Embed Size (px)
Citation preview

Replication initiator DnaA binds at the Caulobactercentromere and enables chromosome segregationPaola E. Mera, Virginia S. Kalogeraki, and Lucy Shapiro1
Department of Developmental Biology, Beckman Center, Stanford University School of Medicine, Palo Alto, CA 94305
Contributed by Lucy Shapiro, October 1, 2014 (sent for review September 5, 2014; reviewed by James W. Gober and Matthew K. Waldor)
During cell division, multiple processes are highly coordinated tofaithfully generate genetically equivalent daughter cells. In bacte-ria, the mechanisms that underlie the coordination of chromosomereplication and segregation are poorly understood. Here, wereport that the conserved replication initiator, DnaA, can mediatechromosome segregation independent of replication initiation. Itdoes so by binding directly to the parS centromere region of thechromosome, and mutations that alter this interaction result incells that display aberrant centromere translocation and cell divi-sion. We propose that DnaA serves to coordinate bacterial DNAreplication with the onset of chromosome segregation.
DnaA | chromosome segregation | replication | Caulobacter | centromere
Cell division requires the faithful transmission of genetic in-formation to each daughter cell. Thus, in all forms of life,
multiple mechanisms cooperate to ensure that DNA synthesisand chromosome segregation are temporally controlled and co-ordinated. Unlike eukaryotes, in which chromosomes are fullyreplicated and organized into higher order structures beforesegregation (1), most bacteria segregate their chromosomesprogressively during replication (2). DnaA is a conserved bac-terial protein responsible for the initiation of DNA synthesis atthe chromosomal origin of replication (ori) (3, 4). The mecha-nism by which chromosome segregation is initiated in bacteria isless well understood.Although the factors responsible for DNA replication are highly
conserved among bacterial species, multiple mechanisms havebeen proposed to account for chromosome segregation (5). In theG1 phase of the Caulobacter crescentus cell cycle, the centromericregion of the chromosome (parS) is tethered to one pole of thecell (Fig. 1A). Upon the swarmer to stalked cell transition, repli-cation initiates with replisome assembly at the origin of replica-tion. The Par system in Caulobacter includes parS and twopartitioning proteins, ParA and ParB. The ParB protein binds toparS (6, 7), which, in turn, interacts with the nucleoid-associatedParA ATPase to effect centromere movement (8–11).In Vibrio cholerae and Bacillus subtilis, the chromosome parti-
tioning protein ParA (Soj) has been reported to regulate replica-tion initiation by directly interacting with the DnaA replicationinitiator protein, suggesting a connection between segregation andthe initiation of replication (12–14). However, the signals thattrigger the Par system to initiate chromosome segregation are notknown. By generating Caulobacter strains that express limitedconcentrations of DnaA, we sought to determine whether repli-cation initiation is a prerequisite for the translocation of the cen-tromere complex. Under these conditions, we were able to detecttranslocation of the chromosome in the absence of replication. Weshow that DnaA binds directly within the parS region and thataltering binding of DnaA to parS leads to compromised chromo-some segregation. These results suggest that, in Caulobacter, DnaAplays a direct role in the initiation of chromosome segregation.
ResultsDnaA-Dependent Chromosome Translocation. In Caulobacter, theParB partitioning protein directly occupies the parS site, found8 kb from ori (7). In cells expressing a functional CFP-ParB fu-sion protein, the number of fluorescent foci reflects the copy
number of ori (Fig. 1A). In a wild-type background, initiation ofDNA replication results in the migration of one copy of the CFP-ParB/parS complex toward the opposite end of the cell, culmi-nating in the establishment of a second fluorescent ParB focus atthe distal pole (7, 11) (Fig. 1 A and B). To investigate the de-pendency of chromosome segregation on DNA replication, weimaged the translocation of CFP-ParB/parS foci in cells expressingsubphysiological DnaA concentrations. We constructed aCaulobacterstrain in which transcription of the sole copy of dnaA was controlledby the tightly regulated, xylose-inducible PxylX promoter (15) and inwhich parB was replaced with a translational fusion of the geneencoding CFP to parB. When xylose was present in the growthmedia in this dnaA depletion strain, DnaA was expressed andfluorescent imaging revealed CFP-ParB localized to both cellpoles, indicating replication initiation had occurred (Fig. 1C).When these cells were shifted to media lacking xylose, a singleCFP-ParB focus remained at the stalked pole (Fig. 1D), consis-tent with the requirement of DnaA to initiate replication (16).To characterize the effect of limited DnaA levels on chro-
mosome segregation, we constructed a dnaA depletion strainin which dnaA expression was driven by the leaky, vanillate-inducible PvanA promoter. Growth of this strain in the absence ofvanillate led to an 80% reduction in relative levels of DnaA after1 h, whereas DnaA levels decreased below the detection thresh-old in a PxylX-dnaA strain within 30 min following removal ofinducer (Fig. 1G and Fig. S1). In the presence of vanillate(250 μM), the dynamic localization of CFP-ParB during the cellcycle was indistinguishable from that observed in a wild-typestrain (Fig. 1E). Growth in the absence of vanillate producedcells with a single CFP-ParB focus. Notably, in this background,we found that this single focus translocated to the opposite polein more than 60% of the cells (Fig. 1F).
Significance
DnaA is an essential and conserved bacterial protein that enablesthe initiation of DNA replication. Although it is commonly heldthat the onset of bacterial chromosome segregation dependson the initiation of DNA replication, we have found that inCaulobacter crescentus, chromosome segregation can be in-duced in a DnaA-dependent, yet replication-independent manner.The chromosome replication origin, containing essential DnaAbinding motifs, resides 8 kb from the centromere parS region thatalso contains DnaA binding motifs. The centromere parS regionbound to the ParB partition protein initiates movement across thecell followed by the origin region. Mutations in a centromereDnaA motif that alter DnaA–centromere interaction exhibit aber-rant patterns of ParB/parS translocation, implicating DnaA in theprocess of chromosome segregation.
Author contributions: P.E.M., V.S.K., and L.S. designed research; P.E.M. and V.S.K. per-formed research; P.E.M., V.S.K., and L.S. analyzed data; and P.E.M., V.S.K., and L.S. wrotethe paper.
Reviewers: J.W.G., University of California, Los Angeles; and M.K.W., Harvard MedicalSchool, Brigham and Women’s Hospital.
The authors declare no conflict of interest.1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1418989111/-/DCSupplemental.
