Embed Size (px)
Citation preview

Reproductive success and fry production of the paiche
or pirarucu, Arapaima gigas (Schinz), in the region of
Iquitos, Peru
Jesu¤ s Nu¤ n! ez1,2, Fred Chu-Koo3, Magali Berland1, LambertoAre¤ valo3, Ola¡ Ribeyro3,Fabrice Duponchelle1,2 & Jean Franc" ois Renno1,31IRD UR175 CAVIAR, Montpellier, France2UNFV-FOPCA, Mira£ores, Lima, Peru¤3Instituto de Investigaciones de la Amazon|¤ a Peruana ^ IIAP, Programa para el Uso y Conservacio¤ n del Agua y sus Recursos(AQUAREC), Iquitos, Peru¤
Correspondence: J Nu¤ n! ez,351RUE J.F. BRETON, BP 5095, Montpellier cedex 05, France. E-mail: [email protected]
Abstract
Arapaima gigas (paiche) is the largest scaled ¢shspecies living in the Amazon basin. Its biology isboth fascinating and misunderstood. In a context ofover¢shing, hence reduced natural populations,aquaculture of a ¢sh with such interesting charac-teristics (large size, high growth rate, no intramuscu-lar spines) is an important issue. The developmentof farming production would also reduce the ¢shingpressure on natural populations and allow re-stocking programmes in certain areas. To determinewhat factors may in£uence the reproductive successin captivity, data from breeding reports for 2007^2010 were collected among ¢sh farmers in the regionof Iquitos. In parallel, we carried out physico-chemical measurements in di¡erent ponds wherethese paiches breed, and conducted personal inter-views about the general ¢sh management condi-tions. The results show that reproduction occursthroughout the year but with a higher intensityduring the rainy season. It also highlights farmsthat have performed much better than others, butno single factor except feeding level has been clearlyassociated with reproductive success. The environ-mental control of reproduction in paiche, therefore,remains partly mysterious. To deepen this study, werecommend the systematic sexing of breeders, ex-tending reproductive behavioural studies, and exam-ining the limnological factors involved in frymortality.
Keywords: arapaima, pirarucu, paiche, reproduc-tion, peru, ¢sh culture
Introduction
Arapaima gigas (Schinz) is the largest scaled ¢shspecies living in the Amazon basin (Chu-Koo &Alca" ntara, 2009). It is an emblematic species of theAmazonian ¢sh fauna, also known as the Paiche inPeru and Pirarucu in Brazil. This species is very at-tractive for aquaculture in theAmazon region, owingto its many advantages. Arapaima gigas has the bestgrowth rate among Amazonian cultivated ¢sh spe-cies, of 10^15 kg per year (Bard & Imbiriba, 1986;Pereira Filho & Roubach,2005; Nun! ez, 2009; Rebaza,Rebaza & Deza 2010), and its ¢let, which is verypopular, does not have intramuscular spines. It alsoaccepts low water dissolved oxygen levels, due to itsobligatory aerial breathing. Finally, it has a carnivor-ous diet but is still £exible because it can accommo-date consumption of dead ¢sh or ¢sh feed pellets.However, natural populations have been subjected toexcessive ¢shing in the past decades for human con-sumption and ornamental ¢sh trading (Chu-Koo &Tello, 2010), and already for more than 15 years,A. gigas has become a very uncommon species in theAmazon, except in some natural reserves (PacayaSamiria in Peru or Mamiraua" in Brazil). The Paicheis now in the CITES II list, which means that its trade
Aquaculture Research, 2011, 42, 815^822 doi:10.1111/j.1365-2109.2011.02886.x
r 2011 Blackwell Publishing Ltd 815

