Embed Size (px)
Citation preview

THE JOIXNAI. OF BIOI.OGICAL CHEMISTKY Vol. 251, No. 15. Issue of’ August 10. pp. 4544-4550. 1976
Prmted III U.S.A.
Regulation of the Transmembrane Potential of Isolated Chromaffin Granules by ATP, ATP Analogs, and External pH
(Received for publication, January 27, 1976)
HARVEY B. POLLARD, OREN ZINDER,* PHILIP G. HOFFMAN,* AND OLGA NIKODFJEVIC$
From the Reproduction Research Branch, National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, Maryland 20014
The transmembrane potential of isolated chromaffin granules has been measured using the permeant ions [‘%]methylamine and [?S]thiocyanate, as well as the fluorescent probe, 9-aminoacridine. At pH 7.0, the granule membrane had a Nernst proton potential of -45 mV, inside negative. This potential was sensitive to the external pH, but was unaffected by K’, Na’, CaZ+, Mg’+, or other cations. The pH of zero potential was 6.25 for both methylamine and thiocyanate. Thiocyanate also had a Nernst potential of similar magnitude and sign to that of methylamine at pH 7.0, and was also sensitive to variation in external pH.
MgZ-ATP was found to depolarize the granule membrane by a saturable mechanism with a K, for ATP of 40 /.LM. Ca2+ was only 30%) as effective as Mg *+ in supporting the ATP effect. The pH optimum for this
process was 6.25 and appeared to be accompanied by a marked alkalinization of the granule interior. The specificity for ATP was further tested with structural analogs of ATP and GTP.
The rate of change of membrane potential in response to changes in external pH or MgZ+ATP was estimated using the fluorescent probe 9-aminoacridine. Changes came to completion in less than 1 s. This
suggested that the ATP effects were not dependent on an enzymatic transformation but on an ATP-induced conformational change in the membrane.
We conclude that the chromaffin granule exists in at least two proton permeability states, corresponding to the presence or absence of Mg*+ATP. These states may be related to hormone release from granules and regulation of secretion in uiuo.
Chromaffin granules are 2000 A diameter secretory vesicles from the adrenal medulla which contain large amounts of epinephrine, ATP and specific proteins (1, 2). The isolated vesicles are highly stable in isotonic sucrose, but can release their total soluble contents in 10 min when exposed to MgZ+ATP and chloride ions at 37” and pH 6.2 (3). This ATP-stimulated release process has attracted attention as a model for reactions involved in storage and release of hormones or neurotransmitters in uiuo (l-5). Yet, the biochemical and biophysical basis for the effects of ATP on isolated granules remains unknown.
Chromaffin granule membranes have an ATPase activity which earlier workers have invoked as part of the mechanism
for ATP-stimulated release (4, 5). However, we have recently shown that App(NH)p,’ an ATP analog that is not cleaved by
* Present address, Laboratory of Clinical Biochemistry, Rambam Hospital and the Department of Biochemistry, Haifa Medical School, Haifa, Israel.
+ Present address, Department of Obstetrics and Gynecology, Uni- versity of California School of Medicine, San Francisco, California 94143.
0 Present address, School of Medicine, University of Skopje, Skopje, Yugoslavia.
‘The abbreviations used are: App(NH)p (Gpp(NH)p), adenosine (guanosine) 5’.(/3,y-imino)triphosphate; App(CH,)p, adenosine 5’.(8,.
typical 0-r ATPases (6), can substitute for ATP in the release reaction (3). By contrast, App(NH)p did not support the ATP-dependent catacholamine uptake reaction by granules, which occurred optimally at pH 7.4 (3). These results tended to exclude the ATPase from a direct role in ATP-stimulated release.
Another possible mechanism for ATP action was suggested by recent reports that granules might have a transmembrane potential that was sensitive to ATP. Bashford et al. (7) have
reported that MgZ+ATP enhanced ANS fluorescence in the presence of chromaffin granules or granule membranes. This change was blocked by mitochondrial uncouplers, and it was suggested that the granule membrane ATPase might be associated with a proton flux resulting in generation of both an electrical and chemical potential. We were therefore interested in whether granules had a true transmembrane potential, and whether it might be sensitive to external pH and ATP. It
seemed important to avoid reliance on fluorescent probe studies since these tended to yield only indirect information.
An alternative approach was to measure the transmembrane
y-methylene)triphosphate; ANS, 1-anilinonaphthalene-8-sulfonic acid; Hepes, N-2-hydroxyethylpiperazine-N-2.propanesulfonic acid; SCN, thiocyanate; g-AA, 9.aminoacridine; MEA, methylamine.
4544
by guest on March 18, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Chromaffin Granule Membrane Potential 4545
potential directly using equilibration of tracer permeant anions and cations such as methylamine, or thiocyanate. This method has been applied in the past to red cells (8-12), muscle (13, 14), mitochondria (15-21), chloroplasts (22-24) and microorga- nisms (25-27).
In this paper, we report that isolated chromaf’fin granules have a transmembrane potential that can be measured quanti- tatively with both radioactive methylamine and thiocyanate, as well as with the fluorescent probe, 9-aminoacridine. The potential is independent of exogenous energy sources and is sensitive to ATP as well as other purine trinucleotide analogs. We suggest that this membrane property may be relevant to ATP-stimulated release of catecholamines from isolated gran- ules.
EXPERIMENTAL PROCEDURES
Preparation of Chromaffin Granules-The granules were prepared essentially as described by Taugner (28). Fifteen to twenty pairs of bovine adrenal glands were obtained at the slaughter house and immediately chilled. Within 2 hours of slaughter, the adrenal medullas were dissected free of cortical material and placed in 10 volumes of unbuffered 0.3 M sucrose. The tissue was blended for 5 s in a Waring Blendor and then homogenized in a cylindrical glass homogenizer with a loose fitting motor-driven Teflon pestle. After filtering through four layers of gauze, the unbroken cells and nuclei were removed by centrifugation at 500 x g for 5 min. The supernatant was then centrifuged at 20,000 x g for 30 min. The resulting supernatant was decanted and the upper layer was removed by careful swirling with 5 ml of 0.3 M sucrose. The remainder of the pellet was resuspended in the same solution by hand homogenization in a loose fitting glass homoge- nizer. The resuspended material was centrifuged at 10,000 x g for 30 min and subsequently at 7,000 x g for 30 min with intervening decanting of the supernatant, swirling of the pellet, and resuspension by hand homogenization. The final suspension of the chromaffin granules had a protein concentration of approximately 5 mg/ml and contained 3.0 * 0.5 PM epinephrine/mg of protein.
