Embed Size (px)
Citation preview

46
Eapen Jacob, 2013. Studies on the captive breeding and
reproductive biology of two indigenous ornamental
fishes of the Western Ghats. Ph. D Thesis, Mahatma
Gandhi University, Kottayam, India
CHAPTER 3
REPRODUCTIVE BIOLOGY OF
PUNTIUS POOKODENSIS
3.1 INTRODUCTION
Reproductive biology of many commercially important fishes has been studied,
because of their importance in fish culture and fishery management programmes.
However a review of literature showed that such studies have been selective and the
numerous smaller fishes have been neglected extensively. Smaller fishes are important
in fishery management programmes, because they are prey for larger fish or compete
with them for food (Geevarghese and John, 1983) and they are the unavoidable parts of
many aquatic biotic communities. As far as P. pookodensis is concerned, this is the
pioneering work and other biological aspects are still in dark. Considering its
importance, attempt has been made to study the different aspects of reproductive
biology of this specie.

47
Development of a successful captive propagation technique for freshwater fishes
depends on the raising of a domesticated broodstock (Scott, 1962). In order to begin the
domestication process it is necessary to gain control over all phases of its life cycle,
especially reproduction. The development of a potential broodstock turns to be the
major cost for the producer due to the expense of operating and maintaining facilities as
well as feeding fish for extended periods of growth, sexual maturation and successive
reproductive cycles. Although males are equally important in reproduction, the
availability of eggs in right maturity stage, most often, becomes the limiting factor. In
many species, egg production by females is a year long process but viable milt can be
produced by males in a matter of a few weeks (Bromage et al.,1992).
Knowledge on the reproductive biology of any fish is very important for
productive aquaculture and scientific based fishery management of any of the water
bodies. Manipulation of a fish’s reproductive system under culture or captive conditions
requires an understanding of natural spawning patterns and other influential factors.
Reproductive biology i.e. fecundity, spawning, sex ratio etc. are among the important
aspects of the biology of fishes, which must be understood to explain the variations in
the level of populations as well as to make efforts to increase the amount of harvest
(Azadi and Mamun, 2004). Puntius pookodensis is considered as an important
indigenous ornamental fish of the Western Ghats of India and breeding this fish under
captive conditions will help to produce it commercially. A thorough knowledge on the
reproductive biology of a particular species is very important for the proper management
as well as understanding and predicting the annual changes that occur in the population.
The different reproductive parameters like Gonado Somatic Index (GSI), absolute

48
fecundity, relative fecundity, oocyte diameter frequency analysis, sexual dimorphism
and sex ratio, and size (length / weight) at the time of initial maturity of Puntius
pookodensis have been investigated.
3.2 REVIEW OF LITERATURE
3.2.1 Fecundity
The absolute fecundity is described as the number of, usually mature eggs, filled
with yolk or all vitellogenic oocytes found in the ovary immediately before the
reproduction process (Bagenal, 1973). Fecundity among egg-laying animals is the
number of eggs being readied for the next spawning by a female (Royce, 1972).
Knowledge of fecundity is important for understanding the life history and for modeling
population dynamics of a species (Bruch et al., 2006) and is important in the estimation
of abundance and reproductive potential (Gupta, 1967). Fecundity appears to bear some
broad relationships to the care or nurture accorded to the eggs (Lagler et al., 1967).
Fecundity studies have been considered useful in tracing the different stocks or
populations of the same species of fish in different areas (Gupta, 1967). Knowledge
about fecundity of a fish is essential for evaluating the commercial potentialities of its
stock, life history, practical culture and actual management of the fishery (Lagler, 1956;
Doha and Hye, 1970). Fecundity varies from one species to another, depending on the
environmental conditions, length, age etc. Many researchers have worked on the
fecundity of different fish, such as, Bhuiyan et al. (1993), Kuddus et al. (1996), Alam et
al. (1994), Bhuiyan and Parveen (1998), Kiran and Puttaiah (2003) and Bhuiyan et al.
(2000) and Dobriyal et al. (2000).

49
Though it is easy procedure for estimation of the reproductive potential of any
given fish species, it does not give an accurate picture of the egg release as the fishes
tend to be multiple spawners or even batch spawners. Hence a precise estimation of
fecundity is almost impossible. Absolute fecundity is defined as the number of ripe
eggs found in the female prior to spawning (Bagenal and Braun, 1968). On the other
hand fertility may be defined as the actual number of young ones produced rather than
the number of eggs (Bagenal and Braun, 1968). Both endogenous and exogenous
factors have a profound effect on the fecundity. The variation in fecundity is dependent
on season, climatic condition, environmental habitat, nutritional status and genetic
potential (Bromage et al., 1992).
Estimation of the absolute fecundity of multiple spawners is difficult.
Usually all the yolked ripe or ripening eggs are counted based on the classic work of
Hickling and Rutenberg (1936) for multiple spawners. The estimation of fecundity in
open substrate spawners is also difficult owing to the extremely high fecundities and
protracted spawning periods. On the other hand fecundity estimation of the brood hiders
and nest spawners and all the fishes showing parental care is easy due to low fecundity.
Here, eggs are larger with high survival rate. In case of mouth brooders the fecundity
depends more on the brooding capacity of the parent than on the fertility. The fecundity
is the least in live bearers where the newly born are stronger than those that hatch out of
an egg. The fecundity is inversely related to egg size, yolk content and incubation time.

50
3.2.2 Ova diameter frequency studies
The frequency of reproduction forms the index of the predictability of the
environment and can be elucidated by the ova diameter studies. It has been
demonstrated that by studying the intraovarian egg dimensions of fishes in the ripe
condition or penultimate stage of maturity, it is possible to elucidate the duration of
spawning periods and individual spawning frequency (Clark, 1934; Hickling and
Rutenberg, 1936; De Jong, 1939; June, 1953; Prabhu, 1956; Quasim and Qayyum, 1961;
Grimes and Huntsman, 1980).
Based on oocyte size distribution, Wallace ad Selman (1981) and Dietrich and
Krieger (2008) classified ovaries into three basic types:
(i) ‘Synchronous ovaries’ in which all oocytes develop and ovulate in unison and
there is no replenishment from the earlier stages. Such ovaries are found in species that
spawn once and then die. The oocyte size distribution consists of a single mode
(semelparous fishes.)
(ii) ‘Group synchronous ovaries’, in which at least two size groups of oocytes are
present at the same time; the larger group or clutch usually being more homogenous
than the smaller.
(iii) ‘Asynchronous ovaries’, in which oocytes at all stages of development are
present at the same time. The oocyte size frequency distribution is continuous except in
the ripe stage where, there may be a clear separation of the yolked oocytes.
However, a protracted breeding season in itself does not imply multiple
spawning for each female, as it might simply reflect a lack of population synchrony in

51
gonad development showing asynchronous breeding populations (De Jong, 1939; De
Vlaming, 1983). Also the type of oocyte development is not species specific.
‘Isochronal’ or ‘total’ spawners is the other name given to the group
synchronous spawners while it is ‘partial’, ‘heterochronal’, ‘multiple’ or ‘serial’
spawners for the asynchronous spawners. According to Holden and Raitt (1974) the
oocytes will be shed within a short period - a week or so in the group synchronous
spawners and only a compliment of the yolked oocytes is spawned in case of the
asynchronous spawners. De Vlaming (1983) suggested that the multiple spawning
generally refers to more than one spawning in a season, and ‘fractional spawning’ is
used for species that spawn part of an ovulated clutch.
Based on spawning frequency (Prabhu, 1956; Karekar and Bal, 1960), the fishes
are categorized into four groups. This classification is based upon the works of Hickling
and Rutenbnerg (1936).
(i) Category A: Spawning takes place once in a season during a short duration, the
individual spawning once. Ovary contains a ripe stock distinctly and clearly separated
from immature stock.
(ii) Category B: Spawning takes place once in a season but with longer duration.
Range in size of the ripe ova nearly one half of the total range of intra-ovarian eggs.
(iii) Category C: Spawning more than once during a protracted spawning season.
Ovary with a batch of ripe stock, an immature stock and an intermediate ripening stock
in between the ripe and immature ones.