and use are strictly controlled. Scarce available infor-mation concerns mainly the reproductive biologyin the wild (Guerra, 1980; Saavedra Rojas, QuinteroPinto & Landines Parra 2005); however, it is in theinterest of ¢sh farmers to develop a controlled pro-duction of this species (Pereira Filho & Roubach,2005; Nun! ez, 2009) and one of the collateral ex-pected e¡ects of Paiche domestication is a decreased¢shing pressure on wild populations. Nevertheless,many attempts have been made in recent decades todevelop its production in captivity (Bard & Imbiriba,1986; Imbiriba,1991; Alcantara & Guerra,1992; Guer-ra, Alca" ntara, Padilla, Rebaza,Tello, Ismin! o, Rebaza,Deza, Ascon, Iberico, Montreuil & Limachi 2002;Pereira Filho & Roubach,2005; Saavedra Rojas, Quin-tero Pinto, Lopez Hernandez & Pezzato 2005), but thistask is made di⁄cult by the dispersed knowledge onA. gigas biology in captivity, especially in crucial as-pects like reproduction and feeding.Paiche domestication faces the di⁄culty of gender
determination, which is virtually impossible fromexternal characteristics except during the reproduc-tive period; otherwise, sex has to be determined inblood or plasma samples by biochemical methodsusing vitellogenin or sex steroid ratios (Chu-koo,Dugue¤ , Alva" n Aguilar, Casanova Daza, Alca" ntaraBocanegra, Cha" vezVeintemilla, Duponchelle, Renno,Tello & Nun! ez 2009). Moreover, the ovary is com-posed only by a single naked lobe or gymnovarium(Godinho, Santos, Formagio & Guimaraes-cruz2005; Saavedra Rojas, Quintero Pinto & LandinesParra 2005; Nu¤ n! ez & Duponchelle, 2009) that makesthe practice of canulation to determine oocyte devel-opment, inductionof oocytematurationandarti¢cialfertilization almost impossible. As the production of¢ngerlings actually relies only on spontaneous re-productions in ponds, the knowledge of the optimalrequirements for reproductive success is a prerequi-site to substantially increase the ¢ngerling o¡er inthis region. Nevertheless, a noticeable developmentof A. gigas production is expected as most of theactual ¢ngerling production is destined for aquacul-ture instead of ornamental ¢sh trade as before(Chu-Koo & Tello, 2010). This study aims at analysingthe reproductive data collected from A. gigas farmersin the region of Iquitos between 2007 and 2010.The results reported here represent the ¢rst attemptto provide answers on the magnitude of a sustainedproduction level of ¢ngerlings with the currentknowledge and management conditions. It alsoevaluates the variations of some physicochemicaland environmental factors that could promote or
inhibit A. gigas reproduction in controlled environ-ments.
Material and methods
Fish origin
Between 2000 and 2007, a programme to assist ¢shfarmers in the region of Iquitos enabled the Institutode Investigaciones de la Amazon|¤ a Peruna (IIAP) toprovide six paiche breeders to selected local ¢shfarmers. Arapaima breeders had been bred previouslyin ponds for 3 years in the IIAP’s facilities of Quisto-cocha.Fish reproduction supervising, mandated by law,
started in 2007 and ¢sh farmers had to report to localauthorities all events (births, fryharvest, etc.) that oc-curred in their farms, thus facilitating data collec-tion. Some farms already owned paiches before theassistance programme; they did not originate fromIIAP’s stock but contributed to some reproductiveevents. Arapaima breeder weights varied approxi-mately from 60 to100 kg.
Fish rearing and feeding
The paiches are fed live ¢sh (forage ¢sh), dead ¢sh(trash ¢sh from local market) and sometimes down-graded chicken or chicks, ¢sh or chicken viscera, ¢shfeed pellets and even fruits, although ¢sh-based foodis predominant in every farm.The amount of food percapita (calculated as kg food paiche biomass!1) var-ied greatly among farms, and most farmers could notprecisely quantify how much food they had distribu-ted to ¢sh daily, monthly or even yearly. For this rea-son, wehad to split the farms in three feeding levels:1,small quantity (the forage ¢sh are distributed aboutonce a month); 2, average quantity (the ¢sh are fedabout once aweek) and 3, large quantity (the ¢sh arefed daily). In the ¢rst level, the food is essentially for-age ¢sh consumed progressively by paiches. In theother two levels, the food is mainlycomposed of trash¢sh supplemented, or not, with the other cited food-stu¡s.