16100–16105 | PNAS | November 11, 2014 | vol. 111 | no. 45 www.pnas.org/cgi/doi/10.1073/pnas.1418989111
Dow
nloa
ded
by g
uest
on
Aug
ust 5
, 202
0
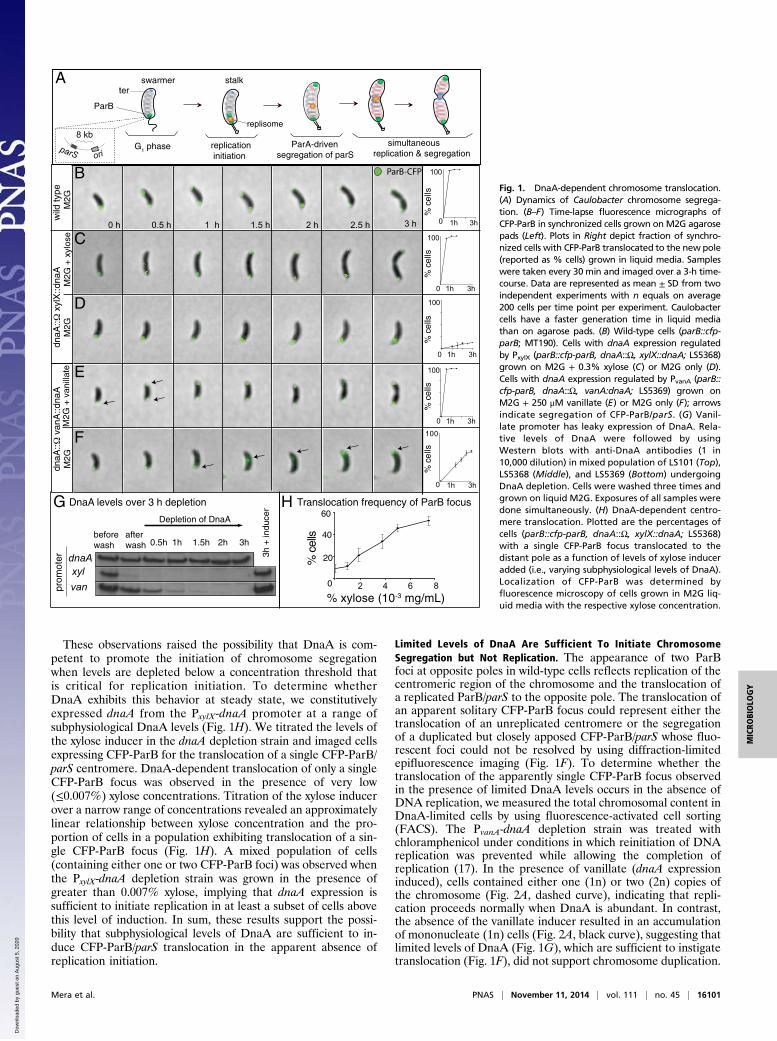
These observations raised the possibility that DnaA is com-petent to promote the initiation of chromosome segregationwhen levels are depleted below a concentration threshold thatis critical for replication initiation. To determine whetherDnaA exhibits this behavior at steady state, we constitutivelyexpressed dnaA from the PxylX-dnaA promoter at a range ofsubphysiological DnaA levels (Fig. 1H). We titrated the levels ofthe xylose inducer in the dnaA depletion strain and imaged cellsexpressing CFP-ParB for the translocation of a single CFP-ParB/parS centromere. DnaA-dependent translocation of only a singleCFP-ParB focus was observed in the presence of very low(≤0.007%) xylose concentrations. Titration of the xylose inducerover a narrow range of concentrations revealed an approximatelylinear relationship between xylose concentration and the pro-portion of cells in a population exhibiting translocation of a sin-gle CFP-ParB focus (Fig. 1H). A mixed population of cells(containing either one or two CFP-ParB foci) was observed whenthe PxylX-dnaA depletion strain was grown in the presence ofgreater than 0.007% xylose, implying that dnaA expression issufficient to initiate replication in at least a subset of cells abovethis level of induction. In sum, these results support the possi-bility that subphysiological levels of DnaA are sufficient to in-duce CFP-ParB/parS translocation in the apparent absence ofreplication initiation.
Limited Levels of DnaA Are Sufficient To Initiate ChromosomeSegregation but Not Replication. The appearance of two ParBfoci at opposite poles in wild-type cells reflects replication of thecentromeric region of the chromosome and the translocation ofa replicated ParB/parS to the opposite pole. The translocation ofan apparent solitary CFP-ParB focus could represent either thetranslocation of an unreplicated centromere or the segregationof a duplicated but closely apposed CFP-ParB/parS whose fluo-rescent foci could not be resolved by using diffraction-limitedepifluorescence imaging (Fig. 1F). To determine whether thetranslocation of the apparently single CFP-ParB focus observedin the presence of limited DnaA levels occurs in the absence ofDNA replication, we measured the total chromosomal content inDnaA-limited cells by using fluorescence-activated cell sorting(FACS). The PvanA-dnaA depletion strain was treated withchloramphenicol under conditions in which reinitiation of DNAreplication was prevented while allowing the completion ofreplication (17). In the presence of vanillate (dnaA expressioninduced), cells contained either one (1n) or two (2n) copies ofthe chromosome (Fig. 2A, dashed curve), indicating that repli-cation proceeds normally when DnaA is abundant. In contrast,the absence of the vanillate inducer resulted in an accumulationof mononucleate (1n) cells (Fig. 2A, black curve), suggesting thatlimited levels of DnaA (Fig. 1G), which are sufficient to instigatetranslocation (Fig. 1F), did not support chromosome duplication.