Measurement of Total Granule Water-Chromaffin granules, equiv- alent to 10 to 15 mg of protein, were suspended in isotonic sucrose and mixed with [SH]dextran (nominal molecular weight = 20,000) as a marker for extra granular medium. The final specific activity was lo3 cpm/pl in a total volume of 1 ml. The granules were then placed in a preweighed glass test tube and centrifuged at 48,000 x g for 1 hour in a Sovall RC-2B centrifuge at 2’. The supernatant was removed and aliquots were taken for determination of specific activity of 13H]dex- tran, as well as density by refractometry. The pellet was weighed and then resuspended by homogenization in exactly 2 ml of distilled water. An aliquot was taken for measurement of [3H]dextran to determine the contribution to wet pellet weight by extragranular medium. The correct granule weight was calculated by subtracting the mass of extragranular medium from the total wet pellet weight. The granules were then lyophilized in the same preweighed glass test tube and the resulting dry product weighed again. The total water in the system was taken to be the difference in weights of the wet and dry pellets, corrected for sucrose left over from the trapped volume.
Measurement of Exchangeable Water-Intact chromaffin granules equivalent to 0.5 mg of protein were suspended and mixed with 3H,O and [“Cldextran (nominal molecular weight = 20,000) in a final volume of 1.00 ml. The specific activities were lo3 cpmlrl final volume for both isotopes. The samples were then centrifuged in a 12.ml plastic test tube at 20,000 x g for 30 min at 2”. The supernatant solutions were then removed with a Pasteur pipette and 50.~1 aliquots were taken for counting to determine the exact specific activity of each sample. Excess medium was removed by swabbing the inside of the tube with a laboratory tissue. The tightly packed granule pellet was then mixed with 0.5 ml of distilled water, frozen at -20” and thawed. This procedure facilitated dispersion of the pellet by agitation in a Vortex mixer to yield an homogeneous solution. A 300.~1 aliquot was taken for counting and 20 ~1 were taken for protein determination. The ex- changeable water volume, V,,,, was taken as the difference between the tritiated water volume and the [“Cldextran volume in the pellet.
Measurement of Thiocyanate and Methylamine Distribution- Chromaffin granules (0.3 to 0.6 mg of protein) were mixed with [3H]dextran (nominal molecular weight = 20,000) and lo-’ M concen-
tration of either [3SS]thiocyanate or [“C]methylamine. The medium also contained 20 rnM buffer (Tris-maleate or Tris-HCl) and was adjusted to an osmotic strength of 300 mosM with sucrose in a final volume of 1.0 ml. The activities of all isotopes were adjusted to 1000 cpm/wl in the final mixture. After 15 min at 2’, all samples were centrifuged at 48,000 x g for 30 min in a SS-34 rotor in a Sorvall RC-2B Superspeed centrifuge. The pellets and supernatants were then proc- essed as described for the determination of V,,, in the preceding section. In separate experiments on the same samples, the exchangea- ble water volume was also determined with [3H]Hz0 and [“Cldextran.
Uptake of Epinephrine by Granules-Chromaffin granules were incubated in isotonic medium containing 500 pg of granule protein, 1 rnM Mg(Ac),, 1 rnM ATP, 20 mM Tris-maleate buffer, pH 7.4 and 1 x 10m5 M [SH]epinephrine, all in a total volume of 1 ml. Osmotic strength was adjusted with sucrose. Samples were incubated for 10 min at 37”, mixed with 2 ml of ice-cold 0.3 M sucrose, and centrifuged at 20,000 x g for 30 min. The pellets were washed once in 12 ml of cold 0.3 M sucrose by decantation. The pellets were resuspended in 0.5 ml of distilled water and aliquots were taken for counting and protein determination. The supernatant was also assayed for epineph- rine to determine the exact specific activity.
Release of Epinephrine by Granules-Chromaffin granules were incubated in an isotonic medium containing 500 pg of granule protein, 1 rnM Mg(Ac),, 1 mM ATP, 50 rnM Hepes-Tris buffer, pH 6.30, and 90 rnM choline chloride in a total volume of 1 ml. Osmotic strength was adjusted with sucrose. Samples were incubated for 10 min at 37”, mixed with 2 ml of ice-cold 0.3 M sucrose and centrifuged at 20,000 x g for 30 min. Aliquots of supernatant solution were assayed for released, endogenous epinephrine. Release rates are expressed in terms of total original protein in the reaction mixture. Experiments were initiated within 15 min of final preparation step.
Fluorescence of9-Aminoacridine in Granules-Chromaffin granules in an isotonic solution were mixed with 0.1 PM 9-aminoacridine under various conditions in the presence and absence of 90 rnM choline chloride. These experiments were done at both 2” and 37”. The sample compartment was either cooled to 2” with circulating ethylene glycol or heated to 37” with water. The temperature in the cuvette was measured with a thermistor.
The reaction was initiated by placing 10 pg of chromaffin granule protein in a temperature-equilibrated solution containing 1 x lo-’ M 9-aminoacridine, 20 mM Tris-maleate or Hepes-Tris buffer, and 90 mM choline chloride where appropriate. The osmotic strength was adjusted to be isotonic with 0.3 M sucrose in all cases. The fluorescence was monitored at 460 nm with excitation at 395 nm in a quartz fluorescence cuvette (l-cm) using an Aminco-Bowman SPF fluorescence spectrome- ter. Measurements of fluorescence were obtained within 1 s of the addition of granules and were followed for up to 5 min subsequently.
The optical density at both 395 and 460 nm was determined and used to make corrections when necessary for self-absorbtion and scattering at the exciting and emitted wavelengths. The optical density at 395 nm was generally less than 0.05 A.
Chemical and Enzymatic Determinations-Protein was determined using the Lowry method (29). The samples were precipitated with 5% trichloroacetic acid prior to measurement, and bovine serum albumin was used as a standard.
