Embed Size (px)
Citation preview

Research ArticleLPS from P gingivalis and Hypoxia IncreasesOxidative Stress in Periodontal Ligament Fibroblasts andContributes to Periodontitis
L Goumllz1 S Memmert1 B Rath-Deschner1 A Jaumlger1 T Appel2 G Baumgarten3
W Goumltz1 and S Frede3
1 Department of Orthodontics Dental Clinic University Hospital of Bonn Welschnonnenstraszlige 17 53111 Bonn Germany2 Center of Dento-Maxillo-Facial Medicine Faculty of Medicine University of Bonn Welschnonnenstraszlige 17 53111 Bonn Germany3 Clinic of Anesthesiology and Intensive Care Medicine University Hospital of Bonn Sigmund-Freud-Straszlige 25 53105 Bonn Germany
Correspondence should be addressed to L Golz linagoelzukbuni-bonnde and S Frede stillafredeukbuni-bonnde
Received 2 July 2014 Accepted 21 August 2014 Published 12 October 2014
Academic Editor Elisabetta Buommino
Copyright copy 2014 L Golz et al This is an open access article distributed under the Creative Commons Attribution License whichpermits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Oxidative stress is characterized by an accumulation of reactive oxygen species (ROS) and plays a key role in the progression ofinflammatory diseases We hypothesize that hypoxic and inflammatory events induce oxidative stress in the periodontal ligament(PDL) by activating NOX4 Human primary PDL fibroblasts were stimulated with lipopolysaccharide from Porphyromonasgingivalis (LPS-PG) a periodontal pathogen bacteriumunder normoxic and hypoxic conditions By quantitative PCR immunoblotimmunostaining and a specific ROS assay we determined the amount of NOX4 ROS and several redox systems Healthy andinflamed periodontal tissues were collected to evaluate NOX4 and redox systems by immunohistochemistryWe found significantlyincreased NOX4 levels after hypoxic or inflammatory stimulation in PDL cells (119875 lt 0001) which was even more pronouncedafter combination of the stimuli This was accompanied by a significant upregulation of ROS and catalase (119875 lt 0001) Howeverprolonged incubation with both stimuli induced a reduction of catalase indicating a collapse of the protective machinery favoringROS increase and the progression of inflammatory oral diseases Analysis of inflamed tissues confirmed our hypothesis Inconclusion we demonstrated that the interplay of NOX4 and redox systems is crucial for ROS formation which plays a pivotalrole during oral diseases
1 Introduction
Oxidative stress describes ametabolic state where the amountof reactive oxygen species (ROS) is strikingly increased abovephysiological levels ROS originate from the incompletereduction of molecular oxygen resulting in the accumulationof oxidants like hydrogen peroxide (H
2O2) or free radicals
like the hyperoxide (superoxide) anion (O2
∙minus) They are con-tinuously generated in cells as byproducts of normal aerobicmetabolic pathways through the oxidative phosphorylationchain Extrinsic factors like radiation hypoxia smokingheat trauma chemotherapy growth factors or cytokinesmayaugment ROS formation [1ndash3] Low or physiological levelsof ROS can activate intracellular signaling pathways relatedto proliferation differentiation and other cellular functions
whereas uncontrolled release may damage DNA lipids orproteins potentially leading to mutagenesis or carcinogenesis[4ndash7] as many cancer cells were shown to have increasedlevels of ROS Thereby ROS are promoting the invasion andprogression of tumors via matrix metalloproteinase (MMP)activation
The primary ROS O2
∙minus is mainly produced bymonoamine oxidases so- called NADPH-oxidase (NOX)proteins at complexes I and III of the electron transportchain [8] NOX proteins are expressed in a variety of tissuesand represent the only known enzyme systemwhose primarybiological function is to produce ROS They catalyze thereduction of oxygen to superoxide using NADPH as anelectron donor [9] These enzymes were first describedin phagocytes where the respiratory burst works as an
Hindawi Publishing CorporationMediators of InflammationVolume 2014 Article ID 986264 13 pageshttpdxdoiorg1011552014986264
2 Mediators of Inflammation
important defense mechanism against bacteria In the lastdecades five NOX enzymes (NOX 1-5) and two dual oxidases(Duox) Duox1 and Duox2 have been discovered which wereshown to be expressed not only in phagocytes but also inepithelial cells adipocytes or cardiac fibroblasts [9 10]
The NADPH-oxidase family member NOX4 originallyRenox was first described in kidney cells where it is highlyexpressed [11] Later on NOX4 was also detected in othercell types such as endothelial cells fibroblasts keratinocytesand osteoclasts [9 11 12] The typically transmembranouslylocated enzyme is upregulated in response to endoplasmaticreticulum stress [13] shear stress [14] hypoxia or ischemia[15 16] In addition cytokines like transforming growthfactor- (TGF-) 1205731 tumor necrosis factor- (TNF-)120572 andinsulin like growth factor- (IGF-) 1 induce NOX4 expression[9 17] Furthermore this enzyme seems to be a part of thecellular oxygen sensingmechanismsThis is supported by thefact that multiple interactions exist between NOX4 hypoxiainducible factor-1 alpha (HIF-1120572) and ROS [18 19] NOX4-derived ROS have been associated with a variety of processesfor example apoptosis insulin signaling cell differentiationand senescence and bone remodeling [9 20 21]
To limit the potentially deleterious effects of ROS severalprotective mechanisms have been evolved In mitochondriametabolic uncoupling is one mechanism to regulate ROSproduction In healthy cells excessive ROS are decomposedby a protective antioxidant machinery According to theircatalyzing function in the ROS detoxification process theseredox enzymes are grouped in different systems or familieslike the superoxide dismutase (SOD) catalase (CAT) theglutathione system or thioredoxin Furthermore there arealso nonenzymatic or small molecules including vitamins (Cand E) pyruvate flavonoids ubiquinone carotinoids andmany plant-derived antioxidants involved in ROS scavenging[6]
Oxidative stress is gaining increasing interest as an impor-tant cofactor in the etiology and pathogenesis of differentoral and dental diseases including pulpal or mucosal inflam-mation tumor development or inflammatory processes likeperiodontitis First evidence for the presence and role of ROSin the periodontium was given by the detection of thesemolecules after respiratory burst by polymorphonuclear lym-phocytes (PMNLs) in periodontal damage [22] Since thenthe implication of ROS and their degrading enzymes SODor CAT in the pathogenesis of periodontal diseases has beensupported by several groups [23ndash26] High ROS levels mayalso be a linking element between obesity diabetes mellitusatherosclerosis and chronic periodontitis [27] In additionthere are many intervention studies investigating antioxi-dants and antioxidant micronutrients in periodontal therapyincluding the use of mouth rinses and toothpastes Someof these studies provided evidence for beneficial effects butthe links between certain antioxidant species and pathogenicmechanisms are still missing [22]This is also the case for theappearance of ROS and redox systems in the healthy humanoral cavity or periodontium
The periodontium consists of the alveolar bone the toothand the gingiva as well as the periodontal ligament (PDL)which connects the tooth and the surrounding alveolar bone
The collagenous structure of the PDL functions as a shock-reabsorbing barrier protecting the tissues against mechan-ical forces [28] These forces may induce local hypoxiaas well as inflammation [29] Studies indicate that peri-odontal pathogenic bacteria like Porphyromonas gingivalis(P gingivalis) trigger the onset and duration of chronicperiodontitis [30] P gingivalis is a gram-negative anaerobicbacterium belonging to the red complex of oral microflorain accordance with their pathogenicity [31] P gingivalis isthe most prominent periodontal pathogen and is knownto be associated with artherosclerosis and cardiovasculardiseases To mimic the microenvironment of the gingivalsulcus which is characterized by low oxygen tension [32]we include hypoxic conditions in our experiments Since Pgingivalis is an anaerobic bacterium the hypoxic gingivalsulci represent an ideal place for the proliferation of thesebacteria Moreover inflammation by itself may contributeto the development of tissue hypoxia due to an increase inoxygen consumption for example by invading immune cells[33]
Resident cells in the periodontium are also involved ininflammatory and immuneprocesses leading to cytokine (ieinterleukin- (IL-) 1120573 or TNF-120572) or chemokine release whichcould further enhance inflammation [34] PDL fibroblasts areresponsible for the maintenance remodeling and repair ofthe extracellular matrix of the PDL Their immune modu-latory capacities were demonstrated in some recent studies[35 36] Local ROS accumulationmay promote the inductionof proinflammatory cytokines with consecutive activation ofmacrophages [37] finally leading to periodontal destructionvia MMPs [38]
In vitro studies evaluating oxidative stress at the cellularlevel have been focused on gingival fibroblasts while PDLcells have rarely been analyzed Under low zinc environmenthuman PDL fibroblasts showed increased oxidative stress[39] Similar results were found after nicotine ethanol ormetal exposure which was abolished after different antioxi-dant treatment [40 41] Stimulation experiments in humangingival fibroblasts (HGFs) support our hypothesis that PDLcells might play a pivotal role during periodontal diseasesThe authors demonstrated that exposure to lipopolysaccha-ride from P gingivalis (LPS-PG) or LPS from Escherichiacoli (LPS-E coli) increased superoxide concentrations [26]the levels and expression of ROS [42] and proinflammatorycytokines in HGFs [43] ROS activation was associated withactivation of the redox-sensitive transcription factor nuclearfactor-kappaB (NF-120581B) [43] which is critically involved inthe regulation of cytokine and MMP expression The impor-tance of NF-120581B was also demonstrated for the interactionbetween NOX4 HIF-1120572 and ROS [18 19]
However evidence for the involvement of PDL cells inbacterial- or hypoxia-induced ROS formation is missingThesame is true for the balance betweenNOX4and redox systemsin the periodontium The objective of the present study wastherefore to define the role of NOX4 and ROS formation inthe PDL especially during inflammatory and hypoxic eventsWe hypothesized that ROS are regulated by hypoxia andinflammatory stimuli in PDL cells and periodontal tissuesdue to an imbalance of NOX4 and redox system activation
Mediators of Inflammation 3
The current study will enhance the knowledge of the roleplayed by oxidative stress in periodontal tissues which mightimprove diagnostic and therapeutic strategies in oral diseases
2 Materials and Methods
21 Isolation and Characterization of PDL Cells Humanperiodontal ligament fibroblasts were harvested from caries-free and periodontally healthy premolars from patients aged12 to 17 years (119899 = 9) which had to be extracted dueto orthodontic reasons Exclusion criteria were acute orchronic inflammation Patients have had nomedication theyreceived local anesthesia during extraction and analgesia afterextraction The approval of the Ethics Committee of theUniversity of Bonn as well as parental and patients allowancewas obtained Cells were harvested as previously described[44] In brief cells were collected from the middle part ofthe tooth root to avoid contamination by cells derived fromthe gingiva and dental germ Using explant technique PDLcells were cultured in Dulbeccorsquos minimal essential medium(DMEM Invitrogen Karlsruhe Germany) supplementedwith 10 fetal bovine serum (FBS Invitrogen) 100 units ofpenicillin and 100mgmL streptomycin (Biochrom BerlinGermany) at 37∘C in a humidified atmosphere of 5 CO
2
Cells from nine different individuals were phenotyped by theanalysis of the known PDL markers (alkaline phosphatase(ALP) osteocalcin (OC) osteopontin (OP) periostin andS100 A4) at 3rd passage [40] After phenotyping three celllines were pooled and used for experiments at 4th or 5thpassage
22 Cell Experiments Cells were seeded (50000 cellswell)on six-well plates and grown to 80 confluence Cellswere cultured under normoxic (21 O
2and 5 CO
2) or
hypoxic conditions (1 O2and 5 CO
2 INCO 153 O
2CO2
incubator Memmert GmbH Schwabach Germany) for 2 48 24 and 48 hours with or without the addition of 1 120583gmLultrapure lipopolysaccharide from Porphyromonas gingivalis(LPS-PG) (Invivogen San Diego USA) Prior to the here-presented study we analyzed the dose-response of PDL cellswith respect to the inducibility of NOX4 and H
2O2release by
LPS-PG treatmentThemaximal inductionwas detected afterstimulation with 1 120583gmL LPS-PG and no further increasewas observed when the LPS-PG concentration was raised
23 Analysis of Gene Expression Cells were lysed with 4Mguanidinthiocyanat (Roth Karlsruhe Germany) and totalRNA was extracted using the acid guanidinium thio-cyanatephenolchloroform extraction method [45] A totalof 1 120583g of RNA was reverse-transcribed into cDNA by the useof 200U ofMoloneymurine leukemia virus (MMLV) reversetranscriptase (Bio-Rad Munich Germany) cDNA expres-sion was detected by real-time PCR (RT-PCR) using specificprimers (18S fwd CGGCTACCACATCCAAGGAA revGCTGGAATTACCGCGGCT and NOX4 fwd TCTGCC-TGTTCATCTGGCTCTCCA rev AGCCAAGAGTGT-TCGGCACATGGGTA) and SYBR Green as fluorescentdye on the ViiA7 detection system (Applied Biosystems
Darmstadt Germany) Two 120583L of cDNA served as templatein a 20120583L reaction mixture containing 10 120583L QuantiTectSYBR Green Master Mix (Qiagen Hilden Germany) Themixture was heated initially to 50∘C for 2min and to 95∘C for15min followed by 40 cycles with denaturation at 95∘C for 15 sand annealing and extension at 60∘C for 1min Amounts ofspecific cDNAwere normalized to the housekeeping gene 18Sand cDNA expression was calculated as relative expression ofthe respective control
24 Immunoblot For protein quantification the Pierce BCAProtein Assay Kit (Thermo Scientific Rockford USA) wasused After addition of 4x sample buffer (50mM Tris pH68 2 (wv) SDS 5 (vv) 2-mercaptoethanol 00125bromophenol blue and 1 glycine) 50 120583g of total cell lysateper lane was subjected to 75 SDS-PAGE and transferredonto a nitrocellulose membrane (02 120583mpore size Schleicheramp Schuell Microscience Dassel Germany) The efficiency ofprotein transfer and equal loading was confirmed by stainingthe membrane with Ponceau S (Sigma Aldrich MunichGermany) Membranes were blocked overnight at 4∘C with5 (wv) nonfat milk in TBS-T (10mM Tris pH 75 100mMNaCl and 005 Tween-20) and washed with TBS-T prior toincubation with a rabbit polyclonal antibody against NOX4(Abcam Cambridge UK) at room temperature for 2 h szlig-Actin (Santa Cruz Biotechnology Santa Cruz USA) servedas marker for equal protein loading After washing with TBS-THRP- (horseradish peroxidase-) conjugated anti-rabbit IgGantibody (NewEnglandBiolabs) was added Immunoreactiveproteins were visualized by chemiluminescence (AmershamChemiluminescence Kit) and exposure to X-ray films (AgfaMortsel Belgium) Densitometrical analysis was performedusing the freely available image-processing software ImageJ143 (httprsbinfonihgovij)
25 Amplex Red Assay For hydrogen peroxide (H2O2)
detection the commercially availableN-acetyl-37-dihydrox-yphenoxazine (Amplex Red) Hydrogen PeroxideperoxidaseAssay Kit (Molecular Probes Eugene OR) was used Thecolorless reagent reacts with hydrogen peroxide (H
2O2)
using horseradish peroxidase (HRP) to catalyze the H2O2-
dependent oxidation of nonfluorescent Amplex Red to flu-orescent resorufin red This assay detects only the releaseof hydrogen peroxide since the size of HRP prevents itfrom entering the mitochondria Therefore PDL cells wereincubated with 500120583L Amplex Red and 01 UmL peroxidasewas added at 37∘C for 30min Absorbance was measuredat 565 nm after 1 2 and 4 hours During the procedureexcitation light intensity was kept to an absolute minimumas recommended by Zhao et al [46]
26 Immunofluorescence For this purpose PDL cells werecultured on cover slips to 60 confluence PDL cells werestained for NOX4 and redox systems under normoxic orhypoxic conditions in the presence and absence of LPS-PGHypoxic conditions were obtained by placing the cells in anincubator where the oxygen concentration can be adjusted
4 Mediators of Inflammation
to 1 O2 balanced with 5 CO
2and N
2in the incuba-
tion atmosphere (INCO 153 O2CO2incubator Memmert
GmbH Schwabach Germany) At the end of the experimentmedium was removed cells were washed three times withPBS and fixed with 250120583L cooled methanolaceton (1 1)at minus20∘C for 10min The solution was removed and cellsremained at room temperature for 10min to dry Afterblocking with 3 BSA-PBS for 10min cells were incubatedwith antibodies (Ab) directed against NOX4 (Abcam) CATor SOD (Abcam) 1 50 in PBS 3 BSA for 2 hours Cellswere washed with PBS before the species-specific secondaryAb (dilution 1 400 in PBS) were added for additional90min Cells were covered on slides with DAPI contain-ing mounting medium (Dako) On the following day cellswere scanned with the Zeiss Axio Imager A1 fluorescencemicroscope (HBO 100 Carl Zeiss Jena Germany) anAxioCam MRc camera (Carl Zeiss) and the AxioVision 47software (Carl Zeiss) Density analysis was performed usingthe freely available image-processing software ImageJ 143(httprsbinfonihgovij) Quantification of relative fluores-cence intensity was performed by measuring the intensity ofeach cell related to cell area To obtain grayscale pictures eachwas processed using the tools ldquocolorrdquo and ldquosplit channelsrdquoin ImageJ which resulted in red green and blue color-fractionated grayscale pictures for each channel Only greenfractionated pictures were used for calculation of the relativefluorescence intensity
27 Immunohistochemistry Periodontal tissues from clini-cally healthy patients (I) as well as those from patientswith the diagnosis of gingivitis (II) and periodontitis (III)were obtained from three different patients for each groupDiagnoses of gingivitis and periodontitis were performedby specialists from the Clinic of Dental Medicine Gin-givitis was characterized by a reversible local inflammatoryreaction of the gingiva visible as redness swelling andbleeding In contrast periodontitis affects four differentstructures gingiva periodontal ligament tooth and alveolarbone Therefore periodontitis was diagnosed by an increaseof probing depth bleeding and attachment loss whichwas supported by radiographs Tissues were fixed in 4phosphate-buffered paraformaldehyde for 24 h followed byhydration and dehydration in an ascending ethanol serieswhich ended in paraffin embedding Afterwards 4 120583m thickparaffin-embedded sections were prepared deparaffinizedrehydrated rinsed with TBS pH 74 for 10min and thensoaked in 70mL methanol containing 700mL of 30 H
2O2
for 45min in the dark to block endogenous peroxidase activ-ity Afterwards tissue sections were rinsed and preincubatedwith TBSBSA 4 for 20min to avoid unspecific backgroundstaining Incubation with monoclonal primary antibodies ofNOX4 (Abcam Cambridge UK) and polyclonal antibody ofcatalase and SOD (Abcam) in a 1 100 working solution ofTBSBSA was performed at 4∘C overnight in a humidifiedchamber Subsequently sections were washed in TBS andincubated with a peroxidase-labeled polymer conjugated toa goat anti-rabbit immunoglobulin provided as a ready-to-use solution (Envision Dako AS Glostrup Denmark)
for 30min in a humidified chamber at room temperatureAntibody complexeswere visualized using diaminobenzidinefor 10min resulting in brown staining Thereafter the slideswere rinsed counterstainedwithMayerrsquos hematoxylin rinsedagain and covered Negative controls were prepared by omis-sion of the primary antibodies as well as both the primaryand secondary antibody from the staining procedures usingTBSBSA instead Finally tissue sections were analyzed usingtheZeissAxioScop 2microscope (Carl Zeiss JenaGermany)
28 Statistical Analysis All experiments were performedin triplicate and mRNA quantification is given as meansplusmn SD Results were compared by ANOVA Dunnettrsquos andTukey-Kramer tests were performed as posttest to calculatewhether differences between treated and control groups weresignificant A 119875 value of 005 was considered as significant
3 Results
31 NOX4 Human primary PDL cells were incubated undernormoxic and hypoxic conditions in the presence or absenceof LPS-PG (1 120583gmL) Treatment with LPS-PG induced time-dependently a significant increase in the mRNA expressionof NOX4 compared to the untreated control (Figure 1(a)) Indetail NOX4mRNA levels weremarkedly increased after twohours of LPS-PG stimulation the increase reached significantlevels (4-fold) after 4 hours (119875 lt 0001) NOX4 mRNA levelspeaked after 8-hour stimulation with LPS-PG and returnedto base line levels afterwards
Hypoxia alone induced the expression of NOX4 in PDLcells but in contrast to the stimulation with LPS-PG themaximum in NOX4 mRNA upregulation was detected after4 hours (119875 lt 0001 Figure 1(a)) Subsequently NOX4mRNA expression decreased however it remained at ahigher level compared to the control group even after 24or 48 hours (Figure 1(a)) The highest increase in NOX4mRNA expression was detected after the combination ofboth stimuli (85 times) which was statistically significantcompared to control and even compared to LPS stimulationunder normoxia (119875 lt 0001 Figure 1(a))
To prove whether the increase in mRNA expression wasfollowed by an increase in protein content immunofluo-rescence stainings for NOX4 were performed NOX4 wasdetectable predominantly in the cytosol and to a slight extentin the nucleus of unstimulated PDL cells (Figure 1(b)) To getmore detailed information about the time course of NOX4protein induction after hypoxic and inflammatory stimula-tion we performed immunoblots After 2 hours only hypoxiaprovoked a significant increase in NOX4 protein (119875 lt 0001)which diminished afterwards (Figure 1(c)) Under normoxiccondition the treatment with LPS-PG induced a significantelevation of NOX4 protein after 4 hours of incubation com-pared to control and hypoxia alone (119875 lt 0001 Figure 1(c))The LPS-PG effect was evenmore pronounced under hypoxiccondition which was statistically significant compared to allthe other groups (119875 lt 0001 Figure 1(c)) After 24 hours ofincubation alterations of NOX4 protein levels were no longerdetectable (data not shown) These findings were in line with
Mediators of Inflammation 5
NADPH-oxidase 4 mRNA
125
100
75
50
25
00minus
minus
minus minus minus+
+ minus + minus + minus + minus + minus + minus + minus + minus + minus +
+ minus minus+ + minus minus+ + minus minus+ + + +
lowastlowast
lowast
lowast
2h 4h 8h 24h 48h
HoxLPS
(a) (b)
(c)
mRN
A ex
pres
sion
com
pare
d to
cont
rol
minusminus
minus minus
+
+
+minusminus
minus minusminus
+
+ +
+
lowast
lowast
Hox
NOX4 protein
Inte
grat
ed d
ensit
y (
) com
pare
dto
cont
rol (
au)
LPS
2h 4h
300
200
100100
50
0
Nox Hoxminus minus+ +
120573-Actin
NOX4
LPS
120573-Actin
NOX4
2h
4h
Figure 1 NOX4 in human primary periodontal ligament (PDL) cells (a) PDL cells were cultured under normoxic or hypoxic condition andstimulated with or without LPS of Porphyromonas gingivalis (1 120583gmL) NOX4 mRNA expression was analyzed after 2 4 8 24 and 48 hStatistical differences were analyzed by one-way ANOVA and post hoc Dunnett and Tukeyrsquos multiple comparison test 119875 lt 005 differenceto control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 9) (b) NOX4 protein was visualized by immunofluorescence as well asimmunohistochemical staining in unstimulated PDL fibroblasts (1 50 antibody from Abcam) (c) Immunoblot data of NOX4 in PDL cellscultured under normoxic or hypoxic condition (Hox) and stimulated with or without LPS of Porphyromonas gingivalis (1 120583gmL) NOX4protein levels were analyzed after 2 and 4 h Statistical analysis of western blot data was performed using the freely available image-processingsoftware ImageJ 143 (httprsbinfonihgovij) Statistical differences were analyzed by one-way ANOVA and post hoc Dunnett and Tukeyrsquosmultiple comparison test 119875 lt 005 difference to control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 3) The scale bars indicate200 120583m in the bright field survey and 50120583m in the immunofluorescence image respectively
our mRNA results even though we detected time-dependentdifferences
32 H2O2Formation To determine hydrogen peroxide gen-
eration we used the commercially available Amplex Red-horseradish peroxidase assay PDL cells displayed a lowconstitutive H
2O2production under normoxic conditions
We observed a significant upregulation of the H2O2con-
centration in the cell culture supernatant after 1 2 and 4hours of stimulation with LPS-PG Hypoxia alone induceda significant increase in H
2O2formation compared to the
time-matched controls (Figure 2) While in the LPS groupthe H
2O2values were the highest after 1 hour and decreased
continuously after 2 and 4 hours (Figure 2) under hypoxiccondition H
2O2increased permanently (Figure 2) Compar-
ing the LPS and the hypoxic effect revealed that hypoxiaalone induced a significantly higher H
2O2accumulation over
the complete observation period (119875 lt 0001) Howeverthe combination of both stimuli elicited the highest H
2O2
concentration after 1 hour which was significant not onlycompared to control but also compared to the LPS and thehypoxia group (Figure 2) After 2 hours the treatment withLPS-PG was no longer able to enhance the hypoxia-inducedH2O2formation it was still significantly increased compared
to control and the LPS stimulation but no longer comparedto the hypoxic groupThis was even more pronounced after 4hours
6 Mediators of Inflammation
2h 4h1h
50000
40000
30000
20000
10000
0
(120583M
)
HoxLPS
minus minus minus minus minus minus
minus
+
+minus +minus +minus +minus +minus +
++ + + +
lowast
lowastlowast lowast
lowast
lowast
H2O2 concentration
Figure 2 Determination of ROS formation (H2O2) using Amplex
Red assay PDL cells were cultured under normoxic (Nox) orhypoxic condition (Hox) and stimulated with or without LPS ofPorphyromonas gingivalis (1 120583gmL) H
2O2was analyzed after 1
2 and 4 hours Statistical differences were analyzed by one-wayANOVA and post hoc Dunnett and Tukeyrsquos multiple comparisontest 119875 lt 005 difference to control lowast119875 lt 005 difference betweengroups (means plusmn SD 119899 = 6)
33 Redox Systems To analyze which protectivemechanismsagainst ROS formation are involved in our setting wedetermined the redox system components catalase (CAT)and superoxide dismutase (SOD) in PDL cells after treat-ment with LPS-PG under normoxic and hypoxic conditionsby immunofluorescence (Figures 3(a)-3(b)) In additionperiodontal tissues from clinically healthy patients (I) aswell as from patients with the diagnosis of gingivitis (II)and periodontitis (III) were obtained from three differentpatients for each group and stained immunohistochemicallyfor NOX4 CAT and SOD (Figures 4 5 and 6)
331 Immunofluorescence of Redox Systems in PDL CellsPDL cells displayed a constitutive cytosolic catalase expres-sion under normoxic conditions Treatment with LPS-PGandhypoxia induced a change in PDL cellmorphologywhichwas first obvious after a 1-hour incubation under hypoxicconditions and after 2 hours with LPS-PG (Figure 3(a)) Asshown in Figure 3(b) analyzing catalase cytoplasm densityin relation to constant cell area revealed a significant upreg-ulation of catalase in PDL cells stimulated with LPS-PG orhypoxia after 1 h (119875 lt 0001 Figure 3(b)) This was also truefor hypoxic conditions after 2 h compared to all other groupsUnder inflammatory stimulation (LPS) a significant increaseof catalase cytoplasm density was detectable after 4 hourscompared to all the other groups (119875 lt 0001 Figure 3(b))Instead the combination of both stimuli diminished thehypoxic as well as LPS-induced effects after 1 2 and 4hours Determining superoxide dismutase protein expressionin PDL cells showed no significant differences at any timepoint (119875 gt 005 data not shown)
332 Immunohistochemistry of NOX4 and Redox Systemsin Periodontal Tissue Sections Periodontal tissues (healthygingiva and periodontal ligament (PDL) gingivitis andperiodontitis) were obtained from three different patients foreach group
NOX4 In the healthy periodontal tissue samples only veryweak immunostaining for NOX4 could be obtained (Figures4(a) and 4(b)) In the PDL weak immunoreactivity witha moderate staining of some fibroblasts was observed Ininflamed tissues from patients suffering from gingivitis orperiodontitis strong immunoreactivity for NOX4 appearedin the cytosol and even stronger in the nuclei of keratinocytesof the basal layer of inflamed gingival epithelium (Figures4(c)-4(d)) In gingival and PDL fibroblasts as well as inimmune cells like leukocytes and in vessels the immunore-activity was primarily restricted to the cytosol and cell walls(Figures 4(c)-4(d) see red arrows)
CAT Nearly no immunostaining could be found in thehealthy gingival epithelium (Figure 5(a)) In the healthyPDL predominantly PDL cells located near to the toothroot surfaces were stained (Figure 5(b) see red arrow) Ingingivitis tissue a weak staining of gingival keratinocytessimilar to healthy tissue could be seen (Figure 5(c)) Insteadin the tissue samples of patients with periodontitis a strongimmunoreactivity was observed in the subepithelial layerprobably referring to nuclei staining of immune cells likeleukocytes (Figure 5(d) see red arrows)
SOD As for healthy tissues gingival epithelium showeda moderate to strong immunoreactivity of keratinocyteswhich was primarily limited to the cytosol (Figure 6(a)see red arrow) In healthy periodontium gingival and PDLfibroblasts were weakly stained (Figure 6(b)) Compared toinflamed tissues basal epithelial cells seemed to be stainedmore intensively in the gingivitis specimen (Figures 6(c)-6(d)) In addition the subepithelium revealed immunore-activity in local cells like gingival fibroblasts and scatteredimmune cells (Figure 6(c)) In periodontitis stained sectionsSOD immunostaining seemed to be limited to the gingivalepithelium (Figure 6(d) see red arrows) In inflamed tissuessubepithelial layers showed no signs of immunoreactivity
4 Discussion
Oxidative stress is discussed as an important cofactor in theetiology and pathogenesis of several oral and dental diseasesfor example in inflammatory processes like periodontitisFirst evidence for the presence and role of ROS in theperiodontium was given by the detection of these moleculesafter respiratory burst by polymorphonuclear lymphocytes(PMNLs) in periodontal damage [22] Since then the impli-cation of ROS and their degrading enzymes SOD or CAT inthe pathogenesis of periodontal diseases has been supportedby several groups [23ndash26] To analyse whether cellular redoxsystems in the periodontium are affected by inflammatoryconditions we investigated the effect of hypoxia and LPSfrom Porphyromonas gingivalis (LPS-PG) on periodontal
Mediators of Inflammation 7
1h 2hCatalase
Nox
Nox + LPS
Hox
Hox + LPS
4h
100120583m 100120583m 100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
(a)
2h 4h
Hox
Inte
grat
ed d
ensit
y (a
u)
Catalase cytoplasm density
LPSminusminus + + minusminus + + minusminus +
minus +minus +minus +minus +minus +minus +
+
lowast
lowast
lowast
lowast
lowastlowast
1h
20000
25000
15000
1000010000
7500
5000
2500
0
(b)
Figure 3 Catalase (CAT) in PDL cells (a) CAT was visualized by immunofluorescence staining (1 50 antibody from Abcam) PDL cellswere cultured under normoxic (Nox) or hypoxic condition (Hox) and stimulated with or without LPS of Porphyromonas gingivalis (1120583gmL)Catalase cytoplasm density (b) was determined in relation to cell area using the freely available image-processing software ImageJ 143(httprsbinfonihgovij) Statistical analysis of immunofluorescence data was analyzed by one-way ANOVA and post hoc Dunnett andTukeyrsquos multiple comparison test 119875 lt 005 difference to control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 6) The scale barsindicate 100 120583m in the immunofluorescence images
8 Mediators of Inflammation
50120583m
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m
50120583m
50120583m
Inflamed
(d)
Figure 4 NADPH-oxidase 4 (NOX4) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Monoclonal primary antibody against NOX4 (Abcam Cambridge UK) was used ina concentration of 1 100 In gingival and PDL fibroblasts as well as in immune cells like leukocytes and in vessels the immunoreactivity wasprimarily restricted to the cytosol and cell walls Red arrows indicate examples for the typical distribution (Figures 4(c) and 4(d)) The scalebars indicate 200120583m in the surveys and 50 120583m in the higher magnifications respectively
ligament cells We showed that both stimuli induced asignificant increase of NOX4 and a significant upregulationof hydrogen peroxide generation NOX enzymes were firstdescribed in phagocytes as being responsible for defensemechanisms against pathogens using NOX-generated ROSmolecules Till then NOX4 expression has been detected innonphagocyte cell types like kidney cells osteoclasts andosteoblasts as well as dermal cells [9 11 47] In PDL cellshypoxia and LPS-PG induced a significant increase of NOX4mRNA The protein expression was mainly located in thecytosol but likewise in the nucleus Our findings were inline with Brown and Griendling who have shown that thetypically transmembrane molecule is localized in differentcell compartments [9]
NOX4 is one of the major sources of ROS generationwhich is highly and ubiquitously expressed compared tothe other family members Mandal and colleagues demon-strated that apart from its antimicrobial function NOX4 isresponsible not only for osteoclast but also for preosteoblastdifferentiation via ROS accumulation reinforcing the ideaof NOX4 as a key regulator in bone remodeling underphysiological ROS amounts [47] Animal studies supportthe hypothesis that massive ROS formation induced bone
resorption via NOX4 [20 21] Whereas physiological levelsof NOX4-generated ROS in PDL cellsmight facilitate alveolarbone remodeling massive ROS accumulation during inflam-matory periodontal diseases might lead to alveolar bonedestruction favoring tooth loss We have demonstrated firstthat NOX4 is upregulated in periodontal ligament fibroblastsduring bacterial and hypoxic events Hence PDL fibroblastsmay be a source of excessive ROS generation and are thereforeof particular importance during periodontal diseases
Our results support current findings determining the roleof oxidative stress in different periodontal pathologies whichimplicate that the balance between NADPH-oxidases andredox systems is crucial to maintain cell and tissue home-ostasis [22ndash26] However there are only a few studies usinghuman periodontal fibroblast in the context of oxidativestress and none of them evaluated the role of NOX4 andcounteracting redox systems during inflammatory conditionswith LPS-PG stimulation and hypoxia An increase of ROSgeneration was detected by San Miguel et al following PDLfibroblast stimulation with nicotine or ethanol which wereabolished after antioxidant treatment [40] They completedtheir results using metal ions like nickel for PDL stimulationwhich increases ROS activity in these cells indicating a role
Mediators of Inflammation 9
200120583m
50120583m
50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 5 Catalase (CAT) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontal ligament (b) andsamples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University of Bonn and parentalas well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against catalase (Abcam Cambridge UK) was used in a concentration of1 100 No immunostaining could be found in the healthy gingival epithelium In the healthy PDL predominantly PDL cells located near tothe tooth root surfaces were stained (Figure 5(b) red arrow) In gingivitis tissue a weak staining of gingival keratinocytes similar to healthytissue could be seen (Figure 5(c)) Instead in the tissue samples of patients with periodontitis a strong immunoreactivity was observed inthe subepithelial layer probably referring to nuclei staining of immune cells like leukocytes Red arrows indicate examples for the typicallocalisation (Figure 5(d)) The scale bars indicate 200 120583m in the surveys and 50120583m in the higher magnifications respectively
in metal cytotoxicity or even nickel allergy [41] Transientmetal ions like nickel and cobalt are known to mimic cellularhypoxia mainly due to the inhibition of prolyl hydrolases(PDHs) and asparaginyl hydrolase (factor-inhibiting HIF(FIH)) The inhibition of hydroxylation leads to HIF-1120572nuclear translocation followed by the transcription of diversegenes which are implicated in hypoxic adaptation immuneresponse or angiogenesis therefore indirectly supporting ourresults under hypoxic condition [48 49]
We could show that hypoxia as well as LPS-PG inducedROS formation by detecting H
2O2using the commercially
available Amplex Red assay The high levels of H2O2released
under hypoxic conditions after 1 and 2 hours in PDL cellsmay be caused by different mechanisms First we founda significant upregulation of the mRNA and protein ofthe H
2O2-generating enzyme NOX4 Second the lack of
molecular oxygen as the final electron acceptor in therespiratory chain may be responsible for the early release of
ROS LPS-PG alone induced acutely a small but significantincrease in H
2O2release and enhanced the hypoxia-induced
generation In human gingival fibroblasts similar results wereobtained showing the increase of ROS and antioxidants afterP gingivalis or LPS stimulation [26 42 43] Interestinglydespite the increase in NOX4 mRNA and protein expressionafter 4 hours of stimulation with LPS-PG under hypoxicconditions the H
2O2generation was significantly reduced
when compared to hypoxia alone An additional inhibitionof the respiratory chain by LPS-PG which was described forcardiac mitochondria [50] is therefore conceivable
To antagonize oxidative stress cells developed severalantioxidative enzyme systems Therefore we measured SODand CAT expression in PDL cells and periodontal tissues byimmunofluorescence and immunohistochemistry SODs aremetalloenzymes which are responsible for the dismutation ofsuperoxide radicals to hydrogen peroxide [51] The reductionof hydrogen peroxide to water and oxygen is subsequently
10 Mediators of Inflammation
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m 50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 6 Superoxide dismutase (SOD) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against SOD (Abcam Cambridge UK) was used ina concentration of 1 100 Gingival epithelium showed a moderate to strong staining of SOD in keratinocytes which was primarily limitedto the cytosol (Figure 6(a) and red arrow depicts the typical localization) In the healthy periodontium gingival and PDL fibroblasts wereweakly stained (Figure 6(b)) In inflamed tissues basal epithelial cells seemed to be stained more intensively in the gingivitis specimen thanin periodontitis (Figures 6(c) and 6(d)) In addition the subepithelium revealed immunoreactivity in local cells like gingival fibroblasts andscattered immune cells (Figure 6(c) see red arrow) In periodontitis SOD immunostaining seemed to be limited to the gingival epithelium(Figure 6(d) see red arrow) The scale bars indicate 200 120583m in the surveys and 50 120583m in the higher magnifications respectively
catalyzed by CATs which are ubiquitously expressed Bothenzymes are essential to maintain cellular redox homeostasisThe present data imply that PDL cells and periodontal tissueshave fundamental defense machineries against bacteria rep-resented by NOX4-dependent inducible ROS generation aswell as protective redox systems This is of great importanceconcerning the high amounts and diversity of the oral floraDifferent studies indicate that redox systems are key playersin the development of periodontal diseases and markers forperiodontal health status [52ndash56] We detected a significantincrease of catalase in PDL cells in response to LPS aswell as hypoxia after 1 and 2 hours In our setting PDLfibroblasts seem to counteracted LPS- and hypoxia-inducedROS formation with the elevation of CAT expression Butprolonged incubation under hypoxic conditions failed toincrease catalase Since ROS generation was still elevated atthese later time points a delay in ROS degradation may be
the explanation Taken together the dramatic elevation inROS formation by the combination of hypoxia and LPS-PGon one hand and the reduction of catalase under the sameconditions on the other hand increase the oxidative stress inthe periodontium favoring the progression of inflammationand bone resorption and finally may lead to tooth loss
The fundamental role of the balance between ROSgeneration and antioxidative mechanisms in periodontalinflammatory processes was supported by our immunohis-tochemical findings In patients suffering from periodontalinflammatory diseases like gingivitis and periodontitis wedemonstrated that NOX4 is upregulated and thus likelyresponsible for H
2O2generation However we detected as
well an increase in catalase abundance in inflamed gingivaland periodontal tissue which was in line with data fromKrifka and colleagues who detected high CAT levels inmonomer-exposed macrophages as a response to oxidative
Mediators of Inflammation 11
stress [57] In our in vitro setting PDL fibroblasts seemto counteract LPS- and hypoxia-induced ROS formationpredominantly with an elevation of CAT expression sincewe could not detect any SOD immunoreactivity The roleand impact of the antioxidative systems SOD and CAT arecontroversially discussed Recent studies measured CAT andSOD activities in gingival tissues of patients with periodon-titis [58 59] Borges Jr et al did not find differences inCAT activities comparing healthy and diseased patients [58]Instead Tonguc et al detected decreased local SOD andCAT activities in periodontitis patients which seemed to beincreased by smoking [59] In contrast in inflamed tissues weobservedhigh amounts of SODwhichwas localized primarilyin epithelial cells Jacoby and Davis determined SOD in peri-odontal soft tissues They detected twice as much SOD activ-ity in the periodontal ligament as in the human skin (dermis)[60] Furthermore immunohistochemical analysis revealedthat SOD is localized in the periphery of matrix collagenfibrils and to the glycocalyx of tissue fibroblasts This wasconfirmed by Skaleric et al [42] which might explain the factthat we did not detect significant differences of SOD levelsafter hypoxic and LPS-PG stimulation intracellularly in PDLfibroblasts Another reason for these results may be causedby the age of the PDL cells used in this study We explantedthe PDL cells from young and oral healthy patients but mostof the immunohistochemical stainings were performed intissues from elderly patients with clearly diagnosed gingivitisor periodontitisTherefore it is possible that in PDL cells fromelderly patients a different expression pattern of antioxidantenzyme systems could be found which further deterioratethe redox state of the periodontal ligament Moreover otherauthors found reduced antioxidants in the body fluids ofpatients with periodontitis indicating a dysregulation whichwould favor disease progression [23 24 61] A variety of invitro and animal studies analyzed the effect of antioxidantsupplement in the context of inflammatory or cytotoxicevents [57 62 63] They have shown that antioxidantsinhibit these processes indicating a therapeutic approachin pathologies like periodontitis First clinical investigationsconfirmed the efficiency of antioxidant micronutrients andadjuncts in periodontal diseases [22 54 56 64 65] Thepresent data indirectly support these studies
Concerning periodontal pathogen invasiveness and viru-lence there is compelling evidence that P gingivalis possessesits own protective antioxidants for example rubrerythrindefending them against the oxidative burst of the host [66]We have shown that its virulence factor LPS-PG inducedhigh amounts of ROS via NOX4 which may result in boneresorption favoring the invasiveness of the pathogen whichis at the same time protected by its own defense machineryThis would further explain the pathogenicity and resistanceof this bacterium strain in patients with periodontitis
5 Conclusion
In this study we have demonstrated that LPS from Porphy-romonas gingivalis (LPS-PG) as well as hypoxia induces aNOX4-dependent increase inH
2O2release in PDLfibroblasts
which may contribute to the development and progressionof periodontal diseases when not balanced by a concomitantincrease in antioxidative systems The dramatic elevationin ROS formation induced by the combination of hypoxiaand LPS-PG and the reduction of catalase under the sameconditions increased the oxidative stress in the periodontiumfavoring the progression of inflammation bone resorptionand possibly tooth loss
Taken together the presented data further explain thedestructive mechanism of hypoxic and inflammatory condi-tions in the periodontium and may improve diagnostic andtherapeutic strategies in periodontal diseases
Conflict of Interests
The authors have no conflict of interests to disclose
Acknowledgments
The authors would like to thank Ms Tina Schaffrath for hertechnical supportThe present studywas supported by a grantfrom the German Research Foundation (Clinical ResearchUnit 208TP7) and the Medical Faculty of the University ofBonn
References
[1] S Reuter S C Gupta M M Chaturvedi and B B AggarwalldquoOxidative stress inflammation and cancer how are theylinkedrdquo Free Radical Biology and Medicine vol 49 no 11 pp1603ndash1616 2010
[2] H Sone H Akanuma and T Fukuda ldquoOxygenomics inenvironmental stressrdquo Redox Report vol 15 no 3 pp 98ndash1142010
[3] K E Wellen and C B Thompson ldquoCellular metabolic stressconsidering how cells respond to nutrient excessrdquo MolecularCell vol 40 no 2 pp 323ndash332 2010
[4] R O Poyton K A Ball and P R Castello ldquoMitochondrialgeneration of free radicals and hypoxic signalingrdquo Trends inEndocrinology andMetabolism vol 20 no 7 pp 332ndash340 2009
[5] F Weinberg and N S Chandel ldquoReactive oxygen species-dependent signaling regulates cancerrdquo Cellular and MolecularLife Sciences vol 66 no 23 pp 3663ndash3673 2009
[6] A Cataldi ldquoCell responses to oxidative stressorsrdquo CurrentPharmaceutical Design vol 16 no 12 pp 1387ndash1395 2010
[7] Z Durackova ldquoSome current insights into oxidative stressrdquoPhysiological Research vol 59 no 4 pp 459ndash469 2010
[8] L A Sena and N S Chandel ldquoPhysiological roles of mitochon-drial reactive oxygen speciesrdquoMolecular Cell vol 48 no 2 pp158ndash166 2012
[9] D I Brown and K K Griendling ldquoNox proteins in signaltransductionrdquo Free Radical Biology amp Medicine vol 47 no 9pp 1239ndash1253 2009
[10] K Bedard and K-H Krause ldquoThe NOX family of ROS-generatingNADPHoxidases physiology and pathophysiologyrdquoPhysiological Reviews vol 87 no 1 pp 245ndash313 2007
[11] K-H Krause ldquoTissue distribution and putative physiologicalfunction of NOX family NADPH oxidasesrdquo Japanese Journal ofInfectious Diseases vol 57 no 5 pp S28ndashS29 2004
12 Mediators of Inflammation
[12] S Yang Y Zhang W Ries and L Key ldquoExpression of Nox4 inosteoclastsrdquo Journal of Cellular Biochemistry vol 92 no 2 pp238ndash248 2004
[13] E Pedruzzi C Guichard V Ollivier et al ldquoNAD(P)H oxidaseNox-4mediates 7-ketocholesterol-induced endoplasmic reticu-lum stress and apoptosis in human aortic smooth muscle cellsrdquoMolecular and Cellular Biology vol 24 no 24 pp 10703ndash107172004
[14] J Hwang A Saha Y C Boo et al ldquoOscillatory shear stress stim-ulates endothelial production of O
2-from p47119901ℎ119900119909-dependent
NAD(P)Hoxidases leading tomonocyte adhesionrdquoThe Journalof Biological Chemistry vol 278 no 47 pp 47291ndash47298 2003
[15] H B Suliman M Ali and C A Piantadosi ldquoSuperoxidedismutase-3 promotes full expression of the EPO response tohypoxiardquo Blood vol 104 no 1 pp 43ndash50 2004
[16] P Vallet Y Charnay K Steger et al ldquoNeuronal expressionof the NADPH oxidase NOX4 and its regulation in mouseexperimental brain ischemiardquo Neuroscience vol 132 no 2 pp233ndash238 2005
[17] K T Moe S Aulia F Jiang et al ldquoDifferential upregulation ofNox homologues of NADPH oxidase by tumor necrosis factor-120572 in human aortic smooth muscle and embryonic kidney cellsrdquoJournal of Cellular andMolecularMedicine vol 10 no 1 pp 231ndash239 2006
[18] I Diebold A Petry J Hess and A Gorlach ldquoThe NADPHoxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1rdquoMolecular Biology of the Cell vol 21 no 12pp 2087ndash2096 2010
[19] K V Tormos and N S Chandel ldquoInter-connection betweenmitochondria and HIFsrdquo Journal of Cellular and MolecularMedicine vol 14 no 4 pp 795ndash804 2010
[20] T Ohnishi K Bandow K Kakimoto M Machigashira TMatsuyama and T Matsuguchi ldquoOxidative stress causes alve-olar bone loss in metabolic syndrome model mice with type 2diabetesrdquo Journal of Periodontal Research vol 44 no 1 pp 43ndash51 2009
[21] C Goettsch A Babelova O Trummer et al ldquoNADPH oxidase4 limits bone mass by promoting osteoclastogenesisrdquo Journal ofClinical Investigation vol 123 no 11 pp 4731ndash4738 2013
[22] I L C Chapple and J B Matthews ldquoThe role of reactive oxygenand antioxidant species in periodontal tissue destructionrdquoPeriodontology 2000 vol 43 no 1 pp 160ndash232 2007
[23] A Guentsch P M Preshaw S Bremer-Streck G KlingerE Glockmann and B W Sigusch ldquoLipid peroxidation andantioxidant activity in saliva of periodontitis patients effect ofsmoking and periodontal treatmentrdquo Clinical Oral Investiga-tions vol 12 no 4 pp 345ndash352 2008
[24] R Agnihotri P Pandurang S U Kamath et al ldquoAssociation ofcigarette smoking with superoxide dismutase enzyme levels insubjected with chonic periodontitisrdquo Journal of Periodontologyvol 80 no 4 pp 657ndash662 2009
[25] D Ekuni T Tomofuji N Tamaki et al ldquoMechanical stimulationof gingiva reduces plasma 8-OHdG level in rat periodontitisrdquoArchives of Oral Biology vol 53 no 4 pp 324ndash329 2008
[26] H Staudte A Guntsch A Volpel and B W Sigusch ldquoVitaminC attenuates the cytotoxic effects of Porphyromonas gingivalison human gingival fibroblastsrdquo Archives of Oral Biology vol 55no 1 pp 40ndash45 2010
[27] P Bullon H N Newman and M Battino ldquoObesity diabetesmellitus atherosclerosis and chronic periodontitis a sharedpathology via oxidative stress andmitochondrial dysfunctionrdquoPeriodontology 2000 vol 64 no 1 pp 139ndash153 2014
[28] G R Naveh V Brumfeld R Shahar and S Weiner ldquoToothperiodontal ligament direct 3D microCT visualization of thecollagen network and how the network changes when the toothis loadedrdquo Journal of Structural Biology vol 181 no 2 pp 108ndash115 2013
[29] Y Kitase M Yokozeki S Fujihara et al ldquoAnalysis of geneexpression profiles in human periodontal ligament cells underhypoxia the protective effect of CC chemokine ligand 2 tooxygen shortagerdquoArchives of Oral Biology vol 54 no 7 pp 618ndash624 2009
[30] G Thornton-Evans P Eke L Wei et al ldquoPeriodontitis amongadults aged ge 30 years - United States 2009-2010rdquo Morbidityand Mortality Weekly Report (MMWR) vol 62 supplement 3pp 129ndash135 2013
[31] S S Socransky C Smith and A D Haffajee ldquoSubgingivalmicrobial profiles in refractory periodontal diseaserdquo Journal ofClinical Periodontology vol 29 no 3 pp 260ndash268 2002
[32] K-T Ng J-P Li K M Ng G L Tipoe W K Leung and M-L Fung ldquoExpression of hypoxia-inducible factor-1120572 in humanperiodontal tissuerdquo Journal of Periodontology vol 82 no 1 pp136ndash141 2011
[33] S Frede U Berchner-Pfannschmidt and J Fandrey ldquoReg-ulation of Hypoxia-inducible factors during inflammationrdquoMethods in Enzymology vol 435 pp 405ndash419 2007
