Embed Size (px)
Citation preview

Requirement for multiple domains of the protein arginine methyltransferase CARM1 in
its transcriptional coactivator function
CATHERINE TEYSSIER1,‡, DAGANG CHEN1,‡,# AND MICHAEL R.
STALLCUP1,2*
Departments of Pathology1 and of Biochemistry and Molecular Biology2, University of
Southern California, Los Angeles, California 90089.
Running title: Functional sub-domains of coactivator CARM1
‡ These authors contributed equally.
# Current address: Deltagen, Inc., 1003 Hamilton Court, Menlo Park, CA 94025-1422
*Corresponding author
Michael R. Stallcup
Department of Pathology, HMR 301
Univ. of Southern California
2011 Zonal Ave.
Los Angeles, CA 90089-9092
Teyssier et al 9-10-02
1
Copyright 2002 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on September 25, 2002 as Manuscript M207623200 by guest on A
pril 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Phone: (323) 442-1289; Fax: (323) 442-3049
E-mail: [email protected]
Teyssier et al 9-10-02
2
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

ABSTRACT
The p160 coactivator complex plays a critical role in transcriptional activation by
nuclear receptors and possibly other classes of DNA-binding transcriptional activators.
The complex contains at least one of the three p160 coactivators (SRC-1, GRIP1/TIF2,
or pCIP/RAC3/ACTR/AIB1/TRAM1), a histone acetyltransferase such as CBP or p300,
and the histone methyltransferase CARM1 (coactivator associated arginine
methyltransferase 1). Methylation of histone H3 and possibly other proteins in the
transcription initiation complex by CARM1 occurs along with acetylation of histones and
other proteins by CBP and p300 to help remodel chromatin structure and recruit RNA
polymerase II. Here we show that other domains of CARM1 are required for the
coactivator function of CARM1 in addition to the methyltransferase activity. The
methyltransferase, GRIP1 binding, and homo-oligomerization activities all reside in the
central region of CARM1, which is highly conserved among the entire protein arginine
methyltransferase family. In addition to this conserved domain, the unique N-terminal
and C-terminal regions of CARM1 were also required for enhancement of transcriptional
activation by nuclear receptors. While the N-terminal region has no known activity at
present, the C-terminal part of CARM1 contains an autonomous activation domain,
suggesting that it interacts with other proteins that help to mediate CARM1 coactivator
function.
Teyssier et al 9-10-02
3
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

INTRODUCTION
Activation of transcription by DNA-binding transcriptional activator proteins is
mediated by coactivators which locally remodel chromatin structure and recruit RNA
polymerase II and its transcription initiation complex to the promoter. Members of the
nuclear receptor (NR)1 family of transcriptional activator proteins, which include the
receptors for steroid and thyroid hormones, retinoids and vitamin D, as well as so-called
orphan receptors (1-3), recruit several different complexes of coactivator proteins to their
target gene promoters (4-8).
One coactivator complex which plays a central role in mediating transcriptional
activation includes at least one of the three related 160-kDa proteins commonly referred
to as p160 coactivators (SRC-1, GRIP1/TIF2, and pCIP/RAC3/ACTR/AIB1/TRAM1).
The p160 coactivators bind directly and in a ligand-dependent manner to the C-terminal
AF2 activation domains of NRs through three LXXLL motifs (where L is a leucine and
X, any amino acid) located in the central part of the p160 polypeptide chain. The C-
terminal region of the p160 coactivators can also interact with the N-terminal AF1
activation domains of some NRs (9-11). The p160 coactivators contribute to transcriptional
activation by bringing other associated coactivator proteins with them to the promoter.
The p160 coactivator complex includes either of the two related proteins p300 and CBP,
which bind to the AD1 activation domain of p160 coactivators (12-14) and function as
coactivators for many DNA-binding transcriptional activators, including NRs (15). Recent
studies have confirmed hormone-dependent recruitment of p160 coactivators, CBP, and
p300 to promoters activated by NRs (16-19). CBP and p300 contribute to chromatin remodeling
by acetylating histones, and also acetylate other components of the transcription initiation
Teyssier et al 9-10-02
4
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

complex (16,20-22). CBP and p300 can also bind directly to basal transcription factors and may
thereby help to assemble the transcription initiation complex (12). Thus multiple domains of
CBP and p300 apparently contribute to chromatin remodeling and recruitment/activation
of RNA polymerase II.
The activation domain AD2, located at the C-terminus of p160 factors, binds
CARM1 which belongs to a family of previously identified arginine specific protein
methyltransferases (PRMTs) (23). CARM1 enhances nuclear receptor function in a p160-
dependent manner in transient transfection assays. CARM1, p300/CBP, and a p160
coactivator can also form a ternary complex which functions synergistically to enhance
NR function and requires the methyltransferase activity of CARM1 to do so (24,25). CARM1
methylates histone H3 at Arg17 and Arg26 in vitro (26), and chromatin immunoprecipitation
studies indicated that CARM1 is specifically recruited to steroid hormone regulated
promoters in vivo in response to the hormone and methylates histone H3 as part of the
transcription initiation process (27,28).
PRMTs are homo-dimeric or homo-oligomeric proteins (29-31) which transfer the methyl
group from S-adenosyl-L-methionine (AdoMet) to the guanidino group of arginines in
protein substrates (32). The enzymatic activity is supported by a catalytic core domain which
is highly conserved among PRMT family members and contains the AdoMet and
arginine binding sites and a barrel-like domain (29). In addition to the conserved core region,
each methyltransferase has a unique N-terminal region of variable size. However,
CARM1 is the only member of the family to harbor unique domains at its N-terminus
and C-terminus. The contribution of these additional domains to the function of the
PRMTs is unknown, although some studies suggested that they may be involved in the
Teyssier et al 9-10-02
5
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

