Embed Size (px)
Citation preview

Research ArticleEffects of Parity and Serum Prolactin Levels on the Incidenceand Regression of DMBA-Induced Tumors in OFA hrhr Rats
Corina V Sasso1 Flavia E Santiano1 Constanza M Loacutepez-Fontana1
Virginia Pistone-Creydt1 Marcelo E Ezquer2 Mariacutea B Hapon3
Graciela A Jahn3 and Rubeacuten W Caroacuten1
1 Laboratory of Hormones and Biology of Cancer Institute of Medicine and Experimental Biology of Cuyo (IMBECU)CONICET CCT-Mendoza Avenida Adrian Ruiz-Leal sn CC855 5500 Mendoza Argentina
2 Institute of Sciences Faculty of Medicine German Clinique University of Development Santiago Chile3 Laboratory of Reproduction and Lactation Institute of Medicine and Experimental Biology of Cuyo (IMBECU)CONICET 5500 Mendoza Argentina
Correspondence should be addressed to Ruben W Caron rcaronmendoza-conicetgobar
Received 4 February 2014 Revised 19 June 2014 Accepted 2 July 2014 Published 17 July 2014
Academic Editor Sheba Mohankumar
Copyright copy 2014 Corina V Sasso et alThis is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Prolactin (PRL) is a key player in the development of mammary cancer We studied the effects of parity or hyperprolactinemia onmammary carcinogenesis inOFAhrhr treatedwith 712-dimethylbenzanthraceneTheywere divided into three groups nulliparous(Null) primiparous (PL after pregnancy and lactation) and hyperprolactinemic rats (I implanted in the arcuate nucleus with 17120573-estradiol) The tumor incidence was similar in the three groups However a higher percentage of regressing tumors was evident inthe PL group Serum PRL mammary development and mammary 120573-casein content were higher in I rats compared to Null Theexpression of hormone receptors was similar in the different groups However mammary tissue from PL rats bearing tumors hadincreased expression of PRL and estrogen alpha receptors compared to rats free of tumors Our results suggest that serumPRL levelsdo not have relevance on the incidence of tumors probably because the low levels of PRL in OFA rats are not further decreasedby PL like in other strains However supraphysiological levels of PRL affect carcinogenesis PL induces regression of the tumorsdue to the differentiation produced on the mammary cells Alterations in the expression of hormonal receptors may be involved inprogression and regression of tumors
1 Introduction
Parity is an effective protective factor against breast cancer inwomen and provides protection against chemically inducedmammary carcinogenesis in rats [1ndash5] Although rodentexperimental data and human epidemiological evidence con-sistently show a protective effect of pregnancy on mammarycarcinogenesis the mechanisms underlying this protectionare still unclear [6] The pregnancy-associated refractorinessof the mammary gland to carcinogenesis is caused in partby lasting phenotypic alterations of the mammary epitheliathat occur during pregnancy and lactation [7]The endocrinemilieu is also a determining factor in the parity-associatedrefractoriness tomammary carcinogenesis It has been shown
that treatment of rats with both estrogen (E2) and pro-
gesterone (P4) for a short period of time protects against
mammary carcinogenesis [5 8]With some controversy prolactin (PRL) has been pro-
posed as a key player in the development of mammarycancer in rodent models Thus a direct correlation betweenserum PRL levels and susceptibility of various rat strainsto induction of mammary tumors by chemical carcinogenshas been suggested [9] There is also a direct correlationbetween drug-induced hyperprolactinemia and increasedtumor growth and between hypoprolactinemia and retardedtumor growth [10]
TheOFAhrhr rats are hairless rats derived fromSprague-Dawley with a genetic deficient lactation caused by an
Hindawi Publishing CorporationBioMed Research InternationalVolume 2014 Article ID 210424 11 pageshttpdxdoiorg1011552014210424
2 BioMed Research International
impaired response to the suckling stimulus and heightenedsusceptibility to stress [11 12] The nature of the mutationhas been shown to be a large intragenic deletion of thedesmoglein-4 gene (Dsg-4) encompassing nine exons [13ndash15] which codifies for a protein belonging to the desmogleinfamily The products of these genes expressed in neural andneuroendocrine tissues are cell-adhesion molecules relatedto cadherins Similarly to the effects of parity on women theOFA rats have a persistent reduction in the concentration ofserum PRL and blunted PRL response to secretagogues
To assess the involvement of serum PRL in the suscepti-bility of the mammary gland to carcinogenesis we used OFAhrhr rats and we compared the effects of parity and lactationor induced hyperprolactinemia onmammary carcinogenesisIn the present study we show that the incidence of mammarytumors induced by DMBA in OFA rats is not influencedby serum PRL and that the progression and regression ofthe tumors are dependent on the extent of differentiationproduced in the mammary gland by the effects of parity andlactation
2 Materials and Methods
21 Animals Virgin OFA hrhr female rats (180ndash200 g) bredin our laboratory were used The animals were kept in alight (lights on 0600ndash2000 h) and temperature (22ndash24∘C)controlled room rat chow (Cargill Cordoba Argentina) andtap water were available ad libitum
Animal maintenance and handling were performedaccording to the NIH guide for the Care and Use of Lab-oratory Animals (NIH publication number 86-23 revised1985 and 1991) and the UK requirements for ethics of animalexperimentation (Animals Scientific Procedures Act 1986)
22 Experimental Design All rats were treated per os witha single dose (15mgrat) of 712-dimethylbenzanthracene(DMBA Sigma Buenos Aires) at 54ndash56 days of age and theywere included at random in one of the three experimentalgroups
(i) controls (nulliparous rats -Null- 119899 = 42) rats exposedto DMBA without subsequent treatment
(ii) pregnancy and lactation (primiparous rats -PL- 119899 =82) twenty-five days after carcinogen administration(80 days of age) rats were caged with fertile maleson proestrus afternoonThe following day all the ratsshowing spermatozoa in their vaginal smears werecaged individually and they were checked for theoccurrence of mammary tumor twice a week duringthe 21 days of pregnancy and afterward for 20 daysof lactation Twenty-four hours after delivery all thelitters were adjusted at 8 pups and were maintainedwith their mothers until weaning As previouslypublished [12] OFA rats have normal fertility but 50of the litters die of malnutrition on early lactationonly 6 of the mothers show normal lactation Thusall pups without signs of being properly nourishedwere replaced for age-matched pups This procedure
did not influence the incidence and the proportion oftumor in regression
(iii) estradiol implants (hyperprolactinemic rats -I- 119899 =37) twenty-five days after carcinogen administra-tion the rats were implanted in the arcuate nucleuswith 17120573-estradiol as previously described [16ndash18]Briefly rats were anesthetized by ip injection ofketamine hydrochloride (40mgkg ip) and xylazine(8mgkg im) and placed in a stereotaxic frameIn order to access the area immediately above themedial PeV-ARC region stainless steel cannulae (ID033mm OD 178mm Small Parts Miami FLA)were bilaterally implanted through burr holes drilledthrough the skull over the target sites The tip of thecannulae was brought to the following coordinatesrelative to the bregma 30mm posterior 85mmventral and 06mm right and left Each cannula wasfilled with approximately 05 plusmn 005mg of crystalline17120573-estradiol (Sigma Chem Co St Louis MO)The upper end of cannulae protruded 2-3mm fromthe skull and was fixed to the bone with dentalacrylic cement (Subiton Surrey UK) The accuracyof cannula placement was checked after euthanasia inthe fixed brains by identification of the needle trackin brain sections In general the tips of the cannulaewere above the ARC-PeV region In previous studieswe have demonstrated that the implant of emptycannulas (or filled with placebo) into the arcuatenucleus has no significant effects on PRL secretion[16ndash18]
All animals were palpated twice a week starting at day30 after DMBA administration and for at least 200 days fortumor detection Incidence was calculated as the percentageof rats that had tumors respect to the total number of rats pergroup The rats were decapitated between 1000 and 1200 hon the day the tumors reached a tumor volume gt1000mm3or at the end of the experiment on day 200 when theydid not develop mammary tumors Trunk blood sampleswere allowed to clot at room temperature Serum was storedat minus20∘C until assayed for hormones determinations Afterdecapitation a piece of normal mammary gland and thetumors were removed for 120573-casein content and histopatho-logical analysis
23 Hormone Assays PRL and GH were measured by adouble-antibody RIA as previously described [19] usingmaterials kindly provided by Dr A F Parlow and theNHPP (National Hormone and Pituitary Program Harbor-UCLAMedical Center Torrance CA USA) Hormones wereradioiodinated using the chloramine-T method Results areexpressed in terms of the rat PRLRP-3 andGHRP-2 standardpreparations Assay sensitivity was 05120583gL and the inter- andintra-assay coefficients of variation were less than 10 forboth hormones
24 120573-Casein Determination Mammary 120573-casein was mea-sured as previously described [20 21] Briefly 200mg of
BioMed Research International 3
Table 1 Sequence of primers used in the respective PCR and molecular size of the amplification product
RNAm Sense 51015840-31015840 Antisense 31015840-51015840 SizeActin CGTGGGCCGCCCTAGGCACCA TTGGCCTTAGGGTTCAGAGGGG 243ER120572 AATTCTGACAATCGACGCCAG GTGCTTCAACATTCTCCCTCCTC 345ER120573 AAAGCCAAGAGAAACGGTGGGCAT GCCAATCATGTGCACCAGTTCCTT 204PRLR AAAGTATCTTGTCCAGACTCGCTG AGCAGTTCTTCAGACTTGCCCTT 279PR CCCACAGGAGTTTGTCAAGCTC TAACTTCAGACATCATTTCCGG 325GHR GAGGAGGTGAACACCATCTTGGGC ACCACCTGCTGGTGTAATGTC 534
mammary tissue was cut into small pieces and homogenizedin 2mL 50mM sodium phosphate buffer 150mMNaCl 01NaN3 01TritonX-100 pH 76with anUltraturrax homoge-
nizerThe homogenates were centrifuged at 600 g for 30minThe supernatants were used for 120573-casein determination by ahomologous radioimmunoassay according to Edery et al [22]as modified in our laboratory [23] All samples were assayedin duplicate The standard curve of rat 120573-casein was between025 and 512 ngmL and the sample values were calculated permg of tissue
25 Tumor and Mammary Gland Histology A small pieceof tumor and inguinal mammary gland (contralateral tothe tumor) from each rat were processed for histopatho-logic studies Sections of 3ndash5120583m thickness were cut witha microtome and stained with hematoxylin-eosin (HampE)to define the histopathological changes in the mammaryglands and to classify tumors according to published criteria[24 25] Images were taken with a Nikon Eclipse E200Microscope fitted with a digital still camera MicrometricSE Premium (Nikon Corp Japan) under 100x 400x and600x magnifications The quantification of the percentagesof stroma mostly composed by adipocytes and epithelialtissue in the mammary gland was performed by measuringthe area occupied in 8ndash10 fields of each preparation fromall rats using the ImageJ 142q software available at the NIHsite (httprsbinfonihgovij) Each area was expressed as apercentage of the whole field as previously published [25 26]
26 RNA Isolation and RT-PCR Analysis Total RNA fromnormal mammary glands and tumors was extracted usingthe Chomczynski-Sacchi method modified by Puissant andHoudebine [27] to determine the expression of the followinghormone receptors E
2receptor120572 (ER120572) E
2receptor120573 (ER120573)
PRL receptor (PRLR) P4receptor (PR) and GH receptor
(GHR) Ten micrograms of total RNA were reverse tran-scribed at 42∘C using random hexamer primers andMoloneymurine leukemia virus RT (InvitrogenLife TechnologiesBuenos Aires Argentina) in a 20120583L reaction mixture Beforeproceeding with the semiquantitative PCR the conditionswere established for each tissue such that the amplificationof the products was in the exponential phase and the assaywas linear with respect to the amount of input RNA
All reactions were carried out for 30 cycles with thefollowing cyclic parameters 95∘C for 1min 62∘C for 1minand 72∘C for 2min and then terminated with a 5minextension at 72∘C RNA samples were assayed for DNA
contamination by PCR without prior reverse transcriptionThe PCR products were analyzed on 15 agarose gelscontaining 05mgmL ethidium bromide and photographedwith a Polaroid camera Band intensities of RT-PCR productswere quantified using the ImageJ 142q software availableat the NIH site (httprsbinfonihgovij) Relative levels ofmRNA were expressed as the ratio of signal intensity for thetarget genes relative to that for 120573-actin The sequence of theprimers used for each gene and the size of the product ofamplification obtained are shown in Table 1
27 Statistical Analysis Values are given as means plusmn SEMof 11ndash47 animals per group All statistical analysis wasperformed using GraphPad Prism 501 software (GraphPadSoftware Inc CA USA) Differences in the distribution ofvariables between the three studied groups were assessedusing oneway analysis of variance (ANOVA I) or Kruskal-Wallis test depending on the normality of the variablesas evaluated by the Kolmogorov-Smirnov test Two-wayanalysis of variance (ANOVA II) was used for analysis ofdifferences between rats with and without mammary tumorsin the three groups Post hoc comparisons between meanswere made by Bonferronirsquos test or Dunnrsquos Multiple Compar-ison test Studentrsquos t-test was used when only two groupswere compared When variances were not homogeneouslogarithmic transformation of data was applied Incidenceand percentages of mammary epithelial areas were analyzedby chi-square Differences were considered significant if theprobability was 5 or less
3 Results
31 Incidence Progression and Regression of MammaryTumors Tumor incidence (Table 2) multiplicity and latency(not shown) were not statistically different among the threegroups However a nonsignificant tendency to lower valueswas observed in the PL rats Interestingly a significant rise(119875 lt 0001) in the percentage of tumors with regression andwith macroscopic signs of necrosis was evident in the PL rats(Table 2)
32 Serum Prolactin and GH Levels As expected serum PRLconcentration at the end of the experiment was significantlyhigher in I rats (119875 lt 00001) compared to Null or PL(Figure 1(a)) No correlation between serum PRL levels atthe end of the experiment and percentages of regression oftumors was found
4 BioMed Research International
Table 2 Number and percentages of rats with and without mammary tumors in the three groups
Without tumors With tumors Regressed tumors Necrotic tumors Regular tumors Nulliparous 12 30 714 0 0 0 0 30 100Pregnancy-lactation 35 47 573 17 362lowast 11 234lowast 19 404Implanted 11 26 703 0 0 0 0 26 100lowast119875 lt 005 compared to Nulliparous or Implanted rats
Null I PL0
100
200
300
400
500
600
Seru
m P
RL (n
gm
L)
lowastlowastlowast
(a)
Null T Null NT I T I NT PL T PL NT PL RegT0
255075
100
500
600
700
Seru
m P
RL (n
gm
L)
lowastlowastlowast
(b)
Figure 1 Serum prolactin (PRL) levels in DMBA-treated rats at the end of the experiment (a) Nulliparous rats (Null) and rats after a cycle ofpregnancy and lactation (PL) had significantly lower serumPRL than rats implantedwith 17-120573 estradiol in the arcuate nucleus (I) (119875 lt 00001)This group is referred to as hyperprolactinemic rats (b) When classified depending the development (T) absence (NT) or regression (PLRegT) ofmammary tumors the implanted rats developing tumors showed significantly higher levels of serumPRL than the rest of the animals(119875 lt 00001) including implanted rats without tumors
No significant differences in serum PRL levels wereobserved in Null or PL rats that developed mammary tumorsfrom those that did not However a statistically significantincrease (119875 lt 001) was found in I rats that bore mammarytumors compared to those that did not (Figure 1(b))
Circulating GH levels were similar in all the groupsregardless of tumor development (results not shown)
33 Mammary Gland Tissue Differentiation and DevelopmentThe effect of pregnancy and partial lactation on the devel-opment of the mammary gland was evaluated by histolog-ical observation and measurement of the areas occupiedby parenchyma or stroma Figure 2 shows representativemicrophotographs of HampE-stained mammary tissue fromNull rats ((a) and (b)) I rats ((c) and (d)) and PL rats ((e)and (f)) The mammary glands of Null rats had a normalappearance with few ducts surrounded by a small amountof fibrous connective tissue and abundant adipocytes Asimilar image was seen in hyperprolactinemic rats but witha higher development of alveolar structures The mammarytissue from primiparous rats showed a more noticeabledevelopment of lobuloalveoli Figure 2(g) shows that thepercentages of alveolar tissue in mammary glands from ratsthat bore tumors were higher than in those rats that did notdevelop cancer particularly evident in I rats (119875 lt 001) andin PL rats (119875 lt 0001)
34 Mammary Content of 120573-Casein As expected the mam-mary content of 120573-casein an indicative of the degree ofdifferentiation was increased (119875 lt 0001) in I rats comparedto Null rats A significant increase in mammary 120573-caseinwas produced in PL rats (119875 lt 0001 Figure 3(a)) even longafter the pregnancy lactation cycle Moreover when the ratswere grouped according to the development of tumors nosignificant differences in mammary 120573-casein were observedin Null rats with and without mammary tumors However Irats or PL rats had higher levels of 120573-casein (119875 lt 005 and119875 lt 0001 resp) if they had developed mammary tumors(Figure 3(b))
35 Expression of Hormone Receptors in the NontumoralMammary Tissue We next investigated the level of expres-sion of hormone receptors in the normal tissue of themammary gland from the three groups The expression ofPRLR was similar in the three groups However mammarytissue from PL rats bearing tumors had increased (119875 lt 001)content of PRLR mRNA compared to rats free of tumors(Figure 4(a)) No difference in the expression of total PRmRNA was found between the three groups of treatmentsirrespective of the development of tumors (not shown)
The overall expression of ER120572 in the mammary gland ofI rats was similar to that of Null or PL rats (data not shown)However the mammary gland of I rats that had developedtumors had a diminished expression of ER120572 in comparisonto Null or PL rats bearing mammary tumors (119875 lt 0001
BioMed Research International 5
100120583m
(a)
100120583m
(b)
100120583m
(c)
100120583m
(d)
100120583m
(e)
100120583m
(f)
0
20
40
60
Mam
mar
y al
veol
ar ar
ea (
)
lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowast
lowastlowast
Null T Null NT I T I NT PL T PL NT PL RegT
(g)
Figure 2 Hyperprolactinemia or pregnancy and lactation modify the ratio parenchymastroma in the mammary gland Representativemicrophotographs (100x) of HampE-stained normal mammary tissue from nulliparous rats ((a) and (b)) hyperprolactinemic rats ((c) and (d))and primiparous rats ((e) and (f)) with development ((a) (c) and (e)) or absence ((b) (d) and (f)) of mammary tumors (g) Quantificationsof the relative percentages of the alveolar area Values representmeanplusmn SEM of 8ndash10 fields of each preparation from 11ndash26 animals per grouplowastlowast
119875 lt 0001 lowastlowastlowast119875 lt 00001 comparing the selected groups Comparisons were performed by ANOVA I Arrows show alveolar structures
Figure 4(b)) On the contrary the expression of ER120572 wasincreased in the mammary gland of tumor-bearing Null orPL rats compared to the respective tumor-free rats (119875 lt 005and 119875 lt 001 respectively Figure 4(b))
No significant differences in the expression of ER120573 wereobserved between mammary glands from Null I or PL rats(results not shown) However grouping the rats according tothe presence or not of tumors showed a higher expression ofER120573 only in the Null group (119875 lt 001 Figure 4(b))
36 Changes Related to the Pregnancy and Lactation Cyclein the Mammary Tumors We compared the expression of
120573-casein in the tumors from the three groups to determinethe influence of the different physiological backgrounds onthe changes related to tumor transformation The mammarytumors fromPL rats produced significantly higher levels of120573-casein than the Null or I rats (Figure 5(a)) Moreover 234of the tumors fromprimiparous rats showedmacroscopic andmicroscopic evidence of necrosis (Table 2)
Both PRLR and ER120572 expressions were similar in tumorsdeveloped in Null PL or I rats (results not shown) Howevertumors from I rats showed undetected expression of ER120573 andan increased expression of PR (Figures 5(b) and 5(c))
6 BioMed Research International
Null I PL0
2
4
6
8M
amm
ary120573
-cas
ein
(ng
mg)
lowastlowastlowast
lowastlowastlowast
(a)
Null T Null NT I T I NT PL T PL NT0
2
4
6
8
lowast
lowast
Mam
mar
y120573-c
asei
n (n
gm
g)
(b)
Figure 3 Mammary 120573-casein content at the end of the experiment from DMBA-treated rats (a) Nulliparous rats (Null) had significantlylower mammary 120573-casein than rats implanted with 17-120573 estradiol in the arcuate nucleus (I) or rats after a cycle of pregnancy and lactation(PL) lowastlowastlowast119875 lt 00001 (b) When classified depending on the development (T) or absence (NT) of mammary tumors the implanted andprimiparous rats developing tumors showed significantly higher levels of mammary 120573-casein than the corresponding tumor-free rats lowast119875 lt005 Comparisons were performed by ANOVA I
Null T Null NT I T I NT PL T PL NT00
05
10
PRLR
expr
essio
n
lowastlowast
(a)
00
02
04
06
08
10
ER ex
pres
sion
Null T Null NT I T I NT PL T PL NT
lowastlowastlowast
lowastlowast
lowastlowast
lowast
ER120572ER120573
(b)
Figure 4 Expression of mRNA for hormones receptors in mammary glands from DMBA-treated rats at the end of the experiment (a)Expression of mRNA for PRLR relative to 120573-actin lowastlowast119875 lt 001 with respect to the corresponding tumor-free group (b) Expression of mRNAfor ER120572 and 120573 relative to 120573-actin lowast119875 lt 005 lowastlowast119875 lt 001 lowastlowastlowast119875 lt 00001 with respect to the corresponding tumor-free groups Comparisonswere performed by ANOVA I
37 Changes in the Mammary Gland Related to the Regressionof Tumors In order to investigate whether the increasedpercentage of regressing tumors in the group of primiparousrats is due to long lasting changes in the hormonal milieuor to the grade of differentiation of the mammary gland wecompared primiparous rats bearing tumors with those with-out tumors and rats with regressing tumors No significantdifferences were found between serum PRL concentrationsfrom the PL rats that did not develop tumors those thatbore nonregressed tumors and those that bore regressedtumors (not shown) However a decrease (119875 lt 001) in themammary content of120573-caseinwas evident in the group of ratsthat developed mammary tumors and had tumor regression
(Figure 6(a)) suggesting a link between regression of thetumors and involution of the mammary gland
PRLR expression was significantly lower in mammaryglands from rats that did not develop tumors but no differentfrom those that had regression of the tumors (Figure 6(b))A similar decrease was observed in the expression of ER120572(Figure 6(c)) No significant differences were found in mam-mary expression of PR or ER120573 (results not shown) betweenthe three groups
As shown in Table 2 more than 20 of the tumors fromPL rats showed evident areas of necrosis Figure 7 shows threerepresentativemicrophotographs stained with HampE of tumorfrom Null I and PL rats which shows necrosis
BioMed Research International 7
Null I PL0
2
4
120573-C
asei
n in
tum
ors (
ngm
g)
lowastlowast
lowast
(a)
Null I PL00
01
02
03
04
05
ER120573
expr
essio
n
lowast
(b)
Null I PL00
05
10
15
PR ex
pres
sion
lowastlowast
(c)
Figure 5 120573-Casein content and expression of mRNA for hormones receptors in tumors from DMBA-treated rats (a) Tumor tissue fromnulliparous rats (Null) contains significantly lower amounts of 120573-casein than tumors from implanted (I) or primiparous (PL) rats (119875 lt 005and 119875 lt 0001 resp) (b)The expression of mRNA of ER120573 is lower in the tumors of I rats than in the other two groups (119875 lt 005) (c) mRNAexpression of PR is significantly higher in I rats with respect to Null or PL rats (119875 lt 001)
4 Discussion
To study the involvement of lactogenic hormones in mam-mary carcinogenesis we compared our Null or PL rats withvirgin rats made hyperprolactinemic through hypothalamicimplants of estradiol In this model serum estradiol remainsat a physiological level [16ndash18] allowing us to investigatethe effects of PRL excluding the well-known actions of highamounts of E
2on mammary carcinogenesis
A correlation between serum PRL levels and susceptibil-ity of various rat strains to chemically induced mammarycarcinogenesis has been suggested [9] Both GH and PRLhave been previously shown to be reduced after parturition ascomparedwith nulliparous age-matched animals [28]More-over parous rats showing almost complete refractorinessto chemical carcinogenesis acquire high susceptibility afterhormonal treatment that increases serum GH and PRL levels[29] In our OFA rats neither serum GH nor circulating PRLdecreased after PL Accordingly even when the incidenceof mammary tumors was slightly lower in primiparousrats the difference respect to nulliparae was not statisticallysignificant suggesting that the cycle of pregnancylactationfailed to prevent tumor development in OFA hrhr rats
This different behavior from the original strain (Sprague-Dawley) [28 29] regarding the lack of protection induced bypregnancy and lactation might be due to the lower levels ofcirculating PRL during lactation in the PLOFA ratsThus thedeficient lactation may have been insufficient to protect themammary tissue from tumoral transformation Moreoverdespite the persistent hyperprolactinaemia produced in I ratsthe incidence of tumors was similar to the other groupssuggesting a null impact of serum PRL levels on mammarycarcinogenesis Interestingly in I rats serum PRL was signif-icantly higher in those rats that developed tumors suggestingthat supraphysiological levels of PRL may be relevant inincreasing the risk of mammary cancer
Our results support the idea that the mammary devel-opment per se is not enough to account for the protectiveeffect of parity In fact the implant of estradiol in the arcuatenucleus produces hyperprolactinemia and luteal phase withhigh levels of circulating P
4and mammary development and
differentiation similar to pregnancy [16 17]Thus a protectiveeffect of the implants dependent on the differentiation of themammary gland could be expected However the incidenceof mammary tumors in the I group was similar to that in the
8 BioMed Research International
PL T PL NT PL RegT0
2
4
6
8
Mam
mar
y120573
-cas
ein
(ng
mg)
lowastlowast
(a)
PL T PL NT PL RegT00
05
10
PRL
R ex
pres
sion lowast
(b)
PL T PL NT PL RegT00
05
10
ER120572
expr
essio
n lowastlowast
(c)
Figure 6 120573-Casein content and expression of mRNA for hormones receptors in mammary glands from primiparous rats (PL) Mammary120573-casein content was lower (119875 lt 001) in mammary glands from PL rats that had tumor regression (PLRegT) than in those that did not havetumors (PLNT) or did have no regressing tumors (PLT) (b) and (c) PRLR and ER120572 expressions were lower (119875 lt 005 and 119875 lt 001 resp) inmammary glands from tumor-free PL rats compared to the other two groups
Null rats suggesting that the sustained hyperprolactinaemiais not enough to increase carcinogenesis and moreoverthe development of the mammary gland achieved in theimplanted rats is not enough to prevent chemical carcinogen-esis In agreement with our results chronic treatment withperphenazine producing hyperprolactinemia and high levelsof P4 did not protect the mammary gland against chemical
induction of tumors [30]It is worth noting that in our model mammary 120573-
casein content paralleled the alveolar area measured in thehistological preparations suggesting that the proliferationof epithelial tissue of the mammary gland is accompaniedof certain differentiation This result is in agreement with aprevious study in normal Wistar rats [26]
Interestingly the percentage of mammary tumors inregression was greatly affected by PL indicating that theclassical protective effect of gestation described by othersmaybe due at least in part to an increased number of tumorsundergoing regression rather than a decrease of the develop-ment of mammary tumors Our overall results suggest that acycle of PL has ldquotoxicrdquo effects on themammary tumors whichcannot be explained only by the milieu of mammogenic