Physicochemical parameters
The physicochemical characteristics of pond waterswere measured within a weekly period for all farmsduring the rainy season (February^March), usingthe LamotteAQ-2 kit (Drop Count Titration, Chester-
Reproductive aspects of ARAPAIMA in captivity J Nu¤ n! ez et al. Aquaculture Research, 2011, 42, 815^822
816 r 2011 Blackwell Publishing Ltd, Aquaculture Research, 42, 815^822

town, MD, USA). The following parameters wereconsidered:carbon dioxide (CO2): absent if values wereo1.25mgmL!1 corresponding to the detectionlimit, present if values were41.25mgmL!1;nitrites (NO2! ): absent if values wereo0.05mgmL!1 corresponding to the detectionlimit; present if values were40.05mgmL!1;ammonia (NH41): absent if values wereo0.02mgmL!1 corresponding to the detectionlimit; present if values were40.02mgmL;pH and temperature were measured using a YSI
s
digital meter (Yellow Springs Instrument Com-pany, Yellow Springs, OH, USA) at 15 cm from thesurface;hardness (mg L!1of CaCO3) levelso60mgmL!1
are considered to be low;transparency was determined by the Secchi discmethod.
Hydromorphological parameters
The environmental parameters susceptible to play arole in the reproductive success of the paiche havebeen recorded in all farms. Six environmental para-meters have been studied: pond area; pond maxi-mum depth; presence of vegetation on pond watersurface; shape of the shoreline (slow or steep slope);soil type (clay, sandy clay or sandy-muddy) and water(£ow through or stagnant water).
Field data collection and analysis
We went to di¡erent ¢sh farms, and there we re-quested to speak to the people who tend the paiches.Then, we explained the reason for our presence, andrequested permission to make measurements of theirpond water quality. During the interviews, we en-quired about the total number of paiches and thefood types and quantities distributed. During the vis-it, we also evaluated the total pond area, soil type andwater surface vegetation.Data were analysed with the free statistical soft-
ware R, version 2.6.2 and Statgraphics Plus (StatPointTechnologies,Washington, DC, USA).
Results
Pond characteristics
The visited farms are located along the road betweenIquitos and Nauta (Fig. 1). Farms are generally
composed of a few ponds of di¡erent sizes andshapes, including small water bodies created in thelower parts of small water streams that drain theabundant rainfall in the area. Most of the farms haverunning water almost all year round. A fewhave lim-ited water £ow or even depend exclusively on rain-falls. The maximum depth is generally comprisedbetween 1.5 and 4m (Table 1) with variable £oatingvegetation cover, and with shorelines generally cov-ered by grass or other small plants, which were ex-ceptionally steep.The physicochemical parameters measured varied