52
(iv) Category D: Spawning extended over a very long period or almost round the
year but intermittently, the individual spawns many times in the spawning season.
Batches of eggs in the ovary are not well differentiated from one another, usually shown
by fishes in tropical structured communities.
3.2.3 Gonadosomatic index
The Gonado Somatic Index (GSI), is one of the most important measures in the
estimation of the reproductive period and maturity condition of a fish. Some others
included maturation dynamics of the ovaries assessed by a maturity stage cycle,
condition factor and visual estimations. The gradually increasing values of GSI gives the
picture about the dynamics of gradual gonadal maturation and the maximum GSI means
the point of maximum reproductive maturity and beyond that a steep fall because of
spawning, the shedding of eggs. Sexually mature fish had high GSI values and in
females these were as much as six times greater than in males (Chellappa et al., 2003).
3.2.4 Size at first maturity or studies on initial sexual maturity
It is the length at which 50% of the fish population is regarded to have attained
gonadal maturity (Kagwade, 1968). This varies not only from species to species but
also within the species, which are subjected to varied internal and external stimuli.
These variations in the length at first maturity may be related to the ecological factors,
food supply and assimilation (Keshava et al., 1988). Usually in fishes, it is the males,
which mature earlier and so remain smaller than the females as greater part of their
energy reserves are diverted towards gonadal growth and development. However, in

53
species exhibiting territoriality and parental care it is the males, which are larger as they
are involved with the care of the eggs and young ones (Nickolsky, 1963). The females
usually mature later so that they can somatically grow larger and produce more number
of viable eggs.
Besides the above factors, population density and quantity and quality of food
available also have a significant role to play in determining the size at first maturity of
the given species. In expanding populations the females tend to mature at an earlier age.
Their maturing earlier owes to the fact that expanding population is usually found in
favourable environment and so attained larger size at a younger age. On the other hand
where the environment is favourable for growth and adult survival, fishes tend to delay
reproduction (Bagenal, 1957). The advantage in such cases is the increase in fecundity,
which is proportionate to the somatic growth of the fish. The size at first reproduction
has an important role in understanding the life history of a species during its evolution
and gives a rough estimation of the ultimate size of the species. A thorough knowledge
on initial sexual maturity has extreme importance in developing techniques for captive
propagation.
3.2.5 Sexual Dimorphism and Sex ratio
Differentiating sex is the primary step in developing a captive breeding
programme for a fish species. Sexual dimorphism depicts the morphological features
with which we can successfully segregate them sexwise. Differences in the selective
pressures experienced by the sexes can ultimately result in the evolution of sexual
dimorphism of morphological traits (Andersson, 1994). Fish exhibits wide variations in

54
the possession of sexual dimorphic characters. In some species, there exists highly
remarkable differences between male and female as they should have been mistaken to
be a different species, while some others are found to be extremely identical in
morphological features.
Determining the sex ratio of a population is tremendously valuable when we go
through the conservation aspects of a threatened species. The existence of a population
in its natural habitats is linked with its reproductive strategies. It includes the timing of
spawning, other breeding behaviour aspects and the number and proportion of male and
female individuals took part in the actual nuptial acts.
3.2.6 Reproductive Strategies
A complete knowledge of the reproductive system and the reproductive biology
of fishes are essential to understand the reproductive strategy of any given species. The
reproductive cycle of different species have developed in response to the fishes’ natural
range and habitat. In order to close the reproductive cycle for a given species it is
necessary to mimic some elements of the natural environment for providing the
appropriate stimuli for the reproductive cycle. The timing of spawning in annual
spawning fish has developed as a response to “ultimate” factors that will maximize the
survival of the eggs and fry. These ultimate factors include the water supply and water
quality, availability of suitable food supply and a reduced number of predators. In order
for the fish to be ready to spawn when the ultimate factors are present, the fish needs to
respond to "proximate" factors or cues to adjust the reproductive cycle to match the
changing environment (Sumpter, 1990). The main objective of a reproductive strategy is

55
to maximise reproductively active offspring in relation to available energy and parental
life expectancy (Wootton, 1984; Roff, 1992; Pianka, 2000). In order to achieve this, fish
follows different strategies and tactics (Balon, 1984). The reproductive strategy of a
species is the overall pattern of reproduction common to individuals of a species,
whereas reproductive tactics are those variations in response to fluctuations in the
environment (Wootton, 1984; Roff, 1996). Fishes exhibit great diversity in reproductive
strategies and associated traits such as breeding system, number of partners, gender role,
spawning habitat, spawning season, fecundity and others (Helfman et al., 1997).
Descriptions of reproductive strategies and the assessment of fecundity are
fundamental topics in the study of biology and population dynamics of fish species
(Hunter et al., 1992). Studies on reproduction, including the assessment of size at first
maturity, fecundity, duration of reproductive season, daily spawning behaviour and
spawning fraction, permit quantification of the reproductive capacity of individual fish.
This increases our knowledge about the state of stock and improves standard assessment
of many commercially valuable fish species thereby define their reproductive strategies.
Moreover, establishment of extensive databases on reproductive parameters with
corresponding data on abiotic factors enables the study of causal relationship between
reproductive potential and environmental variation. This leads to a better understanding
of observed fluctuations in reproductive output and enhances our ability to estimate
recruitment (Kraus et al.,2002).
There are three basic strategies for the production of eggs and their subsequent
spawning (Balon, 1984).

56
• Synchronous spawning : One crop of eggs and sperm are produced and
spawned at one time after which the fish dies.
• Group synchronous spawning : Groups of eggs are produced and
spawned at one time but several cycles of development and spawning can
occur.
• Asynchronous spawning: There is continuous development and spawning
of oocytes.
Different reproductive strategies are in existence and are explained based on different
components of breeding systems, such as number of breeding opportunities, type of
spawning, mating system, gender system, secondary sexual characteristics, spawning
site preparation, place of fertilization, embryonic development and parental care (Murua
and Saborido-Rey, 2003).
3.3 MATERIALS AND METHODS
3.3.1 Fecundity
Fecundity was estimated from samples collected from the wild as well as
hatchery raised specimens. A total number of 214 specimens (128 females and 86
males) were used for the study of different aspects of reproductive biology. The total
length and weight were recorded for each individual to the nearest centimeter (cm) and
gram (g) respectively. The specimens were then dissected out, the ovaries were removed
and the length and weight of the ovaries were recorded to the nearest millimeter (mm)
and milligrams (mg) respectively. The ripe ovaries were preserved in 5% formalin.
These were later weighed after removal of the excess water and also their length and

57
weight were taken. The oocytes were separated by introducing samples of completely
mature ovaries into Gilson’s solution, as modified by Simpson (1951). The ovaries,
being small, were kept whole in Gilson’s fluid with appropriate labels showing standard
length, body weight, ovary weight, etc. Then it was shook vigorously and left to stand
for about 24 hours. The Gilson’s fluid hardened the ova liberated easily as the ovarian
tissue breaks down. The ovary can also be preserved in Gilson’s fluid without any
disadvantage.
Composition of the Gilson’s fluid:
60% alcohol -100 ml; 80% alcohol - 15 ml; Saturated formalin - 20 ml; Water
- 880 ml; Glacial acetic acid - 18 ml.
For calculating the fecundity, gravimetric method (Burd and Howlett, 1974 ; Hunter et.
al., 1985) was adopted where a portion of the ovary accurately weighed and the
subsamples were counted and the same was multiplied by the total weight of the ovaries
using the formula,
Weight of the ovary X Average number of eggs per sub sample
Fecundity =
Average weight of the sub sample
The final figures of fecundity were arrived based on the three subsamples of each ovary.
The fecundity (F) thus computed was correlated to variables such as total length (TL) in
‘cm’ and body weight (W) in ‘g’. Regression analysis (Joshi and Khanna, 1980) was
employed to find out the correlation between fecundity and various parameters such as
total body length, body weight, and ovary weight (Murua et al., 2003)