hormones since we did not find any statistically significantdifference on serum GH (results not shown) or PRL valuesbetween nulliparous or primiparous rats On the contrarythe significantly lower content of 120573-casein in the mammarygland from rats that had regression of their tumors constitutesan interesting support of the idea that involution of thenormal epithelia can be implied in the regression of tumorsespecially so when the tumors of the rats with pregnancyand lactation retain the capability to synthesize 120573-caseinStudies currently under way in our laboratory are focusing onthe expression of several molecular markers involved in theprocess of involution of the mammary gland after lactationin the regressing tumors
We did not find correlation between expression of ARNmof PRLR and serum PRL levels However mammary glandsfrom PL rats that had developed tumors had increasedexpression of PRLR compared to PL rats free of tumorsThis difference can explain the higher alveolar developmentreached in those rats even when the values of circulating PRLat the moment of sacrifice were similar in both groups Theincreased development of alveolar tissue may cause in turnincreased content of 120573-casein It is worth to note that both I
BioMed Research International 9
(a) (b)
(c)
Figure 7 Representative microphotographs (100x) of HampE-stained normal tumors from nulliparous rats (a) hyperprolactinemic rats (b)and primiparous rats showing a noticeable area of necrosis (c) Arrows show necrotic areas
and PL rats had augmented alveolar development comparedto Null rats but the level of differentiation in terms of 120573-casein expression is higher in PL rats supporting a role for theextent of differentiation more than the alveolar developmentin the protective effect of PL
To assess whether the transformation of the mammarycells involves changes in the expression of receptors westudied in parallel the tumors and tissue from the con-tralateral normal mammary gland in the same animals Theexpression of ER120572 increased in tumors of hyperprolactinemicrats compared to the contralateral mammary gland but itwas unmodified in Null or PL rats (results not-shown) Theability of PRL to stimulate the expression of both types of ERhas been shown in the rat corpus luteum in the mammarygland and in the decidua (see [31] for a review) Previousreports suggest that PRL can stimulate ER expression in somebreast cancers [32 33] Moreover PRL can activate ER120572 evenin the absence of estrogenic ligands leading to oncogenesis[34] In our study the expression of ER120573 decreased in thetumors from the three groups compared to their respectivemammary glands (not shown) suggesting a protective rolefor ER120573 This result supports previous studies showing adecrease in the expression of ER120573 in the process of humanbreast cancer progression associatedwith poor differentiation[35] Moreover the expression of ER120572was higher in untrans-formed mammary tissue from Null or PL rats that developedmammary tumors compared with those that did not On thecontrary hyperprolactinemic rats that developed tumors had
lower expression of ER120572 in their mammary tissue This par-ticular result may suggest that when supraphysiological levelsof circulating PRL are constantly acting on the mammarygland tumors can progress even with lower expression ofER120572
In conclusion in our model of carcinogenesis using OFAhrhr rats we showed that serum PRL levels do not seem tohave relevance on the incidence of tumors probably becausethe relative low levels of PRL in those rats are not furtherdecreased by pregnancy and lactation like in other strainsEven though supraphysiological levels of serum PRL mayaffect carcinogenesis
On the other hand PL induces regression of the tumorsmost probably due to the degree of differentiation producedon the mammary cells than to the extent of proliferationreached by the epithelial tissue Alterations in the expressionof hormonal receptors may be involved in both progressionand regression of the tumors
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are deeply indebted to Mrs Elina Guinazu deDi Nasso and Mr Juan Rosales for their excellent technical
10 BioMed Research International
assistance This work was partially supported by grantsfrom Instituto Nacional del Cancer (Ministerio de SaludArgentina) and fromCONICET (Consejo Nacional de Inves-tigaciones Cientıficas y Tecnicas) Argentina
References
[1] P K Chakravarty and D K Sinha ldquoPregnancy inducedmammary tumor specific effector cells are present long afterparturition in a breast cancer model in ratsrdquo Cancer Letters vol154 no 1 pp 1ndash7 2000
[2] J Russo G A Balogh and I H Russo ldquoFull-term pregnancyinduces a specific genomic signature in the human breastrdquoCancer Epidemiology Biomarkers amp Prevention vol 17 no 1 pp51ndash66 2008
[3] J Russo L K Tay and I H Russo ldquoDifferentiation of themammary gland and susceptibility to carcinogenesisrdquo BreastCancer Research and Treatment vol 2 no 1 pp 5ndash73 1982
[4] OMaller KCHansen T R Lyons et al ldquoCollagen architecturein pregnancy-induced protection from breast cancerrdquo Journal ofCell Science vol 126 part 18 pp 4108ndash4110 2013
[5] S K Siwko J Dong M T Lewis H Liu S G Hilsenbeckand Y Li ldquoEvidence that an early pregnancy causes a persistentdecrease in the number of functional mammary epithelial stemcellsmdashimplications for pregnancy-induced protection againstbreast cancerrdquo Stem Cells vol 26 no 12 pp 3205ndash3209 2008
[6] Y Misra P A Bentley J P Bond S Tighe T Hunter and FZhao ldquoMammary gland morphological and gene expressionchanges underlying pregnancy protection of breast cancertumorigenesisrdquo Physiological Genomics vol 44 no 1 pp 76ndash882012
[7] J Russo and I H Russo ldquoInfluence of differentiation and cellkinetics on the susceptibility of the rat mammary gland tocarcinogenesisrdquo Cancer Research vol 40 no 8 pp 2677ndash26871980
[8] R C Guzman J Yang L Rajkumar G Thordarson X Chenand S Nand ldquoHormonal prevention of breast cancer mim-icking the protective effect of pregnancyrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 5 pp 2520ndash2525 1999
[9] A R Bovns R Buchan E N Cole A P M Forrest and KGrifiths ldquoBasal prolactin blood levels in three strains of ratwith differing incidence of 712-Dimethylbenz (a) anthraceneinduced mammary tumoursrdquo European Journal of Cancer vol9 no 3 pp 169ndash171 1973
[10] C W Welsch and H Nagasawa ldquoProlactin and murine mam-mary tumorigenesis a reviewrdquo Cancer Research vol 37 no 4pp 951ndash963 1977
[11] H Cohen I Sabbagh P Guillaumot J Bertrand M Robert-son and H G Friesen ldquoRelationship between bioassay andradioimmunoassay measurement of prolactin in the IPL rat ahypoprolactinemic rat strainrdquo Biology of Reproduction vol 38no 1 pp 84ndash90 1988
[12] S R Valdes A B Penissi R P Deis and G A JahnldquoHormonal profile and reproductive performance in lactationdeficient (OFA hrhr) and normal (Sprague-Dawley) femaleratsrdquo Reproduction vol 133 no 4 pp 827ndash840 2007
[13] H Bazzi A Kljuic A M Christiano and A A PanteleyevldquoIntragenic deletion in theDesmoglein 4 gene underlies the skinphenotype in the Iffa Credo ldquohairlessrdquo ratrdquo Differentiation vol72 no 8 pp 450ndash464 2004
[14] H Kim A A Panteleyev C A B Jahoda Y Ishii and A MChristiano ldquoGenomic organization and analysis of the hairlessgene in four hypotrichotic rat strainsrdquoMammalianGenome vol15 no 12 pp 975ndash981 2004
[15] B Meyer H Bazzi V Zidek et al ldquoA spontaneous mutationin the desmoglein 4 gene underlies hypotrichosis in a newlanceolate hair rat modelrdquo Differentiation vol 72 no 9-10 pp541ndash547 2004
[16] R W Caron and R P Deis ldquoParticipation of opioid and sero-toninergic systems in prolactin secretion induced by hypotha-lamic action of estradiolrdquoNeuroendocrinology vol 64 no 2 pp124ndash130 1996
[17] R W Caron and R P Deis ldquoEstradiol implants in the arcuatenucleus induce lactogenesis in virgin rats Role of progesteronerdquoLife Sciences vol 62 no 3 pp 229ndash237 1997
[18] G R Morel R W Caron G M Console et al ldquoEstrogeninhibits tuberoinfundibular dopaminergic neurons but does notcause irreversible damagerdquo Brain Research Bulletin vol 80 no6 pp 347ndash352 2009
[19] R W Caron A M Salicioni and R P Deis ldquoMifepristonetreatment demonstrates the participation of adrenal glucocorti-coids in the regulation of oestrogen-induced prolactin secretionin ovariectomized ratsrdquo Journal of Steroid Biochemistry andMolecular Biology vol 48 no 4 pp 385ndash389 1994
[20] R P Deis D G Carrizo and G A Jahn ldquoSuckling-inducedprolactin release potentiates mifepristone-induced lactogenesisin pregnant ratsrdquo Journal of Reproduction and Fertility vol 87no 1 pp 147ndash153 1989
[21] R W Caron C A Jahn and R P Deis ldquoLactogenic actionsof different growth hormone preparations in pregnant andlactating ratsrdquo Journal of Endocrinology vol 142 no 3 pp 535ndash545 1994
[22] M Edery L Houdebine J Djiane and P A Kelly ldquoStudies of 120573-casein content of normal and neoplastic rat mammary tissuesby a homologous radioimmunoassayrdquo Molecular and CellularEndocrinology vol 34 no 2 pp 145ndash151 1984
[23] L E Bussmann and R P Deis ldquo120574-Glutamyltransferase activityin mammary gland of pregnant rats and its regulation by ovar-ian hormones prolactin and placental lactogenrdquo BiochemicalJournal vol 223 no 1 pp 275ndash277 1984
[24] J Russo and I H Russo ldquoAtlas and histologic classification oftumors of the rat mammary glandrdquo Journal of Mammary GlandBiology and Neoplasia vol 5 no 2 pp 187ndash200 2000
[25] C M Lopez-Fontana C V Sasso M E Maselli et al ldquoExperi-mental hypothyroidism increases apoptosis in dimethylbenzan-thracene-inducedmammary tumorsrdquoOncologyReports vol 30no 4 pp 1651ndash1660 2013
[26] C M Lopez-Fontana M E Maselli A M Salicioni and R WCaron ldquoThe inhibitory effect of progesterone on lactogenesisduring pregnancy is already evident by mid- to late gestation inrodentsrdquo Reproduction Fertility and Development vol 24 no 5pp 704ndash714 2012
[27] C Puissant and L M Houdebine ldquoAn improvement of thesingle-step method of RNA isolation by acid guanidiniumthiocyanate-phenol-chloroform extractionrdquo BioTechniques vol8 no 2 pp 148ndash149 1990
[28] G Thordarson E Jin R C Guzman S M Swanson S Nandiand F Talamantes ldquoRefractoriness to mammary tumorigenesisin parous rats is it caused by persistent changes in the hormonalenvironment or permanent biochemical alterations in themammary epitheliardquo Carcinogenesis vol 16 no 11 pp 2847ndash2853 1995
BioMed Research International 11
[29] G Thordarson K van Horn R C Guzman S Nandi and FTalamantes ldquoParous rats regain high susceptibility to chemi-cally induced mammary cancer after treatment with variousmammotropic hormonesrdquo Carcinogenesis vol 22 no 7 pp1027ndash1033 2001
[30] L Sivaraman O M Conneely D Medina and B W OMalleyldquop53 is a potential mediator of pregnancy and hormone-induced resistance to mammary carcinogenesisrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 98 no 22 pp 12379ndash12384 2001
[31] J Frasor and G Gibori ldquoProlactin regulation of estrogenreceptor expressionrdquo Trends in Endocrinology and Metabolismvol 14 no 3 pp 118ndash123 2003
[32] M Edery W Imagawa L Larson and S Nandi ldquoRegulation ofestrogen and progesterone receptor levels in mouse mammaryepithelial cells grown in serum-free collagen gel culturesrdquoEndocrinology vol 116 no 1 pp 105ndash112 1985
[33] C J Ormandy R E Hall D L Manning et al ldquoCoexpres-sion and cross-regulation of the prolactin receptor and sexsteroid hormone receptors in breast cancerrdquo Journal of ClinicalEndocrinology and Metabolism vol 82 no 11 pp 3692ndash36991997
[34] K A OrsquoLeary F Jallow D E Rugowski et al ldquoProlactinactivates ER120572 in the absence of ligand in female mammarydevelopment and carcinogenesis in vivordquo Endocrinology vol154 pp 4483ndash4492 2013
[35] B W Park K S Kim M K Heo et al ldquoExpression ofestrogen receptor-120573 in normal mammary and tumor tissues isit protective in breast carcinogenesisrdquo Breast Cancer Researchand Treatment vol 80 no 1 pp 79ndash85 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 BioMed Research International
impaired response to the suckling stimulus and heightenedsusceptibility to stress [11 12] The nature of the mutationhas been shown to be a large intragenic deletion of thedesmoglein-4 gene (Dsg-4) encompassing nine exons [13ndash15] which codifies for a protein belonging to the desmogleinfamily The products of these genes expressed in neural andneuroendocrine tissues are cell-adhesion molecules relatedto cadherins Similarly to the effects of parity on women theOFA rats have a persistent reduction in the concentration ofserum PRL and blunted PRL response to secretagogues
To assess the involvement of serum PRL in the suscepti-bility of the mammary gland to carcinogenesis we used OFAhrhr rats and we compared the effects of parity and lactationor induced hyperprolactinemia onmammary carcinogenesisIn the present study we show that the incidence of mammarytumors induced by DMBA in OFA rats is not influencedby serum PRL and that the progression and regression ofthe tumors are dependent on the extent of differentiationproduced in the mammary gland by the effects of parity andlactation
2 Materials and Methods
21 Animals Virgin OFA hrhr female rats (180ndash200 g) bredin our laboratory were used The animals were kept in alight (lights on 0600ndash2000 h) and temperature (22ndash24∘C)controlled room rat chow (Cargill Cordoba Argentina) andtap water were available ad libitum
Animal maintenance and handling were performedaccording to the NIH guide for the Care and Use of Lab-oratory Animals (NIH publication number 86-23 revised1985 and 1991) and the UK requirements for ethics of animalexperimentation (Animals Scientific Procedures Act 1986)
22 Experimental Design All rats were treated per os witha single dose (15mgrat) of 712-dimethylbenzanthracene(DMBA Sigma Buenos Aires) at 54ndash56 days of age and theywere included at random in one of the three experimentalgroups
(i) controls (nulliparous rats -Null- 119899 = 42) rats exposedto DMBA without subsequent treatment
(ii) pregnancy and lactation (primiparous rats -PL- 119899 =82) twenty-five days after carcinogen administration(80 days of age) rats were caged with fertile maleson proestrus afternoonThe following day all the ratsshowing spermatozoa in their vaginal smears werecaged individually and they were checked for theoccurrence of mammary tumor twice a week duringthe 21 days of pregnancy and afterward for 20 daysof lactation Twenty-four hours after delivery all thelitters were adjusted at 8 pups and were maintainedwith their mothers until weaning As previouslypublished [12] OFA rats have normal fertility but 50of the litters die of malnutrition on early lactationonly 6 of the mothers show normal lactation Thusall pups without signs of being properly nourishedwere replaced for age-matched pups This procedure
did not influence the incidence and the proportion oftumor in regression
(iii) estradiol implants (hyperprolactinemic rats -I- 119899 =37) twenty-five days after carcinogen administra-tion the rats were implanted in the arcuate nucleuswith 17120573-estradiol as previously described [16ndash18]Briefly rats were anesthetized by ip injection ofketamine hydrochloride (40mgkg ip) and xylazine(8mgkg im) and placed in a stereotaxic frameIn order to access the area immediately above themedial PeV-ARC region stainless steel cannulae (ID033mm OD 178mm Small Parts Miami FLA)were bilaterally implanted through burr holes drilledthrough the skull over the target sites The tip of thecannulae was brought to the following coordinatesrelative to the bregma 30mm posterior 85mmventral and 06mm right and left Each cannula wasfilled with approximately 05 plusmn 005mg of crystalline17120573-estradiol (Sigma Chem Co St Louis MO)The upper end of cannulae protruded 2-3mm fromthe skull and was fixed to the bone with dentalacrylic cement (Subiton Surrey UK) The accuracyof cannula placement was checked after euthanasia inthe fixed brains by identification of the needle trackin brain sections In general the tips of the cannulaewere above the ARC-PeV region In previous studieswe have demonstrated that the implant of emptycannulas (or filled with placebo) into the arcuatenucleus has no significant effects on PRL secretion[16ndash18]
All animals were palpated twice a week starting at day30 after DMBA administration and for at least 200 days fortumor detection Incidence was calculated as the percentageof rats that had tumors respect to the total number of rats pergroup The rats were decapitated between 1000 and 1200 hon the day the tumors reached a tumor volume gt1000mm3or at the end of the experiment on day 200 when theydid not develop mammary tumors Trunk blood sampleswere allowed to clot at room temperature Serum was storedat minus20∘C until assayed for hormones determinations Afterdecapitation a piece of normal mammary gland and thetumors were removed for 120573-casein content and histopatho-logical analysis
23 Hormone Assays PRL and GH were measured by adouble-antibody RIA as previously described [19] usingmaterials kindly provided by Dr A F Parlow and theNHPP (National Hormone and Pituitary Program Harbor-UCLAMedical Center Torrance CA USA) Hormones wereradioiodinated using the chloramine-T method Results areexpressed in terms of the rat PRLRP-3 andGHRP-2 standardpreparations Assay sensitivity was 05120583gL and the inter- andintra-assay coefficients of variation were less than 10 forboth hormones
24 120573-Casein Determination Mammary 120573-casein was mea-sured as previously described [20 21] Briefly 200mg of
BioMed Research International 3
Table 1 Sequence of primers used in the respective PCR and molecular size of the amplification product
RNAm Sense 51015840-31015840 Antisense 31015840-51015840 SizeActin CGTGGGCCGCCCTAGGCACCA TTGGCCTTAGGGTTCAGAGGGG 243ER120572 AATTCTGACAATCGACGCCAG GTGCTTCAACATTCTCCCTCCTC 345ER120573 AAAGCCAAGAGAAACGGTGGGCAT GCCAATCATGTGCACCAGTTCCTT 204PRLR AAAGTATCTTGTCCAGACTCGCTG AGCAGTTCTTCAGACTTGCCCTT 279PR CCCACAGGAGTTTGTCAAGCTC TAACTTCAGACATCATTTCCGG 325GHR GAGGAGGTGAACACCATCTTGGGC ACCACCTGCTGGTGTAATGTC 534
mammary tissue was cut into small pieces and homogenizedin 2mL 50mM sodium phosphate buffer 150mMNaCl 01NaN3 01TritonX-100 pH 76with anUltraturrax homoge-
nizerThe homogenates were centrifuged at 600 g for 30minThe supernatants were used for 120573-casein determination by ahomologous radioimmunoassay according to Edery et al [22]as modified in our laboratory [23] All samples were assayedin duplicate The standard curve of rat 120573-casein was between025 and 512 ngmL and the sample values were calculated permg of tissue
25 Tumor and Mammary Gland Histology A small pieceof tumor and inguinal mammary gland (contralateral tothe tumor) from each rat were processed for histopatho-logic studies Sections of 3ndash5120583m thickness were cut witha microtome and stained with hematoxylin-eosin (HampE)to define the histopathological changes in the mammaryglands and to classify tumors according to published criteria[24 25] Images were taken with a Nikon Eclipse E200Microscope fitted with a digital still camera MicrometricSE Premium (Nikon Corp Japan) under 100x 400x and600x magnifications The quantification of the percentagesof stroma mostly composed by adipocytes and epithelialtissue in the mammary gland was performed by measuringthe area occupied in 8ndash10 fields of each preparation fromall rats using the ImageJ 142q software available at the NIHsite (httprsbinfonihgovij) Each area was expressed as apercentage of the whole field as previously published [25 26]
26 RNA Isolation and RT-PCR Analysis Total RNA fromnormal mammary glands and tumors was extracted usingthe Chomczynski-Sacchi method modified by Puissant andHoudebine [27] to determine the expression of the followinghormone receptors E
2receptor120572 (ER120572) E
2receptor120573 (ER120573)
PRL receptor (PRLR) P4receptor (PR) and GH receptor
(GHR) Ten micrograms of total RNA were reverse tran-scribed at 42∘C using random hexamer primers andMoloneymurine leukemia virus RT (InvitrogenLife TechnologiesBuenos Aires Argentina) in a 20120583L reaction mixture Beforeproceeding with the semiquantitative PCR the conditionswere established for each tissue such that the amplificationof the products was in the exponential phase and the assaywas linear with respect to the amount of input RNA
All reactions were carried out for 30 cycles with thefollowing cyclic parameters 95∘C for 1min 62∘C for 1minand 72∘C for 2min and then terminated with a 5minextension at 72∘C RNA samples were assayed for DNA
contamination by PCR without prior reverse transcriptionThe PCR products were analyzed on 15 agarose gelscontaining 05mgmL ethidium bromide and photographedwith a Polaroid camera Band intensities of RT-PCR productswere quantified using the ImageJ 142q software availableat the NIH site (httprsbinfonihgovij) Relative levels ofmRNA were expressed as the ratio of signal intensity for thetarget genes relative to that for 120573-actin The sequence of theprimers used for each gene and the size of the product ofamplification obtained are shown in Table 1
27 Statistical Analysis Values are given as means plusmn SEMof 11ndash47 animals per group All statistical analysis wasperformed using GraphPad Prism 501 software (GraphPadSoftware Inc CA USA) Differences in the distribution ofvariables between the three studied groups were assessedusing oneway analysis of variance (ANOVA I) or Kruskal-Wallis test depending on the normality of the variablesas evaluated by the Kolmogorov-Smirnov test Two-wayanalysis of variance (ANOVA II) was used for analysis ofdifferences between rats with and without mammary tumorsin the three groups Post hoc comparisons between meanswere made by Bonferronirsquos test or Dunnrsquos Multiple Compar-ison test Studentrsquos t-test was used when only two groupswere compared When variances were not homogeneouslogarithmic transformation of data was applied Incidenceand percentages of mammary epithelial areas were analyzedby chi-square Differences were considered significant if theprobability was 5 or less
3 Results
31 Incidence Progression and Regression of MammaryTumors Tumor incidence (Table 2) multiplicity and latency(not shown) were not statistically different among the threegroups However a nonsignificant tendency to lower valueswas observed in the PL rats Interestingly a significant rise(119875 lt 0001) in the percentage of tumors with regression andwith macroscopic signs of necrosis was evident in the PL rats(Table 2)
32 Serum Prolactin and GH Levels As expected serum PRLconcentration at the end of the experiment was significantlyhigher in I rats (119875 lt 00001) compared to Null or PL(Figure 1(a)) No correlation between serum PRL levels atthe end of the experiment and percentages of regression oftumors was found
4 BioMed Research International
Table 2 Number and percentages of rats with and without mammary tumors in the three groups
Without tumors With tumors Regressed tumors Necrotic tumors Regular tumors Nulliparous 12 30 714 0 0 0 0 30 100Pregnancy-lactation 35 47 573 17 362lowast 11 234lowast 19 404Implanted 11 26 703 0 0 0 0 26 100lowast119875 lt 005 compared to Nulliparous or Implanted rats
Null I PL0
100
200
300
400
500
600
Seru
m P
RL (n
gm
L)
lowastlowastlowast
(a)
Null T Null NT I T I NT PL T PL NT PL RegT0
255075
100
500
600
700
Seru
m P
RL (n
gm
L)
lowastlowastlowast
(b)
Figure 1 Serum prolactin (PRL) levels in DMBA-treated rats at the end of the experiment (a) Nulliparous rats (Null) and rats after a cycle ofpregnancy and lactation (PL) had significantly lower serumPRL than rats implantedwith 17-120573 estradiol in the arcuate nucleus (I) (119875 lt 00001)This group is referred to as hyperprolactinemic rats (b) When classified depending the development (T) absence (NT) or regression (PLRegT) ofmammary tumors the implanted rats developing tumors showed significantly higher levels of serumPRL than the rest of the animals(119875 lt 00001) including implanted rats without tumors
No significant differences in serum PRL levels wereobserved in Null or PL rats that developed mammary tumorsfrom those that did not However a statistically significantincrease (119875 lt 001) was found in I rats that bore mammarytumors compared to those that did not (Figure 1(b))
Circulating GH levels were similar in all the groupsregardless of tumor development (results not shown)
33 Mammary Gland Tissue Differentiation and DevelopmentThe effect of pregnancy and partial lactation on the devel-opment of the mammary gland was evaluated by histolog-ical observation and measurement of the areas occupiedby parenchyma or stroma Figure 2 shows representativemicrophotographs of HampE-stained mammary tissue fromNull rats ((a) and (b)) I rats ((c) and (d)) and PL rats ((e)and (f)) The mammary glands of Null rats had a normalappearance with few ducts surrounded by a small amountof fibrous connective tissue and abundant adipocytes Asimilar image was seen in hyperprolactinemic rats but witha higher development of alveolar structures The mammarytissue from primiparous rats showed a more noticeabledevelopment of lobuloalveoli Figure 2(g) shows that thepercentages of alveolar tissue in mammary glands from ratsthat bore tumors were higher than in those rats that did notdevelop cancer particularly evident in I rats (119875 lt 001) andin PL rats (119875 lt 0001)
34 Mammary Content of 120573-Casein As expected the mam-mary content of 120573-casein an indicative of the degree ofdifferentiation was increased (119875 lt 0001) in I rats comparedto Null rats A significant increase in mammary 120573-caseinwas produced in PL rats (119875 lt 0001 Figure 3(a)) even longafter the pregnancy lactation cycle Moreover when the ratswere grouped according to the development of tumors nosignificant differences in mammary 120573-casein were observedin Null rats with and without mammary tumors However Irats or PL rats had higher levels of 120573-casein (119875 lt 005 and119875 lt 0001 resp) if they had developed mammary tumors(Figure 3(b))
35 Expression of Hormone Receptors in the NontumoralMammary Tissue We next investigated the level of expres-sion of hormone receptors in the normal tissue of themammary gland from the three groups The expression ofPRLR was similar in the three groups However mammarytissue from PL rats bearing tumors had increased (119875 lt 001)content of PRLR mRNA compared to rats free of tumors(Figure 4(a)) No difference in the expression of total PRmRNA was found between the three groups of treatmentsirrespective of the development of tumors (not shown)
The overall expression of ER120572 in the mammary gland ofI rats was similar to that of Null or PL rats (data not shown)However the mammary gland of I rats that had developedtumors had a diminished expression of ER120572 in comparisonto Null or PL rats bearing mammary tumors (119875 lt 0001
BioMed Research International 5
100120583m
(a)
100120583m
(b)
100120583m
(c)
100120583m
(d)
100120583m
(e)
100120583m
(f)
0
20
40
60
Mam
mar
y al
veol
ar ar
ea (
)
lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowast
lowastlowast
Null T Null NT I T I NT PL T PL NT PL RegT
(g)
Figure 2 Hyperprolactinemia or pregnancy and lactation modify the ratio parenchymastroma in the mammary gland Representativemicrophotographs (100x) of HampE-stained normal mammary tissue from nulliparous rats ((a) and (b)) hyperprolactinemic rats ((c) and (d))and primiparous rats ((e) and (f)) with development ((a) (c) and (e)) or absence ((b) (d) and (f)) of mammary tumors (g) Quantificationsof the relative percentages of the alveolar area Values representmeanplusmn SEM of 8ndash10 fields of each preparation from 11ndash26 animals per grouplowastlowast
119875 lt 0001 lowastlowastlowast119875 lt 00001 comparing the selected groups Comparisons were performed by ANOVA I Arrows show alveolar structures
Figure 4(b)) On the contrary the expression of ER120572 wasincreased in the mammary gland of tumor-bearing Null orPL rats compared to the respective tumor-free rats (119875 lt 005and 119875 lt 001 respectively Figure 4(b))
No significant differences in the expression of ER120573 wereobserved between mammary glands from Null I or PL rats(results not shown) However grouping the rats according tothe presence or not of tumors showed a higher expression ofER120573 only in the Null group (119875 lt 001 Figure 4(b))
36 Changes Related to the Pregnancy and Lactation Cyclein the Mammary Tumors We compared the expression of
120573-casein in the tumors from the three groups to determinethe influence of the different physiological backgrounds onthe changes related to tumor transformation The mammarytumors fromPL rats produced significantly higher levels of120573-casein than the Null or I rats (Figure 5(a)) Moreover 234of the tumors fromprimiparous rats showedmacroscopic andmicroscopic evidence of necrosis (Table 2)