Epinephrine was determined by the trihydroxy indole reaction (30). Purification on an alumina column was not necessary.
ATP was determined from the optical density at 260 nm. Sucrose concentration and medium density were determined by
refractometry using an Abbe refractometer. Density was determined by pycnometry of a 5.000.ml volume. Radioisotopes were obtained from New England Nuclear (Boston,
Mass.). Scintillation counting was done in a Beckman LS-355 scintilla- tion counter using Aquasol scintillation fluid.
Nucleotide analogs were obtained from either ICN (ADD(NH)D and . . . Gpp(NH)p) or Miles Laboratories and were used without further purification.
All other chemicals were of the highest quality commercially obtainable.
RESULTS
Total and Exchangeable Water-An accurate measure of the internal water volume of the granules was necessary in order to measure the internal concentration of permeant ions. Total water volume, VT,,, was 12.07 &mg of protein, or 83% of the
by guest on March 18, 2018
http://ww
w.jbc.org/
Dow
nloaded from
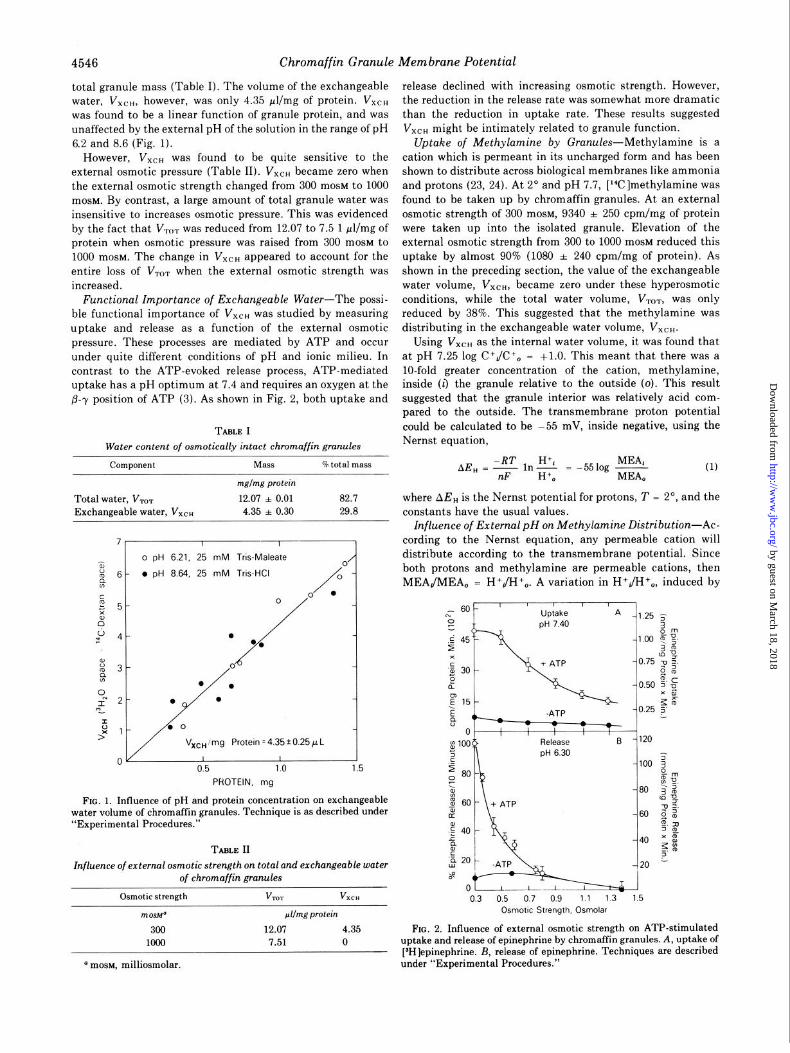
4546 Chromaffin Granule Membrane Potential
total granule mass (Table I). The volume of the exchangeable water, VX~ti, however, was only 4.35 &mg of protein. VXCH was found to be a linear function of granule protein, and was unaffected by the external pH of the solution in the range of pH 6.2 and 8.6 (Fig. 1).
release declined with increasing osmotic strength. However,
However, VXCH was found to be quite sensitive to the external osmotic pressure (Table II). VXCH became zero when the external osmotic strength changed from 300 mOSM to 1000 mosM. By contrast, a large amount of total granule water was insensitive to increases osmotic pressure. This was evidenced by the fact that V,,,, was reduced from 12.07 to 7.5 1 wl/mg of protein when osmotic pressure was raised from 300 mOSM to 1000 mosM. The change in VXCH appeared to account for the entire loss of V,,, when the external osmotic strength was increased.
Functional Importance of Exchangeable Water-The possi- ble functional importance of VXCH was studied by measuring uptake and release as a function of the external osmotic pressure. These processes are mediated by ATP and occur under quite different conditions of pH and ionic milieu. In contrast to the ATP-evoked release process, ATP-mediated uptake has a pH optimum at 7.4 and requires an oxygen at the B--y position of ATP (3). As shown in Fig. 2, both uptake and
TABLE I Water content of osmotically intact chromaffin granules
component
mglmg protein
Total water, V,, 12.07 * 0.01 82.7 where AE, is the Nernst potential for protons, T = 2’, and the Exchangeable water, VXCH 4.35 * 0.30 29.8 constants have the usual values.
7-’ 0 PH
l PH o pH 6.21, 25 mM Tris-Maleate 0’
6 - . pH 8.64, 25 0 mM Tris-HCI / -
0 l
5-
0
4- . I *‘/
00
/ . 0
Vxoimg Protein =4.35*0.25pL Vxoimg Protein = 4.35iO.25pL
I I I 0.5 0.5 1 .o 1 .o 1.5 1.5
PROTEIN, mg PROTEIN, mg
FIG. 1. Influence of pH and protein concentration on exchangeable water volume of chromaffin granules. Technique is as described under “Experimental Procedures.”
TABLE II
Influence of external osmotic strength on total and exchangeable water of chromaffin granules
Osmotic strength VTOT VXCH
mosbP &mg protein
300 12.07 4.35 1000 7.51 0
the reduction in the release rate was somewhat more dramatic than the reduction in uptake rate. These results suggested V,,, might be intimately related to granule function.