0 h 0.5 h 1 h 1.5 h 2 h 2.5 h 3 h
ParB-CFP
M2G
+ x
ylos
eM
2GM
2G +
van
illat
eM
2Gdn
aA::
xyl
X::d
naA
dnaA
:: v
anA
::dna
Aw
ild ty
pe
A
0
100
%ce
ll s
F
M2G
1h 3h0
100
dnaA
Depletion of DnaA
3h
+ in
duce
r
beforewash
afterwash 0.5h 1h 1.5h 2h 3h
G DnaA levels over 3 h depletion H Translocation frequency of ParB focus
% xylose (10-3 mg/mL)2 4 6 80
20
40
60
% c
ells
E
D
C
B
G1 phase replication initiation
ParA-drivensegregation of parS
simultaneousreplication & segregation
ter
ParB
replisome
swarmer stalk
oriparS
8 kb
%ce
ll s%
cell s
%ce
ll s%
c ells
1h 3h
0
100
1h 3h
0
100
1h 3h
0
100
1h 3h
xylvanpr
omot
er
Fig. 1. DnaA-dependent chromosome translocation.(A) Dynamics of Caulobacter chromosome segrega-tion. (B–F) Time-lapse fluorescence micrographs ofCFP-ParB in synchronized cells grown on M2G agarosepads (Left). Plots in Right depict fraction of synchro-nized cells with CFP-ParB translocated to the new pole(reported as % cells) grown in liquid media. Sampleswere taken every 30 min and imaged over a 3-h time-course. Data are represented as mean ± SD from twoindependent experiments with n equals on average200 cells per time point per experiment. Caulobactercells have a faster generation time in liquid mediathan on agarose pads. (B) Wild-type cells (parB::cfp-parB; MT190). Cells with dnaA expression regulatedby PxylX (parB::cfp-parB, dnaA::Ω, xylX::dnaA; LS5368)grown on M2G + 0.3% xylose (C) or M2G only (D).Cells with dnaA expression regulated by PvanA (parB::cfp-parB, dnaA::Ω, vanA:dnaA; LS5369) grown onM2G + 250 μM vanillate (E) or M2G only (F); arrowsindicate segregation of CFP-ParB/parS. (G) Vanil-late promoter has leaky expression of DnaA. Rela-tive levels of DnaA were followed by usingWestern blots with anti-DnaA antibodies (1 in10,000 dilution) in mixed population of LS101 (Top),LS5368 (Middle), and LS5369 (Bottom) undergoingDnaA depletion. Cells were washed three times andgrown on liquid M2G. Exposures of all samples weredone simultaneously. (H) DnaA-dependent centro-mere translocation. Plotted are the percentages ofcells (parB::cfp-parB, dnaA::Ω, xylX::dnaA; LS5368)with a single CFP-ParB focus translocated to thedistant pole as a function of levels of xylose induceradded (i.e., varying subphysiological levels of DnaA).Localization of CFP-ParB was determined byfluorescence microscopy of cells grown in M2G liq-uid media with the respective xylose concentration.
Mera et al. PNAS | November 11, 2014 | vol. 111 | no. 45 | 16101
MICRO
BIOLO
GY
Dow
nloa
ded
by g
uest
on
Aug
ust 5
, 202
0
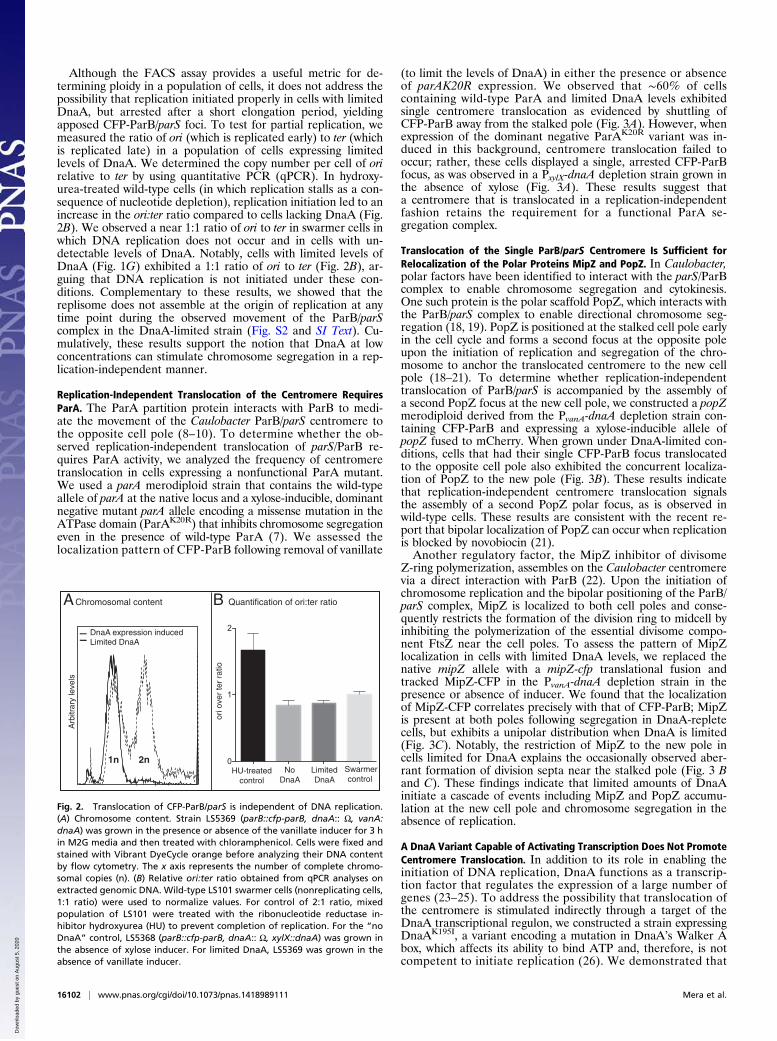
Although the FACS assay provides a useful metric for de-termining ploidy in a population of cells, it does not address thepossibility that replication initiated properly in cells with limitedDnaA, but arrested after a short elongation period, yieldingapposed CFP-ParB/parS foci. To test for partial replication, wemeasured the ratio of ori (which is replicated early) to ter (whichis replicated late) in a population of cells expressing limitedlevels of DnaA. We determined the copy number per cell of orirelative to ter by using quantitative PCR (qPCR). In hydroxy-urea-treated wild-type cells (in which replication stalls as a con-sequence of nucleotide depletion), replication initiation led to anincrease in the ori:ter ratio compared to cells lacking DnaA (Fig.2B). We observed a near 1:1 ratio of ori to ter in swarmer cells inwhich DNA replication does not occur and in cells with un-detectable levels of DnaA. Notably, cells with limited levels ofDnaA (Fig. 1G) exhibited a 1:1 ratio of ori to ter (Fig. 2B), ar-guing that DNA replication is not initiated under these con-ditions. Complementary to these results, we showed that thereplisome does not assemble at the origin of replication at anytime point during the observed movement of the ParB/parScomplex in the DnaA-limited strain (Fig. S2 and SI Text). Cu-mulatively, these results support the notion that DnaA at lowconcentrations can stimulate chromosome segregation in a rep-lication-independent manner.
Replication-Independent Translocation of the Centromere RequiresParA. The ParA partition protein interacts with ParB to medi-ate the movement of the Caulobacter ParB/parS centromere tothe opposite cell pole (8–10). To determine whether the ob-served replication-independent translocation of parS/ParB re-quires ParA activity, we analyzed the frequency of centromeretranslocation in cells expressing a nonfunctional ParA mutant.We used a parA merodiploid strain that contains the wild-typeallele of parA at the native locus and a xylose-inducible, dominantnegative mutant parA allele encoding a missense mutation in theATPase domain (ParAK20R) that inhibits chromosome segregationeven in the presence of wild-type ParA (7). We assessed thelocalization pattern of CFP-ParB following removal of vanillate
(to limit the levels of DnaA) in either the presence or absenceof parAK20R expression. We observed that ∼60% of cellscontaining wild-type ParA and limited DnaA levels exhibitedsingle centromere translocation as evidenced by shuttling ofCFP-ParB away from the stalked pole (Fig. 3A). However, whenexpression of the dominant negative ParAK20R variant was in-duced in this background, centromere translocation failed tooccur; rather, these cells displayed a single, arrested CFP-ParBfocus, as was observed in a PxylX-dnaA depletion strain grown inthe absence of xylose (Fig. 3A). These results suggest thata centromere that is translocated in a replication-independentfashion retains the requirement for a functional ParA se-gregation complex.