Both PRLR and ER120572 expressions were similar in tumorsdeveloped in Null PL or I rats (results not shown) Howevertumors from I rats showed undetected expression of ER120573 andan increased expression of PR (Figures 5(b) and 5(c))
6 BioMed Research International
Null I PL0
2
4
6
8M
amm
ary120573
-cas
ein
(ng
mg)
lowastlowastlowast
lowastlowastlowast
(a)
Null T Null NT I T I NT PL T PL NT0
2
4
6
8
lowast
lowast
Mam
mar
y120573-c
asei
n (n
gm
g)
(b)
Figure 3 Mammary 120573-casein content at the end of the experiment from DMBA-treated rats (a) Nulliparous rats (Null) had significantlylower mammary 120573-casein than rats implanted with 17-120573 estradiol in the arcuate nucleus (I) or rats after a cycle of pregnancy and lactation(PL) lowastlowastlowast119875 lt 00001 (b) When classified depending on the development (T) or absence (NT) of mammary tumors the implanted andprimiparous rats developing tumors showed significantly higher levels of mammary 120573-casein than the corresponding tumor-free rats lowast119875 lt005 Comparisons were performed by ANOVA I
Null T Null NT I T I NT PL T PL NT00
05
10
PRLR
expr
essio
n
lowastlowast
(a)
00
02
04
06
08
10
ER ex
pres
sion
Null T Null NT I T I NT PL T PL NT
lowastlowastlowast
lowastlowast
lowastlowast
lowast
ER120572ER120573
(b)
Figure 4 Expression of mRNA for hormones receptors in mammary glands from DMBA-treated rats at the end of the experiment (a)Expression of mRNA for PRLR relative to 120573-actin lowastlowast119875 lt 001 with respect to the corresponding tumor-free group (b) Expression of mRNAfor ER120572 and 120573 relative to 120573-actin lowast119875 lt 005 lowastlowast119875 lt 001 lowastlowastlowast119875 lt 00001 with respect to the corresponding tumor-free groups Comparisonswere performed by ANOVA I
37 Changes in the Mammary Gland Related to the Regressionof Tumors In order to investigate whether the increasedpercentage of regressing tumors in the group of primiparousrats is due to long lasting changes in the hormonal milieuor to the grade of differentiation of the mammary gland wecompared primiparous rats bearing tumors with those with-out tumors and rats with regressing tumors No significantdifferences were found between serum PRL concentrationsfrom the PL rats that did not develop tumors those thatbore nonregressed tumors and those that bore regressedtumors (not shown) However a decrease (119875 lt 001) in themammary content of120573-caseinwas evident in the group of ratsthat developed mammary tumors and had tumor regression
(Figure 6(a)) suggesting a link between regression of thetumors and involution of the mammary gland
PRLR expression was significantly lower in mammaryglands from rats that did not develop tumors but no differentfrom those that had regression of the tumors (Figure 6(b))A similar decrease was observed in the expression of ER120572(Figure 6(c)) No significant differences were found in mam-mary expression of PR or ER120573 (results not shown) betweenthe three groups
As shown in Table 2 more than 20 of the tumors fromPL rats showed evident areas of necrosis Figure 7 shows threerepresentativemicrophotographs stained with HampE of tumorfrom Null I and PL rats which shows necrosis
BioMed Research International 7
Null I PL0
2
4
120573-C
asei
n in
tum
ors (
ngm
g)
lowastlowast
lowast
(a)
Null I PL00
01
02
03
04
05
ER120573
expr
essio
n
lowast
(b)
Null I PL00
05
10
15
PR ex
pres
sion
lowastlowast
(c)
Figure 5 120573-Casein content and expression of mRNA for hormones receptors in tumors from DMBA-treated rats (a) Tumor tissue fromnulliparous rats (Null) contains significantly lower amounts of 120573-casein than tumors from implanted (I) or primiparous (PL) rats (119875 lt 005and 119875 lt 0001 resp) (b)The expression of mRNA of ER120573 is lower in the tumors of I rats than in the other two groups (119875 lt 005) (c) mRNAexpression of PR is significantly higher in I rats with respect to Null or PL rats (119875 lt 001)
4 Discussion
To study the involvement of lactogenic hormones in mam-mary carcinogenesis we compared our Null or PL rats withvirgin rats made hyperprolactinemic through hypothalamicimplants of estradiol In this model serum estradiol remainsat a physiological level [16ndash18] allowing us to investigatethe effects of PRL excluding the well-known actions of highamounts of E
2on mammary carcinogenesis
A correlation between serum PRL levels and susceptibil-ity of various rat strains to chemically induced mammarycarcinogenesis has been suggested [9] Both GH and PRLhave been previously shown to be reduced after parturition ascomparedwith nulliparous age-matched animals [28]More-over parous rats showing almost complete refractorinessto chemical carcinogenesis acquire high susceptibility afterhormonal treatment that increases serum GH and PRL levels[29] In our OFA rats neither serum GH nor circulating PRLdecreased after PL Accordingly even when the incidenceof mammary tumors was slightly lower in primiparousrats the difference respect to nulliparae was not statisticallysignificant suggesting that the cycle of pregnancylactationfailed to prevent tumor development in OFA hrhr rats
This different behavior from the original strain (Sprague-Dawley) [28 29] regarding the lack of protection induced bypregnancy and lactation might be due to the lower levels ofcirculating PRL during lactation in the PLOFA ratsThus thedeficient lactation may have been insufficient to protect themammary tissue from tumoral transformation Moreoverdespite the persistent hyperprolactinaemia produced in I ratsthe incidence of tumors was similar to the other groupssuggesting a null impact of serum PRL levels on mammarycarcinogenesis Interestingly in I rats serum PRL was signif-icantly higher in those rats that developed tumors suggestingthat supraphysiological levels of PRL may be relevant inincreasing the risk of mammary cancer
Our results support the idea that the mammary devel-opment per se is not enough to account for the protectiveeffect of parity In fact the implant of estradiol in the arcuatenucleus produces hyperprolactinemia and luteal phase withhigh levels of circulating P
4and mammary development and
differentiation similar to pregnancy [16 17]Thus a protectiveeffect of the implants dependent on the differentiation of themammary gland could be expected However the incidenceof mammary tumors in the I group was similar to that in the
8 BioMed Research International
PL T PL NT PL RegT0
2
4
6
8
Mam
mar
y120573
-cas
ein
(ng
mg)
lowastlowast
(a)
PL T PL NT PL RegT00
05
10
PRL
R ex
pres
sion lowast
(b)
PL T PL NT PL RegT00
05
10
ER120572
expr
essio
n lowastlowast
(c)
Figure 6 120573-Casein content and expression of mRNA for hormones receptors in mammary glands from primiparous rats (PL) Mammary120573-casein content was lower (119875 lt 001) in mammary glands from PL rats that had tumor regression (PLRegT) than in those that did not havetumors (PLNT) or did have no regressing tumors (PLT) (b) and (c) PRLR and ER120572 expressions were lower (119875 lt 005 and 119875 lt 001 resp) inmammary glands from tumor-free PL rats compared to the other two groups
Null rats suggesting that the sustained hyperprolactinaemiais not enough to increase carcinogenesis and moreoverthe development of the mammary gland achieved in theimplanted rats is not enough to prevent chemical carcinogen-esis In agreement with our results chronic treatment withperphenazine producing hyperprolactinemia and high levelsof P4 did not protect the mammary gland against chemical
induction of tumors [30]It is worth noting that in our model mammary 120573-
casein content paralleled the alveolar area measured in thehistological preparations suggesting that the proliferationof epithelial tissue of the mammary gland is accompaniedof certain differentiation This result is in agreement with aprevious study in normal Wistar rats [26]
Interestingly the percentage of mammary tumors inregression was greatly affected by PL indicating that theclassical protective effect of gestation described by othersmaybe due at least in part to an increased number of tumorsundergoing regression rather than a decrease of the develop-ment of mammary tumors Our overall results suggest that acycle of PL has ldquotoxicrdquo effects on themammary tumors whichcannot be explained only by the milieu of mammogenic
hormones since we did not find any statistically significantdifference on serum GH (results not shown) or PRL valuesbetween nulliparous or primiparous rats On the contrarythe significantly lower content of 120573-casein in the mammarygland from rats that had regression of their tumors constitutesan interesting support of the idea that involution of thenormal epithelia can be implied in the regression of tumorsespecially so when the tumors of the rats with pregnancyand lactation retain the capability to synthesize 120573-caseinStudies currently under way in our laboratory are focusing onthe expression of several molecular markers involved in theprocess of involution of the mammary gland after lactationin the regressing tumors
We did not find correlation between expression of ARNmof PRLR and serum PRL levels However mammary glandsfrom PL rats that had developed tumors had increasedexpression of PRLR compared to PL rats free of tumorsThis difference can explain the higher alveolar developmentreached in those rats even when the values of circulating PRLat the moment of sacrifice were similar in both groups Theincreased development of alveolar tissue may cause in turnincreased content of 120573-casein It is worth to note that both I
BioMed Research International 9
(a) (b)
(c)
Figure 7 Representative microphotographs (100x) of HampE-stained normal tumors from nulliparous rats (a) hyperprolactinemic rats (b)and primiparous rats showing a noticeable area of necrosis (c) Arrows show necrotic areas
and PL rats had augmented alveolar development comparedto Null rats but the level of differentiation in terms of 120573-casein expression is higher in PL rats supporting a role for theextent of differentiation more than the alveolar developmentin the protective effect of PL
To assess whether the transformation of the mammarycells involves changes in the expression of receptors westudied in parallel the tumors and tissue from the con-tralateral normal mammary gland in the same animals Theexpression of ER120572 increased in tumors of hyperprolactinemicrats compared to the contralateral mammary gland but itwas unmodified in Null or PL rats (results not-shown) Theability of PRL to stimulate the expression of both types of ERhas been shown in the rat corpus luteum in the mammarygland and in the decidua (see [31] for a review) Previousreports suggest that PRL can stimulate ER expression in somebreast cancers [32 33] Moreover PRL can activate ER120572 evenin the absence of estrogenic ligands leading to oncogenesis[34] In our study the expression of ER120573 decreased in thetumors from the three groups compared to their respectivemammary glands (not shown) suggesting a protective rolefor ER120573 This result supports previous studies showing adecrease in the expression of ER120573 in the process of humanbreast cancer progression associatedwith poor differentiation[35] Moreover the expression of ER120572was higher in untrans-formed mammary tissue from Null or PL rats that developedmammary tumors compared with those that did not On thecontrary hyperprolactinemic rats that developed tumors had
lower expression of ER120572 in their mammary tissue This par-ticular result may suggest that when supraphysiological levelsof circulating PRL are constantly acting on the mammarygland tumors can progress even with lower expression ofER120572
In conclusion in our model of carcinogenesis using OFAhrhr rats we showed that serum PRL levels do not seem tohave relevance on the incidence of tumors probably becausethe relative low levels of PRL in those rats are not furtherdecreased by pregnancy and lactation like in other strainsEven though supraphysiological levels of serum PRL mayaffect carcinogenesis
On the other hand PL induces regression of the tumorsmost probably due to the degree of differentiation producedon the mammary cells than to the extent of proliferationreached by the epithelial tissue Alterations in the expressionof hormonal receptors may be involved in both progressionand regression of the tumors
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are deeply indebted to Mrs Elina Guinazu deDi Nasso and Mr Juan Rosales for their excellent technical
10 BioMed Research International
assistance This work was partially supported by grantsfrom Instituto Nacional del Cancer (Ministerio de SaludArgentina) and fromCONICET (Consejo Nacional de Inves-tigaciones Cientıficas y Tecnicas) Argentina
References
[1] P K Chakravarty and D K Sinha ldquoPregnancy inducedmammary tumor specific effector cells are present long afterparturition in a breast cancer model in ratsrdquo Cancer Letters vol154 no 1 pp 1ndash7 2000
[2] J Russo G A Balogh and I H Russo ldquoFull-term pregnancyinduces a specific genomic signature in the human breastrdquoCancer Epidemiology Biomarkers amp Prevention vol 17 no 1 pp51ndash66 2008
[3] J Russo L K Tay and I H Russo ldquoDifferentiation of themammary gland and susceptibility to carcinogenesisrdquo BreastCancer Research and Treatment vol 2 no 1 pp 5ndash73 1982
[4] OMaller KCHansen T R Lyons et al ldquoCollagen architecturein pregnancy-induced protection from breast cancerrdquo Journal ofCell Science vol 126 part 18 pp 4108ndash4110 2013
[5] S K Siwko J Dong M T Lewis H Liu S G Hilsenbeckand Y Li ldquoEvidence that an early pregnancy causes a persistentdecrease in the number of functional mammary epithelial stemcellsmdashimplications for pregnancy-induced protection againstbreast cancerrdquo Stem Cells vol 26 no 12 pp 3205ndash3209 2008
[6] Y Misra P A Bentley J P Bond S Tighe T Hunter and FZhao ldquoMammary gland morphological and gene expressionchanges underlying pregnancy protection of breast cancertumorigenesisrdquo Physiological Genomics vol 44 no 1 pp 76ndash882012
[7] J Russo and I H Russo ldquoInfluence of differentiation and cellkinetics on the susceptibility of the rat mammary gland tocarcinogenesisrdquo Cancer Research vol 40 no 8 pp 2677ndash26871980
[8] R C Guzman J Yang L Rajkumar G Thordarson X Chenand S Nand ldquoHormonal prevention of breast cancer mim-icking the protective effect of pregnancyrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 5 pp 2520ndash2525 1999
[9] A R Bovns R Buchan E N Cole A P M Forrest and KGrifiths ldquoBasal prolactin blood levels in three strains of ratwith differing incidence of 712-Dimethylbenz (a) anthraceneinduced mammary tumoursrdquo European Journal of Cancer vol9 no 3 pp 169ndash171 1973
[10] C W Welsch and H Nagasawa ldquoProlactin and murine mam-mary tumorigenesis a reviewrdquo Cancer Research vol 37 no 4pp 951ndash963 1977
[11] H Cohen I Sabbagh P Guillaumot J Bertrand M Robert-son and H G Friesen ldquoRelationship between bioassay andradioimmunoassay measurement of prolactin in the IPL rat ahypoprolactinemic rat strainrdquo Biology of Reproduction vol 38no 1 pp 84ndash90 1988
[12] S R Valdes A B Penissi R P Deis and G A JahnldquoHormonal profile and reproductive performance in lactationdeficient (OFA hrhr) and normal (Sprague-Dawley) femaleratsrdquo Reproduction vol 133 no 4 pp 827ndash840 2007
[13] H Bazzi A Kljuic A M Christiano and A A PanteleyevldquoIntragenic deletion in theDesmoglein 4 gene underlies the skinphenotype in the Iffa Credo ldquohairlessrdquo ratrdquo Differentiation vol72 no 8 pp 450ndash464 2004
[14] H Kim A A Panteleyev C A B Jahoda Y Ishii and A MChristiano ldquoGenomic organization and analysis of the hairlessgene in four hypotrichotic rat strainsrdquoMammalianGenome vol15 no 12 pp 975ndash981 2004
[15] B Meyer H Bazzi V Zidek et al ldquoA spontaneous mutationin the desmoglein 4 gene underlies hypotrichosis in a newlanceolate hair rat modelrdquo Differentiation vol 72 no 9-10 pp541ndash547 2004
[16] R W Caron and R P Deis ldquoParticipation of opioid and sero-toninergic systems in prolactin secretion induced by hypotha-lamic action of estradiolrdquoNeuroendocrinology vol 64 no 2 pp124ndash130 1996
[17] R W Caron and R P Deis ldquoEstradiol implants in the arcuatenucleus induce lactogenesis in virgin rats Role of progesteronerdquoLife Sciences vol 62 no 3 pp 229ndash237 1997
[18] G R Morel R W Caron G M Console et al ldquoEstrogeninhibits tuberoinfundibular dopaminergic neurons but does notcause irreversible damagerdquo Brain Research Bulletin vol 80 no6 pp 347ndash352 2009
[19] R W Caron A M Salicioni and R P Deis ldquoMifepristonetreatment demonstrates the participation of adrenal glucocorti-coids in the regulation of oestrogen-induced prolactin secretionin ovariectomized ratsrdquo Journal of Steroid Biochemistry andMolecular Biology vol 48 no 4 pp 385ndash389 1994
[20] R P Deis D G Carrizo and G A Jahn ldquoSuckling-inducedprolactin release potentiates mifepristone-induced lactogenesisin pregnant ratsrdquo Journal of Reproduction and Fertility vol 87no 1 pp 147ndash153 1989
[21] R W Caron C A Jahn and R P Deis ldquoLactogenic actionsof different growth hormone preparations in pregnant andlactating ratsrdquo Journal of Endocrinology vol 142 no 3 pp 535ndash545 1994
[22] M Edery L Houdebine J Djiane and P A Kelly ldquoStudies of 120573-casein content of normal and neoplastic rat mammary tissuesby a homologous radioimmunoassayrdquo Molecular and CellularEndocrinology vol 34 no 2 pp 145ndash151 1984
[23] L E Bussmann and R P Deis ldquo120574-Glutamyltransferase activityin mammary gland of pregnant rats and its regulation by ovar-ian hormones prolactin and placental lactogenrdquo BiochemicalJournal vol 223 no 1 pp 275ndash277 1984
[24] J Russo and I H Russo ldquoAtlas and histologic classification oftumors of the rat mammary glandrdquo Journal of Mammary GlandBiology and Neoplasia vol 5 no 2 pp 187ndash200 2000
[25] C M Lopez-Fontana C V Sasso M E Maselli et al ldquoExperi-mental hypothyroidism increases apoptosis in dimethylbenzan-thracene-inducedmammary tumorsrdquoOncologyReports vol 30no 4 pp 1651ndash1660 2013
[26] C M Lopez-Fontana M E Maselli A M Salicioni and R WCaron ldquoThe inhibitory effect of progesterone on lactogenesisduring pregnancy is already evident by mid- to late gestation inrodentsrdquo Reproduction Fertility and Development vol 24 no 5pp 704ndash714 2012
[27] C Puissant and L M Houdebine ldquoAn improvement of thesingle-step method of RNA isolation by acid guanidiniumthiocyanate-phenol-chloroform extractionrdquo BioTechniques vol8 no 2 pp 148ndash149 1990
[28] G Thordarson E Jin R C Guzman S M Swanson S Nandiand F Talamantes ldquoRefractoriness to mammary tumorigenesisin parous rats is it caused by persistent changes in the hormonalenvironment or permanent biochemical alterations in themammary epitheliardquo Carcinogenesis vol 16 no 11 pp 2847ndash2853 1995
BioMed Research International 11
[29] G Thordarson K van Horn R C Guzman S Nandi and FTalamantes ldquoParous rats regain high susceptibility to chemi-cally induced mammary cancer after treatment with variousmammotropic hormonesrdquo Carcinogenesis vol 22 no 7 pp1027ndash1033 2001
[30] L Sivaraman O M Conneely D Medina and B W OMalleyldquop53 is a potential mediator of pregnancy and hormone-induced resistance to mammary carcinogenesisrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 98 no 22 pp 12379ndash12384 2001
[31] J Frasor and G Gibori ldquoProlactin regulation of estrogenreceptor expressionrdquo Trends in Endocrinology and Metabolismvol 14 no 3 pp 118ndash123 2003
[32] M Edery W Imagawa L Larson and S Nandi ldquoRegulation ofestrogen and progesterone receptor levels in mouse mammaryepithelial cells grown in serum-free collagen gel culturesrdquoEndocrinology vol 116 no 1 pp 105ndash112 1985
[33] C J Ormandy R E Hall D L Manning et al ldquoCoexpres-sion and cross-regulation of the prolactin receptor and sexsteroid hormone receptors in breast cancerrdquo Journal of ClinicalEndocrinology and Metabolism vol 82 no 11 pp 3692ndash36991997
[34] K A OrsquoLeary F Jallow D E Rugowski et al ldquoProlactinactivates ER120572 in the absence of ligand in female mammarydevelopment and carcinogenesis in vivordquo Endocrinology vol154 pp 4483ndash4492 2013
[35] B W Park K S Kim M K Heo et al ldquoExpression ofestrogen receptor-120573 in normal mammary and tumor tissues isit protective in breast carcinogenesisrdquo Breast Cancer Researchand Treatment vol 80 no 1 pp 79ndash85 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 3
Table 1 Sequence of primers used in the respective PCR and molecular size of the amplification product
RNAm Sense 51015840-31015840 Antisense 31015840-51015840 SizeActin CGTGGGCCGCCCTAGGCACCA TTGGCCTTAGGGTTCAGAGGGG 243ER120572 AATTCTGACAATCGACGCCAG GTGCTTCAACATTCTCCCTCCTC 345ER120573 AAAGCCAAGAGAAACGGTGGGCAT GCCAATCATGTGCACCAGTTCCTT 204PRLR AAAGTATCTTGTCCAGACTCGCTG AGCAGTTCTTCAGACTTGCCCTT 279PR CCCACAGGAGTTTGTCAAGCTC TAACTTCAGACATCATTTCCGG 325GHR GAGGAGGTGAACACCATCTTGGGC ACCACCTGCTGGTGTAATGTC 534
mammary tissue was cut into small pieces and homogenizedin 2mL 50mM sodium phosphate buffer 150mMNaCl 01NaN3 01TritonX-100 pH 76with anUltraturrax homoge-
nizerThe homogenates were centrifuged at 600 g for 30minThe supernatants were used for 120573-casein determination by ahomologous radioimmunoassay according to Edery et al [22]as modified in our laboratory [23] All samples were assayedin duplicate The standard curve of rat 120573-casein was between025 and 512 ngmL and the sample values were calculated permg of tissue
25 Tumor and Mammary Gland Histology A small pieceof tumor and inguinal mammary gland (contralateral tothe tumor) from each rat were processed for histopatho-logic studies Sections of 3ndash5120583m thickness were cut witha microtome and stained with hematoxylin-eosin (HampE)to define the histopathological changes in the mammaryglands and to classify tumors according to published criteria[24 25] Images were taken with a Nikon Eclipse E200Microscope fitted with a digital still camera MicrometricSE Premium (Nikon Corp Japan) under 100x 400x and600x magnifications The quantification of the percentagesof stroma mostly composed by adipocytes and epithelialtissue in the mammary gland was performed by measuringthe area occupied in 8ndash10 fields of each preparation fromall rats using the ImageJ 142q software available at the NIHsite (httprsbinfonihgovij) Each area was expressed as apercentage of the whole field as previously published [25 26]
26 RNA Isolation and RT-PCR Analysis Total RNA fromnormal mammary glands and tumors was extracted usingthe Chomczynski-Sacchi method modified by Puissant andHoudebine [27] to determine the expression of the followinghormone receptors E
2receptor120572 (ER120572) E
2receptor120573 (ER120573)
PRL receptor (PRLR) P4receptor (PR) and GH receptor
(GHR) Ten micrograms of total RNA were reverse tran-scribed at 42∘C using random hexamer primers andMoloneymurine leukemia virus RT (InvitrogenLife TechnologiesBuenos Aires Argentina) in a 20120583L reaction mixture Beforeproceeding with the semiquantitative PCR the conditionswere established for each tissue such that the amplificationof the products was in the exponential phase and the assaywas linear with respect to the amount of input RNA
All reactions were carried out for 30 cycles with thefollowing cyclic parameters 95∘C for 1min 62∘C for 1minand 72∘C for 2min and then terminated with a 5minextension at 72∘C RNA samples were assayed for DNA
contamination by PCR without prior reverse transcriptionThe PCR products were analyzed on 15 agarose gelscontaining 05mgmL ethidium bromide and photographedwith a Polaroid camera Band intensities of RT-PCR productswere quantified using the ImageJ 142q software availableat the NIH site (httprsbinfonihgovij) Relative levels ofmRNA were expressed as the ratio of signal intensity for thetarget genes relative to that for 120573-actin The sequence of theprimers used for each gene and the size of the product ofamplification obtained are shown in Table 1
27 Statistical Analysis Values are given as means plusmn SEMof 11ndash47 animals per group All statistical analysis wasperformed using GraphPad Prism 501 software (GraphPadSoftware Inc CA USA) Differences in the distribution ofvariables between the three studied groups were assessedusing oneway analysis of variance (ANOVA I) or Kruskal-Wallis test depending on the normality of the variablesas evaluated by the Kolmogorov-Smirnov test Two-wayanalysis of variance (ANOVA II) was used for analysis ofdifferences between rats with and without mammary tumorsin the three groups Post hoc comparisons between meanswere made by Bonferronirsquos test or Dunnrsquos Multiple Compar-ison test Studentrsquos t-test was used when only two groupswere compared When variances were not homogeneouslogarithmic transformation of data was applied Incidenceand percentages of mammary epithelial areas were analyzedby chi-square Differences were considered significant if theprobability was 5 or less
3 Results
31 Incidence Progression and Regression of MammaryTumors Tumor incidence (Table 2) multiplicity and latency(not shown) were not statistically different among the threegroups However a nonsignificant tendency to lower valueswas observed in the PL rats Interestingly a significant rise(119875 lt 0001) in the percentage of tumors with regression andwith macroscopic signs of necrosis was evident in the PL rats(Table 2)
32 Serum Prolactin and GH Levels As expected serum PRLconcentration at the end of the experiment was significantlyhigher in I rats (119875 lt 00001) compared to Null or PL(Figure 1(a)) No correlation between serum PRL levels atthe end of the experiment and percentages of regression oftumors was found
4 BioMed Research International
Table 2 Number and percentages of rats with and without mammary tumors in the three groups
Without tumors With tumors Regressed tumors Necrotic tumors Regular tumors Nulliparous 12 30 714 0 0 0 0 30 100Pregnancy-lactation 35 47 573 17 362lowast 11 234lowast 19 404Implanted 11 26 703 0 0 0 0 26 100lowast119875 lt 005 compared to Nulliparous or Implanted rats
Null I PL0
100
200
300
400
500
600
Seru
m P
RL (n
gm
L)
lowastlowastlowast
(a)
Null T Null NT I T I NT PL T PL NT PL RegT0
255075
100
500
600
700
Seru
m P
RL (n
gm
L)
lowastlowastlowast
(b)
Figure 1 Serum prolactin (PRL) levels in DMBA-treated rats at the end of the experiment (a) Nulliparous rats (Null) and rats after a cycle ofpregnancy and lactation (PL) had significantly lower serumPRL than rats implantedwith 17-120573 estradiol in the arcuate nucleus (I) (119875 lt 00001)This group is referred to as hyperprolactinemic rats (b) When classified depending the development (T) absence (NT) or regression (PLRegT) ofmammary tumors the implanted rats developing tumors showed significantly higher levels of serumPRL than the rest of the animals(119875 lt 00001) including implanted rats without tumors
No significant differences in serum PRL levels wereobserved in Null or PL rats that developed mammary tumorsfrom those that did not However a statistically significantincrease (119875 lt 001) was found in I rats that bore mammarytumors compared to those that did not (Figure 1(b))
Circulating GH levels were similar in all the groupsregardless of tumor development (results not shown)
33 Mammary Gland Tissue Differentiation and DevelopmentThe effect of pregnancy and partial lactation on the devel-opment of the mammary gland was evaluated by histolog-ical observation and measurement of the areas occupiedby parenchyma or stroma Figure 2 shows representativemicrophotographs of HampE-stained mammary tissue fromNull rats ((a) and (b)) I rats ((c) and (d)) and PL rats ((e)and (f)) The mammary glands of Null rats had a normalappearance with few ducts surrounded by a small amountof fibrous connective tissue and abundant adipocytes Asimilar image was seen in hyperprolactinemic rats but witha higher development of alveolar structures The mammarytissue from primiparous rats showed a more noticeabledevelopment of lobuloalveoli Figure 2(g) shows that thepercentages of alveolar tissue in mammary glands from ratsthat bore tumors were higher than in those rats that did notdevelop cancer particularly evident in I rats (119875 lt 001) andin PL rats (119875 lt 0001)
34 Mammary Content of 120573-Casein As expected the mam-mary content of 120573-casein an indicative of the degree ofdifferentiation was increased (119875 lt 0001) in I rats comparedto Null rats A significant increase in mammary 120573-caseinwas produced in PL rats (119875 lt 0001 Figure 3(a)) even longafter the pregnancy lactation cycle Moreover when the ratswere grouped according to the development of tumors nosignificant differences in mammary 120573-casein were observedin Null rats with and without mammary tumors However Irats or PL rats had higher levels of 120573-casein (119875 lt 005 and119875 lt 0001 resp) if they had developed mammary tumors(Figure 3(b))
