquoNramp 2(DCT1DMT1) expressed at the plasma membrane transportsiron and other divalent cations into a calcein-accessible cyto-plasmic poolrdquo Journal of Biological Chemistry vol 275 no 46pp 35738ndash35745 2000
[45] M Okubo K Yamada M Hosoyamada T Shibasaki and HEndou ldquoCadmium transport by human Nramp 2 expressed inXenopus laevis oocytesrdquo Toxicology and Applied Pharmacologyvol 187 no 3 pp 162ndash167 2003
[46] A C Illing A Shawki C L Cunningham and B MackenzieldquoSubstrate profile and metal-ion selectivity of human divalentmetal-ion transporter-1rdquo The Journal of Biological Chemistryvol 287 pp 30485ndash30496 2012
[47] J P Bressler L Olivi J H Cheong Y Kim A Maerten andD Bannon ldquoMetal transporters in intestine and brain theirinvolvement in metal-associated neurotoxicitiesrdquo Human andExperimental Toxicology vol 26 no 3 pp 221ndash229 2007
[48] Z Liu H Li M Soleimani et al ldquoCd2+ versus Zn2+ uptakeby the ZIP8 HCO
3mdashdependent symporter kinetics electro-
genicity and traffickingrdquo Biochemical and Biophysical ResearchCommunications vol 365 no 4 pp 814ndash820 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of



![[3H]Nitrendipine-labeled calcium ... · [3H]Nitrendipine-labeled calciumchannelsdiscriminateinorganic ... Drugs that block these voltage-dependent calcium channels ... The subcellular](https://img.pdfslide.us/doc/110x75/5b92cae009d3f2d1448c069e/3hnitrendipine-labeled-calcium-3hnitrendipine-labeled-calciumchannelsdiscriminateinorganic.jpg)

![Enhancement by Hydrogen Peroxide of Calcium Signals in ...downloads.hindawi.com/journals/omcl/2019/1701478.pdf · calcium channels [18]. Calcium ions released from endoly-sosomal](https://img.pdfslide.us/doc/110x75/5f887ebb417cc311147dd18e/enhancement-by-hydrogen-peroxide-of-calcium-signals-in-calcium-channels-18.jpg)