Uptake of Methylamine by Granules-Methylamine is a cation which is permeant in its uncharged form and has been shown to distribute across biological membranes like ammonia and protons (23, 24). At 2” and pH 7.7, [‘C]methylamine was found to be taken up by chromaffin granules. At an external osmotic strength of 300 mosM, 9340 f 250 cpm/mg of protein were taken up into the isolated granule. Elevation of the external osmotic strength from 300 to 1000 mOSM reduced this uptake by almost 90% (1080 f 240 cpm/mg of protein). As shown in the preceding section, the value of the exchangeable water volume, VXCH, became zero under these hyperosmotic conditions, while the total water volume, VTOT, was only reduced by 38%. This suggested that the methylamine was distributing in the exchangeable water volume, VXCH.
Using VXCH as the internal water volume, it was found that at pH 7.25 log C+JC +0 = +l.O. This meant that there was a lo-fold greater concentration of the cation, methylamine, inside (1) the granule relative to the outside (0). This result suggested that the granule interior was relatively acid com- pared to the outside. The transmembrane proton potential could be calculated to be -55 mV, inside negative, using the Nernst equation,
AE, = 9 MEA,
ln$ = -551og - 0 MEA,
(1)
Influence of External pH on Methylamine Distribution-Ac- cording to the Nernst equation, any permeable cation will distribute according to the transmembrane potential. Since both protons and methylamine are permeable cations, then MEAJMEA, = H+JI-I+,. A variation in H+JI-I+,, induced by
L- 601 ' ' ' IA Uptake
Release pH 6.30
-1.25 5 3
-0.50 0, 5’ u XB bF
-0.25 5 1
03 05 07 09 11 13 1.5 Osmotic Strength. Osmolar
FIG. 2. Influence of external osmotic strength on ATP-stimulated uptake and release of epinephrine by chromaffin granules. A, uptake of [SH]epinephrine. B, release of epinephrine. Techniques are described under “Experimental Procedures.” a mosM, milliosmolar.
by guest on March 18, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Chromaffin Granule Membrane Potential 4547
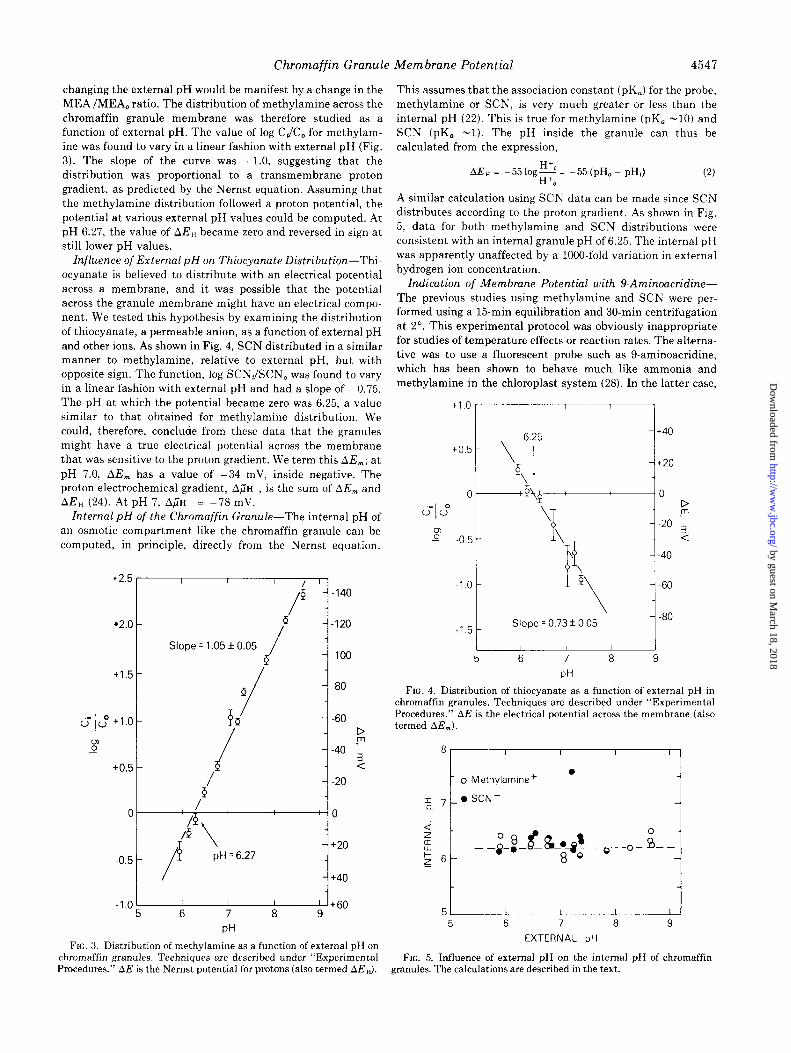
changing the external pH would be manifest by a change in the MEA/MEA, ratio. The distribution of methylamine across the chromaffin granule membrane was therefore studied as a function of external pH. The value of log C,/C, for methylam- ine was found to vary in a linear fashion with external pH (Fig. 3). The slope of the curve was +l.O, suggesting that the distribution was proportional to a transmembrane proton gradient, as predicted by the Nernst equation. Assuming that the methylamine distribution followed a proton potential, the potential at various external pH values could be computed. At pH 6.27, the value of AE, became zero and reversed in sign at still lower pH values.
Znfluence of External pH on Thiocyanate Distribution-Thi- ocyanate is believed to distribute with an electrical potential across a membrane, and it was possible that the potential across the granule membrane might have an electrical compo- nent. We tested this hypothesis by examining the distribution of thiocyanate, a permeable anion, as a function of external pH and other ions. As shown in Fig. 4, SCN distributed in a similar manner to methylamine, relative to external pH, but with opposite sign. The function, log SCNJSCN, was found to vary in a linear fashion with external pH and had a slope of -0.75. The pH at which the potential became zero was 6.25, a value similar to that obtained for methylamine distribution. We could, therefore, conclude from these data that the granules might have a true electrical potential across the membrane that was sensitive to the proton gradient. We term this AE,; at
pH 7.0, AE, has a value of -34 mV, inside negative. The proton electrochemical gradient, A~IH , is the sum of AE, and AE, (24). At pH 7, A;H = -78 mV.