35 Expression of Hormone Receptors in the NontumoralMammary Tissue We next investigated the level of expres-sion of hormone receptors in the normal tissue of themammary gland from the three groups The expression ofPRLR was similar in the three groups However mammarytissue from PL rats bearing tumors had increased (119875 lt 001)content of PRLR mRNA compared to rats free of tumors(Figure 4(a)) No difference in the expression of total PRmRNA was found between the three groups of treatmentsirrespective of the development of tumors (not shown)
The overall expression of ER120572 in the mammary gland ofI rats was similar to that of Null or PL rats (data not shown)However the mammary gland of I rats that had developedtumors had a diminished expression of ER120572 in comparisonto Null or PL rats bearing mammary tumors (119875 lt 0001
BioMed Research International 5
100120583m
(a)
100120583m
(b)
100120583m
(c)
100120583m
(d)
100120583m
(e)
100120583m
(f)
0
20
40
60
Mam
mar
y al
veol
ar ar
ea (
)
lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowast
lowastlowast
Null T Null NT I T I NT PL T PL NT PL RegT
(g)
Figure 2 Hyperprolactinemia or pregnancy and lactation modify the ratio parenchymastroma in the mammary gland Representativemicrophotographs (100x) of HampE-stained normal mammary tissue from nulliparous rats ((a) and (b)) hyperprolactinemic rats ((c) and (d))and primiparous rats ((e) and (f)) with development ((a) (c) and (e)) or absence ((b) (d) and (f)) of mammary tumors (g) Quantificationsof the relative percentages of the alveolar area Values representmeanplusmn SEM of 8ndash10 fields of each preparation from 11ndash26 animals per grouplowastlowast
119875 lt 0001 lowastlowastlowast119875 lt 00001 comparing the selected groups Comparisons were performed by ANOVA I Arrows show alveolar structures
Figure 4(b)) On the contrary the expression of ER120572 wasincreased in the mammary gland of tumor-bearing Null orPL rats compared to the respective tumor-free rats (119875 lt 005and 119875 lt 001 respectively Figure 4(b))
No significant differences in the expression of ER120573 wereobserved between mammary glands from Null I or PL rats(results not shown) However grouping the rats according tothe presence or not of tumors showed a higher expression ofER120573 only in the Null group (119875 lt 001 Figure 4(b))
36 Changes Related to the Pregnancy and Lactation Cyclein the Mammary Tumors We compared the expression of
120573-casein in the tumors from the three groups to determinethe influence of the different physiological backgrounds onthe changes related to tumor transformation The mammarytumors fromPL rats produced significantly higher levels of120573-casein than the Null or I rats (Figure 5(a)) Moreover 234of the tumors fromprimiparous rats showedmacroscopic andmicroscopic evidence of necrosis (Table 2)
Both PRLR and ER120572 expressions were similar in tumorsdeveloped in Null PL or I rats (results not shown) Howevertumors from I rats showed undetected expression of ER120573 andan increased expression of PR (Figures 5(b) and 5(c))
6 BioMed Research International
Null I PL0
2
4
6
8M
amm
ary120573
-cas
ein
(ng
mg)
lowastlowastlowast
lowastlowastlowast
(a)
Null T Null NT I T I NT PL T PL NT0
2
4
6
8
lowast
lowast
Mam
mar
y120573-c
asei
n (n
gm
g)
(b)
Figure 3 Mammary 120573-casein content at the end of the experiment from DMBA-treated rats (a) Nulliparous rats (Null) had significantlylower mammary 120573-casein than rats implanted with 17-120573 estradiol in the arcuate nucleus (I) or rats after a cycle of pregnancy and lactation(PL) lowastlowastlowast119875 lt 00001 (b) When classified depending on the development (T) or absence (NT) of mammary tumors the implanted andprimiparous rats developing tumors showed significantly higher levels of mammary 120573-casein than the corresponding tumor-free rats lowast119875 lt005 Comparisons were performed by ANOVA I
Null T Null NT I T I NT PL T PL NT00
05
10
PRLR
expr
essio
n
lowastlowast
(a)
00
02
04
06
08
10
ER ex
pres
sion
Null T Null NT I T I NT PL T PL NT
lowastlowastlowast
lowastlowast
lowastlowast
lowast
ER120572ER120573
(b)
Figure 4 Expression of mRNA for hormones receptors in mammary glands from DMBA-treated rats at the end of the experiment (a)Expression of mRNA for PRLR relative to 120573-actin lowastlowast119875 lt 001 with respect to the corresponding tumor-free group (b) Expression of mRNAfor ER120572 and 120573 relative to 120573-actin lowast119875 lt 005 lowastlowast119875 lt 001 lowastlowastlowast119875 lt 00001 with respect to the corresponding tumor-free groups Comparisonswere performed by ANOVA I
37 Changes in the Mammary Gland Related to the Regressionof Tumors In order to investigate whether the increasedpercentage of regressing tumors in the group of primiparousrats is due to long lasting changes in the hormonal milieuor to the grade of differentiation of the mammary gland wecompared primiparous rats bearing tumors with those with-out tumors and rats with regressing tumors No significantdifferences were found between serum PRL concentrationsfrom the PL rats that did not develop tumors those thatbore nonregressed tumors and those that bore regressedtumors (not shown) However a decrease (119875 lt 001) in themammary content of120573-caseinwas evident in the group of ratsthat developed mammary tumors and had tumor regression
(Figure 6(a)) suggesting a link between regression of thetumors and involution of the mammary gland
PRLR expression was significantly lower in mammaryglands from rats that did not develop tumors but no differentfrom those that had regression of the tumors (Figure 6(b))A similar decrease was observed in the expression of ER120572(Figure 6(c)) No significant differences were found in mam-mary expression of PR or ER120573 (results not shown) betweenthe three groups
As shown in Table 2 more than 20 of the tumors fromPL rats showed evident areas of necrosis Figure 7 shows threerepresentativemicrophotographs stained with HampE of tumorfrom Null I and PL rats which shows necrosis
BioMed Research International 7
Null I PL0
2
4
120573-C
asei
n in
tum
ors (
ngm
g)
lowastlowast
lowast
(a)
Null I PL00
01
02
03
04
05
ER120573
expr
essio
n
lowast
(b)
Null I PL00
05
10
15
PR ex
pres
sion
lowastlowast
(c)
Figure 5 120573-Casein content and expression of mRNA for hormones receptors in tumors from DMBA-treated rats (a) Tumor tissue fromnulliparous rats (Null) contains significantly lower amounts of 120573-casein than tumors from implanted (I) or primiparous (PL) rats (119875 lt 005and 119875 lt 0001 resp) (b)The expression of mRNA of ER120573 is lower in the tumors of I rats than in the other two groups (119875 lt 005) (c) mRNAexpression of PR is significantly higher in I rats with respect to Null or PL rats (119875 lt 001)
4 Discussion
To study the involvement of lactogenic hormones in mam-mary carcinogenesis we compared our Null or PL rats withvirgin rats made hyperprolactinemic through hypothalamicimplants of estradiol In this model serum estradiol remainsat a physiological level [16ndash18] allowing us to investigatethe effects of PRL excluding the well-known actions of highamounts of E
2on mammary carcinogenesis
A correlation between serum PRL levels and susceptibil-ity of various rat strains to chemically induced mammarycarcinogenesis has been suggested [9] Both GH and PRLhave been previously shown to be reduced after parturition ascomparedwith nulliparous age-matched animals [28]More-over parous rats showing almost complete refractorinessto chemical carcinogenesis acquire high susceptibility afterhormonal treatment that increases serum GH and PRL levels[29] In our OFA rats neither serum GH nor circulating PRLdecreased after PL Accordingly even when the incidenceof mammary tumors was slightly lower in primiparousrats the difference respect to nulliparae was not statisticallysignificant suggesting that the cycle of pregnancylactationfailed to prevent tumor development in OFA hrhr rats
This different behavior from the original strain (Sprague-Dawley) [28 29] regarding the lack of protection induced bypregnancy and lactation might be due to the lower levels ofcirculating PRL during lactation in the PLOFA ratsThus thedeficient lactation may have been insufficient to protect themammary tissue from tumoral transformation Moreoverdespite the persistent hyperprolactinaemia produced in I ratsthe incidence of tumors was similar to the other groupssuggesting a null impact of serum PRL levels on mammarycarcinogenesis Interestingly in I rats serum PRL was signif-icantly higher in those rats that developed tumors suggestingthat supraphysiological levels of PRL may be relevant inincreasing the risk of mammary cancer
Our results support the idea that the mammary devel-opment per se is not enough to account for the protectiveeffect of parity In fact the implant of estradiol in the arcuatenucleus produces hyperprolactinemia and luteal phase withhigh levels of circulating P
4and mammary development and
differentiation similar to pregnancy [16 17]Thus a protectiveeffect of the implants dependent on the differentiation of themammary gland could be expected However the incidenceof mammary tumors in the I group was similar to that in the
8 BioMed Research International
PL T PL NT PL RegT0
2
4
6
8
Mam
mar
y120573
-cas
ein
(ng
mg)
lowastlowast
(a)
PL T PL NT PL RegT00
05
10
PRL
R ex
pres
sion lowast
(b)
PL T PL NT PL RegT00
05
10
ER120572
expr
essio
n lowastlowast
(c)
Figure 6 120573-Casein content and expression of mRNA for hormones receptors in mammary glands from primiparous rats (PL) Mammary120573-casein content was lower (119875 lt 001) in mammary glands from PL rats that had tumor regression (PLRegT) than in those that did not havetumors (PLNT) or did have no regressing tumors (PLT) (b) and (c) PRLR and ER120572 expressions were lower (119875 lt 005 and 119875 lt 001 resp) inmammary glands from tumor-free PL rats compared to the other two groups
Null rats suggesting that the sustained hyperprolactinaemiais not enough to increase carcinogenesis and moreoverthe development of the mammary gland achieved in theimplanted rats is not enough to prevent chemical carcinogen-esis In agreement with our results chronic treatment withperphenazine producing hyperprolactinemia and high levelsof P4 did not protect the mammary gland against chemical
induction of tumors [30]It is worth noting that in our model mammary 120573-
casein content paralleled the alveolar area measured in thehistological preparations suggesting that the proliferationof epithelial tissue of the mammary gland is accompaniedof certain differentiation This result is in agreement with aprevious study in normal Wistar rats [26]
Interestingly the percentage of mammary tumors inregression was greatly affected by PL indicating that theclassical protective effect of gestation described by othersmaybe due at least in part to an increased number of tumorsundergoing regression rather than a decrease of the develop-ment of mammary tumors Our overall results suggest that acycle of PL has ldquotoxicrdquo effects on themammary tumors whichcannot be explained only by the milieu of mammogenic
hormones since we did not find any statistically significantdifference on serum GH (results not shown) or PRL valuesbetween nulliparous or primiparous rats On the contrarythe significantly lower content of 120573-casein in the mammarygland from rats that had regression of their tumors constitutesan interesting support of the idea that involution of thenormal epithelia can be implied in the regression of tumorsespecially so when the tumors of the rats with pregnancyand lactation retain the capability to synthesize 120573-caseinStudies currently under way in our laboratory are focusing onthe expression of several molecular markers involved in theprocess of involution of the mammary gland after lactationin the regressing tumors
We did not find correlation between expression of ARNmof PRLR and serum PRL levels However mammary glandsfrom PL rats that had developed tumors had increasedexpression of PRLR compared to PL rats free of tumorsThis difference can explain the higher alveolar developmentreached in those rats even when the values of circulating PRLat the moment of sacrifice were similar in both groups Theincreased development of alveolar tissue may cause in turnincreased content of 120573-casein It is worth to note that both I
BioMed Research International 9
(a) (b)
(c)
Figure 7 Representative microphotographs (100x) of HampE-stained normal tumors from nulliparous rats (a) hyperprolactinemic rats (b)and primiparous rats showing a noticeable area of necrosis (c) Arrows show necrotic areas
and PL rats had augmented alveolar development comparedto Null rats but the level of differentiation in terms of 120573-casein expression is higher in PL rats supporting a role for theextent of differentiation more than the alveolar developmentin the protective effect of PL
To assess whether the transformation of the mammarycells involves changes in the expression of receptors westudied in parallel the tumors and tissue from the con-tralateral normal mammary gland in the same animals Theexpression of ER120572 increased in tumors of hyperprolactinemicrats compared to the contralateral mammary gland but itwas unmodified in Null or PL rats (results not-shown) Theability of PRL to stimulate the expression of both types of ERhas been shown in the rat corpus luteum in the mammarygland and in the decidua (see [31] for a review) Previousreports suggest that PRL can stimulate ER expression in somebreast cancers [32 33] Moreover PRL can activate ER120572 evenin the absence of estrogenic ligands leading to oncogenesis[34] In our study the expression of ER120573 decreased in thetumors from the three groups compared to their respectivemammary glands (not shown) suggesting a protective rolefor ER120573 This result supports previous studies showing adecrease in the expression of ER120573 in the process of humanbreast cancer progression associatedwith poor differentiation[35] Moreover the expression of ER120572was higher in untrans-formed mammary tissue from Null or PL rats that developedmammary tumors compared with those that did not On thecontrary hyperprolactinemic rats that developed tumors had
lower expression of ER120572 in their mammary tissue This par-ticular result may suggest that when supraphysiological levelsof circulating PRL are constantly acting on the mammarygland tumors can progress even with lower expression ofER120572
In conclusion in our model of carcinogenesis using OFAhrhr rats we showed that serum PRL levels do not seem tohave relevance on the incidence of tumors probably becausethe relative low levels of PRL in those rats are not furtherdecreased by pregnancy and lactation like in other strainsEven though supraphysiological levels of serum PRL mayaffect carcinogenesis
On the other hand PL induces regression of the tumorsmost probably due to the degree of differentiation producedon the mammary cells than to the extent of proliferationreached by the epithelial tissue Alterations in the expressionof hormonal receptors may be involved in both progressionand regression of the tumors
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are deeply indebted to Mrs Elina Guinazu deDi Nasso and Mr Juan Rosales for their excellent technical
10 BioMed Research International
assistance This work was partially supported by grantsfrom Instituto Nacional del Cancer (Ministerio de SaludArgentina) and fromCONICET (Consejo Nacional de Inves-tigaciones Cientıficas y Tecnicas) Argentina
References
[1] P K Chakravarty and D K Sinha ldquoPregnancy inducedmammary tumor specific effector cells are present long afterparturition in a breast cancer model in ratsrdquo Cancer Letters vol154 no 1 pp 1ndash7 2000
[2] J Russo G A Balogh and I H Russo ldquoFull-term pregnancyinduces a specific genomic signature in the human breastrdquoCancer Epidemiology Biomarkers amp Prevention vol 17 no 1 pp51ndash66 2008
[3] J Russo L K Tay and I H Russo ldquoDifferentiation of themammary gland and susceptibility to carcinogenesisrdquo BreastCancer Research and Treatment vol 2 no 1 pp 5ndash73 1982
[4] OMaller KCHansen T R Lyons et al ldquoCollagen architecturein pregnancy-induced protection from breast cancerrdquo Journal ofCell Science vol 126 part 18 pp 4108ndash4110 2013
[5] S K Siwko J Dong M T Lewis H Liu S G Hilsenbeckand Y Li ldquoEvidence that an early pregnancy causes a persistentdecrease in the number of functional mammary epithelial stemcellsmdashimplications for pregnancy-induced protection againstbreast cancerrdquo Stem Cells vol 26 no 12 pp 3205ndash3209 2008
[6] Y Misra P A Bentley J P Bond S Tighe T Hunter and FZhao ldquoMammary gland morphological and gene expressionchanges underlying pregnancy protection of breast cancertumorigenesisrdquo Physiological Genomics vol 44 no 1 pp 76ndash882012
[7] J Russo and I H Russo ldquoInfluence of differentiation and cellkinetics on the susceptibility of the rat mammary gland tocarcinogenesisrdquo Cancer Research vol 40 no 8 pp 2677ndash26871980
[8] R C Guzman J Yang L Rajkumar G Thordarson X Chenand S Nand ldquoHormonal prevention of breast cancer mim-icking the protective effect of pregnancyrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 5 pp 2520ndash2525 1999
[9] A R Bovns R Buchan E N Cole A P M Forrest and KGrifiths ldquoBasal prolactin blood levels in three strains of ratwith differing incidence of 712-Dimethylbenz (a) anthraceneinduced mammary tumoursrdquo European Journal of Cancer vol9 no 3 pp 169ndash171 1973
[10] C W Welsch and H Nagasawa ldquoProlactin and murine mam-mary tumorigenesis a reviewrdquo Cancer Research vol 37 no 4pp 951ndash963 1977
[11] H Cohen I Sabbagh P Guillaumot J Bertrand M Robert-son and H G Friesen ldquoRelationship between bioassay andradioimmunoassay measurement of prolactin in the IPL rat ahypoprolactinemic rat strainrdquo Biology of Reproduction vol 38no 1 pp 84ndash90 1988
[12] S R Valdes A B Penissi R P Deis and G A JahnldquoHormonal profile and reproductive performance in lactationdeficient (OFA hrhr) and normal (Sprague-Dawley) femaleratsrdquo Reproduction vol 133 no 4 pp 827ndash840 2007
[13] H Bazzi A Kljuic A M Christiano and A A PanteleyevldquoIntragenic deletion in theDesmoglein 4 gene underlies the skinphenotype in the Iffa Credo ldquohairlessrdquo ratrdquo Differentiation vol72 no 8 pp 450ndash464 2004
[14] H Kim A A Panteleyev C A B Jahoda Y Ishii and A MChristiano ldquoGenomic organization and analysis of the hairlessgene in four hypotrichotic rat strainsrdquoMammalianGenome vol15 no 12 pp 975ndash981 2004
[15] B Meyer H Bazzi V Zidek et al ldquoA spontaneous mutationin the desmoglein 4 gene underlies hypotrichosis in a newlanceolate hair rat modelrdquo Differentiation vol 72 no 9-10 pp541ndash547 2004
[16] R W Caron and R P Deis ldquoParticipation of opioid and sero-toninergic systems in prolactin secretion induced by hypotha-lamic action of estradiolrdquoNeuroendocrinology vol 64 no 2 pp124ndash130 1996
[17] R W Caron and R P Deis ldquoEstradiol implants in the arcuatenucleus induce lactogenesis in virgin rats Role of progesteronerdquoLife Sciences vol 62 no 3 pp 229ndash237 1997
[18] G R Morel R W Caron G M Console et al ldquoEstrogeninhibits tuberoinfundibular dopaminergic neurons but does notcause irreversible damagerdquo Brain Research Bulletin vol 80 no6 pp 347ndash352 2009
[19] R W Caron A M Salicioni and R P Deis ldquoMifepristonetreatment demonstrates the participation of adrenal glucocorti-coids in the regulation of oestrogen-induced prolactin secretionin ovariectomized ratsrdquo Journal of Steroid Biochemistry andMolecular Biology vol 48 no 4 pp 385ndash389 1994
[20] R P Deis D G Carrizo and G A Jahn ldquoSuckling-inducedprolactin release potentiates mifepristone-induced lactogenesisin pregnant ratsrdquo Journal of Reproduction and Fertility vol 87no 1 pp 147ndash153 1989
[21] R W Caron C A Jahn and R P Deis ldquoLactogenic actionsof different growth hormone preparations in pregnant andlactating ratsrdquo Journal of Endocrinology vol 142 no 3 pp 535ndash545 1994
[22] M Edery L Houdebine J Djiane and P A Kelly ldquoStudies of 120573-casein content of normal and neoplastic rat mammary tissuesby a homologous radioimmunoassayrdquo Molecular and CellularEndocrinology vol 34 no 2 pp 145ndash151 1984
[23] L E Bussmann and R P Deis ldquo120574-Glutamyltransferase activityin mammary gland of pregnant rats and its regulation by ovar-ian hormones prolactin and placental lactogenrdquo BiochemicalJournal vol 223 no 1 pp 275ndash277 1984
[24] J Russo and I H Russo ldquoAtlas and histologic classification oftumors of the rat mammary glandrdquo Journal of Mammary GlandBiology and Neoplasia vol 5 no 2 pp 187ndash200 2000
[25] C M Lopez-Fontana C V Sasso M E Maselli et al ldquoExperi-mental hypothyroidism increases apoptosis in dimethylbenzan-thracene-inducedmammary tumorsrdquoOncologyReports vol 30no 4 pp 1651ndash1660 2013
[26] C M Lopez-Fontana M E Maselli A M Salicioni and R WCaron ldquoThe inhibitory effect of progesterone on lactogenesisduring pregnancy is already evident by mid- to late gestation inrodentsrdquo Reproduction Fertility and Development vol 24 no 5pp 704ndash714 2012
[27] C Puissant and L M Houdebine ldquoAn improvement of thesingle-step method of RNA isolation by acid guanidiniumthiocyanate-phenol-chloroform extractionrdquo BioTechniques vol8 no 2 pp 148ndash149 1990
[28] G Thordarson E Jin R C Guzman S M Swanson S Nandiand F Talamantes ldquoRefractoriness to mammary tumorigenesisin parous rats is it caused by persistent changes in the hormonalenvironment or permanent biochemical alterations in themammary epitheliardquo Carcinogenesis vol 16 no 11 pp 2847ndash2853 1995
BioMed Research International 11
[29] G Thordarson K van Horn R C Guzman S Nandi and FTalamantes ldquoParous rats regain high susceptibility to chemi-cally induced mammary cancer after treatment with variousmammotropic hormonesrdquo Carcinogenesis vol 22 no 7 pp1027ndash1033 2001
[30] L Sivaraman O M Conneely D Medina and B W OMalleyldquop53 is a potential mediator of pregnancy and hormone-induced resistance to mammary carcinogenesisrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 98 no 22 pp 12379ndash12384 2001
[31] J Frasor and G Gibori ldquoProlactin regulation of estrogenreceptor expressionrdquo Trends in Endocrinology and Metabolismvol 14 no 3 pp 118ndash123 2003
[32] M Edery W Imagawa L Larson and S Nandi ldquoRegulation ofestrogen and progesterone receptor levels in mouse mammaryepithelial cells grown in serum-free collagen gel culturesrdquoEndocrinology vol 116 no 1 pp 105ndash112 1985
[33] C J Ormandy R E Hall D L Manning et al ldquoCoexpres-sion and cross-regulation of the prolactin receptor and sexsteroid hormone receptors in breast cancerrdquo Journal of ClinicalEndocrinology and Metabolism vol 82 no 11 pp 3692ndash36991997
[34] K A OrsquoLeary F Jallow D E Rugowski et al ldquoProlactinactivates ER120572 in the absence of ligand in female mammarydevelopment and carcinogenesis in vivordquo Endocrinology vol154 pp 4483ndash4492 2013
[35] B W Park K S Kim M K Heo et al ldquoExpression ofestrogen receptor-120573 in normal mammary and tumor tissues isit protective in breast carcinogenesisrdquo Breast Cancer Researchand Treatment vol 80 no 1 pp 79ndash85 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
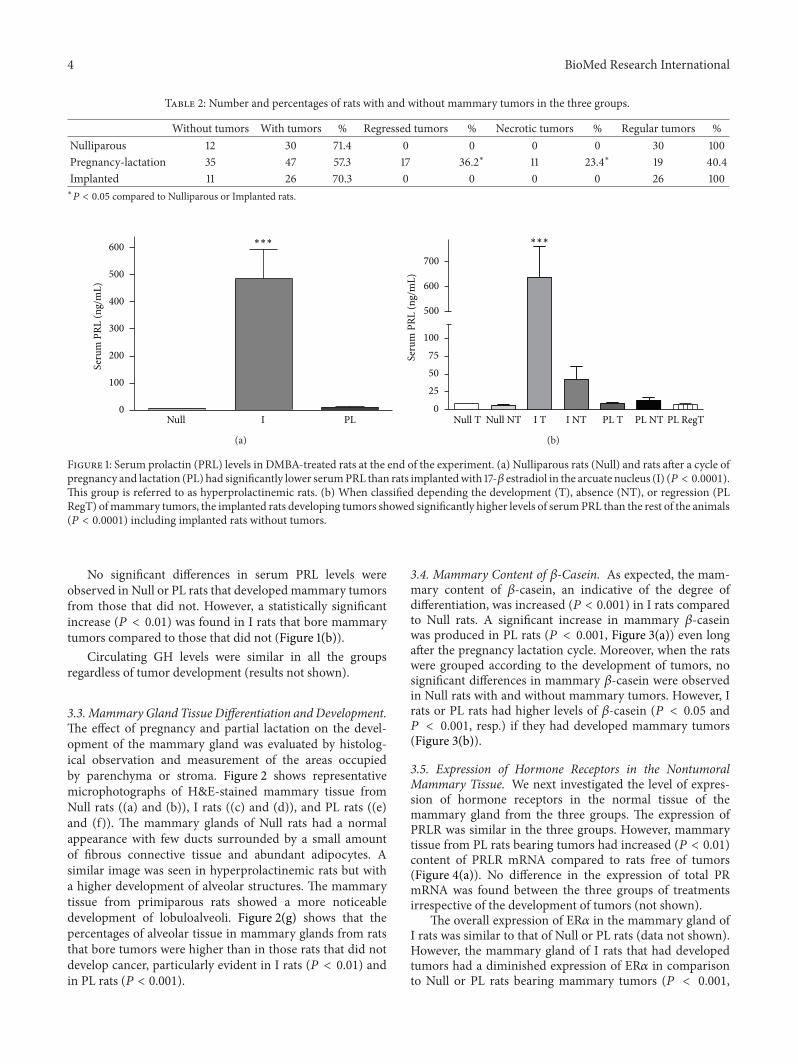
4 BioMed Research International
Table 2 Number and percentages of rats with and without mammary tumors in the three groups
Without tumors With tumors Regressed tumors Necrotic tumors Regular tumors Nulliparous 12 30 714 0 0 0 0 30 100Pregnancy-lactation 35 47 573 17 362lowast 11 234lowast 19 404Implanted 11 26 703 0 0 0 0 26 100lowast119875 lt 005 compared to Nulliparous or Implanted rats
Null I PL0
100
200
300
400
500
600
Seru
m P
RL (n
gm
L)
lowastlowastlowast
(a)
Null T Null NT I T I NT PL T PL NT PL RegT0
255075
100
500
600
700
Seru
m P
RL (n
gm
L)
lowastlowastlowast
(b)
Figure 1 Serum prolactin (PRL) levels in DMBA-treated rats at the end of the experiment (a) Nulliparous rats (Null) and rats after a cycle ofpregnancy and lactation (PL) had significantly lower serumPRL than rats implantedwith 17-120573 estradiol in the arcuate nucleus (I) (119875 lt 00001)This group is referred to as hyperprolactinemic rats (b) When classified depending the development (T) absence (NT) or regression (PLRegT) ofmammary tumors the implanted rats developing tumors showed significantly higher levels of serumPRL than the rest of the animals(119875 lt 00001) including implanted rats without tumors
No significant differences in serum PRL levels wereobserved in Null or PL rats that developed mammary tumorsfrom those that did not However a statistically significantincrease (119875 lt 001) was found in I rats that bore mammarytumors compared to those that did not (Figure 1(b))
Circulating GH levels were similar in all the groupsregardless of tumor development (results not shown)
33 Mammary Gland Tissue Differentiation and DevelopmentThe effect of pregnancy and partial lactation on the devel-opment of the mammary gland was evaluated by histolog-ical observation and measurement of the areas occupiedby parenchyma or stroma Figure 2 shows representativemicrophotographs of HampE-stained mammary tissue fromNull rats ((a) and (b)) I rats ((c) and (d)) and PL rats ((e)and (f)) The mammary glands of Null rats had a normalappearance with few ducts surrounded by a small amountof fibrous connective tissue and abundant adipocytes Asimilar image was seen in hyperprolactinemic rats but witha higher development of alveolar structures The mammarytissue from primiparous rats showed a more noticeabledevelopment of lobuloalveoli Figure 2(g) shows that thepercentages of alveolar tissue in mammary glands from ratsthat bore tumors were higher than in those rats that did notdevelop cancer particularly evident in I rats (119875 lt 001) andin PL rats (119875 lt 0001)
34 Mammary Content of 120573-Casein As expected the mam-mary content of 120573-casein an indicative of the degree ofdifferentiation was increased (119875 lt 0001) in I rats comparedto Null rats A significant increase in mammary 120573-caseinwas produced in PL rats (119875 lt 0001 Figure 3(a)) even longafter the pregnancy lactation cycle Moreover when the ratswere grouped according to the development of tumors nosignificant differences in mammary 120573-casein were observedin Null rats with and without mammary tumors However Irats or PL rats had higher levels of 120573-casein (119875 lt 005 and119875 lt 0001 resp) if they had developed mammary tumors(Figure 3(b))
35 Expression of Hormone Receptors in the NontumoralMammary Tissue We next investigated the level of expres-sion of hormone receptors in the normal tissue of themammary gland from the three groups The expression ofPRLR was similar in the three groups However mammarytissue from PL rats bearing tumors had increased (119875 lt 001)content of PRLR mRNA compared to rats free of tumors(Figure 4(a)) No difference in the expression of total PRmRNA was found between the three groups of treatmentsirrespective of the development of tumors (not shown)
The overall expression of ER120572 in the mammary gland ofI rats was similar to that of Null or PL rats (data not shown)However the mammary gland of I rats that had developedtumors had a diminished expression of ER120572 in comparisonto Null or PL rats bearing mammary tumors (119875 lt 0001
BioMed Research International 5
100120583m
(a)
100120583m
(b)
100120583m
(c)
100120583m
(d)
100120583m
(e)
100120583m
(f)
0
20
40
60
Mam
mar
y al
veol
ar ar
ea (
)
lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowast
lowastlowast
Null T Null NT I T I NT PL T PL NT PL RegT
(g)
Figure 2 Hyperprolactinemia or pregnancy and lactation modify the ratio parenchymastroma in the mammary gland Representativemicrophotographs (100x) of HampE-stained normal mammary tissue from nulliparous rats ((a) and (b)) hyperprolactinemic rats ((c) and (d))and primiparous rats ((e) and (f)) with development ((a) (c) and (e)) or absence ((b) (d) and (f)) of mammary tumors (g) Quantificationsof the relative percentages of the alveolar area Values representmeanplusmn SEM of 8ndash10 fields of each preparation from 11ndash26 animals per grouplowastlowast
119875 lt 0001 lowastlowastlowast119875 lt 00001 comparing the selected groups Comparisons were performed by ANOVA I Arrows show alveolar structures
Figure 4(b)) On the contrary the expression of ER120572 wasincreased in the mammary gland of tumor-bearing Null orPL rats compared to the respective tumor-free rats (119875 lt 005and 119875 lt 001 respectively Figure 4(b))
No significant differences in the expression of ER120573 wereobserved between mammary glands from Null I or PL rats(results not shown) However grouping the rats according tothe presence or not of tumors showed a higher expression ofER120573 only in the Null group (119875 lt 001 Figure 4(b))
