Embed Size (px)
Citation preview

correspondence
nature genetics | volume 40 | number 9 | september 2008 1035
Eeles et al. reply: We recently conducted a genome-wide association study (GWAS) using data from 1,854 individuals with clini-cally detected (not PSA screened) prostate cancer diagnosed at <60 years or with a family history of the disease, and 1,894 population-screened controls with a prostate-specific anti-gen (PSA) of <0.5ng/ml (ref. 1). These were analyzed for 541,129 SNPs using the Illumina Infinium platform. We then evaluated puta-tive associations using a further 3,268 cases and 3,366 controls. After these two stages, associations at seven loci, on chromosomes 3,6,7,10,11,19 and X, reached genome-wide levels of significance (P = 2.7 × 10−8 to P = 8.7 × 10−29). The SNP rs2735839 on chromosome 19 lies between two kallikreins, PSA (encoded by KLK3) and hK2 (encoded by KLK2). It was associated with a per allele OR for prostate cancer of 0.83 (95% CI = 0.75–0.91; Ptrend in stage 2 = 0.0002; Ptrend overall = 2 × 10−18). We also showed that rs2735839 was strongly associated with PSA level, in the direction consistent with the disease association (per allele rise in geometric mean PSA = 1.12., P = 6 × 10−8).
Ahn et al.2 analyzed 24 tag SNPs in the kal-likrein region on chromosome 19 (to include KLK1, KLK2, KLK3 and KLK15) in five stud-ies and found that none showed a significant association with prostate cancer risk. They also confirmed the association between sev-eral SNPs, including rs2735839, and PSA level. They raise the possibility that the asso-ciation found with prostate cancer risk in our study may reflect the selection of subjects on the basis of PSA levels rather than a causal relationship with prostate cancer risk.
It is clear that the selection of controls in stage 1 of our study for low PSA levels did influence the association in stage 1. This is reflected in the minor allele frequency for rs2735839, which is 21.1% in the stage 1 controls, compared with 14–15% in the UK 1958 birth cohort and the CGEMS study (which included both males and females). However, the controls in stage 2 were not highly selected for PSA level. The only selec-tion was to exclude controls with PSA levels of >10 and to require a negative prostatic biopsy if the PSA was >4. The MAF in the stage 2 controls (15.2%) is similar to that in other control populations and indicates that any selection bias at this stage was minimal.
To further evaluate the evidence for this association, we have undertaken an analy-sis of rs2735839 (together with SNPs at the other loci identified in our GWAS) in 13 further case-control studies as part of the PRACTICAL Consortium. These studies comprise 7,370 prostate cancer cases and
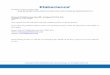
5,342 controls. The estimated per allele OR for prostate cancer associated with rs2735839 was 0.89 (95% CI = 0.83–0.95; P = 0.0007), very close to our original estimate3. There was no evidence of heterogeneity in the OR estimates among studies (Fig. 1). We also note that when data from the five CGEMS studies are combined, the per allele OR is also notably similar (per allele OR = 0.90, 95% CI =0.83–0.90; P = 0.01), although this was not formally significant using the 4-de-gree-of-freedom test given by Ahn et al.2. If the results from our stage 2, PRACTICAL and CGEMS are combined, the overall evi-dence of association reaches genome-wide levels of significance (P < 10−8), demon-strating that, even disregarding our stage 1 result, the association is unlikely to be due to chance. The overall effect size, although modest, is comparable to that seen for other cancer-associated loci.
None of the control series used in PRACTICAL or in CGEMS involved selection for PSA level, and for this reason and those given above, the association seems unlikely to be driven purely by control selection. Selection bias related to case ascertainment is an alter-native possible explanation. We excluded from our GWAS any cases identified through PSA screening, and several of the studies included
in PRACTICAL are drawn from populations where PSA screening has not been used (for example, the study from Finland). Thus, the association is unlikely to be due to PSA screen-ing for asymptomatic disease. PSA testing is, however, also used in the process of diagnosis of symptomatic disease. This raises the possi-bility of a more subtle bias, in that some cases may have raised PSA related to their genotype but not related to their disease. Whether or not this potential bias is significant could be resolved using genotyping in studies based on biopsy of whole populations not driven by the PSA level4 or studies where mortality is the endpoint.
Conversely, there are plausible biological grounds for believing that the association with KLK polymorphisms may be causal. For example, polymorphisms in the promoter of KLK3 are associated with alterations in androgen receptor binding5. Moreover, it is known that PSA level is a long-term predic-tor of prostate cancer risk6, and it is plausible that determinants of PSA level, including genetic determinants, may influence pros-tate cancer risk.