Internal pH of the Chromaffin Granule-The internal pH of an osmotic compartment like the chromaffin granule can be computed, in principle, directly from the Nernst equation.
+2.5 r I 1 I / ‘1
I Slope = 1.05 f 0.05 / -100
+1.5
01: +l.O -
$
+0.5 -
j-80
1 -f -60
1 D .m
-40
! :
-j -20
-1.0 I+60 5 6 7 8 9
PH
FIG. 3. Distribution of methylamine as a function of external pH on chromaffin granules. Techniques are described under “Experimental
This assumes that the association constant (pK,) for the probe, methylamine or SCN, is very much greater or less than the internal pH (22). This is true for methylamine (pK, -10) and SCN (pK, -1). The pH inside the granule can thus be calculated from the expression,
AE, = -55 log: = -55 (pH, - pH,) 0
(2)
A similar calculation using SCN data can be made since SCN distributes according to the proton gradient. As shown in Fig. 5, data for both methylamine and SCN distributions were consistent with an internal granule pH of 6.25. The internal pH was apparently unaffected by a 1000.fold variation in external hydrogen ion concentration.
Indication of Membrane Potential with 9-Aminoacridine- The previous studies using methylamine and SCN were per- formed using a 15-min equilibration and 30-min centrifugation at 2”. This experimental protocol was obviously inappropriate for studies of temperature effects or reaction rates. The alterna- tive was to use a fluorescent probe such as 9-aminoacridine, which has been shown to behave much like ammonia and methylamine in the chloroplast system (28). In the latter case,
+lOr-----1 '
6.25
to.5 ‘I $1
~1.5 t
Slope = 0.73 f 0.05
+40
I
+20
0 D .m
-20
:
-40
-60
-80
- 5 9
PH
t
FIG. 4. Distribution of thiocvanate as a function of external DH in chromaffin granules. Techniques are described under “Experimental Procedures.” AE is the electrical potential across the membrane (also termed AE,)
*r--n
t o Methylamine + l
IaJ t 0 SCN-
5 6 7 8 9
EXTERNAL pH
FIG. 5. Influence of external pH on the internal pH of chromaffin Procedures.” APE is the Nernst potential for protons (also termed AE,). granules. The calculations are described in the text.
by guest on March 18, 2018
http://ww
w.jbc.org/
Dow
nloaded from
4548 Chromaffin Granule Membrane Potential
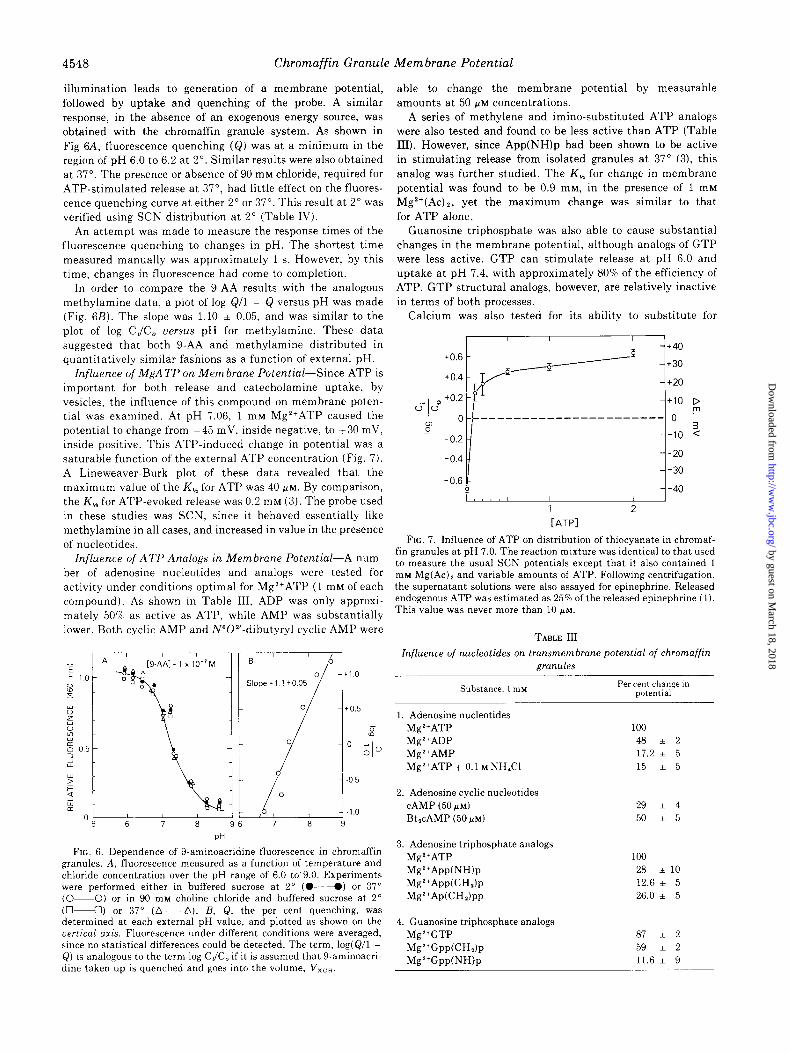
illumination leads to generation of a membrane potential, followed by uptake and quenching of the probe. A similar response, in the absence of an exogenous energy source, was obtained with the chromaffin granule system. As shown in
Fig 6A, fluorescence quenching (Q) was at a minimum in the region of pH 6.0 to 6.2 at 2”. Similar results were also obtained at 37”. The presence or absence of 90 mM chloride, required for ATP-stimulated release at 37”, had little effect on the fluores- cence quenching curve at either 2” or 37”. This result at 2” was verified using SCN distribution at 2” (Table IV).
An attempt was made to measure the response times of the fluorescence quenching to changes in pH. The shortest time measured manually was approximately 1 s. However, by this time, changes in fluorescence had come to completion.
In order to compare the g-AA results with the analogous methylamine data, a plot of log Q/l - Q versus pH was made (Fig. 6B). The slope was 1.10 * 0.05, and was similar to the
plot of log CJC, uersus pH for methylamine. These data suggested that both g-AA and methylamine distributed in quantitatively similar fashions as a function of external pH.