58
Relative fecundity was expressed in terms of number of eggs per unit
length and unit weight of the fish and unit weight of the ovary. The linear relationship
between absolute fecundity and (i) Total length, (ii) Body weight and (iii) Weight of
ovary were computed by regression analysis after log10 transformation of the respective
X and Y values
Log Y = log a + b log X
This is calculated using the least square method and corresponds to an
exponential function of the type : Y = a.Xb. The linear equation was fitted by the method
of least squares, allowing the use of standard statistical procedures for subsequent
analysis. Scatter diagrams of fecundity against total length, fecundity against body
weight and fecundity against ovary weight were drawn. A regression line was fitted on
each scatter diagram by the least squares method (Draper and Smith, 1966).
Absolute fecundity
=
Total length (cm) or total weight (gm) or ovary weight (mg)
Estimation of mathematical equations that represent the relation between
absolute fecundity on one side and total length (cm) or body weight (gm) or ovary
weight (mg) on the other side was done.
Oocyte size-frequency distributions studies could be made use for fecundity
studies also. It is helpful to find out whether a fish shows determinate fecundity or
indeterminate fecundity. In fishes with determinate fecundity, the standing stock of
yolked oocytes (total fecundity) prior to the onset of spawning is considered to be
Relative fecundity (Fr)

59
equivalent to the potential annual fecundity; while, indeterminate fecundity refers to a
species where potential annual fecundity is not fixed before the onset of spawning and
unyolked oocytes continue to be matured and spawned during the spawning season
(Hunter et al., 1992).
3.3.2 Ova-diameter studies
Ova were counted following the direct method with a binocular compound
microscope. The ova diameter measurements were done on a monocular microscope
with 4×10 magnification, after calibrating the ocular micrometer using a stage
micrometer. The ovaries were kept in Gilson’s solution (Simpson, 1951), in order to
disperse the oocytes; then, they were transferred to 70 % alcohol. The distribution of ova
in the ovary was studied by the method of Clark (1934) and Prabhu (1956). A ripe ovary
was arbitrarily divided into three segments along its length, the anterior, middle and
posterior. From each segment a batch of 800-1000 ova was isolated and the diameter of
each ovum was measured. These diameters were grouped in 64 µm intervals and the
percentage distribution of the groups of ova in each region was calculated. This
percentage occurrence in the mature ovary was used for studying the spawning
frequency.
Different methods have been adopted to classify the maturity stages of gonads in
fishes. In the present study the ovary was grouped into five maturity stages following
the scheme of Qayyum and Qasim (1964) and Bhatt (1968, 1970, 1971). The characters
used for the classification of the ovary were appearance, colour, size, state of distension,
relative space occupied in the body cavity, the size of the ova and their yolk content.

60
3.3.3 Gonadosomatic Index
Using the total and ovarian weights, a standard gonadosomatic index (GSI) was
calculated to determine the time of spawning and the general reproductive status of
females. Gonadosomatic index (GSI) of the female fishes of the collected samples was
determined separately following the equation cited by Parameswaran et al. (1974).
Weight of the gonad
GSI= ------------------------------ X 100
Weight of the fish
The studies were conducted under captive conditions on specimens which were
raised in hatchery. Because of the extremely limited distribution of the species most of
the methods adapted for biological studies were non-lethal. Only minimum samples
were formalin preserved in the case of unavoidable circumstances. The fishes which are
found to be the representative of same maturity by visual estimation of different
macroscopic characters were selected, preserved and dissected. The fish were raised at a
temperature range of 26 to 28 0C, which was the room temperature available in the
hatchery.
3.3.4 Size at first maturity
The length at which 50% of the fishes are mature can be considered as the
minimum length at first maturity (Kagwade, 1968). Size at first maturity or size at 50 %
maturity was computed with a total of 112 fishes of which 77 were females (ranging
from 25 mm to 60 mm TL) and 35 fishes were males (ranging from 20 mm to 55 mm

61
TL). The length at first maturity is the size at which 50 % of the population is mature.
The total lengths of all the fishes collected for the study were grouped according to
different length groups. The percentage occurrences of mature fishes (early ripening,
late ripening, ripe and partially spent) for the females and males have been taken. By
plotting the percentage occurrence of mature fish (males and females) against respective
length classes (5mm), the length at which 50% of the fishes were mature was
demarcated.
3.3.5 Sexual Dimorphism and Sex Ratio
Different macroscopic and visual features were made use of for determining the
sex of the individual. They include
1. Overall body coloration (sexual dichromatism)
2. Bulginess of the stomach
3. Behaviour in captive conditions
A total of 95 nos of specimens were made use of for sex determining studies and
they included wild caught individuals right from their natural habitats, and F1 and F2
generations of the wild caught ones. Different nuptial and breeding behavioural gestures
like chasing, following, nubbing etc also could be used for distinguishing the sexes. The
percentage occurrence of each sex in the population was calculated.
The sexes were also determined after dissection based on different macroscopic
characteristics such as vascular irrigation, size, colour and proportion of oocytes per
developmental stages, and rate of abdominal cavity occupied by ovaries.
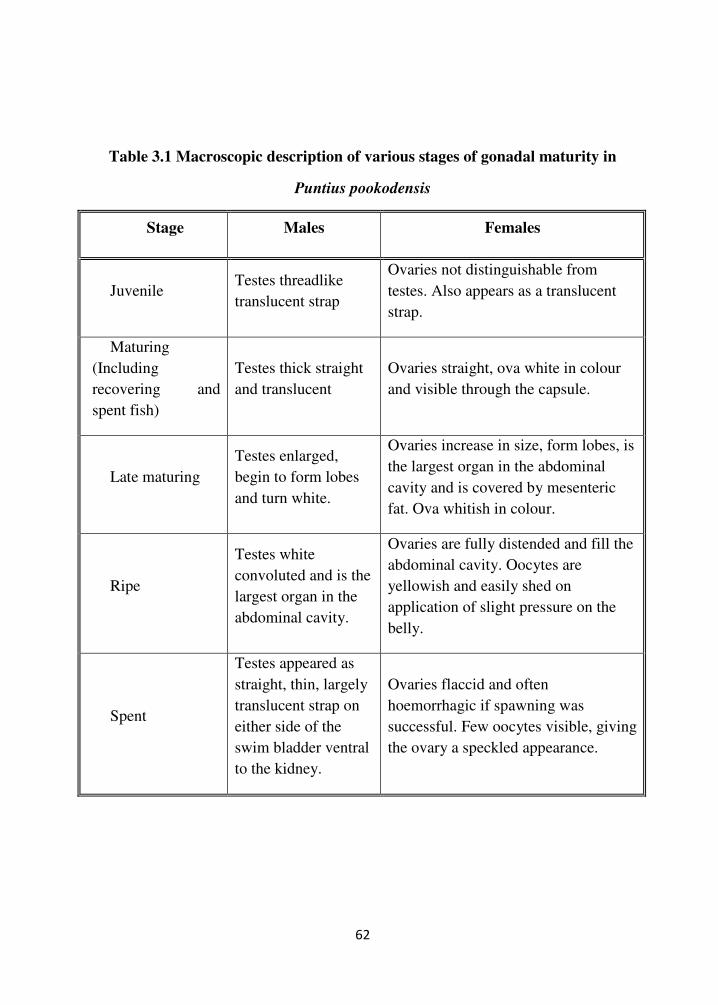
62
Table 3.1 Macroscopic description of various stages of gonadal maturity in
Puntius pookodensis
Stage Males Females
Juvenile Testes threadlike
translucent strap
Ovaries not distinguishable from
testes. Also appears as a translucent
strap.
Maturing
(Including
recovering and
spent fish)
Testes thick straight
and translucent
Ovaries straight, ova white in colour
and visible through the capsule.
Late maturing
Testes enlarged,
begin to form lobes
and turn white.
Ovaries increase in size, form lobes, is
the largest organ in the abdominal
cavity and is covered by mesenteric
fat. Ova whitish in colour.
Ripe
Testes white
convoluted and is the
largest organ in the
abdominal cavity.
Ovaries are fully distended and fill the
abdominal cavity. Oocytes are
yellowish and easily shed on
application of slight pressure on the
belly.
Spent
Testes appeared as
straight, thin, largely
translucent strap on
either side of the
swim bladder ventral
to the kidney.
Ovaries flaccid and often
hoemorrhagic if spawning was
successful. Few oocytes visible, giving
the ovary a speckled appearance.