[34] J Shin S Ji and Y Choi ldquoAbility of oral bacteria to inducetissue-destructive molecules from human neutrophilsrdquo OralDiseases vol 14 no 4 pp 327ndash334 2008
[35] S Takashiba K Naruishi and Y Murayama ldquoPerspectiveof cytokine regulation for periodontal treatment fibroblastbiologyrdquo Journal of Periodontology vol 74 no 1 pp 103ndash1102003
[36] A Konermann D Stabenow P A Knolle S A E HeldJ Deschner and A Jager ldquoRegulatory role of periodontalligament fibroblasts for innate immune cell function and dif-ferentiationrdquo Innate Immunity vol 18 no 5 pp 745ndash752 2012
[37] S Kar S Subbaram P M Carrico and J A Melendez ldquoRedox-control of matrix metalloproteinase-1 a critical link betweenfree radicals matrix remodeling and degenerative diseaserdquoRespiratory Physiology andNeurobiology vol 174 no 3 pp 299ndash306 2010
[38] F Rousset M V C Nguyen L Grange F Morel and B LardyldquoHeme oxygenase-1 regulates matrix metalloproteinase MMP-1secretion and chondrocyte cell death via Nox4NADPH oxidaseactivity in chondrocytesrdquo PLoS ONE vol 8 no 6 Article IDe66478 2013
[39] E Rudolf and M Cervinka ldquoResponses of human gingival andperiodontal fibroblasts to a low-zinc environmentrdquo Alternativesto Laboratory Animals vol 38 no 2 pp 119ndash138 2010
[40] S M San Miguel L A Opperman E P Allen J Zielinski andK K H Svoboda ldquoBioactive polyphenol antioxidants protectoral fibroblasts from ROS-inducing agentsrdquo Archives of OralBiology vol 57 no 12 pp 1657ndash1667 2012
[41] S M San Miguel L A Opperman E P Allen J E Zielinskiand K K H Svoboda ldquoAntioxidant combinations protect oralfibroblasts against metal-induced toxicityrdquo Archives of OralBiology vol 58 no 3 pp 299ndash310 2013
[42] U Skaleric C M Manthey S E Mergenhagen B Gaspircand S M Wahl ldquoSuperoxide release and superoxide dismutaseexpression by human gingival fibroblastsrdquo European Journal ofOral Sciences vol 108 no 2 pp 130ndash135 2000
Mediators of Inflammation 13
[43] D Y Kim J-H Jun H-L Lee et al ldquoN-acetylcysteine preventsLPS-Lnduced pro-inflammatory cytokines andMMP2 produc-tion in gingival fibroblastsrdquoArchives of Pharmacal Research vol30 no 10 pp 1283ndash1292 2007
[44] B Rath-Deschner J Deschner S Reimann A Jager andW Gotz ldquoRegulatory effects of biomechanical strain on theinsulin-like growth factor system in human periodontal cellsrdquoJournal of Biomechanics vol 42 no 15 pp 2584ndash2589 2009
[45] P Chomczynski and N Sacchi ldquoSingle-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextractionrdquo Analytical Biochemistry vol 162 no 1 pp 156ndash1591987
[46] B Zhao K Ranguelova J Jiang and R P Mason ldquoStudies onthe photosensitized reduction of resorufin and implications forthe detection of oxidative stress with Amplex Redrdquo Free RadicalBiology and Medicine vol 51 no 1 pp 153ndash159 2011
[47] C C Mandal S Ganapathy Y Gorin et al ldquoReactive oxygenspecies derived from Nox4 mediate BMP2 gene transcriptionand osteoblast differentiationrdquoBiochemical Journal vol 433 no2 pp 393ndash402 2011
[48] J Fandrey S Frede and W Jelkmann ldquoRole of hydrogenperoxide in hypoxia-induced erythropoietin productionrdquo Bio-chemical Journal vol 303 no 2 pp 507ndash510 1994
[49] F Cervellati C Cervellati A Romani et al ldquoHypoxia inducescell damage via oxidative stress in retinal epithelial cellsrdquo FreeRadical Research vol 48 no 3 pp 303ndash312 2014
[50] V Vanasco N D Magnani M C Cimolai et al ldquoEndotoxemiaimpairs heart mitochondrial function by decreasing electrontransfer ATP synthesis and ATP content without affectingmembrane potentialrdquo Journal of Bioenergetics and Biomem-branes vol 44 no 2 pp 243ndash252 2012
[51] R G Alscher N Erturk and L S Heath ldquoRole of superoxidedismutases (SODs) in controlling oxidative stress in plantsrdquoJournal of Experimental Botany vol 53 no 372 pp 1331ndash13412002
[52] E Baltacıoglu P Yuva G Aydın et al ldquoLipid peroxidation levelsand total oxidantantioxidant status in serum and saliva frompatients with chronic and aggressive periodontitis Oxidativestress index a new biomarker for periodontal diseaserdquo Journalof Periodontology vol 85 no 10 pp 1432ndash1441 2014
[53] I L C Chapple M R Milward N Ling-Mountford et alldquoAdjunctive daily supplementation with encapsulated fruitvegetable and berry juice powder concentrates and clinicalperiodontal outcomes a double-blind RCTrdquo Journal of ClinicalPeriodontology vol 39 no 1 pp 62ndash72 2012
[54] A Jenzsch S Eick F Rassoul R Purschwitz and H JentschldquoNutritional intervention in patients with periodontal diseaseClinical immunological and microbiological variables during12 monthsrdquo British Journal of Nutrition vol 101 no 6 pp 879ndash885 2009
[55] N Novakovic T Todorovic M Rakic et al ldquoSalivary antiox-idants as periodontal biomarkers in evaluation of tissue statusand treatment outcomerdquo Journal of Periodontal Research vol49 no 1 pp 129ndash136 2014
[56] N Singh S C Narula R K Sharma S Tewari and P KSehgal ldquoVitamin e supplementation superoxide dismutasestatus and outcome of scaling and root planing in patients withchronic periodontitis a randomized clinical trialrdquo Journal ofPeriodontology vol 85 no 2 pp 242ndash249 2014
[57] S Krifka K-A Hiller G Spagnuolo A Jewett G Schmalz andH Schweikl ldquoThe influence of glutathione on redox regulation
by antioxidant proteins and apoptosis in macrophages exposedto 2-hydroxyethyl methacrylate (HEMA)rdquo Biomaterials vol 33no 21 pp 5177ndash5186 2012
[58] I Borges Jr E A M Moreira D W Filho T B de Oliveira MB S da Silva and T S Frode ldquoProinflammatory and oxidativestress markers in patients with periodontal diseaserdquo Mediatorsof Inflammation vol 2007 Article ID 45794 5 pages 2007
[59] M O Tonguc O Ozturk R Sutcu et al ldquoThe impact of smok-ing status on antioxidant enzyme activity and malondialdehydelevels in chronic periodontitisrdquo Journal of Periodontology vol82 no 9 pp 1320ndash1328 2011
[60] B H Jacoby and W L Davis ldquoThe electron microscopicimmunolocalization of a copper-zinc superoxide dismutase inassociation with collagen fibers of periodontal soft tissuesrdquoJournal of Periodontology vol 62 no 7 pp 413ndash420 1991
[61] T Konopka K Krol W Kopec and H Gerber ldquoTotal antioxi-dant status and 8-hydroxy-21015840-deoxyguanosine levels in gingi-val and peripheral blood of periodontitis patientsrdquo ArchivumImmunologiae et Therapiae Experimentalis vol 55 no 6 pp417ndash422 2007
[62] M Soory ldquoRedox status in periodontal and systemic inflamma-tory conditions including associated neoplasias antioxidants asadjunctive therapyrdquo Infectious DisordersmdashDrug Targets vol 9no 4 pp 415ndash427 2009
[63] M Soory ldquoOxidative stress inducedmechanisms in the progres-sion of periodontal diseases and cancer a common approach toredox homeostasisrdquo Cancers vol 2 no 2 pp 670ndash692 2010
[64] R V Chandra G Srinivas A A Reddy et al ldquoLocally deliveredantioxidant gel as an adjunct to nonsurgical therapy improvesmeasures of oxidative stress and periodontal diseaserdquo Journal ofPeriodontal and Implant Science vol 43 no 3 pp 121ndash129 2013
[65] M A D S Siqueira R G Fischer N R Pereira et al ldquoEffectsof non-surgical periodontal treatment on the L-arginine-nitricoxide pathway and oxidative status in plateletsrdquo ExperimentalBiology and Medicine vol 238 no 6 pp 713ndash722 2013
[66] P Mydel Y Takahashi H Yumoto et al ldquoRoles of the hostoxidative immune response and bacterial antioxidant rubr-erythrin during Porphyromonas gingivalis infectionrdquo PLoSpathogens vol 2 no 7 p e76 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Mediators of Inflammation
important defense mechanism against bacteria In the lastdecades five NOX enzymes (NOX 1-5) and two dual oxidases(Duox) Duox1 and Duox2 have been discovered which wereshown to be expressed not only in phagocytes but also inepithelial cells adipocytes or cardiac fibroblasts [9 10]
The NADPH-oxidase family member NOX4 originallyRenox was first described in kidney cells where it is highlyexpressed [11] Later on NOX4 was also detected in othercell types such as endothelial cells fibroblasts keratinocytesand osteoclasts [9 11 12] The typically transmembranouslylocated enzyme is upregulated in response to endoplasmaticreticulum stress [13] shear stress [14] hypoxia or ischemia[15 16] In addition cytokines like transforming growthfactor- (TGF-) 1205731 tumor necrosis factor- (TNF-)120572 andinsulin like growth factor- (IGF-) 1 induce NOX4 expression[9 17] Furthermore this enzyme seems to be a part of thecellular oxygen sensingmechanismsThis is supported by thefact that multiple interactions exist between NOX4 hypoxiainducible factor-1 alpha (HIF-1120572) and ROS [18 19] NOX4-derived ROS have been associated with a variety of processesfor example apoptosis insulin signaling cell differentiationand senescence and bone remodeling [9 20 21]
To limit the potentially deleterious effects of ROS severalprotective mechanisms have been evolved In mitochondriametabolic uncoupling is one mechanism to regulate ROSproduction In healthy cells excessive ROS are decomposedby a protective antioxidant machinery According to theircatalyzing function in the ROS detoxification process theseredox enzymes are grouped in different systems or familieslike the superoxide dismutase (SOD) catalase (CAT) theglutathione system or thioredoxin Furthermore there arealso nonenzymatic or small molecules including vitamins (Cand E) pyruvate flavonoids ubiquinone carotinoids andmany plant-derived antioxidants involved in ROS scavenging[6]
Oxidative stress is gaining increasing interest as an impor-tant cofactor in the etiology and pathogenesis of differentoral and dental diseases including pulpal or mucosal inflam-mation tumor development or inflammatory processes likeperiodontitis First evidence for the presence and role of ROSin the periodontium was given by the detection of thesemolecules after respiratory burst by polymorphonuclear lym-phocytes (PMNLs) in periodontal damage [22] Since thenthe implication of ROS and their degrading enzymes SODor CAT in the pathogenesis of periodontal diseases has beensupported by several groups [23ndash26] High ROS levels mayalso be a linking element between obesity diabetes mellitusatherosclerosis and chronic periodontitis [27] In additionthere are many intervention studies investigating antioxi-dants and antioxidant micronutrients in periodontal therapyincluding the use of mouth rinses and toothpastes Someof these studies provided evidence for beneficial effects butthe links between certain antioxidant species and pathogenicmechanisms are still missing [22]This is also the case for theappearance of ROS and redox systems in the healthy humanoral cavity or periodontium
The periodontium consists of the alveolar bone the toothand the gingiva as well as the periodontal ligament (PDL)which connects the tooth and the surrounding alveolar bone
The collagenous structure of the PDL functions as a shock-reabsorbing barrier protecting the tissues against mechan-ical forces [28] These forces may induce local hypoxiaas well as inflammation [29] Studies indicate that peri-odontal pathogenic bacteria like Porphyromonas gingivalis(P gingivalis) trigger the onset and duration of chronicperiodontitis [30] P gingivalis is a gram-negative anaerobicbacterium belonging to the red complex of oral microflorain accordance with their pathogenicity [31] P gingivalis isthe most prominent periodontal pathogen and is knownto be associated with artherosclerosis and cardiovasculardiseases To mimic the microenvironment of the gingivalsulcus which is characterized by low oxygen tension [32]we include hypoxic conditions in our experiments Since Pgingivalis is an anaerobic bacterium the hypoxic gingivalsulci represent an ideal place for the proliferation of thesebacteria Moreover inflammation by itself may contributeto the development of tissue hypoxia due to an increase inoxygen consumption for example by invading immune cells[33]
Resident cells in the periodontium are also involved ininflammatory and immuneprocesses leading to cytokine (ieinterleukin- (IL-) 1120573 or TNF-120572) or chemokine release whichcould further enhance inflammation [34] PDL fibroblasts areresponsible for the maintenance remodeling and repair ofthe extracellular matrix of the PDL Their immune modu-latory capacities were demonstrated in some recent studies[35 36] Local ROS accumulationmay promote the inductionof proinflammatory cytokines with consecutive activation ofmacrophages [37] finally leading to periodontal destructionvia MMPs [38]
In vitro studies evaluating oxidative stress at the cellularlevel have been focused on gingival fibroblasts while PDLcells have rarely been analyzed Under low zinc environmenthuman PDL fibroblasts showed increased oxidative stress[39] Similar results were found after nicotine ethanol ormetal exposure which was abolished after different antioxi-dant treatment [40 41] Stimulation experiments in humangingival fibroblasts (HGFs) support our hypothesis that PDLcells might play a pivotal role during periodontal diseasesThe authors demonstrated that exposure to lipopolysaccha-ride from P gingivalis (LPS-PG) or LPS from Escherichiacoli (LPS-E coli) increased superoxide concentrations [26]the levels and expression of ROS [42] and proinflammatorycytokines in HGFs [43] ROS activation was associated withactivation of the redox-sensitive transcription factor nuclearfactor-kappaB (NF-120581B) [43] which is critically involved inthe regulation of cytokine and MMP expression The impor-tance of NF-120581B was also demonstrated for the interactionbetween NOX4 HIF-1120572 and ROS [18 19]
However evidence for the involvement of PDL cells inbacterial- or hypoxia-induced ROS formation is missingThesame is true for the balance betweenNOX4and redox systemsin the periodontium The objective of the present study wastherefore to define the role of NOX4 and ROS formation inthe PDL especially during inflammatory and hypoxic eventsWe hypothesized that ROS are regulated by hypoxia andinflammatory stimuli in PDL cells and periodontal tissuesdue to an imbalance of NOX4 and redox system activation
Mediators of Inflammation 3
The current study will enhance the knowledge of the roleplayed by oxidative stress in periodontal tissues which mightimprove diagnostic and therapeutic strategies in oral diseases
2 Materials and Methods
21 Isolation and Characterization of PDL Cells Humanperiodontal ligament fibroblasts were harvested from caries-free and periodontally healthy premolars from patients aged12 to 17 years (119899 = 9) which had to be extracted dueto orthodontic reasons Exclusion criteria were acute orchronic inflammation Patients have had nomedication theyreceived local anesthesia during extraction and analgesia afterextraction The approval of the Ethics Committee of theUniversity of Bonn as well as parental and patients allowancewas obtained Cells were harvested as previously described[44] In brief cells were collected from the middle part ofthe tooth root to avoid contamination by cells derived fromthe gingiva and dental germ Using explant technique PDLcells were cultured in Dulbeccorsquos minimal essential medium(DMEM Invitrogen Karlsruhe Germany) supplementedwith 10 fetal bovine serum (FBS Invitrogen) 100 units ofpenicillin and 100mgmL streptomycin (Biochrom BerlinGermany) at 37∘C in a humidified atmosphere of 5 CO
2
Cells from nine different individuals were phenotyped by theanalysis of the known PDL markers (alkaline phosphatase(ALP) osteocalcin (OC) osteopontin (OP) periostin andS100 A4) at 3rd passage [40] After phenotyping three celllines were pooled and used for experiments at 4th or 5thpassage
22 Cell Experiments Cells were seeded (50000 cellswell)on six-well plates and grown to 80 confluence Cellswere cultured under normoxic (21 O
2and 5 CO
2) or
hypoxic conditions (1 O2and 5 CO
2 INCO 153 O
2CO2
incubator Memmert GmbH Schwabach Germany) for 2 48 24 and 48 hours with or without the addition of 1 120583gmLultrapure lipopolysaccharide from Porphyromonas gingivalis(LPS-PG) (Invivogen San Diego USA) Prior to the here-presented study we analyzed the dose-response of PDL cellswith respect to the inducibility of NOX4 and H
2O2release by
LPS-PG treatmentThemaximal inductionwas detected afterstimulation with 1 120583gmL LPS-PG and no further increasewas observed when the LPS-PG concentration was raised
23 Analysis of Gene Expression Cells were lysed with 4Mguanidinthiocyanat (Roth Karlsruhe Germany) and totalRNA was extracted using the acid guanidinium thio-cyanatephenolchloroform extraction method [45] A totalof 1 120583g of RNA was reverse-transcribed into cDNA by the useof 200U ofMoloneymurine leukemia virus (MMLV) reversetranscriptase (Bio-Rad Munich Germany) cDNA expres-sion was detected by real-time PCR (RT-PCR) using specificprimers (18S fwd CGGCTACCACATCCAAGGAA revGCTGGAATTACCGCGGCT and NOX4 fwd TCTGCC-TGTTCATCTGGCTCTCCA rev AGCCAAGAGTGT-TCGGCACATGGGTA) and SYBR Green as fluorescentdye on the ViiA7 detection system (Applied Biosystems
Darmstadt Germany) Two 120583L of cDNA served as templatein a 20120583L reaction mixture containing 10 120583L QuantiTectSYBR Green Master Mix (Qiagen Hilden Germany) Themixture was heated initially to 50∘C for 2min and to 95∘C for15min followed by 40 cycles with denaturation at 95∘C for 15 sand annealing and extension at 60∘C for 1min Amounts ofspecific cDNAwere normalized to the housekeeping gene 18Sand cDNA expression was calculated as relative expression ofthe respective control
24 Immunoblot For protein quantification the Pierce BCAProtein Assay Kit (Thermo Scientific Rockford USA) wasused After addition of 4x sample buffer (50mM Tris pH68 2 (wv) SDS 5 (vv) 2-mercaptoethanol 00125bromophenol blue and 1 glycine) 50 120583g of total cell lysateper lane was subjected to 75 SDS-PAGE and transferredonto a nitrocellulose membrane (02 120583mpore size Schleicheramp Schuell Microscience Dassel Germany) The efficiency ofprotein transfer and equal loading was confirmed by stainingthe membrane with Ponceau S (Sigma Aldrich MunichGermany) Membranes were blocked overnight at 4∘C with5 (wv) nonfat milk in TBS-T (10mM Tris pH 75 100mMNaCl and 005 Tween-20) and washed with TBS-T prior toincubation with a rabbit polyclonal antibody against NOX4(Abcam Cambridge UK) at room temperature for 2 h szlig-Actin (Santa Cruz Biotechnology Santa Cruz USA) servedas marker for equal protein loading After washing with TBS-THRP- (horseradish peroxidase-) conjugated anti-rabbit IgGantibody (NewEnglandBiolabs) was added Immunoreactiveproteins were visualized by chemiluminescence (AmershamChemiluminescence Kit) and exposure to X-ray films (AgfaMortsel Belgium) Densitometrical analysis was performedusing the freely available image-processing software ImageJ143 (httprsbinfonihgovij)
25 Amplex Red Assay For hydrogen peroxide (H2O2)
detection the commercially availableN-acetyl-37-dihydrox-yphenoxazine (Amplex Red) Hydrogen PeroxideperoxidaseAssay Kit (Molecular Probes Eugene OR) was used Thecolorless reagent reacts with hydrogen peroxide (H
2O2)
using horseradish peroxidase (HRP) to catalyze the H2O2-
dependent oxidation of nonfluorescent Amplex Red to flu-orescent resorufin red This assay detects only the releaseof hydrogen peroxide since the size of HRP prevents itfrom entering the mitochondria Therefore PDL cells wereincubated with 500120583L Amplex Red and 01 UmL peroxidasewas added at 37∘C for 30min Absorbance was measuredat 565 nm after 1 2 and 4 hours During the procedureexcitation light intensity was kept to an absolute minimumas recommended by Zhao et al [46]
26 Immunofluorescence For this purpose PDL cells werecultured on cover slips to 60 confluence PDL cells werestained for NOX4 and redox systems under normoxic orhypoxic conditions in the presence and absence of LPS-PGHypoxic conditions were obtained by placing the cells in anincubator where the oxygen concentration can be adjusted
4 Mediators of Inflammation
to 1 O2 balanced with 5 CO
2and N
2in the incuba-
tion atmosphere (INCO 153 O2CO2incubator Memmert
GmbH Schwabach Germany) At the end of the experimentmedium was removed cells were washed three times withPBS and fixed with 250120583L cooled methanolaceton (1 1)at minus20∘C for 10min The solution was removed and cellsremained at room temperature for 10min to dry Afterblocking with 3 BSA-PBS for 10min cells were incubatedwith antibodies (Ab) directed against NOX4 (Abcam) CATor SOD (Abcam) 1 50 in PBS 3 BSA for 2 hours Cellswere washed with PBS before the species-specific secondaryAb (dilution 1 400 in PBS) were added for additional90min Cells were covered on slides with DAPI contain-ing mounting medium (Dako) On the following day cellswere scanned with the Zeiss Axio Imager A1 fluorescencemicroscope (HBO 100 Carl Zeiss Jena Germany) anAxioCam MRc camera (Carl Zeiss) and the AxioVision 47software (Carl Zeiss) Density analysis was performed usingthe freely available image-processing software ImageJ 143(httprsbinfonihgovij) Quantification of relative fluores-cence intensity was performed by measuring the intensity ofeach cell related to cell area To obtain grayscale pictures eachwas processed using the tools ldquocolorrdquo and ldquosplit channelsrdquoin ImageJ which resulted in red green and blue color-fractionated grayscale pictures for each channel Only greenfractionated pictures were used for calculation of the relativefluorescence intensity
27 Immunohistochemistry Periodontal tissues from clini-cally healthy patients (I) as well as those from patientswith the diagnosis of gingivitis (II) and periodontitis (III)were obtained from three different patients for each groupDiagnoses of gingivitis and periodontitis were performedby specialists from the Clinic of Dental Medicine Gin-givitis was characterized by a reversible local inflammatoryreaction of the gingiva visible as redness swelling andbleeding In contrast periodontitis affects four differentstructures gingiva periodontal ligament tooth and alveolarbone Therefore periodontitis was diagnosed by an increaseof probing depth bleeding and attachment loss whichwas supported by radiographs Tissues were fixed in 4phosphate-buffered paraformaldehyde for 24 h followed byhydration and dehydration in an ascending ethanol serieswhich ended in paraffin embedding Afterwards 4 120583m thickparaffin-embedded sections were prepared deparaffinizedrehydrated rinsed with TBS pH 74 for 10min and thensoaked in 70mL methanol containing 700mL of 30 H
2O2
for 45min in the dark to block endogenous peroxidase activ-ity Afterwards tissue sections were rinsed and preincubatedwith TBSBSA 4 for 20min to avoid unspecific backgroundstaining Incubation with monoclonal primary antibodies ofNOX4 (Abcam Cambridge UK) and polyclonal antibody ofcatalase and SOD (Abcam) in a 1 100 working solution ofTBSBSA was performed at 4∘C overnight in a humidifiedchamber Subsequently sections were washed in TBS andincubated with a peroxidase-labeled polymer conjugated toa goat anti-rabbit immunoglobulin provided as a ready-to-use solution (Envision Dako AS Glostrup Denmark)
for 30min in a humidified chamber at room temperatureAntibody complexeswere visualized using diaminobenzidinefor 10min resulting in brown staining Thereafter the slideswere rinsed counterstainedwithMayerrsquos hematoxylin rinsedagain and covered Negative controls were prepared by omis-sion of the primary antibodies as well as both the primaryand secondary antibody from the staining procedures usingTBSBSA instead Finally tissue sections were analyzed usingtheZeissAxioScop 2microscope (Carl Zeiss JenaGermany)
28 Statistical Analysis All experiments were performedin triplicate and mRNA quantification is given as meansplusmn SD Results were compared by ANOVA Dunnettrsquos andTukey-Kramer tests were performed as posttest to calculatewhether differences between treated and control groups weresignificant A 119875 value of 005 was considered as significant
3 Results
31 NOX4 Human primary PDL cells were incubated undernormoxic and hypoxic conditions in the presence or absenceof LPS-PG (1 120583gmL) Treatment with LPS-PG induced time-dependently a significant increase in the mRNA expressionof NOX4 compared to the untreated control (Figure 1(a)) Indetail NOX4mRNA levels weremarkedly increased after twohours of LPS-PG stimulation the increase reached significantlevels (4-fold) after 4 hours (119875 lt 0001) NOX4 mRNA levelspeaked after 8-hour stimulation with LPS-PG and returnedto base line levels afterwards
Hypoxia alone induced the expression of NOX4 in PDLcells but in contrast to the stimulation with LPS-PG themaximum in NOX4 mRNA upregulation was detected after4 hours (119875 lt 0001 Figure 1(a)) Subsequently NOX4mRNA expression decreased however it remained at ahigher level compared to the control group even after 24or 48 hours (Figure 1(a)) The highest increase in NOX4mRNA expression was detected after the combination ofboth stimuli (85 times) which was statistically significantcompared to control and even compared to LPS stimulationunder normoxia (119875 lt 0001 Figure 1(a))
To prove whether the increase in mRNA expression wasfollowed by an increase in protein content immunofluo-rescence stainings for NOX4 were performed NOX4 wasdetectable predominantly in the cytosol and to a slight extentin the nucleus of unstimulated PDL cells (Figure 1(b)) To getmore detailed information about the time course of NOX4protein induction after hypoxic and inflammatory stimula-tion we performed immunoblots After 2 hours only hypoxiaprovoked a significant increase in NOX4 protein (119875 lt 0001)which diminished afterwards (Figure 1(c)) Under normoxiccondition the treatment with LPS-PG induced a significantelevation of NOX4 protein after 4 hours of incubation com-pared to control and hypoxia alone (119875 lt 0001 Figure 1(c))The LPS-PG effect was evenmore pronounced under hypoxiccondition which was statistically significant compared to allthe other groups (119875 lt 0001 Figure 1(c)) After 24 hours ofincubation alterations of NOX4 protein levels were no longerdetectable (data not shown) These findings were in line with
Mediators of Inflammation 5
NADPH-oxidase 4 mRNA
125
100
75
50
25
00minus
minus
minus minus minus+
+ minus + minus + minus + minus + minus + minus + minus + minus + minus +
+ minus minus+ + minus minus+ + minus minus+ + + +
lowastlowast
lowast
lowast
2h 4h 8h 24h 48h
HoxLPS
(a) (b)
(c)
mRN
A ex
pres
sion
com
pare
d to
cont
rol
minusminus
minus minus
+
+
+minusminus
minus minusminus
+
+ +
+
lowast
lowast
Hox
NOX4 protein
Inte
grat
ed d
ensit
y (
) com
pare
dto
cont
rol (
au)
LPS
2h 4h
300
200
100100
50
0
Nox Hoxminus minus+ +
120573-Actin
NOX4
LPS
120573-Actin
NOX4
2h
4h
Figure 1 NOX4 in human primary periodontal ligament (PDL) cells (a) PDL cells were cultured under normoxic or hypoxic condition andstimulated with or without LPS of Porphyromonas gingivalis (1 120583gmL) NOX4 mRNA expression was analyzed after 2 4 8 24 and 48 hStatistical differences were analyzed by one-way ANOVA and post hoc Dunnett and Tukeyrsquos multiple comparison test 119875 lt 005 differenceto control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 9) (b) NOX4 protein was visualized by immunofluorescence as well asimmunohistochemical staining in unstimulated PDL fibroblasts (1 50 antibody from Abcam) (c) Immunoblot data of NOX4 in PDL cellscultured under normoxic or hypoxic condition (Hox) and stimulated with or without LPS of Porphyromonas gingivalis (1 120583gmL) NOX4protein levels were analyzed after 2 and 4 h Statistical analysis of western blot data was performed using the freely available image-processingsoftware ImageJ 143 (httprsbinfonihgovij) Statistical differences were analyzed by one-way ANOVA and post hoc Dunnett and Tukeyrsquosmultiple comparison test 119875 lt 005 difference to control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 3) The scale bars indicate200 120583m in the bright field survey and 50120583m in the immunofluorescence image respectively
our mRNA results even though we detected time-dependentdifferences
32 H2O2Formation To determine hydrogen peroxide gen-
eration we used the commercially available Amplex Red-horseradish peroxidase assay PDL cells displayed a lowconstitutive H
2O2production under normoxic conditions
We observed a significant upregulation of the H2O2con-
centration in the cell culture supernatant after 1 2 and 4hours of stimulation with LPS-PG Hypoxia alone induceda significant increase in H
2O2formation compared to the
time-matched controls (Figure 2) While in the LPS groupthe H
2O2values were the highest after 1 hour and decreased
continuously after 2 and 4 hours (Figure 2) under hypoxiccondition H
2O2increased permanently (Figure 2) Compar-
ing the LPS and the hypoxic effect revealed that hypoxiaalone induced a significantly higher H
2O2accumulation over
the complete observation period (119875 lt 0001) Howeverthe combination of both stimuli elicited the highest H
2O2
concentration after 1 hour which was significant not onlycompared to control but also compared to the LPS and thehypoxia group (Figure 2) After 2 hours the treatment withLPS-PG was no longer able to enhance the hypoxia-inducedH2O2formation it was still significantly increased compared
to control and the LPS stimulation but no longer comparedto the hypoxic groupThis was even more pronounced after 4hours
6 Mediators of Inflammation
2h 4h1h
50000
40000
30000
20000
10000
0
(120583M
)
HoxLPS
minus minus minus minus minus minus
minus
+
+minus +minus +minus +minus +minus +
++ + + +
lowast
lowastlowast lowast
lowast
lowast
H2O2 concentration
Figure 2 Determination of ROS formation (H2O2) using Amplex
Red assay PDL cells were cultured under normoxic (Nox) orhypoxic condition (Hox) and stimulated with or without LPS ofPorphyromonas gingivalis (1 120583gmL) H
2O2was analyzed after 1
2 and 4 hours Statistical differences were analyzed by one-wayANOVA and post hoc Dunnett and Tukeyrsquos multiple comparisontest 119875 lt 005 difference to control lowast119875 lt 005 difference betweengroups (means plusmn SD 119899 = 6)
33 Redox Systems To analyze which protectivemechanismsagainst ROS formation are involved in our setting wedetermined the redox system components catalase (CAT)and superoxide dismutase (SOD) in PDL cells after treat-ment with LPS-PG under normoxic and hypoxic conditionsby immunofluorescence (Figures 3(a)-3(b)) In additionperiodontal tissues from clinically healthy patients (I) aswell as from patients with the diagnosis of gingivitis (II)and periodontitis (III) were obtained from three differentpatients for each group and stained immunohistochemicallyfor NOX4 CAT and SOD (Figures 4 5 and 6)
331 Immunofluorescence of Redox Systems in PDL CellsPDL cells displayed a constitutive cytosolic catalase expres-sion under normoxic conditions Treatment with LPS-PGandhypoxia induced a change in PDL cellmorphologywhichwas first obvious after a 1-hour incubation under hypoxicconditions and after 2 hours with LPS-PG (Figure 3(a)) Asshown in Figure 3(b) analyzing catalase cytoplasm densityin relation to constant cell area revealed a significant upreg-ulation of catalase in PDL cells stimulated with LPS-PG orhypoxia after 1 h (119875 lt 0001 Figure 3(b)) This was also truefor hypoxic conditions after 2 h compared to all other groupsUnder inflammatory stimulation (LPS) a significant increaseof catalase cytoplasm density was detectable after 4 hourscompared to all the other groups (119875 lt 0001 Figure 3(b))Instead the combination of both stimuli diminished thehypoxic as well as LPS-induced effects after 1 2 and 4hours Determining superoxide dismutase protein expressionin PDL cells showed no significant differences at any timepoint (119875 gt 005 data not shown)
332 Immunohistochemistry of NOX4 and Redox Systemsin Periodontal Tissue Sections Periodontal tissues (healthygingiva and periodontal ligament (PDL) gingivitis andperiodontitis) were obtained from three different patients foreach group
NOX4 In the healthy periodontal tissue samples only veryweak immunostaining for NOX4 could be obtained (Figures4(a) and 4(b)) In the PDL weak immunoreactivity witha moderate staining of some fibroblasts was observed Ininflamed tissues from patients suffering from gingivitis orperiodontitis strong immunoreactivity for NOX4 appearedin the cytosol and even stronger in the nuclei of keratinocytesof the basal layer of inflamed gingival epithelium (Figures4(c)-4(d)) In gingival and PDL fibroblasts as well as inimmune cells like leukocytes and in vessels the immunore-activity was primarily restricted to the cytosol and cell walls(Figures 4(c)-4(d) see red arrows)
CAT Nearly no immunostaining could be found in thehealthy gingival epithelium (Figure 5(a)) In the healthyPDL predominantly PDL cells located near to the toothroot surfaces were stained (Figure 5(b) see red arrow) Ingingivitis tissue a weak staining of gingival keratinocytessimilar to healthy tissue could be seen (Figure 5(c)) Insteadin the tissue samples of patients with periodontitis a strongimmunoreactivity was observed in the subepithelial layerprobably referring to nuclei staining of immune cells likeleukocytes (Figure 5(d) see red arrows)
SOD As for healthy tissues gingival epithelium showeda moderate to strong immunoreactivity of keratinocyteswhich was primarily limited to the cytosol (Figure 6(a)see red arrow) In healthy periodontium gingival and PDLfibroblasts were weakly stained (Figure 6(b)) Compared toinflamed tissues basal epithelial cells seemed to be stainedmore intensively in the gingivitis specimen (Figures 6(c)-6(d)) In addition the subepithelium revealed immunore-activity in local cells like gingival fibroblasts and scatteredimmune cells (Figure 6(c)) In periodontitis stained sectionsSOD immunostaining seemed to be limited to the gingivalepithelium (Figure 6(d) see red arrows) In inflamed tissuessubepithelial layers showed no signs of immunoreactivity
4 Discussion
Oxidative stress is discussed as an important cofactor in theetiology and pathogenesis of several oral and dental diseasesfor example in inflammatory processes like periodontitisFirst evidence for the presence and role of ROS in theperiodontium was given by the detection of these moleculesafter respiratory burst by polymorphonuclear lymphocytes(PMNLs) in periodontal damage [22] Since then the impli-cation of ROS and their degrading enzymes SOD or CAT inthe pathogenesis of periodontal diseases has been supportedby several groups [23ndash26] To analyse whether cellular redoxsystems in the periodontium are affected by inflammatoryconditions we investigated the effect of hypoxia and LPSfrom Porphyromonas gingivalis (LPS-PG) on periodontal
Mediators of Inflammation 7
1h 2hCatalase
Nox
Nox + LPS
Hox
Hox + LPS
4h
100120583m 100120583m 100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
(a)
2h 4h
Hox
Inte
grat
ed d
ensit
y (a
u)
Catalase cytoplasm density
LPSminusminus + + minusminus + + minusminus +
minus +minus +minus +minus +minus +minus +
+
lowast
lowast
lowast
lowast
lowastlowast
1h
20000
25000
15000
1000010000
7500
5000
2500
0
(b)
Figure 3 Catalase (CAT) in PDL cells (a) CAT was visualized by immunofluorescence staining (1 50 antibody from Abcam) PDL cellswere cultured under normoxic (Nox) or hypoxic condition (Hox) and stimulated with or without LPS of Porphyromonas gingivalis (1120583gmL)Catalase cytoplasm density (b) was determined in relation to cell area using the freely available image-processing software ImageJ 143(httprsbinfonihgovij) Statistical analysis of immunofluorescence data was analyzed by one-way ANOVA and post hoc Dunnett andTukeyrsquos multiple comparison test 119875 lt 005 difference to control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 6) The scale barsindicate 100 120583m in the immunofluorescence images
8 Mediators of Inflammation
50120583m
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m
50120583m
50120583m
Inflamed
(d)
Figure 4 NADPH-oxidase 4 (NOX4) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Monoclonal primary antibody against NOX4 (Abcam Cambridge UK) was used ina concentration of 1 100 In gingival and PDL fibroblasts as well as in immune cells like leukocytes and in vessels the immunoreactivity wasprimarily restricted to the cytosol and cell walls Red arrows indicate examples for the typical distribution (Figures 4(c) and 4(d)) The scalebars indicate 200120583m in the surveys and 50 120583m in the higher magnifications respectively
ligament cells We showed that both stimuli induced asignificant increase of NOX4 and a significant upregulationof hydrogen peroxide generation NOX enzymes were firstdescribed in phagocytes as being responsible for defensemechanisms against pathogens using NOX-generated ROSmolecules Till then NOX4 expression has been detected innonphagocyte cell types like kidney cells osteoclasts andosteoblasts as well as dermal cells [9 11 47] In PDL cellshypoxia and LPS-PG induced a significant increase of NOX4mRNA The protein expression was mainly located in thecytosol but likewise in the nucleus Our findings were inline with Brown and Griendling who have shown that thetypically transmembrane molecule is localized in differentcell compartments [9]
NOX4 is one of the major sources of ROS generationwhich is highly and ubiquitously expressed compared tothe other family members Mandal and colleagues demon-strated that apart from its antimicrobial function NOX4 isresponsible not only for osteoclast but also for preosteoblastdifferentiation via ROS accumulation reinforcing the ideaof NOX4 as a key regulator in bone remodeling underphysiological ROS amounts [47] Animal studies supportthe hypothesis that massive ROS formation induced bone
resorption via NOX4 [20 21] Whereas physiological levelsof NOX4-generated ROS in PDL cellsmight facilitate alveolarbone remodeling massive ROS accumulation during inflam-matory periodontal diseases might lead to alveolar bonedestruction favoring tooth loss We have demonstrated firstthat NOX4 is upregulated in periodontal ligament fibroblastsduring bacterial and hypoxic events Hence PDL fibroblastsmay be a source of excessive ROS generation and are thereforeof particular importance during periodontal diseases
Our results support current findings determining the roleof oxidative stress in different periodontal pathologies whichimplicate that the balance between NADPH-oxidases andredox systems is crucial to maintain cell and tissue home-ostasis [22ndash26] However there are only a few studies usinghuman periodontal fibroblast in the context of oxidativestress and none of them evaluated the role of NOX4 andcounteracting redox systems during inflammatory conditionswith LPS-PG stimulation and hypoxia An increase of ROSgeneration was detected by San Miguel et al following PDLfibroblast stimulation with nicotine or ethanol which wereabolished after antioxidant treatment [40] They completedtheir results using metal ions like nickel for PDL stimulationwhich increases ROS activity in these cells indicating a role
Mediators of Inflammation 9
200120583m
50120583m
50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 5 Catalase (CAT) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontal ligament (b) andsamples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University of Bonn and parentalas well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against catalase (Abcam Cambridge UK) was used in a concentration of1 100 No immunostaining could be found in the healthy gingival epithelium In the healthy PDL predominantly PDL cells located near tothe tooth root surfaces were stained (Figure 5(b) red arrow) In gingivitis tissue a weak staining of gingival keratinocytes similar to healthytissue could be seen (Figure 5(c)) Instead in the tissue samples of patients with periodontitis a strong immunoreactivity was observed inthe subepithelial layer probably referring to nuclei staining of immune cells like leukocytes Red arrows indicate examples for the typicallocalisation (Figure 5(d)) The scale bars indicate 200 120583m in the surveys and 50120583m in the higher magnifications respectively
in metal cytotoxicity or even nickel allergy [41] Transientmetal ions like nickel and cobalt are known to mimic cellularhypoxia mainly due to the inhibition of prolyl hydrolases(PDHs) and asparaginyl hydrolase (factor-inhibiting HIF(FIH)) The inhibition of hydroxylation leads to HIF-1120572nuclear translocation followed by the transcription of diversegenes which are implicated in hypoxic adaptation immuneresponse or angiogenesis therefore indirectly supporting ourresults under hypoxic condition [48 49]
We could show that hypoxia as well as LPS-PG inducedROS formation by detecting H
2O2using the commercially
available Amplex Red assay The high levels of H2O2released
under hypoxic conditions after 1 and 2 hours in PDL cellsmay be caused by different mechanisms First we founda significant upregulation of the mRNA and protein ofthe H
2O2-generating enzyme NOX4 Second the lack of
molecular oxygen as the final electron acceptor in therespiratory chain may be responsible for the early release of
ROS LPS-PG alone induced acutely a small but significantincrease in H
2O2release and enhanced the hypoxia-induced
generation In human gingival fibroblasts similar results wereobtained showing the increase of ROS and antioxidants afterP gingivalis or LPS stimulation [26 42 43] Interestinglydespite the increase in NOX4 mRNA and protein expressionafter 4 hours of stimulation with LPS-PG under hypoxicconditions the H
2O2generation was significantly reduced
when compared to hypoxia alone An additional inhibitionof the respiratory chain by LPS-PG which was described forcardiac mitochondria [50] is therefore conceivable
To antagonize oxidative stress cells developed severalantioxidative enzyme systems Therefore we measured SODand CAT expression in PDL cells and periodontal tissues byimmunofluorescence and immunohistochemistry SODs aremetalloenzymes which are responsible for the dismutation ofsuperoxide radicals to hydrogen peroxide [51] The reductionof hydrogen peroxide to water and oxygen is subsequently
10 Mediators of Inflammation
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m 50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 6 Superoxide dismutase (SOD) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against SOD (Abcam Cambridge UK) was used ina concentration of 1 100 Gingival epithelium showed a moderate to strong staining of SOD in keratinocytes which was primarily limitedto the cytosol (Figure 6(a) and red arrow depicts the typical localization) In the healthy periodontium gingival and PDL fibroblasts wereweakly stained (Figure 6(b)) In inflamed tissues basal epithelial cells seemed to be stained more intensively in the gingivitis specimen thanin periodontitis (Figures 6(c) and 6(d)) In addition the subepithelium revealed immunoreactivity in local cells like gingival fibroblasts andscattered immune cells (Figure 6(c) see red arrow) In periodontitis SOD immunostaining seemed to be limited to the gingival epithelium(Figure 6(d) see red arrow) The scale bars indicate 200 120583m in the surveys and 50 120583m in the higher magnifications respectively
catalyzed by CATs which are ubiquitously expressed Bothenzymes are essential to maintain cellular redox homeostasisThe present data imply that PDL cells and periodontal tissueshave fundamental defense machineries against bacteria rep-resented by NOX4-dependent inducible ROS generation aswell as protective redox systems This is of great importanceconcerning the high amounts and diversity of the oral floraDifferent studies indicate that redox systems are key playersin the development of periodontal diseases and markers forperiodontal health status [52ndash56] We detected a significantincrease of catalase in PDL cells in response to LPS aswell as hypoxia after 1 and 2 hours In our setting PDLfibroblasts seem to counteracted LPS- and hypoxia-inducedROS formation with the elevation of CAT expression Butprolonged incubation under hypoxic conditions failed toincrease catalase Since ROS generation was still elevated atthese later time points a delay in ROS degradation may be
the explanation Taken together the dramatic elevation inROS formation by the combination of hypoxia and LPS-PGon one hand and the reduction of catalase under the sameconditions on the other hand increase the oxidative stress inthe periodontium favoring the progression of inflammationand bone resorption and finally may lead to tooth loss
The fundamental role of the balance between ROSgeneration and antioxidative mechanisms in periodontalinflammatory processes was supported by our immunohis-tochemical findings In patients suffering from periodontalinflammatory diseases like gingivitis and periodontitis wedemonstrated that NOX4 is upregulated and thus likelyresponsible for H
2O2generation However we detected as
well an increase in catalase abundance in inflamed gingivaland periodontal tissue which was in line with data fromKrifka and colleagues who detected high CAT levels inmonomer-exposed macrophages as a response to oxidative
Mediators of Inflammation 11
stress [57] In our in vitro setting PDL fibroblasts seemto counteract LPS- and hypoxia-induced ROS formationpredominantly with an elevation of CAT expression sincewe could not detect any SOD immunoreactivity The roleand impact of the antioxidative systems SOD and CAT arecontroversially discussed Recent studies measured CAT andSOD activities in gingival tissues of patients with periodon-titis [58 59] Borges Jr et al did not find differences inCAT activities comparing healthy and diseased patients [58]Instead Tonguc et al detected decreased local SOD andCAT activities in periodontitis patients which seemed to beincreased by smoking [59] In contrast in inflamed tissues weobservedhigh amounts of SODwhichwas localized primarilyin epithelial cells Jacoby and Davis determined SOD in peri-odontal soft tissues They detected twice as much SOD activ-ity in the periodontal ligament as in the human skin (dermis)[60] Furthermore immunohistochemical analysis revealedthat SOD is localized in the periphery of matrix collagenfibrils and to the glycocalyx of tissue fibroblasts This wasconfirmed by Skaleric et al [42] which might explain the factthat we did not detect significant differences of SOD levelsafter hypoxic and LPS-PG stimulation intracellularly in PDLfibroblasts Another reason for these results may be causedby the age of the PDL cells used in this study We explantedthe PDL cells from young and oral healthy patients but mostof the immunohistochemical stainings were performed intissues from elderly patients with clearly diagnosed gingivitisor periodontitisTherefore it is possible that in PDL cells fromelderly patients a different expression pattern of antioxidantenzyme systems could be found which further deterioratethe redox state of the periodontal ligament Moreover otherauthors found reduced antioxidants in the body fluids ofpatients with periodontitis indicating a dysregulation whichwould favor disease progression [23 24 61] A variety of invitro and animal studies analyzed the effect of antioxidantsupplement in the context of inflammatory or cytotoxicevents [57 62 63] They have shown that antioxidantsinhibit these processes indicating a therapeutic approachin pathologies like periodontitis First clinical investigationsconfirmed the efficiency of antioxidant micronutrients andadjuncts in periodontal diseases [22 54 56 64 65] Thepresent data indirectly support these studies
Concerning periodontal pathogen invasiveness and viru-lence there is compelling evidence that P gingivalis possessesits own protective antioxidants for example rubrerythrindefending them against the oxidative burst of the host [66]We have shown that its virulence factor LPS-PG inducedhigh amounts of ROS via NOX4 which may result in boneresorption favoring the invasiveness of the pathogen whichis at the same time protected by its own defense machineryThis would further explain the pathogenicity and resistanceof this bacterium strain in patients with periodontitis
5 Conclusion
In this study we have demonstrated that LPS from Porphy-romonas gingivalis (LPS-PG) as well as hypoxia induces aNOX4-dependent increase inH
2O2release in PDLfibroblasts
which may contribute to the development and progressionof periodontal diseases when not balanced by a concomitantincrease in antioxidative systems The dramatic elevationin ROS formation induced by the combination of hypoxiaand LPS-PG and the reduction of catalase under the sameconditions increased the oxidative stress in the periodontiumfavoring the progression of inflammation bone resorptionand possibly tooth loss
Taken together the presented data further explain thedestructive mechanism of hypoxic and inflammatory condi-tions in the periodontium and may improve diagnostic andtherapeutic strategies in periodontal diseases
Conflict of Interests
The authors have no conflict of interests to disclose
Acknowledgments
The authors would like to thank Ms Tina Schaffrath for hertechnical supportThe present studywas supported by a grantfrom the German Research Foundation (Clinical ResearchUnit 208TP7) and the Medical Faculty of the University ofBonn
References
[1] S Reuter S C Gupta M M Chaturvedi and B B AggarwalldquoOxidative stress inflammation and cancer how are theylinkedrdquo Free Radical Biology and Medicine vol 49 no 11 pp1603ndash1616 2010
[2] H Sone H Akanuma and T Fukuda ldquoOxygenomics inenvironmental stressrdquo Redox Report vol 15 no 3 pp 98ndash1142010
[3] K E Wellen and C B Thompson ldquoCellular metabolic stressconsidering how cells respond to nutrient excessrdquo MolecularCell vol 40 no 2 pp 323ndash332 2010
[4] R O Poyton K A Ball and P R Castello ldquoMitochondrialgeneration of free radicals and hypoxic signalingrdquo Trends inEndocrinology andMetabolism vol 20 no 7 pp 332ndash340 2009
[5] F Weinberg and N S Chandel ldquoReactive oxygen species-dependent signaling regulates cancerrdquo Cellular and MolecularLife Sciences vol 66 no 23 pp 3663ndash3673 2009
[6] A Cataldi ldquoCell responses to oxidative stressorsrdquo CurrentPharmaceutical Design vol 16 no 12 pp 1387ndash1395 2010
[7] Z Durackova ldquoSome current insights into oxidative stressrdquoPhysiological Research vol 59 no 4 pp 459ndash469 2010
[8] L A Sena and N S Chandel ldquoPhysiological roles of mitochon-drial reactive oxygen speciesrdquoMolecular Cell vol 48 no 2 pp158ndash166 2012
[9] D I Brown and K K Griendling ldquoNox proteins in signaltransductionrdquo Free Radical Biology amp Medicine vol 47 no 9pp 1239ndash1253 2009
[10] K Bedard and K-H Krause ldquoThe NOX family of ROS-generatingNADPHoxidases physiology and pathophysiologyrdquoPhysiological Reviews vol 87 no 1 pp 245ndash313 2007
[11] K-H Krause ldquoTissue distribution and putative physiologicalfunction of NOX family NADPH oxidasesrdquo Japanese Journal ofInfectious Diseases vol 57 no 5 pp S28ndashS29 2004
12 Mediators of Inflammation
[12] S Yang Y Zhang W Ries and L Key ldquoExpression of Nox4 inosteoclastsrdquo Journal of Cellular Biochemistry vol 92 no 2 pp238ndash248 2004
[13] E Pedruzzi C Guichard V Ollivier et al ldquoNAD(P)H oxidaseNox-4mediates 7-ketocholesterol-induced endoplasmic reticu-lum stress and apoptosis in human aortic smooth muscle cellsrdquoMolecular and Cellular Biology vol 24 no 24 pp 10703ndash107172004
[14] J Hwang A Saha Y C Boo et al ldquoOscillatory shear stress stim-ulates endothelial production of O
2-from p47119901ℎ119900119909-dependent
NAD(P)Hoxidases leading tomonocyte adhesionrdquoThe Journalof Biological Chemistry vol 278 no 47 pp 47291ndash47298 2003
[15] H B Suliman M Ali and C A Piantadosi ldquoSuperoxidedismutase-3 promotes full expression of the EPO response tohypoxiardquo Blood vol 104 no 1 pp 43ndash50 2004
[16] P Vallet Y Charnay K Steger et al ldquoNeuronal expressionof the NADPH oxidase NOX4 and its regulation in mouseexperimental brain ischemiardquo Neuroscience vol 132 no 2 pp233ndash238 2005