Influence of MgA TP on Membrane Potential-Since ATP is important for both release and catecholamine uptake, by vesicles, the influence of this compound on membrane poten- tial was examined. At pH 7.06, 1 mM Mg2+ATP caused the potential to change from -45 mV, inside negative, to +30 mV, inside positive. This ATP-induced change in potential was a saturable function of the external ATP concentration (Fig. 7). A Lineweaver-Burk plot of these data revealed that the maximum value of the K, for ATP was 40 PM. By comparison,
the K, for ATP-evoked release was 0.2 mM (3). The probe used in these studies was SCN, since it behaved essentially like methylamine in all cases, and increased in value in the presence of nucleotides.
Influence of ATP Analogs in Membrane Potential-A num- ber of adenosine nucleotides and analogs were tested for activity under conditions optimal for Mg*+ATP (1 mM of each compound). As shown in Table III, ADP was only approxi-
mately 50’s as active as ATP, while AMP was substantially lower. Both cyclic AMP and N”O*‘-dibutyryl cyclic AMP were
PH
FIG. 6. Dependence of 9-aminoacridine fluorescence in chromaffin granules. A, fluorescence measured as a function of temperature and chloride concentration over the pH range of 6.0 to’9.0. Experiments were performed either in buffered sucrose at 2” (04) or 37” (O-O) or in 90 rn~ choline chloride and buffered sucrose at 2” (04) or 37” (A-A). R, Q, the per cent quenching, was determined at each external pH value, and plotted as shown on the vertical axis. Fluorescence under different conditions were averaged, since no statistical differences could be detected. The term, log(Q/l - Q) is analogous to the term log C,/C, if it is assumed that 9-aminoacri- dine taken up is quenched and goes into the volume, V,,,.
able to change the membrane potential by measurable amounts at 50 WM concentrations.
A series of methylene and imino-substituted ATP analogs were also tested and found to be less active than ATP (Table III). However, since App(NH)p had been shown to be active in stimulating release from isolated granules at 37” (3), this analog was further studied. The K, for change in membrane
potential was found to be 0.9 mM, in the presence of 1 mM Mg’+(Ac)*, yet the maximum change was similar to that for ATP alone.
Guanosine triphosphate was also able to cause substantial changes in the membrane potential, although analogs of GTP were less active. GTP can stimulate release at pH 6.0 and uptake at pH 7.4, with approximately 80% of the efficiency of ATP. GTP structural analogs, however, are relatively inactive in terms of both processes.
Calcium was also tested for its ability to substitute for
c-i- u” $
to.6
+0.4
to.2
0
-0.2
1 2
[ATPI
FIG. 7. Influence of ATP on distribution of thiocyanate in chromaf- fin granules at pH 7.0. The reaction mixture was identical to that used to measure the usual SCN potentials except that it also contained 1 rn~ Mg(Ac), and variable amounts of ATP. Following centrifugation, the supernatant solutions were also assayed for epinephrine. Released endogenous ATP was estimated as 25% of the released epinephrine (1). This value was never more than 10 PM.
TABLE III
Influence of nucleotides on transmembrane potential of chromaffin gFWX&S
Substance, 1 mu Per cent change in potential
1. Adenosine nucleotides Mg*+ATP 100 Mg2+ADP 48 i 2 Mg*+AMP 17.2 i 5 Mg2+ATP + 0.1 M NH,Cl 15 * 5
2. Adenosine cyclic nucleotides CAMP (50 /I’M) BtlcAMP (50 /.LM)
29 * 4
50 * 5
3. Adenosine triphosphate analogs Mg2+ATP MgZ+App(NWp Mg2+App(CHh MgZ+Ap(CH,)pp
100 28 ilO 12.6 * 5 26.0 i 5
4. Guanosine triphosphate analogs Mg2+GTP Mg2+Gpp(CHh
Mg2+GppWWp
87 i 2
59 * 2
11.6 + 9
by guest on March 18, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Chromaffin Granule Membrane Potential 4549
magnesium in the ATP-stimulated potential change. Ca’+ATP had only about one-third the activity of Mg*+ATP. Various monovalent ions, at concentrations as high as 100 mM, had no
effect on the potential (Table IV). pH Optimum for ATP-induced Changes in Potential-The
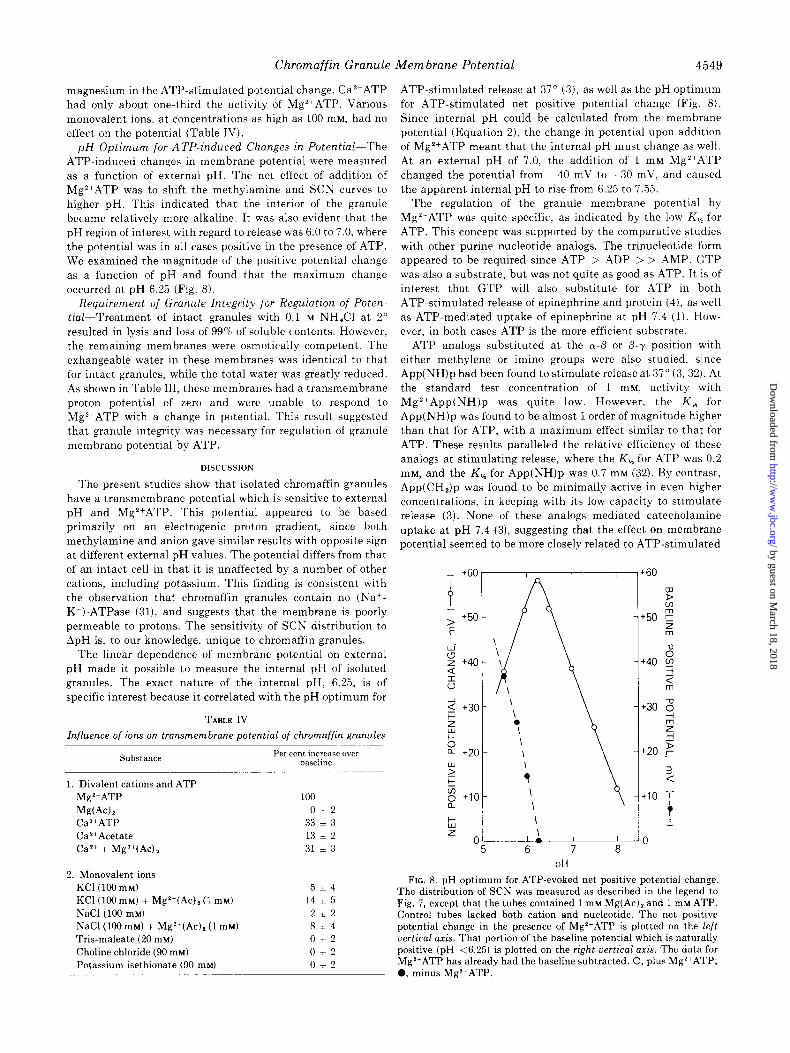
ATP-induced changes in membrane potential were measured as a function of external pH. The net effect of addition of
Mg*+ATP was to shift the methylamine and SCN curves to higher pH. This indicated that the interior of the granule became relatively more alkaline. It was also evident that the pH region of interest with regard to release was 6.0 to 7.0, where the potential was in all cases positive in the presence of ATP. We examined the magnitude of the positive potential change as a function of pH and found that the maximum change occurred at pH 6.25 (Fig. 8).