specificity of protein substrate binding (29,30,33).
Previous studies suggested that while the methyltransferase activity of CARM1 is
required for coactivator function, other unspecified domains also contribute (25).
Furthermore, because the coactivator function of CARM1 depends on p160 coactivators (23,24),
the p160 binding site of CARM1 is also presumably required, but its location has not
been determined. To better understand which activities and domains of CARM1
contribute to its coactivator function, we defined the locations of the methyltransferase,
p160 binding, and homo-oligomerization domains, as well as an autonomous activation
domain, with respect to the conserved central domain (amino acids 150-480) and the
unique N-terminal and C-terminal regions of CARM1. By testing deletion mutants of
CARM1 lacking various domains, we identified multiple regions of CARM1 which are
required for its coactivator function along with the methyltransferase and GRIP1 binding
activities.
MATERIALS AND METHODS
Plasmids. Proteins with N-terminal hemagglutinin A (HA) epitope tags were
expressed in transient mammalian cell transfections and in vitro from vector pSG5.HA
which has SV40 and T7 promoters (23). pSG5.HA GRIP1 (9) and pSG5.HA CARM1 (23) were
previously described, as were pHE0, encoding human estrogen receptor (ER) α (34); the
luciferase reporter plasmids MMTV(ERE)-LUC (20) and GK1, which is controlled by Gal4
response elements (10); and the β-galactosidase (β-gal) reporter plasmid RSV.β-gal (35).
CARM1 deletion mutants were constructed by inserting PCR-amplified CARM1 cDNA
fragments flanked by a 5’ EcoRI site and a 3’ BglII site into EcoRI and BglII sites of
Teyssier et al 9-10-02
6
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

pSG5.HA and into EcoRI and BamHI sites of pM (to make Gal4 DBD fusions) or pVP16
(Clontech) (to make VP16 fusions). pM.GRIP1 (encoding Gal4DBD-GRIP1) was
described previously (9), and pCMV.p300 was kindly provided by Dr. T.-P. Yao (Duke
University). Bacterial expression vectors for glutathione S-transferase (GST) fused to
GRIP1 AD2 (consisting of the GRIP1 C-terminal amino acids 1121-1462) and CARM1
were described previously (9,23).
Protein-protein interaction in vitro. To measure binding of 35S-labeled proteins
synthesized in vitro to GST fusion proteins on glutathione-Sepharose beads, GST pull-
down assays were performed as previously described (36), with the following exceptions:
binding was conducted overnight at 4 °C with 20 µl of the in vitro synthesis reaction in a
150-µl total volume of NETN buffer (0.1% NP-40, 1 mM EDTA, 20 mM Tris pH 8.0,
100 mM NaCl) containing Complete protease inhibitor cocktail (Roche).
Cell Culture and Transfection. Transfections of CV-1 cells (37) were perfomred in
12-well dishes as described previously (25) with 1 µg of total DNA per well. Where
indicated, medium was supplemented with 20 nM estradiol (E2) during the last 30 h of
growth. Luciferase and β-galactosidase activities are shown as the mean and range of
variation of two transfected cell cultures.
Immunoprecipitation of CARM1 from transfected cells and in vitro methylation
assays. COS7 cells (37) were maintained in DMEM supplemented with 10% fetal bovine
serum. One million cells were seeded into 10-cm diameter cell dishes and transfected
with Superfect (Qiagen) according to manufacturer’s protocol with 5 µg of pSG5.HA
plasmid encoding CARM1 wild type or CARM1 mutants. After transfection, cells were
grown in DMEM with 10% fetal bovine serum for 40 h before harvest. Cells were
Teyssier et al 9-10-02
7
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

harvested in lysis buffer containing 50 mM Tris-HCl (pH 8.0), 120 mM NaCl, 0.1%
NP40 and Complete protease inhibitor cocktail (Roche). Cell lysates were frozen at -80
°C, thawed on ice, and clarified by centrifugation before incubation overnight at 4 °C
with monoclonal antibody (clone 3F10, Roche) against HA tag. Pre-blocked protein G-
Sepharose (Amersham Pharmacia Biotech) was then added for 2 h at 4 °C.
Immunoprecipitates were recovered by rapid centrifugation, washed three times with
NETN containing 0.1% NP40 and resuspended in HMT Buffer (20 mM Tris-HCl, pH
8.0, 200 mM NaCl, 0.4 mM EDTA). In vitro histone methylation assays were performed
as follows: 3 µg of histone H3 (Roche) was incubated with immunoprecipitated
methyltransferases and 7 µM S-adenosyl-L-[3H-methyl]methionine (specific activity
14.7 Ci/mmol) in 30 µl HMT buffer for 1 h at 30 °C. Reactions were stopped by addition
of SDS loading buffer and analyzed by 15% SDS-PAGE and fluorography.
Immunoblots. Ten percent of the immunoprecipitated methyltransferases from the
transfected cells (see above) were analyzed by SDS-PAGE on 12% gels. Immunoblotting
was performed as described previously (38) with rat monoclonal antibody 3F10 against the
HA epitope (Roche) at 100 ng/ml as the primary antibody and horseradish peroxidase-
conjugated anti-rat immunoglobin G (Santa Cruz Biotechnology) at 160 ng/ml (1:2,500
dilution) as the secondary antibody.
RESULTS
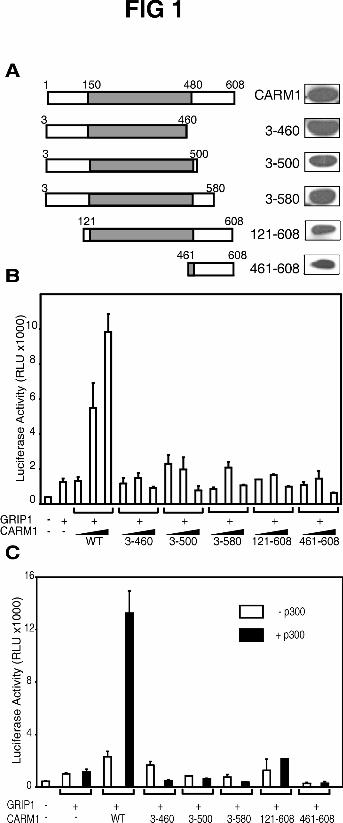
Domains of CARM1 involved in coactivator activity. To determine which parts of
CARM1 are required for its coactivator function, N-terminal and C-terminal truncations
were made near the boundaries between the conserved central domain of CARM1 and its
Teyssier et al 9-10-02
8
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