[17] K T Moe S Aulia F Jiang et al ldquoDifferential upregulation ofNox homologues of NADPH oxidase by tumor necrosis factor-120572 in human aortic smooth muscle and embryonic kidney cellsrdquoJournal of Cellular andMolecularMedicine vol 10 no 1 pp 231ndash239 2006
[18] I Diebold A Petry J Hess and A Gorlach ldquoThe NADPHoxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1rdquoMolecular Biology of the Cell vol 21 no 12pp 2087ndash2096 2010
[19] K V Tormos and N S Chandel ldquoInter-connection betweenmitochondria and HIFsrdquo Journal of Cellular and MolecularMedicine vol 14 no 4 pp 795ndash804 2010
[20] T Ohnishi K Bandow K Kakimoto M Machigashira TMatsuyama and T Matsuguchi ldquoOxidative stress causes alve-olar bone loss in metabolic syndrome model mice with type 2diabetesrdquo Journal of Periodontal Research vol 44 no 1 pp 43ndash51 2009
[21] C Goettsch A Babelova O Trummer et al ldquoNADPH oxidase4 limits bone mass by promoting osteoclastogenesisrdquo Journal ofClinical Investigation vol 123 no 11 pp 4731ndash4738 2013
[22] I L C Chapple and J B Matthews ldquoThe role of reactive oxygenand antioxidant species in periodontal tissue destructionrdquoPeriodontology 2000 vol 43 no 1 pp 160ndash232 2007
[23] A Guentsch P M Preshaw S Bremer-Streck G KlingerE Glockmann and B W Sigusch ldquoLipid peroxidation andantioxidant activity in saliva of periodontitis patients effect ofsmoking and periodontal treatmentrdquo Clinical Oral Investiga-tions vol 12 no 4 pp 345ndash352 2008
[24] R Agnihotri P Pandurang S U Kamath et al ldquoAssociation ofcigarette smoking with superoxide dismutase enzyme levels insubjected with chonic periodontitisrdquo Journal of Periodontologyvol 80 no 4 pp 657ndash662 2009
[25] D Ekuni T Tomofuji N Tamaki et al ldquoMechanical stimulationof gingiva reduces plasma 8-OHdG level in rat periodontitisrdquoArchives of Oral Biology vol 53 no 4 pp 324ndash329 2008
[26] H Staudte A Guntsch A Volpel and B W Sigusch ldquoVitaminC attenuates the cytotoxic effects of Porphyromonas gingivalison human gingival fibroblastsrdquo Archives of Oral Biology vol 55no 1 pp 40ndash45 2010
[27] P Bullon H N Newman and M Battino ldquoObesity diabetesmellitus atherosclerosis and chronic periodontitis a sharedpathology via oxidative stress andmitochondrial dysfunctionrdquoPeriodontology 2000 vol 64 no 1 pp 139ndash153 2014
[28] G R Naveh V Brumfeld R Shahar and S Weiner ldquoToothperiodontal ligament direct 3D microCT visualization of thecollagen network and how the network changes when the toothis loadedrdquo Journal of Structural Biology vol 181 no 2 pp 108ndash115 2013
[29] Y Kitase M Yokozeki S Fujihara et al ldquoAnalysis of geneexpression profiles in human periodontal ligament cells underhypoxia the protective effect of CC chemokine ligand 2 tooxygen shortagerdquoArchives of Oral Biology vol 54 no 7 pp 618ndash624 2009
[30] G Thornton-Evans P Eke L Wei et al ldquoPeriodontitis amongadults aged ge 30 years - United States 2009-2010rdquo Morbidityand Mortality Weekly Report (MMWR) vol 62 supplement 3pp 129ndash135 2013
[31] S S Socransky C Smith and A D Haffajee ldquoSubgingivalmicrobial profiles in refractory periodontal diseaserdquo Journal ofClinical Periodontology vol 29 no 3 pp 260ndash268 2002
[32] K-T Ng J-P Li K M Ng G L Tipoe W K Leung and M-L Fung ldquoExpression of hypoxia-inducible factor-1120572 in humanperiodontal tissuerdquo Journal of Periodontology vol 82 no 1 pp136ndash141 2011
[33] S Frede U Berchner-Pfannschmidt and J Fandrey ldquoReg-ulation of Hypoxia-inducible factors during inflammationrdquoMethods in Enzymology vol 435 pp 405ndash419 2007
[34] J Shin S Ji and Y Choi ldquoAbility of oral bacteria to inducetissue-destructive molecules from human neutrophilsrdquo OralDiseases vol 14 no 4 pp 327ndash334 2008
[35] S Takashiba K Naruishi and Y Murayama ldquoPerspectiveof cytokine regulation for periodontal treatment fibroblastbiologyrdquo Journal of Periodontology vol 74 no 1 pp 103ndash1102003
[36] A Konermann D Stabenow P A Knolle S A E HeldJ Deschner and A Jager ldquoRegulatory role of periodontalligament fibroblasts for innate immune cell function and dif-ferentiationrdquo Innate Immunity vol 18 no 5 pp 745ndash752 2012
[37] S Kar S Subbaram P M Carrico and J A Melendez ldquoRedox-control of matrix metalloproteinase-1 a critical link betweenfree radicals matrix remodeling and degenerative diseaserdquoRespiratory Physiology andNeurobiology vol 174 no 3 pp 299ndash306 2010
[38] F Rousset M V C Nguyen L Grange F Morel and B LardyldquoHeme oxygenase-1 regulates matrix metalloproteinase MMP-1secretion and chondrocyte cell death via Nox4NADPH oxidaseactivity in chondrocytesrdquo PLoS ONE vol 8 no 6 Article IDe66478 2013
[39] E Rudolf and M Cervinka ldquoResponses of human gingival andperiodontal fibroblasts to a low-zinc environmentrdquo Alternativesto Laboratory Animals vol 38 no 2 pp 119ndash138 2010
[40] S M San Miguel L A Opperman E P Allen J Zielinski andK K H Svoboda ldquoBioactive polyphenol antioxidants protectoral fibroblasts from ROS-inducing agentsrdquo Archives of OralBiology vol 57 no 12 pp 1657ndash1667 2012
[41] S M San Miguel L A Opperman E P Allen J E Zielinskiand K K H Svoboda ldquoAntioxidant combinations protect oralfibroblasts against metal-induced toxicityrdquo Archives of OralBiology vol 58 no 3 pp 299ndash310 2013
[42] U Skaleric C M Manthey S E Mergenhagen B Gaspircand S M Wahl ldquoSuperoxide release and superoxide dismutaseexpression by human gingival fibroblastsrdquo European Journal ofOral Sciences vol 108 no 2 pp 130ndash135 2000
Mediators of Inflammation 13
[43] D Y Kim J-H Jun H-L Lee et al ldquoN-acetylcysteine preventsLPS-Lnduced pro-inflammatory cytokines andMMP2 produc-tion in gingival fibroblastsrdquoArchives of Pharmacal Research vol30 no 10 pp 1283ndash1292 2007
[44] B Rath-Deschner J Deschner S Reimann A Jager andW Gotz ldquoRegulatory effects of biomechanical strain on theinsulin-like growth factor system in human periodontal cellsrdquoJournal of Biomechanics vol 42 no 15 pp 2584ndash2589 2009
[45] P Chomczynski and N Sacchi ldquoSingle-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextractionrdquo Analytical Biochemistry vol 162 no 1 pp 156ndash1591987
[46] B Zhao K Ranguelova J Jiang and R P Mason ldquoStudies onthe photosensitized reduction of resorufin and implications forthe detection of oxidative stress with Amplex Redrdquo Free RadicalBiology and Medicine vol 51 no 1 pp 153ndash159 2011
[47] C C Mandal S Ganapathy Y Gorin et al ldquoReactive oxygenspecies derived from Nox4 mediate BMP2 gene transcriptionand osteoblast differentiationrdquoBiochemical Journal vol 433 no2 pp 393ndash402 2011
[48] J Fandrey S Frede and W Jelkmann ldquoRole of hydrogenperoxide in hypoxia-induced erythropoietin productionrdquo Bio-chemical Journal vol 303 no 2 pp 507ndash510 1994
[49] F Cervellati C Cervellati A Romani et al ldquoHypoxia inducescell damage via oxidative stress in retinal epithelial cellsrdquo FreeRadical Research vol 48 no 3 pp 303ndash312 2014
[50] V Vanasco N D Magnani M C Cimolai et al ldquoEndotoxemiaimpairs heart mitochondrial function by decreasing electrontransfer ATP synthesis and ATP content without affectingmembrane potentialrdquo Journal of Bioenergetics and Biomem-branes vol 44 no 2 pp 243ndash252 2012
[51] R G Alscher N Erturk and L S Heath ldquoRole of superoxidedismutases (SODs) in controlling oxidative stress in plantsrdquoJournal of Experimental Botany vol 53 no 372 pp 1331ndash13412002
[52] E Baltacıoglu P Yuva G Aydın et al ldquoLipid peroxidation levelsand total oxidantantioxidant status in serum and saliva frompatients with chronic and aggressive periodontitis Oxidativestress index a new biomarker for periodontal diseaserdquo Journalof Periodontology vol 85 no 10 pp 1432ndash1441 2014
[53] I L C Chapple M R Milward N Ling-Mountford et alldquoAdjunctive daily supplementation with encapsulated fruitvegetable and berry juice powder concentrates and clinicalperiodontal outcomes a double-blind RCTrdquo Journal of ClinicalPeriodontology vol 39 no 1 pp 62ndash72 2012
[54] A Jenzsch S Eick F Rassoul R Purschwitz and H JentschldquoNutritional intervention in patients with periodontal diseaseClinical immunological and microbiological variables during12 monthsrdquo British Journal of Nutrition vol 101 no 6 pp 879ndash885 2009
[55] N Novakovic T Todorovic M Rakic et al ldquoSalivary antiox-idants as periodontal biomarkers in evaluation of tissue statusand treatment outcomerdquo Journal of Periodontal Research vol49 no 1 pp 129ndash136 2014
[56] N Singh S C Narula R K Sharma S Tewari and P KSehgal ldquoVitamin e supplementation superoxide dismutasestatus and outcome of scaling and root planing in patients withchronic periodontitis a randomized clinical trialrdquo Journal ofPeriodontology vol 85 no 2 pp 242ndash249 2014
[57] S Krifka K-A Hiller G Spagnuolo A Jewett G Schmalz andH Schweikl ldquoThe influence of glutathione on redox regulation
by antioxidant proteins and apoptosis in macrophages exposedto 2-hydroxyethyl methacrylate (HEMA)rdquo Biomaterials vol 33no 21 pp 5177ndash5186 2012
[58] I Borges Jr E A M Moreira D W Filho T B de Oliveira MB S da Silva and T S Frode ldquoProinflammatory and oxidativestress markers in patients with periodontal diseaserdquo Mediatorsof Inflammation vol 2007 Article ID 45794 5 pages 2007
[59] M O Tonguc O Ozturk R Sutcu et al ldquoThe impact of smok-ing status on antioxidant enzyme activity and malondialdehydelevels in chronic periodontitisrdquo Journal of Periodontology vol82 no 9 pp 1320ndash1328 2011
[60] B H Jacoby and W L Davis ldquoThe electron microscopicimmunolocalization of a copper-zinc superoxide dismutase inassociation with collagen fibers of periodontal soft tissuesrdquoJournal of Periodontology vol 62 no 7 pp 413ndash420 1991
[61] T Konopka K Krol W Kopec and H Gerber ldquoTotal antioxi-dant status and 8-hydroxy-21015840-deoxyguanosine levels in gingi-val and peripheral blood of periodontitis patientsrdquo ArchivumImmunologiae et Therapiae Experimentalis vol 55 no 6 pp417ndash422 2007
[62] M Soory ldquoRedox status in periodontal and systemic inflamma-tory conditions including associated neoplasias antioxidants asadjunctive therapyrdquo Infectious DisordersmdashDrug Targets vol 9no 4 pp 415ndash427 2009
[63] M Soory ldquoOxidative stress inducedmechanisms in the progres-sion of periodontal diseases and cancer a common approach toredox homeostasisrdquo Cancers vol 2 no 2 pp 670ndash692 2010
[64] R V Chandra G Srinivas A A Reddy et al ldquoLocally deliveredantioxidant gel as an adjunct to nonsurgical therapy improvesmeasures of oxidative stress and periodontal diseaserdquo Journal ofPeriodontal and Implant Science vol 43 no 3 pp 121ndash129 2013
[65] M A D S Siqueira R G Fischer N R Pereira et al ldquoEffectsof non-surgical periodontal treatment on the L-arginine-nitricoxide pathway and oxidative status in plateletsrdquo ExperimentalBiology and Medicine vol 238 no 6 pp 713ndash722 2013
[66] P Mydel Y Takahashi H Yumoto et al ldquoRoles of the hostoxidative immune response and bacterial antioxidant rubr-erythrin during Porphyromonas gingivalis infectionrdquo PLoSpathogens vol 2 no 7 p e76 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 3
The current study will enhance the knowledge of the roleplayed by oxidative stress in periodontal tissues which mightimprove diagnostic and therapeutic strategies in oral diseases
2 Materials and Methods
21 Isolation and Characterization of PDL Cells Humanperiodontal ligament fibroblasts were harvested from caries-free and periodontally healthy premolars from patients aged12 to 17 years (119899 = 9) which had to be extracted dueto orthodontic reasons Exclusion criteria were acute orchronic inflammation Patients have had nomedication theyreceived local anesthesia during extraction and analgesia afterextraction The approval of the Ethics Committee of theUniversity of Bonn as well as parental and patients allowancewas obtained Cells were harvested as previously described[44] In brief cells were collected from the middle part ofthe tooth root to avoid contamination by cells derived fromthe gingiva and dental germ Using explant technique PDLcells were cultured in Dulbeccorsquos minimal essential medium(DMEM Invitrogen Karlsruhe Germany) supplementedwith 10 fetal bovine serum (FBS Invitrogen) 100 units ofpenicillin and 100mgmL streptomycin (Biochrom BerlinGermany) at 37∘C in a humidified atmosphere of 5 CO
2
Cells from nine different individuals were phenotyped by theanalysis of the known PDL markers (alkaline phosphatase(ALP) osteocalcin (OC) osteopontin (OP) periostin andS100 A4) at 3rd passage [40] After phenotyping three celllines were pooled and used for experiments at 4th or 5thpassage
22 Cell Experiments Cells were seeded (50000 cellswell)on six-well plates and grown to 80 confluence Cellswere cultured under normoxic (21 O
2and 5 CO
2) or
hypoxic conditions (1 O2and 5 CO
2 INCO 153 O
2CO2
incubator Memmert GmbH Schwabach Germany) for 2 48 24 and 48 hours with or without the addition of 1 120583gmLultrapure lipopolysaccharide from Porphyromonas gingivalis(LPS-PG) (Invivogen San Diego USA) Prior to the here-presented study we analyzed the dose-response of PDL cellswith respect to the inducibility of NOX4 and H
2O2release by
LPS-PG treatmentThemaximal inductionwas detected afterstimulation with 1 120583gmL LPS-PG and no further increasewas observed when the LPS-PG concentration was raised
23 Analysis of Gene Expression Cells were lysed with 4Mguanidinthiocyanat (Roth Karlsruhe Germany) and totalRNA was extracted using the acid guanidinium thio-cyanatephenolchloroform extraction method [45] A totalof 1 120583g of RNA was reverse-transcribed into cDNA by the useof 200U ofMoloneymurine leukemia virus (MMLV) reversetranscriptase (Bio-Rad Munich Germany) cDNA expres-sion was detected by real-time PCR (RT-PCR) using specificprimers (18S fwd CGGCTACCACATCCAAGGAA revGCTGGAATTACCGCGGCT and NOX4 fwd TCTGCC-TGTTCATCTGGCTCTCCA rev AGCCAAGAGTGT-TCGGCACATGGGTA) and SYBR Green as fluorescentdye on the ViiA7 detection system (Applied Biosystems
Darmstadt Germany) Two 120583L of cDNA served as templatein a 20120583L reaction mixture containing 10 120583L QuantiTectSYBR Green Master Mix (Qiagen Hilden Germany) Themixture was heated initially to 50∘C for 2min and to 95∘C for15min followed by 40 cycles with denaturation at 95∘C for 15 sand annealing and extension at 60∘C for 1min Amounts ofspecific cDNAwere normalized to the housekeeping gene 18Sand cDNA expression was calculated as relative expression ofthe respective control
24 Immunoblot For protein quantification the Pierce BCAProtein Assay Kit (Thermo Scientific Rockford USA) wasused After addition of 4x sample buffer (50mM Tris pH68 2 (wv) SDS 5 (vv) 2-mercaptoethanol 00125bromophenol blue and 1 glycine) 50 120583g of total cell lysateper lane was subjected to 75 SDS-PAGE and transferredonto a nitrocellulose membrane (02 120583mpore size Schleicheramp Schuell Microscience Dassel Germany) The efficiency ofprotein transfer and equal loading was confirmed by stainingthe membrane with Ponceau S (Sigma Aldrich MunichGermany) Membranes were blocked overnight at 4∘C with5 (wv) nonfat milk in TBS-T (10mM Tris pH 75 100mMNaCl and 005 Tween-20) and washed with TBS-T prior toincubation with a rabbit polyclonal antibody against NOX4(Abcam Cambridge UK) at room temperature for 2 h szlig-Actin (Santa Cruz Biotechnology Santa Cruz USA) servedas marker for equal protein loading After washing with TBS-THRP- (horseradish peroxidase-) conjugated anti-rabbit IgGantibody (NewEnglandBiolabs) was added Immunoreactiveproteins were visualized by chemiluminescence (AmershamChemiluminescence Kit) and exposure to X-ray films (AgfaMortsel Belgium) Densitometrical analysis was performedusing the freely available image-processing software ImageJ143 (httprsbinfonihgovij)
25 Amplex Red Assay For hydrogen peroxide (H2O2)
detection the commercially availableN-acetyl-37-dihydrox-yphenoxazine (Amplex Red) Hydrogen PeroxideperoxidaseAssay Kit (Molecular Probes Eugene OR) was used Thecolorless reagent reacts with hydrogen peroxide (H
2O2)
using horseradish peroxidase (HRP) to catalyze the H2O2-
dependent oxidation of nonfluorescent Amplex Red to flu-orescent resorufin red This assay detects only the releaseof hydrogen peroxide since the size of HRP prevents itfrom entering the mitochondria Therefore PDL cells wereincubated with 500120583L Amplex Red and 01 UmL peroxidasewas added at 37∘C for 30min Absorbance was measuredat 565 nm after 1 2 and 4 hours During the procedureexcitation light intensity was kept to an absolute minimumas recommended by Zhao et al [46]
26 Immunofluorescence For this purpose PDL cells werecultured on cover slips to 60 confluence PDL cells werestained for NOX4 and redox systems under normoxic orhypoxic conditions in the presence and absence of LPS-PGHypoxic conditions were obtained by placing the cells in anincubator where the oxygen concentration can be adjusted
4 Mediators of Inflammation
to 1 O2 balanced with 5 CO
2and N
2in the incuba-
tion atmosphere (INCO 153 O2CO2incubator Memmert
GmbH Schwabach Germany) At the end of the experimentmedium was removed cells were washed three times withPBS and fixed with 250120583L cooled methanolaceton (1 1)at minus20∘C for 10min The solution was removed and cellsremained at room temperature for 10min to dry Afterblocking with 3 BSA-PBS for 10min cells were incubatedwith antibodies (Ab) directed against NOX4 (Abcam) CATor SOD (Abcam) 1 50 in PBS 3 BSA for 2 hours Cellswere washed with PBS before the species-specific secondaryAb (dilution 1 400 in PBS) were added for additional90min Cells were covered on slides with DAPI contain-ing mounting medium (Dako) On the following day cellswere scanned with the Zeiss Axio Imager A1 fluorescencemicroscope (HBO 100 Carl Zeiss Jena Germany) anAxioCam MRc camera (Carl Zeiss) and the AxioVision 47software (Carl Zeiss) Density analysis was performed usingthe freely available image-processing software ImageJ 143(httprsbinfonihgovij) Quantification of relative fluores-cence intensity was performed by measuring the intensity ofeach cell related to cell area To obtain grayscale pictures eachwas processed using the tools ldquocolorrdquo and ldquosplit channelsrdquoin ImageJ which resulted in red green and blue color-fractionated grayscale pictures for each channel Only greenfractionated pictures were used for calculation of the relativefluorescence intensity
27 Immunohistochemistry Periodontal tissues from clini-cally healthy patients (I) as well as those from patientswith the diagnosis of gingivitis (II) and periodontitis (III)were obtained from three different patients for each groupDiagnoses of gingivitis and periodontitis were performedby specialists from the Clinic of Dental Medicine Gin-givitis was characterized by a reversible local inflammatoryreaction of the gingiva visible as redness swelling andbleeding In contrast periodontitis affects four differentstructures gingiva periodontal ligament tooth and alveolarbone Therefore periodontitis was diagnosed by an increaseof probing depth bleeding and attachment loss whichwas supported by radiographs Tissues were fixed in 4phosphate-buffered paraformaldehyde for 24 h followed byhydration and dehydration in an ascending ethanol serieswhich ended in paraffin embedding Afterwards 4 120583m thickparaffin-embedded sections were prepared deparaffinizedrehydrated rinsed with TBS pH 74 for 10min and thensoaked in 70mL methanol containing 700mL of 30 H
2O2
for 45min in the dark to block endogenous peroxidase activ-ity Afterwards tissue sections were rinsed and preincubatedwith TBSBSA 4 for 20min to avoid unspecific backgroundstaining Incubation with monoclonal primary antibodies ofNOX4 (Abcam Cambridge UK) and polyclonal antibody ofcatalase and SOD (Abcam) in a 1 100 working solution ofTBSBSA was performed at 4∘C overnight in a humidifiedchamber Subsequently sections were washed in TBS andincubated with a peroxidase-labeled polymer conjugated toa goat anti-rabbit immunoglobulin provided as a ready-to-use solution (Envision Dako AS Glostrup Denmark)
for 30min in a humidified chamber at room temperatureAntibody complexeswere visualized using diaminobenzidinefor 10min resulting in brown staining Thereafter the slideswere rinsed counterstainedwithMayerrsquos hematoxylin rinsedagain and covered Negative controls were prepared by omis-sion of the primary antibodies as well as both the primaryand secondary antibody from the staining procedures usingTBSBSA instead Finally tissue sections were analyzed usingtheZeissAxioScop 2microscope (Carl Zeiss JenaGermany)
28 Statistical Analysis All experiments were performedin triplicate and mRNA quantification is given as meansplusmn SD Results were compared by ANOVA Dunnettrsquos andTukey-Kramer tests were performed as posttest to calculatewhether differences between treated and control groups weresignificant A 119875 value of 005 was considered as significant
3 Results
31 NOX4 Human primary PDL cells were incubated undernormoxic and hypoxic conditions in the presence or absenceof LPS-PG (1 120583gmL) Treatment with LPS-PG induced time-dependently a significant increase in the mRNA expressionof NOX4 compared to the untreated control (Figure 1(a)) Indetail NOX4mRNA levels weremarkedly increased after twohours of LPS-PG stimulation the increase reached significantlevels (4-fold) after 4 hours (119875 lt 0001) NOX4 mRNA levelspeaked after 8-hour stimulation with LPS-PG and returnedto base line levels afterwards
Hypoxia alone induced the expression of NOX4 in PDLcells but in contrast to the stimulation with LPS-PG themaximum in NOX4 mRNA upregulation was detected after4 hours (119875 lt 0001 Figure 1(a)) Subsequently NOX4mRNA expression decreased however it remained at ahigher level compared to the control group even after 24or 48 hours (Figure 1(a)) The highest increase in NOX4mRNA expression was detected after the combination ofboth stimuli (85 times) which was statistically significantcompared to control and even compared to LPS stimulationunder normoxia (119875 lt 0001 Figure 1(a))
To prove whether the increase in mRNA expression wasfollowed by an increase in protein content immunofluo-rescence stainings for NOX4 were performed NOX4 wasdetectable predominantly in the cytosol and to a slight extentin the nucleus of unstimulated PDL cells (Figure 1(b)) To getmore detailed information about the time course of NOX4protein induction after hypoxic and inflammatory stimula-tion we performed immunoblots After 2 hours only hypoxiaprovoked a significant increase in NOX4 protein (119875 lt 0001)which diminished afterwards (Figure 1(c)) Under normoxiccondition the treatment with LPS-PG induced a significantelevation of NOX4 protein after 4 hours of incubation com-pared to control and hypoxia alone (119875 lt 0001 Figure 1(c))The LPS-PG effect was evenmore pronounced under hypoxiccondition which was statistically significant compared to allthe other groups (119875 lt 0001 Figure 1(c)) After 24 hours ofincubation alterations of NOX4 protein levels were no longerdetectable (data not shown) These findings were in line with
Mediators of Inflammation 5
NADPH-oxidase 4 mRNA
125
100
75
50
25
00minus
minus
minus minus minus+
+ minus + minus + minus + minus + minus + minus + minus + minus + minus +
+ minus minus+ + minus minus+ + minus minus+ + + +
lowastlowast
lowast
lowast
2h 4h 8h 24h 48h
HoxLPS
(a) (b)
(c)
mRN
A ex
pres
sion
com
pare
d to
cont
rol
minusminus
minus minus
+
+
+minusminus
minus minusminus
+
+ +
+
lowast
lowast
Hox
NOX4 protein
Inte
grat
ed d
ensit
y (
) com
pare
dto
cont
rol (
au)
LPS
2h 4h
300
200
100100
50
0
Nox Hoxminus minus+ +
120573-Actin
NOX4
LPS
120573-Actin
NOX4
2h
4h
Figure 1 NOX4 in human primary periodontal ligament (PDL) cells (a) PDL cells were cultured under normoxic or hypoxic condition andstimulated with or without LPS of Porphyromonas gingivalis (1 120583gmL) NOX4 mRNA expression was analyzed after 2 4 8 24 and 48 hStatistical differences were analyzed by one-way ANOVA and post hoc Dunnett and Tukeyrsquos multiple comparison test 119875 lt 005 differenceto control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 9) (b) NOX4 protein was visualized by immunofluorescence as well asimmunohistochemical staining in unstimulated PDL fibroblasts (1 50 antibody from Abcam) (c) Immunoblot data of NOX4 in PDL cellscultured under normoxic or hypoxic condition (Hox) and stimulated with or without LPS of Porphyromonas gingivalis (1 120583gmL) NOX4protein levels were analyzed after 2 and 4 h Statistical analysis of western blot data was performed using the freely available image-processingsoftware ImageJ 143 (httprsbinfonihgovij) Statistical differences were analyzed by one-way ANOVA and post hoc Dunnett and Tukeyrsquosmultiple comparison test 119875 lt 005 difference to control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 3) The scale bars indicate200 120583m in the bright field survey and 50120583m in the immunofluorescence image respectively
our mRNA results even though we detected time-dependentdifferences
32 H2O2Formation To determine hydrogen peroxide gen-
eration we used the commercially available Amplex Red-horseradish peroxidase assay PDL cells displayed a lowconstitutive H
2O2production under normoxic conditions
We observed a significant upregulation of the H2O2con-
centration in the cell culture supernatant after 1 2 and 4hours of stimulation with LPS-PG Hypoxia alone induceda significant increase in H
2O2formation compared to the
time-matched controls (Figure 2) While in the LPS groupthe H
2O2values were the highest after 1 hour and decreased
continuously after 2 and 4 hours (Figure 2) under hypoxiccondition H
2O2increased permanently (Figure 2) Compar-
ing the LPS and the hypoxic effect revealed that hypoxiaalone induced a significantly higher H
2O2accumulation over
the complete observation period (119875 lt 0001) Howeverthe combination of both stimuli elicited the highest H
2O2
concentration after 1 hour which was significant not onlycompared to control but also compared to the LPS and thehypoxia group (Figure 2) After 2 hours the treatment withLPS-PG was no longer able to enhance the hypoxia-inducedH2O2formation it was still significantly increased compared
to control and the LPS stimulation but no longer comparedto the hypoxic groupThis was even more pronounced after 4hours
6 Mediators of Inflammation
2h 4h1h
50000
40000
30000
20000
10000
0
(120583M
)
HoxLPS
minus minus minus minus minus minus
minus
+
+minus +minus +minus +minus +minus +
++ + + +
lowast
lowastlowast lowast
lowast
lowast
H2O2 concentration
Figure 2 Determination of ROS formation (H2O2) using Amplex
Red assay PDL cells were cultured under normoxic (Nox) orhypoxic condition (Hox) and stimulated with or without LPS ofPorphyromonas gingivalis (1 120583gmL) H
2O2was analyzed after 1
2 and 4 hours Statistical differences were analyzed by one-wayANOVA and post hoc Dunnett and Tukeyrsquos multiple comparisontest 119875 lt 005 difference to control lowast119875 lt 005 difference betweengroups (means plusmn SD 119899 = 6)
33 Redox Systems To analyze which protectivemechanismsagainst ROS formation are involved in our setting wedetermined the redox system components catalase (CAT)and superoxide dismutase (SOD) in PDL cells after treat-ment with LPS-PG under normoxic and hypoxic conditionsby immunofluorescence (Figures 3(a)-3(b)) In additionperiodontal tissues from clinically healthy patients (I) aswell as from patients with the diagnosis of gingivitis (II)and periodontitis (III) were obtained from three differentpatients for each group and stained immunohistochemicallyfor NOX4 CAT and SOD (Figures 4 5 and 6)
331 Immunofluorescence of Redox Systems in PDL CellsPDL cells displayed a constitutive cytosolic catalase expres-sion under normoxic conditions Treatment with LPS-PGandhypoxia induced a change in PDL cellmorphologywhichwas first obvious after a 1-hour incubation under hypoxicconditions and after 2 hours with LPS-PG (Figure 3(a)) Asshown in Figure 3(b) analyzing catalase cytoplasm densityin relation to constant cell area revealed a significant upreg-ulation of catalase in PDL cells stimulated with LPS-PG orhypoxia after 1 h (119875 lt 0001 Figure 3(b)) This was also truefor hypoxic conditions after 2 h compared to all other groupsUnder inflammatory stimulation (LPS) a significant increaseof catalase cytoplasm density was detectable after 4 hourscompared to all the other groups (119875 lt 0001 Figure 3(b))Instead the combination of both stimuli diminished thehypoxic as well as LPS-induced effects after 1 2 and 4hours Determining superoxide dismutase protein expressionin PDL cells showed no significant differences at any timepoint (119875 gt 005 data not shown)
332 Immunohistochemistry of NOX4 and Redox Systemsin Periodontal Tissue Sections Periodontal tissues (healthygingiva and periodontal ligament (PDL) gingivitis andperiodontitis) were obtained from three different patients foreach group
NOX4 In the healthy periodontal tissue samples only veryweak immunostaining for NOX4 could be obtained (Figures4(a) and 4(b)) In the PDL weak immunoreactivity witha moderate staining of some fibroblasts was observed Ininflamed tissues from patients suffering from gingivitis orperiodontitis strong immunoreactivity for NOX4 appearedin the cytosol and even stronger in the nuclei of keratinocytesof the basal layer of inflamed gingival epithelium (Figures4(c)-4(d)) In gingival and PDL fibroblasts as well as inimmune cells like leukocytes and in vessels the immunore-activity was primarily restricted to the cytosol and cell walls(Figures 4(c)-4(d) see red arrows)
CAT Nearly no immunostaining could be found in thehealthy gingival epithelium (Figure 5(a)) In the healthyPDL predominantly PDL cells located near to the toothroot surfaces were stained (Figure 5(b) see red arrow) Ingingivitis tissue a weak staining of gingival keratinocytessimilar to healthy tissue could be seen (Figure 5(c)) Insteadin the tissue samples of patients with periodontitis a strongimmunoreactivity was observed in the subepithelial layerprobably referring to nuclei staining of immune cells likeleukocytes (Figure 5(d) see red arrows)
SOD As for healthy tissues gingival epithelium showeda moderate to strong immunoreactivity of keratinocyteswhich was primarily limited to the cytosol (Figure 6(a)see red arrow) In healthy periodontium gingival and PDLfibroblasts were weakly stained (Figure 6(b)) Compared toinflamed tissues basal epithelial cells seemed to be stainedmore intensively in the gingivitis specimen (Figures 6(c)-6(d)) In addition the subepithelium revealed immunore-activity in local cells like gingival fibroblasts and scatteredimmune cells (Figure 6(c)) In periodontitis stained sectionsSOD immunostaining seemed to be limited to the gingivalepithelium (Figure 6(d) see red arrows) In inflamed tissuessubepithelial layers showed no signs of immunoreactivity
4 Discussion
Oxidative stress is discussed as an important cofactor in theetiology and pathogenesis of several oral and dental diseasesfor example in inflammatory processes like periodontitisFirst evidence for the presence and role of ROS in theperiodontium was given by the detection of these moleculesafter respiratory burst by polymorphonuclear lymphocytes(PMNLs) in periodontal damage [22] Since then the impli-cation of ROS and their degrading enzymes SOD or CAT inthe pathogenesis of periodontal diseases has been supportedby several groups [23ndash26] To analyse whether cellular redoxsystems in the periodontium are affected by inflammatoryconditions we investigated the effect of hypoxia and LPSfrom Porphyromonas gingivalis (LPS-PG) on periodontal
Mediators of Inflammation 7
1h 2hCatalase
Nox
Nox + LPS
Hox
Hox + LPS
4h
100120583m 100120583m 100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
(a)
2h 4h
Hox
Inte
grat
ed d
ensit
y (a
u)
Catalase cytoplasm density
LPSminusminus + + minusminus + + minusminus +
minus +minus +minus +minus +minus +minus +
+
lowast
lowast
lowast
lowast
lowastlowast
1h
20000
25000
15000
1000010000
7500
5000
2500
0
(b)
Figure 3 Catalase (CAT) in PDL cells (a) CAT was visualized by immunofluorescence staining (1 50 antibody from Abcam) PDL cellswere cultured under normoxic (Nox) or hypoxic condition (Hox) and stimulated with or without LPS of Porphyromonas gingivalis (1120583gmL)Catalase cytoplasm density (b) was determined in relation to cell area using the freely available image-processing software ImageJ 143(httprsbinfonihgovij) Statistical analysis of immunofluorescence data was analyzed by one-way ANOVA and post hoc Dunnett andTukeyrsquos multiple comparison test 119875 lt 005 difference to control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 6) The scale barsindicate 100 120583m in the immunofluorescence images
8 Mediators of Inflammation
50120583m
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m
50120583m
50120583m
Inflamed
(d)
Figure 4 NADPH-oxidase 4 (NOX4) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Monoclonal primary antibody against NOX4 (Abcam Cambridge UK) was used ina concentration of 1 100 In gingival and PDL fibroblasts as well as in immune cells like leukocytes and in vessels the immunoreactivity wasprimarily restricted to the cytosol and cell walls Red arrows indicate examples for the typical distribution (Figures 4(c) and 4(d)) The scalebars indicate 200120583m in the surveys and 50 120583m in the higher magnifications respectively
ligament cells We showed that both stimuli induced asignificant increase of NOX4 and a significant upregulationof hydrogen peroxide generation NOX enzymes were firstdescribed in phagocytes as being responsible for defensemechanisms against pathogens using NOX-generated ROSmolecules Till then NOX4 expression has been detected innonphagocyte cell types like kidney cells osteoclasts andosteoblasts as well as dermal cells [9 11 47] In PDL cellshypoxia and LPS-PG induced a significant increase of NOX4mRNA The protein expression was mainly located in thecytosol but likewise in the nucleus Our findings were inline with Brown and Griendling who have shown that thetypically transmembrane molecule is localized in differentcell compartments [9]
NOX4 is one of the major sources of ROS generationwhich is highly and ubiquitously expressed compared tothe other family members Mandal and colleagues demon-strated that apart from its antimicrobial function NOX4 isresponsible not only for osteoclast but also for preosteoblastdifferentiation via ROS accumulation reinforcing the ideaof NOX4 as a key regulator in bone remodeling underphysiological ROS amounts [47] Animal studies supportthe hypothesis that massive ROS formation induced bone
resorption via NOX4 [20 21] Whereas physiological levelsof NOX4-generated ROS in PDL cellsmight facilitate alveolarbone remodeling massive ROS accumulation during inflam-matory periodontal diseases might lead to alveolar bonedestruction favoring tooth loss We have demonstrated firstthat NOX4 is upregulated in periodontal ligament fibroblastsduring bacterial and hypoxic events Hence PDL fibroblastsmay be a source of excessive ROS generation and are thereforeof particular importance during periodontal diseases
Our results support current findings determining the roleof oxidative stress in different periodontal pathologies whichimplicate that the balance between NADPH-oxidases andredox systems is crucial to maintain cell and tissue home-ostasis [22ndash26] However there are only a few studies usinghuman periodontal fibroblast in the context of oxidativestress and none of them evaluated the role of NOX4 andcounteracting redox systems during inflammatory conditionswith LPS-PG stimulation and hypoxia An increase of ROSgeneration was detected by San Miguel et al following PDLfibroblast stimulation with nicotine or ethanol which wereabolished after antioxidant treatment [40] They completedtheir results using metal ions like nickel for PDL stimulationwhich increases ROS activity in these cells indicating a role
Mediators of Inflammation 9
200120583m
50120583m
50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 5 Catalase (CAT) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontal ligament (b) andsamples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University of Bonn and parentalas well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against catalase (Abcam Cambridge UK) was used in a concentration of1 100 No immunostaining could be found in the healthy gingival epithelium In the healthy PDL predominantly PDL cells located near tothe tooth root surfaces were stained (Figure 5(b) red arrow) In gingivitis tissue a weak staining of gingival keratinocytes similar to healthytissue could be seen (Figure 5(c)) Instead in the tissue samples of patients with periodontitis a strong immunoreactivity was observed inthe subepithelial layer probably referring to nuclei staining of immune cells like leukocytes Red arrows indicate examples for the typicallocalisation (Figure 5(d)) The scale bars indicate 200 120583m in the surveys and 50120583m in the higher magnifications respectively
in metal cytotoxicity or even nickel allergy [41] Transientmetal ions like nickel and cobalt are known to mimic cellularhypoxia mainly due to the inhibition of prolyl hydrolases(PDHs) and asparaginyl hydrolase (factor-inhibiting HIF(FIH)) The inhibition of hydroxylation leads to HIF-1120572nuclear translocation followed by the transcription of diversegenes which are implicated in hypoxic adaptation immuneresponse or angiogenesis therefore indirectly supporting ourresults under hypoxic condition [48 49]
We could show that hypoxia as well as LPS-PG inducedROS formation by detecting H
2O2using the commercially
available Amplex Red assay The high levels of H2O2released
under hypoxic conditions after 1 and 2 hours in PDL cellsmay be caused by different mechanisms First we founda significant upregulation of the mRNA and protein ofthe H
2O2-generating enzyme NOX4 Second the lack of
molecular oxygen as the final electron acceptor in therespiratory chain may be responsible for the early release of
ROS LPS-PG alone induced acutely a small but significantincrease in H
2O2release and enhanced the hypoxia-induced
generation In human gingival fibroblasts similar results wereobtained showing the increase of ROS and antioxidants afterP gingivalis or LPS stimulation [26 42 43] Interestinglydespite the increase in NOX4 mRNA and protein expressionafter 4 hours of stimulation with LPS-PG under hypoxicconditions the H
2O2generation was significantly reduced
when compared to hypoxia alone An additional inhibitionof the respiratory chain by LPS-PG which was described forcardiac mitochondria [50] is therefore conceivable
To antagonize oxidative stress cells developed severalantioxidative enzyme systems Therefore we measured SODand CAT expression in PDL cells and periodontal tissues byimmunofluorescence and immunohistochemistry SODs aremetalloenzymes which are responsible for the dismutation ofsuperoxide radicals to hydrogen peroxide [51] The reductionof hydrogen peroxide to water and oxygen is subsequently
10 Mediators of Inflammation
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m 50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 6 Superoxide dismutase (SOD) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against SOD (Abcam Cambridge UK) was used ina concentration of 1 100 Gingival epithelium showed a moderate to strong staining of SOD in keratinocytes which was primarily limitedto the cytosol (Figure 6(a) and red arrow depicts the typical localization) In the healthy periodontium gingival and PDL fibroblasts wereweakly stained (Figure 6(b)) In inflamed tissues basal epithelial cells seemed to be stained more intensively in the gingivitis specimen thanin periodontitis (Figures 6(c) and 6(d)) In addition the subepithelium revealed immunoreactivity in local cells like gingival fibroblasts andscattered immune cells (Figure 6(c) see red arrow) In periodontitis SOD immunostaining seemed to be limited to the gingival epithelium(Figure 6(d) see red arrow) The scale bars indicate 200 120583m in the surveys and 50 120583m in the higher magnifications respectively
catalyzed by CATs which are ubiquitously expressed Bothenzymes are essential to maintain cellular redox homeostasisThe present data imply that PDL cells and periodontal tissueshave fundamental defense machineries against bacteria rep-resented by NOX4-dependent inducible ROS generation aswell as protective redox systems This is of great importanceconcerning the high amounts and diversity of the oral floraDifferent studies indicate that redox systems are key playersin the development of periodontal diseases and markers forperiodontal health status [52ndash56] We detected a significantincrease of catalase in PDL cells in response to LPS aswell as hypoxia after 1 and 2 hours In our setting PDLfibroblasts seem to counteracted LPS- and hypoxia-inducedROS formation with the elevation of CAT expression Butprolonged incubation under hypoxic conditions failed toincrease catalase Since ROS generation was still elevated atthese later time points a delay in ROS degradation may be
the explanation Taken together the dramatic elevation inROS formation by the combination of hypoxia and LPS-PGon one hand and the reduction of catalase under the sameconditions on the other hand increase the oxidative stress inthe periodontium favoring the progression of inflammationand bone resorption and finally may lead to tooth loss
The fundamental role of the balance between ROSgeneration and antioxidative mechanisms in periodontalinflammatory processes was supported by our immunohis-tochemical findings In patients suffering from periodontalinflammatory diseases like gingivitis and periodontitis wedemonstrated that NOX4 is upregulated and thus likelyresponsible for H
2O2generation However we detected as
well an increase in catalase abundance in inflamed gingivaland periodontal tissue which was in line with data fromKrifka and colleagues who detected high CAT levels inmonomer-exposed macrophages as a response to oxidative
Mediators of Inflammation 11
stress [57] In our in vitro setting PDL fibroblasts seemto counteract LPS- and hypoxia-induced ROS formationpredominantly with an elevation of CAT expression sincewe could not detect any SOD immunoreactivity The roleand impact of the antioxidative systems SOD and CAT arecontroversially discussed Recent studies measured CAT andSOD activities in gingival tissues of patients with periodon-titis [58 59] Borges Jr et al did not find differences inCAT activities comparing healthy and diseased patients [58]Instead Tonguc et al detected decreased local SOD andCAT activities in periodontitis patients which seemed to beincreased by smoking [59] In contrast in inflamed tissues weobservedhigh amounts of SODwhichwas localized primarilyin epithelial cells Jacoby and Davis determined SOD in peri-odontal soft tissues They detected twice as much SOD activ-ity in the periodontal ligament as in the human skin (dermis)[60] Furthermore immunohistochemical analysis revealedthat SOD is localized in the periphery of matrix collagenfibrils and to the glycocalyx of tissue fibroblasts This wasconfirmed by Skaleric et al [42] which might explain the factthat we did not detect significant differences of SOD levelsafter hypoxic and LPS-PG stimulation intracellularly in PDLfibroblasts Another reason for these results may be causedby the age of the PDL cells used in this study We explantedthe PDL cells from young and oral healthy patients but mostof the immunohistochemical stainings were performed intissues from elderly patients with clearly diagnosed gingivitisor periodontitisTherefore it is possible that in PDL cells fromelderly patients a different expression pattern of antioxidantenzyme systems could be found which further deterioratethe redox state of the periodontal ligament Moreover otherauthors found reduced antioxidants in the body fluids ofpatients with periodontitis indicating a dysregulation whichwould favor disease progression [23 24 61] A variety of invitro and animal studies analyzed the effect of antioxidantsupplement in the context of inflammatory or cytotoxicevents [57 62 63] They have shown that antioxidantsinhibit these processes indicating a therapeutic approachin pathologies like periodontitis First clinical investigationsconfirmed the efficiency of antioxidant micronutrients andadjuncts in periodontal diseases [22 54 56 64 65] Thepresent data indirectly support these studies
Concerning periodontal pathogen invasiveness and viru-lence there is compelling evidence that P gingivalis possessesits own protective antioxidants for example rubrerythrindefending them against the oxidative burst of the host [66]We have shown that its virulence factor LPS-PG inducedhigh amounts of ROS via NOX4 which may result in boneresorption favoring the invasiveness of the pathogen whichis at the same time protected by its own defense machineryThis would further explain the pathogenicity and resistanceof this bacterium strain in patients with periodontitis
5 Conclusion
In this study we have demonstrated that LPS from Porphy-romonas gingivalis (LPS-PG) as well as hypoxia induces aNOX4-dependent increase inH
2O2release in PDLfibroblasts
which may contribute to the development and progressionof periodontal diseases when not balanced by a concomitantincrease in antioxidative systems The dramatic elevationin ROS formation induced by the combination of hypoxiaand LPS-PG and the reduction of catalase under the sameconditions increased the oxidative stress in the periodontiumfavoring the progression of inflammation bone resorptionand possibly tooth loss
Taken together the presented data further explain thedestructive mechanism of hypoxic and inflammatory condi-tions in the periodontium and may improve diagnostic andtherapeutic strategies in periodontal diseases
Conflict of Interests
The authors have no conflict of interests to disclose
Acknowledgments
The authors would like to thank Ms Tina Schaffrath for hertechnical supportThe present studywas supported by a grantfrom the German Research Foundation (Clinical ResearchUnit 208TP7) and the Medical Faculty of the University ofBonn
References
[1] S Reuter S C Gupta M M Chaturvedi and B B AggarwalldquoOxidative stress inflammation and cancer how are theylinkedrdquo Free Radical Biology and Medicine vol 49 no 11 pp1603ndash1616 2010
[2] H Sone H Akanuma and T Fukuda ldquoOxygenomics inenvironmental stressrdquo Redox Report vol 15 no 3 pp 98ndash1142010
[3] K E Wellen and C B Thompson ldquoCellular metabolic stressconsidering how cells respond to nutrient excessrdquo MolecularCell vol 40 no 2 pp 323ndash332 2010
[4] R O Poyton K A Ball and P R Castello ldquoMitochondrialgeneration of free radicals and hypoxic signalingrdquo Trends inEndocrinology andMetabolism vol 20 no 7 pp 332ndash340 2009
[5] F Weinberg and N S Chandel ldquoReactive oxygen species-dependent signaling regulates cancerrdquo Cellular and MolecularLife Sciences vol 66 no 23 pp 3663ndash3673 2009
[6] A Cataldi ldquoCell responses to oxidative stressorsrdquo CurrentPharmaceutical Design vol 16 no 12 pp 1387ndash1395 2010
[7] Z Durackova ldquoSome current insights into oxidative stressrdquoPhysiological Research vol 59 no 4 pp 459ndash469 2010
[8] L A Sena and N S Chandel ldquoPhysiological roles of mitochon-drial reactive oxygen speciesrdquoMolecular Cell vol 48 no 2 pp158ndash166 2012
[9] D I Brown and K K Griendling ldquoNox proteins in signaltransductionrdquo Free Radical Biology amp Medicine vol 47 no 9pp 1239ndash1253 2009
[10] K Bedard and K-H Krause ldquoThe NOX family of ROS-generatingNADPHoxidases physiology and pathophysiologyrdquoPhysiological Reviews vol 87 no 1 pp 245ndash313 2007
[11] K-H Krause ldquoTissue distribution and putative physiologicalfunction of NOX family NADPH oxidasesrdquo Japanese Journal ofInfectious Diseases vol 57 no 5 pp S28ndashS29 2004
12 Mediators of Inflammation
[12] S Yang Y Zhang W Ries and L Key ldquoExpression of Nox4 inosteoclastsrdquo Journal of Cellular Biochemistry vol 92 no 2 pp238ndash248 2004
[13] E Pedruzzi C Guichard V Ollivier et al ldquoNAD(P)H oxidaseNox-4mediates 7-ketocholesterol-induced endoplasmic reticu-lum stress and apoptosis in human aortic smooth muscle cellsrdquoMolecular and Cellular Biology vol 24 no 24 pp 10703ndash107172004
[14] J Hwang A Saha Y C Boo et al ldquoOscillatory shear stress stim-ulates endothelial production of O
2-from p47119901ℎ119900119909-dependent
NAD(P)Hoxidases leading tomonocyte adhesionrdquoThe Journalof Biological Chemistry vol 278 no 47 pp 47291ndash47298 2003
[15] H B Suliman M Ali and C A Piantadosi ldquoSuperoxidedismutase-3 promotes full expression of the EPO response tohypoxiardquo Blood vol 104 no 1 pp 43ndash50 2004
[16] P Vallet Y Charnay K Steger et al ldquoNeuronal expressionof the NADPH oxidase NOX4 and its regulation in mouseexperimental brain ischemiardquo Neuroscience vol 132 no 2 pp233ndash238 2005
[17] K T Moe S Aulia F Jiang et al ldquoDifferential upregulation ofNox homologues of NADPH oxidase by tumor necrosis factor-120572 in human aortic smooth muscle and embryonic kidney cellsrdquoJournal of Cellular andMolecularMedicine vol 10 no 1 pp 231ndash239 2006
[18] I Diebold A Petry J Hess and A Gorlach ldquoThe NADPHoxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1rdquoMolecular Biology of the Cell vol 21 no 12pp 2087ndash2096 2010
[19] K V Tormos and N S Chandel ldquoInter-connection betweenmitochondria and HIFsrdquo Journal of Cellular and MolecularMedicine vol 14 no 4 pp 795ndash804 2010
[20] T Ohnishi K Bandow K Kakimoto M Machigashira TMatsuyama and T Matsuguchi ldquoOxidative stress causes alve-olar bone loss in metabolic syndrome model mice with type 2diabetesrdquo Journal of Periodontal Research vol 44 no 1 pp 43ndash51 2009
[21] C Goettsch A Babelova O Trummer et al ldquoNADPH oxidase4 limits bone mass by promoting osteoclastogenesisrdquo Journal ofClinical Investigation vol 123 no 11 pp 4731ndash4738 2013
[22] I L C Chapple and J B Matthews ldquoThe role of reactive oxygenand antioxidant species in periodontal tissue destructionrdquoPeriodontology 2000 vol 43 no 1 pp 160ndash232 2007
[23] A Guentsch P M Preshaw S Bremer-Streck G KlingerE Glockmann and B W Sigusch ldquoLipid peroxidation andantioxidant activity in saliva of periodontitis patients effect ofsmoking and periodontal treatmentrdquo Clinical Oral Investiga-tions vol 12 no 4 pp 345ndash352 2008
[24] R Agnihotri P Pandurang S U Kamath et al ldquoAssociation ofcigarette smoking with superoxide dismutase enzyme levels insubjected with chonic periodontitisrdquo Journal of Periodontologyvol 80 no 4 pp 657ndash662 2009
[25] D Ekuni T Tomofuji N Tamaki et al ldquoMechanical stimulationof gingiva reduces plasma 8-OHdG level in rat periodontitisrdquoArchives of Oral Biology vol 53 no 4 pp 324ndash329 2008
[26] H Staudte A Guntsch A Volpel and B W Sigusch ldquoVitaminC attenuates the cytotoxic effects of Porphyromonas gingivalison human gingival fibroblastsrdquo Archives of Oral Biology vol 55no 1 pp 40ndash45 2010
[27] P Bullon H N Newman and M Battino ldquoObesity diabetesmellitus atherosclerosis and chronic periodontitis a sharedpathology via oxidative stress andmitochondrial dysfunctionrdquoPeriodontology 2000 vol 64 no 1 pp 139ndash153 2014
[28] G R Naveh V Brumfeld R Shahar and S Weiner ldquoToothperiodontal ligament direct 3D microCT visualization of thecollagen network and how the network changes when the toothis loadedrdquo Journal of Structural Biology vol 181 no 2 pp 108ndash115 2013
[29] Y Kitase M Yokozeki S Fujihara et al ldquoAnalysis of geneexpression profiles in human periodontal ligament cells underhypoxia the protective effect of CC chemokine ligand 2 tooxygen shortagerdquoArchives of Oral Biology vol 54 no 7 pp 618ndash624 2009
[30] G Thornton-Evans P Eke L Wei et al ldquoPeriodontitis amongadults aged ge 30 years - United States 2009-2010rdquo Morbidityand Mortality Weekly Report (MMWR) vol 62 supplement 3pp 129ndash135 2013
