Embed Size (px)
Citation preview

154
Regulation of the Clara Cell SecretoryProtein/Uteroglobin Promoter in Lung
MAGNUS NORD,a,b TOBIAS N. CASSEL,b HARALD BRAUN,c
AND GUNTRAM SUSKEc
bDepartment of Medical Nutrition, NOVUM, Karolinska Institute, Huddinge, SwedencInstitut für Molekularbiologie und Tumorforschung, Philipps-Universität Marburg, Marburg, Germany
ABSTRACT: Clara cell secretory protein/uteroglobin (CCSP/UG) is specificallyexpressed in the conducting airway epithelium of the lung in a differentiation-dependent manner. The proximal promoter region of the rodent CCSP/UGgene directs Clara cell specificity. Previously, it was shown that the forkheadtranscription factors HNF-3� and � and the homeodomain factor TTF-1 areimportant transcription factors acting through this region, suggesting thatthey contribute to cell specificity of the CCSP/UG gene. Members of the C/EBPfamily of transcription factors can also interact with elements of the proximalrat and mouse CCSP/UG promoters. The onset of C/EBP� expression in Claracells correlates with the strong increase of CCSP/UG expression. Thus, C/EBP�may play a crucial role for differentiation-dependent CCSP/UG expression.Transfection studies demonstrate that C/EBP� and TTF-1 can synergisticallyactivate the murine CCSP/UG promoter. Altogether, these results suggest thatC/EBP�, TTF-1, and HNF-3 determine the Clara cell–specific, differentiation-dependent expression of the CCSP/UG gene in murine lung. The relative im-portance of these three transcription factors, however, differs in rabbits andhumans.
INTRODUCTION
Regional differentiation of the lung epithelium results in the functionally distinctconducting and respiratory portions of the lung, with specialized cell types servingthe different subfunctions of the respiratory system.1 The bronchioles constitute themost distal part of the conducting airways. Here, the predominant epithelial cells areciliated cells and nonciliated Clara cells. Clara cells represent a well-differentiatedcell type with a high secretory activity. Numerous proteins are secreted from thesecells, with the major secretory product being the Clara cell secretory protein oruteroglobin (CCSP/UG).2 In normal lung, the CCSP/UG gene is specifically ex-pressed at high levels in the bronchiolar Clara cells, as well as in similar cells inbronchi and trachea.3,4 CCSP/UG has been estimated to account for 40% of proteinssecreted from rabbit Clara cells5 and constitutes 2–12% of human bronchoalveolarlavage fluid proteins.6,7
aAddress for correspondence: Magnus Nord, Department of Medical Nutrition, KarolinskaInstitute, NOVUM, Huddinge University Hospital, SE-141 86 Huddinge, Sweden. Voice: +46 85858 37 25; fax: +46 8 711 66 59.
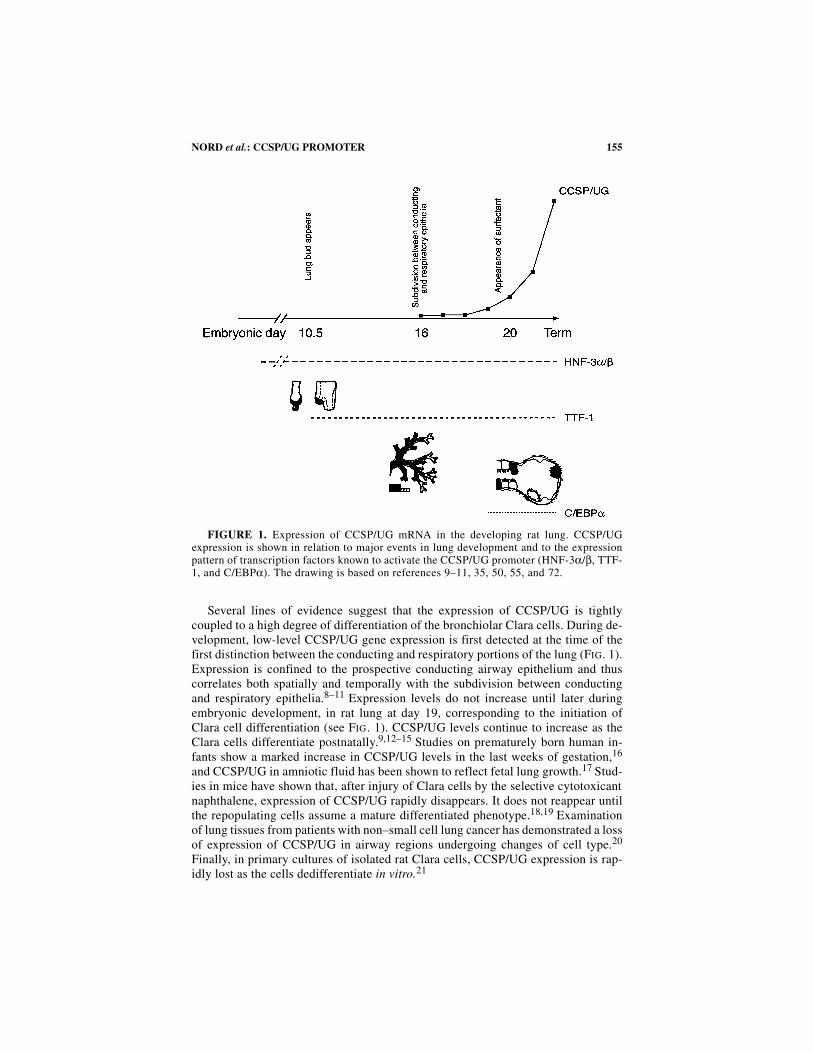
155NORD et al.: CCSP/UG PROMOTER