withina small range (Table1) andwere characteristicof good qualityand soft water. In two cases, pH valuesattained relatively high levels (6.5 an 8.0), whereasthe majority of other farms had pH values around5.0 and hardness never exceeded 25mg L!1. Thesevalues correspond to acidic soft waters typical ofthese environments. Water transparency (Table 1)was generally low (mean of 33 cm). This low trans-parency is also typical of local waters and sometimesafter rainfall the sediment content contributes to dra-matically decrease water transparency. Carbon diox-ide, NO2! and NH41 levels were, in most cases,below the detection limits of the measurement kitand never reached critical values for ¢sh, indicatingthat rearing conditions were compatible with theknown species preferenda.
Reproductive activity
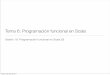
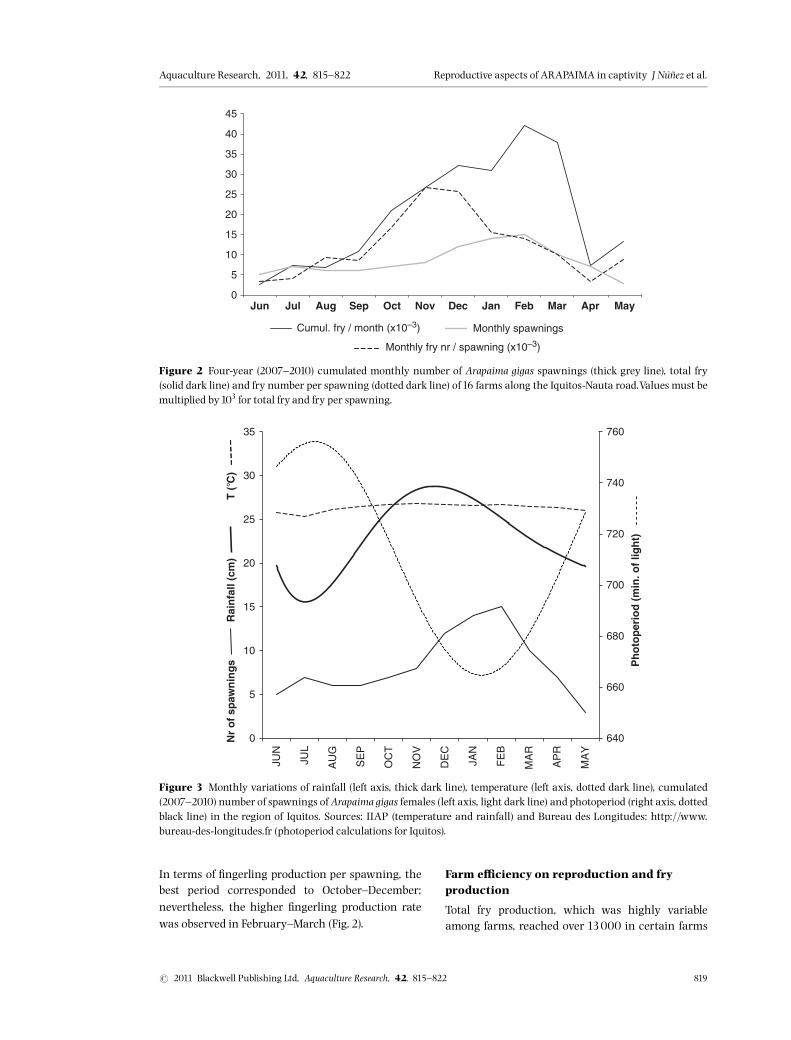
The reproductive activity of A. gigas was determinedduring the period 2007^2010 on the monthly num-ber of spawning events reported by the farmers.Spawning generally occurred in a shallow area ofthe pond in a small depression or nest dug by thetwo brooders. Reproductions occurred all yearround, although a marked seasonality was apparent(Fig. 2). The maximum number of spawning eventswas observed between December and April (73 outof 100 spawnings over the 4-year period), corre-spondingmainly to the rainy season, short photoper-iod and higher temperatures (Fig. 3). During the dryseason (May^September), reproductive activity (Fig.2) was lower (27 out of100 spawnings).
Seasonality of fry production
The seasonality of fry production (Fig. 2) was morepronounced than that of spawning activity. Themonthly fry production peaked during October^
Aquaculture Research, 2011, 42, 815^822 Reproductive aspects of ARAPAIMA in captivity J Nu¤ n! ez et al.
r 2011 Blackwell Publishing Ltd, Aquaculture Research, 42, 815^822 817

March, and a very limited fry production wasobserved during the dry season from April toSeptember, corresponding to longer days and lowertemperatures (Fig. 3). Patterns of fry production andnumber of fry per spawning were quite similar from
April to November, but di¡ered strikingly from De-cember to March (Fig. 2). During this second half ofthe maximum reproductive activity period, themonthly number of ¢ngerlings per spawning de-creased remarkably to reach its minimum in April.
Figure 1 Aerial viewof the ‘Carretera Iquitos-Nauta’ Loreto, Peru. Points indicate the location of the main farms.
Table 1 Summary of some physicochemical and physical characteristics of Arapaima gigas breeding ponds in the region ofIquitos, Peru (Iquitos-Nauta road)
Carbondioxide Nitrites Ammonia pH
Hardness(mgmL! 1)
Transparency(cm)
Temperature( 1C)
Area(m2)
Depth(m)
Minimum 2 0.05 0.2 4.25 7.0 7 27.5 500 1.5
Maximum 22 0.15 3 7.5 25.0 124 32.4 8 000 4
Mean
(" SD)
8.85 0.089 1.06 5.47 15.4 33 29.9 2 923 2.12
6.24 0.121 0.82 0.86 4.32 30 1.46 2 142 0.82
Reproductive aspects of ARAPAIMA in captivity J Nu¤ n! ez et al. Aquaculture Research, 2011, 42, 815^822
818 r 2011 Blackwell Publishing Ltd, Aquaculture Research, 42, 815^822
In terms of ¢ngerling production per spawning, thebest period corresponded to October^December;nevertheless, the higher ¢ngerling production ratewas observed in February^March (Fig. 2).