[31] S S Socransky C Smith and A D Haffajee ldquoSubgingivalmicrobial profiles in refractory periodontal diseaserdquo Journal ofClinical Periodontology vol 29 no 3 pp 260ndash268 2002
[32] K-T Ng J-P Li K M Ng G L Tipoe W K Leung and M-L Fung ldquoExpression of hypoxia-inducible factor-1120572 in humanperiodontal tissuerdquo Journal of Periodontology vol 82 no 1 pp136ndash141 2011
[33] S Frede U Berchner-Pfannschmidt and J Fandrey ldquoReg-ulation of Hypoxia-inducible factors during inflammationrdquoMethods in Enzymology vol 435 pp 405ndash419 2007
[34] J Shin S Ji and Y Choi ldquoAbility of oral bacteria to inducetissue-destructive molecules from human neutrophilsrdquo OralDiseases vol 14 no 4 pp 327ndash334 2008
[35] S Takashiba K Naruishi and Y Murayama ldquoPerspectiveof cytokine regulation for periodontal treatment fibroblastbiologyrdquo Journal of Periodontology vol 74 no 1 pp 103ndash1102003
[36] A Konermann D Stabenow P A Knolle S A E HeldJ Deschner and A Jager ldquoRegulatory role of periodontalligament fibroblasts for innate immune cell function and dif-ferentiationrdquo Innate Immunity vol 18 no 5 pp 745ndash752 2012
[37] S Kar S Subbaram P M Carrico and J A Melendez ldquoRedox-control of matrix metalloproteinase-1 a critical link betweenfree radicals matrix remodeling and degenerative diseaserdquoRespiratory Physiology andNeurobiology vol 174 no 3 pp 299ndash306 2010
[38] F Rousset M V C Nguyen L Grange F Morel and B LardyldquoHeme oxygenase-1 regulates matrix metalloproteinase MMP-1secretion and chondrocyte cell death via Nox4NADPH oxidaseactivity in chondrocytesrdquo PLoS ONE vol 8 no 6 Article IDe66478 2013
[39] E Rudolf and M Cervinka ldquoResponses of human gingival andperiodontal fibroblasts to a low-zinc environmentrdquo Alternativesto Laboratory Animals vol 38 no 2 pp 119ndash138 2010
[40] S M San Miguel L A Opperman E P Allen J Zielinski andK K H Svoboda ldquoBioactive polyphenol antioxidants protectoral fibroblasts from ROS-inducing agentsrdquo Archives of OralBiology vol 57 no 12 pp 1657ndash1667 2012
[41] S M San Miguel L A Opperman E P Allen J E Zielinskiand K K H Svoboda ldquoAntioxidant combinations protect oralfibroblasts against metal-induced toxicityrdquo Archives of OralBiology vol 58 no 3 pp 299ndash310 2013
[42] U Skaleric C M Manthey S E Mergenhagen B Gaspircand S M Wahl ldquoSuperoxide release and superoxide dismutaseexpression by human gingival fibroblastsrdquo European Journal ofOral Sciences vol 108 no 2 pp 130ndash135 2000
Mediators of Inflammation 13
[43] D Y Kim J-H Jun H-L Lee et al ldquoN-acetylcysteine preventsLPS-Lnduced pro-inflammatory cytokines andMMP2 produc-tion in gingival fibroblastsrdquoArchives of Pharmacal Research vol30 no 10 pp 1283ndash1292 2007
[44] B Rath-Deschner J Deschner S Reimann A Jager andW Gotz ldquoRegulatory effects of biomechanical strain on theinsulin-like growth factor system in human periodontal cellsrdquoJournal of Biomechanics vol 42 no 15 pp 2584ndash2589 2009
[45] P Chomczynski and N Sacchi ldquoSingle-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextractionrdquo Analytical Biochemistry vol 162 no 1 pp 156ndash1591987
[46] B Zhao K Ranguelova J Jiang and R P Mason ldquoStudies onthe photosensitized reduction of resorufin and implications forthe detection of oxidative stress with Amplex Redrdquo Free RadicalBiology and Medicine vol 51 no 1 pp 153ndash159 2011
[47] C C Mandal S Ganapathy Y Gorin et al ldquoReactive oxygenspecies derived from Nox4 mediate BMP2 gene transcriptionand osteoblast differentiationrdquoBiochemical Journal vol 433 no2 pp 393ndash402 2011
[48] J Fandrey S Frede and W Jelkmann ldquoRole of hydrogenperoxide in hypoxia-induced erythropoietin productionrdquo Bio-chemical Journal vol 303 no 2 pp 507ndash510 1994
[49] F Cervellati C Cervellati A Romani et al ldquoHypoxia inducescell damage via oxidative stress in retinal epithelial cellsrdquo FreeRadical Research vol 48 no 3 pp 303ndash312 2014
[50] V Vanasco N D Magnani M C Cimolai et al ldquoEndotoxemiaimpairs heart mitochondrial function by decreasing electrontransfer ATP synthesis and ATP content without affectingmembrane potentialrdquo Journal of Bioenergetics and Biomem-branes vol 44 no 2 pp 243ndash252 2012
[51] R G Alscher N Erturk and L S Heath ldquoRole of superoxidedismutases (SODs) in controlling oxidative stress in plantsrdquoJournal of Experimental Botany vol 53 no 372 pp 1331ndash13412002
[52] E Baltacıoglu P Yuva G Aydın et al ldquoLipid peroxidation levelsand total oxidantantioxidant status in serum and saliva frompatients with chronic and aggressive periodontitis Oxidativestress index a new biomarker for periodontal diseaserdquo Journalof Periodontology vol 85 no 10 pp 1432ndash1441 2014
[53] I L C Chapple M R Milward N Ling-Mountford et alldquoAdjunctive daily supplementation with encapsulated fruitvegetable and berry juice powder concentrates and clinicalperiodontal outcomes a double-blind RCTrdquo Journal of ClinicalPeriodontology vol 39 no 1 pp 62ndash72 2012
[54] A Jenzsch S Eick F Rassoul R Purschwitz and H JentschldquoNutritional intervention in patients with periodontal diseaseClinical immunological and microbiological variables during12 monthsrdquo British Journal of Nutrition vol 101 no 6 pp 879ndash885 2009
[55] N Novakovic T Todorovic M Rakic et al ldquoSalivary antiox-idants as periodontal biomarkers in evaluation of tissue statusand treatment outcomerdquo Journal of Periodontal Research vol49 no 1 pp 129ndash136 2014
[56] N Singh S C Narula R K Sharma S Tewari and P KSehgal ldquoVitamin e supplementation superoxide dismutasestatus and outcome of scaling and root planing in patients withchronic periodontitis a randomized clinical trialrdquo Journal ofPeriodontology vol 85 no 2 pp 242ndash249 2014
[57] S Krifka K-A Hiller G Spagnuolo A Jewett G Schmalz andH Schweikl ldquoThe influence of glutathione on redox regulation
by antioxidant proteins and apoptosis in macrophages exposedto 2-hydroxyethyl methacrylate (HEMA)rdquo Biomaterials vol 33no 21 pp 5177ndash5186 2012
[58] I Borges Jr E A M Moreira D W Filho T B de Oliveira MB S da Silva and T S Frode ldquoProinflammatory and oxidativestress markers in patients with periodontal diseaserdquo Mediatorsof Inflammation vol 2007 Article ID 45794 5 pages 2007
[59] M O Tonguc O Ozturk R Sutcu et al ldquoThe impact of smok-ing status on antioxidant enzyme activity and malondialdehydelevels in chronic periodontitisrdquo Journal of Periodontology vol82 no 9 pp 1320ndash1328 2011
[60] B H Jacoby and W L Davis ldquoThe electron microscopicimmunolocalization of a copper-zinc superoxide dismutase inassociation with collagen fibers of periodontal soft tissuesrdquoJournal of Periodontology vol 62 no 7 pp 413ndash420 1991
[61] T Konopka K Krol W Kopec and H Gerber ldquoTotal antioxi-dant status and 8-hydroxy-21015840-deoxyguanosine levels in gingi-val and peripheral blood of periodontitis patientsrdquo ArchivumImmunologiae et Therapiae Experimentalis vol 55 no 6 pp417ndash422 2007
[62] M Soory ldquoRedox status in periodontal and systemic inflamma-tory conditions including associated neoplasias antioxidants asadjunctive therapyrdquo Infectious DisordersmdashDrug Targets vol 9no 4 pp 415ndash427 2009
[63] M Soory ldquoOxidative stress inducedmechanisms in the progres-sion of periodontal diseases and cancer a common approach toredox homeostasisrdquo Cancers vol 2 no 2 pp 670ndash692 2010
[64] R V Chandra G Srinivas A A Reddy et al ldquoLocally deliveredantioxidant gel as an adjunct to nonsurgical therapy improvesmeasures of oxidative stress and periodontal diseaserdquo Journal ofPeriodontal and Implant Science vol 43 no 3 pp 121ndash129 2013
[65] M A D S Siqueira R G Fischer N R Pereira et al ldquoEffectsof non-surgical periodontal treatment on the L-arginine-nitricoxide pathway and oxidative status in plateletsrdquo ExperimentalBiology and Medicine vol 238 no 6 pp 713ndash722 2013
[66] P Mydel Y Takahashi H Yumoto et al ldquoRoles of the hostoxidative immune response and bacterial antioxidant rubr-erythrin during Porphyromonas gingivalis infectionrdquo PLoSpathogens vol 2 no 7 p e76 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Mediators of Inflammation
to 1 O2 balanced with 5 CO
2and N
2in the incuba-
tion atmosphere (INCO 153 O2CO2incubator Memmert
GmbH Schwabach Germany) At the end of the experimentmedium was removed cells were washed three times withPBS and fixed with 250120583L cooled methanolaceton (1 1)at minus20∘C for 10min The solution was removed and cellsremained at room temperature for 10min to dry Afterblocking with 3 BSA-PBS for 10min cells were incubatedwith antibodies (Ab) directed against NOX4 (Abcam) CATor SOD (Abcam) 1 50 in PBS 3 BSA for 2 hours Cellswere washed with PBS before the species-specific secondaryAb (dilution 1 400 in PBS) were added for additional90min Cells were covered on slides with DAPI contain-ing mounting medium (Dako) On the following day cellswere scanned with the Zeiss Axio Imager A1 fluorescencemicroscope (HBO 100 Carl Zeiss Jena Germany) anAxioCam MRc camera (Carl Zeiss) and the AxioVision 47software (Carl Zeiss) Density analysis was performed usingthe freely available image-processing software ImageJ 143(httprsbinfonihgovij) Quantification of relative fluores-cence intensity was performed by measuring the intensity ofeach cell related to cell area To obtain grayscale pictures eachwas processed using the tools ldquocolorrdquo and ldquosplit channelsrdquoin ImageJ which resulted in red green and blue color-fractionated grayscale pictures for each channel Only greenfractionated pictures were used for calculation of the relativefluorescence intensity
27 Immunohistochemistry Periodontal tissues from clini-cally healthy patients (I) as well as those from patientswith the diagnosis of gingivitis (II) and periodontitis (III)were obtained from three different patients for each groupDiagnoses of gingivitis and periodontitis were performedby specialists from the Clinic of Dental Medicine Gin-givitis was characterized by a reversible local inflammatoryreaction of the gingiva visible as redness swelling andbleeding In contrast periodontitis affects four differentstructures gingiva periodontal ligament tooth and alveolarbone Therefore periodontitis was diagnosed by an increaseof probing depth bleeding and attachment loss whichwas supported by radiographs Tissues were fixed in 4phosphate-buffered paraformaldehyde for 24 h followed byhydration and dehydration in an ascending ethanol serieswhich ended in paraffin embedding Afterwards 4 120583m thickparaffin-embedded sections were prepared deparaffinizedrehydrated rinsed with TBS pH 74 for 10min and thensoaked in 70mL methanol containing 700mL of 30 H
2O2
for 45min in the dark to block endogenous peroxidase activ-ity Afterwards tissue sections were rinsed and preincubatedwith TBSBSA 4 for 20min to avoid unspecific backgroundstaining Incubation with monoclonal primary antibodies ofNOX4 (Abcam Cambridge UK) and polyclonal antibody ofcatalase and SOD (Abcam) in a 1 100 working solution ofTBSBSA was performed at 4∘C overnight in a humidifiedchamber Subsequently sections were washed in TBS andincubated with a peroxidase-labeled polymer conjugated toa goat anti-rabbit immunoglobulin provided as a ready-to-use solution (Envision Dako AS Glostrup Denmark)
for 30min in a humidified chamber at room temperatureAntibody complexeswere visualized using diaminobenzidinefor 10min resulting in brown staining Thereafter the slideswere rinsed counterstainedwithMayerrsquos hematoxylin rinsedagain and covered Negative controls were prepared by omis-sion of the primary antibodies as well as both the primaryand secondary antibody from the staining procedures usingTBSBSA instead Finally tissue sections were analyzed usingtheZeissAxioScop 2microscope (Carl Zeiss JenaGermany)
28 Statistical Analysis All experiments were performedin triplicate and mRNA quantification is given as meansplusmn SD Results were compared by ANOVA Dunnettrsquos andTukey-Kramer tests were performed as posttest to calculatewhether differences between treated and control groups weresignificant A 119875 value of 005 was considered as significant
3 Results
31 NOX4 Human primary PDL cells were incubated undernormoxic and hypoxic conditions in the presence or absenceof LPS-PG (1 120583gmL) Treatment with LPS-PG induced time-dependently a significant increase in the mRNA expressionof NOX4 compared to the untreated control (Figure 1(a)) Indetail NOX4mRNA levels weremarkedly increased after twohours of LPS-PG stimulation the increase reached significantlevels (4-fold) after 4 hours (119875 lt 0001) NOX4 mRNA levelspeaked after 8-hour stimulation with LPS-PG and returnedto base line levels afterwards
Hypoxia alone induced the expression of NOX4 in PDLcells but in contrast to the stimulation with LPS-PG themaximum in NOX4 mRNA upregulation was detected after4 hours (119875 lt 0001 Figure 1(a)) Subsequently NOX4mRNA expression decreased however it remained at ahigher level compared to the control group even after 24or 48 hours (Figure 1(a)) The highest increase in NOX4mRNA expression was detected after the combination ofboth stimuli (85 times) which was statistically significantcompared to control and even compared to LPS stimulationunder normoxia (119875 lt 0001 Figure 1(a))
To prove whether the increase in mRNA expression wasfollowed by an increase in protein content immunofluo-rescence stainings for NOX4 were performed NOX4 wasdetectable predominantly in the cytosol and to a slight extentin the nucleus of unstimulated PDL cells (Figure 1(b)) To getmore detailed information about the time course of NOX4protein induction after hypoxic and inflammatory stimula-tion we performed immunoblots After 2 hours only hypoxiaprovoked a significant increase in NOX4 protein (119875 lt 0001)which diminished afterwards (Figure 1(c)) Under normoxiccondition the treatment with LPS-PG induced a significantelevation of NOX4 protein after 4 hours of incubation com-pared to control and hypoxia alone (119875 lt 0001 Figure 1(c))The LPS-PG effect was evenmore pronounced under hypoxiccondition which was statistically significant compared to allthe other groups (119875 lt 0001 Figure 1(c)) After 24 hours ofincubation alterations of NOX4 protein levels were no longerdetectable (data not shown) These findings were in line with
Mediators of Inflammation 5
NADPH-oxidase 4 mRNA
125
100
75
50
25
00minus
minus
minus minus minus+
+ minus + minus + minus + minus + minus + minus + minus + minus + minus +
+ minus minus+ + minus minus+ + minus minus+ + + +
lowastlowast
lowast
lowast
2h 4h 8h 24h 48h
HoxLPS
(a) (b)
(c)
mRN
A ex
pres
sion
com
pare
d to
cont
rol
minusminus
minus minus
+
+
+minusminus
minus minusminus
+
+ +
+
lowast
lowast
Hox
NOX4 protein
Inte
grat
ed d
ensit
y (
) com
pare
dto
cont
rol (
au)
LPS
2h 4h
300
200
100100
50
0
Nox Hoxminus minus+ +
120573-Actin
NOX4
LPS
120573-Actin
NOX4
2h
4h
Figure 1 NOX4 in human primary periodontal ligament (PDL) cells (a) PDL cells were cultured under normoxic or hypoxic condition andstimulated with or without LPS of Porphyromonas gingivalis (1 120583gmL) NOX4 mRNA expression was analyzed after 2 4 8 24 and 48 hStatistical differences were analyzed by one-way ANOVA and post hoc Dunnett and Tukeyrsquos multiple comparison test 119875 lt 005 differenceto control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 9) (b) NOX4 protein was visualized by immunofluorescence as well asimmunohistochemical staining in unstimulated PDL fibroblasts (1 50 antibody from Abcam) (c) Immunoblot data of NOX4 in PDL cellscultured under normoxic or hypoxic condition (Hox) and stimulated with or without LPS of Porphyromonas gingivalis (1 120583gmL) NOX4protein levels were analyzed after 2 and 4 h Statistical analysis of western blot data was performed using the freely available image-processingsoftware ImageJ 143 (httprsbinfonihgovij) Statistical differences were analyzed by one-way ANOVA and post hoc Dunnett and Tukeyrsquosmultiple comparison test 119875 lt 005 difference to control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 3) The scale bars indicate200 120583m in the bright field survey and 50120583m in the immunofluorescence image respectively
our mRNA results even though we detected time-dependentdifferences
32 H2O2Formation To determine hydrogen peroxide gen-
eration we used the commercially available Amplex Red-horseradish peroxidase assay PDL cells displayed a lowconstitutive H
2O2production under normoxic conditions
We observed a significant upregulation of the H2O2con-
centration in the cell culture supernatant after 1 2 and 4hours of stimulation with LPS-PG Hypoxia alone induceda significant increase in H
2O2formation compared to the
time-matched controls (Figure 2) While in the LPS groupthe H
2O2values were the highest after 1 hour and decreased
continuously after 2 and 4 hours (Figure 2) under hypoxiccondition H
2O2increased permanently (Figure 2) Compar-
ing the LPS and the hypoxic effect revealed that hypoxiaalone induced a significantly higher H
2O2accumulation over
the complete observation period (119875 lt 0001) Howeverthe combination of both stimuli elicited the highest H
2O2
concentration after 1 hour which was significant not onlycompared to control but also compared to the LPS and thehypoxia group (Figure 2) After 2 hours the treatment withLPS-PG was no longer able to enhance the hypoxia-inducedH2O2formation it was still significantly increased compared
to control and the LPS stimulation but no longer comparedto the hypoxic groupThis was even more pronounced after 4hours
6 Mediators of Inflammation
2h 4h1h
50000
40000
30000
20000
10000
0
(120583M
)
HoxLPS
minus minus minus minus minus minus
minus
+
+minus +minus +minus +minus +minus +
++ + + +
lowast
lowastlowast lowast
lowast
lowast
H2O2 concentration
Figure 2 Determination of ROS formation (H2O2) using Amplex
Red assay PDL cells were cultured under normoxic (Nox) orhypoxic condition (Hox) and stimulated with or without LPS ofPorphyromonas gingivalis (1 120583gmL) H
2O2was analyzed after 1
2 and 4 hours Statistical differences were analyzed by one-wayANOVA and post hoc Dunnett and Tukeyrsquos multiple comparisontest 119875 lt 005 difference to control lowast119875 lt 005 difference betweengroups (means plusmn SD 119899 = 6)
33 Redox Systems To analyze which protectivemechanismsagainst ROS formation are involved in our setting wedetermined the redox system components catalase (CAT)and superoxide dismutase (SOD) in PDL cells after treat-ment with LPS-PG under normoxic and hypoxic conditionsby immunofluorescence (Figures 3(a)-3(b)) In additionperiodontal tissues from clinically healthy patients (I) aswell as from patients with the diagnosis of gingivitis (II)and periodontitis (III) were obtained from three differentpatients for each group and stained immunohistochemicallyfor NOX4 CAT and SOD (Figures 4 5 and 6)
331 Immunofluorescence of Redox Systems in PDL CellsPDL cells displayed a constitutive cytosolic catalase expres-sion under normoxic conditions Treatment with LPS-PGandhypoxia induced a change in PDL cellmorphologywhichwas first obvious after a 1-hour incubation under hypoxicconditions and after 2 hours with LPS-PG (Figure 3(a)) Asshown in Figure 3(b) analyzing catalase cytoplasm densityin relation to constant cell area revealed a significant upreg-ulation of catalase in PDL cells stimulated with LPS-PG orhypoxia after 1 h (119875 lt 0001 Figure 3(b)) This was also truefor hypoxic conditions after 2 h compared to all other groupsUnder inflammatory stimulation (LPS) a significant increaseof catalase cytoplasm density was detectable after 4 hourscompared to all the other groups (119875 lt 0001 Figure 3(b))Instead the combination of both stimuli diminished thehypoxic as well as LPS-induced effects after 1 2 and 4hours Determining superoxide dismutase protein expressionin PDL cells showed no significant differences at any timepoint (119875 gt 005 data not shown)
332 Immunohistochemistry of NOX4 and Redox Systemsin Periodontal Tissue Sections Periodontal tissues (healthygingiva and periodontal ligament (PDL) gingivitis andperiodontitis) were obtained from three different patients foreach group
NOX4 In the healthy periodontal tissue samples only veryweak immunostaining for NOX4 could be obtained (Figures4(a) and 4(b)) In the PDL weak immunoreactivity witha moderate staining of some fibroblasts was observed Ininflamed tissues from patients suffering from gingivitis orperiodontitis strong immunoreactivity for NOX4 appearedin the cytosol and even stronger in the nuclei of keratinocytesof the basal layer of inflamed gingival epithelium (Figures4(c)-4(d)) In gingival and PDL fibroblasts as well as inimmune cells like leukocytes and in vessels the immunore-activity was primarily restricted to the cytosol and cell walls(Figures 4(c)-4(d) see red arrows)
CAT Nearly no immunostaining could be found in thehealthy gingival epithelium (Figure 5(a)) In the healthyPDL predominantly PDL cells located near to the toothroot surfaces were stained (Figure 5(b) see red arrow) Ingingivitis tissue a weak staining of gingival keratinocytessimilar to healthy tissue could be seen (Figure 5(c)) Insteadin the tissue samples of patients with periodontitis a strongimmunoreactivity was observed in the subepithelial layerprobably referring to nuclei staining of immune cells likeleukocytes (Figure 5(d) see red arrows)
SOD As for healthy tissues gingival epithelium showeda moderate to strong immunoreactivity of keratinocyteswhich was primarily limited to the cytosol (Figure 6(a)see red arrow) In healthy periodontium gingival and PDLfibroblasts were weakly stained (Figure 6(b)) Compared toinflamed tissues basal epithelial cells seemed to be stainedmore intensively in the gingivitis specimen (Figures 6(c)-6(d)) In addition the subepithelium revealed immunore-activity in local cells like gingival fibroblasts and scatteredimmune cells (Figure 6(c)) In periodontitis stained sectionsSOD immunostaining seemed to be limited to the gingivalepithelium (Figure 6(d) see red arrows) In inflamed tissuessubepithelial layers showed no signs of immunoreactivity
4 Discussion
Oxidative stress is discussed as an important cofactor in theetiology and pathogenesis of several oral and dental diseasesfor example in inflammatory processes like periodontitisFirst evidence for the presence and role of ROS in theperiodontium was given by the detection of these moleculesafter respiratory burst by polymorphonuclear lymphocytes(PMNLs) in periodontal damage [22] Since then the impli-cation of ROS and their degrading enzymes SOD or CAT inthe pathogenesis of periodontal diseases has been supportedby several groups [23ndash26] To analyse whether cellular redoxsystems in the periodontium are affected by inflammatoryconditions we investigated the effect of hypoxia and LPSfrom Porphyromonas gingivalis (LPS-PG) on periodontal
Mediators of Inflammation 7
1h 2hCatalase
Nox
Nox + LPS
Hox
Hox + LPS
4h
100120583m 100120583m 100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
(a)
2h 4h
Hox
Inte
grat
ed d
ensit
y (a
u)
Catalase cytoplasm density
LPSminusminus + + minusminus + + minusminus +
minus +minus +minus +minus +minus +minus +
+
lowast
lowast
lowast
lowast
lowastlowast
1h
20000
25000
15000
1000010000
7500
5000
2500
0
(b)
Figure 3 Catalase (CAT) in PDL cells (a) CAT was visualized by immunofluorescence staining (1 50 antibody from Abcam) PDL cellswere cultured under normoxic (Nox) or hypoxic condition (Hox) and stimulated with or without LPS of Porphyromonas gingivalis (1120583gmL)Catalase cytoplasm density (b) was determined in relation to cell area using the freely available image-processing software ImageJ 143(httprsbinfonihgovij) Statistical analysis of immunofluorescence data was analyzed by one-way ANOVA and post hoc Dunnett andTukeyrsquos multiple comparison test 119875 lt 005 difference to control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 6) The scale barsindicate 100 120583m in the immunofluorescence images
8 Mediators of Inflammation
50120583m
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m
50120583m
50120583m
Inflamed
(d)
Figure 4 NADPH-oxidase 4 (NOX4) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Monoclonal primary antibody against NOX4 (Abcam Cambridge UK) was used ina concentration of 1 100 In gingival and PDL fibroblasts as well as in immune cells like leukocytes and in vessels the immunoreactivity wasprimarily restricted to the cytosol and cell walls Red arrows indicate examples for the typical distribution (Figures 4(c) and 4(d)) The scalebars indicate 200120583m in the surveys and 50 120583m in the higher magnifications respectively
ligament cells We showed that both stimuli induced asignificant increase of NOX4 and a significant upregulationof hydrogen peroxide generation NOX enzymes were firstdescribed in phagocytes as being responsible for defensemechanisms against pathogens using NOX-generated ROSmolecules Till then NOX4 expression has been detected innonphagocyte cell types like kidney cells osteoclasts andosteoblasts as well as dermal cells [9 11 47] In PDL cellshypoxia and LPS-PG induced a significant increase of NOX4mRNA The protein expression was mainly located in thecytosol but likewise in the nucleus Our findings were inline with Brown and Griendling who have shown that thetypically transmembrane molecule is localized in differentcell compartments [9]
NOX4 is one of the major sources of ROS generationwhich is highly and ubiquitously expressed compared tothe other family members Mandal and colleagues demon-strated that apart from its antimicrobial function NOX4 isresponsible not only for osteoclast but also for preosteoblastdifferentiation via ROS accumulation reinforcing the ideaof NOX4 as a key regulator in bone remodeling underphysiological ROS amounts [47] Animal studies supportthe hypothesis that massive ROS formation induced bone
resorption via NOX4 [20 21] Whereas physiological levelsof NOX4-generated ROS in PDL cellsmight facilitate alveolarbone remodeling massive ROS accumulation during inflam-matory periodontal diseases might lead to alveolar bonedestruction favoring tooth loss We have demonstrated firstthat NOX4 is upregulated in periodontal ligament fibroblastsduring bacterial and hypoxic events Hence PDL fibroblastsmay be a source of excessive ROS generation and are thereforeof particular importance during periodontal diseases
Our results support current findings determining the roleof oxidative stress in different periodontal pathologies whichimplicate that the balance between NADPH-oxidases andredox systems is crucial to maintain cell and tissue home-ostasis [22ndash26] However there are only a few studies usinghuman periodontal fibroblast in the context of oxidativestress and none of them evaluated the role of NOX4 andcounteracting redox systems during inflammatory conditionswith LPS-PG stimulation and hypoxia An increase of ROSgeneration was detected by San Miguel et al following PDLfibroblast stimulation with nicotine or ethanol which wereabolished after antioxidant treatment [40] They completedtheir results using metal ions like nickel for PDL stimulationwhich increases ROS activity in these cells indicating a role
Mediators of Inflammation 9
200120583m
50120583m
50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 5 Catalase (CAT) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontal ligament (b) andsamples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University of Bonn and parentalas well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against catalase (Abcam Cambridge UK) was used in a concentration of1 100 No immunostaining could be found in the healthy gingival epithelium In the healthy PDL predominantly PDL cells located near tothe tooth root surfaces were stained (Figure 5(b) red arrow) In gingivitis tissue a weak staining of gingival keratinocytes similar to healthytissue could be seen (Figure 5(c)) Instead in the tissue samples of patients with periodontitis a strong immunoreactivity was observed inthe subepithelial layer probably referring to nuclei staining of immune cells like leukocytes Red arrows indicate examples for the typicallocalisation (Figure 5(d)) The scale bars indicate 200 120583m in the surveys and 50120583m in the higher magnifications respectively
in metal cytotoxicity or even nickel allergy [41] Transientmetal ions like nickel and cobalt are known to mimic cellularhypoxia mainly due to the inhibition of prolyl hydrolases(PDHs) and asparaginyl hydrolase (factor-inhibiting HIF(FIH)) The inhibition of hydroxylation leads to HIF-1120572nuclear translocation followed by the transcription of diversegenes which are implicated in hypoxic adaptation immuneresponse or angiogenesis therefore indirectly supporting ourresults under hypoxic condition [48 49]
We could show that hypoxia as well as LPS-PG inducedROS formation by detecting H
2O2using the commercially
available Amplex Red assay The high levels of H2O2released
under hypoxic conditions after 1 and 2 hours in PDL cellsmay be caused by different mechanisms First we founda significant upregulation of the mRNA and protein ofthe H
2O2-generating enzyme NOX4 Second the lack of
molecular oxygen as the final electron acceptor in therespiratory chain may be responsible for the early release of
ROS LPS-PG alone induced acutely a small but significantincrease in H
2O2release and enhanced the hypoxia-induced
generation In human gingival fibroblasts similar results wereobtained showing the increase of ROS and antioxidants afterP gingivalis or LPS stimulation [26 42 43] Interestinglydespite the increase in NOX4 mRNA and protein expressionafter 4 hours of stimulation with LPS-PG under hypoxicconditions the H
2O2generation was significantly reduced
when compared to hypoxia alone An additional inhibitionof the respiratory chain by LPS-PG which was described forcardiac mitochondria [50] is therefore conceivable
To antagonize oxidative stress cells developed severalantioxidative enzyme systems Therefore we measured SODand CAT expression in PDL cells and periodontal tissues byimmunofluorescence and immunohistochemistry SODs aremetalloenzymes which are responsible for the dismutation ofsuperoxide radicals to hydrogen peroxide [51] The reductionof hydrogen peroxide to water and oxygen is subsequently
10 Mediators of Inflammation
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m 50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 6 Superoxide dismutase (SOD) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against SOD (Abcam Cambridge UK) was used ina concentration of 1 100 Gingival epithelium showed a moderate to strong staining of SOD in keratinocytes which was primarily limitedto the cytosol (Figure 6(a) and red arrow depicts the typical localization) In the healthy periodontium gingival and PDL fibroblasts wereweakly stained (Figure 6(b)) In inflamed tissues basal epithelial cells seemed to be stained more intensively in the gingivitis specimen thanin periodontitis (Figures 6(c) and 6(d)) In addition the subepithelium revealed immunoreactivity in local cells like gingival fibroblasts andscattered immune cells (Figure 6(c) see red arrow) In periodontitis SOD immunostaining seemed to be limited to the gingival epithelium(Figure 6(d) see red arrow) The scale bars indicate 200 120583m in the surveys and 50 120583m in the higher magnifications respectively
catalyzed by CATs which are ubiquitously expressed Bothenzymes are essential to maintain cellular redox homeostasisThe present data imply that PDL cells and periodontal tissueshave fundamental defense machineries against bacteria rep-resented by NOX4-dependent inducible ROS generation aswell as protective redox systems This is of great importanceconcerning the high amounts and diversity of the oral floraDifferent studies indicate that redox systems are key playersin the development of periodontal diseases and markers forperiodontal health status [52ndash56] We detected a significantincrease of catalase in PDL cells in response to LPS aswell as hypoxia after 1 and 2 hours In our setting PDLfibroblasts seem to counteracted LPS- and hypoxia-inducedROS formation with the elevation of CAT expression Butprolonged incubation under hypoxic conditions failed toincrease catalase Since ROS generation was still elevated atthese later time points a delay in ROS degradation may be
the explanation Taken together the dramatic elevation inROS formation by the combination of hypoxia and LPS-PGon one hand and the reduction of catalase under the sameconditions on the other hand increase the oxidative stress inthe periodontium favoring the progression of inflammationand bone resorption and finally may lead to tooth loss
The fundamental role of the balance between ROSgeneration and antioxidative mechanisms in periodontalinflammatory processes was supported by our immunohis-tochemical findings In patients suffering from periodontalinflammatory diseases like gingivitis and periodontitis wedemonstrated that NOX4 is upregulated and thus likelyresponsible for H
2O2generation However we detected as
well an increase in catalase abundance in inflamed gingivaland periodontal tissue which was in line with data fromKrifka and colleagues who detected high CAT levels inmonomer-exposed macrophages as a response to oxidative
Mediators of Inflammation 11
stress [57] In our in vitro setting PDL fibroblasts seemto counteract LPS- and hypoxia-induced ROS formationpredominantly with an elevation of CAT expression sincewe could not detect any SOD immunoreactivity The roleand impact of the antioxidative systems SOD and CAT arecontroversially discussed Recent studies measured CAT andSOD activities in gingival tissues of patients with periodon-titis [58 59] Borges Jr et al did not find differences inCAT activities comparing healthy and diseased patients [58]Instead Tonguc et al detected decreased local SOD andCAT activities in periodontitis patients which seemed to beincreased by smoking [59] In contrast in inflamed tissues weobservedhigh amounts of SODwhichwas localized primarilyin epithelial cells Jacoby and Davis determined SOD in peri-odontal soft tissues They detected twice as much SOD activ-ity in the periodontal ligament as in the human skin (dermis)[60] Furthermore immunohistochemical analysis revealedthat SOD is localized in the periphery of matrix collagenfibrils and to the glycocalyx of tissue fibroblasts This wasconfirmed by Skaleric et al [42] which might explain the factthat we did not detect significant differences of SOD levelsafter hypoxic and LPS-PG stimulation intracellularly in PDLfibroblasts Another reason for these results may be causedby the age of the PDL cells used in this study We explantedthe PDL cells from young and oral healthy patients but mostof the immunohistochemical stainings were performed intissues from elderly patients with clearly diagnosed gingivitisor periodontitisTherefore it is possible that in PDL cells fromelderly patients a different expression pattern of antioxidantenzyme systems could be found which further deterioratethe redox state of the periodontal ligament Moreover otherauthors found reduced antioxidants in the body fluids ofpatients with periodontitis indicating a dysregulation whichwould favor disease progression [23 24 61] A variety of invitro and animal studies analyzed the effect of antioxidantsupplement in the context of inflammatory or cytotoxicevents [57 62 63] They have shown that antioxidantsinhibit these processes indicating a therapeutic approachin pathologies like periodontitis First clinical investigationsconfirmed the efficiency of antioxidant micronutrients andadjuncts in periodontal diseases [22 54 56 64 65] Thepresent data indirectly support these studies
Concerning periodontal pathogen invasiveness and viru-lence there is compelling evidence that P gingivalis possessesits own protective antioxidants for example rubrerythrindefending them against the oxidative burst of the host [66]We have shown that its virulence factor LPS-PG inducedhigh amounts of ROS via NOX4 which may result in boneresorption favoring the invasiveness of the pathogen whichis at the same time protected by its own defense machineryThis would further explain the pathogenicity and resistanceof this bacterium strain in patients with periodontitis
5 Conclusion
In this study we have demonstrated that LPS from Porphy-romonas gingivalis (LPS-PG) as well as hypoxia induces aNOX4-dependent increase inH
2O2release in PDLfibroblasts
which may contribute to the development and progressionof periodontal diseases when not balanced by a concomitantincrease in antioxidative systems The dramatic elevationin ROS formation induced by the combination of hypoxiaand LPS-PG and the reduction of catalase under the sameconditions increased the oxidative stress in the periodontiumfavoring the progression of inflammation bone resorptionand possibly tooth loss
Taken together the presented data further explain thedestructive mechanism of hypoxic and inflammatory condi-tions in the periodontium and may improve diagnostic andtherapeutic strategies in periodontal diseases
Conflict of Interests
The authors have no conflict of interests to disclose
Acknowledgments
The authors would like to thank Ms Tina Schaffrath for hertechnical supportThe present studywas supported by a grantfrom the German Research Foundation (Clinical ResearchUnit 208TP7) and the Medical Faculty of the University ofBonn
References
[1] S Reuter S C Gupta M M Chaturvedi and B B AggarwalldquoOxidative stress inflammation and cancer how are theylinkedrdquo Free Radical Biology and Medicine vol 49 no 11 pp1603ndash1616 2010
[2] H Sone H Akanuma and T Fukuda ldquoOxygenomics inenvironmental stressrdquo Redox Report vol 15 no 3 pp 98ndash1142010
[3] K E Wellen and C B Thompson ldquoCellular metabolic stressconsidering how cells respond to nutrient excessrdquo MolecularCell vol 40 no 2 pp 323ndash332 2010
[4] R O Poyton K A Ball and P R Castello ldquoMitochondrialgeneration of free radicals and hypoxic signalingrdquo Trends inEndocrinology andMetabolism vol 20 no 7 pp 332ndash340 2009
[5] F Weinberg and N S Chandel ldquoReactive oxygen species-dependent signaling regulates cancerrdquo Cellular and MolecularLife Sciences vol 66 no 23 pp 3663ndash3673 2009
[6] A Cataldi ldquoCell responses to oxidative stressorsrdquo CurrentPharmaceutical Design vol 16 no 12 pp 1387ndash1395 2010
[7] Z Durackova ldquoSome current insights into oxidative stressrdquoPhysiological Research vol 59 no 4 pp 459ndash469 2010
[8] L A Sena and N S Chandel ldquoPhysiological roles of mitochon-drial reactive oxygen speciesrdquoMolecular Cell vol 48 no 2 pp158ndash166 2012
[9] D I Brown and K K Griendling ldquoNox proteins in signaltransductionrdquo Free Radical Biology amp Medicine vol 47 no 9pp 1239ndash1253 2009
[10] K Bedard and K-H Krause ldquoThe NOX family of ROS-generatingNADPHoxidases physiology and pathophysiologyrdquoPhysiological Reviews vol 87 no 1 pp 245ndash313 2007
[11] K-H Krause ldquoTissue distribution and putative physiologicalfunction of NOX family NADPH oxidasesrdquo Japanese Journal ofInfectious Diseases vol 57 no 5 pp S28ndashS29 2004
12 Mediators of Inflammation
[12] S Yang Y Zhang W Ries and L Key ldquoExpression of Nox4 inosteoclastsrdquo Journal of Cellular Biochemistry vol 92 no 2 pp238ndash248 2004
[13] E Pedruzzi C Guichard V Ollivier et al ldquoNAD(P)H oxidaseNox-4mediates 7-ketocholesterol-induced endoplasmic reticu-lum stress and apoptosis in human aortic smooth muscle cellsrdquoMolecular and Cellular Biology vol 24 no 24 pp 10703ndash107172004
[14] J Hwang A Saha Y C Boo et al ldquoOscillatory shear stress stim-ulates endothelial production of O
2-from p47119901ℎ119900119909-dependent
NAD(P)Hoxidases leading tomonocyte adhesionrdquoThe Journalof Biological Chemistry vol 278 no 47 pp 47291ndash47298 2003
[15] H B Suliman M Ali and C A Piantadosi ldquoSuperoxidedismutase-3 promotes full expression of the EPO response tohypoxiardquo Blood vol 104 no 1 pp 43ndash50 2004
[16] P Vallet Y Charnay K Steger et al ldquoNeuronal expressionof the NADPH oxidase NOX4 and its regulation in mouseexperimental brain ischemiardquo Neuroscience vol 132 no 2 pp233ndash238 2005
[17] K T Moe S Aulia F Jiang et al ldquoDifferential upregulation ofNox homologues of NADPH oxidase by tumor necrosis factor-120572 in human aortic smooth muscle and embryonic kidney cellsrdquoJournal of Cellular andMolecularMedicine vol 10 no 1 pp 231ndash239 2006
[18] I Diebold A Petry J Hess and A Gorlach ldquoThe NADPHoxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1rdquoMolecular Biology of the Cell vol 21 no 12pp 2087ndash2096 2010
[19] K V Tormos and N S Chandel ldquoInter-connection betweenmitochondria and HIFsrdquo Journal of Cellular and MolecularMedicine vol 14 no 4 pp 795ndash804 2010
[20] T Ohnishi K Bandow K Kakimoto M Machigashira TMatsuyama and T Matsuguchi ldquoOxidative stress causes alve-olar bone loss in metabolic syndrome model mice with type 2diabetesrdquo Journal of Periodontal Research vol 44 no 1 pp 43ndash51 2009
[21] C Goettsch A Babelova O Trummer et al ldquoNADPH oxidase4 limits bone mass by promoting osteoclastogenesisrdquo Journal ofClinical Investigation vol 123 no 11 pp 4731ndash4738 2013
[22] I L C Chapple and J B Matthews ldquoThe role of reactive oxygenand antioxidant species in periodontal tissue destructionrdquoPeriodontology 2000 vol 43 no 1 pp 160ndash232 2007
[23] A Guentsch P M Preshaw S Bremer-Streck G KlingerE Glockmann and B W Sigusch ldquoLipid peroxidation andantioxidant activity in saliva of periodontitis patients effect ofsmoking and periodontal treatmentrdquo Clinical Oral Investiga-tions vol 12 no 4 pp 345ndash352 2008
[24] R Agnihotri P Pandurang S U Kamath et al ldquoAssociation ofcigarette smoking with superoxide dismutase enzyme levels insubjected with chonic periodontitisrdquo Journal of Periodontologyvol 80 no 4 pp 657ndash662 2009
[25] D Ekuni T Tomofuji N Tamaki et al ldquoMechanical stimulationof gingiva reduces plasma 8-OHdG level in rat periodontitisrdquoArchives of Oral Biology vol 53 no 4 pp 324ndash329 2008
[26] H Staudte A Guntsch A Volpel and B W Sigusch ldquoVitaminC attenuates the cytotoxic effects of Porphyromonas gingivalison human gingival fibroblastsrdquo Archives of Oral Biology vol 55no 1 pp 40ndash45 2010
[27] P Bullon H N Newman and M Battino ldquoObesity diabetesmellitus atherosclerosis and chronic periodontitis a sharedpathology via oxidative stress andmitochondrial dysfunctionrdquoPeriodontology 2000 vol 64 no 1 pp 139ndash153 2014
[28] G R Naveh V Brumfeld R Shahar and S Weiner ldquoToothperiodontal ligament direct 3D microCT visualization of thecollagen network and how the network changes when the toothis loadedrdquo Journal of Structural Biology vol 181 no 2 pp 108ndash115 2013
[29] Y Kitase M Yokozeki S Fujihara et al ldquoAnalysis of geneexpression profiles in human periodontal ligament cells underhypoxia the protective effect of CC chemokine ligand 2 tooxygen shortagerdquoArchives of Oral Biology vol 54 no 7 pp 618ndash624 2009
[30] G Thornton-Evans P Eke L Wei et al ldquoPeriodontitis amongadults aged ge 30 years - United States 2009-2010rdquo Morbidityand Mortality Weekly Report (MMWR) vol 62 supplement 3pp 129ndash135 2013
[31] S S Socransky C Smith and A D Haffajee ldquoSubgingivalmicrobial profiles in refractory periodontal diseaserdquo Journal ofClinical Periodontology vol 29 no 3 pp 260ndash268 2002
[32] K-T Ng J-P Li K M Ng G L Tipoe W K Leung and M-L Fung ldquoExpression of hypoxia-inducible factor-1120572 in humanperiodontal tissuerdquo Journal of Periodontology vol 82 no 1 pp136ndash141 2011
[33] S Frede U Berchner-Pfannschmidt and J Fandrey ldquoReg-ulation of Hypoxia-inducible factors during inflammationrdquoMethods in Enzymology vol 435 pp 405ndash419 2007
[34] J Shin S Ji and Y Choi ldquoAbility of oral bacteria to inducetissue-destructive molecules from human neutrophilsrdquo OralDiseases vol 14 no 4 pp 327ndash334 2008
[35] S Takashiba K Naruishi and Y Murayama ldquoPerspectiveof cytokine regulation for periodontal treatment fibroblastbiologyrdquo Journal of Periodontology vol 74 no 1 pp 103ndash1102003
[36] A Konermann D Stabenow P A Knolle S A E HeldJ Deschner and A Jager ldquoRegulatory role of periodontalligament fibroblasts for innate immune cell function and dif-ferentiationrdquo Innate Immunity vol 18 no 5 pp 745ndash752 2012
[37] S Kar S Subbaram P M Carrico and J A Melendez ldquoRedox-control of matrix metalloproteinase-1 a critical link betweenfree radicals matrix remodeling and degenerative diseaserdquoRespiratory Physiology andNeurobiology vol 174 no 3 pp 299ndash306 2010
[38] F Rousset M V C Nguyen L Grange F Morel and B LardyldquoHeme oxygenase-1 regulates matrix metalloproteinase MMP-1secretion and chondrocyte cell death via Nox4NADPH oxidaseactivity in chondrocytesrdquo PLoS ONE vol 8 no 6 Article IDe66478 2013
[39] E Rudolf and M Cervinka ldquoResponses of human gingival andperiodontal fibroblasts to a low-zinc environmentrdquo Alternativesto Laboratory Animals vol 38 no 2 pp 119ndash138 2010
[40] S M San Miguel L A Opperman E P Allen J Zielinski andK K H Svoboda ldquoBioactive polyphenol antioxidants protectoral fibroblasts from ROS-inducing agentsrdquo Archives of OralBiology vol 57 no 12 pp 1657ndash1667 2012
[41] S M San Miguel L A Opperman E P Allen J E Zielinskiand K K H Svoboda ldquoAntioxidant combinations protect oralfibroblasts against metal-induced toxicityrdquo Archives of OralBiology vol 58 no 3 pp 299ndash310 2013
[42] U Skaleric C M Manthey S E Mergenhagen B Gaspircand S M Wahl ldquoSuperoxide release and superoxide dismutaseexpression by human gingival fibroblastsrdquo European Journal ofOral Sciences vol 108 no 2 pp 130ndash135 2000
Mediators of Inflammation 13
[43] D Y Kim J-H Jun H-L Lee et al ldquoN-acetylcysteine preventsLPS-Lnduced pro-inflammatory cytokines andMMP2 produc-tion in gingival fibroblastsrdquoArchives of Pharmacal Research vol30 no 10 pp 1283ndash1292 2007
[44] B Rath-Deschner J Deschner S Reimann A Jager andW Gotz ldquoRegulatory effects of biomechanical strain on theinsulin-like growth factor system in human periodontal cellsrdquoJournal of Biomechanics vol 42 no 15 pp 2584ndash2589 2009
[45] P Chomczynski and N Sacchi ldquoSingle-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextractionrdquo Analytical Biochemistry vol 162 no 1 pp 156ndash1591987
[46] B Zhao K Ranguelova J Jiang and R P Mason ldquoStudies onthe photosensitized reduction of resorufin and implications forthe detection of oxidative stress with Amplex Redrdquo Free RadicalBiology and Medicine vol 51 no 1 pp 153ndash159 2011
[47] C C Mandal S Ganapathy Y Gorin et al ldquoReactive oxygenspecies derived from Nox4 mediate BMP2 gene transcriptionand osteoblast differentiationrdquoBiochemical Journal vol 433 no2 pp 393ndash402 2011
[48] J Fandrey S Frede and W Jelkmann ldquoRole of hydrogenperoxide in hypoxia-induced erythropoietin productionrdquo Bio-chemical Journal vol 303 no 2 pp 507ndash510 1994
[49] F Cervellati C Cervellati A Romani et al ldquoHypoxia inducescell damage via oxidative stress in retinal epithelial cellsrdquo FreeRadical Research vol 48 no 3 pp 303ndash312 2014
[50] V Vanasco N D Magnani M C Cimolai et al ldquoEndotoxemiaimpairs heart mitochondrial function by decreasing electrontransfer ATP synthesis and ATP content without affectingmembrane potentialrdquo Journal of Bioenergetics and Biomem-branes vol 44 no 2 pp 243ndash252 2012
[51] R G Alscher N Erturk and L S Heath ldquoRole of superoxidedismutases (SODs) in controlling oxidative stress in plantsrdquoJournal of Experimental Botany vol 53 no 372 pp 1331ndash13412002
[52] E Baltacıoglu P Yuva G Aydın et al ldquoLipid peroxidation levelsand total oxidantantioxidant status in serum and saliva frompatients with chronic and aggressive periodontitis Oxidativestress index a new biomarker for periodontal diseaserdquo Journalof Periodontology vol 85 no 10 pp 1432ndash1441 2014
[53] I L C Chapple M R Milward N Ling-Mountford et alldquoAdjunctive daily supplementation with encapsulated fruitvegetable and berry juice powder concentrates and clinicalperiodontal outcomes a double-blind RCTrdquo Journal of ClinicalPeriodontology vol 39 no 1 pp 62ndash72 2012
[54] A Jenzsch S Eick F Rassoul R Purschwitz and H JentschldquoNutritional intervention in patients with periodontal diseaseClinical immunological and microbiological variables during12 monthsrdquo British Journal of Nutrition vol 101 no 6 pp 879ndash885 2009
[55] N Novakovic T Todorovic M Rakic et al ldquoSalivary antiox-idants as periodontal biomarkers in evaluation of tissue statusand treatment outcomerdquo Journal of Periodontal Research vol49 no 1 pp 129ndash136 2014
[56] N Singh S C Narula R K Sharma S Tewari and P KSehgal ldquoVitamin e supplementation superoxide dismutasestatus and outcome of scaling and root planing in patients withchronic periodontitis a randomized clinical trialrdquo Journal ofPeriodontology vol 85 no 2 pp 242ndash249 2014
[57] S Krifka K-A Hiller G Spagnuolo A Jewett G Schmalz andH Schweikl ldquoThe influence of glutathione on redox regulation
by antioxidant proteins and apoptosis in macrophages exposedto 2-hydroxyethyl methacrylate (HEMA)rdquo Biomaterials vol 33no 21 pp 5177ndash5186 2012
[58] I Borges Jr E A M Moreira D W Filho T B de Oliveira MB S da Silva and T S Frode ldquoProinflammatory and oxidativestress markers in patients with periodontal diseaserdquo Mediatorsof Inflammation vol 2007 Article ID 45794 5 pages 2007
[59] M O Tonguc O Ozturk R Sutcu et al ldquoThe impact of smok-ing status on antioxidant enzyme activity and malondialdehydelevels in chronic periodontitisrdquo Journal of Periodontology vol82 no 9 pp 1320ndash1328 2011
[60] B H Jacoby and W L Davis ldquoThe electron microscopicimmunolocalization of a copper-zinc superoxide dismutase inassociation with collagen fibers of periodontal soft tissuesrdquoJournal of Periodontology vol 62 no 7 pp 413ndash420 1991
[61] T Konopka K Krol W Kopec and H Gerber ldquoTotal antioxi-dant status and 8-hydroxy-21015840-deoxyguanosine levels in gingi-val and peripheral blood of periodontitis patientsrdquo ArchivumImmunologiae et Therapiae Experimentalis vol 55 no 6 pp417ndash422 2007
[62] M Soory ldquoRedox status in periodontal and systemic inflamma-tory conditions including associated neoplasias antioxidants asadjunctive therapyrdquo Infectious DisordersmdashDrug Targets vol 9no 4 pp 415ndash427 2009
[63] M Soory ldquoOxidative stress inducedmechanisms in the progres-sion of periodontal diseases and cancer a common approach toredox homeostasisrdquo Cancers vol 2 no 2 pp 670ndash692 2010
[64] R V Chandra G Srinivas A A Reddy et al ldquoLocally deliveredantioxidant gel as an adjunct to nonsurgical therapy improvesmeasures of oxidative stress and periodontal diseaserdquo Journal ofPeriodontal and Implant Science vol 43 no 3 pp 121ndash129 2013
[65] M A D S Siqueira R G Fischer N R Pereira et al ldquoEffectsof non-surgical periodontal treatment on the L-arginine-nitricoxide pathway and oxidative status in plateletsrdquo ExperimentalBiology and Medicine vol 238 no 6 pp 713ndash722 2013
[66] P Mydel Y Takahashi H Yumoto et al ldquoRoles of the hostoxidative immune response and bacterial antioxidant rubr-erythrin during Porphyromonas gingivalis infectionrdquo PLoSpathogens vol 2 no 7 p e76 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
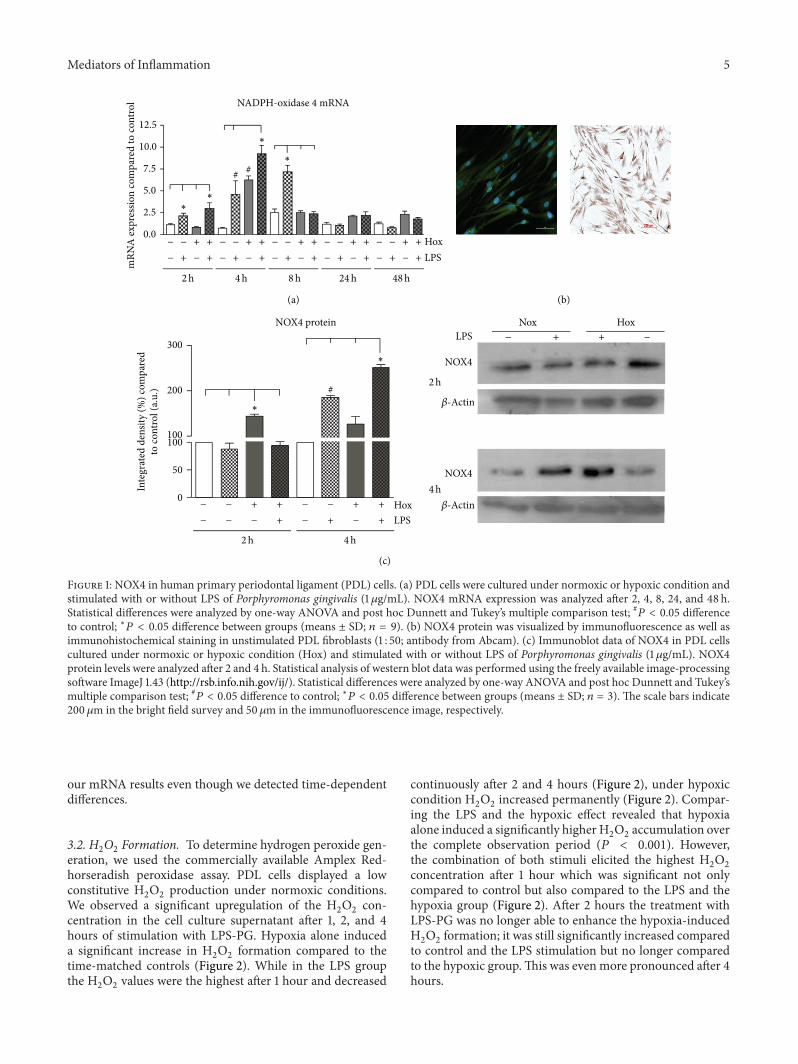
Mediators of Inflammation 5
NADPH-oxidase 4 mRNA
125
100
75
50
25
00minus
minus
minus minus minus+
+ minus + minus + minus + minus + minus + minus + minus + minus + minus +
+ minus minus+ + minus minus+ + minus minus+ + + +
lowastlowast
lowast
lowast
2h 4h 8h 24h 48h
HoxLPS
(a) (b)
(c)
mRN
A ex
pres
sion
com
pare
d to
cont
rol
minusminus
minus minus
+
+
+minusminus
minus minusminus
+
+ +
+
lowast
lowast
Hox
NOX4 protein
Inte
grat
ed d
ensit
y (
) com
pare
dto
cont
rol (
au)
LPS
2h 4h
300
200
100100
50
0
Nox Hoxminus minus+ +
120573-Actin
NOX4
LPS
120573-Actin
NOX4
2h
4h