Translocation of the Single ParB/parS Centromere Is Sufficient forRelocalization of the Polar Proteins MipZ and PopZ. In Caulobacter,polar factors have been identified to interact with the parS/ParBcomplex to enable chromosome segregation and cytokinesis.One such protein is the polar scaffold PopZ, which interacts withthe ParB/parS complex to enable directional chromosome seg-regation (18, 19). PopZ is positioned at the stalked cell pole earlyin the cell cycle and forms a second focus at the opposite poleupon the initiation of replication and segregation of the chro-mosome to anchor the translocated centromere to the new cellpole (18–21). To determine whether replication-independenttranslocation of ParB/parS is accompanied by the assembly ofa second PopZ focus at the new cell pole, we constructed a popZmerodiploid derived from the PvanA-dnaA depletion strain con-taining CFP-ParB and expressing a xylose-inducible allele ofpopZ fused to mCherry. When grown under DnaA-limited con-ditions, cells that had their single CFP-ParB focus translocatedto the opposite cell pole also exhibited the concurrent localiza-tion of PopZ to the new pole (Fig. 3B). These results indicatethat replication-independent centromere translocation signalsthe assembly of a second PopZ polar focus, as is observed inwild-type cells. These results are consistent with the recent re-port that bipolar localization of PopZ can occur when replicationis blocked by novobiocin (21).Another regulatory factor, the MipZ inhibitor of divisome
Z-ring polymerization, assembles on the Caulobacter centromerevia a direct interaction with ParB (22). Upon the initiation ofchromosome replication and the bipolar positioning of the ParB/parS complex, MipZ is localized to both cell poles and conse-quently restricts the formation of the division ring to midcell byinhibiting the polymerization of the essential divisome compo-nent FtsZ near the cell poles. To assess the pattern of MipZlocalization in cells with limited DnaA levels, we replaced thenative mipZ allele with a mipZ-cfp translational fusion andtracked MipZ-CFP in the PvanA-dnaA depletion strain in thepresence or absence of inducer. We found that the localizationof MipZ-CFP correlates precisely with that of CFP-ParB; MipZis present at both poles following segregation in DnaA-repletecells, but exhibits a unipolar distribution when DnaA is limited(Fig. 3C). Notably, the restriction of MipZ to the new pole incells limited for DnaA explains the occasionally observed aber-rant formation of division septa near the stalked pole (Fig. 3 Band C). These findings indicate that limited amounts of DnaAinitiate a cascade of events including MipZ and PopZ accumu-lation at the new cell pole and chromosome segregation in theabsence of replication.
A DnaA Variant Capable of Activating Transcription Does Not PromoteCentromere Translocation. In addition to its role in enabling theinitiation of DNA replication, DnaA functions as a transcrip-tion factor that regulates the expression of a large number ofgenes (23–25). To address the possibility that translocation ofthe centromere is stimulated indirectly through a target of theDnaA transcriptional regulon, we constructed a strain expressingDnaAK195I, a variant encoding a mutation in DnaA’s Walker Abox, which affects its ability to bind ATP and, therefore, is notcompetent to initiate replication (26). We demonstrated that
Arb
itrar
y le
vels
1n 2n
DnaA expression inducedLimited DnaA
ori o
ver
ter
ratio
LimitedDnaA
NoDnaA
HU-treatedcontrol
Swarmercontrol
A Chromosomal content B Quantification of ori:ter ratio
0
1
2
Fig. 2. Translocation of CFP-ParB/parS is independent of DNA replication.(A) Chromosome content. Strain LS5369 (parB::cfp-parB, dnaA:: Ω, vanA:dnaA) was grown in the presence or absence of the vanillate inducer for 3 hin M2G media and then treated with chloramphenicol. Cells were fixed andstained with Vibrant DyeCycle orange before analyzing their DNA contentby flow cytometry. The x axis represents the number of complete chromo-somal copies (n). (B) Relative ori:ter ratio obtained from qPCR analyses onextracted genomic DNA. Wild-type LS101 swarmer cells (nonreplicating cells,1:1 ratio) were used to normalize values. For control of 2:1 ratio, mixedpopulation of LS101 were treated with the ribonucleotide reductase in-hibitor hydroxyurea (HU) to prevent completion of replication. For the “noDnaA” control, LS5368 (parB::cfp-parB, dnaA:: Ω, xylX::dnaA) was grown inthe absence of xylose inducer. For limited DnaA, LS5369 was grown in theabsence of vanillate inducer.
16102 | www.pnas.org/cgi/doi/10.1073/pnas.1418989111 Mera et al.