Farm e⁄ciency on reproduction and fryproduction
Total fry production, which was highly variableamong farms, reached over 13000 in certain farms
640
660
680
700
720
740
760
0
5
10
15
20
25
30
35
JUN
JUL
AU
G
SE
P
OC
T
NO
V
DE
C
JAN
FE
B
MA
R
AP
R
MA
Y
Pho
tope
riod
(min
. of l
ight
)
Nr
of s
paw
ning
sR
ainf
all (
cm)
T (°
C)
Figure 3 Monthly variations of rainfall (left axis, thick dark line), temperature (left axis, dotted dark line), cumulated(2007^2010) number of spawnings of Arapaima gigas females (left axis, light dark line) and photoperiod (right axis, dottedblack line) in the region of Iquitos. Sources: IIAP (temperature and rainfall) and Bureau des Longitudes: http://www.bureau-des-longitudes.fr (photoperiod calculations for Iquitos).
0
5
10
15
20
25
30
35
40
45
Jun Jul Aug Sep Oct Nov Dec Jan Feb Mar Apr May
Cumul. fry / month (x10–3)
Monthly fry nr / spawning (x10–3)
Monthly spawnings
Figure 2 Four-year (2007^2010) cumulated monthly number of Arapaima gigas spawnings (thick grey line), total fry(solid dark line) and fry number per spawning (dotted dark line) of16 farms along the Iquitos-Nauta road.Values must bemultiplied by103 for total fry and fry per spawning.
Aquaculture Research, 2011, 42, 815^822 Reproductive aspects of ARAPAIMA in captivity J Nu¤ n! ez et al.
r 2011 Blackwell Publishing Ltd, Aquaculture Research, 42, 815^822 819
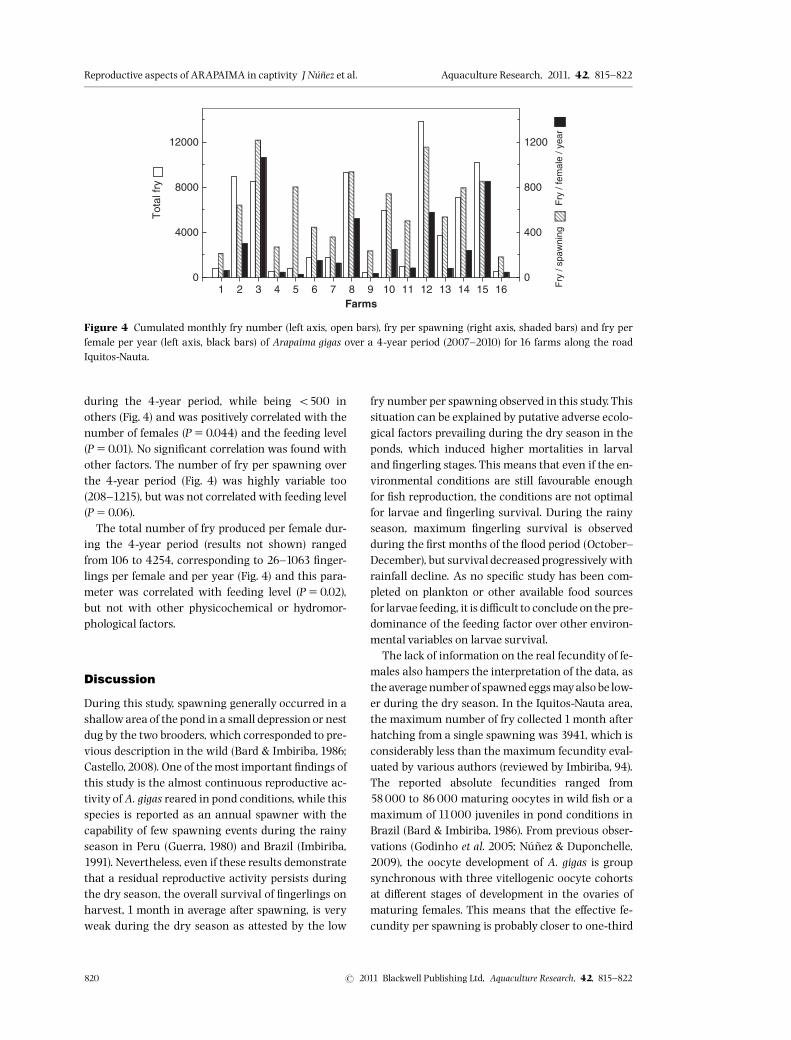
during the 4-year period, while being o500 inothers (Fig. 4) and was positively correlated with thenumber of females (P50.044) and the feeding level(P50.01). No signi¢cant correlation was found withother factors. The number of fry per spawning overthe 4-year period (Fig. 4) was highly variable too(208^1215), but was not correlated with feeding level(P50.06).The total number of fry produced per female dur-
ing the 4-year period (results not shown) rangedfrom 106 to 4254, corresponding to 26^1063 ¢nger-lings per female and per year (Fig. 4) and this para-meter was correlated with feeding level (P50.02),but not with other physicochemical or hydromor-phological factors.