Figure 1 NOX4 in human primary periodontal ligament (PDL) cells (a) PDL cells were cultured under normoxic or hypoxic condition andstimulated with or without LPS of Porphyromonas gingivalis (1 120583gmL) NOX4 mRNA expression was analyzed after 2 4 8 24 and 48 hStatistical differences were analyzed by one-way ANOVA and post hoc Dunnett and Tukeyrsquos multiple comparison test 119875 lt 005 differenceto control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 9) (b) NOX4 protein was visualized by immunofluorescence as well asimmunohistochemical staining in unstimulated PDL fibroblasts (1 50 antibody from Abcam) (c) Immunoblot data of NOX4 in PDL cellscultured under normoxic or hypoxic condition (Hox) and stimulated with or without LPS of Porphyromonas gingivalis (1 120583gmL) NOX4protein levels were analyzed after 2 and 4 h Statistical analysis of western blot data was performed using the freely available image-processingsoftware ImageJ 143 (httprsbinfonihgovij) Statistical differences were analyzed by one-way ANOVA and post hoc Dunnett and Tukeyrsquosmultiple comparison test 119875 lt 005 difference to control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 3) The scale bars indicate200 120583m in the bright field survey and 50120583m in the immunofluorescence image respectively
our mRNA results even though we detected time-dependentdifferences
32 H2O2Formation To determine hydrogen peroxide gen-
eration we used the commercially available Amplex Red-horseradish peroxidase assay PDL cells displayed a lowconstitutive H
2O2production under normoxic conditions
We observed a significant upregulation of the H2O2con-
centration in the cell culture supernatant after 1 2 and 4hours of stimulation with LPS-PG Hypoxia alone induceda significant increase in H
2O2formation compared to the
time-matched controls (Figure 2) While in the LPS groupthe H
2O2values were the highest after 1 hour and decreased
continuously after 2 and 4 hours (Figure 2) under hypoxiccondition H
2O2increased permanently (Figure 2) Compar-
ing the LPS and the hypoxic effect revealed that hypoxiaalone induced a significantly higher H
2O2accumulation over
the complete observation period (119875 lt 0001) Howeverthe combination of both stimuli elicited the highest H
2O2
concentration after 1 hour which was significant not onlycompared to control but also compared to the LPS and thehypoxia group (Figure 2) After 2 hours the treatment withLPS-PG was no longer able to enhance the hypoxia-inducedH2O2formation it was still significantly increased compared
to control and the LPS stimulation but no longer comparedto the hypoxic groupThis was even more pronounced after 4hours
6 Mediators of Inflammation
2h 4h1h
50000
40000
30000
20000
10000
0
(120583M
)
HoxLPS
minus minus minus minus minus minus
minus
+
+minus +minus +minus +minus +minus +
++ + + +
lowast
lowastlowast lowast
lowast
lowast
H2O2 concentration
Figure 2 Determination of ROS formation (H2O2) using Amplex
Red assay PDL cells were cultured under normoxic (Nox) orhypoxic condition (Hox) and stimulated with or without LPS ofPorphyromonas gingivalis (1 120583gmL) H
2O2was analyzed after 1
2 and 4 hours Statistical differences were analyzed by one-wayANOVA and post hoc Dunnett and Tukeyrsquos multiple comparisontest 119875 lt 005 difference to control lowast119875 lt 005 difference betweengroups (means plusmn SD 119899 = 6)
33 Redox Systems To analyze which protectivemechanismsagainst ROS formation are involved in our setting wedetermined the redox system components catalase (CAT)and superoxide dismutase (SOD) in PDL cells after treat-ment with LPS-PG under normoxic and hypoxic conditionsby immunofluorescence (Figures 3(a)-3(b)) In additionperiodontal tissues from clinically healthy patients (I) aswell as from patients with the diagnosis of gingivitis (II)and periodontitis (III) were obtained from three differentpatients for each group and stained immunohistochemicallyfor NOX4 CAT and SOD (Figures 4 5 and 6)
331 Immunofluorescence of Redox Systems in PDL CellsPDL cells displayed a constitutive cytosolic catalase expres-sion under normoxic conditions Treatment with LPS-PGandhypoxia induced a change in PDL cellmorphologywhichwas first obvious after a 1-hour incubation under hypoxicconditions and after 2 hours with LPS-PG (Figure 3(a)) Asshown in Figure 3(b) analyzing catalase cytoplasm densityin relation to constant cell area revealed a significant upreg-ulation of catalase in PDL cells stimulated with LPS-PG orhypoxia after 1 h (119875 lt 0001 Figure 3(b)) This was also truefor hypoxic conditions after 2 h compared to all other groupsUnder inflammatory stimulation (LPS) a significant increaseof catalase cytoplasm density was detectable after 4 hourscompared to all the other groups (119875 lt 0001 Figure 3(b))Instead the combination of both stimuli diminished thehypoxic as well as LPS-induced effects after 1 2 and 4hours Determining superoxide dismutase protein expressionin PDL cells showed no significant differences at any timepoint (119875 gt 005 data not shown)
332 Immunohistochemistry of NOX4 and Redox Systemsin Periodontal Tissue Sections Periodontal tissues (healthygingiva and periodontal ligament (PDL) gingivitis andperiodontitis) were obtained from three different patients foreach group
NOX4 In the healthy periodontal tissue samples only veryweak immunostaining for NOX4 could be obtained (Figures4(a) and 4(b)) In the PDL weak immunoreactivity witha moderate staining of some fibroblasts was observed Ininflamed tissues from patients suffering from gingivitis orperiodontitis strong immunoreactivity for NOX4 appearedin the cytosol and even stronger in the nuclei of keratinocytesof the basal layer of inflamed gingival epithelium (Figures4(c)-4(d)) In gingival and PDL fibroblasts as well as inimmune cells like leukocytes and in vessels the immunore-activity was primarily restricted to the cytosol and cell walls(Figures 4(c)-4(d) see red arrows)
CAT Nearly no immunostaining could be found in thehealthy gingival epithelium (Figure 5(a)) In the healthyPDL predominantly PDL cells located near to the toothroot surfaces were stained (Figure 5(b) see red arrow) Ingingivitis tissue a weak staining of gingival keratinocytessimilar to healthy tissue could be seen (Figure 5(c)) Insteadin the tissue samples of patients with periodontitis a strongimmunoreactivity was observed in the subepithelial layerprobably referring to nuclei staining of immune cells likeleukocytes (Figure 5(d) see red arrows)
SOD As for healthy tissues gingival epithelium showeda moderate to strong immunoreactivity of keratinocyteswhich was primarily limited to the cytosol (Figure 6(a)see red arrow) In healthy periodontium gingival and PDLfibroblasts were weakly stained (Figure 6(b)) Compared toinflamed tissues basal epithelial cells seemed to be stainedmore intensively in the gingivitis specimen (Figures 6(c)-6(d)) In addition the subepithelium revealed immunore-activity in local cells like gingival fibroblasts and scatteredimmune cells (Figure 6(c)) In periodontitis stained sectionsSOD immunostaining seemed to be limited to the gingivalepithelium (Figure 6(d) see red arrows) In inflamed tissuessubepithelial layers showed no signs of immunoreactivity
4 Discussion
Oxidative stress is discussed as an important cofactor in theetiology and pathogenesis of several oral and dental diseasesfor example in inflammatory processes like periodontitisFirst evidence for the presence and role of ROS in theperiodontium was given by the detection of these moleculesafter respiratory burst by polymorphonuclear lymphocytes(PMNLs) in periodontal damage [22] Since then the impli-cation of ROS and their degrading enzymes SOD or CAT inthe pathogenesis of periodontal diseases has been supportedby several groups [23ndash26] To analyse whether cellular redoxsystems in the periodontium are affected by inflammatoryconditions we investigated the effect of hypoxia and LPSfrom Porphyromonas gingivalis (LPS-PG) on periodontal
Mediators of Inflammation 7
1h 2hCatalase
Nox
Nox + LPS
Hox
Hox + LPS
4h
100120583m 100120583m 100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
(a)
2h 4h
Hox
Inte
grat
ed d
ensit
y (a
u)
Catalase cytoplasm density
LPSminusminus + + minusminus + + minusminus +
minus +minus +minus +minus +minus +minus +
+
lowast
lowast
lowast
lowast
lowastlowast
1h
20000
25000
15000
1000010000
7500
5000
2500
0
(b)
Figure 3 Catalase (CAT) in PDL cells (a) CAT was visualized by immunofluorescence staining (1 50 antibody from Abcam) PDL cellswere cultured under normoxic (Nox) or hypoxic condition (Hox) and stimulated with or without LPS of Porphyromonas gingivalis (1120583gmL)Catalase cytoplasm density (b) was determined in relation to cell area using the freely available image-processing software ImageJ 143(httprsbinfonihgovij) Statistical analysis of immunofluorescence data was analyzed by one-way ANOVA and post hoc Dunnett andTukeyrsquos multiple comparison test 119875 lt 005 difference to control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 6) The scale barsindicate 100 120583m in the immunofluorescence images
8 Mediators of Inflammation
50120583m
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m
50120583m
50120583m
Inflamed
(d)
Figure 4 NADPH-oxidase 4 (NOX4) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Monoclonal primary antibody against NOX4 (Abcam Cambridge UK) was used ina concentration of 1 100 In gingival and PDL fibroblasts as well as in immune cells like leukocytes and in vessels the immunoreactivity wasprimarily restricted to the cytosol and cell walls Red arrows indicate examples for the typical distribution (Figures 4(c) and 4(d)) The scalebars indicate 200120583m in the surveys and 50 120583m in the higher magnifications respectively
ligament cells We showed that both stimuli induced asignificant increase of NOX4 and a significant upregulationof hydrogen peroxide generation NOX enzymes were firstdescribed in phagocytes as being responsible for defensemechanisms against pathogens using NOX-generated ROSmolecules Till then NOX4 expression has been detected innonphagocyte cell types like kidney cells osteoclasts andosteoblasts as well as dermal cells [9 11 47] In PDL cellshypoxia and LPS-PG induced a significant increase of NOX4mRNA The protein expression was mainly located in thecytosol but likewise in the nucleus Our findings were inline with Brown and Griendling who have shown that thetypically transmembrane molecule is localized in differentcell compartments [9]
NOX4 is one of the major sources of ROS generationwhich is highly and ubiquitously expressed compared tothe other family members Mandal and colleagues demon-strated that apart from its antimicrobial function NOX4 isresponsible not only for osteoclast but also for preosteoblastdifferentiation via ROS accumulation reinforcing the ideaof NOX4 as a key regulator in bone remodeling underphysiological ROS amounts [47] Animal studies supportthe hypothesis that massive ROS formation induced bone
resorption via NOX4 [20 21] Whereas physiological levelsof NOX4-generated ROS in PDL cellsmight facilitate alveolarbone remodeling massive ROS accumulation during inflam-matory periodontal diseases might lead to alveolar bonedestruction favoring tooth loss We have demonstrated firstthat NOX4 is upregulated in periodontal ligament fibroblastsduring bacterial and hypoxic events Hence PDL fibroblastsmay be a source of excessive ROS generation and are thereforeof particular importance during periodontal diseases
Our results support current findings determining the roleof oxidative stress in different periodontal pathologies whichimplicate that the balance between NADPH-oxidases andredox systems is crucial to maintain cell and tissue home-ostasis [22ndash26] However there are only a few studies usinghuman periodontal fibroblast in the context of oxidativestress and none of them evaluated the role of NOX4 andcounteracting redox systems during inflammatory conditionswith LPS-PG stimulation and hypoxia An increase of ROSgeneration was detected by San Miguel et al following PDLfibroblast stimulation with nicotine or ethanol which wereabolished after antioxidant treatment [40] They completedtheir results using metal ions like nickel for PDL stimulationwhich increases ROS activity in these cells indicating a role
Mediators of Inflammation 9
200120583m
50120583m
50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 5 Catalase (CAT) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontal ligament (b) andsamples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University of Bonn and parentalas well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against catalase (Abcam Cambridge UK) was used in a concentration of1 100 No immunostaining could be found in the healthy gingival epithelium In the healthy PDL predominantly PDL cells located near tothe tooth root surfaces were stained (Figure 5(b) red arrow) In gingivitis tissue a weak staining of gingival keratinocytes similar to healthytissue could be seen (Figure 5(c)) Instead in the tissue samples of patients with periodontitis a strong immunoreactivity was observed inthe subepithelial layer probably referring to nuclei staining of immune cells like leukocytes Red arrows indicate examples for the typicallocalisation (Figure 5(d)) The scale bars indicate 200 120583m in the surveys and 50120583m in the higher magnifications respectively
in metal cytotoxicity or even nickel allergy [41] Transientmetal ions like nickel and cobalt are known to mimic cellularhypoxia mainly due to the inhibition of prolyl hydrolases(PDHs) and asparaginyl hydrolase (factor-inhibiting HIF(FIH)) The inhibition of hydroxylation leads to HIF-1120572nuclear translocation followed by the transcription of diversegenes which are implicated in hypoxic adaptation immuneresponse or angiogenesis therefore indirectly supporting ourresults under hypoxic condition [48 49]
We could show that hypoxia as well as LPS-PG inducedROS formation by detecting H
2O2using the commercially
available Amplex Red assay The high levels of H2O2released
under hypoxic conditions after 1 and 2 hours in PDL cellsmay be caused by different mechanisms First we founda significant upregulation of the mRNA and protein ofthe H
2O2-generating enzyme NOX4 Second the lack of
molecular oxygen as the final electron acceptor in therespiratory chain may be responsible for the early release of
ROS LPS-PG alone induced acutely a small but significantincrease in H
2O2release and enhanced the hypoxia-induced
generation In human gingival fibroblasts similar results wereobtained showing the increase of ROS and antioxidants afterP gingivalis or LPS stimulation [26 42 43] Interestinglydespite the increase in NOX4 mRNA and protein expressionafter 4 hours of stimulation with LPS-PG under hypoxicconditions the H
2O2generation was significantly reduced
when compared to hypoxia alone An additional inhibitionof the respiratory chain by LPS-PG which was described forcardiac mitochondria [50] is therefore conceivable
To antagonize oxidative stress cells developed severalantioxidative enzyme systems Therefore we measured SODand CAT expression in PDL cells and periodontal tissues byimmunofluorescence and immunohistochemistry SODs aremetalloenzymes which are responsible for the dismutation ofsuperoxide radicals to hydrogen peroxide [51] The reductionof hydrogen peroxide to water and oxygen is subsequently
10 Mediators of Inflammation
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m 50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 6 Superoxide dismutase (SOD) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against SOD (Abcam Cambridge UK) was used ina concentration of 1 100 Gingival epithelium showed a moderate to strong staining of SOD in keratinocytes which was primarily limitedto the cytosol (Figure 6(a) and red arrow depicts the typical localization) In the healthy periodontium gingival and PDL fibroblasts wereweakly stained (Figure 6(b)) In inflamed tissues basal epithelial cells seemed to be stained more intensively in the gingivitis specimen thanin periodontitis (Figures 6(c) and 6(d)) In addition the subepithelium revealed immunoreactivity in local cells like gingival fibroblasts andscattered immune cells (Figure 6(c) see red arrow) In periodontitis SOD immunostaining seemed to be limited to the gingival epithelium(Figure 6(d) see red arrow) The scale bars indicate 200 120583m in the surveys and 50 120583m in the higher magnifications respectively
catalyzed by CATs which are ubiquitously expressed Bothenzymes are essential to maintain cellular redox homeostasisThe present data imply that PDL cells and periodontal tissueshave fundamental defense machineries against bacteria rep-resented by NOX4-dependent inducible ROS generation aswell as protective redox systems This is of great importanceconcerning the high amounts and diversity of the oral floraDifferent studies indicate that redox systems are key playersin the development of periodontal diseases and markers forperiodontal health status [52ndash56] We detected a significantincrease of catalase in PDL cells in response to LPS aswell as hypoxia after 1 and 2 hours In our setting PDLfibroblasts seem to counteracted LPS- and hypoxia-inducedROS formation with the elevation of CAT expression Butprolonged incubation under hypoxic conditions failed toincrease catalase Since ROS generation was still elevated atthese later time points a delay in ROS degradation may be
the explanation Taken together the dramatic elevation inROS formation by the combination of hypoxia and LPS-PGon one hand and the reduction of catalase under the sameconditions on the other hand increase the oxidative stress inthe periodontium favoring the progression of inflammationand bone resorption and finally may lead to tooth loss
The fundamental role of the balance between ROSgeneration and antioxidative mechanisms in periodontalinflammatory processes was supported by our immunohis-tochemical findings In patients suffering from periodontalinflammatory diseases like gingivitis and periodontitis wedemonstrated that NOX4 is upregulated and thus likelyresponsible for H
2O2generation However we detected as
well an increase in catalase abundance in inflamed gingivaland periodontal tissue which was in line with data fromKrifka and colleagues who detected high CAT levels inmonomer-exposed macrophages as a response to oxidative
Mediators of Inflammation 11
stress [57] In our in vitro setting PDL fibroblasts seemto counteract LPS- and hypoxia-induced ROS formationpredominantly with an elevation of CAT expression sincewe could not detect any SOD immunoreactivity The roleand impact of the antioxidative systems SOD and CAT arecontroversially discussed Recent studies measured CAT andSOD activities in gingival tissues of patients with periodon-titis [58 59] Borges Jr et al did not find differences inCAT activities comparing healthy and diseased patients [58]Instead Tonguc et al detected decreased local SOD andCAT activities in periodontitis patients which seemed to beincreased by smoking [59] In contrast in inflamed tissues weobservedhigh amounts of SODwhichwas localized primarilyin epithelial cells Jacoby and Davis determined SOD in peri-odontal soft tissues They detected twice as much SOD activ-ity in the periodontal ligament as in the human skin (dermis)[60] Furthermore immunohistochemical analysis revealedthat SOD is localized in the periphery of matrix collagenfibrils and to the glycocalyx of tissue fibroblasts This wasconfirmed by Skaleric et al [42] which might explain the factthat we did not detect significant differences of SOD levelsafter hypoxic and LPS-PG stimulation intracellularly in PDLfibroblasts Another reason for these results may be causedby the age of the PDL cells used in this study We explantedthe PDL cells from young and oral healthy patients but mostof the immunohistochemical stainings were performed intissues from elderly patients with clearly diagnosed gingivitisor periodontitisTherefore it is possible that in PDL cells fromelderly patients a different expression pattern of antioxidantenzyme systems could be found which further deterioratethe redox state of the periodontal ligament Moreover otherauthors found reduced antioxidants in the body fluids ofpatients with periodontitis indicating a dysregulation whichwould favor disease progression [23 24 61] A variety of invitro and animal studies analyzed the effect of antioxidantsupplement in the context of inflammatory or cytotoxicevents [57 62 63] They have shown that antioxidantsinhibit these processes indicating a therapeutic approachin pathologies like periodontitis First clinical investigationsconfirmed the efficiency of antioxidant micronutrients andadjuncts in periodontal diseases [22 54 56 64 65] Thepresent data indirectly support these studies
Concerning periodontal pathogen invasiveness and viru-lence there is compelling evidence that P gingivalis possessesits own protective antioxidants for example rubrerythrindefending them against the oxidative burst of the host [66]We have shown that its virulence factor LPS-PG inducedhigh amounts of ROS via NOX4 which may result in boneresorption favoring the invasiveness of the pathogen whichis at the same time protected by its own defense machineryThis would further explain the pathogenicity and resistanceof this bacterium strain in patients with periodontitis
5 Conclusion
In this study we have demonstrated that LPS from Porphy-romonas gingivalis (LPS-PG) as well as hypoxia induces aNOX4-dependent increase inH
2O2release in PDLfibroblasts
which may contribute to the development and progressionof periodontal diseases when not balanced by a concomitantincrease in antioxidative systems The dramatic elevationin ROS formation induced by the combination of hypoxiaand LPS-PG and the reduction of catalase under the sameconditions increased the oxidative stress in the periodontiumfavoring the progression of inflammation bone resorptionand possibly tooth loss
Taken together the presented data further explain thedestructive mechanism of hypoxic and inflammatory condi-tions in the periodontium and may improve diagnostic andtherapeutic strategies in periodontal diseases
Conflict of Interests
The authors have no conflict of interests to disclose
Acknowledgments
The authors would like to thank Ms Tina Schaffrath for hertechnical supportThe present studywas supported by a grantfrom the German Research Foundation (Clinical ResearchUnit 208TP7) and the Medical Faculty of the University ofBonn
References
[1] S Reuter S C Gupta M M Chaturvedi and B B AggarwalldquoOxidative stress inflammation and cancer how are theylinkedrdquo Free Radical Biology and Medicine vol 49 no 11 pp1603ndash1616 2010
[2] H Sone H Akanuma and T Fukuda ldquoOxygenomics inenvironmental stressrdquo Redox Report vol 15 no 3 pp 98ndash1142010
[3] K E Wellen and C B Thompson ldquoCellular metabolic stressconsidering how cells respond to nutrient excessrdquo MolecularCell vol 40 no 2 pp 323ndash332 2010
[4] R O Poyton K A Ball and P R Castello ldquoMitochondrialgeneration of free radicals and hypoxic signalingrdquo Trends inEndocrinology andMetabolism vol 20 no 7 pp 332ndash340 2009
[5] F Weinberg and N S Chandel ldquoReactive oxygen species-dependent signaling regulates cancerrdquo Cellular and MolecularLife Sciences vol 66 no 23 pp 3663ndash3673 2009
[6] A Cataldi ldquoCell responses to oxidative stressorsrdquo CurrentPharmaceutical Design vol 16 no 12 pp 1387ndash1395 2010
[7] Z Durackova ldquoSome current insights into oxidative stressrdquoPhysiological Research vol 59 no 4 pp 459ndash469 2010
[8] L A Sena and N S Chandel ldquoPhysiological roles of mitochon-drial reactive oxygen speciesrdquoMolecular Cell vol 48 no 2 pp158ndash166 2012
[9] D I Brown and K K Griendling ldquoNox proteins in signaltransductionrdquo Free Radical Biology amp Medicine vol 47 no 9pp 1239ndash1253 2009
[10] K Bedard and K-H Krause ldquoThe NOX family of ROS-generatingNADPHoxidases physiology and pathophysiologyrdquoPhysiological Reviews vol 87 no 1 pp 245ndash313 2007
[11] K-H Krause ldquoTissue distribution and putative physiologicalfunction of NOX family NADPH oxidasesrdquo Japanese Journal ofInfectious Diseases vol 57 no 5 pp S28ndashS29 2004
12 Mediators of Inflammation
[12] S Yang Y Zhang W Ries and L Key ldquoExpression of Nox4 inosteoclastsrdquo Journal of Cellular Biochemistry vol 92 no 2 pp238ndash248 2004
[13] E Pedruzzi C Guichard V Ollivier et al ldquoNAD(P)H oxidaseNox-4mediates 7-ketocholesterol-induced endoplasmic reticu-lum stress and apoptosis in human aortic smooth muscle cellsrdquoMolecular and Cellular Biology vol 24 no 24 pp 10703ndash107172004
[14] J Hwang A Saha Y C Boo et al ldquoOscillatory shear stress stim-ulates endothelial production of O
2-from p47119901ℎ119900119909-dependent
NAD(P)Hoxidases leading tomonocyte adhesionrdquoThe Journalof Biological Chemistry vol 278 no 47 pp 47291ndash47298 2003
[15] H B Suliman M Ali and C A Piantadosi ldquoSuperoxidedismutase-3 promotes full expression of the EPO response tohypoxiardquo Blood vol 104 no 1 pp 43ndash50 2004
[16] P Vallet Y Charnay K Steger et al ldquoNeuronal expressionof the NADPH oxidase NOX4 and its regulation in mouseexperimental brain ischemiardquo Neuroscience vol 132 no 2 pp233ndash238 2005
[17] K T Moe S Aulia F Jiang et al ldquoDifferential upregulation ofNox homologues of NADPH oxidase by tumor necrosis factor-120572 in human aortic smooth muscle and embryonic kidney cellsrdquoJournal of Cellular andMolecularMedicine vol 10 no 1 pp 231ndash239 2006
[18] I Diebold A Petry J Hess and A Gorlach ldquoThe NADPHoxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1rdquoMolecular Biology of the Cell vol 21 no 12pp 2087ndash2096 2010
[19] K V Tormos and N S Chandel ldquoInter-connection betweenmitochondria and HIFsrdquo Journal of Cellular and MolecularMedicine vol 14 no 4 pp 795ndash804 2010
[20] T Ohnishi K Bandow K Kakimoto M Machigashira TMatsuyama and T Matsuguchi ldquoOxidative stress causes alve-olar bone loss in metabolic syndrome model mice with type 2diabetesrdquo Journal of Periodontal Research vol 44 no 1 pp 43ndash51 2009
[21] C Goettsch A Babelova O Trummer et al ldquoNADPH oxidase4 limits bone mass by promoting osteoclastogenesisrdquo Journal ofClinical Investigation vol 123 no 11 pp 4731ndash4738 2013
[22] I L C Chapple and J B Matthews ldquoThe role of reactive oxygenand antioxidant species in periodontal tissue destructionrdquoPeriodontology 2000 vol 43 no 1 pp 160ndash232 2007
[23] A Guentsch P M Preshaw S Bremer-Streck G KlingerE Glockmann and B W Sigusch ldquoLipid peroxidation andantioxidant activity in saliva of periodontitis patients effect ofsmoking and periodontal treatmentrdquo Clinical Oral Investiga-tions vol 12 no 4 pp 345ndash352 2008
[24] R Agnihotri P Pandurang S U Kamath et al ldquoAssociation ofcigarette smoking with superoxide dismutase enzyme levels insubjected with chonic periodontitisrdquo Journal of Periodontologyvol 80 no 4 pp 657ndash662 2009
[25] D Ekuni T Tomofuji N Tamaki et al ldquoMechanical stimulationof gingiva reduces plasma 8-OHdG level in rat periodontitisrdquoArchives of Oral Biology vol 53 no 4 pp 324ndash329 2008
[26] H Staudte A Guntsch A Volpel and B W Sigusch ldquoVitaminC attenuates the cytotoxic effects of Porphyromonas gingivalison human gingival fibroblastsrdquo Archives of Oral Biology vol 55no 1 pp 40ndash45 2010
[27] P Bullon H N Newman and M Battino ldquoObesity diabetesmellitus atherosclerosis and chronic periodontitis a sharedpathology via oxidative stress andmitochondrial dysfunctionrdquoPeriodontology 2000 vol 64 no 1 pp 139ndash153 2014
[28] G R Naveh V Brumfeld R Shahar and S Weiner ldquoToothperiodontal ligament direct 3D microCT visualization of thecollagen network and how the network changes when the toothis loadedrdquo Journal of Structural Biology vol 181 no 2 pp 108ndash115 2013
[29] Y Kitase M Yokozeki S Fujihara et al ldquoAnalysis of geneexpression profiles in human periodontal ligament cells underhypoxia the protective effect of CC chemokine ligand 2 tooxygen shortagerdquoArchives of Oral Biology vol 54 no 7 pp 618ndash624 2009
[30] G Thornton-Evans P Eke L Wei et al ldquoPeriodontitis amongadults aged ge 30 years - United States 2009-2010rdquo Morbidityand Mortality Weekly Report (MMWR) vol 62 supplement 3pp 129ndash135 2013
[31] S S Socransky C Smith and A D Haffajee ldquoSubgingivalmicrobial profiles in refractory periodontal diseaserdquo Journal ofClinical Periodontology vol 29 no 3 pp 260ndash268 2002
[32] K-T Ng J-P Li K M Ng G L Tipoe W K Leung and M-L Fung ldquoExpression of hypoxia-inducible factor-1120572 in humanperiodontal tissuerdquo Journal of Periodontology vol 82 no 1 pp136ndash141 2011
[33] S Frede U Berchner-Pfannschmidt and J Fandrey ldquoReg-ulation of Hypoxia-inducible factors during inflammationrdquoMethods in Enzymology vol 435 pp 405ndash419 2007
[34] J Shin S Ji and Y Choi ldquoAbility of oral bacteria to inducetissue-destructive molecules from human neutrophilsrdquo OralDiseases vol 14 no 4 pp 327ndash334 2008
[35] S Takashiba K Naruishi and Y Murayama ldquoPerspectiveof cytokine regulation for periodontal treatment fibroblastbiologyrdquo Journal of Periodontology vol 74 no 1 pp 103ndash1102003
[36] A Konermann D Stabenow P A Knolle S A E HeldJ Deschner and A Jager ldquoRegulatory role of periodontalligament fibroblasts for innate immune cell function and dif-ferentiationrdquo Innate Immunity vol 18 no 5 pp 745ndash752 2012
[37] S Kar S Subbaram P M Carrico and J A Melendez ldquoRedox-control of matrix metalloproteinase-1 a critical link betweenfree radicals matrix remodeling and degenerative diseaserdquoRespiratory Physiology andNeurobiology vol 174 no 3 pp 299ndash306 2010
[38] F Rousset M V C Nguyen L Grange F Morel and B LardyldquoHeme oxygenase-1 regulates matrix metalloproteinase MMP-1secretion and chondrocyte cell death via Nox4NADPH oxidaseactivity in chondrocytesrdquo PLoS ONE vol 8 no 6 Article IDe66478 2013
[39] E Rudolf and M Cervinka ldquoResponses of human gingival andperiodontal fibroblasts to a low-zinc environmentrdquo Alternativesto Laboratory Animals vol 38 no 2 pp 119ndash138 2010
[40] S M San Miguel L A Opperman E P Allen J Zielinski andK K H Svoboda ldquoBioactive polyphenol antioxidants protectoral fibroblasts from ROS-inducing agentsrdquo Archives of OralBiology vol 57 no 12 pp 1657ndash1667 2012
[41] S M San Miguel L A Opperman E P Allen J E Zielinskiand K K H Svoboda ldquoAntioxidant combinations protect oralfibroblasts against metal-induced toxicityrdquo Archives of OralBiology vol 58 no 3 pp 299ndash310 2013
[42] U Skaleric C M Manthey S E Mergenhagen B Gaspircand S M Wahl ldquoSuperoxide release and superoxide dismutaseexpression by human gingival fibroblastsrdquo European Journal ofOral Sciences vol 108 no 2 pp 130ndash135 2000
Mediators of Inflammation 13
[43] D Y Kim J-H Jun H-L Lee et al ldquoN-acetylcysteine preventsLPS-Lnduced pro-inflammatory cytokines andMMP2 produc-tion in gingival fibroblastsrdquoArchives of Pharmacal Research vol30 no 10 pp 1283ndash1292 2007
[44] B Rath-Deschner J Deschner S Reimann A Jager andW Gotz ldquoRegulatory effects of biomechanical strain on theinsulin-like growth factor system in human periodontal cellsrdquoJournal of Biomechanics vol 42 no 15 pp 2584ndash2589 2009
[45] P Chomczynski and N Sacchi ldquoSingle-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextractionrdquo Analytical Biochemistry vol 162 no 1 pp 156ndash1591987
[46] B Zhao K Ranguelova J Jiang and R P Mason ldquoStudies onthe photosensitized reduction of resorufin and implications forthe detection of oxidative stress with Amplex Redrdquo Free RadicalBiology and Medicine vol 51 no 1 pp 153ndash159 2011
[47] C C Mandal S Ganapathy Y Gorin et al ldquoReactive oxygenspecies derived from Nox4 mediate BMP2 gene transcriptionand osteoblast differentiationrdquoBiochemical Journal vol 433 no2 pp 393ndash402 2011
[48] J Fandrey S Frede and W Jelkmann ldquoRole of hydrogenperoxide in hypoxia-induced erythropoietin productionrdquo Bio-chemical Journal vol 303 no 2 pp 507ndash510 1994
[49] F Cervellati C Cervellati A Romani et al ldquoHypoxia inducescell damage via oxidative stress in retinal epithelial cellsrdquo FreeRadical Research vol 48 no 3 pp 303ndash312 2014
[50] V Vanasco N D Magnani M C Cimolai et al ldquoEndotoxemiaimpairs heart mitochondrial function by decreasing electrontransfer ATP synthesis and ATP content without affectingmembrane potentialrdquo Journal of Bioenergetics and Biomem-branes vol 44 no 2 pp 243ndash252 2012
[51] R G Alscher N Erturk and L S Heath ldquoRole of superoxidedismutases (SODs) in controlling oxidative stress in plantsrdquoJournal of Experimental Botany vol 53 no 372 pp 1331ndash13412002
[52] E Baltacıoglu P Yuva G Aydın et al ldquoLipid peroxidation levelsand total oxidantantioxidant status in serum and saliva frompatients with chronic and aggressive periodontitis Oxidativestress index a new biomarker for periodontal diseaserdquo Journalof Periodontology vol 85 no 10 pp 1432ndash1441 2014
[53] I L C Chapple M R Milward N Ling-Mountford et alldquoAdjunctive daily supplementation with encapsulated fruitvegetable and berry juice powder concentrates and clinicalperiodontal outcomes a double-blind RCTrdquo Journal of ClinicalPeriodontology vol 39 no 1 pp 62ndash72 2012
[54] A Jenzsch S Eick F Rassoul R Purschwitz and H JentschldquoNutritional intervention in patients with periodontal diseaseClinical immunological and microbiological variables during12 monthsrdquo British Journal of Nutrition vol 101 no 6 pp 879ndash885 2009
[55] N Novakovic T Todorovic M Rakic et al ldquoSalivary antiox-idants as periodontal biomarkers in evaluation of tissue statusand treatment outcomerdquo Journal of Periodontal Research vol49 no 1 pp 129ndash136 2014
[56] N Singh S C Narula R K Sharma S Tewari and P KSehgal ldquoVitamin e supplementation superoxide dismutasestatus and outcome of scaling and root planing in patients withchronic periodontitis a randomized clinical trialrdquo Journal ofPeriodontology vol 85 no 2 pp 242ndash249 2014
[57] S Krifka K-A Hiller G Spagnuolo A Jewett G Schmalz andH Schweikl ldquoThe influence of glutathione on redox regulation
by antioxidant proteins and apoptosis in macrophages exposedto 2-hydroxyethyl methacrylate (HEMA)rdquo Biomaterials vol 33no 21 pp 5177ndash5186 2012
[58] I Borges Jr E A M Moreira D W Filho T B de Oliveira MB S da Silva and T S Frode ldquoProinflammatory and oxidativestress markers in patients with periodontal diseaserdquo Mediatorsof Inflammation vol 2007 Article ID 45794 5 pages 2007
[59] M O Tonguc O Ozturk R Sutcu et al ldquoThe impact of smok-ing status on antioxidant enzyme activity and malondialdehydelevels in chronic periodontitisrdquo Journal of Periodontology vol82 no 9 pp 1320ndash1328 2011
[60] B H Jacoby and W L Davis ldquoThe electron microscopicimmunolocalization of a copper-zinc superoxide dismutase inassociation with collagen fibers of periodontal soft tissuesrdquoJournal of Periodontology vol 62 no 7 pp 413ndash420 1991
[61] T Konopka K Krol W Kopec and H Gerber ldquoTotal antioxi-dant status and 8-hydroxy-21015840-deoxyguanosine levels in gingi-val and peripheral blood of periodontitis patientsrdquo ArchivumImmunologiae et Therapiae Experimentalis vol 55 no 6 pp417ndash422 2007
[62] M Soory ldquoRedox status in periodontal and systemic inflamma-tory conditions including associated neoplasias antioxidants asadjunctive therapyrdquo Infectious DisordersmdashDrug Targets vol 9no 4 pp 415ndash427 2009
[63] M Soory ldquoOxidative stress inducedmechanisms in the progres-sion of periodontal diseases and cancer a common approach toredox homeostasisrdquo Cancers vol 2 no 2 pp 670ndash692 2010
[64] R V Chandra G Srinivas A A Reddy et al ldquoLocally deliveredantioxidant gel as an adjunct to nonsurgical therapy improvesmeasures of oxidative stress and periodontal diseaserdquo Journal ofPeriodontal and Implant Science vol 43 no 3 pp 121ndash129 2013
[65] M A D S Siqueira R G Fischer N R Pereira et al ldquoEffectsof non-surgical periodontal treatment on the L-arginine-nitricoxide pathway and oxidative status in plateletsrdquo ExperimentalBiology and Medicine vol 238 no 6 pp 713ndash722 2013
[66] P Mydel Y Takahashi H Yumoto et al ldquoRoles of the hostoxidative immune response and bacterial antioxidant rubr-erythrin during Porphyromonas gingivalis infectionrdquo PLoSpathogens vol 2 no 7 p e76 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
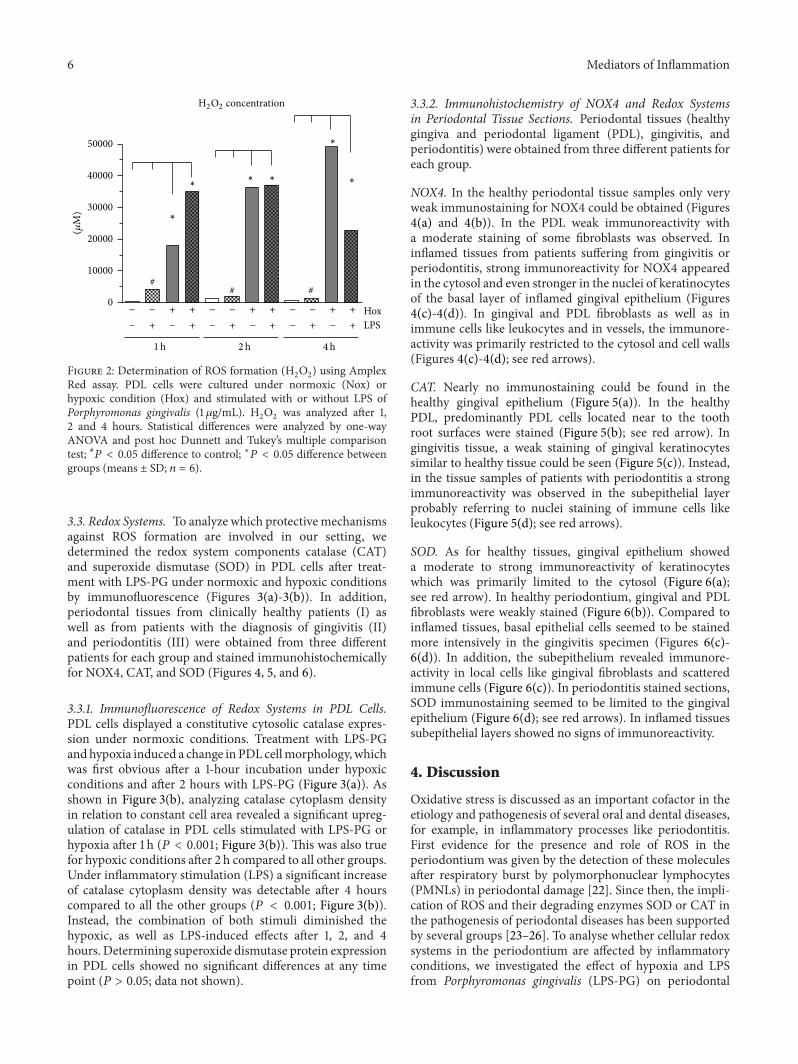
6 Mediators of Inflammation
2h 4h1h
50000
40000
30000
20000
10000
0
(120583M
)
HoxLPS
minus minus minus minus minus minus
minus
+
+minus +minus +minus +minus +minus +
++ + + +
lowast
lowastlowast lowast
lowast
lowast
H2O2 concentration
Figure 2 Determination of ROS formation (H2O2) using Amplex
Red assay PDL cells were cultured under normoxic (Nox) orhypoxic condition (Hox) and stimulated with or without LPS ofPorphyromonas gingivalis (1 120583gmL) H
2O2was analyzed after 1
2 and 4 hours Statistical differences were analyzed by one-wayANOVA and post hoc Dunnett and Tukeyrsquos multiple comparisontest 119875 lt 005 difference to control lowast119875 lt 005 difference betweengroups (means plusmn SD 119899 = 6)
33 Redox Systems To analyze which protectivemechanismsagainst ROS formation are involved in our setting wedetermined the redox system components catalase (CAT)and superoxide dismutase (SOD) in PDL cells after treat-ment with LPS-PG under normoxic and hypoxic conditionsby immunofluorescence (Figures 3(a)-3(b)) In additionperiodontal tissues from clinically healthy patients (I) aswell as from patients with the diagnosis of gingivitis (II)and periodontitis (III) were obtained from three differentpatients for each group and stained immunohistochemicallyfor NOX4 CAT and SOD (Figures 4 5 and 6)
331 Immunofluorescence of Redox Systems in PDL CellsPDL cells displayed a constitutive cytosolic catalase expres-sion under normoxic conditions Treatment with LPS-PGandhypoxia induced a change in PDL cellmorphologywhichwas first obvious after a 1-hour incubation under hypoxicconditions and after 2 hours with LPS-PG (Figure 3(a)) Asshown in Figure 3(b) analyzing catalase cytoplasm densityin relation to constant cell area revealed a significant upreg-ulation of catalase in PDL cells stimulated with LPS-PG orhypoxia after 1 h (119875 lt 0001 Figure 3(b)) This was also truefor hypoxic conditions after 2 h compared to all other groupsUnder inflammatory stimulation (LPS) a significant increaseof catalase cytoplasm density was detectable after 4 hourscompared to all the other groups (119875 lt 0001 Figure 3(b))Instead the combination of both stimuli diminished thehypoxic as well as LPS-induced effects after 1 2 and 4hours Determining superoxide dismutase protein expressionin PDL cells showed no significant differences at any timepoint (119875 gt 005 data not shown)
332 Immunohistochemistry of NOX4 and Redox Systemsin Periodontal Tissue Sections Periodontal tissues (healthygingiva and periodontal ligament (PDL) gingivitis andperiodontitis) were obtained from three different patients foreach group
NOX4 In the healthy periodontal tissue samples only veryweak immunostaining for NOX4 could be obtained (Figures4(a) and 4(b)) In the PDL weak immunoreactivity witha moderate staining of some fibroblasts was observed Ininflamed tissues from patients suffering from gingivitis orperiodontitis strong immunoreactivity for NOX4 appearedin the cytosol and even stronger in the nuclei of keratinocytesof the basal layer of inflamed gingival epithelium (Figures4(c)-4(d)) In gingival and PDL fibroblasts as well as inimmune cells like leukocytes and in vessels the immunore-activity was primarily restricted to the cytosol and cell walls(Figures 4(c)-4(d) see red arrows)
CAT Nearly no immunostaining could be found in thehealthy gingival epithelium (Figure 5(a)) In the healthyPDL predominantly PDL cells located near to the toothroot surfaces were stained (Figure 5(b) see red arrow) Ingingivitis tissue a weak staining of gingival keratinocytessimilar to healthy tissue could be seen (Figure 5(c)) Insteadin the tissue samples of patients with periodontitis a strongimmunoreactivity was observed in the subepithelial layerprobably referring to nuclei staining of immune cells likeleukocytes (Figure 5(d) see red arrows)
SOD As for healthy tissues gingival epithelium showeda moderate to strong immunoreactivity of keratinocyteswhich was primarily limited to the cytosol (Figure 6(a)see red arrow) In healthy periodontium gingival and PDLfibroblasts were weakly stained (Figure 6(b)) Compared toinflamed tissues basal epithelial cells seemed to be stainedmore intensively in the gingivitis specimen (Figures 6(c)-6(d)) In addition the subepithelium revealed immunore-activity in local cells like gingival fibroblasts and scatteredimmune cells (Figure 6(c)) In periodontitis stained sectionsSOD immunostaining seemed to be limited to the gingivalepithelium (Figure 6(d) see red arrows) In inflamed tissuessubepithelial layers showed no signs of immunoreactivity
4 Discussion
Oxidative stress is discussed as an important cofactor in theetiology and pathogenesis of several oral and dental diseasesfor example in inflammatory processes like periodontitisFirst evidence for the presence and role of ROS in theperiodontium was given by the detection of these moleculesafter respiratory burst by polymorphonuclear lymphocytes(PMNLs) in periodontal damage [22] Since then the impli-cation of ROS and their degrading enzymes SOD or CAT inthe pathogenesis of periodontal diseases has been supportedby several groups [23ndash26] To analyse whether cellular redoxsystems in the periodontium are affected by inflammatoryconditions we investigated the effect of hypoxia and LPSfrom Porphyromonas gingivalis (LPS-PG) on periodontal
Mediators of Inflammation 7
1h 2hCatalase
Nox
Nox + LPS
Hox
Hox + LPS
4h
100120583m 100120583m 100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
(a)
2h 4h
Hox
Inte
grat
ed d
ensit
y (a
u)
Catalase cytoplasm density
LPSminusminus + + minusminus + + minusminus +
minus +minus +minus +minus +minus +minus +
+
lowast
lowast
lowast
lowast
lowastlowast
1h
20000
25000
15000
1000010000
7500
5000
2500
0
(b)
Figure 3 Catalase (CAT) in PDL cells (a) CAT was visualized by immunofluorescence staining (1 50 antibody from Abcam) PDL cellswere cultured under normoxic (Nox) or hypoxic condition (Hox) and stimulated with or without LPS of Porphyromonas gingivalis (1120583gmL)Catalase cytoplasm density (b) was determined in relation to cell area using the freely available image-processing software ImageJ 143(httprsbinfonihgovij) Statistical analysis of immunofluorescence data was analyzed by one-way ANOVA and post hoc Dunnett andTukeyrsquos multiple comparison test 119875 lt 005 difference to control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 6) The scale barsindicate 100 120583m in the immunofluorescence images
8 Mediators of Inflammation
50120583m
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m
50120583m
50120583m
Inflamed
(d)
Figure 4 NADPH-oxidase 4 (NOX4) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Monoclonal primary antibody against NOX4 (Abcam Cambridge UK) was used ina concentration of 1 100 In gingival and PDL fibroblasts as well as in immune cells like leukocytes and in vessels the immunoreactivity wasprimarily restricted to the cytosol and cell walls Red arrows indicate examples for the typical distribution (Figures 4(c) and 4(d)) The scalebars indicate 200120583m in the surveys and 50 120583m in the higher magnifications respectively
ligament cells We showed that both stimuli induced asignificant increase of NOX4 and a significant upregulationof hydrogen peroxide generation NOX enzymes were firstdescribed in phagocytes as being responsible for defensemechanisms against pathogens using NOX-generated ROSmolecules Till then NOX4 expression has been detected innonphagocyte cell types like kidney cells osteoclasts andosteoblasts as well as dermal cells [9 11 47] In PDL cellshypoxia and LPS-PG induced a significant increase of NOX4mRNA The protein expression was mainly located in thecytosol but likewise in the nucleus Our findings were inline with Brown and Griendling who have shown that thetypically transmembrane molecule is localized in differentcell compartments [9]
NOX4 is one of the major sources of ROS generationwhich is highly and ubiquitously expressed compared tothe other family members Mandal and colleagues demon-strated that apart from its antimicrobial function NOX4 isresponsible not only for osteoclast but also for preosteoblastdifferentiation via ROS accumulation reinforcing the ideaof NOX4 as a key regulator in bone remodeling underphysiological ROS amounts [47] Animal studies supportthe hypothesis that massive ROS formation induced bone
resorption via NOX4 [20 21] Whereas physiological levelsof NOX4-generated ROS in PDL cellsmight facilitate alveolarbone remodeling massive ROS accumulation during inflam-matory periodontal diseases might lead to alveolar bonedestruction favoring tooth loss We have demonstrated firstthat NOX4 is upregulated in periodontal ligament fibroblastsduring bacterial and hypoxic events Hence PDL fibroblastsmay be a source of excessive ROS generation and are thereforeof particular importance during periodontal diseases
Our results support current findings determining the roleof oxidative stress in different periodontal pathologies whichimplicate that the balance between NADPH-oxidases andredox systems is crucial to maintain cell and tissue home-ostasis [22ndash26] However there are only a few studies usinghuman periodontal fibroblast in the context of oxidativestress and none of them evaluated the role of NOX4 andcounteracting redox systems during inflammatory conditionswith LPS-PG stimulation and hypoxia An increase of ROSgeneration was detected by San Miguel et al following PDLfibroblast stimulation with nicotine or ethanol which wereabolished after antioxidant treatment [40] They completedtheir results using metal ions like nickel for PDL stimulationwhich increases ROS activity in these cells indicating a role
Mediators of Inflammation 9
200120583m
50120583m
50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 5 Catalase (CAT) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontal ligament (b) andsamples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University of Bonn and parentalas well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against catalase (Abcam Cambridge UK) was used in a concentration of1 100 No immunostaining could be found in the healthy gingival epithelium In the healthy PDL predominantly PDL cells located near tothe tooth root surfaces were stained (Figure 5(b) red arrow) In gingivitis tissue a weak staining of gingival keratinocytes similar to healthytissue could be seen (Figure 5(c)) Instead in the tissue samples of patients with periodontitis a strong immunoreactivity was observed inthe subepithelial layer probably referring to nuclei staining of immune cells like leukocytes Red arrows indicate examples for the typicallocalisation (Figure 5(d)) The scale bars indicate 200 120583m in the surveys and 50120583m in the higher magnifications respectively
in metal cytotoxicity or even nickel allergy [41] Transientmetal ions like nickel and cobalt are known to mimic cellularhypoxia mainly due to the inhibition of prolyl hydrolases(PDHs) and asparaginyl hydrolase (factor-inhibiting HIF(FIH)) The inhibition of hydroxylation leads to HIF-1120572nuclear translocation followed by the transcription of diversegenes which are implicated in hypoxic adaptation immuneresponse or angiogenesis therefore indirectly supporting ourresults under hypoxic condition [48 49]
We could show that hypoxia as well as LPS-PG inducedROS formation by detecting H
2O2using the commercially
available Amplex Red assay The high levels of H2O2released
under hypoxic conditions after 1 and 2 hours in PDL cellsmay be caused by different mechanisms First we founda significant upregulation of the mRNA and protein ofthe H
2O2-generating enzyme NOX4 Second the lack of
molecular oxygen as the final electron acceptor in therespiratory chain may be responsible for the early release of
ROS LPS-PG alone induced acutely a small but significantincrease in H
2O2release and enhanced the hypoxia-induced
generation In human gingival fibroblasts similar results wereobtained showing the increase of ROS and antioxidants afterP gingivalis or LPS stimulation [26 42 43] Interestinglydespite the increase in NOX4 mRNA and protein expressionafter 4 hours of stimulation with LPS-PG under hypoxicconditions the H
2O2generation was significantly reduced
when compared to hypoxia alone An additional inhibitionof the respiratory chain by LPS-PG which was described forcardiac mitochondria [50] is therefore conceivable
To antagonize oxidative stress cells developed severalantioxidative enzyme systems Therefore we measured SODand CAT expression in PDL cells and periodontal tissues byimmunofluorescence and immunohistochemistry SODs aremetalloenzymes which are responsible for the dismutation ofsuperoxide radicals to hydrogen peroxide [51] The reductionof hydrogen peroxide to water and oxygen is subsequently
10 Mediators of Inflammation
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m 50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 6 Superoxide dismutase (SOD) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against SOD (Abcam Cambridge UK) was used ina concentration of 1 100 Gingival epithelium showed a moderate to strong staining of SOD in keratinocytes which was primarily limitedto the cytosol (Figure 6(a) and red arrow depicts the typical localization) In the healthy periodontium gingival and PDL fibroblasts wereweakly stained (Figure 6(b)) In inflamed tissues basal epithelial cells seemed to be stained more intensively in the gingivitis specimen thanin periodontitis (Figures 6(c) and 6(d)) In addition the subepithelium revealed immunoreactivity in local cells like gingival fibroblasts andscattered immune cells (Figure 6(c) see red arrow) In periodontitis SOD immunostaining seemed to be limited to the gingival epithelium(Figure 6(d) see red arrow) The scale bars indicate 200 120583m in the surveys and 50 120583m in the higher magnifications respectively
catalyzed by CATs which are ubiquitously expressed Bothenzymes are essential to maintain cellular redox homeostasisThe present data imply that PDL cells and periodontal tissueshave fundamental defense machineries against bacteria rep-resented by NOX4-dependent inducible ROS generation aswell as protective redox systems This is of great importanceconcerning the high amounts and diversity of the oral floraDifferent studies indicate that redox systems are key playersin the development of periodontal diseases and markers forperiodontal health status [52ndash56] We detected a significantincrease of catalase in PDL cells in response to LPS aswell as hypoxia after 1 and 2 hours In our setting PDLfibroblasts seem to counteracted LPS- and hypoxia-inducedROS formation with the elevation of CAT expression Butprolonged incubation under hypoxic conditions failed toincrease catalase Since ROS generation was still elevated atthese later time points a delay in ROS degradation may be
the explanation Taken together the dramatic elevation inROS formation by the combination of hypoxia and LPS-PGon one hand and the reduction of catalase under the sameconditions on the other hand increase the oxidative stress inthe periodontium favoring the progression of inflammationand bone resorption and finally may lead to tooth loss
The fundamental role of the balance between ROSgeneration and antioxidative mechanisms in periodontalinflammatory processes was supported by our immunohis-tochemical findings In patients suffering from periodontalinflammatory diseases like gingivitis and periodontitis wedemonstrated that NOX4 is upregulated and thus likelyresponsible for H
2O2generation However we detected as