36 Changes Related to the Pregnancy and Lactation Cyclein the Mammary Tumors We compared the expression of
120573-casein in the tumors from the three groups to determinethe influence of the different physiological backgrounds onthe changes related to tumor transformation The mammarytumors fromPL rats produced significantly higher levels of120573-casein than the Null or I rats (Figure 5(a)) Moreover 234of the tumors fromprimiparous rats showedmacroscopic andmicroscopic evidence of necrosis (Table 2)
Both PRLR and ER120572 expressions were similar in tumorsdeveloped in Null PL or I rats (results not shown) Howevertumors from I rats showed undetected expression of ER120573 andan increased expression of PR (Figures 5(b) and 5(c))
6 BioMed Research International
Null I PL0
2
4
6
8M
amm
ary120573
-cas
ein
(ng
mg)
lowastlowastlowast
lowastlowastlowast
(a)
Null T Null NT I T I NT PL T PL NT0
2
4
6
8
lowast
lowast
Mam
mar
y120573-c
asei
n (n
gm
g)
(b)
Figure 3 Mammary 120573-casein content at the end of the experiment from DMBA-treated rats (a) Nulliparous rats (Null) had significantlylower mammary 120573-casein than rats implanted with 17-120573 estradiol in the arcuate nucleus (I) or rats after a cycle of pregnancy and lactation(PL) lowastlowastlowast119875 lt 00001 (b) When classified depending on the development (T) or absence (NT) of mammary tumors the implanted andprimiparous rats developing tumors showed significantly higher levels of mammary 120573-casein than the corresponding tumor-free rats lowast119875 lt005 Comparisons were performed by ANOVA I
Null T Null NT I T I NT PL T PL NT00
05
10
PRLR
expr
essio
n
lowastlowast
(a)
00
02
04
06
08
10
ER ex
pres
sion
Null T Null NT I T I NT PL T PL NT
lowastlowastlowast
lowastlowast
lowastlowast
lowast
ER120572ER120573
(b)
Figure 4 Expression of mRNA for hormones receptors in mammary glands from DMBA-treated rats at the end of the experiment (a)Expression of mRNA for PRLR relative to 120573-actin lowastlowast119875 lt 001 with respect to the corresponding tumor-free group (b) Expression of mRNAfor ER120572 and 120573 relative to 120573-actin lowast119875 lt 005 lowastlowast119875 lt 001 lowastlowastlowast119875 lt 00001 with respect to the corresponding tumor-free groups Comparisonswere performed by ANOVA I
37 Changes in the Mammary Gland Related to the Regressionof Tumors In order to investigate whether the increasedpercentage of regressing tumors in the group of primiparousrats is due to long lasting changes in the hormonal milieuor to the grade of differentiation of the mammary gland wecompared primiparous rats bearing tumors with those with-out tumors and rats with regressing tumors No significantdifferences were found between serum PRL concentrationsfrom the PL rats that did not develop tumors those thatbore nonregressed tumors and those that bore regressedtumors (not shown) However a decrease (119875 lt 001) in themammary content of120573-caseinwas evident in the group of ratsthat developed mammary tumors and had tumor regression
(Figure 6(a)) suggesting a link between regression of thetumors and involution of the mammary gland
PRLR expression was significantly lower in mammaryglands from rats that did not develop tumors but no differentfrom those that had regression of the tumors (Figure 6(b))A similar decrease was observed in the expression of ER120572(Figure 6(c)) No significant differences were found in mam-mary expression of PR or ER120573 (results not shown) betweenthe three groups
As shown in Table 2 more than 20 of the tumors fromPL rats showed evident areas of necrosis Figure 7 shows threerepresentativemicrophotographs stained with HampE of tumorfrom Null I and PL rats which shows necrosis
BioMed Research International 7
Null I PL0
2
4
120573-C
asei
n in
tum
ors (
ngm
g)
lowastlowast
lowast
(a)
Null I PL00
01
02
03
04
05
ER120573
expr
essio
n
lowast
(b)
Null I PL00
05
10
15
PR ex
pres
sion
lowastlowast
(c)
Figure 5 120573-Casein content and expression of mRNA for hormones receptors in tumors from DMBA-treated rats (a) Tumor tissue fromnulliparous rats (Null) contains significantly lower amounts of 120573-casein than tumors from implanted (I) or primiparous (PL) rats (119875 lt 005and 119875 lt 0001 resp) (b)The expression of mRNA of ER120573 is lower in the tumors of I rats than in the other two groups (119875 lt 005) (c) mRNAexpression of PR is significantly higher in I rats with respect to Null or PL rats (119875 lt 001)
4 Discussion
To study the involvement of lactogenic hormones in mam-mary carcinogenesis we compared our Null or PL rats withvirgin rats made hyperprolactinemic through hypothalamicimplants of estradiol In this model serum estradiol remainsat a physiological level [16ndash18] allowing us to investigatethe effects of PRL excluding the well-known actions of highamounts of E
2on mammary carcinogenesis
A correlation between serum PRL levels and susceptibil-ity of various rat strains to chemically induced mammarycarcinogenesis has been suggested [9] Both GH and PRLhave been previously shown to be reduced after parturition ascomparedwith nulliparous age-matched animals [28]More-over parous rats showing almost complete refractorinessto chemical carcinogenesis acquire high susceptibility afterhormonal treatment that increases serum GH and PRL levels[29] In our OFA rats neither serum GH nor circulating PRLdecreased after PL Accordingly even when the incidenceof mammary tumors was slightly lower in primiparousrats the difference respect to nulliparae was not statisticallysignificant suggesting that the cycle of pregnancylactationfailed to prevent tumor development in OFA hrhr rats
This different behavior from the original strain (Sprague-Dawley) [28 29] regarding the lack of protection induced bypregnancy and lactation might be due to the lower levels ofcirculating PRL during lactation in the PLOFA ratsThus thedeficient lactation may have been insufficient to protect themammary tissue from tumoral transformation Moreoverdespite the persistent hyperprolactinaemia produced in I ratsthe incidence of tumors was similar to the other groupssuggesting a null impact of serum PRL levels on mammarycarcinogenesis Interestingly in I rats serum PRL was signif-icantly higher in those rats that developed tumors suggestingthat supraphysiological levels of PRL may be relevant inincreasing the risk of mammary cancer
Our results support the idea that the mammary devel-opment per se is not enough to account for the protectiveeffect of parity In fact the implant of estradiol in the arcuatenucleus produces hyperprolactinemia and luteal phase withhigh levels of circulating P
4and mammary development and
differentiation similar to pregnancy [16 17]Thus a protectiveeffect of the implants dependent on the differentiation of themammary gland could be expected However the incidenceof mammary tumors in the I group was similar to that in the
8 BioMed Research International
PL T PL NT PL RegT0
2
4
6
8
Mam
mar
y120573
-cas
ein
(ng
mg)
lowastlowast
(a)
PL T PL NT PL RegT00
05
10
PRL
R ex
pres
sion lowast
(b)
PL T PL NT PL RegT00
05
10
ER120572
expr
essio
n lowastlowast
(c)
Figure 6 120573-Casein content and expression of mRNA for hormones receptors in mammary glands from primiparous rats (PL) Mammary120573-casein content was lower (119875 lt 001) in mammary glands from PL rats that had tumor regression (PLRegT) than in those that did not havetumors (PLNT) or did have no regressing tumors (PLT) (b) and (c) PRLR and ER120572 expressions were lower (119875 lt 005 and 119875 lt 001 resp) inmammary glands from tumor-free PL rats compared to the other two groups
Null rats suggesting that the sustained hyperprolactinaemiais not enough to increase carcinogenesis and moreoverthe development of the mammary gland achieved in theimplanted rats is not enough to prevent chemical carcinogen-esis In agreement with our results chronic treatment withperphenazine producing hyperprolactinemia and high levelsof P4 did not protect the mammary gland against chemical
induction of tumors [30]It is worth noting that in our model mammary 120573-
casein content paralleled the alveolar area measured in thehistological preparations suggesting that the proliferationof epithelial tissue of the mammary gland is accompaniedof certain differentiation This result is in agreement with aprevious study in normal Wistar rats [26]
Interestingly the percentage of mammary tumors inregression was greatly affected by PL indicating that theclassical protective effect of gestation described by othersmaybe due at least in part to an increased number of tumorsundergoing regression rather than a decrease of the develop-ment of mammary tumors Our overall results suggest that acycle of PL has ldquotoxicrdquo effects on themammary tumors whichcannot be explained only by the milieu of mammogenic
hormones since we did not find any statistically significantdifference on serum GH (results not shown) or PRL valuesbetween nulliparous or primiparous rats On the contrarythe significantly lower content of 120573-casein in the mammarygland from rats that had regression of their tumors constitutesan interesting support of the idea that involution of thenormal epithelia can be implied in the regression of tumorsespecially so when the tumors of the rats with pregnancyand lactation retain the capability to synthesize 120573-caseinStudies currently under way in our laboratory are focusing onthe expression of several molecular markers involved in theprocess of involution of the mammary gland after lactationin the regressing tumors
We did not find correlation between expression of ARNmof PRLR and serum PRL levels However mammary glandsfrom PL rats that had developed tumors had increasedexpression of PRLR compared to PL rats free of tumorsThis difference can explain the higher alveolar developmentreached in those rats even when the values of circulating PRLat the moment of sacrifice were similar in both groups Theincreased development of alveolar tissue may cause in turnincreased content of 120573-casein It is worth to note that both I
BioMed Research International 9
(a) (b)
(c)
Figure 7 Representative microphotographs (100x) of HampE-stained normal tumors from nulliparous rats (a) hyperprolactinemic rats (b)and primiparous rats showing a noticeable area of necrosis (c) Arrows show necrotic areas
and PL rats had augmented alveolar development comparedto Null rats but the level of differentiation in terms of 120573-casein expression is higher in PL rats supporting a role for theextent of differentiation more than the alveolar developmentin the protective effect of PL
To assess whether the transformation of the mammarycells involves changes in the expression of receptors westudied in parallel the tumors and tissue from the con-tralateral normal mammary gland in the same animals Theexpression of ER120572 increased in tumors of hyperprolactinemicrats compared to the contralateral mammary gland but itwas unmodified in Null or PL rats (results not-shown) Theability of PRL to stimulate the expression of both types of ERhas been shown in the rat corpus luteum in the mammarygland and in the decidua (see [31] for a review) Previousreports suggest that PRL can stimulate ER expression in somebreast cancers [32 33] Moreover PRL can activate ER120572 evenin the absence of estrogenic ligands leading to oncogenesis[34] In our study the expression of ER120573 decreased in thetumors from the three groups compared to their respectivemammary glands (not shown) suggesting a protective rolefor ER120573 This result supports previous studies showing adecrease in the expression of ER120573 in the process of humanbreast cancer progression associatedwith poor differentiation[35] Moreover the expression of ER120572was higher in untrans-formed mammary tissue from Null or PL rats that developedmammary tumors compared with those that did not On thecontrary hyperprolactinemic rats that developed tumors had
lower expression of ER120572 in their mammary tissue This par-ticular result may suggest that when supraphysiological levelsof circulating PRL are constantly acting on the mammarygland tumors can progress even with lower expression ofER120572
In conclusion in our model of carcinogenesis using OFAhrhr rats we showed that serum PRL levels do not seem tohave relevance on the incidence of tumors probably becausethe relative low levels of PRL in those rats are not furtherdecreased by pregnancy and lactation like in other strainsEven though supraphysiological levels of serum PRL mayaffect carcinogenesis
On the other hand PL induces regression of the tumorsmost probably due to the degree of differentiation producedon the mammary cells than to the extent of proliferationreached by the epithelial tissue Alterations in the expressionof hormonal receptors may be involved in both progressionand regression of the tumors
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are deeply indebted to Mrs Elina Guinazu deDi Nasso and Mr Juan Rosales for their excellent technical
10 BioMed Research International
assistance This work was partially supported by grantsfrom Instituto Nacional del Cancer (Ministerio de SaludArgentina) and fromCONICET (Consejo Nacional de Inves-tigaciones Cientıficas y Tecnicas) Argentina
References
[1] P K Chakravarty and D K Sinha ldquoPregnancy inducedmammary tumor specific effector cells are present long afterparturition in a breast cancer model in ratsrdquo Cancer Letters vol154 no 1 pp 1ndash7 2000
[2] J Russo G A Balogh and I H Russo ldquoFull-term pregnancyinduces a specific genomic signature in the human breastrdquoCancer Epidemiology Biomarkers amp Prevention vol 17 no 1 pp51ndash66 2008
[3] J Russo L K Tay and I H Russo ldquoDifferentiation of themammary gland and susceptibility to carcinogenesisrdquo BreastCancer Research and Treatment vol 2 no 1 pp 5ndash73 1982
[4] OMaller KCHansen T R Lyons et al ldquoCollagen architecturein pregnancy-induced protection from breast cancerrdquo Journal ofCell Science vol 126 part 18 pp 4108ndash4110 2013
[5] S K Siwko J Dong M T Lewis H Liu S G Hilsenbeckand Y Li ldquoEvidence that an early pregnancy causes a persistentdecrease in the number of functional mammary epithelial stemcellsmdashimplications for pregnancy-induced protection againstbreast cancerrdquo Stem Cells vol 26 no 12 pp 3205ndash3209 2008
[6] Y Misra P A Bentley J P Bond S Tighe T Hunter and FZhao ldquoMammary gland morphological and gene expressionchanges underlying pregnancy protection of breast cancertumorigenesisrdquo Physiological Genomics vol 44 no 1 pp 76ndash882012
[7] J Russo and I H Russo ldquoInfluence of differentiation and cellkinetics on the susceptibility of the rat mammary gland tocarcinogenesisrdquo Cancer Research vol 40 no 8 pp 2677ndash26871980
[8] R C Guzman J Yang L Rajkumar G Thordarson X Chenand S Nand ldquoHormonal prevention of breast cancer mim-icking the protective effect of pregnancyrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 5 pp 2520ndash2525 1999
[9] A R Bovns R Buchan E N Cole A P M Forrest and KGrifiths ldquoBasal prolactin blood levels in three strains of ratwith differing incidence of 712-Dimethylbenz (a) anthraceneinduced mammary tumoursrdquo European Journal of Cancer vol9 no 3 pp 169ndash171 1973
[10] C W Welsch and H Nagasawa ldquoProlactin and murine mam-mary tumorigenesis a reviewrdquo Cancer Research vol 37 no 4pp 951ndash963 1977
[11] H Cohen I Sabbagh P Guillaumot J Bertrand M Robert-son and H G Friesen ldquoRelationship between bioassay andradioimmunoassay measurement of prolactin in the IPL rat ahypoprolactinemic rat strainrdquo Biology of Reproduction vol 38no 1 pp 84ndash90 1988
[12] S R Valdes A B Penissi R P Deis and G A JahnldquoHormonal profile and reproductive performance in lactationdeficient (OFA hrhr) and normal (Sprague-Dawley) femaleratsrdquo Reproduction vol 133 no 4 pp 827ndash840 2007
[13] H Bazzi A Kljuic A M Christiano and A A PanteleyevldquoIntragenic deletion in theDesmoglein 4 gene underlies the skinphenotype in the Iffa Credo ldquohairlessrdquo ratrdquo Differentiation vol72 no 8 pp 450ndash464 2004
[14] H Kim A A Panteleyev C A B Jahoda Y Ishii and A MChristiano ldquoGenomic organization and analysis of the hairlessgene in four hypotrichotic rat strainsrdquoMammalianGenome vol15 no 12 pp 975ndash981 2004
[15] B Meyer H Bazzi V Zidek et al ldquoA spontaneous mutationin the desmoglein 4 gene underlies hypotrichosis in a newlanceolate hair rat modelrdquo Differentiation vol 72 no 9-10 pp541ndash547 2004
[16] R W Caron and R P Deis ldquoParticipation of opioid and sero-toninergic systems in prolactin secretion induced by hypotha-lamic action of estradiolrdquoNeuroendocrinology vol 64 no 2 pp124ndash130 1996
[17] R W Caron and R P Deis ldquoEstradiol implants in the arcuatenucleus induce lactogenesis in virgin rats Role of progesteronerdquoLife Sciences vol 62 no 3 pp 229ndash237 1997
[18] G R Morel R W Caron G M Console et al ldquoEstrogeninhibits tuberoinfundibular dopaminergic neurons but does notcause irreversible damagerdquo Brain Research Bulletin vol 80 no6 pp 347ndash352 2009
[19] R W Caron A M Salicioni and R P Deis ldquoMifepristonetreatment demonstrates the participation of adrenal glucocorti-coids in the regulation of oestrogen-induced prolactin secretionin ovariectomized ratsrdquo Journal of Steroid Biochemistry andMolecular Biology vol 48 no 4 pp 385ndash389 1994
[20] R P Deis D G Carrizo and G A Jahn ldquoSuckling-inducedprolactin release potentiates mifepristone-induced lactogenesisin pregnant ratsrdquo Journal of Reproduction and Fertility vol 87no 1 pp 147ndash153 1989
[21] R W Caron C A Jahn and R P Deis ldquoLactogenic actionsof different growth hormone preparations in pregnant andlactating ratsrdquo Journal of Endocrinology vol 142 no 3 pp 535ndash545 1994
[22] M Edery L Houdebine J Djiane and P A Kelly ldquoStudies of 120573-casein content of normal and neoplastic rat mammary tissuesby a homologous radioimmunoassayrdquo Molecular and CellularEndocrinology vol 34 no 2 pp 145ndash151 1984
[23] L E Bussmann and R P Deis ldquo120574-Glutamyltransferase activityin mammary gland of pregnant rats and its regulation by ovar-ian hormones prolactin and placental lactogenrdquo BiochemicalJournal vol 223 no 1 pp 275ndash277 1984
[24] J Russo and I H Russo ldquoAtlas and histologic classification oftumors of the rat mammary glandrdquo Journal of Mammary GlandBiology and Neoplasia vol 5 no 2 pp 187ndash200 2000
[25] C M Lopez-Fontana C V Sasso M E Maselli et al ldquoExperi-mental hypothyroidism increases apoptosis in dimethylbenzan-thracene-inducedmammary tumorsrdquoOncologyReports vol 30no 4 pp 1651ndash1660 2013
[26] C M Lopez-Fontana M E Maselli A M Salicioni and R WCaron ldquoThe inhibitory effect of progesterone on lactogenesisduring pregnancy is already evident by mid- to late gestation inrodentsrdquo Reproduction Fertility and Development vol 24 no 5pp 704ndash714 2012
[27] C Puissant and L M Houdebine ldquoAn improvement of thesingle-step method of RNA isolation by acid guanidiniumthiocyanate-phenol-chloroform extractionrdquo BioTechniques vol8 no 2 pp 148ndash149 1990
[28] G Thordarson E Jin R C Guzman S M Swanson S Nandiand F Talamantes ldquoRefractoriness to mammary tumorigenesisin parous rats is it caused by persistent changes in the hormonalenvironment or permanent biochemical alterations in themammary epitheliardquo Carcinogenesis vol 16 no 11 pp 2847ndash2853 1995
BioMed Research International 11
[29] G Thordarson K van Horn R C Guzman S Nandi and FTalamantes ldquoParous rats regain high susceptibility to chemi-cally induced mammary cancer after treatment with variousmammotropic hormonesrdquo Carcinogenesis vol 22 no 7 pp1027ndash1033 2001
[30] L Sivaraman O M Conneely D Medina and B W OMalleyldquop53 is a potential mediator of pregnancy and hormone-induced resistance to mammary carcinogenesisrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 98 no 22 pp 12379ndash12384 2001
[31] J Frasor and G Gibori ldquoProlactin regulation of estrogenreceptor expressionrdquo Trends in Endocrinology and Metabolismvol 14 no 3 pp 118ndash123 2003
[32] M Edery W Imagawa L Larson and S Nandi ldquoRegulation ofestrogen and progesterone receptor levels in mouse mammaryepithelial cells grown in serum-free collagen gel culturesrdquoEndocrinology vol 116 no 1 pp 105ndash112 1985
[33] C J Ormandy R E Hall D L Manning et al ldquoCoexpres-sion and cross-regulation of the prolactin receptor and sexsteroid hormone receptors in breast cancerrdquo Journal of ClinicalEndocrinology and Metabolism vol 82 no 11 pp 3692ndash36991997
[34] K A OrsquoLeary F Jallow D E Rugowski et al ldquoProlactinactivates ER120572 in the absence of ligand in female mammarydevelopment and carcinogenesis in vivordquo Endocrinology vol154 pp 4483ndash4492 2013
[35] B W Park K S Kim M K Heo et al ldquoExpression ofestrogen receptor-120573 in normal mammary and tumor tissues isit protective in breast carcinogenesisrdquo Breast Cancer Researchand Treatment vol 80 no 1 pp 79ndash85 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 5
100120583m
(a)
100120583m
(b)
100120583m
(c)
100120583m
(d)
100120583m
(e)
100120583m
(f)
0
20
40
60
Mam
mar
y al
veol
ar ar
ea (
)
lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowast
lowastlowast
Null T Null NT I T I NT PL T PL NT PL RegT
(g)
Figure 2 Hyperprolactinemia or pregnancy and lactation modify the ratio parenchymastroma in the mammary gland Representativemicrophotographs (100x) of HampE-stained normal mammary tissue from nulliparous rats ((a) and (b)) hyperprolactinemic rats ((c) and (d))and primiparous rats ((e) and (f)) with development ((a) (c) and (e)) or absence ((b) (d) and (f)) of mammary tumors (g) Quantificationsof the relative percentages of the alveolar area Values representmeanplusmn SEM of 8ndash10 fields of each preparation from 11ndash26 animals per grouplowastlowast
119875 lt 0001 lowastlowastlowast119875 lt 00001 comparing the selected groups Comparisons were performed by ANOVA I Arrows show alveolar structures
Figure 4(b)) On the contrary the expression of ER120572 wasincreased in the mammary gland of tumor-bearing Null orPL rats compared to the respective tumor-free rats (119875 lt 005and 119875 lt 001 respectively Figure 4(b))
No significant differences in the expression of ER120573 wereobserved between mammary glands from Null I or PL rats(results not shown) However grouping the rats according tothe presence or not of tumors showed a higher expression ofER120573 only in the Null group (119875 lt 001 Figure 4(b))
36 Changes Related to the Pregnancy and Lactation Cyclein the Mammary Tumors We compared the expression of
120573-casein in the tumors from the three groups to determinethe influence of the different physiological backgrounds onthe changes related to tumor transformation The mammarytumors fromPL rats produced significantly higher levels of120573-casein than the Null or I rats (Figure 5(a)) Moreover 234of the tumors fromprimiparous rats showedmacroscopic andmicroscopic evidence of necrosis (Table 2)
Both PRLR and ER120572 expressions were similar in tumorsdeveloped in Null PL or I rats (results not shown) Howevertumors from I rats showed undetected expression of ER120573 andan increased expression of PR (Figures 5(b) and 5(c))
6 BioMed Research International
Null I PL0
2
4
6
8M
amm
ary120573
-cas
ein
(ng
mg)
lowastlowastlowast
lowastlowastlowast
(a)
Null T Null NT I T I NT PL T PL NT0
2
4
6
8
lowast
lowast
Mam
mar
y120573-c
asei
n (n
gm
g)
(b)
Figure 3 Mammary 120573-casein content at the end of the experiment from DMBA-treated rats (a) Nulliparous rats (Null) had significantlylower mammary 120573-casein than rats implanted with 17-120573 estradiol in the arcuate nucleus (I) or rats after a cycle of pregnancy and lactation(PL) lowastlowastlowast119875 lt 00001 (b) When classified depending on the development (T) or absence (NT) of mammary tumors the implanted andprimiparous rats developing tumors showed significantly higher levels of mammary 120573-casein than the corresponding tumor-free rats lowast119875 lt005 Comparisons were performed by ANOVA I
Null T Null NT I T I NT PL T PL NT00
05
10
PRLR
expr
essio
n
lowastlowast
(a)
00
02
04
06
08
10
ER ex
pres
sion
Null T Null NT I T I NT PL T PL NT
lowastlowastlowast
lowastlowast
lowastlowast
lowast
ER120572ER120573
(b)
Figure 4 Expression of mRNA for hormones receptors in mammary glands from DMBA-treated rats at the end of the experiment (a)Expression of mRNA for PRLR relative to 120573-actin lowastlowast119875 lt 001 with respect to the corresponding tumor-free group (b) Expression of mRNAfor ER120572 and 120573 relative to 120573-actin lowast119875 lt 005 lowastlowast119875 lt 001 lowastlowastlowast119875 lt 00001 with respect to the corresponding tumor-free groups Comparisonswere performed by ANOVA I
37 Changes in the Mammary Gland Related to the Regressionof Tumors In order to investigate whether the increasedpercentage of regressing tumors in the group of primiparousrats is due to long lasting changes in the hormonal milieuor to the grade of differentiation of the mammary gland wecompared primiparous rats bearing tumors with those with-out tumors and rats with regressing tumors No significantdifferences were found between serum PRL concentrationsfrom the PL rats that did not develop tumors those thatbore nonregressed tumors and those that bore regressedtumors (not shown) However a decrease (119875 lt 001) in themammary content of120573-caseinwas evident in the group of ratsthat developed mammary tumors and had tumor regression
(Figure 6(a)) suggesting a link between regression of thetumors and involution of the mammary gland
PRLR expression was significantly lower in mammaryglands from rats that did not develop tumors but no differentfrom those that had regression of the tumors (Figure 6(b))A similar decrease was observed in the expression of ER120572(Figure 6(c)) No significant differences were found in mam-mary expression of PR or ER120573 (results not shown) betweenthe three groups
As shown in Table 2 more than 20 of the tumors fromPL rats showed evident areas of necrosis Figure 7 shows threerepresentativemicrophotographs stained with HampE of tumorfrom Null I and PL rats which shows necrosis
BioMed Research International 7
Null I PL0
2
4
120573-C
asei
n in
tum
ors (
ngm
g)
lowastlowast
lowast
(a)
Null I PL00
01
02
03
04
05
ER120573
expr
essio
n
lowast
(b)
Null I PL00
05
10
15
PR ex
pres
sion
lowastlowast
(c)
Figure 5 120573-Casein content and expression of mRNA for hormones receptors in tumors from DMBA-treated rats (a) Tumor tissue fromnulliparous rats (Null) contains significantly lower amounts of 120573-casein than tumors from implanted (I) or primiparous (PL) rats (119875 lt 005and 119875 lt 0001 resp) (b)The expression of mRNA of ER120573 is lower in the tumors of I rats than in the other two groups (119875 lt 005) (c) mRNAexpression of PR is significantly higher in I rats with respect to Null or PL rats (119875 lt 001)
4 Discussion
To study the involvement of lactogenic hormones in mam-mary carcinogenesis we compared our Null or PL rats withvirgin rats made hyperprolactinemic through hypothalamicimplants of estradiol In this model serum estradiol remainsat a physiological level [16ndash18] allowing us to investigatethe effects of PRL excluding the well-known actions of highamounts of E
2on mammary carcinogenesis
A correlation between serum PRL levels and susceptibil-ity of various rat strains to chemically induced mammarycarcinogenesis has been suggested [9] Both GH and PRLhave been previously shown to be reduced after parturition ascomparedwith nulliparous age-matched animals [28]More-over parous rats showing almost complete refractorinessto chemical carcinogenesis acquire high susceptibility afterhormonal treatment that increases serum GH and PRL levels[29] In our OFA rats neither serum GH nor circulating PRLdecreased after PL Accordingly even when the incidenceof mammary tumors was slightly lower in primiparousrats the difference respect to nulliparae was not statisticallysignificant suggesting that the cycle of pregnancylactationfailed to prevent tumor development in OFA hrhr rats
This different behavior from the original strain (Sprague-Dawley) [28 29] regarding the lack of protection induced bypregnancy and lactation might be due to the lower levels ofcirculating PRL during lactation in the PLOFA ratsThus thedeficient lactation may have been insufficient to protect themammary tissue from tumoral transformation Moreoverdespite the persistent hyperprolactinaemia produced in I ratsthe incidence of tumors was similar to the other groupssuggesting a null impact of serum PRL levels on mammarycarcinogenesis Interestingly in I rats serum PRL was signif-icantly higher in those rats that developed tumors suggestingthat supraphysiological levels of PRL may be relevant inincreasing the risk of mammary cancer
Our results support the idea that the mammary devel-opment per se is not enough to account for the protectiveeffect of parity In fact the implant of estradiol in the arcuatenucleus produces hyperprolactinemia and luteal phase withhigh levels of circulating P