Discussion
During this study, spawning generally occurred in ashallowarea of the pond in a small depression or nestdug by the two brooders, which corresponded to pre-vious description in the wild (Bard & Imbiriba, 1986;Castello,2008). One of the most important ¢ndings ofthis study is the almost continuous reproductive ac-tivity of A. gigas reared in pond conditions, while thisspecies is reported as an annual spawner with thecapability of few spawning events during the rainyseason in Peru (Guerra, 1980) and Brazil (Imbiriba,1991). Nevertheless, even if these results demonstratethat a residual reproductive activity persists duringthe dry season, the overall survival of ¢ngerlings onharvest, 1 month in average after spawning, is veryweak during the dry season as attested by the low
fry number per spawning observed in this study.Thissituation can be explained by putative adverse ecolo-gical factors prevailing during the dry season in theponds, which induced higher mortalities in larvaland ¢ngerling stages. This means that even if the en-vironmental conditions are still favourable enoughfor ¢sh reproduction, the conditions are not optimalfor larvae and ¢ngerling survival. During the rainyseason, maximum ¢ngerling survival is observedduring the ¢rst months of the £ood period (October^December), but survival decreased progressively withrainfall decline. As no speci¢c study has been com-pleted on plankton or other available food sourcesfor larvae feeding, it is di⁄cult to conclude on the pre-dominance of the feeding factor over other environ-mental variables on larvae survival.The lack of information on the real fecundity of fe-
males also hampers the interpretation of the data, asthe average number of spawned eggsmayalso be low-er during the dry season. In the Iquitos-Nauta area,the maximum number of fry collected 1month afterhatching from a single spawning was 3941, which isconsiderably less than the maximum fecundity eval-uated by various authors (reviewed by Imbiriba, 94).The reported absolute fecundities ranged from58000 to 86000 maturing oocytes in wild ¢sh or amaximum of 11000 juveniles in pond conditions inBrazil (Bard & Imbiriba, 1986). From previous obser-vations (Godinho et al. 2005; Nu¤ n! ez & Duponchelle,2009), the oocyte development of A. gigas is groupsynchronous with three vitellogenic oocyte cohortsat di¡erent stages of development in the ovaries ofmaturing females. This means that the e¡ective fe-cundity per spawning is probably closer to one-third
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 160
4000
8000
12000
0
400
800
1200
Farms
Fry
/ sp
awni
ngF
ry /
fem
ale
/ yea
r
Tot
al fr
y
Figure 4 Cumulated monthly fry number (left axis, open bars), fry per spawning (right axis, shaded bars) and fry perfemale per year (left axis, black bars) of Arapaima gigas over a 4-year period (2007^2010) for 16 farms along the roadIquitos-Nauta.
Reproductive aspects of ARAPAIMA in captivity J Nu¤ n! ez et al. Aquaculture Research, 2011, 42, 815^822
820 r 2011 Blackwell Publishing Ltd, Aquaculture Research, 42, 815^822

of the reported maximum fecundity or could beequivalent to the oocyte number of the larger oocytediameter cohort. Even if one takes into account thishypothesis (i.e. around 20000 spawned eggs), the to-tal number of hatched larvae is very far below this¢gure in view of our observations during this 4-yearstudy. The probability that such high rates of eggsand young stages mortality exist in a species provid-ing elaborated bi-parental care seems unlikely. Thisspecies may experience high rates of atresia just be-fore spawningor poor egg survival during the embry-ogenesis stage, occurring in the nest at the bottom ofthe pond, which might partly account for why esti-mated fecundities on ovaries in ¢nal maturationstage are much higher than the observed number ofguarded fry. Clearly, additional work is needed toshed light on this issue and to better understand thereproductive potential of theA. gigas in captivity.The numberof fry produced per female and per year