well an increase in catalase abundance in inflamed gingivaland periodontal tissue which was in line with data fromKrifka and colleagues who detected high CAT levels inmonomer-exposed macrophages as a response to oxidative
Mediators of Inflammation 11
stress [57] In our in vitro setting PDL fibroblasts seemto counteract LPS- and hypoxia-induced ROS formationpredominantly with an elevation of CAT expression sincewe could not detect any SOD immunoreactivity The roleand impact of the antioxidative systems SOD and CAT arecontroversially discussed Recent studies measured CAT andSOD activities in gingival tissues of patients with periodon-titis [58 59] Borges Jr et al did not find differences inCAT activities comparing healthy and diseased patients [58]Instead Tonguc et al detected decreased local SOD andCAT activities in periodontitis patients which seemed to beincreased by smoking [59] In contrast in inflamed tissues weobservedhigh amounts of SODwhichwas localized primarilyin epithelial cells Jacoby and Davis determined SOD in peri-odontal soft tissues They detected twice as much SOD activ-ity in the periodontal ligament as in the human skin (dermis)[60] Furthermore immunohistochemical analysis revealedthat SOD is localized in the periphery of matrix collagenfibrils and to the glycocalyx of tissue fibroblasts This wasconfirmed by Skaleric et al [42] which might explain the factthat we did not detect significant differences of SOD levelsafter hypoxic and LPS-PG stimulation intracellularly in PDLfibroblasts Another reason for these results may be causedby the age of the PDL cells used in this study We explantedthe PDL cells from young and oral healthy patients but mostof the immunohistochemical stainings were performed intissues from elderly patients with clearly diagnosed gingivitisor periodontitisTherefore it is possible that in PDL cells fromelderly patients a different expression pattern of antioxidantenzyme systems could be found which further deterioratethe redox state of the periodontal ligament Moreover otherauthors found reduced antioxidants in the body fluids ofpatients with periodontitis indicating a dysregulation whichwould favor disease progression [23 24 61] A variety of invitro and animal studies analyzed the effect of antioxidantsupplement in the context of inflammatory or cytotoxicevents [57 62 63] They have shown that antioxidantsinhibit these processes indicating a therapeutic approachin pathologies like periodontitis First clinical investigationsconfirmed the efficiency of antioxidant micronutrients andadjuncts in periodontal diseases [22 54 56 64 65] Thepresent data indirectly support these studies
Concerning periodontal pathogen invasiveness and viru-lence there is compelling evidence that P gingivalis possessesits own protective antioxidants for example rubrerythrindefending them against the oxidative burst of the host [66]We have shown that its virulence factor LPS-PG inducedhigh amounts of ROS via NOX4 which may result in boneresorption favoring the invasiveness of the pathogen whichis at the same time protected by its own defense machineryThis would further explain the pathogenicity and resistanceof this bacterium strain in patients with periodontitis
5 Conclusion
In this study we have demonstrated that LPS from Porphy-romonas gingivalis (LPS-PG) as well as hypoxia induces aNOX4-dependent increase inH
2O2release in PDLfibroblasts
which may contribute to the development and progressionof periodontal diseases when not balanced by a concomitantincrease in antioxidative systems The dramatic elevationin ROS formation induced by the combination of hypoxiaand LPS-PG and the reduction of catalase under the sameconditions increased the oxidative stress in the periodontiumfavoring the progression of inflammation bone resorptionand possibly tooth loss
Taken together the presented data further explain thedestructive mechanism of hypoxic and inflammatory condi-tions in the periodontium and may improve diagnostic andtherapeutic strategies in periodontal diseases
Conflict of Interests
The authors have no conflict of interests to disclose
Acknowledgments
The authors would like to thank Ms Tina Schaffrath for hertechnical supportThe present studywas supported by a grantfrom the German Research Foundation (Clinical ResearchUnit 208TP7) and the Medical Faculty of the University ofBonn
References
[1] S Reuter S C Gupta M M Chaturvedi and B B AggarwalldquoOxidative stress inflammation and cancer how are theylinkedrdquo Free Radical Biology and Medicine vol 49 no 11 pp1603ndash1616 2010
[2] H Sone H Akanuma and T Fukuda ldquoOxygenomics inenvironmental stressrdquo Redox Report vol 15 no 3 pp 98ndash1142010
[3] K E Wellen and C B Thompson ldquoCellular metabolic stressconsidering how cells respond to nutrient excessrdquo MolecularCell vol 40 no 2 pp 323ndash332 2010
[4] R O Poyton K A Ball and P R Castello ldquoMitochondrialgeneration of free radicals and hypoxic signalingrdquo Trends inEndocrinology andMetabolism vol 20 no 7 pp 332ndash340 2009
[5] F Weinberg and N S Chandel ldquoReactive oxygen species-dependent signaling regulates cancerrdquo Cellular and MolecularLife Sciences vol 66 no 23 pp 3663ndash3673 2009
[6] A Cataldi ldquoCell responses to oxidative stressorsrdquo CurrentPharmaceutical Design vol 16 no 12 pp 1387ndash1395 2010
[7] Z Durackova ldquoSome current insights into oxidative stressrdquoPhysiological Research vol 59 no 4 pp 459ndash469 2010
[8] L A Sena and N S Chandel ldquoPhysiological roles of mitochon-drial reactive oxygen speciesrdquoMolecular Cell vol 48 no 2 pp158ndash166 2012
[9] D I Brown and K K Griendling ldquoNox proteins in signaltransductionrdquo Free Radical Biology amp Medicine vol 47 no 9pp 1239ndash1253 2009
[10] K Bedard and K-H Krause ldquoThe NOX family of ROS-generatingNADPHoxidases physiology and pathophysiologyrdquoPhysiological Reviews vol 87 no 1 pp 245ndash313 2007
[11] K-H Krause ldquoTissue distribution and putative physiologicalfunction of NOX family NADPH oxidasesrdquo Japanese Journal ofInfectious Diseases vol 57 no 5 pp S28ndashS29 2004
12 Mediators of Inflammation
[12] S Yang Y Zhang W Ries and L Key ldquoExpression of Nox4 inosteoclastsrdquo Journal of Cellular Biochemistry vol 92 no 2 pp238ndash248 2004
[13] E Pedruzzi C Guichard V Ollivier et al ldquoNAD(P)H oxidaseNox-4mediates 7-ketocholesterol-induced endoplasmic reticu-lum stress and apoptosis in human aortic smooth muscle cellsrdquoMolecular and Cellular Biology vol 24 no 24 pp 10703ndash107172004
[14] J Hwang A Saha Y C Boo et al ldquoOscillatory shear stress stim-ulates endothelial production of O
2-from p47119901ℎ119900119909-dependent
NAD(P)Hoxidases leading tomonocyte adhesionrdquoThe Journalof Biological Chemistry vol 278 no 47 pp 47291ndash47298 2003
[15] H B Suliman M Ali and C A Piantadosi ldquoSuperoxidedismutase-3 promotes full expression of the EPO response tohypoxiardquo Blood vol 104 no 1 pp 43ndash50 2004
[16] P Vallet Y Charnay K Steger et al ldquoNeuronal expressionof the NADPH oxidase NOX4 and its regulation in mouseexperimental brain ischemiardquo Neuroscience vol 132 no 2 pp233ndash238 2005
[17] K T Moe S Aulia F Jiang et al ldquoDifferential upregulation ofNox homologues of NADPH oxidase by tumor necrosis factor-120572 in human aortic smooth muscle and embryonic kidney cellsrdquoJournal of Cellular andMolecularMedicine vol 10 no 1 pp 231ndash239 2006
[18] I Diebold A Petry J Hess and A Gorlach ldquoThe NADPHoxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1rdquoMolecular Biology of the Cell vol 21 no 12pp 2087ndash2096 2010
[19] K V Tormos and N S Chandel ldquoInter-connection betweenmitochondria and HIFsrdquo Journal of Cellular and MolecularMedicine vol 14 no 4 pp 795ndash804 2010
[20] T Ohnishi K Bandow K Kakimoto M Machigashira TMatsuyama and T Matsuguchi ldquoOxidative stress causes alve-olar bone loss in metabolic syndrome model mice with type 2diabetesrdquo Journal of Periodontal Research vol 44 no 1 pp 43ndash51 2009
[21] C Goettsch A Babelova O Trummer et al ldquoNADPH oxidase4 limits bone mass by promoting osteoclastogenesisrdquo Journal ofClinical Investigation vol 123 no 11 pp 4731ndash4738 2013
[22] I L C Chapple and J B Matthews ldquoThe role of reactive oxygenand antioxidant species in periodontal tissue destructionrdquoPeriodontology 2000 vol 43 no 1 pp 160ndash232 2007
[23] A Guentsch P M Preshaw S Bremer-Streck G KlingerE Glockmann and B W Sigusch ldquoLipid peroxidation andantioxidant activity in saliva of periodontitis patients effect ofsmoking and periodontal treatmentrdquo Clinical Oral Investiga-tions vol 12 no 4 pp 345ndash352 2008
[24] R Agnihotri P Pandurang S U Kamath et al ldquoAssociation ofcigarette smoking with superoxide dismutase enzyme levels insubjected with chonic periodontitisrdquo Journal of Periodontologyvol 80 no 4 pp 657ndash662 2009
[25] D Ekuni T Tomofuji N Tamaki et al ldquoMechanical stimulationof gingiva reduces plasma 8-OHdG level in rat periodontitisrdquoArchives of Oral Biology vol 53 no 4 pp 324ndash329 2008
[26] H Staudte A Guntsch A Volpel and B W Sigusch ldquoVitaminC attenuates the cytotoxic effects of Porphyromonas gingivalison human gingival fibroblastsrdquo Archives of Oral Biology vol 55no 1 pp 40ndash45 2010
[27] P Bullon H N Newman and M Battino ldquoObesity diabetesmellitus atherosclerosis and chronic periodontitis a sharedpathology via oxidative stress andmitochondrial dysfunctionrdquoPeriodontology 2000 vol 64 no 1 pp 139ndash153 2014
[28] G R Naveh V Brumfeld R Shahar and S Weiner ldquoToothperiodontal ligament direct 3D microCT visualization of thecollagen network and how the network changes when the toothis loadedrdquo Journal of Structural Biology vol 181 no 2 pp 108ndash115 2013
[29] Y Kitase M Yokozeki S Fujihara et al ldquoAnalysis of geneexpression profiles in human periodontal ligament cells underhypoxia the protective effect of CC chemokine ligand 2 tooxygen shortagerdquoArchives of Oral Biology vol 54 no 7 pp 618ndash624 2009
[30] G Thornton-Evans P Eke L Wei et al ldquoPeriodontitis amongadults aged ge 30 years - United States 2009-2010rdquo Morbidityand Mortality Weekly Report (MMWR) vol 62 supplement 3pp 129ndash135 2013
[31] S S Socransky C Smith and A D Haffajee ldquoSubgingivalmicrobial profiles in refractory periodontal diseaserdquo Journal ofClinical Periodontology vol 29 no 3 pp 260ndash268 2002
[32] K-T Ng J-P Li K M Ng G L Tipoe W K Leung and M-L Fung ldquoExpression of hypoxia-inducible factor-1120572 in humanperiodontal tissuerdquo Journal of Periodontology vol 82 no 1 pp136ndash141 2011
[33] S Frede U Berchner-Pfannschmidt and J Fandrey ldquoReg-ulation of Hypoxia-inducible factors during inflammationrdquoMethods in Enzymology vol 435 pp 405ndash419 2007
[34] J Shin S Ji and Y Choi ldquoAbility of oral bacteria to inducetissue-destructive molecules from human neutrophilsrdquo OralDiseases vol 14 no 4 pp 327ndash334 2008
[35] S Takashiba K Naruishi and Y Murayama ldquoPerspectiveof cytokine regulation for periodontal treatment fibroblastbiologyrdquo Journal of Periodontology vol 74 no 1 pp 103ndash1102003
[36] A Konermann D Stabenow P A Knolle S A E HeldJ Deschner and A Jager ldquoRegulatory role of periodontalligament fibroblasts for innate immune cell function and dif-ferentiationrdquo Innate Immunity vol 18 no 5 pp 745ndash752 2012
[37] S Kar S Subbaram P M Carrico and J A Melendez ldquoRedox-control of matrix metalloproteinase-1 a critical link betweenfree radicals matrix remodeling and degenerative diseaserdquoRespiratory Physiology andNeurobiology vol 174 no 3 pp 299ndash306 2010
[38] F Rousset M V C Nguyen L Grange F Morel and B LardyldquoHeme oxygenase-1 regulates matrix metalloproteinase MMP-1secretion and chondrocyte cell death via Nox4NADPH oxidaseactivity in chondrocytesrdquo PLoS ONE vol 8 no 6 Article IDe66478 2013
[39] E Rudolf and M Cervinka ldquoResponses of human gingival andperiodontal fibroblasts to a low-zinc environmentrdquo Alternativesto Laboratory Animals vol 38 no 2 pp 119ndash138 2010
[40] S M San Miguel L A Opperman E P Allen J Zielinski andK K H Svoboda ldquoBioactive polyphenol antioxidants protectoral fibroblasts from ROS-inducing agentsrdquo Archives of OralBiology vol 57 no 12 pp 1657ndash1667 2012
[41] S M San Miguel L A Opperman E P Allen J E Zielinskiand K K H Svoboda ldquoAntioxidant combinations protect oralfibroblasts against metal-induced toxicityrdquo Archives of OralBiology vol 58 no 3 pp 299ndash310 2013
[42] U Skaleric C M Manthey S E Mergenhagen B Gaspircand S M Wahl ldquoSuperoxide release and superoxide dismutaseexpression by human gingival fibroblastsrdquo European Journal ofOral Sciences vol 108 no 2 pp 130ndash135 2000
Mediators of Inflammation 13
[43] D Y Kim J-H Jun H-L Lee et al ldquoN-acetylcysteine preventsLPS-Lnduced pro-inflammatory cytokines andMMP2 produc-tion in gingival fibroblastsrdquoArchives of Pharmacal Research vol30 no 10 pp 1283ndash1292 2007
[44] B Rath-Deschner J Deschner S Reimann A Jager andW Gotz ldquoRegulatory effects of biomechanical strain on theinsulin-like growth factor system in human periodontal cellsrdquoJournal of Biomechanics vol 42 no 15 pp 2584ndash2589 2009
[45] P Chomczynski and N Sacchi ldquoSingle-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextractionrdquo Analytical Biochemistry vol 162 no 1 pp 156ndash1591987
[46] B Zhao K Ranguelova J Jiang and R P Mason ldquoStudies onthe photosensitized reduction of resorufin and implications forthe detection of oxidative stress with Amplex Redrdquo Free RadicalBiology and Medicine vol 51 no 1 pp 153ndash159 2011
[47] C C Mandal S Ganapathy Y Gorin et al ldquoReactive oxygenspecies derived from Nox4 mediate BMP2 gene transcriptionand osteoblast differentiationrdquoBiochemical Journal vol 433 no2 pp 393ndash402 2011
[48] J Fandrey S Frede and W Jelkmann ldquoRole of hydrogenperoxide in hypoxia-induced erythropoietin productionrdquo Bio-chemical Journal vol 303 no 2 pp 507ndash510 1994
[49] F Cervellati C Cervellati A Romani et al ldquoHypoxia inducescell damage via oxidative stress in retinal epithelial cellsrdquo FreeRadical Research vol 48 no 3 pp 303ndash312 2014
[50] V Vanasco N D Magnani M C Cimolai et al ldquoEndotoxemiaimpairs heart mitochondrial function by decreasing electrontransfer ATP synthesis and ATP content without affectingmembrane potentialrdquo Journal of Bioenergetics and Biomem-branes vol 44 no 2 pp 243ndash252 2012
[51] R G Alscher N Erturk and L S Heath ldquoRole of superoxidedismutases (SODs) in controlling oxidative stress in plantsrdquoJournal of Experimental Botany vol 53 no 372 pp 1331ndash13412002
[52] E Baltacıoglu P Yuva G Aydın et al ldquoLipid peroxidation levelsand total oxidantantioxidant status in serum and saliva frompatients with chronic and aggressive periodontitis Oxidativestress index a new biomarker for periodontal diseaserdquo Journalof Periodontology vol 85 no 10 pp 1432ndash1441 2014
[53] I L C Chapple M R Milward N Ling-Mountford et alldquoAdjunctive daily supplementation with encapsulated fruitvegetable and berry juice powder concentrates and clinicalperiodontal outcomes a double-blind RCTrdquo Journal of ClinicalPeriodontology vol 39 no 1 pp 62ndash72 2012
[54] A Jenzsch S Eick F Rassoul R Purschwitz and H JentschldquoNutritional intervention in patients with periodontal diseaseClinical immunological and microbiological variables during12 monthsrdquo British Journal of Nutrition vol 101 no 6 pp 879ndash885 2009
[55] N Novakovic T Todorovic M Rakic et al ldquoSalivary antiox-idants as periodontal biomarkers in evaluation of tissue statusand treatment outcomerdquo Journal of Periodontal Research vol49 no 1 pp 129ndash136 2014
[56] N Singh S C Narula R K Sharma S Tewari and P KSehgal ldquoVitamin e supplementation superoxide dismutasestatus and outcome of scaling and root planing in patients withchronic periodontitis a randomized clinical trialrdquo Journal ofPeriodontology vol 85 no 2 pp 242ndash249 2014
[57] S Krifka K-A Hiller G Spagnuolo A Jewett G Schmalz andH Schweikl ldquoThe influence of glutathione on redox regulation
by antioxidant proteins and apoptosis in macrophages exposedto 2-hydroxyethyl methacrylate (HEMA)rdquo Biomaterials vol 33no 21 pp 5177ndash5186 2012
[58] I Borges Jr E A M Moreira D W Filho T B de Oliveira MB S da Silva and T S Frode ldquoProinflammatory and oxidativestress markers in patients with periodontal diseaserdquo Mediatorsof Inflammation vol 2007 Article ID 45794 5 pages 2007
[59] M O Tonguc O Ozturk R Sutcu et al ldquoThe impact of smok-ing status on antioxidant enzyme activity and malondialdehydelevels in chronic periodontitisrdquo Journal of Periodontology vol82 no 9 pp 1320ndash1328 2011
[60] B H Jacoby and W L Davis ldquoThe electron microscopicimmunolocalization of a copper-zinc superoxide dismutase inassociation with collagen fibers of periodontal soft tissuesrdquoJournal of Periodontology vol 62 no 7 pp 413ndash420 1991
[61] T Konopka K Krol W Kopec and H Gerber ldquoTotal antioxi-dant status and 8-hydroxy-21015840-deoxyguanosine levels in gingi-val and peripheral blood of periodontitis patientsrdquo ArchivumImmunologiae et Therapiae Experimentalis vol 55 no 6 pp417ndash422 2007
[62] M Soory ldquoRedox status in periodontal and systemic inflamma-tory conditions including associated neoplasias antioxidants asadjunctive therapyrdquo Infectious DisordersmdashDrug Targets vol 9no 4 pp 415ndash427 2009
[63] M Soory ldquoOxidative stress inducedmechanisms in the progres-sion of periodontal diseases and cancer a common approach toredox homeostasisrdquo Cancers vol 2 no 2 pp 670ndash692 2010
[64] R V Chandra G Srinivas A A Reddy et al ldquoLocally deliveredantioxidant gel as an adjunct to nonsurgical therapy improvesmeasures of oxidative stress and periodontal diseaserdquo Journal ofPeriodontal and Implant Science vol 43 no 3 pp 121ndash129 2013
[65] M A D S Siqueira R G Fischer N R Pereira et al ldquoEffectsof non-surgical periodontal treatment on the L-arginine-nitricoxide pathway and oxidative status in plateletsrdquo ExperimentalBiology and Medicine vol 238 no 6 pp 713ndash722 2013
[66] P Mydel Y Takahashi H Yumoto et al ldquoRoles of the hostoxidative immune response and bacterial antioxidant rubr-erythrin during Porphyromonas gingivalis infectionrdquo PLoSpathogens vol 2 no 7 p e76 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
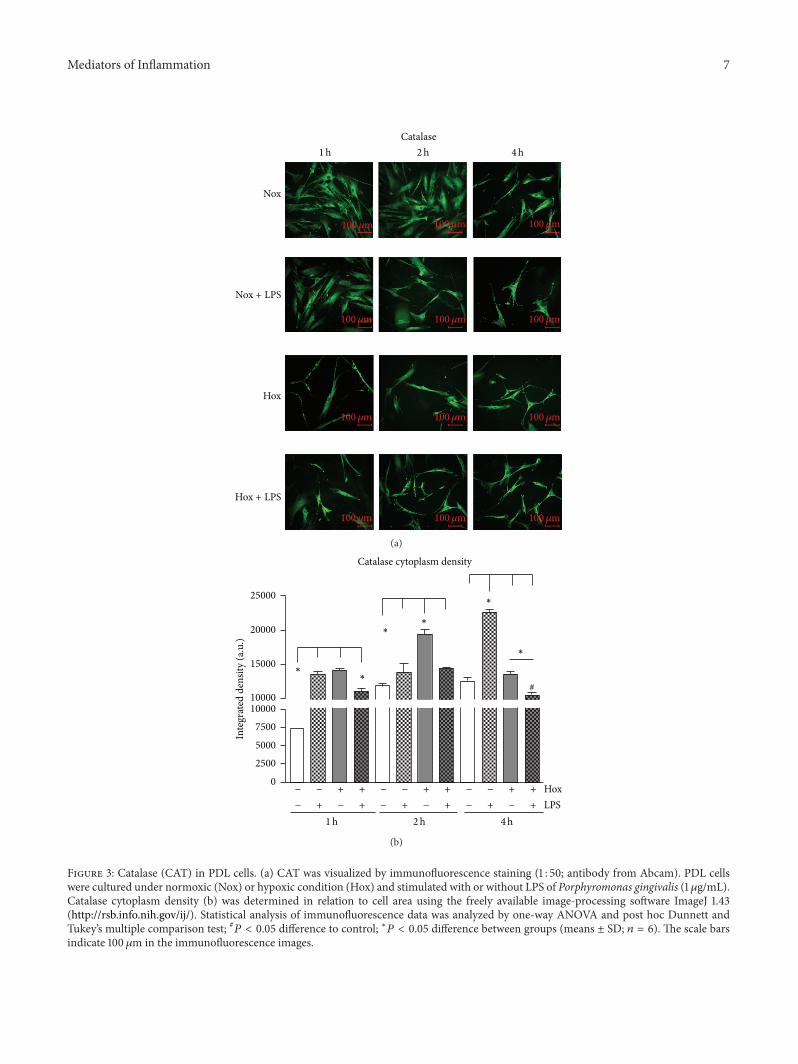
Mediators of Inflammation 7
1h 2hCatalase
Nox
Nox + LPS
Hox
Hox + LPS
4h
100120583m 100120583m 100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
100120583m100120583m100120583m
(a)
2h 4h
Hox
Inte
grat
ed d
ensit
y (a
u)
Catalase cytoplasm density
LPSminusminus + + minusminus + + minusminus +
minus +minus +minus +minus +minus +minus +
+
lowast
lowast
lowast
lowast
lowastlowast
1h
20000
25000
15000
1000010000
7500
5000
2500
0
(b)
Figure 3 Catalase (CAT) in PDL cells (a) CAT was visualized by immunofluorescence staining (1 50 antibody from Abcam) PDL cellswere cultured under normoxic (Nox) or hypoxic condition (Hox) and stimulated with or without LPS of Porphyromonas gingivalis (1120583gmL)Catalase cytoplasm density (b) was determined in relation to cell area using the freely available image-processing software ImageJ 143(httprsbinfonihgovij) Statistical analysis of immunofluorescence data was analyzed by one-way ANOVA and post hoc Dunnett andTukeyrsquos multiple comparison test 119875 lt 005 difference to control lowast119875 lt 005 difference between groups (means plusmn SD 119899 = 6) The scale barsindicate 100 120583m in the immunofluorescence images
8 Mediators of Inflammation
50120583m
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m
50120583m
50120583m
Inflamed
(d)
Figure 4 NADPH-oxidase 4 (NOX4) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Monoclonal primary antibody against NOX4 (Abcam Cambridge UK) was used ina concentration of 1 100 In gingival and PDL fibroblasts as well as in immune cells like leukocytes and in vessels the immunoreactivity wasprimarily restricted to the cytosol and cell walls Red arrows indicate examples for the typical distribution (Figures 4(c) and 4(d)) The scalebars indicate 200120583m in the surveys and 50 120583m in the higher magnifications respectively
ligament cells We showed that both stimuli induced asignificant increase of NOX4 and a significant upregulationof hydrogen peroxide generation NOX enzymes were firstdescribed in phagocytes as being responsible for defensemechanisms against pathogens using NOX-generated ROSmolecules Till then NOX4 expression has been detected innonphagocyte cell types like kidney cells osteoclasts andosteoblasts as well as dermal cells [9 11 47] In PDL cellshypoxia and LPS-PG induced a significant increase of NOX4mRNA The protein expression was mainly located in thecytosol but likewise in the nucleus Our findings were inline with Brown and Griendling who have shown that thetypically transmembrane molecule is localized in differentcell compartments [9]
NOX4 is one of the major sources of ROS generationwhich is highly and ubiquitously expressed compared tothe other family members Mandal and colleagues demon-strated that apart from its antimicrobial function NOX4 isresponsible not only for osteoclast but also for preosteoblastdifferentiation via ROS accumulation reinforcing the ideaof NOX4 as a key regulator in bone remodeling underphysiological ROS amounts [47] Animal studies supportthe hypothesis that massive ROS formation induced bone
resorption via NOX4 [20 21] Whereas physiological levelsof NOX4-generated ROS in PDL cellsmight facilitate alveolarbone remodeling massive ROS accumulation during inflam-matory periodontal diseases might lead to alveolar bonedestruction favoring tooth loss We have demonstrated firstthat NOX4 is upregulated in periodontal ligament fibroblastsduring bacterial and hypoxic events Hence PDL fibroblastsmay be a source of excessive ROS generation and are thereforeof particular importance during periodontal diseases
Our results support current findings determining the roleof oxidative stress in different periodontal pathologies whichimplicate that the balance between NADPH-oxidases andredox systems is crucial to maintain cell and tissue home-ostasis [22ndash26] However there are only a few studies usinghuman periodontal fibroblast in the context of oxidativestress and none of them evaluated the role of NOX4 andcounteracting redox systems during inflammatory conditionswith LPS-PG stimulation and hypoxia An increase of ROSgeneration was detected by San Miguel et al following PDLfibroblast stimulation with nicotine or ethanol which wereabolished after antioxidant treatment [40] They completedtheir results using metal ions like nickel for PDL stimulationwhich increases ROS activity in these cells indicating a role
Mediators of Inflammation 9
200120583m
50120583m
50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 5 Catalase (CAT) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontal ligament (b) andsamples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University of Bonn and parentalas well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against catalase (Abcam Cambridge UK) was used in a concentration of1 100 No immunostaining could be found in the healthy gingival epithelium In the healthy PDL predominantly PDL cells located near tothe tooth root surfaces were stained (Figure 5(b) red arrow) In gingivitis tissue a weak staining of gingival keratinocytes similar to healthytissue could be seen (Figure 5(c)) Instead in the tissue samples of patients with periodontitis a strong immunoreactivity was observed inthe subepithelial layer probably referring to nuclei staining of immune cells like leukocytes Red arrows indicate examples for the typicallocalisation (Figure 5(d)) The scale bars indicate 200 120583m in the surveys and 50120583m in the higher magnifications respectively
in metal cytotoxicity or even nickel allergy [41] Transientmetal ions like nickel and cobalt are known to mimic cellularhypoxia mainly due to the inhibition of prolyl hydrolases(PDHs) and asparaginyl hydrolase (factor-inhibiting HIF(FIH)) The inhibition of hydroxylation leads to HIF-1120572nuclear translocation followed by the transcription of diversegenes which are implicated in hypoxic adaptation immuneresponse or angiogenesis therefore indirectly supporting ourresults under hypoxic condition [48 49]
We could show that hypoxia as well as LPS-PG inducedROS formation by detecting H
2O2using the commercially
available Amplex Red assay The high levels of H2O2released
under hypoxic conditions after 1 and 2 hours in PDL cellsmay be caused by different mechanisms First we founda significant upregulation of the mRNA and protein ofthe H
2O2-generating enzyme NOX4 Second the lack of
molecular oxygen as the final electron acceptor in therespiratory chain may be responsible for the early release of
ROS LPS-PG alone induced acutely a small but significantincrease in H
2O2release and enhanced the hypoxia-induced
generation In human gingival fibroblasts similar results wereobtained showing the increase of ROS and antioxidants afterP gingivalis or LPS stimulation [26 42 43] Interestinglydespite the increase in NOX4 mRNA and protein expressionafter 4 hours of stimulation with LPS-PG under hypoxicconditions the H
2O2generation was significantly reduced
when compared to hypoxia alone An additional inhibitionof the respiratory chain by LPS-PG which was described forcardiac mitochondria [50] is therefore conceivable
To antagonize oxidative stress cells developed severalantioxidative enzyme systems Therefore we measured SODand CAT expression in PDL cells and periodontal tissues byimmunofluorescence and immunohistochemistry SODs aremetalloenzymes which are responsible for the dismutation ofsuperoxide radicals to hydrogen peroxide [51] The reductionof hydrogen peroxide to water and oxygen is subsequently
10 Mediators of Inflammation
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m 50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 6 Superoxide dismutase (SOD) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against SOD (Abcam Cambridge UK) was used ina concentration of 1 100 Gingival epithelium showed a moderate to strong staining of SOD in keratinocytes which was primarily limitedto the cytosol (Figure 6(a) and red arrow depicts the typical localization) In the healthy periodontium gingival and PDL fibroblasts wereweakly stained (Figure 6(b)) In inflamed tissues basal epithelial cells seemed to be stained more intensively in the gingivitis specimen thanin periodontitis (Figures 6(c) and 6(d)) In addition the subepithelium revealed immunoreactivity in local cells like gingival fibroblasts andscattered immune cells (Figure 6(c) see red arrow) In periodontitis SOD immunostaining seemed to be limited to the gingival epithelium(Figure 6(d) see red arrow) The scale bars indicate 200 120583m in the surveys and 50 120583m in the higher magnifications respectively
catalyzed by CATs which are ubiquitously expressed Bothenzymes are essential to maintain cellular redox homeostasisThe present data imply that PDL cells and periodontal tissueshave fundamental defense machineries against bacteria rep-resented by NOX4-dependent inducible ROS generation aswell as protective redox systems This is of great importanceconcerning the high amounts and diversity of the oral floraDifferent studies indicate that redox systems are key playersin the development of periodontal diseases and markers forperiodontal health status [52ndash56] We detected a significantincrease of catalase in PDL cells in response to LPS aswell as hypoxia after 1 and 2 hours In our setting PDLfibroblasts seem to counteracted LPS- and hypoxia-inducedROS formation with the elevation of CAT expression Butprolonged incubation under hypoxic conditions failed toincrease catalase Since ROS generation was still elevated atthese later time points a delay in ROS degradation may be
the explanation Taken together the dramatic elevation inROS formation by the combination of hypoxia and LPS-PGon one hand and the reduction of catalase under the sameconditions on the other hand increase the oxidative stress inthe periodontium favoring the progression of inflammationand bone resorption and finally may lead to tooth loss
The fundamental role of the balance between ROSgeneration and antioxidative mechanisms in periodontalinflammatory processes was supported by our immunohis-tochemical findings In patients suffering from periodontalinflammatory diseases like gingivitis and periodontitis wedemonstrated that NOX4 is upregulated and thus likelyresponsible for H
2O2generation However we detected as
well an increase in catalase abundance in inflamed gingivaland periodontal tissue which was in line with data fromKrifka and colleagues who detected high CAT levels inmonomer-exposed macrophages as a response to oxidative
Mediators of Inflammation 11
stress [57] In our in vitro setting PDL fibroblasts seemto counteract LPS- and hypoxia-induced ROS formationpredominantly with an elevation of CAT expression sincewe could not detect any SOD immunoreactivity The roleand impact of the antioxidative systems SOD and CAT arecontroversially discussed Recent studies measured CAT andSOD activities in gingival tissues of patients with periodon-titis [58 59] Borges Jr et al did not find differences inCAT activities comparing healthy and diseased patients [58]Instead Tonguc et al detected decreased local SOD andCAT activities in periodontitis patients which seemed to beincreased by smoking [59] In contrast in inflamed tissues weobservedhigh amounts of SODwhichwas localized primarilyin epithelial cells Jacoby and Davis determined SOD in peri-odontal soft tissues They detected twice as much SOD activ-ity in the periodontal ligament as in the human skin (dermis)[60] Furthermore immunohistochemical analysis revealedthat SOD is localized in the periphery of matrix collagenfibrils and to the glycocalyx of tissue fibroblasts This wasconfirmed by Skaleric et al [42] which might explain the factthat we did not detect significant differences of SOD levelsafter hypoxic and LPS-PG stimulation intracellularly in PDLfibroblasts Another reason for these results may be causedby the age of the PDL cells used in this study We explantedthe PDL cells from young and oral healthy patients but mostof the immunohistochemical stainings were performed intissues from elderly patients with clearly diagnosed gingivitisor periodontitisTherefore it is possible that in PDL cells fromelderly patients a different expression pattern of antioxidantenzyme systems could be found which further deterioratethe redox state of the periodontal ligament Moreover otherauthors found reduced antioxidants in the body fluids ofpatients with periodontitis indicating a dysregulation whichwould favor disease progression [23 24 61] A variety of invitro and animal studies analyzed the effect of antioxidantsupplement in the context of inflammatory or cytotoxicevents [57 62 63] They have shown that antioxidantsinhibit these processes indicating a therapeutic approachin pathologies like periodontitis First clinical investigationsconfirmed the efficiency of antioxidant micronutrients andadjuncts in periodontal diseases [22 54 56 64 65] Thepresent data indirectly support these studies
Concerning periodontal pathogen invasiveness and viru-lence there is compelling evidence that P gingivalis possessesits own protective antioxidants for example rubrerythrindefending them against the oxidative burst of the host [66]We have shown that its virulence factor LPS-PG inducedhigh amounts of ROS via NOX4 which may result in boneresorption favoring the invasiveness of the pathogen whichis at the same time protected by its own defense machineryThis would further explain the pathogenicity and resistanceof this bacterium strain in patients with periodontitis
5 Conclusion
In this study we have demonstrated that LPS from Porphy-romonas gingivalis (LPS-PG) as well as hypoxia induces aNOX4-dependent increase inH
2O2release in PDLfibroblasts
which may contribute to the development and progressionof periodontal diseases when not balanced by a concomitantincrease in antioxidative systems The dramatic elevationin ROS formation induced by the combination of hypoxiaand LPS-PG and the reduction of catalase under the sameconditions increased the oxidative stress in the periodontiumfavoring the progression of inflammation bone resorptionand possibly tooth loss
Taken together the presented data further explain thedestructive mechanism of hypoxic and inflammatory condi-tions in the periodontium and may improve diagnostic andtherapeutic strategies in periodontal diseases
Conflict of Interests
The authors have no conflict of interests to disclose
Acknowledgments
The authors would like to thank Ms Tina Schaffrath for hertechnical supportThe present studywas supported by a grantfrom the German Research Foundation (Clinical ResearchUnit 208TP7) and the Medical Faculty of the University ofBonn
References
[1] S Reuter S C Gupta M M Chaturvedi and B B AggarwalldquoOxidative stress inflammation and cancer how are theylinkedrdquo Free Radical Biology and Medicine vol 49 no 11 pp1603ndash1616 2010
[2] H Sone H Akanuma and T Fukuda ldquoOxygenomics inenvironmental stressrdquo Redox Report vol 15 no 3 pp 98ndash1142010
[3] K E Wellen and C B Thompson ldquoCellular metabolic stressconsidering how cells respond to nutrient excessrdquo MolecularCell vol 40 no 2 pp 323ndash332 2010
[4] R O Poyton K A Ball and P R Castello ldquoMitochondrialgeneration of free radicals and hypoxic signalingrdquo Trends inEndocrinology andMetabolism vol 20 no 7 pp 332ndash340 2009
[5] F Weinberg and N S Chandel ldquoReactive oxygen species-dependent signaling regulates cancerrdquo Cellular and MolecularLife Sciences vol 66 no 23 pp 3663ndash3673 2009
[6] A Cataldi ldquoCell responses to oxidative stressorsrdquo CurrentPharmaceutical Design vol 16 no 12 pp 1387ndash1395 2010
[7] Z Durackova ldquoSome current insights into oxidative stressrdquoPhysiological Research vol 59 no 4 pp 459ndash469 2010
[8] L A Sena and N S Chandel ldquoPhysiological roles of mitochon-drial reactive oxygen speciesrdquoMolecular Cell vol 48 no 2 pp158ndash166 2012
[9] D I Brown and K K Griendling ldquoNox proteins in signaltransductionrdquo Free Radical Biology amp Medicine vol 47 no 9pp 1239ndash1253 2009
[10] K Bedard and K-H Krause ldquoThe NOX family of ROS-generatingNADPHoxidases physiology and pathophysiologyrdquoPhysiological Reviews vol 87 no 1 pp 245ndash313 2007
[11] K-H Krause ldquoTissue distribution and putative physiologicalfunction of NOX family NADPH oxidasesrdquo Japanese Journal ofInfectious Diseases vol 57 no 5 pp S28ndashS29 2004
12 Mediators of Inflammation
[12] S Yang Y Zhang W Ries and L Key ldquoExpression of Nox4 inosteoclastsrdquo Journal of Cellular Biochemistry vol 92 no 2 pp238ndash248 2004
[13] E Pedruzzi C Guichard V Ollivier et al ldquoNAD(P)H oxidaseNox-4mediates 7-ketocholesterol-induced endoplasmic reticu-lum stress and apoptosis in human aortic smooth muscle cellsrdquoMolecular and Cellular Biology vol 24 no 24 pp 10703ndash107172004
[14] J Hwang A Saha Y C Boo et al ldquoOscillatory shear stress stim-ulates endothelial production of O
2-from p47119901ℎ119900119909-dependent
NAD(P)Hoxidases leading tomonocyte adhesionrdquoThe Journalof Biological Chemistry vol 278 no 47 pp 47291ndash47298 2003
[15] H B Suliman M Ali and C A Piantadosi ldquoSuperoxidedismutase-3 promotes full expression of the EPO response tohypoxiardquo Blood vol 104 no 1 pp 43ndash50 2004
[16] P Vallet Y Charnay K Steger et al ldquoNeuronal expressionof the NADPH oxidase NOX4 and its regulation in mouseexperimental brain ischemiardquo Neuroscience vol 132 no 2 pp233ndash238 2005
[17] K T Moe S Aulia F Jiang et al ldquoDifferential upregulation ofNox homologues of NADPH oxidase by tumor necrosis factor-120572 in human aortic smooth muscle and embryonic kidney cellsrdquoJournal of Cellular andMolecularMedicine vol 10 no 1 pp 231ndash239 2006
[18] I Diebold A Petry J Hess and A Gorlach ldquoThe NADPHoxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1rdquoMolecular Biology of the Cell vol 21 no 12pp 2087ndash2096 2010
[19] K V Tormos and N S Chandel ldquoInter-connection betweenmitochondria and HIFsrdquo Journal of Cellular and MolecularMedicine vol 14 no 4 pp 795ndash804 2010
[20] T Ohnishi K Bandow K Kakimoto M Machigashira TMatsuyama and T Matsuguchi ldquoOxidative stress causes alve-olar bone loss in metabolic syndrome model mice with type 2diabetesrdquo Journal of Periodontal Research vol 44 no 1 pp 43ndash51 2009
[21] C Goettsch A Babelova O Trummer et al ldquoNADPH oxidase4 limits bone mass by promoting osteoclastogenesisrdquo Journal ofClinical Investigation vol 123 no 11 pp 4731ndash4738 2013
[22] I L C Chapple and J B Matthews ldquoThe role of reactive oxygenand antioxidant species in periodontal tissue destructionrdquoPeriodontology 2000 vol 43 no 1 pp 160ndash232 2007
[23] A Guentsch P M Preshaw S Bremer-Streck G KlingerE Glockmann and B W Sigusch ldquoLipid peroxidation andantioxidant activity in saliva of periodontitis patients effect ofsmoking and periodontal treatmentrdquo Clinical Oral Investiga-tions vol 12 no 4 pp 345ndash352 2008
[24] R Agnihotri P Pandurang S U Kamath et al ldquoAssociation ofcigarette smoking with superoxide dismutase enzyme levels insubjected with chonic periodontitisrdquo Journal of Periodontologyvol 80 no 4 pp 657ndash662 2009
[25] D Ekuni T Tomofuji N Tamaki et al ldquoMechanical stimulationof gingiva reduces plasma 8-OHdG level in rat periodontitisrdquoArchives of Oral Biology vol 53 no 4 pp 324ndash329 2008
[26] H Staudte A Guntsch A Volpel and B W Sigusch ldquoVitaminC attenuates the cytotoxic effects of Porphyromonas gingivalison human gingival fibroblastsrdquo Archives of Oral Biology vol 55no 1 pp 40ndash45 2010
[27] P Bullon H N Newman and M Battino ldquoObesity diabetesmellitus atherosclerosis and chronic periodontitis a sharedpathology via oxidative stress andmitochondrial dysfunctionrdquoPeriodontology 2000 vol 64 no 1 pp 139ndash153 2014
[28] G R Naveh V Brumfeld R Shahar and S Weiner ldquoToothperiodontal ligament direct 3D microCT visualization of thecollagen network and how the network changes when the toothis loadedrdquo Journal of Structural Biology vol 181 no 2 pp 108ndash115 2013
[29] Y Kitase M Yokozeki S Fujihara et al ldquoAnalysis of geneexpression profiles in human periodontal ligament cells underhypoxia the protective effect of CC chemokine ligand 2 tooxygen shortagerdquoArchives of Oral Biology vol 54 no 7 pp 618ndash624 2009
[30] G Thornton-Evans P Eke L Wei et al ldquoPeriodontitis amongadults aged ge 30 years - United States 2009-2010rdquo Morbidityand Mortality Weekly Report (MMWR) vol 62 supplement 3pp 129ndash135 2013
[31] S S Socransky C Smith and A D Haffajee ldquoSubgingivalmicrobial profiles in refractory periodontal diseaserdquo Journal ofClinical Periodontology vol 29 no 3 pp 260ndash268 2002
[32] K-T Ng J-P Li K M Ng G L Tipoe W K Leung and M-L Fung ldquoExpression of hypoxia-inducible factor-1120572 in humanperiodontal tissuerdquo Journal of Periodontology vol 82 no 1 pp136ndash141 2011
[33] S Frede U Berchner-Pfannschmidt and J Fandrey ldquoReg-ulation of Hypoxia-inducible factors during inflammationrdquoMethods in Enzymology vol 435 pp 405ndash419 2007
[34] J Shin S Ji and Y Choi ldquoAbility of oral bacteria to inducetissue-destructive molecules from human neutrophilsrdquo OralDiseases vol 14 no 4 pp 327ndash334 2008
[35] S Takashiba K Naruishi and Y Murayama ldquoPerspectiveof cytokine regulation for periodontal treatment fibroblastbiologyrdquo Journal of Periodontology vol 74 no 1 pp 103ndash1102003
[36] A Konermann D Stabenow P A Knolle S A E HeldJ Deschner and A Jager ldquoRegulatory role of periodontalligament fibroblasts for innate immune cell function and dif-ferentiationrdquo Innate Immunity vol 18 no 5 pp 745ndash752 2012
[37] S Kar S Subbaram P M Carrico and J A Melendez ldquoRedox-control of matrix metalloproteinase-1 a critical link betweenfree radicals matrix remodeling and degenerative diseaserdquoRespiratory Physiology andNeurobiology vol 174 no 3 pp 299ndash306 2010
[38] F Rousset M V C Nguyen L Grange F Morel and B LardyldquoHeme oxygenase-1 regulates matrix metalloproteinase MMP-1secretion and chondrocyte cell death via Nox4NADPH oxidaseactivity in chondrocytesrdquo PLoS ONE vol 8 no 6 Article IDe66478 2013
[39] E Rudolf and M Cervinka ldquoResponses of human gingival andperiodontal fibroblasts to a low-zinc environmentrdquo Alternativesto Laboratory Animals vol 38 no 2 pp 119ndash138 2010
[40] S M San Miguel L A Opperman E P Allen J Zielinski andK K H Svoboda ldquoBioactive polyphenol antioxidants protectoral fibroblasts from ROS-inducing agentsrdquo Archives of OralBiology vol 57 no 12 pp 1657ndash1667 2012
[41] S M San Miguel L A Opperman E P Allen J E Zielinskiand K K H Svoboda ldquoAntioxidant combinations protect oralfibroblasts against metal-induced toxicityrdquo Archives of OralBiology vol 58 no 3 pp 299ndash310 2013
[42] U Skaleric C M Manthey S E Mergenhagen B Gaspircand S M Wahl ldquoSuperoxide release and superoxide dismutaseexpression by human gingival fibroblastsrdquo European Journal ofOral Sciences vol 108 no 2 pp 130ndash135 2000
Mediators of Inflammation 13
[43] D Y Kim J-H Jun H-L Lee et al ldquoN-acetylcysteine preventsLPS-Lnduced pro-inflammatory cytokines andMMP2 produc-tion in gingival fibroblastsrdquoArchives of Pharmacal Research vol30 no 10 pp 1283ndash1292 2007
[44] B Rath-Deschner J Deschner S Reimann A Jager andW Gotz ldquoRegulatory effects of biomechanical strain on theinsulin-like growth factor system in human periodontal cellsrdquoJournal of Biomechanics vol 42 no 15 pp 2584ndash2589 2009
[45] P Chomczynski and N Sacchi ldquoSingle-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextractionrdquo Analytical Biochemistry vol 162 no 1 pp 156ndash1591987
[46] B Zhao K Ranguelova J Jiang and R P Mason ldquoStudies onthe photosensitized reduction of resorufin and implications forthe detection of oxidative stress with Amplex Redrdquo Free RadicalBiology and Medicine vol 51 no 1 pp 153ndash159 2011
[47] C C Mandal S Ganapathy Y Gorin et al ldquoReactive oxygenspecies derived from Nox4 mediate BMP2 gene transcriptionand osteoblast differentiationrdquoBiochemical Journal vol 433 no2 pp 393ndash402 2011
[48] J Fandrey S Frede and W Jelkmann ldquoRole of hydrogenperoxide in hypoxia-induced erythropoietin productionrdquo Bio-chemical Journal vol 303 no 2 pp 507ndash510 1994
[49] F Cervellati C Cervellati A Romani et al ldquoHypoxia inducescell damage via oxidative stress in retinal epithelial cellsrdquo FreeRadical Research vol 48 no 3 pp 303ndash312 2014
[50] V Vanasco N D Magnani M C Cimolai et al ldquoEndotoxemiaimpairs heart mitochondrial function by decreasing electrontransfer ATP synthesis and ATP content without affectingmembrane potentialrdquo Journal of Bioenergetics and Biomem-branes vol 44 no 2 pp 243ndash252 2012
[51] R G Alscher N Erturk and L S Heath ldquoRole of superoxidedismutases (SODs) in controlling oxidative stress in plantsrdquoJournal of Experimental Botany vol 53 no 372 pp 1331ndash13412002
[52] E Baltacıoglu P Yuva G Aydın et al ldquoLipid peroxidation levelsand total oxidantantioxidant status in serum and saliva frompatients with chronic and aggressive periodontitis Oxidativestress index a new biomarker for periodontal diseaserdquo Journalof Periodontology vol 85 no 10 pp 1432ndash1441 2014
[53] I L C Chapple M R Milward N Ling-Mountford et alldquoAdjunctive daily supplementation with encapsulated fruitvegetable and berry juice powder concentrates and clinicalperiodontal outcomes a double-blind RCTrdquo Journal of ClinicalPeriodontology vol 39 no 1 pp 62ndash72 2012
[54] A Jenzsch S Eick F Rassoul R Purschwitz and H JentschldquoNutritional intervention in patients with periodontal diseaseClinical immunological and microbiological variables during12 monthsrdquo British Journal of Nutrition vol 101 no 6 pp 879ndash885 2009
[55] N Novakovic T Todorovic M Rakic et al ldquoSalivary antiox-idants as periodontal biomarkers in evaluation of tissue statusand treatment outcomerdquo Journal of Periodontal Research vol49 no 1 pp 129ndash136 2014
[56] N Singh S C Narula R K Sharma S Tewari and P KSehgal ldquoVitamin e supplementation superoxide dismutasestatus and outcome of scaling and root planing in patients withchronic periodontitis a randomized clinical trialrdquo Journal ofPeriodontology vol 85 no 2 pp 242ndash249 2014
[57] S Krifka K-A Hiller G Spagnuolo A Jewett G Schmalz andH Schweikl ldquoThe influence of glutathione on redox regulation
by antioxidant proteins and apoptosis in macrophages exposedto 2-hydroxyethyl methacrylate (HEMA)rdquo Biomaterials vol 33no 21 pp 5177ndash5186 2012
[58] I Borges Jr E A M Moreira D W Filho T B de Oliveira MB S da Silva and T S Frode ldquoProinflammatory and oxidativestress markers in patients with periodontal diseaserdquo Mediatorsof Inflammation vol 2007 Article ID 45794 5 pages 2007
[59] M O Tonguc O Ozturk R Sutcu et al ldquoThe impact of smok-ing status on antioxidant enzyme activity and malondialdehydelevels in chronic periodontitisrdquo Journal of Periodontology vol82 no 9 pp 1320ndash1328 2011
[60] B H Jacoby and W L Davis ldquoThe electron microscopicimmunolocalization of a copper-zinc superoxide dismutase inassociation with collagen fibers of periodontal soft tissuesrdquoJournal of Periodontology vol 62 no 7 pp 413ndash420 1991
[61] T Konopka K Krol W Kopec and H Gerber ldquoTotal antioxi-dant status and 8-hydroxy-21015840-deoxyguanosine levels in gingi-val and peripheral blood of periodontitis patientsrdquo ArchivumImmunologiae et Therapiae Experimentalis vol 55 no 6 pp417ndash422 2007
[62] M Soory ldquoRedox status in periodontal and systemic inflamma-tory conditions including associated neoplasias antioxidants asadjunctive therapyrdquo Infectious DisordersmdashDrug Targets vol 9no 4 pp 415ndash427 2009
[63] M Soory ldquoOxidative stress inducedmechanisms in the progres-sion of periodontal diseases and cancer a common approach toredox homeostasisrdquo Cancers vol 2 no 2 pp 670ndash692 2010
[64] R V Chandra G Srinivas A A Reddy et al ldquoLocally deliveredantioxidant gel as an adjunct to nonsurgical therapy improvesmeasures of oxidative stress and periodontal diseaserdquo Journal ofPeriodontal and Implant Science vol 43 no 3 pp 121ndash129 2013
[65] M A D S Siqueira R G Fischer N R Pereira et al ldquoEffectsof non-surgical periodontal treatment on the L-arginine-nitricoxide pathway and oxidative status in plateletsrdquo ExperimentalBiology and Medicine vol 238 no 6 pp 713ndash722 2013
[66] P Mydel Y Takahashi H Yumoto et al ldquoRoles of the hostoxidative immune response and bacterial antioxidant rubr-erythrin during Porphyromonas gingivalis infectionrdquo PLoSpathogens vol 2 no 7 p e76 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 Mediators of Inflammation
50120583m
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m
50120583m
50120583m
Inflamed
(d)
Figure 4 NADPH-oxidase 4 (NOX4) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Monoclonal primary antibody against NOX4 (Abcam Cambridge UK) was used ina concentration of 1 100 In gingival and PDL fibroblasts as well as in immune cells like leukocytes and in vessels the immunoreactivity wasprimarily restricted to the cytosol and cell walls Red arrows indicate examples for the typical distribution (Figures 4(c) and 4(d)) The scalebars indicate 200120583m in the surveys and 50 120583m in the higher magnifications respectively
ligament cells We showed that both stimuli induced asignificant increase of NOX4 and a significant upregulationof hydrogen peroxide generation NOX enzymes were firstdescribed in phagocytes as being responsible for defensemechanisms against pathogens using NOX-generated ROSmolecules Till then NOX4 expression has been detected innonphagocyte cell types like kidney cells osteoclasts andosteoblasts as well as dermal cells [9 11 47] In PDL cellshypoxia and LPS-PG induced a significant increase of NOX4mRNA The protein expression was mainly located in thecytosol but likewise in the nucleus Our findings were inline with Brown and Griendling who have shown that thetypically transmembrane molecule is localized in differentcell compartments [9]
NOX4 is one of the major sources of ROS generationwhich is highly and ubiquitously expressed compared tothe other family members Mandal and colleagues demon-strated that apart from its antimicrobial function NOX4 isresponsible not only for osteoclast but also for preosteoblastdifferentiation via ROS accumulation reinforcing the ideaof NOX4 as a key regulator in bone remodeling underphysiological ROS amounts [47] Animal studies supportthe hypothesis that massive ROS formation induced bone
resorption via NOX4 [20 21] Whereas physiological levelsof NOX4-generated ROS in PDL cellsmight facilitate alveolarbone remodeling massive ROS accumulation during inflam-matory periodontal diseases might lead to alveolar bonedestruction favoring tooth loss We have demonstrated firstthat NOX4 is upregulated in periodontal ligament fibroblastsduring bacterial and hypoxic events Hence PDL fibroblastsmay be a source of excessive ROS generation and are thereforeof particular importance during periodontal diseases
Our results support current findings determining the roleof oxidative stress in different periodontal pathologies whichimplicate that the balance between NADPH-oxidases andredox systems is crucial to maintain cell and tissue home-ostasis [22ndash26] However there are only a few studies usinghuman periodontal fibroblast in the context of oxidativestress and none of them evaluated the role of NOX4 andcounteracting redox systems during inflammatory conditionswith LPS-PG stimulation and hypoxia An increase of ROSgeneration was detected by San Miguel et al following PDLfibroblast stimulation with nicotine or ethanol which wereabolished after antioxidant treatment [40] They completedtheir results using metal ions like nickel for PDL stimulationwhich increases ROS activity in these cells indicating a role
Mediators of Inflammation 9
200120583m
50120583m
50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 5 Catalase (CAT) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontal ligament (b) andsamples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University of Bonn and parentalas well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against catalase (Abcam Cambridge UK) was used in a concentration of1 100 No immunostaining could be found in the healthy gingival epithelium In the healthy PDL predominantly PDL cells located near tothe tooth root surfaces were stained (Figure 5(b) red arrow) In gingivitis tissue a weak staining of gingival keratinocytes similar to healthytissue could be seen (Figure 5(c)) Instead in the tissue samples of patients with periodontitis a strong immunoreactivity was observed inthe subepithelial layer probably referring to nuclei staining of immune cells like leukocytes Red arrows indicate examples for the typicallocalisation (Figure 5(d)) The scale bars indicate 200 120583m in the surveys and 50120583m in the higher magnifications respectively
in metal cytotoxicity or even nickel allergy [41] Transientmetal ions like nickel and cobalt are known to mimic cellularhypoxia mainly due to the inhibition of prolyl hydrolases(PDHs) and asparaginyl hydrolase (factor-inhibiting HIF(FIH)) The inhibition of hydroxylation leads to HIF-1120572nuclear translocation followed by the transcription of diversegenes which are implicated in hypoxic adaptation immuneresponse or angiogenesis therefore indirectly supporting ourresults under hypoxic condition [48 49]
We could show that hypoxia as well as LPS-PG inducedROS formation by detecting H
2O2using the commercially
available Amplex Red assay The high levels of H2O2released
under hypoxic conditions after 1 and 2 hours in PDL cellsmay be caused by different mechanisms First we founda significant upregulation of the mRNA and protein ofthe H