4and mammary development and
differentiation similar to pregnancy [16 17]Thus a protectiveeffect of the implants dependent on the differentiation of themammary gland could be expected However the incidenceof mammary tumors in the I group was similar to that in the
8 BioMed Research International
PL T PL NT PL RegT0
2
4
6
8
Mam
mar
y120573
-cas
ein
(ng
mg)
lowastlowast
(a)
PL T PL NT PL RegT00
05
10
PRL
R ex
pres
sion lowast
(b)
PL T PL NT PL RegT00
05
10
ER120572
expr
essio
n lowastlowast
(c)
Figure 6 120573-Casein content and expression of mRNA for hormones receptors in mammary glands from primiparous rats (PL) Mammary120573-casein content was lower (119875 lt 001) in mammary glands from PL rats that had tumor regression (PLRegT) than in those that did not havetumors (PLNT) or did have no regressing tumors (PLT) (b) and (c) PRLR and ER120572 expressions were lower (119875 lt 005 and 119875 lt 001 resp) inmammary glands from tumor-free PL rats compared to the other two groups
Null rats suggesting that the sustained hyperprolactinaemiais not enough to increase carcinogenesis and moreoverthe development of the mammary gland achieved in theimplanted rats is not enough to prevent chemical carcinogen-esis In agreement with our results chronic treatment withperphenazine producing hyperprolactinemia and high levelsof P4 did not protect the mammary gland against chemical
induction of tumors [30]It is worth noting that in our model mammary 120573-
casein content paralleled the alveolar area measured in thehistological preparations suggesting that the proliferationof epithelial tissue of the mammary gland is accompaniedof certain differentiation This result is in agreement with aprevious study in normal Wistar rats [26]
Interestingly the percentage of mammary tumors inregression was greatly affected by PL indicating that theclassical protective effect of gestation described by othersmaybe due at least in part to an increased number of tumorsundergoing regression rather than a decrease of the develop-ment of mammary tumors Our overall results suggest that acycle of PL has ldquotoxicrdquo effects on themammary tumors whichcannot be explained only by the milieu of mammogenic
hormones since we did not find any statistically significantdifference on serum GH (results not shown) or PRL valuesbetween nulliparous or primiparous rats On the contrarythe significantly lower content of 120573-casein in the mammarygland from rats that had regression of their tumors constitutesan interesting support of the idea that involution of thenormal epithelia can be implied in the regression of tumorsespecially so when the tumors of the rats with pregnancyand lactation retain the capability to synthesize 120573-caseinStudies currently under way in our laboratory are focusing onthe expression of several molecular markers involved in theprocess of involution of the mammary gland after lactationin the regressing tumors
We did not find correlation between expression of ARNmof PRLR and serum PRL levels However mammary glandsfrom PL rats that had developed tumors had increasedexpression of PRLR compared to PL rats free of tumorsThis difference can explain the higher alveolar developmentreached in those rats even when the values of circulating PRLat the moment of sacrifice were similar in both groups Theincreased development of alveolar tissue may cause in turnincreased content of 120573-casein It is worth to note that both I
BioMed Research International 9
(a) (b)
(c)
Figure 7 Representative microphotographs (100x) of HampE-stained normal tumors from nulliparous rats (a) hyperprolactinemic rats (b)and primiparous rats showing a noticeable area of necrosis (c) Arrows show necrotic areas
and PL rats had augmented alveolar development comparedto Null rats but the level of differentiation in terms of 120573-casein expression is higher in PL rats supporting a role for theextent of differentiation more than the alveolar developmentin the protective effect of PL
To assess whether the transformation of the mammarycells involves changes in the expression of receptors westudied in parallel the tumors and tissue from the con-tralateral normal mammary gland in the same animals Theexpression of ER120572 increased in tumors of hyperprolactinemicrats compared to the contralateral mammary gland but itwas unmodified in Null or PL rats (results not-shown) Theability of PRL to stimulate the expression of both types of ERhas been shown in the rat corpus luteum in the mammarygland and in the decidua (see [31] for a review) Previousreports suggest that PRL can stimulate ER expression in somebreast cancers [32 33] Moreover PRL can activate ER120572 evenin the absence of estrogenic ligands leading to oncogenesis[34] In our study the expression of ER120573 decreased in thetumors from the three groups compared to their respectivemammary glands (not shown) suggesting a protective rolefor ER120573 This result supports previous studies showing adecrease in the expression of ER120573 in the process of humanbreast cancer progression associatedwith poor differentiation[35] Moreover the expression of ER120572was higher in untrans-formed mammary tissue from Null or PL rats that developedmammary tumors compared with those that did not On thecontrary hyperprolactinemic rats that developed tumors had
lower expression of ER120572 in their mammary tissue This par-ticular result may suggest that when supraphysiological levelsof circulating PRL are constantly acting on the mammarygland tumors can progress even with lower expression ofER120572
In conclusion in our model of carcinogenesis using OFAhrhr rats we showed that serum PRL levels do not seem tohave relevance on the incidence of tumors probably becausethe relative low levels of PRL in those rats are not furtherdecreased by pregnancy and lactation like in other strainsEven though supraphysiological levels of serum PRL mayaffect carcinogenesis
On the other hand PL induces regression of the tumorsmost probably due to the degree of differentiation producedon the mammary cells than to the extent of proliferationreached by the epithelial tissue Alterations in the expressionof hormonal receptors may be involved in both progressionand regression of the tumors
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are deeply indebted to Mrs Elina Guinazu deDi Nasso and Mr Juan Rosales for their excellent technical
10 BioMed Research International
assistance This work was partially supported by grantsfrom Instituto Nacional del Cancer (Ministerio de SaludArgentina) and fromCONICET (Consejo Nacional de Inves-tigaciones Cientıficas y Tecnicas) Argentina
References
[1] P K Chakravarty and D K Sinha ldquoPregnancy inducedmammary tumor specific effector cells are present long afterparturition in a breast cancer model in ratsrdquo Cancer Letters vol154 no 1 pp 1ndash7 2000
[2] J Russo G A Balogh and I H Russo ldquoFull-term pregnancyinduces a specific genomic signature in the human breastrdquoCancer Epidemiology Biomarkers amp Prevention vol 17 no 1 pp51ndash66 2008
[3] J Russo L K Tay and I H Russo ldquoDifferentiation of themammary gland and susceptibility to carcinogenesisrdquo BreastCancer Research and Treatment vol 2 no 1 pp 5ndash73 1982
[4] OMaller KCHansen T R Lyons et al ldquoCollagen architecturein pregnancy-induced protection from breast cancerrdquo Journal ofCell Science vol 126 part 18 pp 4108ndash4110 2013
[5] S K Siwko J Dong M T Lewis H Liu S G Hilsenbeckand Y Li ldquoEvidence that an early pregnancy causes a persistentdecrease in the number of functional mammary epithelial stemcellsmdashimplications for pregnancy-induced protection againstbreast cancerrdquo Stem Cells vol 26 no 12 pp 3205ndash3209 2008
[6] Y Misra P A Bentley J P Bond S Tighe T Hunter and FZhao ldquoMammary gland morphological and gene expressionchanges underlying pregnancy protection of breast cancertumorigenesisrdquo Physiological Genomics vol 44 no 1 pp 76ndash882012
[7] J Russo and I H Russo ldquoInfluence of differentiation and cellkinetics on the susceptibility of the rat mammary gland tocarcinogenesisrdquo Cancer Research vol 40 no 8 pp 2677ndash26871980
[8] R C Guzman J Yang L Rajkumar G Thordarson X Chenand S Nand ldquoHormonal prevention of breast cancer mim-icking the protective effect of pregnancyrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 5 pp 2520ndash2525 1999
[9] A R Bovns R Buchan E N Cole A P M Forrest and KGrifiths ldquoBasal prolactin blood levels in three strains of ratwith differing incidence of 712-Dimethylbenz (a) anthraceneinduced mammary tumoursrdquo European Journal of Cancer vol9 no 3 pp 169ndash171 1973
[10] C W Welsch and H Nagasawa ldquoProlactin and murine mam-mary tumorigenesis a reviewrdquo Cancer Research vol 37 no 4pp 951ndash963 1977
[11] H Cohen I Sabbagh P Guillaumot J Bertrand M Robert-son and H G Friesen ldquoRelationship between bioassay andradioimmunoassay measurement of prolactin in the IPL rat ahypoprolactinemic rat strainrdquo Biology of Reproduction vol 38no 1 pp 84ndash90 1988
[12] S R Valdes A B Penissi R P Deis and G A JahnldquoHormonal profile and reproductive performance in lactationdeficient (OFA hrhr) and normal (Sprague-Dawley) femaleratsrdquo Reproduction vol 133 no 4 pp 827ndash840 2007
[13] H Bazzi A Kljuic A M Christiano and A A PanteleyevldquoIntragenic deletion in theDesmoglein 4 gene underlies the skinphenotype in the Iffa Credo ldquohairlessrdquo ratrdquo Differentiation vol72 no 8 pp 450ndash464 2004
[14] H Kim A A Panteleyev C A B Jahoda Y Ishii and A MChristiano ldquoGenomic organization and analysis of the hairlessgene in four hypotrichotic rat strainsrdquoMammalianGenome vol15 no 12 pp 975ndash981 2004
[15] B Meyer H Bazzi V Zidek et al ldquoA spontaneous mutationin the desmoglein 4 gene underlies hypotrichosis in a newlanceolate hair rat modelrdquo Differentiation vol 72 no 9-10 pp541ndash547 2004
[16] R W Caron and R P Deis ldquoParticipation of opioid and sero-toninergic systems in prolactin secretion induced by hypotha-lamic action of estradiolrdquoNeuroendocrinology vol 64 no 2 pp124ndash130 1996
[17] R W Caron and R P Deis ldquoEstradiol implants in the arcuatenucleus induce lactogenesis in virgin rats Role of progesteronerdquoLife Sciences vol 62 no 3 pp 229ndash237 1997
[18] G R Morel R W Caron G M Console et al ldquoEstrogeninhibits tuberoinfundibular dopaminergic neurons but does notcause irreversible damagerdquo Brain Research Bulletin vol 80 no6 pp 347ndash352 2009
[19] R W Caron A M Salicioni and R P Deis ldquoMifepristonetreatment demonstrates the participation of adrenal glucocorti-coids in the regulation of oestrogen-induced prolactin secretionin ovariectomized ratsrdquo Journal of Steroid Biochemistry andMolecular Biology vol 48 no 4 pp 385ndash389 1994
[20] R P Deis D G Carrizo and G A Jahn ldquoSuckling-inducedprolactin release potentiates mifepristone-induced lactogenesisin pregnant ratsrdquo Journal of Reproduction and Fertility vol 87no 1 pp 147ndash153 1989
[21] R W Caron C A Jahn and R P Deis ldquoLactogenic actionsof different growth hormone preparations in pregnant andlactating ratsrdquo Journal of Endocrinology vol 142 no 3 pp 535ndash545 1994
[22] M Edery L Houdebine J Djiane and P A Kelly ldquoStudies of 120573-casein content of normal and neoplastic rat mammary tissuesby a homologous radioimmunoassayrdquo Molecular and CellularEndocrinology vol 34 no 2 pp 145ndash151 1984
[23] L E Bussmann and R P Deis ldquo120574-Glutamyltransferase activityin mammary gland of pregnant rats and its regulation by ovar-ian hormones prolactin and placental lactogenrdquo BiochemicalJournal vol 223 no 1 pp 275ndash277 1984
[24] J Russo and I H Russo ldquoAtlas and histologic classification oftumors of the rat mammary glandrdquo Journal of Mammary GlandBiology and Neoplasia vol 5 no 2 pp 187ndash200 2000
[25] C M Lopez-Fontana C V Sasso M E Maselli et al ldquoExperi-mental hypothyroidism increases apoptosis in dimethylbenzan-thracene-inducedmammary tumorsrdquoOncologyReports vol 30no 4 pp 1651ndash1660 2013
[26] C M Lopez-Fontana M E Maselli A M Salicioni and R WCaron ldquoThe inhibitory effect of progesterone on lactogenesisduring pregnancy is already evident by mid- to late gestation inrodentsrdquo Reproduction Fertility and Development vol 24 no 5pp 704ndash714 2012
[27] C Puissant and L M Houdebine ldquoAn improvement of thesingle-step method of RNA isolation by acid guanidiniumthiocyanate-phenol-chloroform extractionrdquo BioTechniques vol8 no 2 pp 148ndash149 1990
[28] G Thordarson E Jin R C Guzman S M Swanson S Nandiand F Talamantes ldquoRefractoriness to mammary tumorigenesisin parous rats is it caused by persistent changes in the hormonalenvironment or permanent biochemical alterations in themammary epitheliardquo Carcinogenesis vol 16 no 11 pp 2847ndash2853 1995
BioMed Research International 11
[29] G Thordarson K van Horn R C Guzman S Nandi and FTalamantes ldquoParous rats regain high susceptibility to chemi-cally induced mammary cancer after treatment with variousmammotropic hormonesrdquo Carcinogenesis vol 22 no 7 pp1027ndash1033 2001
[30] L Sivaraman O M Conneely D Medina and B W OMalleyldquop53 is a potential mediator of pregnancy and hormone-induced resistance to mammary carcinogenesisrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 98 no 22 pp 12379ndash12384 2001
[31] J Frasor and G Gibori ldquoProlactin regulation of estrogenreceptor expressionrdquo Trends in Endocrinology and Metabolismvol 14 no 3 pp 118ndash123 2003
[32] M Edery W Imagawa L Larson and S Nandi ldquoRegulation ofestrogen and progesterone receptor levels in mouse mammaryepithelial cells grown in serum-free collagen gel culturesrdquoEndocrinology vol 116 no 1 pp 105ndash112 1985
[33] C J Ormandy R E Hall D L Manning et al ldquoCoexpres-sion and cross-regulation of the prolactin receptor and sexsteroid hormone receptors in breast cancerrdquo Journal of ClinicalEndocrinology and Metabolism vol 82 no 11 pp 3692ndash36991997
[34] K A OrsquoLeary F Jallow D E Rugowski et al ldquoProlactinactivates ER120572 in the absence of ligand in female mammarydevelopment and carcinogenesis in vivordquo Endocrinology vol154 pp 4483ndash4492 2013
[35] B W Park K S Kim M K Heo et al ldquoExpression ofestrogen receptor-120573 in normal mammary and tumor tissues isit protective in breast carcinogenesisrdquo Breast Cancer Researchand Treatment vol 80 no 1 pp 79ndash85 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
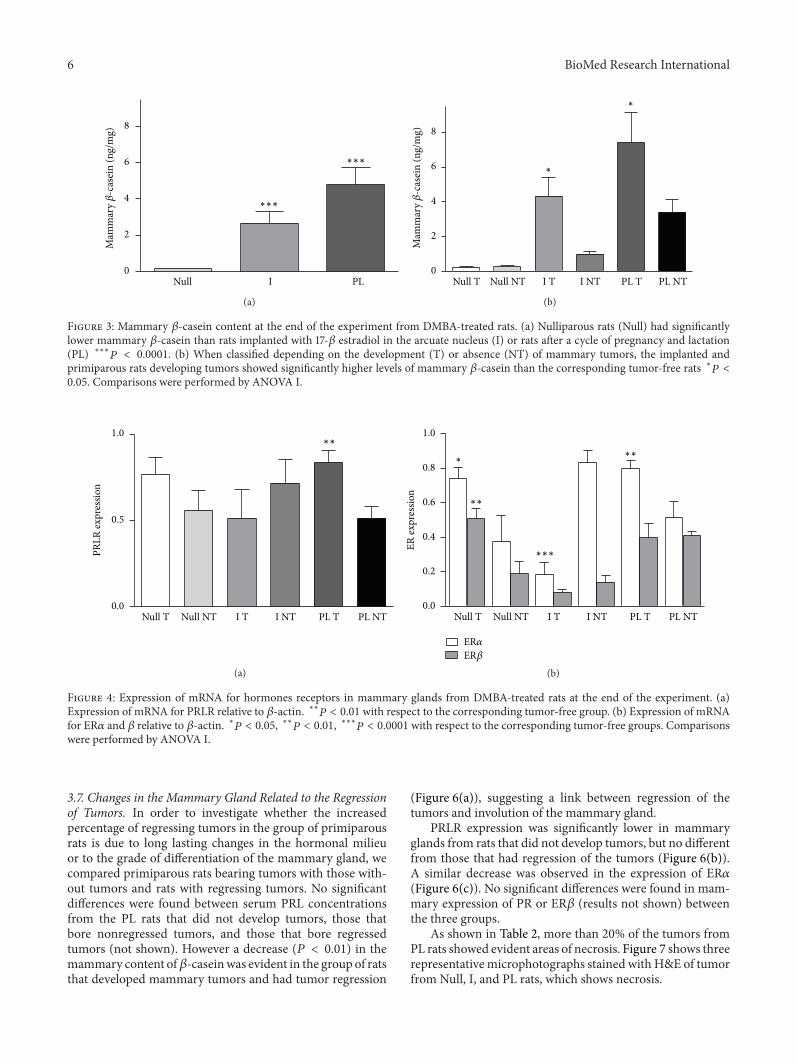
6 BioMed Research International
Null I PL0
2
4
6
8M
amm
ary120573
-cas
ein
(ng
mg)
lowastlowastlowast
lowastlowastlowast
(a)
Null T Null NT I T I NT PL T PL NT0
2
4
6
8
lowast
lowast
Mam
mar
y120573-c
asei
n (n
gm
g)
(b)
Figure 3 Mammary 120573-casein content at the end of the experiment from DMBA-treated rats (a) Nulliparous rats (Null) had significantlylower mammary 120573-casein than rats implanted with 17-120573 estradiol in the arcuate nucleus (I) or rats after a cycle of pregnancy and lactation(PL) lowastlowastlowast119875 lt 00001 (b) When classified depending on the development (T) or absence (NT) of mammary tumors the implanted andprimiparous rats developing tumors showed significantly higher levels of mammary 120573-casein than the corresponding tumor-free rats lowast119875 lt005 Comparisons were performed by ANOVA I
Null T Null NT I T I NT PL T PL NT00
05
10
PRLR
expr
essio
n
lowastlowast
(a)
00
02
04
06
08
10
ER ex
pres
sion
Null T Null NT I T I NT PL T PL NT
lowastlowastlowast
lowastlowast
lowastlowast
lowast
ER120572ER120573
(b)
Figure 4 Expression of mRNA for hormones receptors in mammary glands from DMBA-treated rats at the end of the experiment (a)Expression of mRNA for PRLR relative to 120573-actin lowastlowast119875 lt 001 with respect to the corresponding tumor-free group (b) Expression of mRNAfor ER120572 and 120573 relative to 120573-actin lowast119875 lt 005 lowastlowast119875 lt 001 lowastlowastlowast119875 lt 00001 with respect to the corresponding tumor-free groups Comparisonswere performed by ANOVA I
37 Changes in the Mammary Gland Related to the Regressionof Tumors In order to investigate whether the increasedpercentage of regressing tumors in the group of primiparousrats is due to long lasting changes in the hormonal milieuor to the grade of differentiation of the mammary gland wecompared primiparous rats bearing tumors with those with-out tumors and rats with regressing tumors No significantdifferences were found between serum PRL concentrationsfrom the PL rats that did not develop tumors those thatbore nonregressed tumors and those that bore regressedtumors (not shown) However a decrease (119875 lt 001) in themammary content of120573-caseinwas evident in the group of ratsthat developed mammary tumors and had tumor regression
(Figure 6(a)) suggesting a link between regression of thetumors and involution of the mammary gland
PRLR expression was significantly lower in mammaryglands from rats that did not develop tumors but no differentfrom those that had regression of the tumors (Figure 6(b))A similar decrease was observed in the expression of ER120572(Figure 6(c)) No significant differences were found in mam-mary expression of PR or ER120573 (results not shown) betweenthe three groups
As shown in Table 2 more than 20 of the tumors fromPL rats showed evident areas of necrosis Figure 7 shows threerepresentativemicrophotographs stained with HampE of tumorfrom Null I and PL rats which shows necrosis
BioMed Research International 7
Null I PL0
2
4
120573-C
asei
n in
tum
ors (
ngm
g)
lowastlowast
lowast
(a)
Null I PL00
01
02
03
04
05
ER120573
expr
essio
n
lowast
(b)
Null I PL00
05
10
15
PR ex
pres
sion
lowastlowast
(c)
Figure 5 120573-Casein content and expression of mRNA for hormones receptors in tumors from DMBA-treated rats (a) Tumor tissue fromnulliparous rats (Null) contains significantly lower amounts of 120573-casein than tumors from implanted (I) or primiparous (PL) rats (119875 lt 005and 119875 lt 0001 resp) (b)The expression of mRNA of ER120573 is lower in the tumors of I rats than in the other two groups (119875 lt 005) (c) mRNAexpression of PR is significantly higher in I rats with respect to Null or PL rats (119875 lt 001)
4 Discussion
To study the involvement of lactogenic hormones in mam-mary carcinogenesis we compared our Null or PL rats withvirgin rats made hyperprolactinemic through hypothalamicimplants of estradiol In this model serum estradiol remainsat a physiological level [16ndash18] allowing us to investigatethe effects of PRL excluding the well-known actions of highamounts of E
2on mammary carcinogenesis
A correlation between serum PRL levels and susceptibil-ity of various rat strains to chemically induced mammarycarcinogenesis has been suggested [9] Both GH and PRLhave been previously shown to be reduced after parturition ascomparedwith nulliparous age-matched animals [28]More-over parous rats showing almost complete refractorinessto chemical carcinogenesis acquire high susceptibility afterhormonal treatment that increases serum GH and PRL levels[29] In our OFA rats neither serum GH nor circulating PRLdecreased after PL Accordingly even when the incidenceof mammary tumors was slightly lower in primiparousrats the difference respect to nulliparae was not statisticallysignificant suggesting that the cycle of pregnancylactationfailed to prevent tumor development in OFA hrhr rats
This different behavior from the original strain (Sprague-Dawley) [28 29] regarding the lack of protection induced bypregnancy and lactation might be due to the lower levels ofcirculating PRL during lactation in the PLOFA ratsThus thedeficient lactation may have been insufficient to protect themammary tissue from tumoral transformation Moreoverdespite the persistent hyperprolactinaemia produced in I ratsthe incidence of tumors was similar to the other groupssuggesting a null impact of serum PRL levels on mammarycarcinogenesis Interestingly in I rats serum PRL was signif-icantly higher in those rats that developed tumors suggestingthat supraphysiological levels of PRL may be relevant inincreasing the risk of mammary cancer
Our results support the idea that the mammary devel-opment per se is not enough to account for the protectiveeffect of parity In fact the implant of estradiol in the arcuatenucleus produces hyperprolactinemia and luteal phase withhigh levels of circulating P
4and mammary development and
differentiation similar to pregnancy [16 17]Thus a protectiveeffect of the implants dependent on the differentiation of themammary gland could be expected However the incidenceof mammary tumors in the I group was similar to that in the
8 BioMed Research International
PL T PL NT PL RegT0
2
4
6
8
Mam
mar
y120573
-cas
ein
(ng
mg)
lowastlowast
(a)
PL T PL NT PL RegT00
05
10
PRL
R ex
pres
sion lowast
(b)
PL T PL NT PL RegT00
05
10
ER120572
expr
essio
n lowastlowast
(c)
Figure 6 120573-Casein content and expression of mRNA for hormones receptors in mammary glands from primiparous rats (PL) Mammary120573-casein content was lower (119875 lt 001) in mammary glands from PL rats that had tumor regression (PLRegT) than in those that did not havetumors (PLNT) or did have no regressing tumors (PLT) (b) and (c) PRLR and ER120572 expressions were lower (119875 lt 005 and 119875 lt 001 resp) inmammary glands from tumor-free PL rats compared to the other two groups
Null rats suggesting that the sustained hyperprolactinaemiais not enough to increase carcinogenesis and moreoverthe development of the mammary gland achieved in theimplanted rats is not enough to prevent chemical carcinogen-esis In agreement with our results chronic treatment withperphenazine producing hyperprolactinemia and high levelsof P4 did not protect the mammary gland against chemical
induction of tumors [30]It is worth noting that in our model mammary 120573-
casein content paralleled the alveolar area measured in thehistological preparations suggesting that the proliferationof epithelial tissue of the mammary gland is accompaniedof certain differentiation This result is in agreement with aprevious study in normal Wistar rats [26]
Interestingly the percentage of mammary tumors inregression was greatly affected by PL indicating that theclassical protective effect of gestation described by othersmaybe due at least in part to an increased number of tumorsundergoing regression rather than a decrease of the develop-ment of mammary tumors Our overall results suggest that acycle of PL has ldquotoxicrdquo effects on themammary tumors whichcannot be explained only by the milieu of mammogenic
hormones since we did not find any statistically significantdifference on serum GH (results not shown) or PRL valuesbetween nulliparous or primiparous rats On the contrarythe significantly lower content of 120573-casein in the mammarygland from rats that had regression of their tumors constitutesan interesting support of the idea that involution of thenormal epithelia can be implied in the regression of tumorsespecially so when the tumors of the rats with pregnancyand lactation retain the capability to synthesize 120573-caseinStudies currently under way in our laboratory are focusing onthe expression of several molecular markers involved in theprocess of involution of the mammary gland after lactationin the regressing tumors
We did not find correlation between expression of ARNmof PRLR and serum PRL levels However mammary glandsfrom PL rats that had developed tumors had increasedexpression of PRLR compared to PL rats free of tumorsThis difference can explain the higher alveolar developmentreached in those rats even when the values of circulating PRLat the moment of sacrifice were similar in both groups Theincreased development of alveolar tissue may cause in turnincreased content of 120573-casein It is worth to note that both I
BioMed Research International 9
(a) (b)
(c)
Figure 7 Representative microphotographs (100x) of HampE-stained normal tumors from nulliparous rats (a) hyperprolactinemic rats (b)and primiparous rats showing a noticeable area of necrosis (c) Arrows show necrotic areas
and PL rats had augmented alveolar development comparedto Null rats but the level of differentiation in terms of 120573-casein expression is higher in PL rats supporting a role for theextent of differentiation more than the alveolar developmentin the protective effect of PL
To assess whether the transformation of the mammarycells involves changes in the expression of receptors westudied in parallel the tumors and tissue from the con-tralateral normal mammary gland in the same animals Theexpression of ER120572 increased in tumors of hyperprolactinemicrats compared to the contralateral mammary gland but itwas unmodified in Null or PL rats (results not-shown) Theability of PRL to stimulate the expression of both types of ERhas been shown in the rat corpus luteum in the mammarygland and in the decidua (see [31] for a review) Previousreports suggest that PRL can stimulate ER expression in somebreast cancers [32 33] Moreover PRL can activate ER120572 evenin the absence of estrogenic ligands leading to oncogenesis[34] In our study the expression of ER120573 decreased in thetumors from the three groups compared to their respectivemammary glands (not shown) suggesting a protective rolefor ER120573 This result supports previous studies showing adecrease in the expression of ER120573 in the process of humanbreast cancer progression associatedwith poor differentiation[35] Moreover the expression of ER120572was higher in untrans-formed mammary tissue from Null or PL rats that developedmammary tumors compared with those that did not On thecontrary hyperprolactinemic rats that developed tumors had
lower expression of ER120572 in their mammary tissue This par-ticular result may suggest that when supraphysiological levelsof circulating PRL are constantly acting on the mammarygland tumors can progress even with lower expression ofER120572
In conclusion in our model of carcinogenesis using OFAhrhr rats we showed that serum PRL levels do not seem tohave relevance on the incidence of tumors probably becausethe relative low levels of PRL in those rats are not furtherdecreased by pregnancy and lactation like in other strainsEven though supraphysiological levels of serum PRL mayaffect carcinogenesis
On the other hand PL induces regression of the tumorsmost probably due to the degree of differentiation producedon the mammary cells than to the extent of proliferationreached by the epithelial tissue Alterations in the expressionof hormonal receptors may be involved in both progressionand regression of the tumors
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are deeply indebted to Mrs Elina Guinazu deDi Nasso and Mr Juan Rosales for their excellent technical
10 BioMed Research International
assistance This work was partially supported by grantsfrom Instituto Nacional del Cancer (Ministerio de SaludArgentina) and fromCONICET (Consejo Nacional de Inves-tigaciones Cientıficas y Tecnicas) Argentina
References
[1] P K Chakravarty and D K Sinha ldquoPregnancy inducedmammary tumor specific effector cells are present long afterparturition in a breast cancer model in ratsrdquo Cancer Letters vol154 no 1 pp 1ndash7 2000
[2] J Russo G A Balogh and I H Russo ldquoFull-term pregnancyinduces a specific genomic signature in the human breastrdquoCancer Epidemiology Biomarkers amp Prevention vol 17 no 1 pp51ndash66 2008
[3] J Russo L K Tay and I H Russo ldquoDifferentiation of themammary gland and susceptibility to carcinogenesisrdquo BreastCancer Research and Treatment vol 2 no 1 pp 5ndash73 1982
[4] OMaller KCHansen T R Lyons et al ldquoCollagen architecturein pregnancy-induced protection from breast cancerrdquo Journal ofCell Science vol 126 part 18 pp 4108ndash4110 2013
[5] S K Siwko J Dong M T Lewis H Liu S G Hilsenbeckand Y Li ldquoEvidence that an early pregnancy causes a persistentdecrease in the number of functional mammary epithelial stemcellsmdashimplications for pregnancy-induced protection againstbreast cancerrdquo Stem Cells vol 26 no 12 pp 3205ndash3209 2008
[6] Y Misra P A Bentley J P Bond S Tighe T Hunter and FZhao ldquoMammary gland morphological and gene expressionchanges underlying pregnancy protection of breast cancertumorigenesisrdquo Physiological Genomics vol 44 no 1 pp 76ndash882012
[7] J Russo and I H Russo ldquoInfluence of differentiation and cellkinetics on the susceptibility of the rat mammary gland tocarcinogenesisrdquo Cancer Research vol 40 no 8 pp 2677ndash26871980
[8] R C Guzman J Yang L Rajkumar G Thordarson X Chenand S Nand ldquoHormonal prevention of breast cancer mim-icking the protective effect of pregnancyrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 5 pp 2520ndash2525 1999
[9] A R Bovns R Buchan E N Cole A P M Forrest and KGrifiths ldquoBasal prolactin blood levels in three strains of ratwith differing incidence of 712-Dimethylbenz (a) anthraceneinduced mammary tumoursrdquo European Journal of Cancer vol9 no 3 pp 169ndash171 1973