has been positively correlated with the feeding level;the farms where the food distributionwas very low ex-hibited also a lower number of spawning. Among allother factors studied, none appeared to play a key roleon the number of spawning or the total number of fryproduced. In particular, the presence or absence of£oating vegetation seemed to have no in£uence in thereproductive activity of A. gigas in captivity. Concern-ing NH41, the absence of correlationwith the numberof spawning indicated that NH41 had no direct e¡ectwithin themeasured range on female reproductive ac-tivity, and did not a¡ect ¢sh behaviour becauseA. gigascan tolerate much higher concentrations than thoseobserved in this study (Cavero, Pereira-¢lho, Bordin-hon, Fonseca, Ituassu¤ , Roubach & Ono 2004).Regardless of the number of spawning events, the
quantity of fry produced per brood varied greatlyfrom one farm to another. These di¡erences can beexplained by lower fecundities or di¡erential mortal-ities during the1-month average period before ¢nger-ling harvest. The most likely hypothesis to explainthese variable mortalities is that feeding of fry de-pends greatly on the productivity of ponds. As juve-niles and ¢ngerlings feeding in the wild consistsmainly of zooplankton, insects, insect larvae andsmall gastropods (Oliveira, Poleto & Venere 2005),the main causes of mortality are probably an insu⁄-cient food availability. This hypothesis needs to betested by a detailed study of the limnological para-meters and plankton productivity in the di¡erentfarming environments. If con¢rmed, considering thehuge variations observed on harvested fry amongfarms, ¢ngerling production could be improved by
properly fertilizing the ponds to allow optimal pri-mary and secondary production.The results described in this study emphasize the
high impact of feeding level on reproductive activityand probablyalso on the survival of young stages andit appeared that in most farms the feeding level of thebreeders was insu⁄cient. The importance of nutri-tional aspects has been also reported by (SaavedraRojas, Quintero Pinto, Lopez Hernandez et al. 2005)for the quantity and quality of fry produced in captiv-ity, but this is the ¢rst experimental observation of di-rect in£uence of the feeding level on reproductiveactivity of A. gigas. In the Iquitos-Nauta area, feedingof A. gigas is achieved mainly with frozen or live ¢sh,this feed appeared to be e¡ective for breeding stocksat 2% of biomass per day, although a more detailedstudy is needed to establish the exact amount andquality of food that would allow for optimal repro-ductive rate and fry survival in this species.In conclusion, this study shows that under good
management conditions including su⁄cient foodsupply, the production of A. gigas ¢ngerlings in cap-tivity can be substantial (more than 1000 fry per fe-male per year) and holds a good potential forimprovement, despite the low fecundity of this spe-cies. These results also substantiate the possibility ofsustained fry production almost during 8 monthsyear round from September to April. Finally, ourstudy con¢rms the possibility of aquaculture devel-opment of A. gigas through an improvement ofbroodstock management.
Acknowledgements
This study was carried out as part of the scienti¢ccollaborations between the IIAP and the Institut deRecherche pour le De¤ veloppement (IRD), both partof the research network known as RIIA (Red de In-vestigacio¤ n sobre la Ictiofauna Amazo¤ nica, http://www.riiaamazonia.org).The authors wish to thankall the farmowners and
people who contributed to the ¢eld work.This work has been funded in part by the Peruvian
research programme INCAGRO.This is a publication IRD-DIVA-ISEM 2011-042.
Reference
Alcantara F. & Guerra H. (1992) Cultivo de paiche, Arapaimagigas, utilizando bujurqui, Cichlasoma bimaculatum, comopresa. Folia Amazo¤ nica 4,129^139.
Aquaculture Research, 2011, 42, 815^822 Reproductive aspects of ARAPAIMA in captivity J Nu¤ n! ez et al.
r 2011 Blackwell Publishing Ltd, Aquaculture Research, 42, 815^822 821

Bard J. & Imbiriba E.P. (1986) Piscicultura de pirarucu, Arapai-ma gigas. No.52. EMBRAPA-CPATU, Bele¤ m, Brazil pp.52.
Castello L. (2008) Nesting habitat of Arapaima gigas (Schinz)in Amazonian £oodplains. Journal of Fish Biology 72,1520^1528.
Cavero B.A.S., Pereira-Filho M., Bordinhon A.M., FonsecaF.A.L., Ituassu¤ D.R., Roubach R. & Ono E.A. (2004) Toler-a“ ncia de juvenis de pirarucu aoaumento da concentrac# a! ode amoŒ nia em ambiente con¢nado. Pesquisa Agropecua?r-ia Brasileira 39, 513–516.