2O2-generating enzyme NOX4 Second the lack of
molecular oxygen as the final electron acceptor in therespiratory chain may be responsible for the early release of
ROS LPS-PG alone induced acutely a small but significantincrease in H
2O2release and enhanced the hypoxia-induced
generation In human gingival fibroblasts similar results wereobtained showing the increase of ROS and antioxidants afterP gingivalis or LPS stimulation [26 42 43] Interestinglydespite the increase in NOX4 mRNA and protein expressionafter 4 hours of stimulation with LPS-PG under hypoxicconditions the H
2O2generation was significantly reduced
when compared to hypoxia alone An additional inhibitionof the respiratory chain by LPS-PG which was described forcardiac mitochondria [50] is therefore conceivable
To antagonize oxidative stress cells developed severalantioxidative enzyme systems Therefore we measured SODand CAT expression in PDL cells and periodontal tissues byimmunofluorescence and immunohistochemistry SODs aremetalloenzymes which are responsible for the dismutation ofsuperoxide radicals to hydrogen peroxide [51] The reductionof hydrogen peroxide to water and oxygen is subsequently
10 Mediators of Inflammation
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m 50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 6 Superoxide dismutase (SOD) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against SOD (Abcam Cambridge UK) was used ina concentration of 1 100 Gingival epithelium showed a moderate to strong staining of SOD in keratinocytes which was primarily limitedto the cytosol (Figure 6(a) and red arrow depicts the typical localization) In the healthy periodontium gingival and PDL fibroblasts wereweakly stained (Figure 6(b)) In inflamed tissues basal epithelial cells seemed to be stained more intensively in the gingivitis specimen thanin periodontitis (Figures 6(c) and 6(d)) In addition the subepithelium revealed immunoreactivity in local cells like gingival fibroblasts andscattered immune cells (Figure 6(c) see red arrow) In periodontitis SOD immunostaining seemed to be limited to the gingival epithelium(Figure 6(d) see red arrow) The scale bars indicate 200 120583m in the surveys and 50 120583m in the higher magnifications respectively
catalyzed by CATs which are ubiquitously expressed Bothenzymes are essential to maintain cellular redox homeostasisThe present data imply that PDL cells and periodontal tissueshave fundamental defense machineries against bacteria rep-resented by NOX4-dependent inducible ROS generation aswell as protective redox systems This is of great importanceconcerning the high amounts and diversity of the oral floraDifferent studies indicate that redox systems are key playersin the development of periodontal diseases and markers forperiodontal health status [52ndash56] We detected a significantincrease of catalase in PDL cells in response to LPS aswell as hypoxia after 1 and 2 hours In our setting PDLfibroblasts seem to counteracted LPS- and hypoxia-inducedROS formation with the elevation of CAT expression Butprolonged incubation under hypoxic conditions failed toincrease catalase Since ROS generation was still elevated atthese later time points a delay in ROS degradation may be
the explanation Taken together the dramatic elevation inROS formation by the combination of hypoxia and LPS-PGon one hand and the reduction of catalase under the sameconditions on the other hand increase the oxidative stress inthe periodontium favoring the progression of inflammationand bone resorption and finally may lead to tooth loss
The fundamental role of the balance between ROSgeneration and antioxidative mechanisms in periodontalinflammatory processes was supported by our immunohis-tochemical findings In patients suffering from periodontalinflammatory diseases like gingivitis and periodontitis wedemonstrated that NOX4 is upregulated and thus likelyresponsible for H
2O2generation However we detected as
well an increase in catalase abundance in inflamed gingivaland periodontal tissue which was in line with data fromKrifka and colleagues who detected high CAT levels inmonomer-exposed macrophages as a response to oxidative
Mediators of Inflammation 11
stress [57] In our in vitro setting PDL fibroblasts seemto counteract LPS- and hypoxia-induced ROS formationpredominantly with an elevation of CAT expression sincewe could not detect any SOD immunoreactivity The roleand impact of the antioxidative systems SOD and CAT arecontroversially discussed Recent studies measured CAT andSOD activities in gingival tissues of patients with periodon-titis [58 59] Borges Jr et al did not find differences inCAT activities comparing healthy and diseased patients [58]Instead Tonguc et al detected decreased local SOD andCAT activities in periodontitis patients which seemed to beincreased by smoking [59] In contrast in inflamed tissues weobservedhigh amounts of SODwhichwas localized primarilyin epithelial cells Jacoby and Davis determined SOD in peri-odontal soft tissues They detected twice as much SOD activ-ity in the periodontal ligament as in the human skin (dermis)[60] Furthermore immunohistochemical analysis revealedthat SOD is localized in the periphery of matrix collagenfibrils and to the glycocalyx of tissue fibroblasts This wasconfirmed by Skaleric et al [42] which might explain the factthat we did not detect significant differences of SOD levelsafter hypoxic and LPS-PG stimulation intracellularly in PDLfibroblasts Another reason for these results may be causedby the age of the PDL cells used in this study We explantedthe PDL cells from young and oral healthy patients but mostof the immunohistochemical stainings were performed intissues from elderly patients with clearly diagnosed gingivitisor periodontitisTherefore it is possible that in PDL cells fromelderly patients a different expression pattern of antioxidantenzyme systems could be found which further deterioratethe redox state of the periodontal ligament Moreover otherauthors found reduced antioxidants in the body fluids ofpatients with periodontitis indicating a dysregulation whichwould favor disease progression [23 24 61] A variety of invitro and animal studies analyzed the effect of antioxidantsupplement in the context of inflammatory or cytotoxicevents [57 62 63] They have shown that antioxidantsinhibit these processes indicating a therapeutic approachin pathologies like periodontitis First clinical investigationsconfirmed the efficiency of antioxidant micronutrients andadjuncts in periodontal diseases [22 54 56 64 65] Thepresent data indirectly support these studies
Concerning periodontal pathogen invasiveness and viru-lence there is compelling evidence that P gingivalis possessesits own protective antioxidants for example rubrerythrindefending them against the oxidative burst of the host [66]We have shown that its virulence factor LPS-PG inducedhigh amounts of ROS via NOX4 which may result in boneresorption favoring the invasiveness of the pathogen whichis at the same time protected by its own defense machineryThis would further explain the pathogenicity and resistanceof this bacterium strain in patients with periodontitis
5 Conclusion
In this study we have demonstrated that LPS from Porphy-romonas gingivalis (LPS-PG) as well as hypoxia induces aNOX4-dependent increase inH
2O2release in PDLfibroblasts
which may contribute to the development and progressionof periodontal diseases when not balanced by a concomitantincrease in antioxidative systems The dramatic elevationin ROS formation induced by the combination of hypoxiaand LPS-PG and the reduction of catalase under the sameconditions increased the oxidative stress in the periodontiumfavoring the progression of inflammation bone resorptionand possibly tooth loss
Taken together the presented data further explain thedestructive mechanism of hypoxic and inflammatory condi-tions in the periodontium and may improve diagnostic andtherapeutic strategies in periodontal diseases
Conflict of Interests
The authors have no conflict of interests to disclose
Acknowledgments
The authors would like to thank Ms Tina Schaffrath for hertechnical supportThe present studywas supported by a grantfrom the German Research Foundation (Clinical ResearchUnit 208TP7) and the Medical Faculty of the University ofBonn
References
[1] S Reuter S C Gupta M M Chaturvedi and B B AggarwalldquoOxidative stress inflammation and cancer how are theylinkedrdquo Free Radical Biology and Medicine vol 49 no 11 pp1603ndash1616 2010
[2] H Sone H Akanuma and T Fukuda ldquoOxygenomics inenvironmental stressrdquo Redox Report vol 15 no 3 pp 98ndash1142010
[3] K E Wellen and C B Thompson ldquoCellular metabolic stressconsidering how cells respond to nutrient excessrdquo MolecularCell vol 40 no 2 pp 323ndash332 2010
[4] R O Poyton K A Ball and P R Castello ldquoMitochondrialgeneration of free radicals and hypoxic signalingrdquo Trends inEndocrinology andMetabolism vol 20 no 7 pp 332ndash340 2009
[5] F Weinberg and N S Chandel ldquoReactive oxygen species-dependent signaling regulates cancerrdquo Cellular and MolecularLife Sciences vol 66 no 23 pp 3663ndash3673 2009
[6] A Cataldi ldquoCell responses to oxidative stressorsrdquo CurrentPharmaceutical Design vol 16 no 12 pp 1387ndash1395 2010
[7] Z Durackova ldquoSome current insights into oxidative stressrdquoPhysiological Research vol 59 no 4 pp 459ndash469 2010
[8] L A Sena and N S Chandel ldquoPhysiological roles of mitochon-drial reactive oxygen speciesrdquoMolecular Cell vol 48 no 2 pp158ndash166 2012
[9] D I Brown and K K Griendling ldquoNox proteins in signaltransductionrdquo Free Radical Biology amp Medicine vol 47 no 9pp 1239ndash1253 2009
[10] K Bedard and K-H Krause ldquoThe NOX family of ROS-generatingNADPHoxidases physiology and pathophysiologyrdquoPhysiological Reviews vol 87 no 1 pp 245ndash313 2007
[11] K-H Krause ldquoTissue distribution and putative physiologicalfunction of NOX family NADPH oxidasesrdquo Japanese Journal ofInfectious Diseases vol 57 no 5 pp S28ndashS29 2004
12 Mediators of Inflammation
[12] S Yang Y Zhang W Ries and L Key ldquoExpression of Nox4 inosteoclastsrdquo Journal of Cellular Biochemistry vol 92 no 2 pp238ndash248 2004
[13] E Pedruzzi C Guichard V Ollivier et al ldquoNAD(P)H oxidaseNox-4mediates 7-ketocholesterol-induced endoplasmic reticu-lum stress and apoptosis in human aortic smooth muscle cellsrdquoMolecular and Cellular Biology vol 24 no 24 pp 10703ndash107172004
[14] J Hwang A Saha Y C Boo et al ldquoOscillatory shear stress stim-ulates endothelial production of O
2-from p47119901ℎ119900119909-dependent
NAD(P)Hoxidases leading tomonocyte adhesionrdquoThe Journalof Biological Chemistry vol 278 no 47 pp 47291ndash47298 2003
[15] H B Suliman M Ali and C A Piantadosi ldquoSuperoxidedismutase-3 promotes full expression of the EPO response tohypoxiardquo Blood vol 104 no 1 pp 43ndash50 2004
[16] P Vallet Y Charnay K Steger et al ldquoNeuronal expressionof the NADPH oxidase NOX4 and its regulation in mouseexperimental brain ischemiardquo Neuroscience vol 132 no 2 pp233ndash238 2005
[17] K T Moe S Aulia F Jiang et al ldquoDifferential upregulation ofNox homologues of NADPH oxidase by tumor necrosis factor-120572 in human aortic smooth muscle and embryonic kidney cellsrdquoJournal of Cellular andMolecularMedicine vol 10 no 1 pp 231ndash239 2006
[18] I Diebold A Petry J Hess and A Gorlach ldquoThe NADPHoxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1rdquoMolecular Biology of the Cell vol 21 no 12pp 2087ndash2096 2010
[19] K V Tormos and N S Chandel ldquoInter-connection betweenmitochondria and HIFsrdquo Journal of Cellular and MolecularMedicine vol 14 no 4 pp 795ndash804 2010
[20] T Ohnishi K Bandow K Kakimoto M Machigashira TMatsuyama and T Matsuguchi ldquoOxidative stress causes alve-olar bone loss in metabolic syndrome model mice with type 2diabetesrdquo Journal of Periodontal Research vol 44 no 1 pp 43ndash51 2009
[21] C Goettsch A Babelova O Trummer et al ldquoNADPH oxidase4 limits bone mass by promoting osteoclastogenesisrdquo Journal ofClinical Investigation vol 123 no 11 pp 4731ndash4738 2013
[22] I L C Chapple and J B Matthews ldquoThe role of reactive oxygenand antioxidant species in periodontal tissue destructionrdquoPeriodontology 2000 vol 43 no 1 pp 160ndash232 2007
[23] A Guentsch P M Preshaw S Bremer-Streck G KlingerE Glockmann and B W Sigusch ldquoLipid peroxidation andantioxidant activity in saliva of periodontitis patients effect ofsmoking and periodontal treatmentrdquo Clinical Oral Investiga-tions vol 12 no 4 pp 345ndash352 2008
[24] R Agnihotri P Pandurang S U Kamath et al ldquoAssociation ofcigarette smoking with superoxide dismutase enzyme levels insubjected with chonic periodontitisrdquo Journal of Periodontologyvol 80 no 4 pp 657ndash662 2009
[25] D Ekuni T Tomofuji N Tamaki et al ldquoMechanical stimulationof gingiva reduces plasma 8-OHdG level in rat periodontitisrdquoArchives of Oral Biology vol 53 no 4 pp 324ndash329 2008
[26] H Staudte A Guntsch A Volpel and B W Sigusch ldquoVitaminC attenuates the cytotoxic effects of Porphyromonas gingivalison human gingival fibroblastsrdquo Archives of Oral Biology vol 55no 1 pp 40ndash45 2010
[27] P Bullon H N Newman and M Battino ldquoObesity diabetesmellitus atherosclerosis and chronic periodontitis a sharedpathology via oxidative stress andmitochondrial dysfunctionrdquoPeriodontology 2000 vol 64 no 1 pp 139ndash153 2014
[28] G R Naveh V Brumfeld R Shahar and S Weiner ldquoToothperiodontal ligament direct 3D microCT visualization of thecollagen network and how the network changes when the toothis loadedrdquo Journal of Structural Biology vol 181 no 2 pp 108ndash115 2013
[29] Y Kitase M Yokozeki S Fujihara et al ldquoAnalysis of geneexpression profiles in human periodontal ligament cells underhypoxia the protective effect of CC chemokine ligand 2 tooxygen shortagerdquoArchives of Oral Biology vol 54 no 7 pp 618ndash624 2009
[30] G Thornton-Evans P Eke L Wei et al ldquoPeriodontitis amongadults aged ge 30 years - United States 2009-2010rdquo Morbidityand Mortality Weekly Report (MMWR) vol 62 supplement 3pp 129ndash135 2013
[31] S S Socransky C Smith and A D Haffajee ldquoSubgingivalmicrobial profiles in refractory periodontal diseaserdquo Journal ofClinical Periodontology vol 29 no 3 pp 260ndash268 2002
[32] K-T Ng J-P Li K M Ng G L Tipoe W K Leung and M-L Fung ldquoExpression of hypoxia-inducible factor-1120572 in humanperiodontal tissuerdquo Journal of Periodontology vol 82 no 1 pp136ndash141 2011
[33] S Frede U Berchner-Pfannschmidt and J Fandrey ldquoReg-ulation of Hypoxia-inducible factors during inflammationrdquoMethods in Enzymology vol 435 pp 405ndash419 2007
[34] J Shin S Ji and Y Choi ldquoAbility of oral bacteria to inducetissue-destructive molecules from human neutrophilsrdquo OralDiseases vol 14 no 4 pp 327ndash334 2008
[35] S Takashiba K Naruishi and Y Murayama ldquoPerspectiveof cytokine regulation for periodontal treatment fibroblastbiologyrdquo Journal of Periodontology vol 74 no 1 pp 103ndash1102003
[36] A Konermann D Stabenow P A Knolle S A E HeldJ Deschner and A Jager ldquoRegulatory role of periodontalligament fibroblasts for innate immune cell function and dif-ferentiationrdquo Innate Immunity vol 18 no 5 pp 745ndash752 2012
[37] S Kar S Subbaram P M Carrico and J A Melendez ldquoRedox-control of matrix metalloproteinase-1 a critical link betweenfree radicals matrix remodeling and degenerative diseaserdquoRespiratory Physiology andNeurobiology vol 174 no 3 pp 299ndash306 2010
[38] F Rousset M V C Nguyen L Grange F Morel and B LardyldquoHeme oxygenase-1 regulates matrix metalloproteinase MMP-1secretion and chondrocyte cell death via Nox4NADPH oxidaseactivity in chondrocytesrdquo PLoS ONE vol 8 no 6 Article IDe66478 2013
[39] E Rudolf and M Cervinka ldquoResponses of human gingival andperiodontal fibroblasts to a low-zinc environmentrdquo Alternativesto Laboratory Animals vol 38 no 2 pp 119ndash138 2010
[40] S M San Miguel L A Opperman E P Allen J Zielinski andK K H Svoboda ldquoBioactive polyphenol antioxidants protectoral fibroblasts from ROS-inducing agentsrdquo Archives of OralBiology vol 57 no 12 pp 1657ndash1667 2012
[41] S M San Miguel L A Opperman E P Allen J E Zielinskiand K K H Svoboda ldquoAntioxidant combinations protect oralfibroblasts against metal-induced toxicityrdquo Archives of OralBiology vol 58 no 3 pp 299ndash310 2013
[42] U Skaleric C M Manthey S E Mergenhagen B Gaspircand S M Wahl ldquoSuperoxide release and superoxide dismutaseexpression by human gingival fibroblastsrdquo European Journal ofOral Sciences vol 108 no 2 pp 130ndash135 2000
Mediators of Inflammation 13
[43] D Y Kim J-H Jun H-L Lee et al ldquoN-acetylcysteine preventsLPS-Lnduced pro-inflammatory cytokines andMMP2 produc-tion in gingival fibroblastsrdquoArchives of Pharmacal Research vol30 no 10 pp 1283ndash1292 2007
[44] B Rath-Deschner J Deschner S Reimann A Jager andW Gotz ldquoRegulatory effects of biomechanical strain on theinsulin-like growth factor system in human periodontal cellsrdquoJournal of Biomechanics vol 42 no 15 pp 2584ndash2589 2009
[45] P Chomczynski and N Sacchi ldquoSingle-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextractionrdquo Analytical Biochemistry vol 162 no 1 pp 156ndash1591987
[46] B Zhao K Ranguelova J Jiang and R P Mason ldquoStudies onthe photosensitized reduction of resorufin and implications forthe detection of oxidative stress with Amplex Redrdquo Free RadicalBiology and Medicine vol 51 no 1 pp 153ndash159 2011
[47] C C Mandal S Ganapathy Y Gorin et al ldquoReactive oxygenspecies derived from Nox4 mediate BMP2 gene transcriptionand osteoblast differentiationrdquoBiochemical Journal vol 433 no2 pp 393ndash402 2011
[48] J Fandrey S Frede and W Jelkmann ldquoRole of hydrogenperoxide in hypoxia-induced erythropoietin productionrdquo Bio-chemical Journal vol 303 no 2 pp 507ndash510 1994
[49] F Cervellati C Cervellati A Romani et al ldquoHypoxia inducescell damage via oxidative stress in retinal epithelial cellsrdquo FreeRadical Research vol 48 no 3 pp 303ndash312 2014
[50] V Vanasco N D Magnani M C Cimolai et al ldquoEndotoxemiaimpairs heart mitochondrial function by decreasing electrontransfer ATP synthesis and ATP content without affectingmembrane potentialrdquo Journal of Bioenergetics and Biomem-branes vol 44 no 2 pp 243ndash252 2012
[51] R G Alscher N Erturk and L S Heath ldquoRole of superoxidedismutases (SODs) in controlling oxidative stress in plantsrdquoJournal of Experimental Botany vol 53 no 372 pp 1331ndash13412002
[52] E Baltacıoglu P Yuva G Aydın et al ldquoLipid peroxidation levelsand total oxidantantioxidant status in serum and saliva frompatients with chronic and aggressive periodontitis Oxidativestress index a new biomarker for periodontal diseaserdquo Journalof Periodontology vol 85 no 10 pp 1432ndash1441 2014
[53] I L C Chapple M R Milward N Ling-Mountford et alldquoAdjunctive daily supplementation with encapsulated fruitvegetable and berry juice powder concentrates and clinicalperiodontal outcomes a double-blind RCTrdquo Journal of ClinicalPeriodontology vol 39 no 1 pp 62ndash72 2012
[54] A Jenzsch S Eick F Rassoul R Purschwitz and H JentschldquoNutritional intervention in patients with periodontal diseaseClinical immunological and microbiological variables during12 monthsrdquo British Journal of Nutrition vol 101 no 6 pp 879ndash885 2009
[55] N Novakovic T Todorovic M Rakic et al ldquoSalivary antiox-idants as periodontal biomarkers in evaluation of tissue statusand treatment outcomerdquo Journal of Periodontal Research vol49 no 1 pp 129ndash136 2014
[56] N Singh S C Narula R K Sharma S Tewari and P KSehgal ldquoVitamin e supplementation superoxide dismutasestatus and outcome of scaling and root planing in patients withchronic periodontitis a randomized clinical trialrdquo Journal ofPeriodontology vol 85 no 2 pp 242ndash249 2014
[57] S Krifka K-A Hiller G Spagnuolo A Jewett G Schmalz andH Schweikl ldquoThe influence of glutathione on redox regulation
by antioxidant proteins and apoptosis in macrophages exposedto 2-hydroxyethyl methacrylate (HEMA)rdquo Biomaterials vol 33no 21 pp 5177ndash5186 2012
[58] I Borges Jr E A M Moreira D W Filho T B de Oliveira MB S da Silva and T S Frode ldquoProinflammatory and oxidativestress markers in patients with periodontal diseaserdquo Mediatorsof Inflammation vol 2007 Article ID 45794 5 pages 2007
[59] M O Tonguc O Ozturk R Sutcu et al ldquoThe impact of smok-ing status on antioxidant enzyme activity and malondialdehydelevels in chronic periodontitisrdquo Journal of Periodontology vol82 no 9 pp 1320ndash1328 2011
[60] B H Jacoby and W L Davis ldquoThe electron microscopicimmunolocalization of a copper-zinc superoxide dismutase inassociation with collagen fibers of periodontal soft tissuesrdquoJournal of Periodontology vol 62 no 7 pp 413ndash420 1991
[61] T Konopka K Krol W Kopec and H Gerber ldquoTotal antioxi-dant status and 8-hydroxy-21015840-deoxyguanosine levels in gingi-val and peripheral blood of periodontitis patientsrdquo ArchivumImmunologiae et Therapiae Experimentalis vol 55 no 6 pp417ndash422 2007
[62] M Soory ldquoRedox status in periodontal and systemic inflamma-tory conditions including associated neoplasias antioxidants asadjunctive therapyrdquo Infectious DisordersmdashDrug Targets vol 9no 4 pp 415ndash427 2009
[63] M Soory ldquoOxidative stress inducedmechanisms in the progres-sion of periodontal diseases and cancer a common approach toredox homeostasisrdquo Cancers vol 2 no 2 pp 670ndash692 2010
[64] R V Chandra G Srinivas A A Reddy et al ldquoLocally deliveredantioxidant gel as an adjunct to nonsurgical therapy improvesmeasures of oxidative stress and periodontal diseaserdquo Journal ofPeriodontal and Implant Science vol 43 no 3 pp 121ndash129 2013
[65] M A D S Siqueira R G Fischer N R Pereira et al ldquoEffectsof non-surgical periodontal treatment on the L-arginine-nitricoxide pathway and oxidative status in plateletsrdquo ExperimentalBiology and Medicine vol 238 no 6 pp 713ndash722 2013
[66] P Mydel Y Takahashi H Yumoto et al ldquoRoles of the hostoxidative immune response and bacterial antioxidant rubr-erythrin during Porphyromonas gingivalis infectionrdquo PLoSpathogens vol 2 no 7 p e76 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
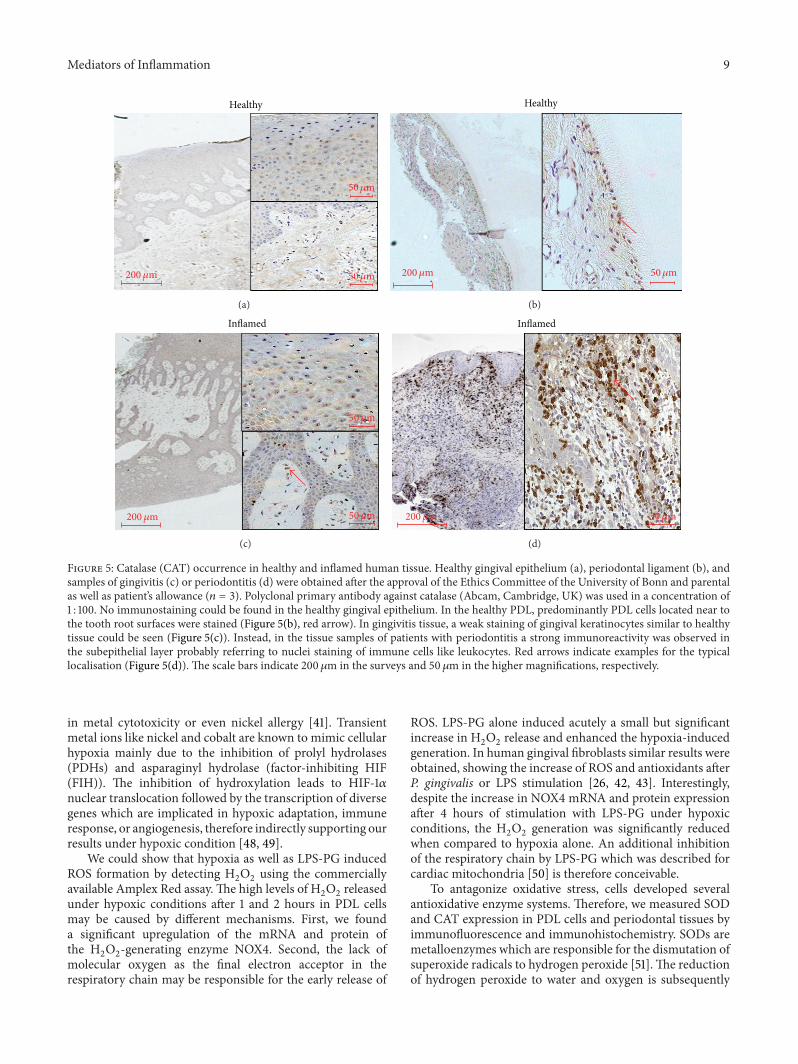
Mediators of Inflammation 9
200120583m
50120583m
50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m
50120583m
50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 5 Catalase (CAT) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontal ligament (b) andsamples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University of Bonn and parentalas well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against catalase (Abcam Cambridge UK) was used in a concentration of1 100 No immunostaining could be found in the healthy gingival epithelium In the healthy PDL predominantly PDL cells located near tothe tooth root surfaces were stained (Figure 5(b) red arrow) In gingivitis tissue a weak staining of gingival keratinocytes similar to healthytissue could be seen (Figure 5(c)) Instead in the tissue samples of patients with periodontitis a strong immunoreactivity was observed inthe subepithelial layer probably referring to nuclei staining of immune cells like leukocytes Red arrows indicate examples for the typicallocalisation (Figure 5(d)) The scale bars indicate 200 120583m in the surveys and 50120583m in the higher magnifications respectively
in metal cytotoxicity or even nickel allergy [41] Transientmetal ions like nickel and cobalt are known to mimic cellularhypoxia mainly due to the inhibition of prolyl hydrolases(PDHs) and asparaginyl hydrolase (factor-inhibiting HIF(FIH)) The inhibition of hydroxylation leads to HIF-1120572nuclear translocation followed by the transcription of diversegenes which are implicated in hypoxic adaptation immuneresponse or angiogenesis therefore indirectly supporting ourresults under hypoxic condition [48 49]
We could show that hypoxia as well as LPS-PG inducedROS formation by detecting H
2O2using the commercially
available Amplex Red assay The high levels of H2O2released
under hypoxic conditions after 1 and 2 hours in PDL cellsmay be caused by different mechanisms First we founda significant upregulation of the mRNA and protein ofthe H
2O2-generating enzyme NOX4 Second the lack of
molecular oxygen as the final electron acceptor in therespiratory chain may be responsible for the early release of
ROS LPS-PG alone induced acutely a small but significantincrease in H
2O2release and enhanced the hypoxia-induced
generation In human gingival fibroblasts similar results wereobtained showing the increase of ROS and antioxidants afterP gingivalis or LPS stimulation [26 42 43] Interestinglydespite the increase in NOX4 mRNA and protein expressionafter 4 hours of stimulation with LPS-PG under hypoxicconditions the H
2O2generation was significantly reduced
when compared to hypoxia alone An additional inhibitionof the respiratory chain by LPS-PG which was described forcardiac mitochondria [50] is therefore conceivable
To antagonize oxidative stress cells developed severalantioxidative enzyme systems Therefore we measured SODand CAT expression in PDL cells and periodontal tissues byimmunofluorescence and immunohistochemistry SODs aremetalloenzymes which are responsible for the dismutation ofsuperoxide radicals to hydrogen peroxide [51] The reductionof hydrogen peroxide to water and oxygen is subsequently
10 Mediators of Inflammation
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m 50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 6 Superoxide dismutase (SOD) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against SOD (Abcam Cambridge UK) was used ina concentration of 1 100 Gingival epithelium showed a moderate to strong staining of SOD in keratinocytes which was primarily limitedto the cytosol (Figure 6(a) and red arrow depicts the typical localization) In the healthy periodontium gingival and PDL fibroblasts wereweakly stained (Figure 6(b)) In inflamed tissues basal epithelial cells seemed to be stained more intensively in the gingivitis specimen thanin periodontitis (Figures 6(c) and 6(d)) In addition the subepithelium revealed immunoreactivity in local cells like gingival fibroblasts andscattered immune cells (Figure 6(c) see red arrow) In periodontitis SOD immunostaining seemed to be limited to the gingival epithelium(Figure 6(d) see red arrow) The scale bars indicate 200 120583m in the surveys and 50 120583m in the higher magnifications respectively
catalyzed by CATs which are ubiquitously expressed Bothenzymes are essential to maintain cellular redox homeostasisThe present data imply that PDL cells and periodontal tissueshave fundamental defense machineries against bacteria rep-resented by NOX4-dependent inducible ROS generation aswell as protective redox systems This is of great importanceconcerning the high amounts and diversity of the oral floraDifferent studies indicate that redox systems are key playersin the development of periodontal diseases and markers forperiodontal health status [52ndash56] We detected a significantincrease of catalase in PDL cells in response to LPS aswell as hypoxia after 1 and 2 hours In our setting PDLfibroblasts seem to counteracted LPS- and hypoxia-inducedROS formation with the elevation of CAT expression Butprolonged incubation under hypoxic conditions failed toincrease catalase Since ROS generation was still elevated atthese later time points a delay in ROS degradation may be
the explanation Taken together the dramatic elevation inROS formation by the combination of hypoxia and LPS-PGon one hand and the reduction of catalase under the sameconditions on the other hand increase the oxidative stress inthe periodontium favoring the progression of inflammationand bone resorption and finally may lead to tooth loss
The fundamental role of the balance between ROSgeneration and antioxidative mechanisms in periodontalinflammatory processes was supported by our immunohis-tochemical findings In patients suffering from periodontalinflammatory diseases like gingivitis and periodontitis wedemonstrated that NOX4 is upregulated and thus likelyresponsible for H
2O2generation However we detected as
well an increase in catalase abundance in inflamed gingivaland periodontal tissue which was in line with data fromKrifka and colleagues who detected high CAT levels inmonomer-exposed macrophages as a response to oxidative
Mediators of Inflammation 11
stress [57] In our in vitro setting PDL fibroblasts seemto counteract LPS- and hypoxia-induced ROS formationpredominantly with an elevation of CAT expression sincewe could not detect any SOD immunoreactivity The roleand impact of the antioxidative systems SOD and CAT arecontroversially discussed Recent studies measured CAT andSOD activities in gingival tissues of patients with periodon-titis [58 59] Borges Jr et al did not find differences inCAT activities comparing healthy and diseased patients [58]Instead Tonguc et al detected decreased local SOD andCAT activities in periodontitis patients which seemed to beincreased by smoking [59] In contrast in inflamed tissues weobservedhigh amounts of SODwhichwas localized primarilyin epithelial cells Jacoby and Davis determined SOD in peri-odontal soft tissues They detected twice as much SOD activ-ity in the periodontal ligament as in the human skin (dermis)[60] Furthermore immunohistochemical analysis revealedthat SOD is localized in the periphery of matrix collagenfibrils and to the glycocalyx of tissue fibroblasts This wasconfirmed by Skaleric et al [42] which might explain the factthat we did not detect significant differences of SOD levelsafter hypoxic and LPS-PG stimulation intracellularly in PDLfibroblasts Another reason for these results may be causedby the age of the PDL cells used in this study We explantedthe PDL cells from young and oral healthy patients but mostof the immunohistochemical stainings were performed intissues from elderly patients with clearly diagnosed gingivitisor periodontitisTherefore it is possible that in PDL cells fromelderly patients a different expression pattern of antioxidantenzyme systems could be found which further deterioratethe redox state of the periodontal ligament Moreover otherauthors found reduced antioxidants in the body fluids ofpatients with periodontitis indicating a dysregulation whichwould favor disease progression [23 24 61] A variety of invitro and animal studies analyzed the effect of antioxidantsupplement in the context of inflammatory or cytotoxicevents [57 62 63] They have shown that antioxidantsinhibit these processes indicating a therapeutic approachin pathologies like periodontitis First clinical investigationsconfirmed the efficiency of antioxidant micronutrients andadjuncts in periodontal diseases [22 54 56 64 65] Thepresent data indirectly support these studies
Concerning periodontal pathogen invasiveness and viru-lence there is compelling evidence that P gingivalis possessesits own protective antioxidants for example rubrerythrindefending them against the oxidative burst of the host [66]We have shown that its virulence factor LPS-PG inducedhigh amounts of ROS via NOX4 which may result in boneresorption favoring the invasiveness of the pathogen whichis at the same time protected by its own defense machineryThis would further explain the pathogenicity and resistanceof this bacterium strain in patients with periodontitis
5 Conclusion
In this study we have demonstrated that LPS from Porphy-romonas gingivalis (LPS-PG) as well as hypoxia induces aNOX4-dependent increase inH
2O2release in PDLfibroblasts
which may contribute to the development and progressionof periodontal diseases when not balanced by a concomitantincrease in antioxidative systems The dramatic elevationin ROS formation induced by the combination of hypoxiaand LPS-PG and the reduction of catalase under the sameconditions increased the oxidative stress in the periodontiumfavoring the progression of inflammation bone resorptionand possibly tooth loss
Taken together the presented data further explain thedestructive mechanism of hypoxic and inflammatory condi-tions in the periodontium and may improve diagnostic andtherapeutic strategies in periodontal diseases
Conflict of Interests
The authors have no conflict of interests to disclose
Acknowledgments
The authors would like to thank Ms Tina Schaffrath for hertechnical supportThe present studywas supported by a grantfrom the German Research Foundation (Clinical ResearchUnit 208TP7) and the Medical Faculty of the University ofBonn
References
[1] S Reuter S C Gupta M M Chaturvedi and B B AggarwalldquoOxidative stress inflammation and cancer how are theylinkedrdquo Free Radical Biology and Medicine vol 49 no 11 pp1603ndash1616 2010
[2] H Sone H Akanuma and T Fukuda ldquoOxygenomics inenvironmental stressrdquo Redox Report vol 15 no 3 pp 98ndash1142010
[3] K E Wellen and C B Thompson ldquoCellular metabolic stressconsidering how cells respond to nutrient excessrdquo MolecularCell vol 40 no 2 pp 323ndash332 2010
[4] R O Poyton K A Ball and P R Castello ldquoMitochondrialgeneration of free radicals and hypoxic signalingrdquo Trends inEndocrinology andMetabolism vol 20 no 7 pp 332ndash340 2009
[5] F Weinberg and N S Chandel ldquoReactive oxygen species-dependent signaling regulates cancerrdquo Cellular and MolecularLife Sciences vol 66 no 23 pp 3663ndash3673 2009
[6] A Cataldi ldquoCell responses to oxidative stressorsrdquo CurrentPharmaceutical Design vol 16 no 12 pp 1387ndash1395 2010
[7] Z Durackova ldquoSome current insights into oxidative stressrdquoPhysiological Research vol 59 no 4 pp 459ndash469 2010
[8] L A Sena and N S Chandel ldquoPhysiological roles of mitochon-drial reactive oxygen speciesrdquoMolecular Cell vol 48 no 2 pp158ndash166 2012
[9] D I Brown and K K Griendling ldquoNox proteins in signaltransductionrdquo Free Radical Biology amp Medicine vol 47 no 9pp 1239ndash1253 2009
[10] K Bedard and K-H Krause ldquoThe NOX family of ROS-generatingNADPHoxidases physiology and pathophysiologyrdquoPhysiological Reviews vol 87 no 1 pp 245ndash313 2007
[11] K-H Krause ldquoTissue distribution and putative physiologicalfunction of NOX family NADPH oxidasesrdquo Japanese Journal ofInfectious Diseases vol 57 no 5 pp S28ndashS29 2004
12 Mediators of Inflammation
[12] S Yang Y Zhang W Ries and L Key ldquoExpression of Nox4 inosteoclastsrdquo Journal of Cellular Biochemistry vol 92 no 2 pp238ndash248 2004
[13] E Pedruzzi C Guichard V Ollivier et al ldquoNAD(P)H oxidaseNox-4mediates 7-ketocholesterol-induced endoplasmic reticu-lum stress and apoptosis in human aortic smooth muscle cellsrdquoMolecular and Cellular Biology vol 24 no 24 pp 10703ndash107172004
[14] J Hwang A Saha Y C Boo et al ldquoOscillatory shear stress stim-ulates endothelial production of O
2-from p47119901ℎ119900119909-dependent
NAD(P)Hoxidases leading tomonocyte adhesionrdquoThe Journalof Biological Chemistry vol 278 no 47 pp 47291ndash47298 2003
[15] H B Suliman M Ali and C A Piantadosi ldquoSuperoxidedismutase-3 promotes full expression of the EPO response tohypoxiardquo Blood vol 104 no 1 pp 43ndash50 2004
[16] P Vallet Y Charnay K Steger et al ldquoNeuronal expressionof the NADPH oxidase NOX4 and its regulation in mouseexperimental brain ischemiardquo Neuroscience vol 132 no 2 pp233ndash238 2005
[17] K T Moe S Aulia F Jiang et al ldquoDifferential upregulation ofNox homologues of NADPH oxidase by tumor necrosis factor-120572 in human aortic smooth muscle and embryonic kidney cellsrdquoJournal of Cellular andMolecularMedicine vol 10 no 1 pp 231ndash239 2006
[18] I Diebold A Petry J Hess and A Gorlach ldquoThe NADPHoxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1rdquoMolecular Biology of the Cell vol 21 no 12pp 2087ndash2096 2010
[19] K V Tormos and N S Chandel ldquoInter-connection betweenmitochondria and HIFsrdquo Journal of Cellular and MolecularMedicine vol 14 no 4 pp 795ndash804 2010
[20] T Ohnishi K Bandow K Kakimoto M Machigashira TMatsuyama and T Matsuguchi ldquoOxidative stress causes alve-olar bone loss in metabolic syndrome model mice with type 2diabetesrdquo Journal of Periodontal Research vol 44 no 1 pp 43ndash51 2009
[21] C Goettsch A Babelova O Trummer et al ldquoNADPH oxidase4 limits bone mass by promoting osteoclastogenesisrdquo Journal ofClinical Investigation vol 123 no 11 pp 4731ndash4738 2013
[22] I L C Chapple and J B Matthews ldquoThe role of reactive oxygenand antioxidant species in periodontal tissue destructionrdquoPeriodontology 2000 vol 43 no 1 pp 160ndash232 2007
[23] A Guentsch P M Preshaw S Bremer-Streck G KlingerE Glockmann and B W Sigusch ldquoLipid peroxidation andantioxidant activity in saliva of periodontitis patients effect ofsmoking and periodontal treatmentrdquo Clinical Oral Investiga-tions vol 12 no 4 pp 345ndash352 2008
[24] R Agnihotri P Pandurang S U Kamath et al ldquoAssociation ofcigarette smoking with superoxide dismutase enzyme levels insubjected with chonic periodontitisrdquo Journal of Periodontologyvol 80 no 4 pp 657ndash662 2009
[25] D Ekuni T Tomofuji N Tamaki et al ldquoMechanical stimulationof gingiva reduces plasma 8-OHdG level in rat periodontitisrdquoArchives of Oral Biology vol 53 no 4 pp 324ndash329 2008
[26] H Staudte A Guntsch A Volpel and B W Sigusch ldquoVitaminC attenuates the cytotoxic effects of Porphyromonas gingivalison human gingival fibroblastsrdquo Archives of Oral Biology vol 55no 1 pp 40ndash45 2010
[27] P Bullon H N Newman and M Battino ldquoObesity diabetesmellitus atherosclerosis and chronic periodontitis a sharedpathology via oxidative stress andmitochondrial dysfunctionrdquoPeriodontology 2000 vol 64 no 1 pp 139ndash153 2014
[28] G R Naveh V Brumfeld R Shahar and S Weiner ldquoToothperiodontal ligament direct 3D microCT visualization of thecollagen network and how the network changes when the toothis loadedrdquo Journal of Structural Biology vol 181 no 2 pp 108ndash115 2013
[29] Y Kitase M Yokozeki S Fujihara et al ldquoAnalysis of geneexpression profiles in human periodontal ligament cells underhypoxia the protective effect of CC chemokine ligand 2 tooxygen shortagerdquoArchives of Oral Biology vol 54 no 7 pp 618ndash624 2009
[30] G Thornton-Evans P Eke L Wei et al ldquoPeriodontitis amongadults aged ge 30 years - United States 2009-2010rdquo Morbidityand Mortality Weekly Report (MMWR) vol 62 supplement 3pp 129ndash135 2013
[31] S S Socransky C Smith and A D Haffajee ldquoSubgingivalmicrobial profiles in refractory periodontal diseaserdquo Journal ofClinical Periodontology vol 29 no 3 pp 260ndash268 2002
[32] K-T Ng J-P Li K M Ng G L Tipoe W K Leung and M-L Fung ldquoExpression of hypoxia-inducible factor-1120572 in humanperiodontal tissuerdquo Journal of Periodontology vol 82 no 1 pp136ndash141 2011
[33] S Frede U Berchner-Pfannschmidt and J Fandrey ldquoReg-ulation of Hypoxia-inducible factors during inflammationrdquoMethods in Enzymology vol 435 pp 405ndash419 2007
[34] J Shin S Ji and Y Choi ldquoAbility of oral bacteria to inducetissue-destructive molecules from human neutrophilsrdquo OralDiseases vol 14 no 4 pp 327ndash334 2008
[35] S Takashiba K Naruishi and Y Murayama ldquoPerspectiveof cytokine regulation for periodontal treatment fibroblastbiologyrdquo Journal of Periodontology vol 74 no 1 pp 103ndash1102003
[36] A Konermann D Stabenow P A Knolle S A E HeldJ Deschner and A Jager ldquoRegulatory role of periodontalligament fibroblasts for innate immune cell function and dif-ferentiationrdquo Innate Immunity vol 18 no 5 pp 745ndash752 2012
[37] S Kar S Subbaram P M Carrico and J A Melendez ldquoRedox-control of matrix metalloproteinase-1 a critical link betweenfree radicals matrix remodeling and degenerative diseaserdquoRespiratory Physiology andNeurobiology vol 174 no 3 pp 299ndash306 2010
[38] F Rousset M V C Nguyen L Grange F Morel and B LardyldquoHeme oxygenase-1 regulates matrix metalloproteinase MMP-1secretion and chondrocyte cell death via Nox4NADPH oxidaseactivity in chondrocytesrdquo PLoS ONE vol 8 no 6 Article IDe66478 2013
[39] E Rudolf and M Cervinka ldquoResponses of human gingival andperiodontal fibroblasts to a low-zinc environmentrdquo Alternativesto Laboratory Animals vol 38 no 2 pp 119ndash138 2010
[40] S M San Miguel L A Opperman E P Allen J Zielinski andK K H Svoboda ldquoBioactive polyphenol antioxidants protectoral fibroblasts from ROS-inducing agentsrdquo Archives of OralBiology vol 57 no 12 pp 1657ndash1667 2012
[41] S M San Miguel L A Opperman E P Allen J E Zielinskiand K K H Svoboda ldquoAntioxidant combinations protect oralfibroblasts against metal-induced toxicityrdquo Archives of OralBiology vol 58 no 3 pp 299ndash310 2013
[42] U Skaleric C M Manthey S E Mergenhagen B Gaspircand S M Wahl ldquoSuperoxide release and superoxide dismutaseexpression by human gingival fibroblastsrdquo European Journal ofOral Sciences vol 108 no 2 pp 130ndash135 2000
Mediators of Inflammation 13
[43] D Y Kim J-H Jun H-L Lee et al ldquoN-acetylcysteine preventsLPS-Lnduced pro-inflammatory cytokines andMMP2 produc-tion in gingival fibroblastsrdquoArchives of Pharmacal Research vol30 no 10 pp 1283ndash1292 2007
[44] B Rath-Deschner J Deschner S Reimann A Jager andW Gotz ldquoRegulatory effects of biomechanical strain on theinsulin-like growth factor system in human periodontal cellsrdquoJournal of Biomechanics vol 42 no 15 pp 2584ndash2589 2009
[45] P Chomczynski and N Sacchi ldquoSingle-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextractionrdquo Analytical Biochemistry vol 162 no 1 pp 156ndash1591987
[46] B Zhao K Ranguelova J Jiang and R P Mason ldquoStudies onthe photosensitized reduction of resorufin and implications forthe detection of oxidative stress with Amplex Redrdquo Free RadicalBiology and Medicine vol 51 no 1 pp 153ndash159 2011
[47] C C Mandal S Ganapathy Y Gorin et al ldquoReactive oxygenspecies derived from Nox4 mediate BMP2 gene transcriptionand osteoblast differentiationrdquoBiochemical Journal vol 433 no2 pp 393ndash402 2011
[48] J Fandrey S Frede and W Jelkmann ldquoRole of hydrogenperoxide in hypoxia-induced erythropoietin productionrdquo Bio-chemical Journal vol 303 no 2 pp 507ndash510 1994
[49] F Cervellati C Cervellati A Romani et al ldquoHypoxia inducescell damage via oxidative stress in retinal epithelial cellsrdquo FreeRadical Research vol 48 no 3 pp 303ndash312 2014
[50] V Vanasco N D Magnani M C Cimolai et al ldquoEndotoxemiaimpairs heart mitochondrial function by decreasing electrontransfer ATP synthesis and ATP content without affectingmembrane potentialrdquo Journal of Bioenergetics and Biomem-branes vol 44 no 2 pp 243ndash252 2012
[51] R G Alscher N Erturk and L S Heath ldquoRole of superoxidedismutases (SODs) in controlling oxidative stress in plantsrdquoJournal of Experimental Botany vol 53 no 372 pp 1331ndash13412002
[52] E Baltacıoglu P Yuva G Aydın et al ldquoLipid peroxidation levelsand total oxidantantioxidant status in serum and saliva frompatients with chronic and aggressive periodontitis Oxidativestress index a new biomarker for periodontal diseaserdquo Journalof Periodontology vol 85 no 10 pp 1432ndash1441 2014
[53] I L C Chapple M R Milward N Ling-Mountford et alldquoAdjunctive daily supplementation with encapsulated fruitvegetable and berry juice powder concentrates and clinicalperiodontal outcomes a double-blind RCTrdquo Journal of ClinicalPeriodontology vol 39 no 1 pp 62ndash72 2012
[54] A Jenzsch S Eick F Rassoul R Purschwitz and H JentschldquoNutritional intervention in patients with periodontal diseaseClinical immunological and microbiological variables during12 monthsrdquo British Journal of Nutrition vol 101 no 6 pp 879ndash885 2009
[55] N Novakovic T Todorovic M Rakic et al ldquoSalivary antiox-idants as periodontal biomarkers in evaluation of tissue statusand treatment outcomerdquo Journal of Periodontal Research vol49 no 1 pp 129ndash136 2014
[56] N Singh S C Narula R K Sharma S Tewari and P KSehgal ldquoVitamin e supplementation superoxide dismutasestatus and outcome of scaling and root planing in patients withchronic periodontitis a randomized clinical trialrdquo Journal ofPeriodontology vol 85 no 2 pp 242ndash249 2014
[57] S Krifka K-A Hiller G Spagnuolo A Jewett G Schmalz andH Schweikl ldquoThe influence of glutathione on redox regulation
by antioxidant proteins and apoptosis in macrophages exposedto 2-hydroxyethyl methacrylate (HEMA)rdquo Biomaterials vol 33no 21 pp 5177ndash5186 2012
[58] I Borges Jr E A M Moreira D W Filho T B de Oliveira MB S da Silva and T S Frode ldquoProinflammatory and oxidativestress markers in patients with periodontal diseaserdquo Mediatorsof Inflammation vol 2007 Article ID 45794 5 pages 2007
[59] M O Tonguc O Ozturk R Sutcu et al ldquoThe impact of smok-ing status on antioxidant enzyme activity and malondialdehydelevels in chronic periodontitisrdquo Journal of Periodontology vol82 no 9 pp 1320ndash1328 2011
[60] B H Jacoby and W L Davis ldquoThe electron microscopicimmunolocalization of a copper-zinc superoxide dismutase inassociation with collagen fibers of periodontal soft tissuesrdquoJournal of Periodontology vol 62 no 7 pp 413ndash420 1991
[61] T Konopka K Krol W Kopec and H Gerber ldquoTotal antioxi-dant status and 8-hydroxy-21015840-deoxyguanosine levels in gingi-val and peripheral blood of periodontitis patientsrdquo ArchivumImmunologiae et Therapiae Experimentalis vol 55 no 6 pp417ndash422 2007
[62] M Soory ldquoRedox status in periodontal and systemic inflamma-tory conditions including associated neoplasias antioxidants asadjunctive therapyrdquo Infectious DisordersmdashDrug Targets vol 9no 4 pp 415ndash427 2009
[63] M Soory ldquoOxidative stress inducedmechanisms in the progres-sion of periodontal diseases and cancer a common approach toredox homeostasisrdquo Cancers vol 2 no 2 pp 670ndash692 2010
[64] R V Chandra G Srinivas A A Reddy et al ldquoLocally deliveredantioxidant gel as an adjunct to nonsurgical therapy improvesmeasures of oxidative stress and periodontal diseaserdquo Journal ofPeriodontal and Implant Science vol 43 no 3 pp 121ndash129 2013
[65] M A D S Siqueira R G Fischer N R Pereira et al ldquoEffectsof non-surgical periodontal treatment on the L-arginine-nitricoxide pathway and oxidative status in plateletsrdquo ExperimentalBiology and Medicine vol 238 no 6 pp 713ndash722 2013
[66] P Mydel Y Takahashi H Yumoto et al ldquoRoles of the hostoxidative immune response and bacterial antioxidant rubr-erythrin during Porphyromonas gingivalis infectionrdquo PLoSpathogens vol 2 no 7 p e76 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
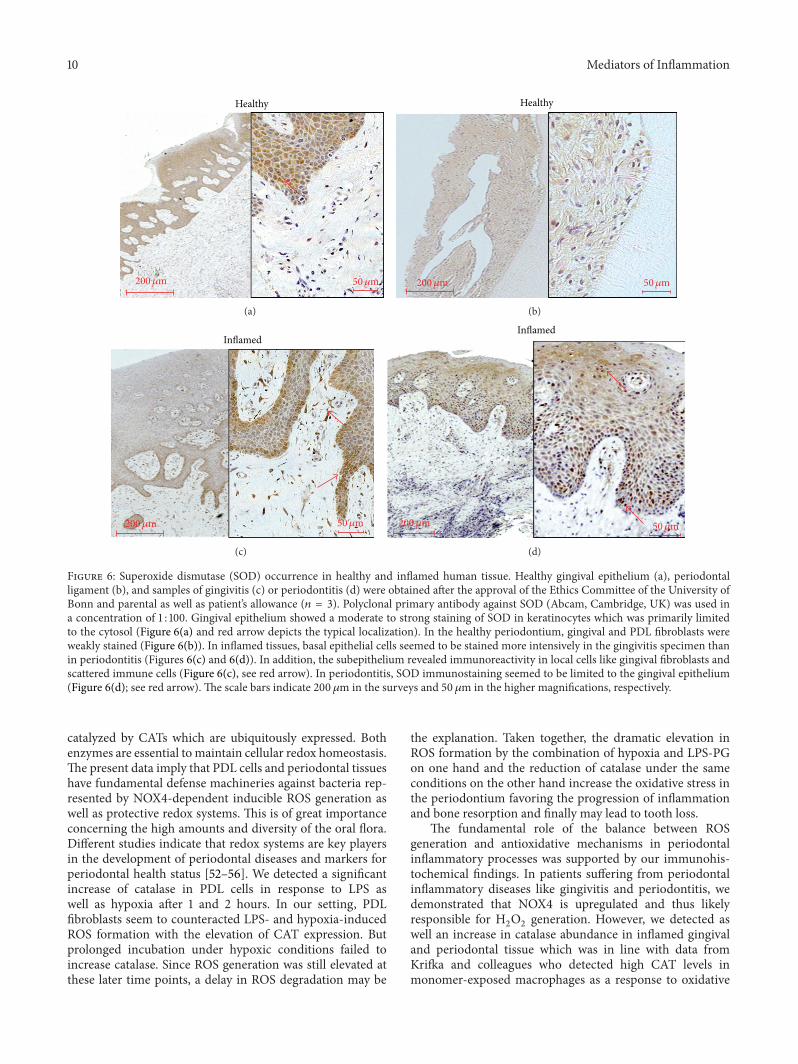
10 Mediators of Inflammation
200120583m 50120583m
Healthy
(a)
200120583m 50120583m
Healthy
(b)
200120583m 50120583m
Inflamed
(c)
200120583m 50120583m
Inflamed
(d)
Figure 6 Superoxide dismutase (SOD) occurrence in healthy and inflamed human tissue Healthy gingival epithelium (a) periodontalligament (b) and samples of gingivitis (c) or periodontitis (d) were obtained after the approval of the Ethics Committee of the University ofBonn and parental as well as patientrsquos allowance (119899 = 3) Polyclonal primary antibody against SOD (Abcam Cambridge UK) was used ina concentration of 1 100 Gingival epithelium showed a moderate to strong staining of SOD in keratinocytes which was primarily limitedto the cytosol (Figure 6(a) and red arrow depicts the typical localization) In the healthy periodontium gingival and PDL fibroblasts wereweakly stained (Figure 6(b)) In inflamed tissues basal epithelial cells seemed to be stained more intensively in the gingivitis specimen thanin periodontitis (Figures 6(c) and 6(d)) In addition the subepithelium revealed immunoreactivity in local cells like gingival fibroblasts andscattered immune cells (Figure 6(c) see red arrow) In periodontitis SOD immunostaining seemed to be limited to the gingival epithelium(Figure 6(d) see red arrow) The scale bars indicate 200 120583m in the surveys and 50 120583m in the higher magnifications respectively
catalyzed by CATs which are ubiquitously expressed Bothenzymes are essential to maintain cellular redox homeostasisThe present data imply that PDL cells and periodontal tissueshave fundamental defense machineries against bacteria rep-resented by NOX4-dependent inducible ROS generation aswell as protective redox systems This is of great importanceconcerning the high amounts and diversity of the oral floraDifferent studies indicate that redox systems are key playersin the development of periodontal diseases and markers forperiodontal health status [52ndash56] We detected a significantincrease of catalase in PDL cells in response to LPS aswell as hypoxia after 1 and 2 hours In our setting PDLfibroblasts seem to counteracted LPS- and hypoxia-inducedROS formation with the elevation of CAT expression Butprolonged incubation under hypoxic conditions failed toincrease catalase Since ROS generation was still elevated atthese later time points a delay in ROS degradation may be
the explanation Taken together the dramatic elevation inROS formation by the combination of hypoxia and LPS-PGon one hand and the reduction of catalase under the sameconditions on the other hand increase the oxidative stress inthe periodontium favoring the progression of inflammationand bone resorption and finally may lead to tooth loss
The fundamental role of the balance between ROSgeneration and antioxidative mechanisms in periodontalinflammatory processes was supported by our immunohis-tochemical findings In patients suffering from periodontalinflammatory diseases like gingivitis and periodontitis wedemonstrated that NOX4 is upregulated and thus likelyresponsible for H
2O2generation However we detected as
well an increase in catalase abundance in inflamed gingivaland periodontal tissue which was in line with data fromKrifka and colleagues who detected high CAT levels inmonomer-exposed macrophages as a response to oxidative
Mediators of Inflammation 11
stress [57] In our in vitro setting PDL fibroblasts seemto counteract LPS- and hypoxia-induced ROS formationpredominantly with an elevation of CAT expression sincewe could not detect any SOD immunoreactivity The roleand impact of the antioxidative systems SOD and CAT arecontroversially discussed Recent studies measured CAT andSOD activities in gingival tissues of patients with periodon-titis [58 59] Borges Jr et al did not find differences inCAT activities comparing healthy and diseased patients [58]Instead Tonguc et al detected decreased local SOD andCAT activities in periodontitis patients which seemed to beincreased by smoking [59] In contrast in inflamed tissues weobservedhigh amounts of SODwhichwas localized primarilyin epithelial cells Jacoby and Davis determined SOD in peri-odontal soft tissues They detected twice as much SOD activ-ity in the periodontal ligament as in the human skin (dermis)[60] Furthermore immunohistochemical analysis revealedthat SOD is localized in the periphery of matrix collagenfibrils and to the glycocalyx of tissue fibroblasts This wasconfirmed by Skaleric et al [42] which might explain the factthat we did not detect significant differences of SOD levelsafter hypoxic and LPS-PG stimulation intracellularly in PDLfibroblasts Another reason for these results may be causedby the age of the PDL cells used in this study We explantedthe PDL cells from young and oral healthy patients but mostof the immunohistochemical stainings were performed intissues from elderly patients with clearly diagnosed gingivitisor periodontitisTherefore it is possible that in PDL cells fromelderly patients a different expression pattern of antioxidantenzyme systems could be found which further deterioratethe redox state of the periodontal ligament Moreover otherauthors found reduced antioxidants in the body fluids ofpatients with periodontitis indicating a dysregulation whichwould favor disease progression [23 24 61] A variety of invitro and animal studies analyzed the effect of antioxidantsupplement in the context of inflammatory or cytotoxicevents [57 62 63] They have shown that antioxidantsinhibit these processes indicating a therapeutic approachin pathologies like periodontitis First clinical investigationsconfirmed the efficiency of antioxidant micronutrients andadjuncts in periodontal diseases [22 54 56 64 65] Thepresent data indirectly support these studies