Rosalind Eeles1,2, Graham Giles3,4, David Neal5,6, Kenneth Muir7 & Douglas F Easton8, for the PRACTICAL Consortium9
OVERALL
TAMPERE
MAYO
USC WhiteUSC HispanicNC−CCPC White
NC−CCPC BlackUSC Black
MCCS
FHCRC
QLDMONTREAL
HaPCS
VALAISULMUKGPCS
PROTECT
10.5 1 1.5 2
Odds ratio
Figure 1 Forest plot giving the per-allele odds ratios between rs2735839 and prostate cancer for each study in PRACTICAL. Horizontal lines represent 95% confidence intervals. The area of each square is inversely proportional to the variance of the log(odds ratio). The diamond represents the combined odds ratio and 95% confidence interval, estimated by logistic regression adjusted for study.
©20
08 N
atur
e P
ublis
hing
Gro
up
http
://w
ww
.nat
ure.
com
/nat
ureg
enet
ics

correspondence
1Translational Cancer Genetics Team, The Institute of Cancer Research, 15 Cotswold Road, Sutton, Surrey SM2 5NG, UK. 2The Royal Marsden Hospital NHS Foundation Trust, Downs Road, Sutton, Surrey SM2 5PT, UK. 3Cancer Epidemiology Centre, The Cancer Council Victoria, 1 Rathdowne Street, Carlton VIC 3053, Australia. 4Centre for Molecular, Environmental, Genetic and Analytic Epidemiology, The University of Melbourne, 723 Swanston Street, Carlton VIC 3053, Australia. 5Surgical Oncology (Uro-Oncology S4), Departments of Oncology and Surgery, University of Cambridge, Box 279, Addenbrooke’s Hospital, Hills Road, Cambridge CB2 2QQ, UK. 6Cancer Research UK Cambridge Research Institute, Li Ka Shing Centre, Robinson Way, Cambridge CB2 0RE, UK. 7University of Nottingham Medical School, Queens Medical Centre, Nottingham, NG7 2UH, UK. 8Cancer Research UK Genetic Epidemiology Unit, University of Cambridge Strangeways Laboratory, Worts Causeway,
Cambridge, CB1 8RN, UK. 9A full list of authors is provided in the Supplementary Note online. Correspondence should be addressed to R.E. ([email protected]).
Note: Supplementary information is available on the Nature Genetics website.
ACKNOWLEDGMENTSThis work was supported by Cancer Research UK Grant C5047/A3354. D.F.E. is a Principal Research Fellow of Cancer Research UK. We would also like to thank the following for funding support: The Institute of Cancer Research and The Everyman Campaign, The Prostate Cancer Research Foundation, Prostate Research Campaign UK, The National Cancer Research Network UK, The National Cancer Research Institute (NCRI) UK, grants from the National Health and Medical Research Council, Australia (209057, 251533, 450104), VicHealth, The Cancer Council Victoria, The Whitten Foundation and Tattersall’s. The ProtecT study is ongoing and is funded by the Health Technology Assessment Programme (projects 96/20/06, 96/20/99). The ProtecT trial and
its linked ProMPT and CAP (Comparison Arm for ProtecT) studies are supported by Department of Health, England, Cancer Research UK grant number C522/A8649, Medical Research Council of England grant number G0500966, ID 75466 and The NCRI, UK. The epidemiological data for ProtecT were generated though funding from the Southwest National Health Service Research and Development. The views and opinions expressed herein are those of the authors and do not necessarily reflect those of the Department of Health of England.
COMpETiNG iNTErESTS STATEMENTThe authors declare competing financial interests: details accompany the full-text HTML version of the paper at http://www.nature.com/naturegenetics/.
1. Eeles, R.A. et al. Nat. Genet. 40, 316–321 (2008).2. Ahn, J. et al. Nat. Genet. 40, 1032–1036 (2008). 3. Kote-Jarai, Z. et al. Cancer Epidemiol. Biomarkers
Prev. (in the press).4. Thompson, I.M. et al. J. Clin. Oncol. 25, 3076–3081
(2007).5. Lai, J. et al. Carcinogenesis 28, 1032–1039
(2007).6. Lilja, H. et al. J. Clin. Oncol. 25, 431–436 (2007).
1036 volume 40 | number 9 | september 2008 | nature genetics
©20
08 N
atur
e P
ublis
hing
Gro
up
http
://w
ww
.nat
ure.
com
/nat
ureg
enet
ics