63
3.3.6 Reproductive strategy
The different reproductive strategies of a fish species can be explained based on
the studies on different reproductive biological aspects like fecundity, ova diameter
frequency studies. Study on captive breeding and breeding behavioural patterns also
give picture of different reproductive strategies of a species.
3.4 RESULTS
3.4.1 Fecundity
Absolute fecundity (Fa)
In this study the absolute fecundity of Puntius pookodensis ranged from 375-
823. The highest absolute fecundity was observed as 858 for a specimen weighing 1.8
g. The average absolute fecundity was found to be 722.33
Relative fecundity (Fr)
The relative fecundity values ranged from 121.71-164.6 per centimeter (cm)
body length and from 349.36-767.57 per gram (g) body weight of fish. Fecundity
showed wide variations as far as the body weight is concerned. The species at a body
weight of 0.555 g showed the maximum relative fecundity, calculated as 720.5 per gram
body weight and a least was observed with specimens of 2.267 g with a count of 349.36
per gram body weight. The ovary weight ranged from 70-310 mg. The relative fecundity
values ranged from 2.55 - 7.05 per milligram (mg) ovary weight.
64
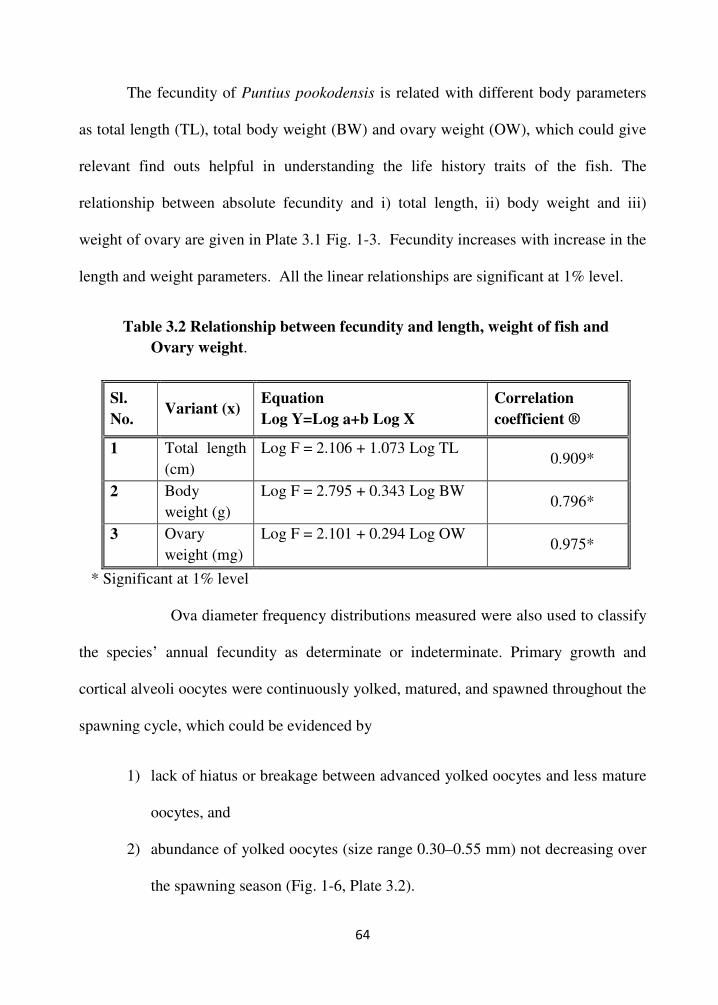
The fecundity of Puntius pookodensis is related with different body parameters
as total length (TL), total body weight (BW) and ovary weight (OW), which could give
relevant find outs helpful in understanding the life history traits of the fish. The
relationship between absolute fecundity and i) total length, ii) body weight and iii)
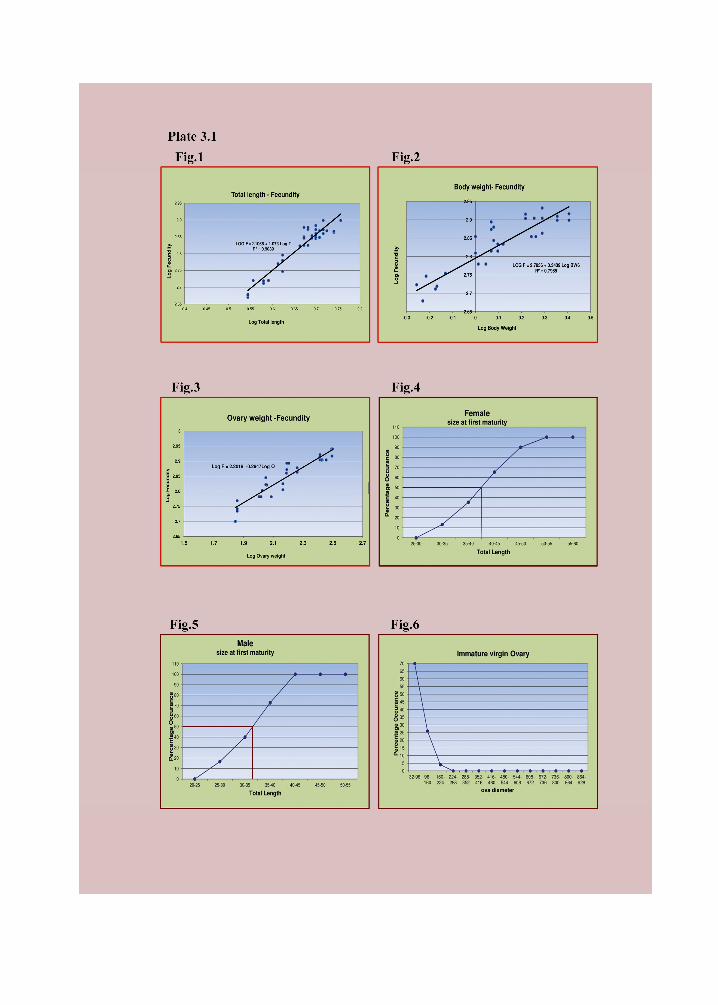
weight of ovary are given in Plate 3.1 Fig. 1-3. Fecundity increases with increase in the
length and weight parameters. All the linear relationships are significant at 1% level.
Table 3.2 Relationship between fecundity and length, weight of fish and
Ovary weight.
Sl.
No. Variant (x)
Equation
Log Y=Log a+b Log X
Correlation
coefficient ®
1 Total length
(cm)
Log F = 2.106 + 1.073 Log TL 0.909*
2 Body
weight (g)
Log F = 2.795 + 0.343 Log BW 0.796*
3 Ovary
weight (mg)
Log F = 2.101 + 0.294 Log OW 0.975*
* Significant at 1% level
Ova diameter frequency distributions measured were also used to classify
the species’ annual fecundity as determinate or indeterminate. Primary growth and
cortical alveoli oocytes were continuously yolked, matured, and spawned throughout the
spawning cycle, which could be evidenced by
1) lack of hiatus or breakage between advanced yolked oocytes and less mature
oocytes, and
2) abundance of yolked oocytes (size range 0.30–0.55 mm) not decreasing over
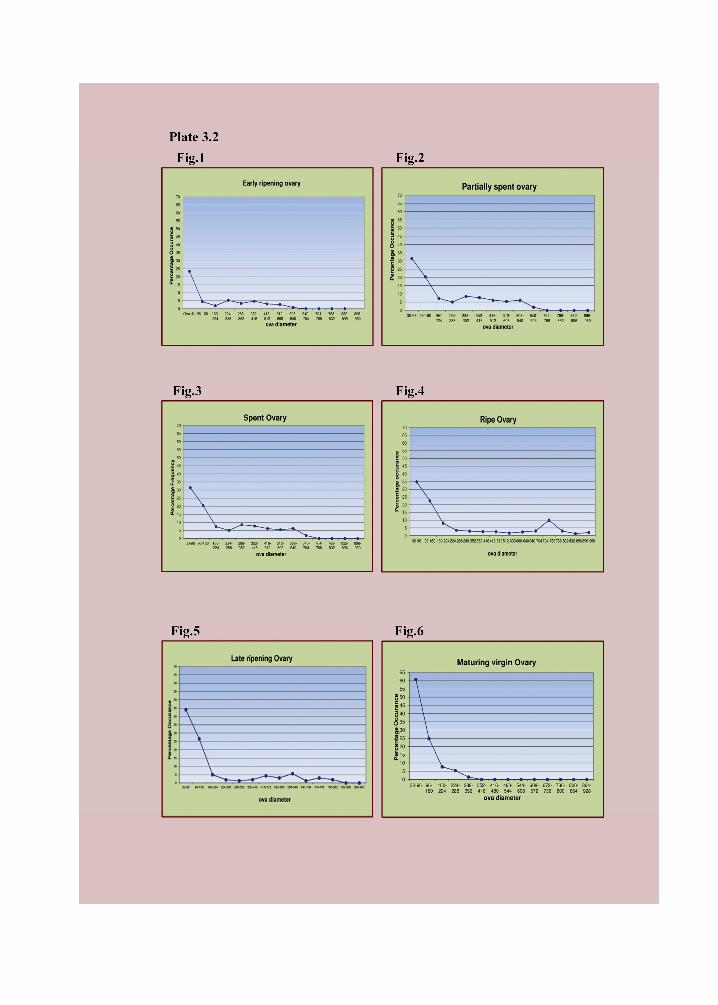
the spawning season (Fig. 1-6, Plate 3.2).