unique N-terminal (amino acids 1-150) and C-terminal (amino acids 480-608) regions
(Fig. 1A). The coactivator activity of CARM1 and its mutants were tested with ER by
transient transfection in CV-1 cells under two different conditions: with relatively high
levels of transfected ER expression vector, where CARM1 cooperates with a p160
coactivator (23); and at very low levels of transfected ER expression vector, where CARM1
functions synergistically with a p160 coactivator and p300, such that all three of these
coactivators must be co-expressed with ER to achieve efficient activation of an
estrogen-dependent reporter gene (25). In both cases, the activity observed was shown
previously to be completely dependent on the exposure of the cells to estradiol to activate
ER.
At the higher level of ER vector (5 ng), GRIP1 expression enhanced reporter gene
activation by ligand bound ER, and co-expression of full length CARM1 resulted in a
further enhancement approximately in proportion to the amount of CARM1 expression
vector used (Fig. 1B). However, mutants lacking the N-terminal part of CARM1 (mutant
121-608) or the C-terminal part (mutants 3-460, 3-500, 3-580) had no effect on the
reporter gene expression mediated by ER and GRIP1, suggesting that both ends of
CARM1 contribute to its coactivator activity.
With a low concentration of ER expression vector (0.1 ng), p300 had no effect on
the transcriptional activity observed with ER and GRIP1, and CARM1 (in the absence of
p300) caused an enhancement of only 2-fold (Fig. 1C). However as previously described
(25), expression of both CARM1 and p300 with GRIP1 resulted in a dramatic increase of
ER-dependent reporter gene activity. Deletions of the unique N- or C-terminal part of
CARM1 severely impaired the synergistic effect.
Teyssier et al 9-10-02
9
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Thus the unique N-terminal and C-terminal regions of CARM1 are required for
coactivator activity with ER under both tested conditions. The expression level of each
mutant was similar to that of full length CARM1, with the exception of CARM1(121-
608) (Fig 1A). The lack of coactivator activity of the CARM1(121-608) fragment could
be partly due to its lower expression. We also recognized that the deletions may disrupt
the protein structure and thereby impair the function of domains that are still present in
the mutant protein. To test this possibility, we examined whether other known functions
of CARM1 remained intact in the various mutants. This line of experimentation also
allowed us to assign specific functions of CARM1 to specific domains of the protein and
thereby to explore the mechanisms by which the unique N-terminal and C-terminal
regions and the conserved central region of CARM1 contribute to the coactivator activity.
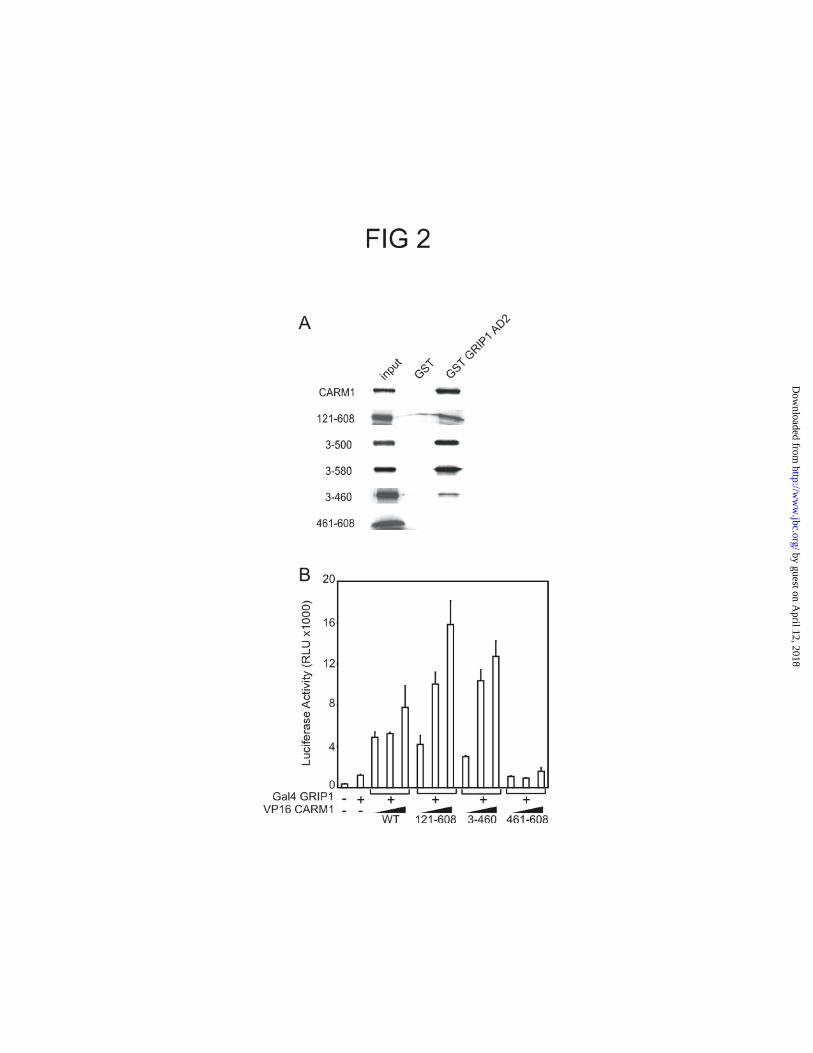
The GRIP1 binding domain of CARM1 is located in the conserved central region.
The lack of coactivator activity of the CARM1 mutants could be due to their inability to
bind GRIP1. Binding between GRIP1 and CARM1 is required for CARM1 coactivator
function (24), and the GRIP1 binding domain of CARM1 has not been mapped. Full length
CARM1 and its deletion mutants were translated in vitro in the presence of
[35S]methionine and incubated with either GST or GST-GRIP1 AD2 fusion protein
(consisting of the GRIP1 C-terminal amino acids 1122-1462), which were preloaded on
glutathione Sepharose beads. None of the CARM1 proteins bound to GST, but full length
CARM1 and several CARM1 fragments bound GST-GRIP1 AD2 (Fig. 2A). Deletion of
the N-terminus (amino acids 1 to 120) or the C-terminus (amino acids 501-608 or 581-
608) of CARM1 did not impair binding to GRIP1 AD2, showing that these domains are
not required for the in vitro interaction. The CARM1 mutant lacking amino acids 461 to
Teyssier et al 9-10-02
10
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