Several lines of evidence suggest that the expression of CCSP/UG is tightlycoupled to a high degree of differentiation of the bronchiolar Clara cells. During de-velopment, low-level CCSP/UG gene expression is first detected at the time of thefirst distinction between the conducting and respiratory portions of the lung (FIG. 1).Expression is confined to the prospective conducting airway epithelium and thuscorrelates both spatially and temporally with the subdivision between conductingand respiratory epithelia.8–11 Expression levels do not increase until later duringembryonic development, in rat lung at day 19, corresponding to the initiation ofClara cell differentiation (see FIG. 1). CCSP/UG levels continue to increase as theClara cells differentiate postnatally.9,12–15 Studies on prematurely born human in-fants show a marked increase in CCSP/UG levels in the last weeks of gestation,16
and CCSP/UG in amniotic fluid has been shown to reflect fetal lung growth.17 Stud-ies in mice have shown that, after injury of Clara cells by the selective cytotoxicantnaphthalene, expression of CCSP/UG rapidly disappears. It does not reappear untilthe repopulating cells assume a mature differentiated phenotype.18,19 Examinationof lung tissues from patients with non–small cell lung cancer has demonstrated a lossof expression of CCSP/UG in airway regions undergoing changes of cell type.20
Finally, in primary cultures of isolated rat Clara cells, CCSP/UG expression is rap-idly lost as the cells dedifferentiate in vitro.21
FIGURE 1. Expression of CCSP/UG mRNA in the developing rat lung. CCSP/UGexpression is shown in relation to major events in lung development and to the expressionpattern of transcription factors known to activate the CCSP/UG promoter (HNF-3α/β, TTF-1, and C/EBPα). The drawing is based on references 9–11, 35, 50, 55, and 72.

156 ANNALS NEW YORK ACADEMY OF SCIENCES
In this review, we will focus on transcription factors that determine the Claracell–specific, differentiation-dependent expression of CCSP/UG in lung. We will notdiscuss the influence of hormones and other signaling factors as this already hasbeen addressed in several previous reviews22,23 (also see articles in this volume).
REGULATION OF LUNG AND CLARA CELL–SPECIFICEXPRESSION OF CCSP/UG
Studies in transgenic mice have demonstrated that cis-acting elements directinglung tissue and Clara cell–specific expression of CCSP/UG reside within 2.3 kb ofthe 5′-flanking region of the rat gene.24,25 Experiments utilizing the Clara cell–likehuman lung adenocarcinoma cell line NCI-H441 as a model have demonstrated thatcis-acting elements within the first 175 bp of the rat CCSP/UG promoter can confercell specificity.24 An in vivo 5′-deletion analysis of the murine CCSP/UG gene intransgenic mice revealed that 166 bp of the mouse promoter is sufficient for directingClara cell–specific expression.26 Within this region of the rodent promoters, cell andlung specific interactions of nuclear proteins occur.24,27–29 Two binding sites forHNF-3/forkhead, or winged helix, transcription factors contribute to the activity ofthis promoter region in lung epithelial cells30,31 (see FIG. 2). In addition, bindingsites for the lung-enriched homeodomain transcription factor TTF-1 are presentwithin this region32–34 (FIG. 2). These studies suggest that factors from the HNF-3/forkhead transcription factor family, along with TTF-1, are crucial for the cell-specific expression of CCSP/UG in lung.