Dow
nloa
ded
by g
uest
on
Aug
ust 5
, 202
0
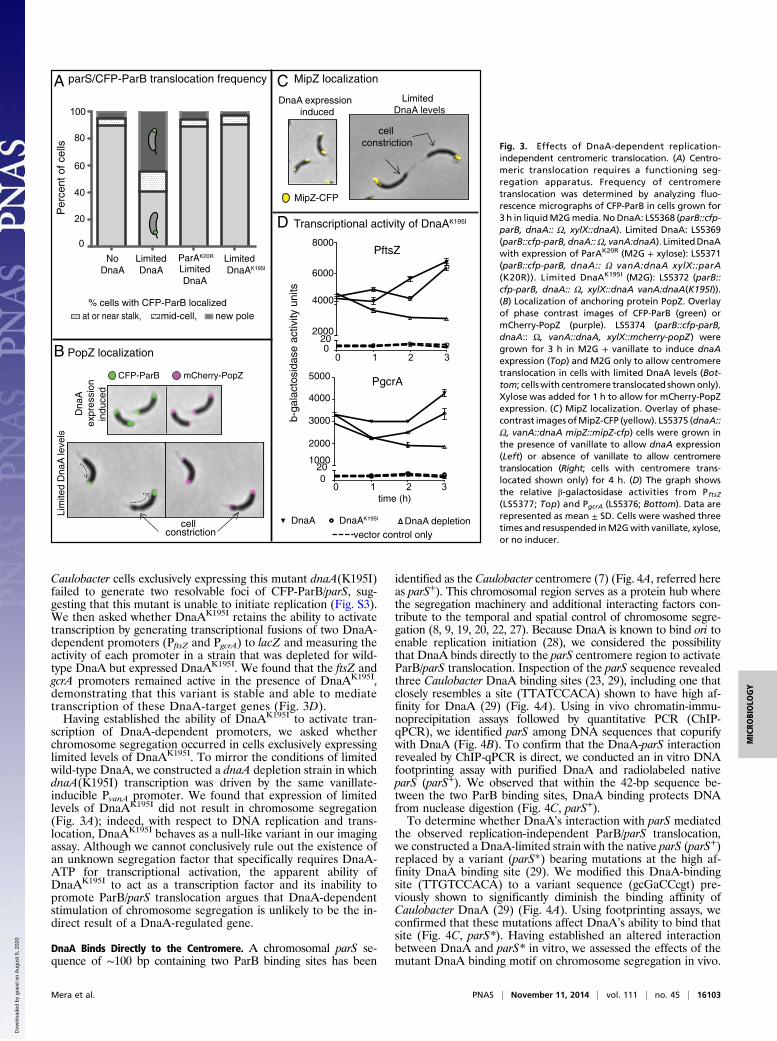
Caulobacter cells exclusively expressing this mutant dnaA(K195I)failed to generate two resolvable foci of CFP-ParB/parS, sug-gesting that this mutant is unable to initiate replication (Fig. S3).We then asked whether DnaAK195I retains the ability to activatetranscription by generating transcriptional fusions of two DnaA-dependent promoters (PftsZ and PgcrA) to lacZ and measuring theactivity of each promoter in a strain that was depleted for wild-type DnaA but expressed DnaAK195I. We found that the ftsZ andgcrA promoters remained active in the presence of DnaAK195I,demonstrating that this variant is stable and able to mediatetranscription of these DnaA-target genes (Fig. 3D).Having established the ability of DnaAK195I to activate tran-
scription of DnaA-dependent promoters, we asked whetherchromosome segregation occurred in cells exclusively expressinglimited levels of DnaAK195I. To mirror the conditions of limitedwild-type DnaA, we constructed a dnaA depletion strain in whichdnaA(K195I) transcription was driven by the same vanillate-inducible PvanA promoter. We found that expression of limitedlevels of DnaAK195I did not result in chromosome segregation(Fig. 3A); indeed, with respect to DNA replication and trans-location, DnaAK195I behaves as a null-like variant in our imagingassay. Although we cannot conclusively rule out the existence ofan unknown segregation factor that specifically requires DnaA-ATP for transcriptional activation, the apparent ability ofDnaAK195I to act as a transcription factor and its inability topromote ParB/parS translocation argues that DnaA-dependentstimulation of chromosome segregation is unlikely to be the in-direct result of a DnaA-regulated gene.
DnaA Binds Directly to the Centromere. A chromosomal parS se-quence of ∼100 bp containing two ParB binding sites has been
identified as the Caulobacter centromere (7) (Fig. 4A, referred hereas parS+). This chromosomal region serves as a protein hub wherethe segregation machinery and additional interacting factors con-tribute to the temporal and spatial control of chromosome segre-gation (8, 9, 19, 20, 22, 27). Because DnaA is known to bind ori toenable replication initiation (28), we considered the possibilitythat DnaA binds directly to the parS centromere region to activateParB/parS translocation. Inspection of the parS sequence revealedthree Caulobacter DnaA binding sites (23, 29), including one thatclosely resembles a site (TTATCCACA) shown to have high af-finity for DnaA (29) (Fig. 4A). Using in vivo chromatin-immu-noprecipitation assays followed by quantitative PCR (ChIP-qPCR), we identified parS among DNA sequences that copurifywith DnaA (Fig. 4B). To confirm that the DnaA-parS interactionrevealed by ChIP-qPCR is direct, we conducted an in vitro DNAfootprinting assay with purified DnaA and radiolabeled nativeparS (parS+). We observed that within the 42-bp sequence be-tween the two ParB binding sites, DnaA binding protects DNAfrom nuclease digestion (Fig. 4C, parS+).To determine whether DnaA’s interaction with parS mediated
the observed replication-independent ParB/parS translocation,we constructed a DnaA-limited strain with the native parS (parS+)replaced by a variant (parS*) bearing mutations at the high af-finity DnaA binding site (29). We modified this DnaA-bindingsite (TTGTCCACA) to a variant sequence (gcGaCCcgt) pre-viously shown to significantly diminish the binding affinity ofCaulobacter DnaA (29) (Fig. 4A). Using footprinting assays, weconfirmed that these mutations affect DnaA’s ability to bind thatsite (Fig. 4C, parS*). Having established an altered interactionbetween DnaA and parS* in vitro, we assessed the effects of themutant DnaA binding motif on chromosome segregation in vivo.
Per
cent
of c
ells
0
20
40
60
80
100
NoDnaA
LimitedDnaA
ParAK20R Limited DnaAK195I
CFP-ParB mCherry-PopZ
A parS/CFP-ParB translocation frequency
B PopZ localization
D Transcriptional activity of DnaAK195I
% cells with CFP-ParB localized new poleat or near stalk, mid-cell,
PftsZ
0 1 2
202000
4000
6000
8000
0
0 1 2 3
1000
2000
3000
4000
5000
020
time (h)
b-ga
lact
osid
ase
activ
ity u
nits
3
DnaA DnaAK195I DnaA depletion
Lim
ited
Dna
A le
vels
D
naA
expr
essi
on
ind
uced
vector control only
PgcrA
cell constriction
LimitedDnaA
C MipZ localization
MipZ-CFP
Limited DnaA levels
DnaA expression induced
cell constriction Fig. 3. Effects of DnaA-dependent replication-
independent centromeric translocation. (A) Centro-meric translocation requires a functioning seg-regation apparatus. Frequency of centromeretranslocation was determined by analyzing fluo-rescence micrographs of CFP-ParB in cells grown for3 h in liquidM2Gmedia. NoDnaA: LS5368 (parB::cfp-parB, dnaA:: Ω, xylX::dnaA). Limited DnaA: LS5369(parB::cfp-parB, dnaA::Ω, vanA:dnaA). LimitedDnaAwith expression of ParAK20R (M2G + xylose): LS5371(parB::cfp-parB, dnaA:: Ω vanA:dnaA xylX::parA(K20R)). Limited DnaAK195I (M2G): LS5372 (parB::cfp-parB, dnaA:: Ω, xylX::dnaA vanA:dnaA(K195I)).(B) Localization of anchoring protein PopZ. Overlayof phase contrast images of CFP-ParB (green) ormCherry-PopZ (purple). LS5374 (parB::cfp-parB,dnaA:: Ω, vanA::dnaA, xylX::mcherry-popZ) weregrown for 3 h in M2G + vanillate to induce dnaAexpression (Top) and M2G only to allow centromeretranslocation in cells with limited DnaA levels (Bot-tom; cellswith centromere translocated shownonly).Xylose was added for 1 h to allow for mCherry-PopZexpression. (C) MipZ localization. Overlay of phase-contrast images ofMipZ-CFP (yellow). LS5375 (dnaA::Ω, vanA::dnaA mipZ::mipZ-cfp) cells were grown inthe presence of vanillate to allow dnaA expression(Left) or absence of vanillate to allow centromeretranslocation (Right; cells with centromere trans-located shown only) for 4 h. (D) The graph showsthe relative β-galactosidase activities from PftsZ(LS5377; Top) and PgcrA (LS5376; Bottom). Data arerepresented as mean ± SD. Cells were washed threetimes and resuspended inM2Gwith vanillate, xylose,or no inducer.