[10] C W Welsch and H Nagasawa ldquoProlactin and murine mam-mary tumorigenesis a reviewrdquo Cancer Research vol 37 no 4pp 951ndash963 1977
[11] H Cohen I Sabbagh P Guillaumot J Bertrand M Robert-son and H G Friesen ldquoRelationship between bioassay andradioimmunoassay measurement of prolactin in the IPL rat ahypoprolactinemic rat strainrdquo Biology of Reproduction vol 38no 1 pp 84ndash90 1988
[12] S R Valdes A B Penissi R P Deis and G A JahnldquoHormonal profile and reproductive performance in lactationdeficient (OFA hrhr) and normal (Sprague-Dawley) femaleratsrdquo Reproduction vol 133 no 4 pp 827ndash840 2007
[13] H Bazzi A Kljuic A M Christiano and A A PanteleyevldquoIntragenic deletion in theDesmoglein 4 gene underlies the skinphenotype in the Iffa Credo ldquohairlessrdquo ratrdquo Differentiation vol72 no 8 pp 450ndash464 2004
[14] H Kim A A Panteleyev C A B Jahoda Y Ishii and A MChristiano ldquoGenomic organization and analysis of the hairlessgene in four hypotrichotic rat strainsrdquoMammalianGenome vol15 no 12 pp 975ndash981 2004
[15] B Meyer H Bazzi V Zidek et al ldquoA spontaneous mutationin the desmoglein 4 gene underlies hypotrichosis in a newlanceolate hair rat modelrdquo Differentiation vol 72 no 9-10 pp541ndash547 2004
[16] R W Caron and R P Deis ldquoParticipation of opioid and sero-toninergic systems in prolactin secretion induced by hypotha-lamic action of estradiolrdquoNeuroendocrinology vol 64 no 2 pp124ndash130 1996
[17] R W Caron and R P Deis ldquoEstradiol implants in the arcuatenucleus induce lactogenesis in virgin rats Role of progesteronerdquoLife Sciences vol 62 no 3 pp 229ndash237 1997
[18] G R Morel R W Caron G M Console et al ldquoEstrogeninhibits tuberoinfundibular dopaminergic neurons but does notcause irreversible damagerdquo Brain Research Bulletin vol 80 no6 pp 347ndash352 2009
[19] R W Caron A M Salicioni and R P Deis ldquoMifepristonetreatment demonstrates the participation of adrenal glucocorti-coids in the regulation of oestrogen-induced prolactin secretionin ovariectomized ratsrdquo Journal of Steroid Biochemistry andMolecular Biology vol 48 no 4 pp 385ndash389 1994
[20] R P Deis D G Carrizo and G A Jahn ldquoSuckling-inducedprolactin release potentiates mifepristone-induced lactogenesisin pregnant ratsrdquo Journal of Reproduction and Fertility vol 87no 1 pp 147ndash153 1989
[21] R W Caron C A Jahn and R P Deis ldquoLactogenic actionsof different growth hormone preparations in pregnant andlactating ratsrdquo Journal of Endocrinology vol 142 no 3 pp 535ndash545 1994
[22] M Edery L Houdebine J Djiane and P A Kelly ldquoStudies of 120573-casein content of normal and neoplastic rat mammary tissuesby a homologous radioimmunoassayrdquo Molecular and CellularEndocrinology vol 34 no 2 pp 145ndash151 1984
[23] L E Bussmann and R P Deis ldquo120574-Glutamyltransferase activityin mammary gland of pregnant rats and its regulation by ovar-ian hormones prolactin and placental lactogenrdquo BiochemicalJournal vol 223 no 1 pp 275ndash277 1984
[24] J Russo and I H Russo ldquoAtlas and histologic classification oftumors of the rat mammary glandrdquo Journal of Mammary GlandBiology and Neoplasia vol 5 no 2 pp 187ndash200 2000
[25] C M Lopez-Fontana C V Sasso M E Maselli et al ldquoExperi-mental hypothyroidism increases apoptosis in dimethylbenzan-thracene-inducedmammary tumorsrdquoOncologyReports vol 30no 4 pp 1651ndash1660 2013
[26] C M Lopez-Fontana M E Maselli A M Salicioni and R WCaron ldquoThe inhibitory effect of progesterone on lactogenesisduring pregnancy is already evident by mid- to late gestation inrodentsrdquo Reproduction Fertility and Development vol 24 no 5pp 704ndash714 2012
[27] C Puissant and L M Houdebine ldquoAn improvement of thesingle-step method of RNA isolation by acid guanidiniumthiocyanate-phenol-chloroform extractionrdquo BioTechniques vol8 no 2 pp 148ndash149 1990
[28] G Thordarson E Jin R C Guzman S M Swanson S Nandiand F Talamantes ldquoRefractoriness to mammary tumorigenesisin parous rats is it caused by persistent changes in the hormonalenvironment or permanent biochemical alterations in themammary epitheliardquo Carcinogenesis vol 16 no 11 pp 2847ndash2853 1995
BioMed Research International 11
[29] G Thordarson K van Horn R C Guzman S Nandi and FTalamantes ldquoParous rats regain high susceptibility to chemi-cally induced mammary cancer after treatment with variousmammotropic hormonesrdquo Carcinogenesis vol 22 no 7 pp1027ndash1033 2001
[30] L Sivaraman O M Conneely D Medina and B W OMalleyldquop53 is a potential mediator of pregnancy and hormone-induced resistance to mammary carcinogenesisrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 98 no 22 pp 12379ndash12384 2001
[31] J Frasor and G Gibori ldquoProlactin regulation of estrogenreceptor expressionrdquo Trends in Endocrinology and Metabolismvol 14 no 3 pp 118ndash123 2003
[32] M Edery W Imagawa L Larson and S Nandi ldquoRegulation ofestrogen and progesterone receptor levels in mouse mammaryepithelial cells grown in serum-free collagen gel culturesrdquoEndocrinology vol 116 no 1 pp 105ndash112 1985
[33] C J Ormandy R E Hall D L Manning et al ldquoCoexpres-sion and cross-regulation of the prolactin receptor and sexsteroid hormone receptors in breast cancerrdquo Journal of ClinicalEndocrinology and Metabolism vol 82 no 11 pp 3692ndash36991997
[34] K A OrsquoLeary F Jallow D E Rugowski et al ldquoProlactinactivates ER120572 in the absence of ligand in female mammarydevelopment and carcinogenesis in vivordquo Endocrinology vol154 pp 4483ndash4492 2013
[35] B W Park K S Kim M K Heo et al ldquoExpression ofestrogen receptor-120573 in normal mammary and tumor tissues isit protective in breast carcinogenesisrdquo Breast Cancer Researchand Treatment vol 80 no 1 pp 79ndash85 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 7
Null I PL0
2
4
120573-C
asei
n in
tum
ors (
ngm
g)
lowastlowast
lowast
(a)
Null I PL00
01
02
03
04
05
ER120573
expr
essio
n
lowast
(b)
Null I PL00
05
10
15
PR ex
pres
sion
lowastlowast
(c)
Figure 5 120573-Casein content and expression of mRNA for hormones receptors in tumors from DMBA-treated rats (a) Tumor tissue fromnulliparous rats (Null) contains significantly lower amounts of 120573-casein than tumors from implanted (I) or primiparous (PL) rats (119875 lt 005and 119875 lt 0001 resp) (b)The expression of mRNA of ER120573 is lower in the tumors of I rats than in the other two groups (119875 lt 005) (c) mRNAexpression of PR is significantly higher in I rats with respect to Null or PL rats (119875 lt 001)
4 Discussion
To study the involvement of lactogenic hormones in mam-mary carcinogenesis we compared our Null or PL rats withvirgin rats made hyperprolactinemic through hypothalamicimplants of estradiol In this model serum estradiol remainsat a physiological level [16ndash18] allowing us to investigatethe effects of PRL excluding the well-known actions of highamounts of E
2on mammary carcinogenesis
A correlation between serum PRL levels and susceptibil-ity of various rat strains to chemically induced mammarycarcinogenesis has been suggested [9] Both GH and PRLhave been previously shown to be reduced after parturition ascomparedwith nulliparous age-matched animals [28]More-over parous rats showing almost complete refractorinessto chemical carcinogenesis acquire high susceptibility afterhormonal treatment that increases serum GH and PRL levels[29] In our OFA rats neither serum GH nor circulating PRLdecreased after PL Accordingly even when the incidenceof mammary tumors was slightly lower in primiparousrats the difference respect to nulliparae was not statisticallysignificant suggesting that the cycle of pregnancylactationfailed to prevent tumor development in OFA hrhr rats
This different behavior from the original strain (Sprague-Dawley) [28 29] regarding the lack of protection induced bypregnancy and lactation might be due to the lower levels ofcirculating PRL during lactation in the PLOFA ratsThus thedeficient lactation may have been insufficient to protect themammary tissue from tumoral transformation Moreoverdespite the persistent hyperprolactinaemia produced in I ratsthe incidence of tumors was similar to the other groupssuggesting a null impact of serum PRL levels on mammarycarcinogenesis Interestingly in I rats serum PRL was signif-icantly higher in those rats that developed tumors suggestingthat supraphysiological levels of PRL may be relevant inincreasing the risk of mammary cancer
Our results support the idea that the mammary devel-opment per se is not enough to account for the protectiveeffect of parity In fact the implant of estradiol in the arcuatenucleus produces hyperprolactinemia and luteal phase withhigh levels of circulating P
4and mammary development and
differentiation similar to pregnancy [16 17]Thus a protectiveeffect of the implants dependent on the differentiation of themammary gland could be expected However the incidenceof mammary tumors in the I group was similar to that in the
8 BioMed Research International
PL T PL NT PL RegT0
2
4
6
8
Mam
mar
y120573
-cas
ein
(ng
mg)
lowastlowast
(a)
PL T PL NT PL RegT00
05
10
PRL
R ex
pres
sion lowast
(b)
PL T PL NT PL RegT00
05
10
ER120572
expr
essio
n lowastlowast
(c)
Figure 6 120573-Casein content and expression of mRNA for hormones receptors in mammary glands from primiparous rats (PL) Mammary120573-casein content was lower (119875 lt 001) in mammary glands from PL rats that had tumor regression (PLRegT) than in those that did not havetumors (PLNT) or did have no regressing tumors (PLT) (b) and (c) PRLR and ER120572 expressions were lower (119875 lt 005 and 119875 lt 001 resp) inmammary glands from tumor-free PL rats compared to the other two groups
Null rats suggesting that the sustained hyperprolactinaemiais not enough to increase carcinogenesis and moreoverthe development of the mammary gland achieved in theimplanted rats is not enough to prevent chemical carcinogen-esis In agreement with our results chronic treatment withperphenazine producing hyperprolactinemia and high levelsof P4 did not protect the mammary gland against chemical
induction of tumors [30]It is worth noting that in our model mammary 120573-
casein content paralleled the alveolar area measured in thehistological preparations suggesting that the proliferationof epithelial tissue of the mammary gland is accompaniedof certain differentiation This result is in agreement with aprevious study in normal Wistar rats [26]
Interestingly the percentage of mammary tumors inregression was greatly affected by PL indicating that theclassical protective effect of gestation described by othersmaybe due at least in part to an increased number of tumorsundergoing regression rather than a decrease of the develop-ment of mammary tumors Our overall results suggest that acycle of PL has ldquotoxicrdquo effects on themammary tumors whichcannot be explained only by the milieu of mammogenic
hormones since we did not find any statistically significantdifference on serum GH (results not shown) or PRL valuesbetween nulliparous or primiparous rats On the contrarythe significantly lower content of 120573-casein in the mammarygland from rats that had regression of their tumors constitutesan interesting support of the idea that involution of thenormal epithelia can be implied in the regression of tumorsespecially so when the tumors of the rats with pregnancyand lactation retain the capability to synthesize 120573-caseinStudies currently under way in our laboratory are focusing onthe expression of several molecular markers involved in theprocess of involution of the mammary gland after lactationin the regressing tumors
We did not find correlation between expression of ARNmof PRLR and serum PRL levels However mammary glandsfrom PL rats that had developed tumors had increasedexpression of PRLR compared to PL rats free of tumorsThis difference can explain the higher alveolar developmentreached in those rats even when the values of circulating PRLat the moment of sacrifice were similar in both groups Theincreased development of alveolar tissue may cause in turnincreased content of 120573-casein It is worth to note that both I
BioMed Research International 9
(a) (b)
(c)
Figure 7 Representative microphotographs (100x) of HampE-stained normal tumors from nulliparous rats (a) hyperprolactinemic rats (b)and primiparous rats showing a noticeable area of necrosis (c) Arrows show necrotic areas
and PL rats had augmented alveolar development comparedto Null rats but the level of differentiation in terms of 120573-casein expression is higher in PL rats supporting a role for theextent of differentiation more than the alveolar developmentin the protective effect of PL
To assess whether the transformation of the mammarycells involves changes in the expression of receptors westudied in parallel the tumors and tissue from the con-tralateral normal mammary gland in the same animals Theexpression of ER120572 increased in tumors of hyperprolactinemicrats compared to the contralateral mammary gland but itwas unmodified in Null or PL rats (results not-shown) Theability of PRL to stimulate the expression of both types of ERhas been shown in the rat corpus luteum in the mammarygland and in the decidua (see [31] for a review) Previousreports suggest that PRL can stimulate ER expression in somebreast cancers [32 33] Moreover PRL can activate ER120572 evenin the absence of estrogenic ligands leading to oncogenesis[34] In our study the expression of ER120573 decreased in thetumors from the three groups compared to their respectivemammary glands (not shown) suggesting a protective rolefor ER120573 This result supports previous studies showing adecrease in the expression of ER120573 in the process of humanbreast cancer progression associatedwith poor differentiation[35] Moreover the expression of ER120572was higher in untrans-formed mammary tissue from Null or PL rats that developedmammary tumors compared with those that did not On thecontrary hyperprolactinemic rats that developed tumors had
lower expression of ER120572 in their mammary tissue This par-ticular result may suggest that when supraphysiological levelsof circulating PRL are constantly acting on the mammarygland tumors can progress even with lower expression ofER120572
In conclusion in our model of carcinogenesis using OFAhrhr rats we showed that serum PRL levels do not seem tohave relevance on the incidence of tumors probably becausethe relative low levels of PRL in those rats are not furtherdecreased by pregnancy and lactation like in other strainsEven though supraphysiological levels of serum PRL mayaffect carcinogenesis
On the other hand PL induces regression of the tumorsmost probably due to the degree of differentiation producedon the mammary cells than to the extent of proliferationreached by the epithelial tissue Alterations in the expressionof hormonal receptors may be involved in both progressionand regression of the tumors
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are deeply indebted to Mrs Elina Guinazu deDi Nasso and Mr Juan Rosales for their excellent technical
10 BioMed Research International
assistance This work was partially supported by grantsfrom Instituto Nacional del Cancer (Ministerio de SaludArgentina) and fromCONICET (Consejo Nacional de Inves-tigaciones Cientıficas y Tecnicas) Argentina
References
[1] P K Chakravarty and D K Sinha ldquoPregnancy inducedmammary tumor specific effector cells are present long afterparturition in a breast cancer model in ratsrdquo Cancer Letters vol154 no 1 pp 1ndash7 2000
[2] J Russo G A Balogh and I H Russo ldquoFull-term pregnancyinduces a specific genomic signature in the human breastrdquoCancer Epidemiology Biomarkers amp Prevention vol 17 no 1 pp51ndash66 2008
[3] J Russo L K Tay and I H Russo ldquoDifferentiation of themammary gland and susceptibility to carcinogenesisrdquo BreastCancer Research and Treatment vol 2 no 1 pp 5ndash73 1982
[4] OMaller KCHansen T R Lyons et al ldquoCollagen architecturein pregnancy-induced protection from breast cancerrdquo Journal ofCell Science vol 126 part 18 pp 4108ndash4110 2013
[5] S K Siwko J Dong M T Lewis H Liu S G Hilsenbeckand Y Li ldquoEvidence that an early pregnancy causes a persistentdecrease in the number of functional mammary epithelial stemcellsmdashimplications for pregnancy-induced protection againstbreast cancerrdquo Stem Cells vol 26 no 12 pp 3205ndash3209 2008
[6] Y Misra P A Bentley J P Bond S Tighe T Hunter and FZhao ldquoMammary gland morphological and gene expressionchanges underlying pregnancy protection of breast cancertumorigenesisrdquo Physiological Genomics vol 44 no 1 pp 76ndash882012
[7] J Russo and I H Russo ldquoInfluence of differentiation and cellkinetics on the susceptibility of the rat mammary gland tocarcinogenesisrdquo Cancer Research vol 40 no 8 pp 2677ndash26871980
[8] R C Guzman J Yang L Rajkumar G Thordarson X Chenand S Nand ldquoHormonal prevention of breast cancer mim-icking the protective effect of pregnancyrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 5 pp 2520ndash2525 1999
[9] A R Bovns R Buchan E N Cole A P M Forrest and KGrifiths ldquoBasal prolactin blood levels in three strains of ratwith differing incidence of 712-Dimethylbenz (a) anthraceneinduced mammary tumoursrdquo European Journal of Cancer vol9 no 3 pp 169ndash171 1973
[10] C W Welsch and H Nagasawa ldquoProlactin and murine mam-mary tumorigenesis a reviewrdquo Cancer Research vol 37 no 4pp 951ndash963 1977
[11] H Cohen I Sabbagh P Guillaumot J Bertrand M Robert-son and H G Friesen ldquoRelationship between bioassay andradioimmunoassay measurement of prolactin in the IPL rat ahypoprolactinemic rat strainrdquo Biology of Reproduction vol 38no 1 pp 84ndash90 1988
[12] S R Valdes A B Penissi R P Deis and G A JahnldquoHormonal profile and reproductive performance in lactationdeficient (OFA hrhr) and normal (Sprague-Dawley) femaleratsrdquo Reproduction vol 133 no 4 pp 827ndash840 2007
[13] H Bazzi A Kljuic A M Christiano and A A PanteleyevldquoIntragenic deletion in theDesmoglein 4 gene underlies the skinphenotype in the Iffa Credo ldquohairlessrdquo ratrdquo Differentiation vol72 no 8 pp 450ndash464 2004
[14] H Kim A A Panteleyev C A B Jahoda Y Ishii and A MChristiano ldquoGenomic organization and analysis of the hairlessgene in four hypotrichotic rat strainsrdquoMammalianGenome vol15 no 12 pp 975ndash981 2004
[15] B Meyer H Bazzi V Zidek et al ldquoA spontaneous mutationin the desmoglein 4 gene underlies hypotrichosis in a newlanceolate hair rat modelrdquo Differentiation vol 72 no 9-10 pp541ndash547 2004
[16] R W Caron and R P Deis ldquoParticipation of opioid and sero-toninergic systems in prolactin secretion induced by hypotha-lamic action of estradiolrdquoNeuroendocrinology vol 64 no 2 pp124ndash130 1996
[17] R W Caron and R P Deis ldquoEstradiol implants in the arcuatenucleus induce lactogenesis in virgin rats Role of progesteronerdquoLife Sciences vol 62 no 3 pp 229ndash237 1997
[18] G R Morel R W Caron G M Console et al ldquoEstrogeninhibits tuberoinfundibular dopaminergic neurons but does notcause irreversible damagerdquo Brain Research Bulletin vol 80 no6 pp 347ndash352 2009
[19] R W Caron A M Salicioni and R P Deis ldquoMifepristonetreatment demonstrates the participation of adrenal glucocorti-coids in the regulation of oestrogen-induced prolactin secretionin ovariectomized ratsrdquo Journal of Steroid Biochemistry andMolecular Biology vol 48 no 4 pp 385ndash389 1994
[20] R P Deis D G Carrizo and G A Jahn ldquoSuckling-inducedprolactin release potentiates mifepristone-induced lactogenesisin pregnant ratsrdquo Journal of Reproduction and Fertility vol 87no 1 pp 147ndash153 1989
[21] R W Caron C A Jahn and R P Deis ldquoLactogenic actionsof different growth hormone preparations in pregnant andlactating ratsrdquo Journal of Endocrinology vol 142 no 3 pp 535ndash545 1994
[22] M Edery L Houdebine J Djiane and P A Kelly ldquoStudies of 120573-casein content of normal and neoplastic rat mammary tissuesby a homologous radioimmunoassayrdquo Molecular and CellularEndocrinology vol 34 no 2 pp 145ndash151 1984
[23] L E Bussmann and R P Deis ldquo120574-Glutamyltransferase activityin mammary gland of pregnant rats and its regulation by ovar-ian hormones prolactin and placental lactogenrdquo BiochemicalJournal vol 223 no 1 pp 275ndash277 1984
[24] J Russo and I H Russo ldquoAtlas and histologic classification oftumors of the rat mammary glandrdquo Journal of Mammary GlandBiology and Neoplasia vol 5 no 2 pp 187ndash200 2000
[25] C M Lopez-Fontana C V Sasso M E Maselli et al ldquoExperi-mental hypothyroidism increases apoptosis in dimethylbenzan-thracene-inducedmammary tumorsrdquoOncologyReports vol 30no 4 pp 1651ndash1660 2013
[26] C M Lopez-Fontana M E Maselli A M Salicioni and R WCaron ldquoThe inhibitory effect of progesterone on lactogenesisduring pregnancy is already evident by mid- to late gestation inrodentsrdquo Reproduction Fertility and Development vol 24 no 5pp 704ndash714 2012
[27] C Puissant and L M Houdebine ldquoAn improvement of thesingle-step method of RNA isolation by acid guanidiniumthiocyanate-phenol-chloroform extractionrdquo BioTechniques vol8 no 2 pp 148ndash149 1990
[28] G Thordarson E Jin R C Guzman S M Swanson S Nandiand F Talamantes ldquoRefractoriness to mammary tumorigenesisin parous rats is it caused by persistent changes in the hormonalenvironment or permanent biochemical alterations in themammary epitheliardquo Carcinogenesis vol 16 no 11 pp 2847ndash2853 1995
BioMed Research International 11
[29] G Thordarson K van Horn R C Guzman S Nandi and FTalamantes ldquoParous rats regain high susceptibility to chemi-cally induced mammary cancer after treatment with variousmammotropic hormonesrdquo Carcinogenesis vol 22 no 7 pp1027ndash1033 2001
[30] L Sivaraman O M Conneely D Medina and B W OMalleyldquop53 is a potential mediator of pregnancy and hormone-induced resistance to mammary carcinogenesisrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 98 no 22 pp 12379ndash12384 2001
[31] J Frasor and G Gibori ldquoProlactin regulation of estrogenreceptor expressionrdquo Trends in Endocrinology and Metabolismvol 14 no 3 pp 118ndash123 2003
[32] M Edery W Imagawa L Larson and S Nandi ldquoRegulation ofestrogen and progesterone receptor levels in mouse mammaryepithelial cells grown in serum-free collagen gel culturesrdquoEndocrinology vol 116 no 1 pp 105ndash112 1985
[33] C J Ormandy R E Hall D L Manning et al ldquoCoexpres-sion and cross-regulation of the prolactin receptor and sexsteroid hormone receptors in breast cancerrdquo Journal of ClinicalEndocrinology and Metabolism vol 82 no 11 pp 3692ndash36991997
[34] K A OrsquoLeary F Jallow D E Rugowski et al ldquoProlactinactivates ER120572 in the absence of ligand in female mammarydevelopment and carcinogenesis in vivordquo Endocrinology vol154 pp 4483ndash4492 2013
[35] B W Park K S Kim M K Heo et al ldquoExpression ofestrogen receptor-120573 in normal mammary and tumor tissues isit protective in breast carcinogenesisrdquo Breast Cancer Researchand Treatment vol 80 no 1 pp 79ndash85 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
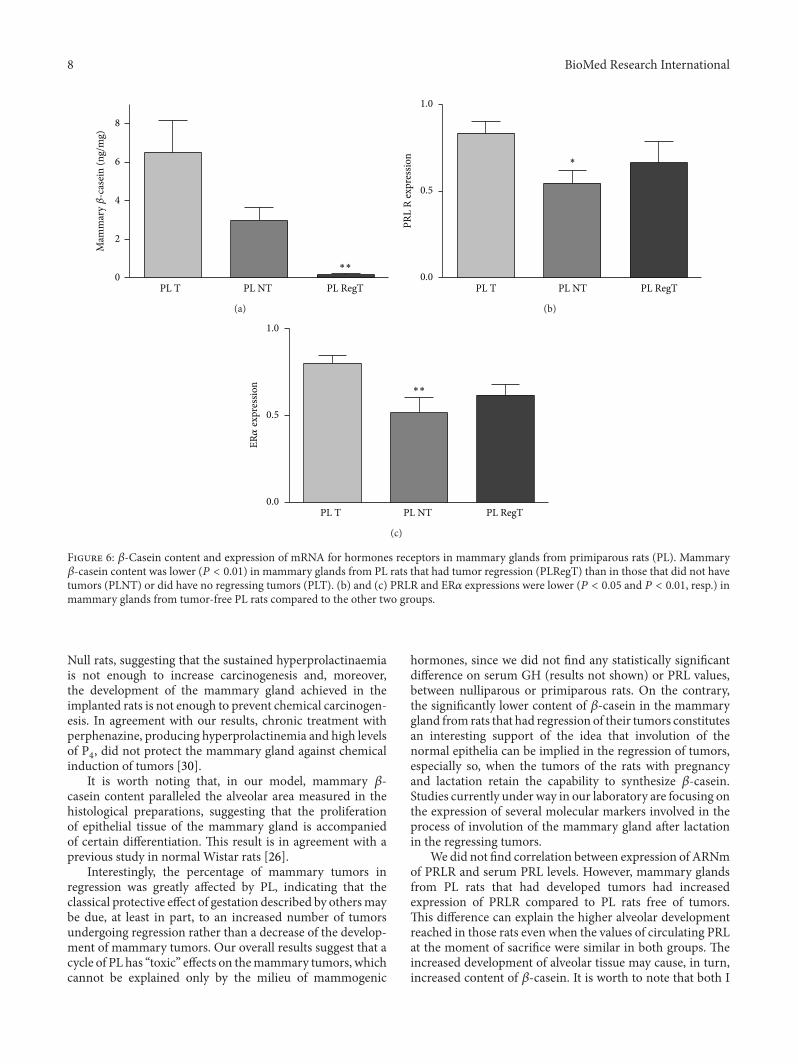
8 BioMed Research International
PL T PL NT PL RegT0
2
4
6
8
Mam
mar
y120573
-cas
ein
(ng
mg)
lowastlowast
(a)
PL T PL NT PL RegT00
05
10
PRL
R ex
pres
sion lowast
(b)
PL T PL NT PL RegT00
05
10
ER120572
expr
essio
n lowastlowast
(c)
Figure 6 120573-Casein content and expression of mRNA for hormones receptors in mammary glands from primiparous rats (PL) Mammary120573-casein content was lower (119875 lt 001) in mammary glands from PL rats that had tumor regression (PLRegT) than in those that did not havetumors (PLNT) or did have no regressing tumors (PLT) (b) and (c) PRLR and ER120572 expressions were lower (119875 lt 005 and 119875 lt 001 resp) inmammary glands from tumor-free PL rats compared to the other two groups
Null rats suggesting that the sustained hyperprolactinaemiais not enough to increase carcinogenesis and moreoverthe development of the mammary gland achieved in theimplanted rats is not enough to prevent chemical carcinogen-esis In agreement with our results chronic treatment withperphenazine producing hyperprolactinemia and high levelsof P4 did not protect the mammary gland against chemical
induction of tumors [30]It is worth noting that in our model mammary 120573-
casein content paralleled the alveolar area measured in thehistological preparations suggesting that the proliferationof epithelial tissue of the mammary gland is accompaniedof certain differentiation This result is in agreement with aprevious study in normal Wistar rats [26]
Interestingly the percentage of mammary tumors inregression was greatly affected by PL indicating that theclassical protective effect of gestation described by othersmaybe due at least in part to an increased number of tumorsundergoing regression rather than a decrease of the develop-ment of mammary tumors Our overall results suggest that acycle of PL has ldquotoxicrdquo effects on themammary tumors whichcannot be explained only by the milieu of mammogenic
hormones since we did not find any statistically significantdifference on serum GH (results not shown) or PRL valuesbetween nulliparous or primiparous rats On the contrarythe significantly lower content of 120573-casein in the mammarygland from rats that had regression of their tumors constitutesan interesting support of the idea that involution of thenormal epithelia can be implied in the regression of tumorsespecially so when the tumors of the rats with pregnancyand lactation retain the capability to synthesize 120573-caseinStudies currently under way in our laboratory are focusing onthe expression of several molecular markers involved in theprocess of involution of the mammary gland after lactationin the regressing tumors
We did not find correlation between expression of ARNmof PRLR and serum PRL levels However mammary glandsfrom PL rats that had developed tumors had increasedexpression of PRLR compared to PL rats free of tumorsThis difference can explain the higher alveolar developmentreached in those rats even when the values of circulating PRLat the moment of sacrifice were similar in both groups Theincreased development of alveolar tissue may cause in turnincreased content of 120573-casein It is worth to note that both I
BioMed Research International 9
(a) (b)
(c)
Figure 7 Representative microphotographs (100x) of HampE-stained normal tumors from nulliparous rats (a) hyperprolactinemic rats (b)and primiparous rats showing a noticeable area of necrosis (c) Arrows show necrotic areas
and PL rats had augmented alveolar development comparedto Null rats but the level of differentiation in terms of 120573-casein expression is higher in PL rats supporting a role for theextent of differentiation more than the alveolar developmentin the protective effect of PL
To assess whether the transformation of the mammarycells involves changes in the expression of receptors westudied in parallel the tumors and tissue from the con-tralateral normal mammary gland in the same animals Theexpression of ER120572 increased in tumors of hyperprolactinemicrats compared to the contralateral mammary gland but itwas unmodified in Null or PL rats (results not-shown) Theability of PRL to stimulate the expression of both types of ERhas been shown in the rat corpus luteum in the mammarygland and in the decidua (see [31] for a review) Previousreports suggest that PRL can stimulate ER expression in somebreast cancers [32 33] Moreover PRL can activate ER120572 evenin the absence of estrogenic ligands leading to oncogenesis[34] In our study the expression of ER120573 decreased in thetumors from the three groups compared to their respectivemammary glands (not shown) suggesting a protective rolefor ER120573 This result supports previous studies showing adecrease in the expression of ER120573 in the process of humanbreast cancer progression associatedwith poor differentiation[35] Moreover the expression of ER120572was higher in untrans-formed mammary tissue from Null or PL rats that developedmammary tumors compared with those that did not On thecontrary hyperprolactinemic rats that developed tumors had
lower expression of ER120572 in their mammary tissue This par-ticular result may suggest that when supraphysiological levelsof circulating PRL are constantly acting on the mammarygland tumors can progress even with lower expression ofER120572
In conclusion in our model of carcinogenesis using OFAhrhr rats we showed that serum PRL levels do not seem tohave relevance on the incidence of tumors probably becausethe relative low levels of PRL in those rats are not furtherdecreased by pregnancy and lactation like in other strainsEven though supraphysiological levels of serum PRL mayaffect carcinogenesis
On the other hand PL induces regression of the tumorsmost probably due to the degree of differentiation producedon the mammary cells than to the extent of proliferationreached by the epithelial tissue Alterations in the expressionof hormonal receptors may be involved in both progressionand regression of the tumors