Concerning periodontal pathogen invasiveness and viru-lence there is compelling evidence that P gingivalis possessesits own protective antioxidants for example rubrerythrindefending them against the oxidative burst of the host [66]We have shown that its virulence factor LPS-PG inducedhigh amounts of ROS via NOX4 which may result in boneresorption favoring the invasiveness of the pathogen whichis at the same time protected by its own defense machineryThis would further explain the pathogenicity and resistanceof this bacterium strain in patients with periodontitis
5 Conclusion
In this study we have demonstrated that LPS from Porphy-romonas gingivalis (LPS-PG) as well as hypoxia induces aNOX4-dependent increase inH
2O2release in PDLfibroblasts
which may contribute to the development and progressionof periodontal diseases when not balanced by a concomitantincrease in antioxidative systems The dramatic elevationin ROS formation induced by the combination of hypoxiaand LPS-PG and the reduction of catalase under the sameconditions increased the oxidative stress in the periodontiumfavoring the progression of inflammation bone resorptionand possibly tooth loss
Taken together the presented data further explain thedestructive mechanism of hypoxic and inflammatory condi-tions in the periodontium and may improve diagnostic andtherapeutic strategies in periodontal diseases
Conflict of Interests
The authors have no conflict of interests to disclose
Acknowledgments
The authors would like to thank Ms Tina Schaffrath for hertechnical supportThe present studywas supported by a grantfrom the German Research Foundation (Clinical ResearchUnit 208TP7) and the Medical Faculty of the University ofBonn
References
[1] S Reuter S C Gupta M M Chaturvedi and B B AggarwalldquoOxidative stress inflammation and cancer how are theylinkedrdquo Free Radical Biology and Medicine vol 49 no 11 pp1603ndash1616 2010
[2] H Sone H Akanuma and T Fukuda ldquoOxygenomics inenvironmental stressrdquo Redox Report vol 15 no 3 pp 98ndash1142010
[3] K E Wellen and C B Thompson ldquoCellular metabolic stressconsidering how cells respond to nutrient excessrdquo MolecularCell vol 40 no 2 pp 323ndash332 2010
[4] R O Poyton K A Ball and P R Castello ldquoMitochondrialgeneration of free radicals and hypoxic signalingrdquo Trends inEndocrinology andMetabolism vol 20 no 7 pp 332ndash340 2009
[5] F Weinberg and N S Chandel ldquoReactive oxygen species-dependent signaling regulates cancerrdquo Cellular and MolecularLife Sciences vol 66 no 23 pp 3663ndash3673 2009
[6] A Cataldi ldquoCell responses to oxidative stressorsrdquo CurrentPharmaceutical Design vol 16 no 12 pp 1387ndash1395 2010
[7] Z Durackova ldquoSome current insights into oxidative stressrdquoPhysiological Research vol 59 no 4 pp 459ndash469 2010
[8] L A Sena and N S Chandel ldquoPhysiological roles of mitochon-drial reactive oxygen speciesrdquoMolecular Cell vol 48 no 2 pp158ndash166 2012
[9] D I Brown and K K Griendling ldquoNox proteins in signaltransductionrdquo Free Radical Biology amp Medicine vol 47 no 9pp 1239ndash1253 2009
[10] K Bedard and K-H Krause ldquoThe NOX family of ROS-generatingNADPHoxidases physiology and pathophysiologyrdquoPhysiological Reviews vol 87 no 1 pp 245ndash313 2007
[11] K-H Krause ldquoTissue distribution and putative physiologicalfunction of NOX family NADPH oxidasesrdquo Japanese Journal ofInfectious Diseases vol 57 no 5 pp S28ndashS29 2004
12 Mediators of Inflammation
[12] S Yang Y Zhang W Ries and L Key ldquoExpression of Nox4 inosteoclastsrdquo Journal of Cellular Biochemistry vol 92 no 2 pp238ndash248 2004
[13] E Pedruzzi C Guichard V Ollivier et al ldquoNAD(P)H oxidaseNox-4mediates 7-ketocholesterol-induced endoplasmic reticu-lum stress and apoptosis in human aortic smooth muscle cellsrdquoMolecular and Cellular Biology vol 24 no 24 pp 10703ndash107172004
[14] J Hwang A Saha Y C Boo et al ldquoOscillatory shear stress stim-ulates endothelial production of O
2-from p47119901ℎ119900119909-dependent
NAD(P)Hoxidases leading tomonocyte adhesionrdquoThe Journalof Biological Chemistry vol 278 no 47 pp 47291ndash47298 2003
[15] H B Suliman M Ali and C A Piantadosi ldquoSuperoxidedismutase-3 promotes full expression of the EPO response tohypoxiardquo Blood vol 104 no 1 pp 43ndash50 2004
[16] P Vallet Y Charnay K Steger et al ldquoNeuronal expressionof the NADPH oxidase NOX4 and its regulation in mouseexperimental brain ischemiardquo Neuroscience vol 132 no 2 pp233ndash238 2005
[17] K T Moe S Aulia F Jiang et al ldquoDifferential upregulation ofNox homologues of NADPH oxidase by tumor necrosis factor-120572 in human aortic smooth muscle and embryonic kidney cellsrdquoJournal of Cellular andMolecularMedicine vol 10 no 1 pp 231ndash239 2006
[18] I Diebold A Petry J Hess and A Gorlach ldquoThe NADPHoxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1rdquoMolecular Biology of the Cell vol 21 no 12pp 2087ndash2096 2010
[19] K V Tormos and N S Chandel ldquoInter-connection betweenmitochondria and HIFsrdquo Journal of Cellular and MolecularMedicine vol 14 no 4 pp 795ndash804 2010
[20] T Ohnishi K Bandow K Kakimoto M Machigashira TMatsuyama and T Matsuguchi ldquoOxidative stress causes alve-olar bone loss in metabolic syndrome model mice with type 2diabetesrdquo Journal of Periodontal Research vol 44 no 1 pp 43ndash51 2009
[21] C Goettsch A Babelova O Trummer et al ldquoNADPH oxidase4 limits bone mass by promoting osteoclastogenesisrdquo Journal ofClinical Investigation vol 123 no 11 pp 4731ndash4738 2013
[22] I L C Chapple and J B Matthews ldquoThe role of reactive oxygenand antioxidant species in periodontal tissue destructionrdquoPeriodontology 2000 vol 43 no 1 pp 160ndash232 2007
[23] A Guentsch P M Preshaw S Bremer-Streck G KlingerE Glockmann and B W Sigusch ldquoLipid peroxidation andantioxidant activity in saliva of periodontitis patients effect ofsmoking and periodontal treatmentrdquo Clinical Oral Investiga-tions vol 12 no 4 pp 345ndash352 2008
[24] R Agnihotri P Pandurang S U Kamath et al ldquoAssociation ofcigarette smoking with superoxide dismutase enzyme levels insubjected with chonic periodontitisrdquo Journal of Periodontologyvol 80 no 4 pp 657ndash662 2009
[25] D Ekuni T Tomofuji N Tamaki et al ldquoMechanical stimulationof gingiva reduces plasma 8-OHdG level in rat periodontitisrdquoArchives of Oral Biology vol 53 no 4 pp 324ndash329 2008
[26] H Staudte A Guntsch A Volpel and B W Sigusch ldquoVitaminC attenuates the cytotoxic effects of Porphyromonas gingivalison human gingival fibroblastsrdquo Archives of Oral Biology vol 55no 1 pp 40ndash45 2010
[27] P Bullon H N Newman and M Battino ldquoObesity diabetesmellitus atherosclerosis and chronic periodontitis a sharedpathology via oxidative stress andmitochondrial dysfunctionrdquoPeriodontology 2000 vol 64 no 1 pp 139ndash153 2014
[28] G R Naveh V Brumfeld R Shahar and S Weiner ldquoToothperiodontal ligament direct 3D microCT visualization of thecollagen network and how the network changes when the toothis loadedrdquo Journal of Structural Biology vol 181 no 2 pp 108ndash115 2013
[29] Y Kitase M Yokozeki S Fujihara et al ldquoAnalysis of geneexpression profiles in human periodontal ligament cells underhypoxia the protective effect of CC chemokine ligand 2 tooxygen shortagerdquoArchives of Oral Biology vol 54 no 7 pp 618ndash624 2009
[30] G Thornton-Evans P Eke L Wei et al ldquoPeriodontitis amongadults aged ge 30 years - United States 2009-2010rdquo Morbidityand Mortality Weekly Report (MMWR) vol 62 supplement 3pp 129ndash135 2013
[31] S S Socransky C Smith and A D Haffajee ldquoSubgingivalmicrobial profiles in refractory periodontal diseaserdquo Journal ofClinical Periodontology vol 29 no 3 pp 260ndash268 2002
[32] K-T Ng J-P Li K M Ng G L Tipoe W K Leung and M-L Fung ldquoExpression of hypoxia-inducible factor-1120572 in humanperiodontal tissuerdquo Journal of Periodontology vol 82 no 1 pp136ndash141 2011
[33] S Frede U Berchner-Pfannschmidt and J Fandrey ldquoReg-ulation of Hypoxia-inducible factors during inflammationrdquoMethods in Enzymology vol 435 pp 405ndash419 2007
[34] J Shin S Ji and Y Choi ldquoAbility of oral bacteria to inducetissue-destructive molecules from human neutrophilsrdquo OralDiseases vol 14 no 4 pp 327ndash334 2008
[35] S Takashiba K Naruishi and Y Murayama ldquoPerspectiveof cytokine regulation for periodontal treatment fibroblastbiologyrdquo Journal of Periodontology vol 74 no 1 pp 103ndash1102003
[36] A Konermann D Stabenow P A Knolle S A E HeldJ Deschner and A Jager ldquoRegulatory role of periodontalligament fibroblasts for innate immune cell function and dif-ferentiationrdquo Innate Immunity vol 18 no 5 pp 745ndash752 2012
[37] S Kar S Subbaram P M Carrico and J A Melendez ldquoRedox-control of matrix metalloproteinase-1 a critical link betweenfree radicals matrix remodeling and degenerative diseaserdquoRespiratory Physiology andNeurobiology vol 174 no 3 pp 299ndash306 2010
[38] F Rousset M V C Nguyen L Grange F Morel and B LardyldquoHeme oxygenase-1 regulates matrix metalloproteinase MMP-1secretion and chondrocyte cell death via Nox4NADPH oxidaseactivity in chondrocytesrdquo PLoS ONE vol 8 no 6 Article IDe66478 2013
[39] E Rudolf and M Cervinka ldquoResponses of human gingival andperiodontal fibroblasts to a low-zinc environmentrdquo Alternativesto Laboratory Animals vol 38 no 2 pp 119ndash138 2010
[40] S M San Miguel L A Opperman E P Allen J Zielinski andK K H Svoboda ldquoBioactive polyphenol antioxidants protectoral fibroblasts from ROS-inducing agentsrdquo Archives of OralBiology vol 57 no 12 pp 1657ndash1667 2012
[41] S M San Miguel L A Opperman E P Allen J E Zielinskiand K K H Svoboda ldquoAntioxidant combinations protect oralfibroblasts against metal-induced toxicityrdquo Archives of OralBiology vol 58 no 3 pp 299ndash310 2013
[42] U Skaleric C M Manthey S E Mergenhagen B Gaspircand S M Wahl ldquoSuperoxide release and superoxide dismutaseexpression by human gingival fibroblastsrdquo European Journal ofOral Sciences vol 108 no 2 pp 130ndash135 2000
Mediators of Inflammation 13
[43] D Y Kim J-H Jun H-L Lee et al ldquoN-acetylcysteine preventsLPS-Lnduced pro-inflammatory cytokines andMMP2 produc-tion in gingival fibroblastsrdquoArchives of Pharmacal Research vol30 no 10 pp 1283ndash1292 2007
[44] B Rath-Deschner J Deschner S Reimann A Jager andW Gotz ldquoRegulatory effects of biomechanical strain on theinsulin-like growth factor system in human periodontal cellsrdquoJournal of Biomechanics vol 42 no 15 pp 2584ndash2589 2009
[45] P Chomczynski and N Sacchi ldquoSingle-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextractionrdquo Analytical Biochemistry vol 162 no 1 pp 156ndash1591987
[46] B Zhao K Ranguelova J Jiang and R P Mason ldquoStudies onthe photosensitized reduction of resorufin and implications forthe detection of oxidative stress with Amplex Redrdquo Free RadicalBiology and Medicine vol 51 no 1 pp 153ndash159 2011
[47] C C Mandal S Ganapathy Y Gorin et al ldquoReactive oxygenspecies derived from Nox4 mediate BMP2 gene transcriptionand osteoblast differentiationrdquoBiochemical Journal vol 433 no2 pp 393ndash402 2011
[48] J Fandrey S Frede and W Jelkmann ldquoRole of hydrogenperoxide in hypoxia-induced erythropoietin productionrdquo Bio-chemical Journal vol 303 no 2 pp 507ndash510 1994
[49] F Cervellati C Cervellati A Romani et al ldquoHypoxia inducescell damage via oxidative stress in retinal epithelial cellsrdquo FreeRadical Research vol 48 no 3 pp 303ndash312 2014
[50] V Vanasco N D Magnani M C Cimolai et al ldquoEndotoxemiaimpairs heart mitochondrial function by decreasing electrontransfer ATP synthesis and ATP content without affectingmembrane potentialrdquo Journal of Bioenergetics and Biomem-branes vol 44 no 2 pp 243ndash252 2012
[51] R G Alscher N Erturk and L S Heath ldquoRole of superoxidedismutases (SODs) in controlling oxidative stress in plantsrdquoJournal of Experimental Botany vol 53 no 372 pp 1331ndash13412002
[52] E Baltacıoglu P Yuva G Aydın et al ldquoLipid peroxidation levelsand total oxidantantioxidant status in serum and saliva frompatients with chronic and aggressive periodontitis Oxidativestress index a new biomarker for periodontal diseaserdquo Journalof Periodontology vol 85 no 10 pp 1432ndash1441 2014
[53] I L C Chapple M R Milward N Ling-Mountford et alldquoAdjunctive daily supplementation with encapsulated fruitvegetable and berry juice powder concentrates and clinicalperiodontal outcomes a double-blind RCTrdquo Journal of ClinicalPeriodontology vol 39 no 1 pp 62ndash72 2012
[54] A Jenzsch S Eick F Rassoul R Purschwitz and H JentschldquoNutritional intervention in patients with periodontal diseaseClinical immunological and microbiological variables during12 monthsrdquo British Journal of Nutrition vol 101 no 6 pp 879ndash885 2009
[55] N Novakovic T Todorovic M Rakic et al ldquoSalivary antiox-idants as periodontal biomarkers in evaluation of tissue statusand treatment outcomerdquo Journal of Periodontal Research vol49 no 1 pp 129ndash136 2014
[56] N Singh S C Narula R K Sharma S Tewari and P KSehgal ldquoVitamin e supplementation superoxide dismutasestatus and outcome of scaling and root planing in patients withchronic periodontitis a randomized clinical trialrdquo Journal ofPeriodontology vol 85 no 2 pp 242ndash249 2014
[57] S Krifka K-A Hiller G Spagnuolo A Jewett G Schmalz andH Schweikl ldquoThe influence of glutathione on redox regulation
by antioxidant proteins and apoptosis in macrophages exposedto 2-hydroxyethyl methacrylate (HEMA)rdquo Biomaterials vol 33no 21 pp 5177ndash5186 2012
[58] I Borges Jr E A M Moreira D W Filho T B de Oliveira MB S da Silva and T S Frode ldquoProinflammatory and oxidativestress markers in patients with periodontal diseaserdquo Mediatorsof Inflammation vol 2007 Article ID 45794 5 pages 2007
[59] M O Tonguc O Ozturk R Sutcu et al ldquoThe impact of smok-ing status on antioxidant enzyme activity and malondialdehydelevels in chronic periodontitisrdquo Journal of Periodontology vol82 no 9 pp 1320ndash1328 2011
[60] B H Jacoby and W L Davis ldquoThe electron microscopicimmunolocalization of a copper-zinc superoxide dismutase inassociation with collagen fibers of periodontal soft tissuesrdquoJournal of Periodontology vol 62 no 7 pp 413ndash420 1991
[61] T Konopka K Krol W Kopec and H Gerber ldquoTotal antioxi-dant status and 8-hydroxy-21015840-deoxyguanosine levels in gingi-val and peripheral blood of periodontitis patientsrdquo ArchivumImmunologiae et Therapiae Experimentalis vol 55 no 6 pp417ndash422 2007
[62] M Soory ldquoRedox status in periodontal and systemic inflamma-tory conditions including associated neoplasias antioxidants asadjunctive therapyrdquo Infectious DisordersmdashDrug Targets vol 9no 4 pp 415ndash427 2009
[63] M Soory ldquoOxidative stress inducedmechanisms in the progres-sion of periodontal diseases and cancer a common approach toredox homeostasisrdquo Cancers vol 2 no 2 pp 670ndash692 2010
[64] R V Chandra G Srinivas A A Reddy et al ldquoLocally deliveredantioxidant gel as an adjunct to nonsurgical therapy improvesmeasures of oxidative stress and periodontal diseaserdquo Journal ofPeriodontal and Implant Science vol 43 no 3 pp 121ndash129 2013
[65] M A D S Siqueira R G Fischer N R Pereira et al ldquoEffectsof non-surgical periodontal treatment on the L-arginine-nitricoxide pathway and oxidative status in plateletsrdquo ExperimentalBiology and Medicine vol 238 no 6 pp 713ndash722 2013
[66] P Mydel Y Takahashi H Yumoto et al ldquoRoles of the hostoxidative immune response and bacterial antioxidant rubr-erythrin during Porphyromonas gingivalis infectionrdquo PLoSpathogens vol 2 no 7 p e76 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 11
stress [57] In our in vitro setting PDL fibroblasts seemto counteract LPS- and hypoxia-induced ROS formationpredominantly with an elevation of CAT expression sincewe could not detect any SOD immunoreactivity The roleand impact of the antioxidative systems SOD and CAT arecontroversially discussed Recent studies measured CAT andSOD activities in gingival tissues of patients with periodon-titis [58 59] Borges Jr et al did not find differences inCAT activities comparing healthy and diseased patients [58]Instead Tonguc et al detected decreased local SOD andCAT activities in periodontitis patients which seemed to beincreased by smoking [59] In contrast in inflamed tissues weobservedhigh amounts of SODwhichwas localized primarilyin epithelial cells Jacoby and Davis determined SOD in peri-odontal soft tissues They detected twice as much SOD activ-ity in the periodontal ligament as in the human skin (dermis)[60] Furthermore immunohistochemical analysis revealedthat SOD is localized in the periphery of matrix collagenfibrils and to the glycocalyx of tissue fibroblasts This wasconfirmed by Skaleric et al [42] which might explain the factthat we did not detect significant differences of SOD levelsafter hypoxic and LPS-PG stimulation intracellularly in PDLfibroblasts Another reason for these results may be causedby the age of the PDL cells used in this study We explantedthe PDL cells from young and oral healthy patients but mostof the immunohistochemical stainings were performed intissues from elderly patients with clearly diagnosed gingivitisor periodontitisTherefore it is possible that in PDL cells fromelderly patients a different expression pattern of antioxidantenzyme systems could be found which further deterioratethe redox state of the periodontal ligament Moreover otherauthors found reduced antioxidants in the body fluids ofpatients with periodontitis indicating a dysregulation whichwould favor disease progression [23 24 61] A variety of invitro and animal studies analyzed the effect of antioxidantsupplement in the context of inflammatory or cytotoxicevents [57 62 63] They have shown that antioxidantsinhibit these processes indicating a therapeutic approachin pathologies like periodontitis First clinical investigationsconfirmed the efficiency of antioxidant micronutrients andadjuncts in periodontal diseases [22 54 56 64 65] Thepresent data indirectly support these studies
Concerning periodontal pathogen invasiveness and viru-lence there is compelling evidence that P gingivalis possessesits own protective antioxidants for example rubrerythrindefending them against the oxidative burst of the host [66]We have shown that its virulence factor LPS-PG inducedhigh amounts of ROS via NOX4 which may result in boneresorption favoring the invasiveness of the pathogen whichis at the same time protected by its own defense machineryThis would further explain the pathogenicity and resistanceof this bacterium strain in patients with periodontitis
5 Conclusion
In this study we have demonstrated that LPS from Porphy-romonas gingivalis (LPS-PG) as well as hypoxia induces aNOX4-dependent increase inH
2O2release in PDLfibroblasts
which may contribute to the development and progressionof periodontal diseases when not balanced by a concomitantincrease in antioxidative systems The dramatic elevationin ROS formation induced by the combination of hypoxiaand LPS-PG and the reduction of catalase under the sameconditions increased the oxidative stress in the periodontiumfavoring the progression of inflammation bone resorptionand possibly tooth loss
Taken together the presented data further explain thedestructive mechanism of hypoxic and inflammatory condi-tions in the periodontium and may improve diagnostic andtherapeutic strategies in periodontal diseases
Conflict of Interests
The authors have no conflict of interests to disclose
Acknowledgments
The authors would like to thank Ms Tina Schaffrath for hertechnical supportThe present studywas supported by a grantfrom the German Research Foundation (Clinical ResearchUnit 208TP7) and the Medical Faculty of the University ofBonn
References
[1] S Reuter S C Gupta M M Chaturvedi and B B AggarwalldquoOxidative stress inflammation and cancer how are theylinkedrdquo Free Radical Biology and Medicine vol 49 no 11 pp1603ndash1616 2010
[2] H Sone H Akanuma and T Fukuda ldquoOxygenomics inenvironmental stressrdquo Redox Report vol 15 no 3 pp 98ndash1142010
[3] K E Wellen and C B Thompson ldquoCellular metabolic stressconsidering how cells respond to nutrient excessrdquo MolecularCell vol 40 no 2 pp 323ndash332 2010
[4] R O Poyton K A Ball and P R Castello ldquoMitochondrialgeneration of free radicals and hypoxic signalingrdquo Trends inEndocrinology andMetabolism vol 20 no 7 pp 332ndash340 2009
[5] F Weinberg and N S Chandel ldquoReactive oxygen species-dependent signaling regulates cancerrdquo Cellular and MolecularLife Sciences vol 66 no 23 pp 3663ndash3673 2009
[6] A Cataldi ldquoCell responses to oxidative stressorsrdquo CurrentPharmaceutical Design vol 16 no 12 pp 1387ndash1395 2010
[7] Z Durackova ldquoSome current insights into oxidative stressrdquoPhysiological Research vol 59 no 4 pp 459ndash469 2010
[8] L A Sena and N S Chandel ldquoPhysiological roles of mitochon-drial reactive oxygen speciesrdquoMolecular Cell vol 48 no 2 pp158ndash166 2012
[9] D I Brown and K K Griendling ldquoNox proteins in signaltransductionrdquo Free Radical Biology amp Medicine vol 47 no 9pp 1239ndash1253 2009
[10] K Bedard and K-H Krause ldquoThe NOX family of ROS-generatingNADPHoxidases physiology and pathophysiologyrdquoPhysiological Reviews vol 87 no 1 pp 245ndash313 2007
[11] K-H Krause ldquoTissue distribution and putative physiologicalfunction of NOX family NADPH oxidasesrdquo Japanese Journal ofInfectious Diseases vol 57 no 5 pp S28ndashS29 2004
12 Mediators of Inflammation
[12] S Yang Y Zhang W Ries and L Key ldquoExpression of Nox4 inosteoclastsrdquo Journal of Cellular Biochemistry vol 92 no 2 pp238ndash248 2004
[13] E Pedruzzi C Guichard V Ollivier et al ldquoNAD(P)H oxidaseNox-4mediates 7-ketocholesterol-induced endoplasmic reticu-lum stress and apoptosis in human aortic smooth muscle cellsrdquoMolecular and Cellular Biology vol 24 no 24 pp 10703ndash107172004
[14] J Hwang A Saha Y C Boo et al ldquoOscillatory shear stress stim-ulates endothelial production of O
2-from p47119901ℎ119900119909-dependent
NAD(P)Hoxidases leading tomonocyte adhesionrdquoThe Journalof Biological Chemistry vol 278 no 47 pp 47291ndash47298 2003
[15] H B Suliman M Ali and C A Piantadosi ldquoSuperoxidedismutase-3 promotes full expression of the EPO response tohypoxiardquo Blood vol 104 no 1 pp 43ndash50 2004
[16] P Vallet Y Charnay K Steger et al ldquoNeuronal expressionof the NADPH oxidase NOX4 and its regulation in mouseexperimental brain ischemiardquo Neuroscience vol 132 no 2 pp233ndash238 2005
[17] K T Moe S Aulia F Jiang et al ldquoDifferential upregulation ofNox homologues of NADPH oxidase by tumor necrosis factor-120572 in human aortic smooth muscle and embryonic kidney cellsrdquoJournal of Cellular andMolecularMedicine vol 10 no 1 pp 231ndash239 2006
[18] I Diebold A Petry J Hess and A Gorlach ldquoThe NADPHoxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1rdquoMolecular Biology of the Cell vol 21 no 12pp 2087ndash2096 2010
[19] K V Tormos and N S Chandel ldquoInter-connection betweenmitochondria and HIFsrdquo Journal of Cellular and MolecularMedicine vol 14 no 4 pp 795ndash804 2010
[20] T Ohnishi K Bandow K Kakimoto M Machigashira TMatsuyama and T Matsuguchi ldquoOxidative stress causes alve-olar bone loss in metabolic syndrome model mice with type 2diabetesrdquo Journal of Periodontal Research vol 44 no 1 pp 43ndash51 2009
[21] C Goettsch A Babelova O Trummer et al ldquoNADPH oxidase4 limits bone mass by promoting osteoclastogenesisrdquo Journal ofClinical Investigation vol 123 no 11 pp 4731ndash4738 2013
[22] I L C Chapple and J B Matthews ldquoThe role of reactive oxygenand antioxidant species in periodontal tissue destructionrdquoPeriodontology 2000 vol 43 no 1 pp 160ndash232 2007
[23] A Guentsch P M Preshaw S Bremer-Streck G KlingerE Glockmann and B W Sigusch ldquoLipid peroxidation andantioxidant activity in saliva of periodontitis patients effect ofsmoking and periodontal treatmentrdquo Clinical Oral Investiga-tions vol 12 no 4 pp 345ndash352 2008
[24] R Agnihotri P Pandurang S U Kamath et al ldquoAssociation ofcigarette smoking with superoxide dismutase enzyme levels insubjected with chonic periodontitisrdquo Journal of Periodontologyvol 80 no 4 pp 657ndash662 2009
[25] D Ekuni T Tomofuji N Tamaki et al ldquoMechanical stimulationof gingiva reduces plasma 8-OHdG level in rat periodontitisrdquoArchives of Oral Biology vol 53 no 4 pp 324ndash329 2008
[26] H Staudte A Guntsch A Volpel and B W Sigusch ldquoVitaminC attenuates the cytotoxic effects of Porphyromonas gingivalison human gingival fibroblastsrdquo Archives of Oral Biology vol 55no 1 pp 40ndash45 2010
[27] P Bullon H N Newman and M Battino ldquoObesity diabetesmellitus atherosclerosis and chronic periodontitis a sharedpathology via oxidative stress andmitochondrial dysfunctionrdquoPeriodontology 2000 vol 64 no 1 pp 139ndash153 2014
[28] G R Naveh V Brumfeld R Shahar and S Weiner ldquoToothperiodontal ligament direct 3D microCT visualization of thecollagen network and how the network changes when the toothis loadedrdquo Journal of Structural Biology vol 181 no 2 pp 108ndash115 2013
[29] Y Kitase M Yokozeki S Fujihara et al ldquoAnalysis of geneexpression profiles in human periodontal ligament cells underhypoxia the protective effect of CC chemokine ligand 2 tooxygen shortagerdquoArchives of Oral Biology vol 54 no 7 pp 618ndash624 2009
[30] G Thornton-Evans P Eke L Wei et al ldquoPeriodontitis amongadults aged ge 30 years - United States 2009-2010rdquo Morbidityand Mortality Weekly Report (MMWR) vol 62 supplement 3pp 129ndash135 2013
[31] S S Socransky C Smith and A D Haffajee ldquoSubgingivalmicrobial profiles in refractory periodontal diseaserdquo Journal ofClinical Periodontology vol 29 no 3 pp 260ndash268 2002
[32] K-T Ng J-P Li K M Ng G L Tipoe W K Leung and M-L Fung ldquoExpression of hypoxia-inducible factor-1120572 in humanperiodontal tissuerdquo Journal of Periodontology vol 82 no 1 pp136ndash141 2011
[33] S Frede U Berchner-Pfannschmidt and J Fandrey ldquoReg-ulation of Hypoxia-inducible factors during inflammationrdquoMethods in Enzymology vol 435 pp 405ndash419 2007
[34] J Shin S Ji and Y Choi ldquoAbility of oral bacteria to inducetissue-destructive molecules from human neutrophilsrdquo OralDiseases vol 14 no 4 pp 327ndash334 2008
[35] S Takashiba K Naruishi and Y Murayama ldquoPerspectiveof cytokine regulation for periodontal treatment fibroblastbiologyrdquo Journal of Periodontology vol 74 no 1 pp 103ndash1102003
[36] A Konermann D Stabenow P A Knolle S A E HeldJ Deschner and A Jager ldquoRegulatory role of periodontalligament fibroblasts for innate immune cell function and dif-ferentiationrdquo Innate Immunity vol 18 no 5 pp 745ndash752 2012
[37] S Kar S Subbaram P M Carrico and J A Melendez ldquoRedox-control of matrix metalloproteinase-1 a critical link betweenfree radicals matrix remodeling and degenerative diseaserdquoRespiratory Physiology andNeurobiology vol 174 no 3 pp 299ndash306 2010
[38] F Rousset M V C Nguyen L Grange F Morel and B LardyldquoHeme oxygenase-1 regulates matrix metalloproteinase MMP-1secretion and chondrocyte cell death via Nox4NADPH oxidaseactivity in chondrocytesrdquo PLoS ONE vol 8 no 6 Article IDe66478 2013
[39] E Rudolf and M Cervinka ldquoResponses of human gingival andperiodontal fibroblasts to a low-zinc environmentrdquo Alternativesto Laboratory Animals vol 38 no 2 pp 119ndash138 2010
[40] S M San Miguel L A Opperman E P Allen J Zielinski andK K H Svoboda ldquoBioactive polyphenol antioxidants protectoral fibroblasts from ROS-inducing agentsrdquo Archives of OralBiology vol 57 no 12 pp 1657ndash1667 2012
[41] S M San Miguel L A Opperman E P Allen J E Zielinskiand K K H Svoboda ldquoAntioxidant combinations protect oralfibroblasts against metal-induced toxicityrdquo Archives of OralBiology vol 58 no 3 pp 299ndash310 2013
[42] U Skaleric C M Manthey S E Mergenhagen B Gaspircand S M Wahl ldquoSuperoxide release and superoxide dismutaseexpression by human gingival fibroblastsrdquo European Journal ofOral Sciences vol 108 no 2 pp 130ndash135 2000
Mediators of Inflammation 13
[43] D Y Kim J-H Jun H-L Lee et al ldquoN-acetylcysteine preventsLPS-Lnduced pro-inflammatory cytokines andMMP2 produc-tion in gingival fibroblastsrdquoArchives of Pharmacal Research vol30 no 10 pp 1283ndash1292 2007
[44] B Rath-Deschner J Deschner S Reimann A Jager andW Gotz ldquoRegulatory effects of biomechanical strain on theinsulin-like growth factor system in human periodontal cellsrdquoJournal of Biomechanics vol 42 no 15 pp 2584ndash2589 2009
[45] P Chomczynski and N Sacchi ldquoSingle-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextractionrdquo Analytical Biochemistry vol 162 no 1 pp 156ndash1591987
[46] B Zhao K Ranguelova J Jiang and R P Mason ldquoStudies onthe photosensitized reduction of resorufin and implications forthe detection of oxidative stress with Amplex Redrdquo Free RadicalBiology and Medicine vol 51 no 1 pp 153ndash159 2011
[47] C C Mandal S Ganapathy Y Gorin et al ldquoReactive oxygenspecies derived from Nox4 mediate BMP2 gene transcriptionand osteoblast differentiationrdquoBiochemical Journal vol 433 no2 pp 393ndash402 2011
[48] J Fandrey S Frede and W Jelkmann ldquoRole of hydrogenperoxide in hypoxia-induced erythropoietin productionrdquo Bio-chemical Journal vol 303 no 2 pp 507ndash510 1994
[49] F Cervellati C Cervellati A Romani et al ldquoHypoxia inducescell damage via oxidative stress in retinal epithelial cellsrdquo FreeRadical Research vol 48 no 3 pp 303ndash312 2014
[50] V Vanasco N D Magnani M C Cimolai et al ldquoEndotoxemiaimpairs heart mitochondrial function by decreasing electrontransfer ATP synthesis and ATP content without affectingmembrane potentialrdquo Journal of Bioenergetics and Biomem-branes vol 44 no 2 pp 243ndash252 2012
[51] R G Alscher N Erturk and L S Heath ldquoRole of superoxidedismutases (SODs) in controlling oxidative stress in plantsrdquoJournal of Experimental Botany vol 53 no 372 pp 1331ndash13412002
[52] E Baltacıoglu P Yuva G Aydın et al ldquoLipid peroxidation levelsand total oxidantantioxidant status in serum and saliva frompatients with chronic and aggressive periodontitis Oxidativestress index a new biomarker for periodontal diseaserdquo Journalof Periodontology vol 85 no 10 pp 1432ndash1441 2014
[53] I L C Chapple M R Milward N Ling-Mountford et alldquoAdjunctive daily supplementation with encapsulated fruitvegetable and berry juice powder concentrates and clinicalperiodontal outcomes a double-blind RCTrdquo Journal of ClinicalPeriodontology vol 39 no 1 pp 62ndash72 2012
[54] A Jenzsch S Eick F Rassoul R Purschwitz and H JentschldquoNutritional intervention in patients with periodontal diseaseClinical immunological and microbiological variables during12 monthsrdquo British Journal of Nutrition vol 101 no 6 pp 879ndash885 2009
[55] N Novakovic T Todorovic M Rakic et al ldquoSalivary antiox-idants as periodontal biomarkers in evaluation of tissue statusand treatment outcomerdquo Journal of Periodontal Research vol49 no 1 pp 129ndash136 2014
[56] N Singh S C Narula R K Sharma S Tewari and P KSehgal ldquoVitamin e supplementation superoxide dismutasestatus and outcome of scaling and root planing in patients withchronic periodontitis a randomized clinical trialrdquo Journal ofPeriodontology vol 85 no 2 pp 242ndash249 2014
[57] S Krifka K-A Hiller G Spagnuolo A Jewett G Schmalz andH Schweikl ldquoThe influence of glutathione on redox regulation
by antioxidant proteins and apoptosis in macrophages exposedto 2-hydroxyethyl methacrylate (HEMA)rdquo Biomaterials vol 33no 21 pp 5177ndash5186 2012
[58] I Borges Jr E A M Moreira D W Filho T B de Oliveira MB S da Silva and T S Frode ldquoProinflammatory and oxidativestress markers in patients with periodontal diseaserdquo Mediatorsof Inflammation vol 2007 Article ID 45794 5 pages 2007
[59] M O Tonguc O Ozturk R Sutcu et al ldquoThe impact of smok-ing status on antioxidant enzyme activity and malondialdehydelevels in chronic periodontitisrdquo Journal of Periodontology vol82 no 9 pp 1320ndash1328 2011
[60] B H Jacoby and W L Davis ldquoThe electron microscopicimmunolocalization of a copper-zinc superoxide dismutase inassociation with collagen fibers of periodontal soft tissuesrdquoJournal of Periodontology vol 62 no 7 pp 413ndash420 1991
[61] T Konopka K Krol W Kopec and H Gerber ldquoTotal antioxi-dant status and 8-hydroxy-21015840-deoxyguanosine levels in gingi-val and peripheral blood of periodontitis patientsrdquo ArchivumImmunologiae et Therapiae Experimentalis vol 55 no 6 pp417ndash422 2007
[62] M Soory ldquoRedox status in periodontal and systemic inflamma-tory conditions including associated neoplasias antioxidants asadjunctive therapyrdquo Infectious DisordersmdashDrug Targets vol 9no 4 pp 415ndash427 2009
[63] M Soory ldquoOxidative stress inducedmechanisms in the progres-sion of periodontal diseases and cancer a common approach toredox homeostasisrdquo Cancers vol 2 no 2 pp 670ndash692 2010
[64] R V Chandra G Srinivas A A Reddy et al ldquoLocally deliveredantioxidant gel as an adjunct to nonsurgical therapy improvesmeasures of oxidative stress and periodontal diseaserdquo Journal ofPeriodontal and Implant Science vol 43 no 3 pp 121ndash129 2013
[65] M A D S Siqueira R G Fischer N R Pereira et al ldquoEffectsof non-surgical periodontal treatment on the L-arginine-nitricoxide pathway and oxidative status in plateletsrdquo ExperimentalBiology and Medicine vol 238 no 6 pp 713ndash722 2013
[66] P Mydel Y Takahashi H Yumoto et al ldquoRoles of the hostoxidative immune response and bacterial antioxidant rubr-erythrin during Porphyromonas gingivalis infectionrdquo PLoSpathogens vol 2 no 7 p e76 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 Mediators of Inflammation
[12] S Yang Y Zhang W Ries and L Key ldquoExpression of Nox4 inosteoclastsrdquo Journal of Cellular Biochemistry vol 92 no 2 pp238ndash248 2004
[13] E Pedruzzi C Guichard V Ollivier et al ldquoNAD(P)H oxidaseNox-4mediates 7-ketocholesterol-induced endoplasmic reticu-lum stress and apoptosis in human aortic smooth muscle cellsrdquoMolecular and Cellular Biology vol 24 no 24 pp 10703ndash107172004
[14] J Hwang A Saha Y C Boo et al ldquoOscillatory shear stress stim-ulates endothelial production of O
2-from p47119901ℎ119900119909-dependent
NAD(P)Hoxidases leading tomonocyte adhesionrdquoThe Journalof Biological Chemistry vol 278 no 47 pp 47291ndash47298 2003
[15] H B Suliman M Ali and C A Piantadosi ldquoSuperoxidedismutase-3 promotes full expression of the EPO response tohypoxiardquo Blood vol 104 no 1 pp 43ndash50 2004
[16] P Vallet Y Charnay K Steger et al ldquoNeuronal expressionof the NADPH oxidase NOX4 and its regulation in mouseexperimental brain ischemiardquo Neuroscience vol 132 no 2 pp233ndash238 2005
[17] K T Moe S Aulia F Jiang et al ldquoDifferential upregulation ofNox homologues of NADPH oxidase by tumor necrosis factor-120572 in human aortic smooth muscle and embryonic kidney cellsrdquoJournal of Cellular andMolecularMedicine vol 10 no 1 pp 231ndash239 2006
[18] I Diebold A Petry J Hess and A Gorlach ldquoThe NADPHoxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1rdquoMolecular Biology of the Cell vol 21 no 12pp 2087ndash2096 2010
[19] K V Tormos and N S Chandel ldquoInter-connection betweenmitochondria and HIFsrdquo Journal of Cellular and MolecularMedicine vol 14 no 4 pp 795ndash804 2010
[20] T Ohnishi K Bandow K Kakimoto M Machigashira TMatsuyama and T Matsuguchi ldquoOxidative stress causes alve-olar bone loss in metabolic syndrome model mice with type 2diabetesrdquo Journal of Periodontal Research vol 44 no 1 pp 43ndash51 2009
[21] C Goettsch A Babelova O Trummer et al ldquoNADPH oxidase4 limits bone mass by promoting osteoclastogenesisrdquo Journal ofClinical Investigation vol 123 no 11 pp 4731ndash4738 2013
[22] I L C Chapple and J B Matthews ldquoThe role of reactive oxygenand antioxidant species in periodontal tissue destructionrdquoPeriodontology 2000 vol 43 no 1 pp 160ndash232 2007
[23] A Guentsch P M Preshaw S Bremer-Streck G KlingerE Glockmann and B W Sigusch ldquoLipid peroxidation andantioxidant activity in saliva of periodontitis patients effect ofsmoking and periodontal treatmentrdquo Clinical Oral Investiga-tions vol 12 no 4 pp 345ndash352 2008
[24] R Agnihotri P Pandurang S U Kamath et al ldquoAssociation ofcigarette smoking with superoxide dismutase enzyme levels insubjected with chonic periodontitisrdquo Journal of Periodontologyvol 80 no 4 pp 657ndash662 2009
[25] D Ekuni T Tomofuji N Tamaki et al ldquoMechanical stimulationof gingiva reduces plasma 8-OHdG level in rat periodontitisrdquoArchives of Oral Biology vol 53 no 4 pp 324ndash329 2008
[26] H Staudte A Guntsch A Volpel and B W Sigusch ldquoVitaminC attenuates the cytotoxic effects of Porphyromonas gingivalison human gingival fibroblastsrdquo Archives of Oral Biology vol 55no 1 pp 40ndash45 2010
[27] P Bullon H N Newman and M Battino ldquoObesity diabetesmellitus atherosclerosis and chronic periodontitis a sharedpathology via oxidative stress andmitochondrial dysfunctionrdquoPeriodontology 2000 vol 64 no 1 pp 139ndash153 2014
[28] G R Naveh V Brumfeld R Shahar and S Weiner ldquoToothperiodontal ligament direct 3D microCT visualization of thecollagen network and how the network changes when the toothis loadedrdquo Journal of Structural Biology vol 181 no 2 pp 108ndash115 2013
[29] Y Kitase M Yokozeki S Fujihara et al ldquoAnalysis of geneexpression profiles in human periodontal ligament cells underhypoxia the protective effect of CC chemokine ligand 2 tooxygen shortagerdquoArchives of Oral Biology vol 54 no 7 pp 618ndash624 2009
[30] G Thornton-Evans P Eke L Wei et al ldquoPeriodontitis amongadults aged ge 30 years - United States 2009-2010rdquo Morbidityand Mortality Weekly Report (MMWR) vol 62 supplement 3pp 129ndash135 2013
[31] S S Socransky C Smith and A D Haffajee ldquoSubgingivalmicrobial profiles in refractory periodontal diseaserdquo Journal ofClinical Periodontology vol 29 no 3 pp 260ndash268 2002
[32] K-T Ng J-P Li K M Ng G L Tipoe W K Leung and M-L Fung ldquoExpression of hypoxia-inducible factor-1120572 in humanperiodontal tissuerdquo Journal of Periodontology vol 82 no 1 pp136ndash141 2011
[33] S Frede U Berchner-Pfannschmidt and J Fandrey ldquoReg-ulation of Hypoxia-inducible factors during inflammationrdquoMethods in Enzymology vol 435 pp 405ndash419 2007
[34] J Shin S Ji and Y Choi ldquoAbility of oral bacteria to inducetissue-destructive molecules from human neutrophilsrdquo OralDiseases vol 14 no 4 pp 327ndash334 2008
[35] S Takashiba K Naruishi and Y Murayama ldquoPerspectiveof cytokine regulation for periodontal treatment fibroblastbiologyrdquo Journal of Periodontology vol 74 no 1 pp 103ndash1102003
[36] A Konermann D Stabenow P A Knolle S A E HeldJ Deschner and A Jager ldquoRegulatory role of periodontalligament fibroblasts for innate immune cell function and dif-ferentiationrdquo Innate Immunity vol 18 no 5 pp 745ndash752 2012
[37] S Kar S Subbaram P M Carrico and J A Melendez ldquoRedox-control of matrix metalloproteinase-1 a critical link betweenfree radicals matrix remodeling and degenerative diseaserdquoRespiratory Physiology andNeurobiology vol 174 no 3 pp 299ndash306 2010
[38] F Rousset M V C Nguyen L Grange F Morel and B LardyldquoHeme oxygenase-1 regulates matrix metalloproteinase MMP-1secretion and chondrocyte cell death via Nox4NADPH oxidaseactivity in chondrocytesrdquo PLoS ONE vol 8 no 6 Article IDe66478 2013
[39] E Rudolf and M Cervinka ldquoResponses of human gingival andperiodontal fibroblasts to a low-zinc environmentrdquo Alternativesto Laboratory Animals vol 38 no 2 pp 119ndash138 2010
[40] S M San Miguel L A Opperman E P Allen J Zielinski andK K H Svoboda ldquoBioactive polyphenol antioxidants protectoral fibroblasts from ROS-inducing agentsrdquo Archives of OralBiology vol 57 no 12 pp 1657ndash1667 2012
[41] S M San Miguel L A Opperman E P Allen J E Zielinskiand K K H Svoboda ldquoAntioxidant combinations protect oralfibroblasts against metal-induced toxicityrdquo Archives of OralBiology vol 58 no 3 pp 299ndash310 2013
[42] U Skaleric C M Manthey S E Mergenhagen B Gaspircand S M Wahl ldquoSuperoxide release and superoxide dismutaseexpression by human gingival fibroblastsrdquo European Journal ofOral Sciences vol 108 no 2 pp 130ndash135 2000
Mediators of Inflammation 13
[43] D Y Kim J-H Jun H-L Lee et al ldquoN-acetylcysteine preventsLPS-Lnduced pro-inflammatory cytokines andMMP2 produc-tion in gingival fibroblastsrdquoArchives of Pharmacal Research vol30 no 10 pp 1283ndash1292 2007
[44] B Rath-Deschner J Deschner S Reimann A Jager andW Gotz ldquoRegulatory effects of biomechanical strain on theinsulin-like growth factor system in human periodontal cellsrdquoJournal of Biomechanics vol 42 no 15 pp 2584ndash2589 2009
[45] P Chomczynski and N Sacchi ldquoSingle-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextractionrdquo Analytical Biochemistry vol 162 no 1 pp 156ndash1591987
[46] B Zhao K Ranguelova J Jiang and R P Mason ldquoStudies onthe photosensitized reduction of resorufin and implications forthe detection of oxidative stress with Amplex Redrdquo Free RadicalBiology and Medicine vol 51 no 1 pp 153ndash159 2011
[47] C C Mandal S Ganapathy Y Gorin et al ldquoReactive oxygenspecies derived from Nox4 mediate BMP2 gene transcriptionand osteoblast differentiationrdquoBiochemical Journal vol 433 no2 pp 393ndash402 2011
[48] J Fandrey S Frede and W Jelkmann ldquoRole of hydrogenperoxide in hypoxia-induced erythropoietin productionrdquo Bio-chemical Journal vol 303 no 2 pp 507ndash510 1994
[49] F Cervellati C Cervellati A Romani et al ldquoHypoxia inducescell damage via oxidative stress in retinal epithelial cellsrdquo FreeRadical Research vol 48 no 3 pp 303ndash312 2014
[50] V Vanasco N D Magnani M C Cimolai et al ldquoEndotoxemiaimpairs heart mitochondrial function by decreasing electrontransfer ATP synthesis and ATP content without affectingmembrane potentialrdquo Journal of Bioenergetics and Biomem-branes vol 44 no 2 pp 243ndash252 2012
[51] R G Alscher N Erturk and L S Heath ldquoRole of superoxidedismutases (SODs) in controlling oxidative stress in plantsrdquoJournal of Experimental Botany vol 53 no 372 pp 1331ndash13412002
[52] E Baltacıoglu P Yuva G Aydın et al ldquoLipid peroxidation levelsand total oxidantantioxidant status in serum and saliva frompatients with chronic and aggressive periodontitis Oxidativestress index a new biomarker for periodontal diseaserdquo Journalof Periodontology vol 85 no 10 pp 1432ndash1441 2014
[53] I L C Chapple M R Milward N Ling-Mountford et alldquoAdjunctive daily supplementation with encapsulated fruitvegetable and berry juice powder concentrates and clinicalperiodontal outcomes a double-blind RCTrdquo Journal of ClinicalPeriodontology vol 39 no 1 pp 62ndash72 2012
[54] A Jenzsch S Eick F Rassoul R Purschwitz and H JentschldquoNutritional intervention in patients with periodontal diseaseClinical immunological and microbiological variables during12 monthsrdquo British Journal of Nutrition vol 101 no 6 pp 879ndash885 2009
[55] N Novakovic T Todorovic M Rakic et al ldquoSalivary antiox-idants as periodontal biomarkers in evaluation of tissue statusand treatment outcomerdquo Journal of Periodontal Research vol49 no 1 pp 129ndash136 2014
[56] N Singh S C Narula R K Sharma S Tewari and P KSehgal ldquoVitamin e supplementation superoxide dismutasestatus and outcome of scaling and root planing in patients withchronic periodontitis a randomized clinical trialrdquo Journal ofPeriodontology vol 85 no 2 pp 242ndash249 2014
[57] S Krifka K-A Hiller G Spagnuolo A Jewett G Schmalz andH Schweikl ldquoThe influence of glutathione on redox regulation
by antioxidant proteins and apoptosis in macrophages exposedto 2-hydroxyethyl methacrylate (HEMA)rdquo Biomaterials vol 33no 21 pp 5177ndash5186 2012
[58] I Borges Jr E A M Moreira D W Filho T B de Oliveira MB S da Silva and T S Frode ldquoProinflammatory and oxidativestress markers in patients with periodontal diseaserdquo Mediatorsof Inflammation vol 2007 Article ID 45794 5 pages 2007
[59] M O Tonguc O Ozturk R Sutcu et al ldquoThe impact of smok-ing status on antioxidant enzyme activity and malondialdehydelevels in chronic periodontitisrdquo Journal of Periodontology vol82 no 9 pp 1320ndash1328 2011
[60] B H Jacoby and W L Davis ldquoThe electron microscopicimmunolocalization of a copper-zinc superoxide dismutase inassociation with collagen fibers of periodontal soft tissuesrdquoJournal of Periodontology vol 62 no 7 pp 413ndash420 1991
[61] T Konopka K Krol W Kopec and H Gerber ldquoTotal antioxi-dant status and 8-hydroxy-21015840-deoxyguanosine levels in gingi-val and peripheral blood of periodontitis patientsrdquo ArchivumImmunologiae et Therapiae Experimentalis vol 55 no 6 pp417ndash422 2007
[62] M Soory ldquoRedox status in periodontal and systemic inflamma-tory conditions including associated neoplasias antioxidants asadjunctive therapyrdquo Infectious DisordersmdashDrug Targets vol 9no 4 pp 415ndash427 2009
[63] M Soory ldquoOxidative stress inducedmechanisms in the progres-sion of periodontal diseases and cancer a common approach toredox homeostasisrdquo Cancers vol 2 no 2 pp 670ndash692 2010
[64] R V Chandra G Srinivas A A Reddy et al ldquoLocally deliveredantioxidant gel as an adjunct to nonsurgical therapy improvesmeasures of oxidative stress and periodontal diseaserdquo Journal ofPeriodontal and Implant Science vol 43 no 3 pp 121ndash129 2013
[65] M A D S Siqueira R G Fischer N R Pereira et al ldquoEffectsof non-surgical periodontal treatment on the L-arginine-nitricoxide pathway and oxidative status in plateletsrdquo ExperimentalBiology and Medicine vol 238 no 6 pp 713ndash722 2013
[66] P Mydel Y Takahashi H Yumoto et al ldquoRoles of the hostoxidative immune response and bacterial antioxidant rubr-erythrin during Porphyromonas gingivalis infectionrdquo PLoSpathogens vol 2 no 7 p e76 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 13
[43] D Y Kim J-H Jun H-L Lee et al ldquoN-acetylcysteine preventsLPS-Lnduced pro-inflammatory cytokines andMMP2 produc-tion in gingival fibroblastsrdquoArchives of Pharmacal Research vol30 no 10 pp 1283ndash1292 2007
[44] B Rath-Deschner J Deschner S Reimann A Jager andW Gotz ldquoRegulatory effects of biomechanical strain on theinsulin-like growth factor system in human periodontal cellsrdquoJournal of Biomechanics vol 42 no 15 pp 2584ndash2589 2009
[45] P Chomczynski and N Sacchi ldquoSingle-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextractionrdquo Analytical Biochemistry vol 162 no 1 pp 156ndash1591987
[46] B Zhao K Ranguelova J Jiang and R P Mason ldquoStudies onthe photosensitized reduction of resorufin and implications forthe detection of oxidative stress with Amplex Redrdquo Free RadicalBiology and Medicine vol 51 no 1 pp 153ndash159 2011
[47] C C Mandal S Ganapathy Y Gorin et al ldquoReactive oxygenspecies derived from Nox4 mediate BMP2 gene transcriptionand osteoblast differentiationrdquoBiochemical Journal vol 433 no2 pp 393ndash402 2011
[48] J Fandrey S Frede and W Jelkmann ldquoRole of hydrogenperoxide in hypoxia-induced erythropoietin productionrdquo Bio-chemical Journal vol 303 no 2 pp 507ndash510 1994
[49] F Cervellati C Cervellati A Romani et al ldquoHypoxia inducescell damage via oxidative stress in retinal epithelial cellsrdquo FreeRadical Research vol 48 no 3 pp 303ndash312 2014
[50] V Vanasco N D Magnani M C Cimolai et al ldquoEndotoxemiaimpairs heart mitochondrial function by decreasing electrontransfer ATP synthesis and ATP content without affectingmembrane potentialrdquo Journal of Bioenergetics and Biomem-branes vol 44 no 2 pp 243ndash252 2012
[51] R G Alscher N Erturk and L S Heath ldquoRole of superoxidedismutases (SODs) in controlling oxidative stress in plantsrdquoJournal of Experimental Botany vol 53 no 372 pp 1331ndash13412002
[52] E Baltacıoglu P Yuva G Aydın et al ldquoLipid peroxidation levelsand total oxidantantioxidant status in serum and saliva frompatients with chronic and aggressive periodontitis Oxidativestress index a new biomarker for periodontal diseaserdquo Journalof Periodontology vol 85 no 10 pp 1432ndash1441 2014
[53] I L C Chapple M R Milward N Ling-Mountford et alldquoAdjunctive daily supplementation with encapsulated fruitvegetable and berry juice powder concentrates and clinicalperiodontal outcomes a double-blind RCTrdquo Journal of ClinicalPeriodontology vol 39 no 1 pp 62ndash72 2012
[54] A Jenzsch S Eick F Rassoul R Purschwitz and H JentschldquoNutritional intervention in patients with periodontal diseaseClinical immunological and microbiological variables during12 monthsrdquo British Journal of Nutrition vol 101 no 6 pp 879ndash885 2009
[55] N Novakovic T Todorovic M Rakic et al ldquoSalivary antiox-idants as periodontal biomarkers in evaluation of tissue statusand treatment outcomerdquo Journal of Periodontal Research vol49 no 1 pp 129ndash136 2014
[56] N Singh S C Narula R K Sharma S Tewari and P KSehgal ldquoVitamin e supplementation superoxide dismutasestatus and outcome of scaling and root planing in patients withchronic periodontitis a randomized clinical trialrdquo Journal ofPeriodontology vol 85 no 2 pp 242ndash249 2014
[57] S Krifka K-A Hiller G Spagnuolo A Jewett G Schmalz andH Schweikl ldquoThe influence of glutathione on redox regulation
by antioxidant proteins and apoptosis in macrophages exposedto 2-hydroxyethyl methacrylate (HEMA)rdquo Biomaterials vol 33no 21 pp 5177ndash5186 2012
[58] I Borges Jr E A M Moreira D W Filho T B de Oliveira MB S da Silva and T S Frode ldquoProinflammatory and oxidativestress markers in patients with periodontal diseaserdquo Mediatorsof Inflammation vol 2007 Article ID 45794 5 pages 2007
[59] M O Tonguc O Ozturk R Sutcu et al ldquoThe impact of smok-ing status on antioxidant enzyme activity and malondialdehydelevels in chronic periodontitisrdquo Journal of Periodontology vol82 no 9 pp 1320ndash1328 2011
[60] B H Jacoby and W L Davis ldquoThe electron microscopicimmunolocalization of a copper-zinc superoxide dismutase inassociation with collagen fibers of periodontal soft tissuesrdquoJournal of Periodontology vol 62 no 7 pp 413ndash420 1991
[61] T Konopka K Krol W Kopec and H Gerber ldquoTotal antioxi-dant status and 8-hydroxy-21015840-deoxyguanosine levels in gingi-val and peripheral blood of periodontitis patientsrdquo ArchivumImmunologiae et Therapiae Experimentalis vol 55 no 6 pp417ndash422 2007
[62] M Soory ldquoRedox status in periodontal and systemic inflamma-tory conditions including associated neoplasias antioxidants asadjunctive therapyrdquo Infectious DisordersmdashDrug Targets vol 9no 4 pp 415ndash427 2009
[63] M Soory ldquoOxidative stress inducedmechanisms in the progres-sion of periodontal diseases and cancer a common approach toredox homeostasisrdquo Cancers vol 2 no 2 pp 670ndash692 2010
[64] R V Chandra G Srinivas A A Reddy et al ldquoLocally deliveredantioxidant gel as an adjunct to nonsurgical therapy improvesmeasures of oxidative stress and periodontal diseaserdquo Journal ofPeriodontal and Implant Science vol 43 no 3 pp 121ndash129 2013
[65] M A D S Siqueira R G Fischer N R Pereira et al ldquoEffectsof non-surgical periodontal treatment on the L-arginine-nitricoxide pathway and oxidative status in plateletsrdquo ExperimentalBiology and Medicine vol 238 no 6 pp 713ndash722 2013
[66] P Mydel Y Takahashi H Yumoto et al ldquoRoles of the hostoxidative immune response and bacterial antioxidant rubr-erythrin during Porphyromonas gingivalis infectionrdquo PLoSpathogens vol 2 no 7 p e76 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom





