608 was still retained by GST-GRIP1 AD2 but to a lesser extent. However, the CARM1
C-terminal region alone was not able to bind GRIP1 AD2. A mammalian two-hybrid
assay confirmed these results (Fig. 2B). These results localized the GRIP1 binding
domain of CARM1 within the central conserved domain (amino acids 121-460) and
demonstrated that the unique N- and C-terminal regions of CARM1 are neither
necessary nor sufficient for the interaction with GRIP1.
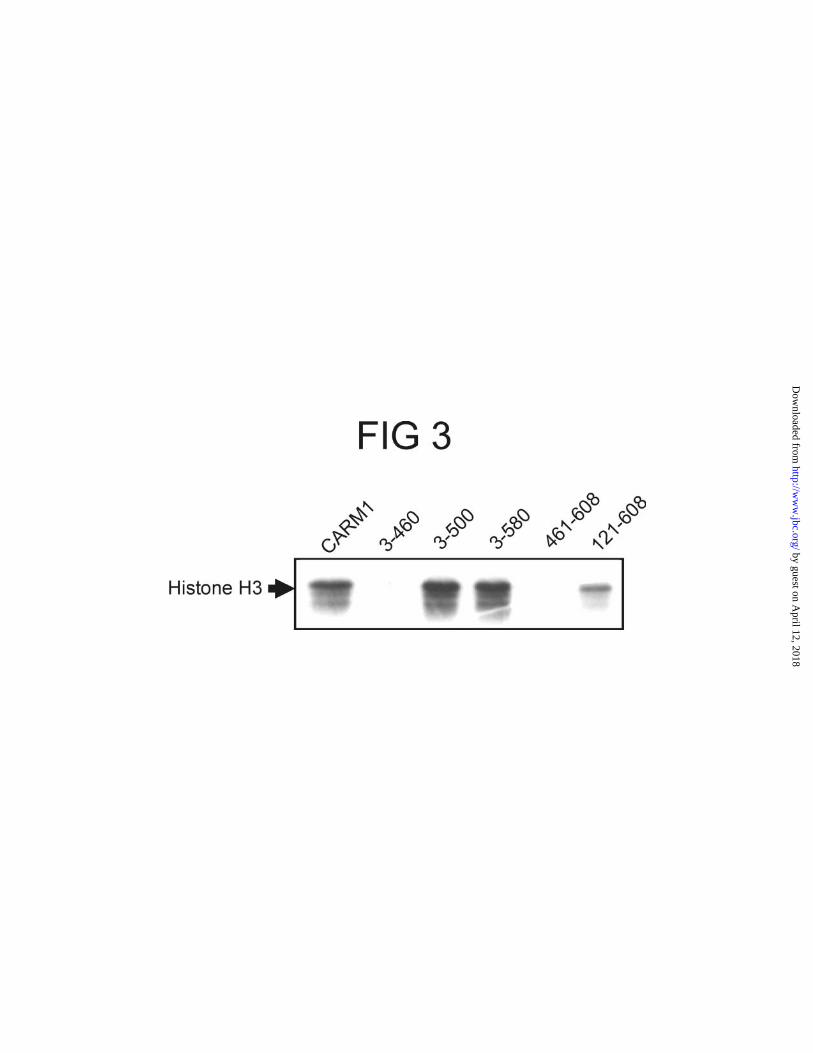
The unique CARM1 ends are not required for the methyltransferase activity. Full
length CARM1 and its deletion mutants were expressed in COS7 cells by transient
transfection, isolated by immunoprecipitation, and incubated with histone H3 in the
presence of [3H-methyl]AdoMet; methylated histone H3 was detected by SDS-PAGE
and fluorography (Fig. 3). The mutant lacking the N-terminal part of CARM1 (1-120) or
the C-terminal region (501-608) still methylated histone H3 efficiently, showing that
these unique domains were not required for the enzymatic activity of CARM1, at least in
vitro. The lower activity of CARM1 121-608 was due to its lower expression level (Fig.
1A). However, the mutant lacking amino acids 461-608, which includes a small portion
of the central conserved domain, was inactive. The unique C-terminal region (amino
acids 461-608) by itself did not exhibit any enzymatic activity. Thus, the
methyltransferase activity resides within amino acids 121-500.
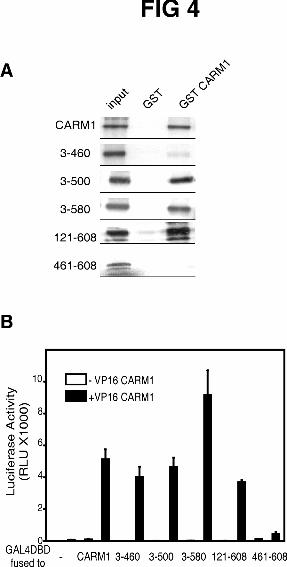
The conserved central part of CARM1 also contains its homo-oligomerization
domain. To localize the homo-oligomerization domain, we tested the ability of the
CARM1 deletion mutants to interact with wild type CARM1. All mutants lacking N-
terminal and C-terminal amino acids were retained by the bead-bound GST-CARM1
fusion protein, although CARM1(3-460) bound very weakly compared with wild type
Teyssier et al 9-10-02
11
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CARM1 (Fig. 4A); thus the unique ends of CARM1 are not required for its homo-
oligomerization in vitro. Furthermore, by itself the C-terminal fragment (amino acids
461-608) did not bind to GST-CARM1. Mammalian two-hybrid assays produced very
similar results in vivo (Fig. 4B). We, therefore, located the CARM1 homo-
oligomerization domain in the conserved central part of the protein along with the
methyltransferase and GRIP1 binding activities.
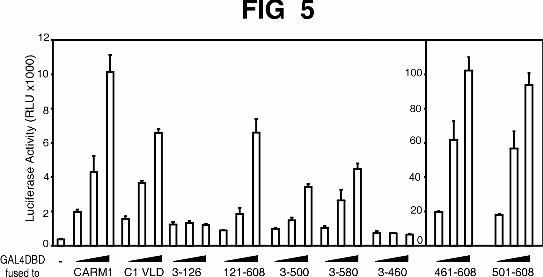
The unique C-terminal region of CARM1 is a transcriptional activation domain.
Since the unique N-terminal and C-terminal regions of CARM1 were important for its
coactivator function but played no role in its methyltransferase, GRIP1-binding, or
homo-oligomerization activities, we tested whether these unique regions might contain
an autonomous activation function. CARM1 mutants fused to the Gal4 DBD were tested
for their ability to activate expression of a reporter gene controlled by Gal4 response
elements (Fig. 5). Fusion of full length CARM1 to Gal4 DBD enhanced the reporter gene
expression, indicating the presence of an autonomous transactivation domain somewhere
in the CARM1 protein. A mutant deficient in methyltransferase activity (C1 VLD) (23) fused
to Gal4 DBD also increased the transcription driven by Gal4 response elements. This
suggests that the methyltransferase activity of CARM1 is not necessary for the observed
transcriptional activation activity. A mutant constituted only by the N-terminal part of
CARM1 (amino acids 3-126) had little or no ability to increase the reporter gene
expression. The mutant 121-608 was almost as effective as full length CARM1, thus
showing that the N-terminal part was not involved in the activity. Mutants lacking the
C-terminal part of CARM1 (mutants 3-500; 3-580) were also active but to a lesser
extent than full length CARM1. Moreover, deletion of residues 461 to 608 totally
Teyssier et al 9-10-02
12
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