Mera et al. PNAS | November 11, 2014 | vol. 111 | no. 45 | 16103
MICRO
BIOLO
GY
Dow
nloa
ded
by g
uest
on
Aug
ust 5
, 202
0
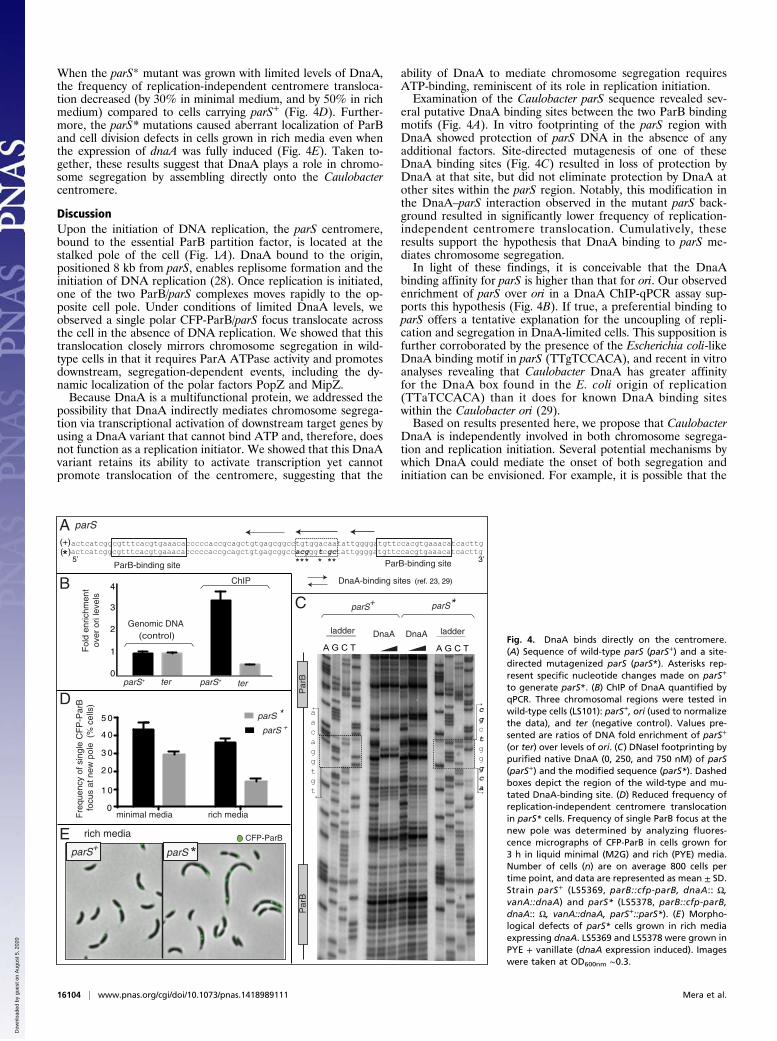
When the parS* mutant was grown with limited levels of DnaA,the frequency of replication-independent centromere transloca-tion decreased (by 30% in minimal medium, and by 50% in richmedium) compared to cells carrying parS+ (Fig. 4D). Further-more, the parS* mutations caused aberrant localization of ParBand cell division defects in cells grown in rich media even whenthe expression of dnaA was fully induced (Fig. 4E). Taken to-gether, these results suggest that DnaA plays a role in chromo-some segregation by assembling directly onto the Caulobactercentromere.
DiscussionUpon the initiation of DNA replication, the parS centromere,bound to the essential ParB partition factor, is located at thestalked pole of the cell (Fig. 1A). DnaA bound to the origin,positioned 8 kb from parS, enables replisome formation and theinitiation of DNA replication (28). Once replication is initiated,one of the two ParB/parS complexes moves rapidly to the op-posite cell pole. Under conditions of limited DnaA levels, weobserved a single polar CFP-ParB/parS focus translocate acrossthe cell in the absence of DNA replication. We showed that thistranslocation closely mirrors chromosome segregation in wild-type cells in that it requires ParA ATPase activity and promotesdownstream, segregation-dependent events, including the dy-namic localization of the polar factors PopZ and MipZ.Because DnaA is a multifunctional protein, we addressed the
possibility that DnaA indirectly mediates chromosome segrega-tion via transcriptional activation of downstream target genes byusing a DnaA variant that cannot bind ATP and, therefore, doesnot function as a replication initiator. We showed that this DnaAvariant retains its ability to activate transcription yet cannotpromote translocation of the centromere, suggesting that the
ability of DnaA to mediate chromosome segregation requiresATP-binding, reminiscent of its role in replication initiation.Examination of the Caulobacter parS sequence revealed sev-
eral putative DnaA binding sites between the two ParB bindingmotifs (Fig. 4A). In vitro footprinting of the parS region withDnaA showed protection of parS DNA in the absence of anyadditional factors. Site-directed mutagenesis of one of theseDnaA binding sites (Fig. 4C) resulted in loss of protection byDnaA at that site, but did not eliminate protection by DnaA atother sites within the parS region. Notably, this modification inthe DnaA–parS interaction observed in the mutant parS back-ground resulted in significantly lower frequency of replication-independent centromere translocation. Cumulatively, theseresults support the hypothesis that DnaA binding to parS me-diates chromosome segregation.In light of these findings, it is conceivable that the DnaA
binding affinity for parS is higher than that for ori. Our observedenrichment of parS over ori in a DnaA ChIP-qPCR assay sup-ports this hypothesis (Fig. 4B). If true, a preferential binding toparS offers a tentative explanation for the uncoupling of repli-cation and segregation in DnaA-limited cells. This supposition isfurther corroborated by the presence of the Escherichia coli-likeDnaA binding motif in parS (TTgTCCACA), and recent in vitroanalyses revealing that Caulobacter DnaA has greater affinityfor the DnaA box found in the E. coli origin of replication(TTaTCCACA) than it does for known DnaA binding siteswithin the Caulobacter ori (29).Based on results presented here, we propose that Caulobacter
DnaA is independently involved in both chromosome segrega-tion and replication initiation. Several potential mechanisms bywhich DnaA could mediate the onset of both segregation andinitiation can be envisioned. For example, it is possible that the
actcatcggcgtttcacgtgaaacacccccaccgcagctgtgagcggccacgggtcgctattggggatgttccacgtgaaacatcacttgactcatcggcgtttcacgtgaaacacccccaccgcagctgtgagcggcctgtggacaatattggggatgttccacgtgaaacatcacttg
** * ***
DnaA-binding sites
ParB-binding siteParB-binding site
(ref. 23, 29)
CFP-ParB
ter
Genomic DNA (control)
ChIP
Fol
d en
richm
ent
ove
r or
i lev
els
parS+ terparS+
4
2
1
3
0
0
1 0
2 0
3 0
4 0
5 0
minimal media rich media
parS +
Fre
quen
cy o
f sin
gle
CF
P-P
arB
focu
s at
new
pol
e (
% c
ells
)
parS *
CB
A parS
D
E rich media
A G C T
parS+ parS*
DnaA DnaAladder ladder
A G C T
cgctgggca
aacaggtgt
Par
BP
arB
3’5’
(+)
*( )
parS+ *parS
Fig. 4. DnaA binds directly on the centromere.(A) Sequence of wild-type parS (parS+) and a site-directed mutagenized parS (parS*). Asterisks rep-resent specific nucleotide changes made on parS+
to generate parS*. (B) ChIP of DnaA quantified byqPCR. Three chromosomal regions were tested inwild-type cells (LS101): parS+, ori (used to normalizethe data), and ter (negative control). Values pre-sented are ratios of DNA fold enrichment of parS+