Chu-Koo F. & Alca" ntara F. (2009) Paiche dome¤ stico en laAmazon|¤ a: perspectivas de una crianza sostenible. PescaResponsable 57,32^33.
Chu-Koo F. & Tello S. (2010) Produccio¤ n de semilla de Paicheen Peru¤ . Infopesca Internacional 41,30^35.
Chu-Koo F., Dugue¤ R., Alva" n Aguilar M., Casanova Daza A.,Alca" ntara Bocanegra F., Cha" vezVeintemilla C., Duponch-elle F., Renno J.F., Tello S. & Nun! ez J. (2009) Genderdetermination in the Paiche or Pirarucu (Arapaima gigas)using plasma vitellogenin, 17b-estradiol, and 11-ketotes-tosterone levels. Fish Physiology and Biochemistry 35,125^136.
Godinho H.P., Santos J.E., Formagio P.S. & Guimaraes-CruzR.J. (2005) Gonadal morphology and reproductive traitsof theAmazonian ¢sh Arapaima gigas (Schinz,1822). ActaZoologica 86, 289^294.
Guerra F.H. (1980) Desarollo sexual del paiche, Arapaimagigas, en las zonas reservadas del estado (R|¤ o Pacaya Sa-miria) 1971^1975. In : Informe IMARPE No. 67, pp. 20.Instituto del Mar del Peru, Callao, Peru.
Guerra H., Alca" ntara F., Padilla P., Rebaza M.,Tello S., Ismi-n! o R., Rebaza C., Deza S., Ascon G., Iberico J., MontreuilV.& Limachi L. (2002) Produccio¤ n y manejo de alevinos depaiche. : IIAP, Iquitos, Peru¤ .
Imbiriba E.P. (1991) Produc" a! ao emanjo de alevinos de pirarucu,Arapaima gigas (Cuvier).Technical Report EMBRAPA-CPA-TU,57,19pp.
Nun! ez J. (2009) Domestication de nouvelles espe' ces d’inte¤ reŒ taquacole en Amazonie. Cahiers Agriculture18,136^143.
Nu¤ n! ez J. & Duponchelle F. (2009) Towards a universal scaleto assess sexual maturation and related life history traitsin oviparous teleost ¢shes. Fish Physiology and Biochemis-try 35,167^180.
OliveiraV.D., Poleto S.L. & Venere P.C. (2005) Feeding of juve-nile pirarucu (Arapaima gigas, Arapaimidae) in their nat-ural environment, lago Quatro Bocas, Araguaiana-MT,Brazil. Neotropical Ichthyology 3,312^314.
Pereira Filho M. & Roubach R. (2005) Pirarucu, Arapaimagigas. In Espe¤ cies nativas para piscicultura no Brasil (ed.by H. Rosa Nascimento), pp. 37–62. UFSM, Santa
Maria, Brasil.
Rebaza M., Rebaza C. & Deza S. (2010) Densidad de siembrapara cultivos de Paiche en jaulas £otantes. Aquavisio¤ n 6,26^27.
SaavedraRojas E.A., QuinteroPintoL.G. &LandinesParraM.A.(2005) Aspectos reproductivos. In: Biolog|¤ a y cultivo del pir-arucu¤ Arapaima gigas Schinz, 1822. Pisces: Arapaimidae.Bases para un aprovechamiento sostenible ^ Aspectos Repro-ductivos (ed. by A.I. Sanabria, I.C. Beltran & P.V.
Daza), pp. 31–40. INCODER/UNC, Bogota, Colombia.
Saavedra Rojas E.A., Quintero Pinto L.G., Lopez HernandezN. & Pezzato L.E. (2005) Nutricio¤ n y alimentacio¤ n del pir-arucu¤ Arapaima gigas (Schinz,1882). In Biolog|¤ a y cultivodel pirarucu¤ Arapaima gigas Schinz,1822. Pisces: Arapaimi-dae. Bases para un aprovechamiento sostenible ^ AspectosReproductivos (ed. by A.I. Sanabria, I.C. Beltran &
P.V. Daza), pp. 41–58. INCODER/UNC, Bogota,
Colombia.
Reproductive aspects of ARAPAIMA in captivity J Nu¤ n! ez et al. Aquaculture Research, 2011, 42, 815^822
822 r 2011 Blackwell Publishing Ltd, Aquaculture Research, 42, 815^822