abolished the autonomous transactivation activity of CARM1. Finally, the CARM1
fragments 461-608 and 501-608 exhibited activity ten times that of wild type CARM1,
indicating that CARM1 contains a strong autonomous activation domain in its unique C-
terminal region (Fig. 5, right panel).
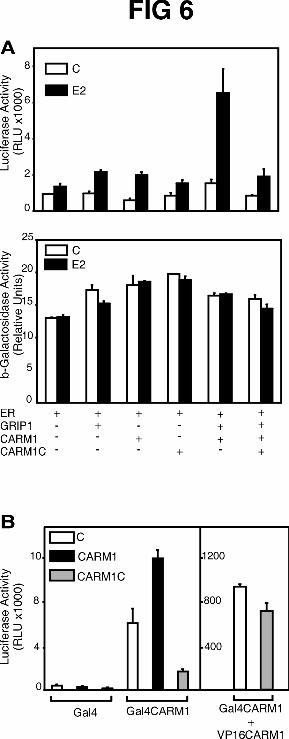
The C-terminal activation domain of CARM1 may contribute to coactivator
function through protein-protein interactions with some important component of the
transcription machinery. If so, over-expression of the isolated C-terminal domain might
inhibit the coactivator function of full length CARM1 by competing with CARM1 for the
interaction with this transcription machinery component. CARM1-C (amino acids 461-
608) strongly inhibited the coactivator effect of CARM1 on the hormone-dependent,
ER-mediated activation of reporter gene expression (Fig. 6A, upper panel). CARM1-C
had little or no effect on the basal ER activity observed in the absence of hormone. The
specificity of the inhibitory effect was also demonstrated by the fact that CARM1-C had
no effect on the expression of a RSV promoter-driven β-galactosidase reporter gene,
which was tested in a parallel experiment (lower panel). CARM1-C also inhibited the
autonomous transactivation activity of full length CARM1 fused to Gal4 DBD (Fig. 6B,
left panel). In contrast full length CARM1, used as a positive control, enhanced the
activity of the Gal4-CARM1 fusion protein, presumably through homo-oligomerization.
In a mammalian two-hybrid assay, the interaction of Gal4-CARM1 with VP16-CARM1
was only slightly inhibited by CARM1-C (Fig. 6B, right panel), consistent with our
previous finding that CARM1-C cannot bind to full length CARM1 (Fig. 4). Thus, the
negative effect of CARM1-C on the autonomous activation activity of CARM1 is not
due to a disruption of the homo-oligomer but could rather be due to a competition of
Teyssier et al 9-10-02
13
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CARM1-C with CARM1 for the interaction with another transcription factor that binds
CARM1-C.
DISCUSSION
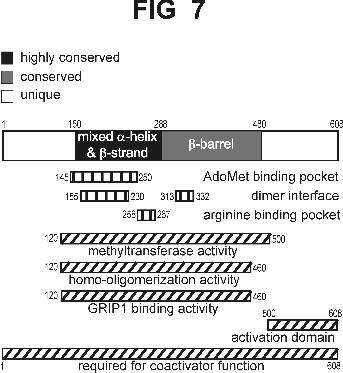
Multiple functions of the conserved central region of CARM1. CARM1 belongs
to the PRMT family of arginine-specific protein methyltransferases which share a
conserved core region of about 330 amino acids that contains the methyltransferase
activity. X-ray crystallography of mammalian PRMT3 and yeast Rmt1/Hmt1
demonstrated that the conserved region forms two separate structural domains which
combine to form the active enzyme (29,39) (Fig. 7). The N-terminal part, which is the
most highly conserved in primary amino acid sequence among family members, is
composed of mixed α-helices and β-strands. The C-terminal part of the core forms an
elongated nine-stranded β-barrel structure. Homo-dimerization is apparently required to
form an active enzyme. The dimer interface is formed by reciprocal contact between the
α/β region of one monomer and a tri-helical arm extending from the surface of the β-
barrel structure of the other monomer. The AdoMet binding pocket is formed by the α/β
region. The arginine residue of the protein substrate binds in an acidic pocket containing
two glutamate residues which interact directly with the two terminal amino groups of the
arginine side chain. The portions of the protein substrate surrounding the target arginine
residue are predicted to fit in a groove between the α/β region and the β-barrel structure
(29,39).
As expected from the three-dimensional structure and the high degree of
conservation among PRMT members, our studies located the methyltransferase and
Teyssier et al 9-10-02
14
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

homo-oligomerization activities of CARM1 approximately within the conserved region
(amino acids 150-480 of CARM1) (Figs. 3, 4, & 7). A C-terminal deletion to amino acid
460, which removed the last β-strand of the β-barrel structure, eliminated the
methyltransferase activity and thus demonstrated that the entire conserved barrel structure
is required for methyltransferase activity. The C-terminal β-strand may contribute to
structural integrity of the entire domain or could help to form the protein substrate
binding groove. The same deletion mutant retained partial-to-full homo-oligomerization
and GRIP1 binding activity (Figs. 2 & 4), indicating that structural integrity was not
completely disrupted. Our finding that the central conserved region of CARM1 also
contained the GRIP1 binding activity (Fig. 2 & 7) is consistent with previous findings
that multiple members of the PRMT family can bind to the C-terminal region of GRIP1
(25,36). The GRIP1 binding activity of CARM1 is undoubtedly required for the
coactivator function of CARM1, since we previously showed that the presence of GRIP1
and its C-terminal CARM1-binding region are required for the coactivator function of
CARM1 (23,24).
While CARM1 shares homology with the PRMT family throughout the
methyltransferase domain, CARM1 has a unique set of protein substrates, including
histone H3 and p300/CBP (23,40). In addition, CARM1 is the only PRMT member tested
to date which can cooperate synergistically with p300, CBP, or p/CAF to enhance
transcriptional activation by NRs (25). Previous studies have shown that the coactivator
function of CARM1 depends on its methyltransferase activity (23,25). Moreover,
chromatin immunoprecipitation assays demonstrated that steroid hormones stimulate
recruitment of CARM1 and methylation of histone H3 in a CARM1-specific manner at
Teyssier et al 9-10-02
15
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