Requirement of Granule Integrity for Regulation of Poten- tial-Treatment of intact granules with 0.1 M NH,Cl at 2” resulted in lysis and loss of 99% of soluble contents. However, the remaining membranes were osmotically competent. The exhangeable water in these membranes was identical to that for intact granules, while the total water was greatly reduced. As shown in Table III, these membranes had a transmembrane proton potential of zero and were unable to respond to MgZ+ATP with a change in potential. This result suggested
that granule integrity was necessary for regulation of granule membrane potential by ATP.
DISCUSSION
The present studies show that isolated chromaffin granules have a transmembrane potential which is sensitive to external
pH and Mg’+ATP. This potential appeared to be based primarily on an electrogenic proton gradient, since both methylamine and anion gave similar results with opposite sign at different external pH values. The potential differs from that
of an intact cell in that it is unaffected by a number of other cations, including potassium. This finding is consistent with the observation that chromaffin granules contain no (Nat- K+)-ATPase (31), and suggests that the membrane is poorly permeable to protons. The sensitivity of SCN distribution to ApH is, to our knowledge, unique to chromaffin granules.
The linear dependence of membrane potential on external pH made it possible to measure the internal pH of isolated granules. The exact nature of the internal pH, 6.25, is of specific interest because it correlated with the pH optimum for
TABLE IV
Influence of ions on transmembrane potential of chromaffin granules
Substance Per cent increase 0~21
baseline
1. Divalent cations and ATP Mg*+ATP Mg(Ac)z CaZ+ATP Ca2+Acetate CaZ+ + MgZ+(Ac) Z
100
O&t2 33 l 3
13 l 2
31 * 3
2. Monovalent ions KC1 (100 mu) KC1 (100 mu) + MgZ+(Ac), (1 mu) NaCl (100 mu) NaCl(100 mM) + MgZ+(Ac), (1 mM)
Tris-maleate (20 mM) Choline chloride (90 mM) Potassium isethionate (90 mM)
5+4
14 * 5
212 8~4
0+2
012
0+2
ATP-stimulated release at 37” (3), as well as the pH optimum for ATP-stimulated net positive potential change (Fig. 8). Since internal pH could be calculated from the membrane
potential (Equation 2), the change in potential upon addition of Mg*+ATP meant that the internal pH must change as well. At an external pH of 7.0, the addition of 1 mM Mg’+ATP changed the potential from -40 mV to +30 mV, and caused the apparent internal pH to rise from 6.25 to 7.55.
The regulation of the granule membrane potential by Mg’+ATP was quite specific, as indicated by the low K, for ATP. This concept was supported by the comparative studies with other purine nucleotide analogs. The trinucleotide form appeared to be required since ATP > ADP > > AMP. GTP was also a substrate, but was not quite as good as ATP. It is of interest that GTP will also substitute for ATP in both
ATP-stimulated release of epinephrine and protein (4), as well as ATP-mediated uptake of epinephrine at pH 7.4 (1). How- ever, in both cases ATP is the more efficient substrate.
ATP analogs substituted at the a-P or P-y position with
either methylene or imino groups were also studied, since App(NH)p had been found to stimulate release at 37” (3, 32). At the standard test concentration of 1 mM, activity with MgZ+App(NH)p was quite low. However, the K, for App(NH)p was found to be almost 1 order of magnitude higher than that for ATP, with a maximum effect similar to that for ATP. These results paralleled the relative efficiency of these analogs at stimulating release, where the K, for ATP was 0.2 mM, and the K, for App(NH)p was 0.7 mM (32). By contrast, App(CH,)p was found to be minimally active in even higher concentrations, in keeping with its low capacity to stimulate release (3). None of these analogs mediated catecholamine uptake at pH 7.4 (3), suggesting that the effect on membrane
potential seemed to be more closely related to ATP-stimulated
Y F : z 0 +10 +lO a
7
t
L L z
0 0 5 6 7 8
PH
FIG. 8. pH optimum for ATP-evoked net positive potential change. The distribution of SCN was measured as described in the legend to Fig. 7, except that the tubes contained 1 mM Mg(Ac), and 1 mM ATP. Control tubes lacked both cation and nucleotide. The net positive potential change in the presence of Mg*+ATP is plotted on the left vertical axis. That portion of the baseline potential which is naturally positive (pH <6.25) is plotted on the right uertical axis. The data for Mgl+ATP has already had the baseline subtracted. 0, plus Mg*+ATP; 0, minus Mg’+ATP.
by guest on March 18, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4550 Chromaffin Granule Membrane Potential
release than to uptake.
These data also suggested that the change in membrane potential induced by ATP may not require cleavage of the substrate. Instead, nucleotide binding may be all that is required. Evidence for this concept includes the observation
that the process occurs at 2”, where enzymatic reactions are
slow, as well as the fact that changes in potential can be induced by stable analogs of ATP, such as App(NH)p. This concept is further substantiated by the quick response time of the transmembrane potential to changes in external pH using the fluorescent probe, 9-aminoacridine.