65
This type of development defines Puntius pookodensis as having indeterminate
fecundity. In the present study, it has been found that the number of eggs increases
linearly with the increase of body weight, body length, standard length and gonadal
weight.
3.4.2 Distribution of ova in the ovary
The frequency distribution of egg diameter of Puntius pookodensis was studied
in connection with the fecundity studies. The frequency distribution of eggs diameter in
the ripe ovary is divided into three batches. The first batch included the eggs diameters
ranged from 32 µ to 224µ. This batch represented the stock of immature eggs which
were very small and transparent with polygonal shape. This batch is considered as the
oocyte stock and it was represented throughout the whole year period. The second batch,
the ripening stock, included the vitelline eggs with a considerable amount of
cytoplasmic growth and yolk formation. Their diameters ranged from 225 µ to 640 µ.
The third batch included the more advanced ova, which are yolky and ripe transparent.
The diameters of ova of this batch ranged from 641 µ to 896 µ.
The distribution of oocyte-diameter frequency showed the presence of a reserve
stock of eggs of all stages of maturity throughout, which suggested that the fish is a
continuous spawner. For the percentage strength of the different size groups of ova,
there is a little difference between anterior, middle and posterior segments of each lobe
with two size groups of 50 -100 µm and 250-300 µm diameter dominant. The mean
percentage size frequency distribution of the ova showed that the two size groups of ova
of diameter 50 -100 µm and 250-300 µm are dominant. The actual percentage values of

66
the different diameter ranges of the ova showed that the size group of 100-250 µm
decline from the anterior to the posterior end of the ovary. In the smaller ova of the size
group 50-100 µm, the pattern was the reverse with an increase in their percentage
strength from the anterior to the posterior; in the other size groups the increase and
decrease was variable in the three regions.
During first month of its reproductive cycle the majority of the eggs were within
the first and second batches. At the end of the first month, two new batches of larger
eggs started to appear with a relatively small percent, 3 % and the mean GSI value
increased to 9.56. During the second month, the ovary represented multiple size batches
of oocytes. During the third, ovary reached its maximum growth and the GSI values
touched the highest point of 15.7. The egg diameters of these samples ranged from 32 µ
to 896 µ. The discharge process at this month was accompanied by a decrease in the GSI
value during late third month, which reached the value of 8.
The frequency distribution of egg diameter for P.pookodensis through six
months indicated that the fish discharged its ripe ova in batches during the spawning
period and withdrew from the egg stock to undergo a continuous maturation process
successfully. Moreover, the presence of more than two modes of egg size, indicated
fractional spawning character and a long spawning season. Two patterns of oocyte
development were common among multiple spawning fishes, viz., group synchronous
and asynchronous oocyte development. In P. pookodensis, oocyte size-frequency
distributions showed no distinct gaps in development, or modes of oocytes, except for

67
hydrated oocytes (ripe stock) (640 µ to 896 µ), a finding that indicated asynchronous
oocyte development and the species was found to exhibit a multiple spawning pattern.
Ova diameter frequency showed that the immature and ripe stocks have
single batch of ova, but the ripening stock showed many batches which were not well
differentiated from each other. There was a large stock of immature ova constituting to
about 67.25% of the total ova count and ranged over a diameter of 32-224 µm with
maximum value at 32-96 µm range. The ripe stock was about 17.34 % with ova size
going up from 640 µm and with a mode at 704-768 µm diameter class. The maximum
size of ova diameter recorded was 896 µm. There was always the presence of large
percentage of immature stock in any developmental stage of the ovary and a
distinguishable stock of ripe ova in the ripe ovary.
The ripening stock contributed to the remaining 15.42 % ranging from 224 - 640
µm. This stock consisted of many batches with different stages of maturity. This
indicated that the spawning may be extended over a very long period of almost round
the year with the individual spawning intermittently.
In P. pookodensis three peaks are very well evident without complete separation
between the groups in the case of a ripe ovary, one peak lies in continuation to the other.
It also means in a ripe ovary eggs of all stages are present at any time of a year. It
clearly showed that the fish is strictly a continuous spawner rather than an annual or
seasonal spawner.

68
3.4.3 Classification of the Maturity stages
The ovaries were classified into the following maturity stages.
(i) Stage I, Immature : The ovary was slender, thread-like, white and
semitransparent, occupying less than one quarter of the body cavity. Sex was difficult to
be determined by naked eye in this stage. The ova were transparent and spherical
without yolk and with a large nucleus in the centre. The mode of ova was in the range
94-124 µm with the largest measuring 214 µm in diameter.
(ii) Stage II, Maturing virgins or recovered spent: The ovaries were large and
broader, occupying one half of the body cavity, colour was light yellow. Ova with two
modes (Plate 3.2 Fig 6), in the range of 125-154 µm and 185-214 µm with largest ovum
measuring 244 µm. The older ova were opaque and light yellow due to the
commencement of yolk formation and the younger ova were colourless and transparent.
(iii) Stage III, Maturing : Ovaries were larger and occupy about three - quarters of
the body cavity; colour, distinct yellow. The entire ovary was flaccid with the outer
membrane slightly distended and thinner than in the previous stage. The older ova are
yolky and opaque, their mode is 275-304 µm (Plate 3.2 Fig 6) and the largest ovum
measures 364 µm.
(iv) Stage IV, Ripe : The ovaries were further enlarged and occupied almost the
entire body cavity, they are bright yellowish, with a brownish tinge invaded with blood
vessels. The outer membrane was further distended and thinner so that the ova were
visible externally. The ovary was turgid and a slight pressure could break the outer
membrane. The ova were fully yolked and spherical in shape. At this stage the ovary