The HNF-3/forkhead transcription factor, hepatocyte nuclear factor-3α (HNF-3α), is first expressed in the murine developing endoderm from the mid- to late-primitive streak stage of gastrulation (around 8 days postcoitum). Subsequently, it isfound in all endoderm-derived structures, including lung35,36 (FIG. 1). In the adultlung, HNF-3α expression is confined to the conducting airway epithelium.28,37 Thissuggests that HNF-3α could be a determinant of bronchiolar cell-specific expressionof CCSP/UG. In accordance, HNF-3α activates the CCSP/UG promoter through twoHNF-3/forkhead sites in the proximal promoter27,28,30,31 (FIG. 2). Two additionalHNF-3 factors exist, HNF-3β and HNF-3γ, of which HNF-3β is also expressed inlung. HNF-3β expression is turned on already at the onset of gastrulation (FIG. 1).Subsequently, it is also expressed in all endoderm-derived structures.35,36 This haslead to the proposition that HNF-3α and β are involved in the specification of theendoderm. This hypothesis is supported by the results of HNF-3β gene targeting inmice. In HNF-3β(−/−) embryos, gut morphogenesis is severely affected, and orga-
FIGURE 2. Transcription factors acting through the proximal murine CCSP/UG pro-moter region. Positions of transcription factor binding sites in the murine CCSP/UG promot-er (numbering is based on reference 29).

157NORD et al.: CCSP/UG PROMOTER
nized node and notochord formation is absent.38,39 In adult lung, immunohistochem-ical studies have revealed expression of HNF-3β in bronchiolar cells, includingClara cells, as well as in alveolar type II cells.40 In situ hybridization studies forHNF-3β mRNA, however, have produced conflicting results regarding the expres-sion in alveolar and bronchiolar cells.28,37 Similar to HNF-3α, HNF-3β regulatesCCSP/UG expression through two HNF-3/forkhead sites in the proximal CCSP/UGpromoter.27,28,30,41
Several other lung-enriched HNF-3/forkhead transcription factors exist, most no-tably the HNF-3/forkhead homologues (HFH) 4 and 8. HFH-4 is expressed in thebronchiolar epithelium. Initial studies in liver cell lines showed that HFH-4 could ac-tivate the CCSP/UG promoter.42 However, more recent studies in other cell modelshave failed to demonstrate activation by HFH-4 (reference 41 and unpublishedobservations). During development, the onset of HFH-4 expression spatially andtemporally correlates with the distinction between the prospective conducting andrespiratory epithelia43 (FIG. 1). However, HFH-4 is expressed in ciliated cells only,and knockout and transgenic experiments have uncovered a key role for this factorin ciliogenesis.44–47 The human homologue of HFH-8, FREAC-1, activates theCCSP/UG promoter in lung epithelial cell lines.48 However, in adult mice, HFH-8expression is confined to the endothelium and connective fibroblasts of the alveolarregion.49 Thus, it seems unlikely that HFH-4 and -8 are involved in conferring Claracell specificity of the CCSP/UG gene. However, the possibility that another yet un-characterized lung or Clara cell–enriched HNF-3/forkhead transcription factor is ofimportance for the cell-specific expression of CCSP/UG still has to be considered.
The homeodomain transcription factor TTF-1 (thyroid transcription factor 1, alsoknown as NKx2.1 or T/EBP) is expressed in the prospective lung epithelium fromthe onset of lung development, that is, the appearance of the lung bud on the ventralforegut (at day 10.5 of rat development; see FIG. 1).50 As development proceeds, ex-pression of TTF-1 continues in the pulmonary epithelium. In the adult murine lung,both conducting and respiratory epithelial cells express TTF-1, excluding alveolartype I cells.40 The occurrence of TTF-1, however, has been described in type I cell–like cell lines.51 That TTF-1 plays a crucial role in lung development was demon-strated by gene targeting experiments. In TTF-1-deficient mice, the respiratory sys-tem is severely malformed, with virtually absent lungs.52,53 Together, this suggestsa key role for TTF-1 in specification of the lung epithelial cell lineage. The murineCCSP/UG promoter contains two TTF-1 binding sites within the 166-bp proximalpromoter region that confers Clara cell–specific expression (see FIG. 2). In addition,one major and two minor TTF-1 regulatory sites are present in upstream promoterregions. Transgenic studies suggest that the upstream sites are important for high-level CCSP/UG expression in vivo.26,32 In the rat CCSP/UG promoter, TTF-1 regu-latory sites have been identified at positions corresponding to the two proximal sitesand the major distal TTF-1 site.33,34
In summary, the data outlined above suggest that the transcription factors HNF-3 and TTF-1 are crucial for Clara cell–specific expression of the rodent CCSP/UGgenes. Both transcription factors are also important for the expression of other lung-specific genes, such as the surfactant apoproteins (recently reviewed in, e.g., refer-ence 54). Taken together with the results of gene targeting, this makes evident thatHNF-3 and TTF-1 play a key role for the specification of the pulmonary epithelium.