(or ter) over levels of ori. (C) DNaseI footprinting bypurified native DnaA (0, 250, and 750 nM) of parS(parS+) and the modified sequence (parS*). Dashedboxes depict the region of the wild-type and mu-tated DnaA-binding site. (D) Reduced frequency ofreplication-independent centromere translocationin parS* cells. Frequency of single ParB focus at thenew pole was determined by analyzing fluores-cence micrographs of CFP-ParB in cells grown for3 h in liquid minimal (M2G) and rich (PYE) media.Number of cells (n) are on average 800 cells pertime point, and data are represented as mean ± SD.Strain parS+ (LS5369, parB::cfp-parB, dnaA:: Ω,vanA::dnaA) and parS* (LS5378, parB::cfp-parB,dnaA:: Ω, vanA::dnaA, parS+::parS*). (E) Morpho-logical defects of parS* cells grown in rich mediaexpressing dnaA. LS5369 and LS5378 were grown inPYE + vanillate (dnaA expression induced). Imageswere taken at OD600nm ∼0.3.
16104 | www.pnas.org/cgi/doi/10.1073/pnas.1418989111 Mera et al.
Dow
nloa
ded
by g
uest
on
Aug
ust 5
, 202
0

proximity between ori and parS in Caulobacter (∼8 kb; ref. 7)enables the assembly of DnaA into a polymeric structure thatlinks both chromosomal loci to integrate the processes of repli-cation and segregation. However, any model positing a DnaA-directed interaction between ori and parS must account for theobservation that parS can be artificially relocated hundreds ofkilobases away from ori and segregation is still maintained (7).Alternatively, it is conceivable that the DNA bending ability ofDnaA (30) is coopted to modify the structure of parS so as to in-fluence the structure and/or composition of the multicomponent,centromeric protein/DNA complex. A similar mechanism has beenproposed for the parS/ParAB partitioning system of P1 plasmid inE. coli in which the DNA-bending protein, IHF (integration hostfactor), binds between ParB-binding sites and bends P1 parS,resulting in a high affinity protein–DNA complex (31, 32).The results presented here apply to the replication and seg-
regation of the Caulobacter chromosome. However, the use ofreplication initiators to coordinate replication with segregationcould be a strategy used by other bacterial species. For example,in the opportunistic pathogen Burkholderia cenocepacia, DnaAbinding sites close to parS sites have been reported in at least twoof its four chromosomes (33). Although demonstration of DnaAbinding sites coincident with verified parS sites are requiredfor other bacterial species, a similar strategy to coordinate rep-lication with segregation has been observed with the eukaryoticreplication initiator complex (ORC). Independent of DNAreplication, components of ORC directly affect chromosomesegregation by properly localizing cohesion complexes of
sister chromatids and by regulating the formation of proteincomplexes required for microtubule organization (34). In fact,the human Orc2 protein localizes to the centrosome, centromeres,and heterochromatin, suggesting that replication initiator proteinsfunction to coordinate the entire chromosome inheritance cycle(35). These findings demonstrate that even when replication andsegregation are separated in time and space, cells maintain factorsthat coordinate these two events. These observations, along withthe results presented here, posit a conserved function of replica-tion initiators as key coordinators of DNA synthesis and chro-mosome segregation in both eukaryotes and bacteria.
MethodsAll Caulobacter strains used were derived from the wild-type strainCB15N (NA1000) and grown at 28 °C in either minimal or rich medium.Swarmer cells were isolated from mixed population cultures using percolldensity centrifugation protocol (36). Fluorescence microscopy followedby image processing with Adobe Photoshop and manual cell countingwas used to determine intracellular location of labeled loci. Detailsregarding experimental protocols, reagents, and data analyses areincluded in SI Methods.
ACKNOWLEDGMENTS. We thank Antonio Iniesta for the initial observationof mini-cells; Dante Ricci, W. Seth Childers, Jared Schrader, Tom Mann, andMichael Melfi for careful revisions of this manuscript; Shripa Patel forassistance with qPCR; and members of L.S. and McAdams laboratories formany helpful discussions. This work was supported by NIH Grants R01GM51426 (to L.S.) and F32 GM 097839 (to P.E.M.).
1. Nasmyth K (2002) Segregating sister genomes: The molecular biology of chromosomeseparation. Science 297(5581):559–565.
2. Reyes-Lamothe R, Nicolas E, Sherratt DJ (2012) Chromosome replication and segre-gation in bacteria. Annu Rev Genet 46:121–143.
3. Skarstad K, Katayama T (2013) Regulating DNA replication in bacteria. Cold SpringHarb Perspect Biol 5(4):a012922.
4. Kornberg A, Baker TA (1992) DNA Replication (Freeman, New York), 2nd Ed.5. Wang X, Montero Llopis P, Rudner DZ (2013) Organization and segregation of bac-
terial chromosomes. Nat Rev Genet 14(3):191–203.6. Mohl DA, Gober JW (1997) Cell cycle-dependent polar localization of chromosome
partitioning proteins in Caulobacter crescentus. Cell 88(5):675–684.7. Toro E, Hong SH, McAdams HH, Shapiro L (2008) Caulobacter requires a dedicated
mechanism to initiate chromosome segregation. Proc Natl Acad Sci USA 105(40):15435–15440.