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are deeply indebted to Mrs Elina Guinazu deDi Nasso and Mr Juan Rosales for their excellent technical
10 BioMed Research International
assistance This work was partially supported by grantsfrom Instituto Nacional del Cancer (Ministerio de SaludArgentina) and fromCONICET (Consejo Nacional de Inves-tigaciones Cientıficas y Tecnicas) Argentina
References
[1] P K Chakravarty and D K Sinha ldquoPregnancy inducedmammary tumor specific effector cells are present long afterparturition in a breast cancer model in ratsrdquo Cancer Letters vol154 no 1 pp 1ndash7 2000
[2] J Russo G A Balogh and I H Russo ldquoFull-term pregnancyinduces a specific genomic signature in the human breastrdquoCancer Epidemiology Biomarkers amp Prevention vol 17 no 1 pp51ndash66 2008
[3] J Russo L K Tay and I H Russo ldquoDifferentiation of themammary gland and susceptibility to carcinogenesisrdquo BreastCancer Research and Treatment vol 2 no 1 pp 5ndash73 1982
[4] OMaller KCHansen T R Lyons et al ldquoCollagen architecturein pregnancy-induced protection from breast cancerrdquo Journal ofCell Science vol 126 part 18 pp 4108ndash4110 2013
[5] S K Siwko J Dong M T Lewis H Liu S G Hilsenbeckand Y Li ldquoEvidence that an early pregnancy causes a persistentdecrease in the number of functional mammary epithelial stemcellsmdashimplications for pregnancy-induced protection againstbreast cancerrdquo Stem Cells vol 26 no 12 pp 3205ndash3209 2008
[6] Y Misra P A Bentley J P Bond S Tighe T Hunter and FZhao ldquoMammary gland morphological and gene expressionchanges underlying pregnancy protection of breast cancertumorigenesisrdquo Physiological Genomics vol 44 no 1 pp 76ndash882012
[7] J Russo and I H Russo ldquoInfluence of differentiation and cellkinetics on the susceptibility of the rat mammary gland tocarcinogenesisrdquo Cancer Research vol 40 no 8 pp 2677ndash26871980
[8] R C Guzman J Yang L Rajkumar G Thordarson X Chenand S Nand ldquoHormonal prevention of breast cancer mim-icking the protective effect of pregnancyrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 5 pp 2520ndash2525 1999
[9] A R Bovns R Buchan E N Cole A P M Forrest and KGrifiths ldquoBasal prolactin blood levels in three strains of ratwith differing incidence of 712-Dimethylbenz (a) anthraceneinduced mammary tumoursrdquo European Journal of Cancer vol9 no 3 pp 169ndash171 1973
[10] C W Welsch and H Nagasawa ldquoProlactin and murine mam-mary tumorigenesis a reviewrdquo Cancer Research vol 37 no 4pp 951ndash963 1977
[11] H Cohen I Sabbagh P Guillaumot J Bertrand M Robert-son and H G Friesen ldquoRelationship between bioassay andradioimmunoassay measurement of prolactin in the IPL rat ahypoprolactinemic rat strainrdquo Biology of Reproduction vol 38no 1 pp 84ndash90 1988
[12] S R Valdes A B Penissi R P Deis and G A JahnldquoHormonal profile and reproductive performance in lactationdeficient (OFA hrhr) and normal (Sprague-Dawley) femaleratsrdquo Reproduction vol 133 no 4 pp 827ndash840 2007
[13] H Bazzi A Kljuic A M Christiano and A A PanteleyevldquoIntragenic deletion in theDesmoglein 4 gene underlies the skinphenotype in the Iffa Credo ldquohairlessrdquo ratrdquo Differentiation vol72 no 8 pp 450ndash464 2004
[14] H Kim A A Panteleyev C A B Jahoda Y Ishii and A MChristiano ldquoGenomic organization and analysis of the hairlessgene in four hypotrichotic rat strainsrdquoMammalianGenome vol15 no 12 pp 975ndash981 2004
[15] B Meyer H Bazzi V Zidek et al ldquoA spontaneous mutationin the desmoglein 4 gene underlies hypotrichosis in a newlanceolate hair rat modelrdquo Differentiation vol 72 no 9-10 pp541ndash547 2004
[16] R W Caron and R P Deis ldquoParticipation of opioid and sero-toninergic systems in prolactin secretion induced by hypotha-lamic action of estradiolrdquoNeuroendocrinology vol 64 no 2 pp124ndash130 1996
[17] R W Caron and R P Deis ldquoEstradiol implants in the arcuatenucleus induce lactogenesis in virgin rats Role of progesteronerdquoLife Sciences vol 62 no 3 pp 229ndash237 1997
[18] G R Morel R W Caron G M Console et al ldquoEstrogeninhibits tuberoinfundibular dopaminergic neurons but does notcause irreversible damagerdquo Brain Research Bulletin vol 80 no6 pp 347ndash352 2009
[19] R W Caron A M Salicioni and R P Deis ldquoMifepristonetreatment demonstrates the participation of adrenal glucocorti-coids in the regulation of oestrogen-induced prolactin secretionin ovariectomized ratsrdquo Journal of Steroid Biochemistry andMolecular Biology vol 48 no 4 pp 385ndash389 1994
[20] R P Deis D G Carrizo and G A Jahn ldquoSuckling-inducedprolactin release potentiates mifepristone-induced lactogenesisin pregnant ratsrdquo Journal of Reproduction and Fertility vol 87no 1 pp 147ndash153 1989
[21] R W Caron C A Jahn and R P Deis ldquoLactogenic actionsof different growth hormone preparations in pregnant andlactating ratsrdquo Journal of Endocrinology vol 142 no 3 pp 535ndash545 1994
[22] M Edery L Houdebine J Djiane and P A Kelly ldquoStudies of 120573-casein content of normal and neoplastic rat mammary tissuesby a homologous radioimmunoassayrdquo Molecular and CellularEndocrinology vol 34 no 2 pp 145ndash151 1984
[23] L E Bussmann and R P Deis ldquo120574-Glutamyltransferase activityin mammary gland of pregnant rats and its regulation by ovar-ian hormones prolactin and placental lactogenrdquo BiochemicalJournal vol 223 no 1 pp 275ndash277 1984
[24] J Russo and I H Russo ldquoAtlas and histologic classification oftumors of the rat mammary glandrdquo Journal of Mammary GlandBiology and Neoplasia vol 5 no 2 pp 187ndash200 2000
[25] C M Lopez-Fontana C V Sasso M E Maselli et al ldquoExperi-mental hypothyroidism increases apoptosis in dimethylbenzan-thracene-inducedmammary tumorsrdquoOncologyReports vol 30no 4 pp 1651ndash1660 2013
[26] C M Lopez-Fontana M E Maselli A M Salicioni and R WCaron ldquoThe inhibitory effect of progesterone on lactogenesisduring pregnancy is already evident by mid- to late gestation inrodentsrdquo Reproduction Fertility and Development vol 24 no 5pp 704ndash714 2012
[27] C Puissant and L M Houdebine ldquoAn improvement of thesingle-step method of RNA isolation by acid guanidiniumthiocyanate-phenol-chloroform extractionrdquo BioTechniques vol8 no 2 pp 148ndash149 1990
[28] G Thordarson E Jin R C Guzman S M Swanson S Nandiand F Talamantes ldquoRefractoriness to mammary tumorigenesisin parous rats is it caused by persistent changes in the hormonalenvironment or permanent biochemical alterations in themammary epitheliardquo Carcinogenesis vol 16 no 11 pp 2847ndash2853 1995
BioMed Research International 11
[29] G Thordarson K van Horn R C Guzman S Nandi and FTalamantes ldquoParous rats regain high susceptibility to chemi-cally induced mammary cancer after treatment with variousmammotropic hormonesrdquo Carcinogenesis vol 22 no 7 pp1027ndash1033 2001
[30] L Sivaraman O M Conneely D Medina and B W OMalleyldquop53 is a potential mediator of pregnancy and hormone-induced resistance to mammary carcinogenesisrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 98 no 22 pp 12379ndash12384 2001
[31] J Frasor and G Gibori ldquoProlactin regulation of estrogenreceptor expressionrdquo Trends in Endocrinology and Metabolismvol 14 no 3 pp 118ndash123 2003
[32] M Edery W Imagawa L Larson and S Nandi ldquoRegulation ofestrogen and progesterone receptor levels in mouse mammaryepithelial cells grown in serum-free collagen gel culturesrdquoEndocrinology vol 116 no 1 pp 105ndash112 1985
[33] C J Ormandy R E Hall D L Manning et al ldquoCoexpres-sion and cross-regulation of the prolactin receptor and sexsteroid hormone receptors in breast cancerrdquo Journal of ClinicalEndocrinology and Metabolism vol 82 no 11 pp 3692ndash36991997
[34] K A OrsquoLeary F Jallow D E Rugowski et al ldquoProlactinactivates ER120572 in the absence of ligand in female mammarydevelopment and carcinogenesis in vivordquo Endocrinology vol154 pp 4483ndash4492 2013
[35] B W Park K S Kim M K Heo et al ldquoExpression ofestrogen receptor-120573 in normal mammary and tumor tissues isit protective in breast carcinogenesisrdquo Breast Cancer Researchand Treatment vol 80 no 1 pp 79ndash85 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 9
(a) (b)
(c)
Figure 7 Representative microphotographs (100x) of HampE-stained normal tumors from nulliparous rats (a) hyperprolactinemic rats (b)and primiparous rats showing a noticeable area of necrosis (c) Arrows show necrotic areas
and PL rats had augmented alveolar development comparedto Null rats but the level of differentiation in terms of 120573-casein expression is higher in PL rats supporting a role for theextent of differentiation more than the alveolar developmentin the protective effect of PL
To assess whether the transformation of the mammarycells involves changes in the expression of receptors westudied in parallel the tumors and tissue from the con-tralateral normal mammary gland in the same animals Theexpression of ER120572 increased in tumors of hyperprolactinemicrats compared to the contralateral mammary gland but itwas unmodified in Null or PL rats (results not-shown) Theability of PRL to stimulate the expression of both types of ERhas been shown in the rat corpus luteum in the mammarygland and in the decidua (see [31] for a review) Previousreports suggest that PRL can stimulate ER expression in somebreast cancers [32 33] Moreover PRL can activate ER120572 evenin the absence of estrogenic ligands leading to oncogenesis[34] In our study the expression of ER120573 decreased in thetumors from the three groups compared to their respectivemammary glands (not shown) suggesting a protective rolefor ER120573 This result supports previous studies showing adecrease in the expression of ER120573 in the process of humanbreast cancer progression associatedwith poor differentiation[35] Moreover the expression of ER120572was higher in untrans-formed mammary tissue from Null or PL rats that developedmammary tumors compared with those that did not On thecontrary hyperprolactinemic rats that developed tumors had
lower expression of ER120572 in their mammary tissue This par-ticular result may suggest that when supraphysiological levelsof circulating PRL are constantly acting on the mammarygland tumors can progress even with lower expression ofER120572
In conclusion in our model of carcinogenesis using OFAhrhr rats we showed that serum PRL levels do not seem tohave relevance on the incidence of tumors probably becausethe relative low levels of PRL in those rats are not furtherdecreased by pregnancy and lactation like in other strainsEven though supraphysiological levels of serum PRL mayaffect carcinogenesis
On the other hand PL induces regression of the tumorsmost probably due to the degree of differentiation producedon the mammary cells than to the extent of proliferationreached by the epithelial tissue Alterations in the expressionof hormonal receptors may be involved in both progressionand regression of the tumors
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are deeply indebted to Mrs Elina Guinazu deDi Nasso and Mr Juan Rosales for their excellent technical
10 BioMed Research International
assistance This work was partially supported by grantsfrom Instituto Nacional del Cancer (Ministerio de SaludArgentina) and fromCONICET (Consejo Nacional de Inves-tigaciones Cientıficas y Tecnicas) Argentina
References
[1] P K Chakravarty and D K Sinha ldquoPregnancy inducedmammary tumor specific effector cells are present long afterparturition in a breast cancer model in ratsrdquo Cancer Letters vol154 no 1 pp 1ndash7 2000
[2] J Russo G A Balogh and I H Russo ldquoFull-term pregnancyinduces a specific genomic signature in the human breastrdquoCancer Epidemiology Biomarkers amp Prevention vol 17 no 1 pp51ndash66 2008
[3] J Russo L K Tay and I H Russo ldquoDifferentiation of themammary gland and susceptibility to carcinogenesisrdquo BreastCancer Research and Treatment vol 2 no 1 pp 5ndash73 1982
[4] OMaller KCHansen T R Lyons et al ldquoCollagen architecturein pregnancy-induced protection from breast cancerrdquo Journal ofCell Science vol 126 part 18 pp 4108ndash4110 2013
[5] S K Siwko J Dong M T Lewis H Liu S G Hilsenbeckand Y Li ldquoEvidence that an early pregnancy causes a persistentdecrease in the number of functional mammary epithelial stemcellsmdashimplications for pregnancy-induced protection againstbreast cancerrdquo Stem Cells vol 26 no 12 pp 3205ndash3209 2008
[6] Y Misra P A Bentley J P Bond S Tighe T Hunter and FZhao ldquoMammary gland morphological and gene expressionchanges underlying pregnancy protection of breast cancertumorigenesisrdquo Physiological Genomics vol 44 no 1 pp 76ndash882012
[7] J Russo and I H Russo ldquoInfluence of differentiation and cellkinetics on the susceptibility of the rat mammary gland tocarcinogenesisrdquo Cancer Research vol 40 no 8 pp 2677ndash26871980
[8] R C Guzman J Yang L Rajkumar G Thordarson X Chenand S Nand ldquoHormonal prevention of breast cancer mim-icking the protective effect of pregnancyrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 5 pp 2520ndash2525 1999
[9] A R Bovns R Buchan E N Cole A P M Forrest and KGrifiths ldquoBasal prolactin blood levels in three strains of ratwith differing incidence of 712-Dimethylbenz (a) anthraceneinduced mammary tumoursrdquo European Journal of Cancer vol9 no 3 pp 169ndash171 1973
[10] C W Welsch and H Nagasawa ldquoProlactin and murine mam-mary tumorigenesis a reviewrdquo Cancer Research vol 37 no 4pp 951ndash963 1977
[11] H Cohen I Sabbagh P Guillaumot J Bertrand M Robert-son and H G Friesen ldquoRelationship between bioassay andradioimmunoassay measurement of prolactin in the IPL rat ahypoprolactinemic rat strainrdquo Biology of Reproduction vol 38no 1 pp 84ndash90 1988
[12] S R Valdes A B Penissi R P Deis and G A JahnldquoHormonal profile and reproductive performance in lactationdeficient (OFA hrhr) and normal (Sprague-Dawley) femaleratsrdquo Reproduction vol 133 no 4 pp 827ndash840 2007
[13] H Bazzi A Kljuic A M Christiano and A A PanteleyevldquoIntragenic deletion in theDesmoglein 4 gene underlies the skinphenotype in the Iffa Credo ldquohairlessrdquo ratrdquo Differentiation vol72 no 8 pp 450ndash464 2004
[14] H Kim A A Panteleyev C A B Jahoda Y Ishii and A MChristiano ldquoGenomic organization and analysis of the hairlessgene in four hypotrichotic rat strainsrdquoMammalianGenome vol15 no 12 pp 975ndash981 2004
[15] B Meyer H Bazzi V Zidek et al ldquoA spontaneous mutationin the desmoglein 4 gene underlies hypotrichosis in a newlanceolate hair rat modelrdquo Differentiation vol 72 no 9-10 pp541ndash547 2004
[16] R W Caron and R P Deis ldquoParticipation of opioid and sero-toninergic systems in prolactin secretion induced by hypotha-lamic action of estradiolrdquoNeuroendocrinology vol 64 no 2 pp124ndash130 1996
[17] R W Caron and R P Deis ldquoEstradiol implants in the arcuatenucleus induce lactogenesis in virgin rats Role of progesteronerdquoLife Sciences vol 62 no 3 pp 229ndash237 1997
[18] G R Morel R W Caron G M Console et al ldquoEstrogeninhibits tuberoinfundibular dopaminergic neurons but does notcause irreversible damagerdquo Brain Research Bulletin vol 80 no6 pp 347ndash352 2009
[19] R W Caron A M Salicioni and R P Deis ldquoMifepristonetreatment demonstrates the participation of adrenal glucocorti-coids in the regulation of oestrogen-induced prolactin secretionin ovariectomized ratsrdquo Journal of Steroid Biochemistry andMolecular Biology vol 48 no 4 pp 385ndash389 1994
[20] R P Deis D G Carrizo and G A Jahn ldquoSuckling-inducedprolactin release potentiates mifepristone-induced lactogenesisin pregnant ratsrdquo Journal of Reproduction and Fertility vol 87no 1 pp 147ndash153 1989
[21] R W Caron C A Jahn and R P Deis ldquoLactogenic actionsof different growth hormone preparations in pregnant andlactating ratsrdquo Journal of Endocrinology vol 142 no 3 pp 535ndash545 1994
[22] M Edery L Houdebine J Djiane and P A Kelly ldquoStudies of 120573-casein content of normal and neoplastic rat mammary tissuesby a homologous radioimmunoassayrdquo Molecular and CellularEndocrinology vol 34 no 2 pp 145ndash151 1984
[23] L E Bussmann and R P Deis ldquo120574-Glutamyltransferase activityin mammary gland of pregnant rats and its regulation by ovar-ian hormones prolactin and placental lactogenrdquo BiochemicalJournal vol 223 no 1 pp 275ndash277 1984
[24] J Russo and I H Russo ldquoAtlas and histologic classification oftumors of the rat mammary glandrdquo Journal of Mammary GlandBiology and Neoplasia vol 5 no 2 pp 187ndash200 2000
[25] C M Lopez-Fontana C V Sasso M E Maselli et al ldquoExperi-mental hypothyroidism increases apoptosis in dimethylbenzan-thracene-inducedmammary tumorsrdquoOncologyReports vol 30no 4 pp 1651ndash1660 2013
[26] C M Lopez-Fontana M E Maselli A M Salicioni and R WCaron ldquoThe inhibitory effect of progesterone on lactogenesisduring pregnancy is already evident by mid- to late gestation inrodentsrdquo Reproduction Fertility and Development vol 24 no 5pp 704ndash714 2012
[27] C Puissant and L M Houdebine ldquoAn improvement of thesingle-step method of RNA isolation by acid guanidiniumthiocyanate-phenol-chloroform extractionrdquo BioTechniques vol8 no 2 pp 148ndash149 1990
[28] G Thordarson E Jin R C Guzman S M Swanson S Nandiand F Talamantes ldquoRefractoriness to mammary tumorigenesisin parous rats is it caused by persistent changes in the hormonalenvironment or permanent biochemical alterations in themammary epitheliardquo Carcinogenesis vol 16 no 11 pp 2847ndash2853 1995
BioMed Research International 11
[29] G Thordarson K van Horn R C Guzman S Nandi and FTalamantes ldquoParous rats regain high susceptibility to chemi-cally induced mammary cancer after treatment with variousmammotropic hormonesrdquo Carcinogenesis vol 22 no 7 pp1027ndash1033 2001
[30] L Sivaraman O M Conneely D Medina and B W OMalleyldquop53 is a potential mediator of pregnancy and hormone-induced resistance to mammary carcinogenesisrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 98 no 22 pp 12379ndash12384 2001
[31] J Frasor and G Gibori ldquoProlactin regulation of estrogenreceptor expressionrdquo Trends in Endocrinology and Metabolismvol 14 no 3 pp 118ndash123 2003
[32] M Edery W Imagawa L Larson and S Nandi ldquoRegulation ofestrogen and progesterone receptor levels in mouse mammaryepithelial cells grown in serum-free collagen gel culturesrdquoEndocrinology vol 116 no 1 pp 105ndash112 1985
[33] C J Ormandy R E Hall D L Manning et al ldquoCoexpres-sion and cross-regulation of the prolactin receptor and sexsteroid hormone receptors in breast cancerrdquo Journal of ClinicalEndocrinology and Metabolism vol 82 no 11 pp 3692ndash36991997
[34] K A OrsquoLeary F Jallow D E Rugowski et al ldquoProlactinactivates ER120572 in the absence of ligand in female mammarydevelopment and carcinogenesis in vivordquo Endocrinology vol154 pp 4483ndash4492 2013
[35] B W Park K S Kim M K Heo et al ldquoExpression ofestrogen receptor-120573 in normal mammary and tumor tissues isit protective in breast carcinogenesisrdquo Breast Cancer Researchand Treatment vol 80 no 1 pp 79ndash85 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 BioMed Research International
assistance This work was partially supported by grantsfrom Instituto Nacional del Cancer (Ministerio de SaludArgentina) and fromCONICET (Consejo Nacional de Inves-tigaciones Cientıficas y Tecnicas) Argentina
References
[1] P K Chakravarty and D K Sinha ldquoPregnancy inducedmammary tumor specific effector cells are present long afterparturition in a breast cancer model in ratsrdquo Cancer Letters vol154 no 1 pp 1ndash7 2000
[2] J Russo G A Balogh and I H Russo ldquoFull-term pregnancyinduces a specific genomic signature in the human breastrdquoCancer Epidemiology Biomarkers amp Prevention vol 17 no 1 pp51ndash66 2008
[3] J Russo L K Tay and I H Russo ldquoDifferentiation of themammary gland and susceptibility to carcinogenesisrdquo BreastCancer Research and Treatment vol 2 no 1 pp 5ndash73 1982
[4] OMaller KCHansen T R Lyons et al ldquoCollagen architecturein pregnancy-induced protection from breast cancerrdquo Journal ofCell Science vol 126 part 18 pp 4108ndash4110 2013
[5] S K Siwko J Dong M T Lewis H Liu S G Hilsenbeckand Y Li ldquoEvidence that an early pregnancy causes a persistentdecrease in the number of functional mammary epithelial stemcellsmdashimplications for pregnancy-induced protection againstbreast cancerrdquo Stem Cells vol 26 no 12 pp 3205ndash3209 2008
[6] Y Misra P A Bentley J P Bond S Tighe T Hunter and FZhao ldquoMammary gland morphological and gene expressionchanges underlying pregnancy protection of breast cancertumorigenesisrdquo Physiological Genomics vol 44 no 1 pp 76ndash882012
[7] J Russo and I H Russo ldquoInfluence of differentiation and cellkinetics on the susceptibility of the rat mammary gland tocarcinogenesisrdquo Cancer Research vol 40 no 8 pp 2677ndash26871980
[8] R C Guzman J Yang L Rajkumar G Thordarson X Chenand S Nand ldquoHormonal prevention of breast cancer mim-icking the protective effect of pregnancyrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 5 pp 2520ndash2525 1999
[9] A R Bovns R Buchan E N Cole A P M Forrest and KGrifiths ldquoBasal prolactin blood levels in three strains of ratwith differing incidence of 712-Dimethylbenz (a) anthraceneinduced mammary tumoursrdquo European Journal of Cancer vol9 no 3 pp 169ndash171 1973
[10] C W Welsch and H Nagasawa ldquoProlactin and murine mam-mary tumorigenesis a reviewrdquo Cancer Research vol 37 no 4pp 951ndash963 1977
[11] H Cohen I Sabbagh P Guillaumot J Bertrand M Robert-son and H G Friesen ldquoRelationship between bioassay andradioimmunoassay measurement of prolactin in the IPL rat ahypoprolactinemic rat strainrdquo Biology of Reproduction vol 38no 1 pp 84ndash90 1988
[12] S R Valdes A B Penissi R P Deis and G A JahnldquoHormonal profile and reproductive performance in lactationdeficient (OFA hrhr) and normal (Sprague-Dawley) femaleratsrdquo Reproduction vol 133 no 4 pp 827ndash840 2007
[13] H Bazzi A Kljuic A M Christiano and A A PanteleyevldquoIntragenic deletion in theDesmoglein 4 gene underlies the skinphenotype in the Iffa Credo ldquohairlessrdquo ratrdquo Differentiation vol72 no 8 pp 450ndash464 2004
[14] H Kim A A Panteleyev C A B Jahoda Y Ishii and A MChristiano ldquoGenomic organization and analysis of the hairlessgene in four hypotrichotic rat strainsrdquoMammalianGenome vol15 no 12 pp 975ndash981 2004
[15] B Meyer H Bazzi V Zidek et al ldquoA spontaneous mutationin the desmoglein 4 gene underlies hypotrichosis in a newlanceolate hair rat modelrdquo Differentiation vol 72 no 9-10 pp541ndash547 2004
[16] R W Caron and R P Deis ldquoParticipation of opioid and sero-toninergic systems in prolactin secretion induced by hypotha-lamic action of estradiolrdquoNeuroendocrinology vol 64 no 2 pp124ndash130 1996
[17] R W Caron and R P Deis ldquoEstradiol implants in the arcuatenucleus induce lactogenesis in virgin rats Role of progesteronerdquoLife Sciences vol 62 no 3 pp 229ndash237 1997
[18] G R Morel R W Caron G M Console et al ldquoEstrogeninhibits tuberoinfundibular dopaminergic neurons but does notcause irreversible damagerdquo Brain Research Bulletin vol 80 no6 pp 347ndash352 2009
[19] R W Caron A M Salicioni and R P Deis ldquoMifepristonetreatment demonstrates the participation of adrenal glucocorti-coids in the regulation of oestrogen-induced prolactin secretionin ovariectomized ratsrdquo Journal of Steroid Biochemistry andMolecular Biology vol 48 no 4 pp 385ndash389 1994
[20] R P Deis D G Carrizo and G A Jahn ldquoSuckling-inducedprolactin release potentiates mifepristone-induced lactogenesisin pregnant ratsrdquo Journal of Reproduction and Fertility vol 87no 1 pp 147ndash153 1989
[21] R W Caron C A Jahn and R P Deis ldquoLactogenic actionsof different growth hormone preparations in pregnant andlactating ratsrdquo Journal of Endocrinology vol 142 no 3 pp 535ndash545 1994
[22] M Edery L Houdebine J Djiane and P A Kelly ldquoStudies of 120573-casein content of normal and neoplastic rat mammary tissuesby a homologous radioimmunoassayrdquo Molecular and CellularEndocrinology vol 34 no 2 pp 145ndash151 1984
[23] L E Bussmann and R P Deis ldquo120574-Glutamyltransferase activityin mammary gland of pregnant rats and its regulation by ovar-ian hormones prolactin and placental lactogenrdquo BiochemicalJournal vol 223 no 1 pp 275ndash277 1984
[24] J Russo and I H Russo ldquoAtlas and histologic classification oftumors of the rat mammary glandrdquo Journal of Mammary GlandBiology and Neoplasia vol 5 no 2 pp 187ndash200 2000
[25] C M Lopez-Fontana C V Sasso M E Maselli et al ldquoExperi-mental hypothyroidism increases apoptosis in dimethylbenzan-thracene-inducedmammary tumorsrdquoOncologyReports vol 30no 4 pp 1651ndash1660 2013
[26] C M Lopez-Fontana M E Maselli A M Salicioni and R WCaron ldquoThe inhibitory effect of progesterone on lactogenesisduring pregnancy is already evident by mid- to late gestation inrodentsrdquo Reproduction Fertility and Development vol 24 no 5pp 704ndash714 2012
[27] C Puissant and L M Houdebine ldquoAn improvement of thesingle-step method of RNA isolation by acid guanidiniumthiocyanate-phenol-chloroform extractionrdquo BioTechniques vol8 no 2 pp 148ndash149 1990
[28] G Thordarson E Jin R C Guzman S M Swanson S Nandiand F Talamantes ldquoRefractoriness to mammary tumorigenesisin parous rats is it caused by persistent changes in the hormonalenvironment or permanent biochemical alterations in themammary epitheliardquo Carcinogenesis vol 16 no 11 pp 2847ndash2853 1995
BioMed Research International 11
[29] G Thordarson K van Horn R C Guzman S Nandi and FTalamantes ldquoParous rats regain high susceptibility to chemi-cally induced mammary cancer after treatment with variousmammotropic hormonesrdquo Carcinogenesis vol 22 no 7 pp1027ndash1033 2001
[30] L Sivaraman O M Conneely D Medina and B W OMalleyldquop53 is a potential mediator of pregnancy and hormone-induced resistance to mammary carcinogenesisrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 98 no 22 pp 12379ndash12384 2001
[31] J Frasor and G Gibori ldquoProlactin regulation of estrogenreceptor expressionrdquo Trends in Endocrinology and Metabolismvol 14 no 3 pp 118ndash123 2003
[32] M Edery W Imagawa L Larson and S Nandi ldquoRegulation ofestrogen and progesterone receptor levels in mouse mammaryepithelial cells grown in serum-free collagen gel culturesrdquoEndocrinology vol 116 no 1 pp 105ndash112 1985
[33] C J Ormandy R E Hall D L Manning et al ldquoCoexpres-sion and cross-regulation of the prolactin receptor and sexsteroid hormone receptors in breast cancerrdquo Journal of ClinicalEndocrinology and Metabolism vol 82 no 11 pp 3692ndash36991997
[34] K A OrsquoLeary F Jallow D E Rugowski et al ldquoProlactinactivates ER120572 in the absence of ligand in female mammarydevelopment and carcinogenesis in vivordquo Endocrinology vol154 pp 4483ndash4492 2013
[35] B W Park K S Kim M K Heo et al ldquoExpression ofestrogen receptor-120573 in normal mammary and tumor tissues isit protective in breast carcinogenesisrdquo Breast Cancer Researchand Treatment vol 80 no 1 pp 79ndash85 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 11
[29] G Thordarson K van Horn R C Guzman S Nandi and FTalamantes ldquoParous rats regain high susceptibility to chemi-cally induced mammary cancer after treatment with variousmammotropic hormonesrdquo Carcinogenesis vol 22 no 7 pp1027ndash1033 2001
[30] L Sivaraman O M Conneely D Medina and B W OMalleyldquop53 is a potential mediator of pregnancy and hormone-induced resistance to mammary carcinogenesisrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 98 no 22 pp 12379ndash12384 2001
[31] J Frasor and G Gibori ldquoProlactin regulation of estrogenreceptor expressionrdquo Trends in Endocrinology and Metabolismvol 14 no 3 pp 118ndash123 2003
[32] M Edery W Imagawa L Larson and S Nandi ldquoRegulation ofestrogen and progesterone receptor levels in mouse mammaryepithelial cells grown in serum-free collagen gel culturesrdquoEndocrinology vol 116 no 1 pp 105ndash112 1985
[33] C J Ormandy R E Hall D L Manning et al ldquoCoexpres-sion and cross-regulation of the prolactin receptor and sexsteroid hormone receptors in breast cancerrdquo Journal of ClinicalEndocrinology and Metabolism vol 82 no 11 pp 3692ndash36991997
[34] K A OrsquoLeary F Jallow D E Rugowski et al ldquoProlactinactivates ER120572 in the absence of ligand in female mammarydevelopment and carcinogenesis in vivordquo Endocrinology vol154 pp 4483ndash4492 2013
[35] B W Park K S Kim M K Heo et al ldquoExpression ofestrogen receptor-120573 in normal mammary and tumor tissues isit protective in breast carcinogenesisrdquo Breast Cancer Researchand Treatment vol 80 no 1 pp 79ndash85 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom





