promoters of stably integrated, steroid hormone-responsive genes (27,28). Thus, the unique
transcriptional coactivator function of CARM1 is at least partly due to its unique
methyltransferase substrate specificity.
Role of the unique N-terminal and C-terminal regions of CARM1 in its coactivator
function. Deletion of the unique N-terminal or C-terminal part of CARM1 totally
abolished its coactivator function (Fig. 1), but had no effect on its ability to bind GRIP1
(Fig. 2), methylate histone H3 (Fig. 3), or form homo-oligomers (Fig. 4). Thus the
unique N-terminal and C-terminal regions must contribute to the coactivator function of
CARM1 through a novel mechanism not involving any of these three activities. To date
little is known about the functions of the unique N-termini of PRMT family members;
these unique N-termini vary greatly in length as well as sequence (29). It has been proposed
that the relatively short N-terminus of mammalian PRMT1 and yeast Rmt1/Hmt1 may
contribute to methyltransferase substrate specificity by interacting with regions of the
substrate protein distinct from the sequence immediately surrounding the target arginine
residue (29,30). In addition, deletion of the relatively long unique N-terminus of PRMT3 altered
its substrate specificity in vitro (33). However, mutants of CARM1 lacking the unique N-
terminal or C-terminal regions appeared to be unaffected in their ability to methylate
histone H3 (Fig. 3) and several other protein substrates that we tested (our unpublished
results). We also found that the N-terminus of CARM1 did not contribute to the
autonomous transcriptional activation activity of CARM1 (Fig. 5). Thus the mechanism
by which the N-terminal region of CARM1 contributes to coactivator function remains
unclear.
While the unique C-terminal part of CARM1 was not required for the GRIP1
Teyssier et al 9-10-02
16
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

binding, methyltransferase, or homo-oligomerization activities, it contains a strong
autonomous activation domain (Fig. 5), which presumably explains why this domain is
necessary for the coactivator function of CARM1. CARM1 mutants lacking this domain
were almost devoid of a transcriptional activation activity when fused to Gal4 DBD,
indicating that the C-terminus is responsible for most or all of the autonomous
transactivation activity observed in full length CARM1. The ineffective autonomous
transcriptional activation activity associated with the mutants lacking the C-terminal
region indicates that the methyltransferase activity per se is not sufficient for
transcriptional activation (Fig. 5). PRMT1, which lacks a unique C-terminal domain and
has a very short unique N-terminal domain, also exhibits no autonomous activation
activity when fused to Gal4 DBD (36). Thus, although CARM1 methylates histone H3 and
PRMT1 methylates histone H4, the simple recruitment of a histone methyltransferase
activity to the promoter of a transient reporter gene is not sufficient to activate
transcription. This is also consistent with our findings that the unique terminal domains of
CARM1 are required in addition to the methyltransferase activity for the coactivator
function of CARM1.
As a model for CARM1 function, we propose that CARM1 is recruited to the
promoter through its interaction with the C-terminal domain of a p160 coactivator. The
autonomous activation activity of the CARM1 C-terminal domain collaborates with the
methyltransferase activity of the central domain and possibly an unknown activity in the
unique N-terminal domain, to mediate the coactivator function of CARM1. The
methyltransferase activity is responsible for the methylation of histone H3, which
presumably contributes to chromatin remodeling. In addition, CARM1 may methylate
Teyssier et al 9-10-02
17
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

other protein components of the transcription machinery. We propose that the
autonomous activation activity of the C-terminal domain is due to its ability to interact
with other proteins in the transcription machinery. For example, CARM1-C could
interact with a component of the basal transcription machinery and thereby help to recruit
RNA polymerase II; or CARM1-C could interact with another, currently unknown,
coactivator and thereby recruit or maintain the additional coactivator in the complex with
GRIP1, CARM1, and p300. The ability of the co-expressed C-terminal fragment of
CARM1 to inhibit the coactivator function of full length CARM1 (Fig. 6) supports our
proposal that this region binds an important factor that contributes to the transcriptional
activation process.
Thus, once bound to the promoter, CARM1 contributes to the transcriptional
activation process through multiple downstream signaling mechanisms, i.e. through
methylation of histones and possibly other proteins and through protein-protein
interactions mediated by the unique C-terminal and possibly N-terminal domains. The
use of multiple downstream signaling mechanisms by a single coactivator is not unique to
CARM1. The coactivators p300, CBP, and p/CAF have multiple protein-protein
interaction domains which also contribute to their coactivator function in collaboration
with their histone acetyltransferase activities (12).
ACKNOWLEDGMENTS.
We thank Daniel Gerke for technical assistance. This work was supported by United
States Public Health Service Grant DK55274 from the National Institutes of Health to
M.R.S.
Teyssier et al 9-10-02
18
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