69
contained two batches of ova, one ripe and the other maturing, the latter have started
accumulating yolk. The larger ova are in the range of 335-364 µm (Plate 3.2 Fig 4) and
the largest ovum measures 424 µm.
(v) Stage V, Spent : The ovaries are shrunken, retracted and flesh coloured and
occupying one – third of the body cavity. Ova with a mode of 155-184 µm in diameter
(Plate 3.2 Fig 3), probably representing the second batch of the earlier stage which have
accumulated more yolk. They were opaque and slightly yellow. The larger eggs are 304-
394 µm diameter in size and represent the unspawned ripe eggs. This stage resembled
Stage II, but can be distinguished by the wrinkled and collapsed nature of the outer
membrane and by the presence of a few ripe ova.
3.4.4 Gonadosomatic index
Gonadosomatic index indicated gonadal development and maturity of fish. It
increased with the maturation of fish and declined abruptly thereafter (Parameswarn et
al., 1974). Due to the special conservational status of Puntius pookodensis all the GSI
studies were limited to captive environments available in the hatchery. The maximum
GSI indicated that the fish is mature enough to spawn in captivity, which was 15.7 in the
case of P. pookodensis. The minimum GSI showed that the fish is spent or immature.
The time taken by the fish to attain maximum GSI indicated the time needed by the
species to attain maturity under captivity, which could be of enormous value in raising
the fish in hatcheries. In the case of P. pookodensis, the time difference between the
least and maximum GSI values were found to be at the range of 75 to 90 days for
female. Here the ‘minimum’ indicated the spent stage rather than immature. This could

70
be interpreted as the fish is able to spawn at every 75 to 90 days, if all the conditions are
favourable, and also it is not an annual or seasonal spawner, but it is a continuous
spawner.
3.4.5 Size at first maturity and studies on initial sexual maturity
The length (total length) at first maturity was determined by analyzing the data
relevant to all mature fishes (stage III and above examined). The percentage occurrence
of mature fishes (early ripening, late ripening, ripe and partially spent) was plotted
against different length classes of 5 mm for both the female and male fishes.
While the first mature male fishes appeared in the 25-30 mm (TL) group (16.66
%), the first mature females appeared only in the group of 30-35 mm (13.33 %). All
male fishes were mature on reaching a total length of 40 mm and all female fishes on
reaching a length of 45 mm total length. The size at first maturity for males was 33.5
mm TL (30-35 mm) and 39.5 mm TL for females (35-40 mm). (Plate 3.1 Fig. 4 and 5 )
The smallest mature male is within 25-30 mm length. If the length at
which 50% of the fishes are mature can be considered as the minimum length at first
maturity (Kagwade, 1968), the specimens below 32.5 mm TL for males and 38.5 mm
TL for females were not mature. The present study showed that the smallest mature
male is smaller than that of the mature female in P. pookodensis.
3.4.6 Sexual dimorphism and Sex ratio
Puntius pookodensis did not possess any organ or specialised structures with
which the sexes are distinguishable as some other cyprinids like gold fish, Carassius

71
auratus which has nuptial tubercles and Indian major carps which have characteristic
serrated pectoral fin rays. The sexes of P. pookodensis were distinguishable by making
use of some other methods, as follows.
The male and female fish showed clear differences in body colorations which
could be termed as sexual dichromatism. The colour differences become prominent by
the onset of sexual maturity. The immature fish appeared in a dull greyish silvery. A
sexually mature male develops a pinkish red tinge all over the body. Deeper colour
shades were noticed towards the caudal peduncular region. The colour showed abrupt
changes as the fish became sexually active. The depth of the colour reached in its
maximum during the courtship activities. But a sexually mature female did not develop
any colour change by the onset of sexual maturity. It remained in the same colour
pattern as that of a juvenile.
Another distinguishing character was the bulginess of the stomach. A sexually
mature female exhibited a more swollen and deeper stomach than that of the males.
Other criteria found useful in distinguishing the sexes were the reproductive
behavioural patterns it exhibited during the onset of maturity.
In the present study out of 214 fishes, 128 were males and 86 were females. The
total male and female ratio was 1: 1.48. The percentage occurrence of the males was
found to be 40.1 while the same of the female was 59.8.

72
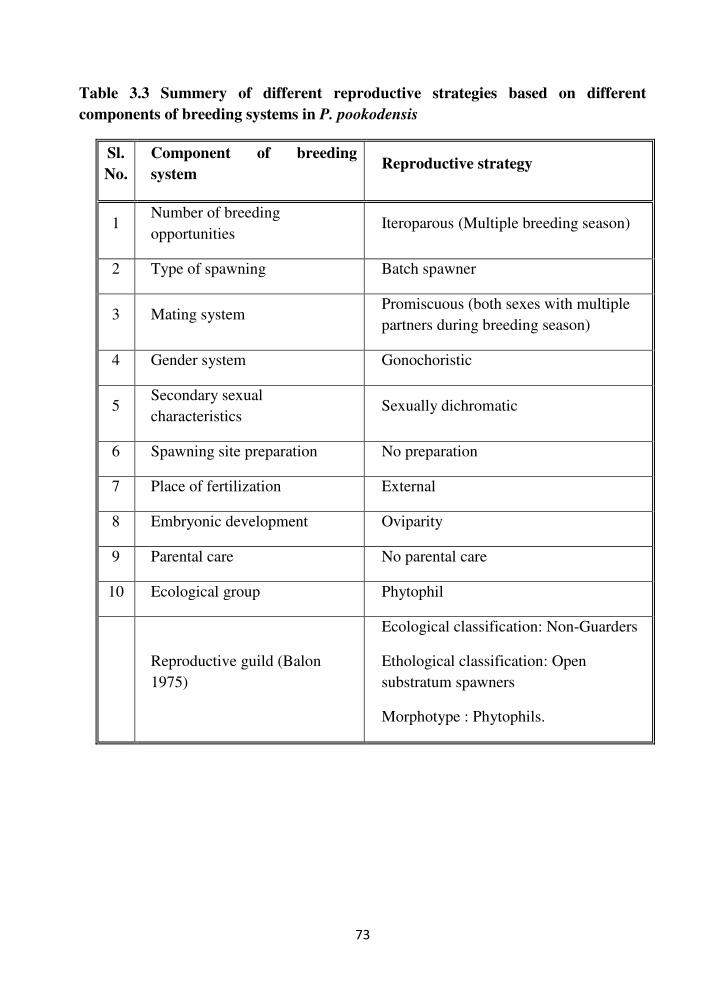
3.4.7 Reproductive strategy
Studies on different aspects of reproductive biology such as fecundity, ova
diameter analysis and gonadosomatic index revealed that the candidate fish species P.
pookdensis is an asynchronous spawner i.e., a continuous development and release of
gametes in the gonads are evident. The fish showed affinity towards aquatic plants or
the presence of plants stimulated the fish to start the spawning activities. So the fish
could be included in the group of phytophils.
In general, the reproductive strategies of P. pookodensis showed that it is an
iteroparous species, i.e., they spawn more than once during their lives and gonochoristic,
that is their sexes were separate and exhibited external fertilization without parental
care. A summary of reproductive strategies shown by P. Pookodensis is shown in table
3.3. The fish possessed an asynchronous type ovary i.e., oocytes of all stages of
development are present without dominant populations.
The species could be categorized as a batch spawner i.e., eggs are recruited and
ovulated from the population of yolked oocytes in several batches over a protracted
period during each spawning season.
73
Table 3.3 Summery of different reproductive strategies based on different
components of breeding systems in P. pookodensis
Sl.
No.
Component of breeding
system Reproductive strategy
1 Number of breeding
opportunities Iteroparous (Multiple breeding season)
2 Type of spawning Batch spawner
3 Mating system Promiscuous (both sexes with multiple
partners during breeding season)
4 Gender system Gonochoristic
5 Secondary sexual
characteristics Sexually dichromatic
6 Spawning site preparation No preparation
7 Place of fertilization External
8 Embryonic development Oviparity
9 Parental care No parental care
10 Ecological group Phytophil
Reproductive guild (Balon
1975)
Ecological classification: Non-Guarders
Ethological classification: Open
substratum spawners
Morphotype : Phytophils.