8. Ptacin JL, et al. (2010) A spindle-like apparatus guides bacterial chromosome segre-gation. Nat Cell Biol 12(8):791–798.
9. Schofield WB, Lim HC, Jacobs-Wagner C (2010) Cell cycle coordination and regulationof bacterial chromosome segregation dynamics by polarly localized proteins. EMBO J29(18):3068–3081.
10. Shebelut CW, Guberman JM, van Teeffelen S, Yakhnina AA, Gitai Z (2010) Caulobacterchromosome segregation is an ordered multistep process. Proc Natl Acad Sci USA107(32):14194–14198.
11. Viollier PH, et al. (2004) Rapid and sequential movement of individual chromosomalloci to specific subcellular locations during bacterial DNA replication. Proc Natl AcadSci USA 101(25):9257–9262.
12. Kadoya R, Baek JH, Sarker A, Chattoraj DK (2011) Participation of chromosome seg-regation protein ParAI of Vibrio cholerae in chromosome replication. J Bacteriol193(7):1504–1514.
13. Scholefield G, Errington J, Murray H (2012) Soj/ParA stalls DNA replication by in-hibiting helix formation of the initiator protein DnaA. EMBO J 31(6):1542–1555.
14. Murray H, Errington J (2008) Dynamic control of the DNA replication initiation pro-tein DnaA by Soj/ParA. Cell 135(1):74–84.
15. Gorbatyuk B, Marczynski GT (2001) Physiological consequences of blocked Caulobactercrescentus dnaA expression, an essential DNA replication gene. Mol Microbiol 40(2):485–497.
16. Marczynski GT, Dingwall A, Shapiro L (1990) Plasmid and chromosomal DNA repli-cation and partitioning during the Caulobacter crescentus cell cycle. J Mol Biol 212(4):709–722.
17. Lesley JA, Shapiro L (2008) SpoT regulates DnaA stability and initiation of DNA rep-lication in carbon-starved Caulobacter crescentus. J Bacteriol 190(20):6867–6880.
18. Bowman GR, et al. (2008) A polymeric protein anchors the chromosomal origin/ParBcomplex at a bacterial cell pole. Cell 134(6):945–955.
19. Ebersbach G, Briegel A, Jensen GJ, Jacobs-Wagner C (2008) A self-associating proteincritical for chromosome attachment, division, and polar organization in caulobacter.Cell 134(6):956–968.
20. Bowman GR, et al. (2010) Caulobacter PopZ forms a polar subdomain dictating se-
quential changes in pole composition and function. Mol Microbiol 76(1):173–189.21. Laloux G, Jacobs-Wagner C (2013) Spatiotemporal control of PopZ localization
through cell cycle-coupled multimerization. J Cell Biol 201(6):827–841.22. Thanbichler M, Shapiro L (2006) MipZ, a spatial regulator coordinating chromosome
segregation with cell division in Caulobacter. Cell 126(1):147–162.23. Hottes AK, Shapiro L, McAdams HH (2005) DnaA coordinates replication initiation
and cell cycle transcription in Caulobacter crescentus. Mol Microbiol 58(5):1340–1353.24. Collier J, Murray SR, Shapiro L (2006) DnaA couples DNA replication and the ex-
pression of two cell cycle master regulators. EMBO J 25(2):346–356.25. Goranov AI, Katz L, Breier AM, Burge CB, Grossman AD (2005) A transcriptional re-
sponse to replication status mediated by the conserved bacterial replication protein
DnaA. Proc Natl Acad Sci USA 102(36):12932–12937.26. Mizushima T, et al. (1998) Site-directed mutational analysis for the ATP binding of
DnaA protein. Functions of two conserved amino acids (Lys-178 and Asp-235) located
in the ATP-binding domain of DnaA protein in vitro and in vivo. J Biol Chem 273(33):
20847–20851.27. Figge RM, Easter J, Gober JW (2003) Productive interaction between the chromosome
partitioning proteins, ParA and ParB, is required for the progression of the cell cycle
in Caulobacter crescentus. Mol Microbiol 47(5):1225–1237.28. Brassinga AK, Marczynski GT (2001) Replication intermediate analysis confirms that
chromosomal replication origin initiates from an unusual intergenic region in
Caulobacter crescentus. Nucleic Acids Res 29(21):4441–4451.29. Taylor JA, Ouimet MC, Wargachuk R, Marczynski GT (2011) The Caulobacter cres-
centus chromosome replication origin evolved two classes of weak DnaA binding
sites. Mol Microbiol 82(2):312–326.30. Schaper S, Messer W (1995) Interaction of the initiator protein DnaA of Escherichia
coli with its DNA target. J Biol Chem 270(29):17622–17626.31. Funnell BE (1988) Participation of Escherichia coli integration host factor in the P1
plasmid partition system. Proc Natl Acad Sci USA 85(18):6657–6661.32. Davis MA, Austin SJ (1988) Recognition of the P1 plasmid centromere analog involves
binding of the ParB protein and is modified by a specific host factor. EMBO J 7(6):
1881–1888.33. Dubarry N, Pasta F, Lane D (2006) ParABS systems of the four replicons of Burkholderia
cenocepacia: New chromosome centromeres confer partition specificity. J Bacteriol
188(4):1489–1496.34. Scholefield G, Veening JW, Murray H (2011) DnaA and ORC: More than DNA repli-
cation initiators. Trends Cell Biol 21(3):188–194.35. Prasanth SG, Prasanth KV, Siddiqui K, Spector DL, Stillman B (2004) Human Orc2 lo-
calizes to centrosomes, centromeres and heterochromatin during chromosome
inheritance. EMBO J 23(13):2651–2663.36. Ely B (1991) Genetics of Caulobacter crescentus. Methods Enzymol 204:372–384.
Mera et al. PNAS | November 11, 2014 | vol. 111 | no. 45 | 16105
MICRO
BIOLO
GY
Dow
nloa
ded
by g
uest
on
Aug
ust 5
, 202
0











![REPLICACIÓN [Modo de compatibilidad]sgpwe.izt.uam.mx/files/users/uami/pacopp/REPLICACION.pdf · •Doble cadena, complementarias y antiparalelas ... Facilitates DnaA act vity](https://img.pdfslide.us/doc/110x75/5b5be0467f8b9a2d458e81db/replicacion-modo-de-compatibilidadsgpweiztuammxfilesusersuamipacopp-.jpg)

