REFERENCES
1. Beato, M., Herrlich, P., and Schutz, G. (1995) Cell 83, 851-857
2. Mangelsdorf, D. J., Thummel, C., Beato, M., Herrlich, P., Schutz, G., Umesono, K., Blumberg, B.,
Kastner, P., Mark, M., and Chambon, P. (1995) Cell 83, 835-839
3. Enmark, E. and Gustafsson, J. A. (1996) Mol.Endocrinol. 10, 1293-1307
4. Glass, C. K. and Rosenfeld, M. G. (2000) Genes Dev. 14, 121-141
5. Xu, L., Glass, C. K., and Rosenfeld, M. G. (1999) Curr.Opin.Genet.Dev. 9, 140-147
6. McKenna, N. J., Lanz, R. B., and O’Malley, B. W. (1999) Endocr.Rev. 20, 321-344
7. Freedman, L. P. (1999) Cell 97, 5-8
8. Leo, C. and Chen, J. D. (2000) Gene 245, 1-11
9. Ma, H., Hong, H., Huang, S. M., Irvine, R. A., Webb, P., Kushner, P. J., Coetzee, G. A., and Stallcup,
M. R. (1999) Mol.Cell. Biol. 19, 6164-6173
10. Webb, P., Nguyen, P., Shinsako, J., Anderson, C., Feng, W., Nguyen, M. P., Chen, D., Huang, S.
M., Subramanian, S., McKinerney, E., Katzenellenbogen, B. S., Stallcup, M. R., and Kushner, P. J.
(1998) Mol.Endocrinol. 12, 1605-1618
11. Onate, S. A., Boonyaratanakornkit, V., Spencer, T. E., Tsai, S. Y., Tsai, M. J., Edwards, D. P., and
O’Malley, B. W. (1998) J.Biol.Chem. 273, 12101-12108
12. Chen, H., Lin, R. J., Schiltz, R. L., Chakravarti, D., Nash, A., Nagy, L., Privalsky, M. L., Nakatani,
Y., and Evans, R. M. (1997) Cell 90, 569-580
13. Li, J., O’Malley, B. W., and Wong, J. (2000) Mol.Cell. Biol. 20, 2031-2042
14. Voegel, J. J., Heine, M. J., Tini, M., Vivat, V., Chambon, P., and Gronemeyer, H. (1998) EMBO J.
Teyssier et al 9-10-02
19
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

17, 507-519
15. Goodman, R. H. and Smolik, S. (2000) Genes Dev. 14, 1553-1577
16. Chen, H., Lin, R. J., Xie, W., Wilpitz, D., and Evans, R. M. (1999) Cell 98, 675-686
17. Shang, Y., Hu, X., DiRenzo, J., Lazar, M. A., and Brown, M. (2000) Cell 103, 843-852
18. Liu, Z., Wong, J., Tsai, S. Y., Tsai, M. J., and O’Malley, B. W. (2001) Proc.Natl.Acad.Sci.U.S.A 98,
12426-12431
19. Kim, M. Y., Hsiao, S. J., and Kraus, W. L. (2001) EMBO J. 20, 6084-6094
20. Korzus, E., Torchia, J., Rose, D. W., Xu, L., Kurokawa, R., McInerney, E. M., Mullen, T. M., Glass,
C. K., and Rosenfeld, M. G. (1998) Science 279, 703-707
21. Gu, W. and Roeder, R. G. (1997) Cell 90, 595-606
22. Imhof, A., Yang, X. J., Ogryzko, V. V., Nakatani, Y., Wolffe, A. P., and Ge, H. (1997) Curr.Biol. 7,
689-692
23. Chen, D., Ma, H., Hong, H., Koh, S. S., Huang, S. M., Schurter, B. T., Aswad, D. W., and Stallcup,
M. R. (1999) Science 284, 2174-2177
24. Chen, D., Huang, S. M., and Stallcup, M. R. (2000) J.Biol.Chem. 275, 40810-40816
25. Lee, Y. H., Koh, S. S., Zhang, X., Cheng, X., and Stallcup, M. R. (2002) Mol.Cell. Biol. 22, 3621-
3632
26. Schurter, B. T., Koh, S. S., Chen, D., Bunick, G. J., Harp, J. M., Hanson, B. L., Henschen-Edman,
A., Mackay, D. R., Stallcup, M. R., and Aswad, D. W. (2001) Biochemistry 40, 5747-5756
27. Ma, H., Baumann, C. T., Li, H., Strahl, B. D., Rice, R., Jelinek, M. A., Aswad, D. W., Allis, C. D.,
Hager, G. L., and Stallcup, M. R. (2001) Curr.Biol. 11, 1981-1985
Teyssier et al 9-10-02
20
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

28. Bauer, U. M., Daujat, S., Nielsen, S. J., Nightingale, K., and Kouzarides, T. (2002) EMBO Rep. 3,
39-44
29. Zhang, X., Zhou, L., and Cheng, X. (2000) EMBO J. 19, 3509-3519
30. McBride, A. E., Weiss, V. H., Kim, H. K., Hogle, J. M., and Silver, P. A. (2000) J.Biol.Chem. 275,
3128-3136
31. Rho, J., Choi, S., Seong, Y. R., Cho, W. K., Kim, S. H., and Im, D. S. (2001) J.Biol.Chem. 276,
11393-11401
32. Lee, H. W., Kim, S., and Paik, W. K. (1977) Biochemistry 16, 78-85
33. Frankel, A. and Clarke, S. (2000) J.Biol.Chem. 275, 32974-32982
34. Green, S., Kumar, V., Theulaz, I., Wahli, W., and Chambon, P. (1988) EMBO J. 7, 3037-3044
35. Hong, H., Kohli, K., Trivedi, A., Johnson, D. L., and Stallcup, M. R. (1996)
Proc.Natl.Acad.Sci.U.S.A. 93, 4948-4952
36. Koh, S. S., Chen, D., Lee, Y. H., and Stallcup, M. R. (2001) J.Biol.Chem. 276, 1089-1098
37. Gluzman, Y. (1981) Cell 23, 175-182
38. Ding, X. F., Anderson, C. M., Ma, H., Hong, H., Uht, R. M., Kushner, P. J., and Stallcup, M. R.
(1998) Mol.Endocrinol. 12, 302-313
39. Weiss, V. H., McBride, A. E., Soriano, M. A., Filman, D. J., Silver, P. A., and Hogle, J. M. (2000)
Nature Struct. Biol. 7, 1165-1171
40. Xu, W., Chen, H., Du, K., Asahara, H., Tini, M., Emerson, B. M., Montminy, M., and Evans, R. M.
(2001) Science 294, 2507-2511
Teyssier et al 9-10-02
21
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