74
3.5 DISCUSSION
3.5.1 Fecundity
The studies on fecundity have great importance, especially for a species in
conservation point of view. It is certainly unavoidable in the case of a species which is
facing threat of extinction or a species which extremely limited in distribution in its
natural habitat. It is difficult to determine the fecundity in females which spawn in
batches and in which oocytes develop asynchronously, i.e. such as the species of fish
under study. The absolute fecundity is described as the number of, usually mature eggs,
filled with yolk or all vitellogenic oocytes found in the ovary immediately before the
reproduction process, which can be laid at spawning sites during the season under study.
The individual or absolute fecundity refers to the number of eggs produced per female
per year (Wootton, 1979), and can also be defined as the number of mature oocytes
present in the ovary immediately before spawning (Bagenal, 1973). In species that has
multiple spawning, it is the number of oocytes destined for spawning i.e., the one that
will mature during the current reproductive cycle, which are usually taken into account
(Aboussouan and Lahaye, 1979). Therefore only oocytes with a diameter greater than
those with the cortical alveoli stage were taken into account, since only these are
considered to have been released in this reproductive cycle. Thus absolute fecundity
tends to increase according to the size and age of the fish. Therefore, in order to
facilitate the comparison, relative fecundity, i.e. the number of eggs per unit weight
(Bagenal, 1978) was also calculated.

75
Although fecundity is described as the number of eggs produced by a female,
there exist a variety of terms describing the different facets of fecundity. Potential
annual fecundity is defined as the total number of advanced yolked oocytes matured per
year, uncorrected for atretic losses (Huter et al., 1992). Annual realised fecundity,
however, is the actual (or real) number of eggs finally released, so it is equal to or lower
than the potential fecundity, since some of the eggs can be reabsorbed through atresia
during spawning, or simply that some of the eggs are not able to be liberated, remaining
in the ovary and being reabsorbed later. Total fecundity is defined as the standing stock
of advanced yolked oocyted at any time (Huter et al., 1992). Batch fecundity is the
number of eggs spawned in each batch, and consequently, the sum of batch fecundities
is the realised annual fecundity. Finally, annual population fecundity is the number of
eggs that all the females in a population spawn in a breeding season (Bagenal, 1978).
The gravimetric method was used in computing the absolute fecundity and
according to May (1967), it is useful in minimizing the error due to sampling. Studies of
fecundity provide information on various aspects of fishery biology such as population
dynamics (Scott, 1962; McFadden et al., 1965; Stauffer, 1976), racial characteristics
(Bagenal, 1966), environmental conditions like rain fall and salinity (Joshi and Khanna,
1980) and stock assessment. Nowadays fish fecundity attains an important role in
aquaculture. In earlier days, fecundity was estimated in mouth brooding cichlid fishes by
the number of young ones produced. Fecundity is very much closer to the body
parameters such as body length, body weight, and ovary length and ovary weight. Many
authors reported that fecundity of fishes increased in relation to body length (Sarkar et
al., 2004). Simpson (1951) established that the fecundity related to cube of its length

76
and it was directly proportional to body weight. Many authors supported Simpson’s
concept (Bagenal, 1957; Sarojini, 1957; Varghese, 1977 and Kurup, 1994).
The exponential value of regression analysis plays a vital role. The values ranged
from 2.3 to 5.3 and frequently exceeded 3. Cotton (1970) reported exponent values
between 3.25 to 3.75 in marine fishes, invariably higher values were recorded in marine
fishes than the freshwater fishes.
In the present study, fecundity showed a linear relationship to body weight. The
fecundity is most significantly correlated with gonadal weight. The correlation
coefficient of fecundity with total length is almost similar to that of fecundity with body
weight. Similar findings were found in six Barbus species studied in SriLanka (De
Silva et al., 1985). Linear relationship has also reported in Opichthus rufs (Casadevall et
al., 2001), Labeo calbasu (Rao and Rao, 1972), Labeo fimbriatus (Rao, 1974 and Khan
and Jhingran, 1975), Labeo desoi (Raina and Bali, 1982 Cirrhinus mrigala and Labeo
rohita (Tripathi, 1989),; Sarojini, 1957 and Gupta, 1967). In general, the correlations are
highly significant, but with much variation, which is far from being fully explained by
the factors total length, body weight and gonad weight. It is likely that other factors
such as age and environmental factors contribute considerably to the variation in
fecundity, independently from body weight and gonad weight (De Silva et al., 1985).
Ovary weight is the most important factor to settle fecundity in fishes. In the
present study a significant relation between fecundity and ovary weight was noticed.
Similar finding was supported in M. cavasius (Sharma, 1989), Channa gachwa (Misra,
1991), Tilapia nilotica (Babikar and Ibrahim, 1979), Lepidocephalus thermalis

77
(Maxwell Samuel, 2002), Sauriendo squamis (Allishman, 2003). L. gonius (Joshi and
Khanna, 1980), Schizothorax richardsonii (Quadri et al., 1983) and Sunder (1986) in S.
longipinnis. The r-value calculated between ovary weight, body weight, ovary length
and body weight exhibited a fair relationship, but ovary weight and body weight were
much more related than the other parameters. Similar trends were reported by Kurup
(1994) in L. dussumieri. The present observation on fecundity revealed that P.
pookodensis has higher fecundity. It is possible to suggest that highest number of eggs
produced were compensated for heavy mortality during early development. Moreover,
higher fecundity is inevitable to supply adequate amount of seeds during breeding
season.
Studies show that repeat spawners have larger fecundity than recruit spawners.
The number of eggs released increased with age and size ranging from 426 for 35 mm
fish to 823 for a fish of size 50 mm. Similar studies have been done on Danio
malabaricus and in Puntius melanostigma (Anna Mercy et al., 2005c).
3.5.2 Ova-diameter studies
The knowledge on the frequency distribution of ova and their diameter is helpful
in predicting the breeding habit and reproductive strategies of a fish species. In the case
of P. pookodensis distribution of oocyte-diameter frequency showed a reserve stock of
eggs of all stages of maturity which suggested that the fish is a continuous spawner.
The quantification of maturity stages is based on the distribution of oocyte stocks in the
different maturity stages. In the present study, it can be seen that the ovaries of Puntius
pookodensis show asynchronous development. ‘Asynchronous ovaries’ show sufficient

78
number of oocytes at various stages of development within the ovary (De Vlaming,
1983). The oocyte size frequency distribution was continuous; however in a ripe ovary
a clear cut mode for the ripe stock was obtained. The results showed that P. pookodensis
has never got a distinct breeding season. The ova diameter distribution studies showed
that it has a continuous maturation process. The availability of quality food may be the
most important criteria in the growth and development of ovary. The occurrence of ova
of different size gradations between immature and ripe ovaries is considered as an
evidence of prolonged spawning period (Sathyanesan 1962).
The ova diameter investigation in the present study indicated that the maximum
size of ripe ova was 896 µm in diameter, which was observed from the third month of
developmental cycle of ovary and it was noticed that at the end of the third month of
ovarian development the fish attained its highest GSI. The egg size was probably related
to the amount of food that the females can metabolize, assimilate and store in each egg
(Neophitou, 1988). Ware (1975) pointed out that egg size was remarkably constant for a
given marine fish species in a given geographical area and this theory could be applied
in the case of P. pookodensis also.
The use of size frequency analysis of oocytes revealed three oocyte groups in the
ovary of P. pookodensis. This indicated that the mature fish discharged its ova in
batches during prolonged spawning period i.e. they are fractional spawners. The
fractional and prolonged spawning season are characteristic features of the tropical and
subtropical fish species (Nikolsky, 1963).