The effect of ATP on the transmembrane potential appeared to be quantitatively distinct from the ATP-induced enhance- ment of ANS fluorescence reported by Bashford et al. (7). For example, the ATP-induced ANS fluorescence changes occurred over a lo-min period, instead of in less than 1 s. The estimated internal pH using ANS was 5.5, while the actual internal pH, estimated with permeant ions, was 6.25. In addition, the ANS fluorescence effect appeared to be directly associated with ATPase, while the transmembrane potential could be changed
with App(NH)p, a nonhydrolyzable ATP analog. Finally, the ANS response was found in both intact and lysed granules, while both transmembrane potential and ATP response were abolished after lysis. However, since lysis, in our case, was accomplished by 100 mM NH,Cl, the loss of potential could have been due to uncoupling by ammonia. These results suggest that the two methods may yield important and comple- mentary information on the action of ATP on the isolated granule.
4n important question at this point is how granule function might relate both to membrane potential and to its modulation by Mg*+ATP. ATP-stimulated release occurred at 37” with a
relatively long time course and a K, of 0.2 mM. However, ATP-mediated membrane depolarization occurred at 2”, with a relatively short time course and a maximum K, of 0.04 mM.
These data are consistent with the possibility that the ATP- mediated changes in membrane potential may actually be preconditions for subsequent events such as release. Changes in potential induced by ATP may occur through regulation of proton flux through the granule membrane, and this regulation may be manifest through changes in conformation of mem-
brane proteins or other components. At the cellular level, the depolarized state may be a precondition for subsequent fusion of granule and plasma membranes and release by exocytosis.
Acknowledgment-The authors wish to thank Dr. H. R. Kaback for valuable discussions related to this work.
1.
2.
3.
4.
5.
6.
7.
8. 9.
10.
11. 12.
13.
14. 15. 16.
17.
18.
19.
20.
REFERENCES
Smith, A. D. (1968) in The Interactiorz of Drugs and Subcell&r Components in Animal Cells Campbell, P. N., ed) pp. 239-294, Churchill, London
Kirshner, N., and Kirshner, A. G. (1971) Phil. Trans. Roy. Sot. Ser. B 261, 279-289
Hoffman, P. G., Zinder, O., Nikodejevik, and Pollard, H. B. (1975) J. Suprumol. Strut. 4,181&184
Oka, M., Ohuchi, I., Yoshida, H., and Imaizumi, R. (1965) Biochim. Biophys. Acta 97,170m171
Poisner, A. M., and Trifaro, J. M. (1967) Mol. Pharmacol. 3, 561-571
Yount, R. G., Ojala, D., and Babcock, D. (1971) Biochemistry 10, 2490-2496
Bashford, C. L., Radda, G. K., and Ritchie, G. A. (1975) FEBS Lett. 50, 21-24
Warburg, E. J. (1922) Biochem J. 16, 153-340 VanSlyke, D. D., Hastings, A. B., Murray, E. D., and Sendroy, J..
Jr. (1925) J. Biol. Chem. 65, 701-728 Henderson, L. J., Bock, A. V., Field, H., and Stoddard, J. L. (1924)
J. Biol. Chem. 59, 379-431 Netter, H. (1934) Pflugers Arch. Ges. Physiol. 234, 680-695 Sims, P. J., Waggoner, A. S., Wang, C.-H., and Hoffman, J. F.
(1974) Biochemistry 13, 3315-3330 Fenn, W. O., Halge, L. F., Sheridan, E., and Flick, J. B. (1944) J.
Gen. Physiol. 28, 53-77 Hill, A. V. (1955) Proc. Roy. Sot. Ser. B. 144, l-22 Mitchell, P., and Moyle, J. (1969) Eur. J. Biochem. 7, 471-484 Liberman, E. A., Topaly, U. P., Tsofina, L. M., Iasaitis, A. A., and
Skulachev, V. P. (1969) Nature 222, 1076-1078 Cockrell, R. S., and Racker, E. (1969) Biochem. Biophys. Res.
Common. 35,414-419 Montal, M., Chance, B., and Lee, C. P. (1950) J. Membrane Biol.
2, 201-234 Papa, S., Guervieri, F., Bernardi, L. R., and Tager, J. M. (1970)
Biochim. Biophys. Acta 197, 100-103 Cockrell, R. S. (1972) Biochem. Biophys. Res. Commun. 46,
1991-1998 21. Addanki, S., Cahill, R. D., and Sotos, J. F. (1968) J. Biol. Chem.
243, 2337-2348 22. Schuldinger, S., Rottenberg, H., Avron, M. (1972) Eur. J.
Biochem. 25, W-70 23. Rottenberg, H., Grunwald, T., and Avron, M. (1972) Eur. J.
Biochem. 25, 54-64 24. Schuldinger, S., Padan, E., Rottenberg, H., Gromel-Elhanan, Z.,
and Avron, M. (1974) FEBS Lett. 49, 174-177 25. Irwin, M. (1925) J. Gen. Physiol. 9, 235-253 26. Osterhout, W. J. V. (1925) J. Gen. Physiol. 8, 131-146 27. Conway, E. J., and Downey, M. (1950) Biochem. J. 47, 355-360 28. Taugner, G. (1972) Biochem. J. 130, 969-973 29. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J.
(1951) J. Biol. Chem. 193, 265-275 30. Anton, A. H., and Sayers, D. F. (1962) J. Pharmacol. Exp. Ther.
138, 360-375 31. Trifaro, J. M., and Warner, M. (1972) Mol. Pharmacol. 8, 159-169 32. Hoffman, P. G., Zinder, O., Banner, W. M., and Pollard, H. B.
(1976) Arch. Biochem. Biophys. in press
by guest on March 18, 2018
http://ww
w.jbc.org/
Dow
nloaded from

H B Pollard, O Zinder, P G Hoffman and O NikodejevicATP, ATP analogs, and external pH.
Regulation of the transmembrane potential of isolated chromaffin granules by
1976, 251:4544-4550.J. Biol. Chem.
http://www.jbc.org/content/251/15/4544Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/251/15/4544.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on March 18, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