FOOTNOTES
1 The abbreviations used are: AD, activation domain; AdoMet, S-adenosyl-L-
methionine; AF, activation function; β-gal, β-galactosidase; CARM1, coactivator
associated arginine methyltransferase 1; CARM1-C, CARM1 amino acids 461-608;
DBD, DNA binding domain; CBP, CREB (cAMP-response element-binding protein)-
binding protein; ER, estrogen receptor α; ERE, estrogen response element; GRIP1,
glucocorticoid receptor interacting protein 1; GST, glutathione S-transferase; HA,
hemagglutinin A; LUC, luciferase coding region; MMTV, mouse mammary tumor virus
promoter; NR(s), nuclear receptor(s); PRMT, protein arginine methyltransferase; RLU,
relative lights units.
FIGURE LEGENDS
FIG. 1. Coactivator function of CARM1 and its deletion mutants with ERα. A, A
schematic representation of CARM1 deletion mutants is shown on the left. The
expression level of each protein is shown on the right, as determined by immunoblot of
transfected COS7 cell extracts, using anti-HA tag antibodies. B, CV1 cells were
transiently transfected with MMTV(ERE)-LUC reporter plasmid (250 ng) and
expression vectors encoding ER (5 ng), GRIP1 (125 ng) and increasing concentrations
(100, 200, and 400 ng) of vectors encoding full length CARM1 or deletion mutants as
indicated. Each mutant protein is identified by the amino acids residues it contains.
Transfected cells were grown in culture medium with 20 nM estradiol, and extracts of the
harvested cells were tested for luciferase activity. C, Transfections were performed as in
B, but amounts of ER and coactivator expression vectors were: ER, 0.1 ng; GRIP1, 125
Teyssier et al 9-10-02
22
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

ng; wild type or mutant CARM1, 250 ng; and p300, 250 ng. The results presented are
from a single experiment representative of 4 independent experiments.
FIG. 2. Location of GRIP1 binding activity in conserved central region of CARM1. A,
[35S]Methionine-labeled CARM1 (wild type or mutant) was incubated with either GST or
GST-GRIP1AD2 fusion proteins preloaded on glutathione-coupled beads. Bound
proteins were eluted and analyzed by SDS-PAGE and fluorography. The input lanes
represent 10% of each 35S-labeled protein used in the binding assay. B, CV1 cells were
transiently transfected with 250 ng of GK1 reporter plasmid, 125 ng of a pM vector
encoding Gal4DBD (left-hand lane) or Gal4DBD-GRIP1 fusion protein (all other lanes),
and varying concentrations (100, 200, and 400 ng) of vectors encoding VP16 fused to
CARM1 or its deletion mutants as indicated. The luciferase activities shown are from a
single experiment which is representative of three independent experiments.
FIG. 3. Histone H3 methylation by wild type and mutant CARM1. COS7 cells were
transfected with vectors encoding HA.CARM1 or its deletion mutants and the
methyltransferase proteins were immunoprecipitated from cell extracts with anti-HA
antibodies. Histone H3 was incubated for 1 h at 30 °C with the immunoprecipitated
CARM1 and [3H-methyl]AdoMet. Methylated histone H3 was detected by SDS-PAGE
and fluorography. The results shown are from a single experiment representative of three
independent experiments.
Teyssier et al 9-10-02
23
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

FIG. 4. Location of homo-oligomerization domain in conserved central region of
CARM1. A, [35S]Methionine-labeled CARM1 or its deletion mutants were incubated
with either GST or GST-CARM1 fusion protein preloaded on glutathione-coupled
beads. Bound proteins were eluted and analyzed as in Fig. 2A. The input lanes represent
10% of each 35S-labeled protein in the binding assay. B, CV1 cells were transiently
transfected with 250 ng of GK1 reporter plasmid, 125 ng of pM vector encoding
Gal4DBD or Gal4DBD fused to wild type or mutant CARM1, and 125 ng of pVP16
vector encoding VP16-CARM1 as indicated. Luciferase activity of the transfected cell
extracts was determined. The results presented are from a single experiment
representative of two independent experiments.
FIG. 5. The CARM1 C-terminal region contains an autonomous activation domain. CV1
cells were transiently transfected with 250 ng of GK1 reporter plasmid, 400 ng of pM
vector encoding Gal4DBD or increasing amounts (100, 200, and 400 ng) of vectors
encoding Gal4DBD fused to CARM1 or its deletion mutants. Luciferase activity of the
transfected cell extracts was determined. Note the different scale on the y-axis of the
right-hand panel. The results shown are from a single experiment representative of three
independent experiments.
FIG. 6. Dominant negative effect of CARM1-C on the coactivator function of CARM1.
A, CV1 cells were transiently transfected with 250 ng of MMTV(ERE)-LUC reporter
plasmid (upper panel) or RSV.β-gal reporter plasmid (lower panel); expression vectors
Teyssier et al 9-10-02
24
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

encoding ER (5 ng) and GRIP1 (125 ng); and 400 ng of a pSG5.HA vector encoding full
length CARM1 or CARM1-C (residues 461-608) or both. Transfected cells were grown
in culture medium with 20 nM estradiol (E2) or with vehicle (C), and extracts of the
harvested cells were tested for luciferase or β-galactosidase activity. B, CV1 cells were
transiently transfected with 250 ng of GK1 reporter plasmid, 250 ng of pM vector
encoding Gal4DBD or Gal4DBD fused to CARM1, 250 ng of pVP16 vector encoding
VP16-CARM1 (right panel only), and 400 ng of pSG5.HA empty vector (C) or
pSG5.HA vector encoding CARM1 or CARM1-C.
FIG. 7. CARM1 structural and functional domains. Regions of CARM1 which are
conserved or unique among the PRMT family are indicate by shading. Structural and
functional features determined from X-ray crystallography studies of PRMT3 and
Rmt1/Hmt1 (29,39) have been superimposed on the homologous regions of CARM1 in
the diagram; these features are labeled inside of the diagram or indicated below the
diagram by the boxes with vertical stripes. The minimum fragments retaining specific
functions as determined in the current study are indicated by the boxes with diagonal
stripes. All numbers refer to the amino acid residues of mouse CARM1.
Teyssier et al 9-10-02
25
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Catherine Teyssier, Dagang Chen and Michael R. Stallcupin its transcriptional coactivator function
Requirement for multiple domains of the protein arginine methyltransferase CARM1
published online September 25, 2002J. Biol. Chem.
10.1074/jbc.M207623200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from




![Arginine...Arginine vasotocin ([8-arginine]-oxytocin) (AVT), the primary antidiuretic principle in submammalian vertebrates, has been reported to be present in mammalian pituitary](https://img.pdfslide.us/doc/110x75/5e81a7e1761a1c6f5832a8ca/arginine-arginine-vasotocin-8-arginine-oxytocin-avt-the-primary-antidiuretic.jpg)