79
Zaki et al., (1995) stated that the analysis of ova diameter for Oblada melanura
revealed that there were nine diameter groups of ova in different samples. Allam (1996)
pointed out that ripe ovaries of Trachinotus ovatus contained three modes of oocytes
(primary, intermediate and most advanced oocytes). Oocytes of both intermediate and
most advanced oocytes were spawned during one spawning season. Massut and Nin
(1997) pointed out that size distribution of oocytes with at least two groups of oocytes in
the ovaries suggested that Coryphaena hippurus is a multiple spawner with an extended
spawning season in the study area. According to Mustafa (1991), the Indian cyprinid
Puntius sophore might spawn twice a year.
3.5.3 Gonadosomatic index
The gonado somatic index was observed to increase with the maturation of fish,
being the maximum during the period of peak maturity and declining abruptly
thereafter, when the fish become spent. The monthly changes in the GSI reflect the
ovarian activity of a fish. The dynamics of gonad maturation is a good tool to indicate
the time in which females are capable of reproducing. P. pookensis showed
comparatively high GSI values. The maximum value of 15.7 was reported in the present
study. This is certainly an indication of huge ripe ovaries at the maximum gonadal
maturity and the body depth of the fully mature female will be considerably great as it
facilitates the visual estimation of ripe fishes in a population. Months with great number
of females with mature oocytes, followed by months with females with empty gonads
were clues to the oocyte laying period, mainly if the macroscopic condition of this ovary
may be confirmed by microscopic analysis (Dias et al., 1998). The maturity stages of

80
females were recorded, according to Vazzoler (1996), counting out the immature gonads
in stage A, since their sex could not be identified macroscopically.
3.5.4 Size at first maturity
In Puntius pookodensis, the size at first maturity was found to be 39.5 mm TL
(40-45 mm) for females and 33.5 mm (30-34 mm) for males. The largest female
obtained during the present study was 57 mm TL while the largest male is 50 mm TL. It
was the female that was larger or at least achieved a larger size. This is a kind of
reproductive strategy to enhance the existence of the race by increasing the fecundity,
directly related to the size of the female fish (Keenleyside, 1991). The females spent
more energy first for somatic growth and from there on, gonadal growth and maturation.
However, the early maturation of the males diverted all the energy reserves towards the
gonadal development and its maturation at a relatively younger age. In Puntius
pookodensis the males attained sexual maturity at a smaller length than females. Similar
observations had been reported in many freshwater fishes like Labeo boggut (Selvaraj et
al., 1972), Barbus sarana (Murthy, 1975), Nemacheilus triangularis (Ritakumari and
Nair, 1979), Puntius denisonii, P. filamentosus, Nemacheilus triangularis and N.
semiarmatus (Anna Mercy et al., 2005 b), Pristolepis marginata (Nisha Raj, 2005) and
Chela fasciata (Divipala, 2008).
Anna Mercy et al. (2005a) reported the size at first maturity of Puntius
melanostigma as 50 mm for males and 55 mm for females. In the case of the African
minnow, Barbus paludinosus sexual maturity was reached within a year at 50.0 mm TL
(Cambray and Burton, 1985). In the case of European minnow, Phoxinus phoxinus the
short lived populations of river Frome in England contained two spawning age groups

81
and the largest fish caught was only 78.0 mm long. The size at first maturity ranged
from 50-55 mm as two year olds (Mills, 1987). Six Barbus species studied in Sri Lanka
had maximum total length of between 42.0 and 101.0 mm and a short life span (De Silva
et al., 1985). In the freshwaters of South Africa out of the 52 Barbus species studied 43
attained maximum fork lengths of less than 150.0 mm (Cambray and Burton, 1985).
The information on initial sexual maturity gives the ornamental fish producers the idea
on the age at which the fish starts to be mature, thus they could prepare the reproduction
and an appropriate nourishment environment for the fish to spawn and obtain the
maximum number of fry, depending on how many female fish are required for this
purpose. In general, heavier fish have heavier gonads, which produce more ova per fish
gonad.
3.5.5 Sexual dimorphism and sex ratio
Sexual dimorphism is widespread across the animal kingdom. Males and females
usually differ not only in reproductive organs, but also in external structures that are not
directly related to reproduction Information about sexual dimorphism is required for
understanding the ecology, behavior, and life history of a species. In
addition, knowledge of sexual dimorphism and its appearance during ontogeny is
indispensable when making morphological comparisons between populations.
Although sexual differences in a variety of external structures have been noted in many
Populations, studies on the sexual difference in fresh water fishes of India are less. A
comprehensive study is by Inasu (2008) in which sexual dimorphism of 26 species of

82
Indian fishes is compiled. Anna Mercy et al. (2005) has described the sexual
dimorphism of 13 species of fresh water fishes of the Western Ghats of India.
Puntius pookodensis exhibited sexual dichromatism rather than sexual
dimorphism. Breeding adults of male Puntius pookodensis had marked sexual
dichromtism. Males had a broad, reddish lateral band just behind the mid line
extending from below the origin of the dorsal fin to the caudal peduncle. Coloration
before the dorsal fin was normally a dusky-grey. The ventro-lateral surfaces were also
reddish. The posterior part of the body became intense red at the time of courtship.
Sexually inactive, adult males retained the lateral band, but the body coloration
was a more uniform light grey. The coloration in the fins was also not as pronounced.
Reproductive females did not have this red colour. The colour change in the body only
became significant after the fish started breeding. This suggests that sexual
dichromatism in the body is a secondary sexual character that may be regulated by
reproductive hormones. Although sex in Puntius pookodensis is genetically determined
it is currently unknown what genes or hormones might regulate secondary sexual
dichromatism of body in this fish. Further analysis of the genetic and developmental
mechanisms that underlie sexual dimorphism in P. pookodensis will be possible by
using the recently established genomic tools (Peichel et al., 2001; Peichel, 2005) and
will provide a complement to ecological studies to discern the functional significance of
sexual dichromtism in P. pookodensis. Similar type of secondary sexual characters were
also observed in P. melnostigma (Mercy et al., 2005).
As previously stated, in the present study the sex ratio of Puntius pookodensis
was found to be 59 % female and 41 % male. This is contrary to that of many other

83
cyprinid species where male populations dominated (Tejavej, 2012). The rearing
temperature ranged from 25 to 27 0C. As the background information on the
requirements for sex determination of Puntius pookodensis was limited, the mechanisms
involved in sex determination have yet to be elucidated. Further studies should,
therefore, be carried out to determine whether Puntius pookodensis has a species-
specific sex determination mechanism or possesses a major gene determining sex and/or
sex chromosomes.
The magnitude and direction of sexual dimorphism of both external and internal
characters can be predicted by examining the reproductive roles of the sexes within a
species and the differences in the reproductive roles occupied by the sexes should
influence patterns of selection and thus should ultimately lead to sex differences in
morphology. (Casselman and Schulte-Hostedde, 2004).
3.5.6 Reproductive strategy
A complete knowledge of the reproductive system and the reproductive biology
of fishes were essential to understand the reproductive strategy of any given species. As
far the natural habitat of P. pookodensis was concerned, the different reproductive
strategies adapted by the species were well suited to thrive on the particular
environment. The wide spectrum of reproductive strategies supports a diversity of
adaptive processes by which species have adapted and populated in the environment. In
addition to aiding in assessing fecundity, understanding these mechanisms of
reproduction could also lead to greater comprehension of the underlying mechanisms of
variable fish recruitment (Murua and Saborido-Rey, 2003). Reproductive potential is a

84
measure of the capacity of a population to produce viable eggs and larvae, and can be
considered as the main outcome of a reproductive strategy (Murua and Saborido-Rey,
2003) and the reproductive strategies of fishes have long been a central issue in fisheries
biology, ecology, and management (Beverton 1963, 1992; Balon 1975).
P. pookodensis was found to possess an asynchronous type of ovary, because
oocytes of all stages of development are present without dominant populations. The
ovary appeared to be a random mixture of oocytes, at every conceivable stage. Such
ovaries may be found in iteroparous species, with protracted seasons and yolk
accumulation, and hence oocyte development relies mostly on the food available in the
environment at that moment (Hunter and Leong, 1981). Because the fish recruited and
ovulated the yolked oocytes in several batches over a protracted period during each
spawning season, it can be classified as a batch spawner. Batch spawning can be seen as
a strategy to release eggs over a long period of time increasing the survival probability
of offspring (Lambert and Ware, 1984).