158 ANNALS NEW YORK ACADEMY OF SCIENCES
REGULATION OF DIFFERENTIATION-DEPENDENT EXPRESSIONOF CCSP/UG IN LUNG
It seems plausible that HNF-3 and TTF-1 regulate Clara cell–specific expressionof CCSP/UG. However, TTF-1 expression in the developing lung is first detectableat the time that the lung bud forms, and HNF-3α and β are expressed from earlierdevelopmental stages.35,36,50 This is in contrast to the onset of CCSP/UG expres-sion, which occurs later during development with levels increasing as Clara cellsstart to differentiate (FIG. 1). Certainly, other factors need to be involved in theregulation of CCSP/UG expression to account for its developmental timing.
The basic region–leucine zipper transcription factor C/EBPα is expressed in thedeveloping rat lung. Expression is first detected between day 18 and 20,55 in closecorrelation to the increase in CCSP/UG expression9,12 (see FIG. 1). In the adult ratand mouse lung, C/EBPα is expressed at high levels in alveolar type II cells and atlower levels in the bronchiolar Clara cells.21,55,56 As primary cultures of isolated ratClara cells are grown in vitro, CCSP/UG expression is rapidly lost as the cells de-differentiate. In contrast, HNF-3α and TTF-1 expression continues. C/EBPα levels,however, parallel the disappearance of CCSP/UG.21 Taken together, this suggeststhat C/EBPα could be involved in controlling the differentiation-dependent expres-sion of CCSP/UG. In this context, it should be noted that C/EBP factors, and espe-cially C/EBPα, play an important role in controlling differentiation anddifferentiation-dependent processes in other tissues. These tissues include liver(which, similar to lung, is an endodermal organ of foregut origin), adipose tissue,and white blood cells. In liver and fat, C/EBPα is an important regulator of prolifer-ation, cell cycle arrest, and gene expression (recently reviewed in references 57–61).
In accordance with a role for C/EBPα in the differentiation-dependent expressionof CCSP/UG, C/EBPα activates the rat CCSP/UG gene in transient transfections oflung cell lines. Activation occurs through a C/EBP response element in the proximalpromoter.21 In mice, expression of C/EBPδ, another C/EBP family member, is high-est in lung.62 Here, C/EBPδ is expressed at high levels in bronchiolar epithelial cells,including Clara cells, and at lower levels in alveolar epithelial cells.56 Studies in thedeveloping rabbit lung have revealed that, like C/EBPα, C/EBPδ also exhibits adifferentiation-dependent expression pattern.63 In transfections, the murine CCSP/UG promoter is activated by both C/EBPα and δ. Activation is mediated via a re-sponse element residing in the proximal CCSP/UG promoter. The C/EBP responseelement is located 60 to 100 bp upstream of the transcriptional start site and consistsof two C/EBP-binding sites (FIG. 2). The integrity of both sites is necessary for fullfunction of the element, indicating that the two sites form a compound response ele-ment.56 C/EBPδ is a more potent activator of the CCSP/UG promoter than C/EBPα.In accordance with this, C/EBPδ strongly interacts with both binding sites in thecompound response element, whereas C/EBPα interaction with the proximal site isweaker. On both C/EBP-binding sites, heterodimers between C/EBPα and δ arepreferentially formed. Cotransfection of C/EBPα and C/EBPδ together resulted in amore than additive induction as compared to the factors alone. This indicates aregulatory role for the C/EBPα–C/EBPδ heterodimers.56
The finding that C/EBPα and δ can activate the CCSP/UG promoter as well asthe correlation between CCSP/UG expression and C/EBP factors during differenti-

159NORD et al.: CCSP/UG PROMOTER
ation suggest that these C/EBP factors play an important role in differentiation-dependent processes in the bronchiolar epithelium of the lung. Such a hypothesis isfurther supported by findings showing that another Clara cell differentiation marker,the P450-enzyme CYP2B1, is also regulated by C/EBPα and δ in lung epithelialcells.64 A more general role for C/EBPα in regulating pulmonary epithelial differen-tiation is suggested by histological examinations of lungs from C/EBPα(−/−) knock-out mice. C/EBPα-deficient lungs demonstrate alveolar abnormalities withhyperproliferation of epithelial cells.65 No abnormalities in the bronchiolar epitheli-um were reported in these mice. A compensation for C/EBPα deficiency by C/EBPδmay occur in bronchioles of C/EBPα knockout mice since expression of C/EBPδ ishigh in this part of the lung. In the alveolar epithelium, where C/EBPδ expression islower, such compensation may not be possible.56 That such compensations canoccur has been demonstrated in the liver of C/EBPα(−/−) mice, where C/EBPβ cansubstitute for the loss of C/EBPα.66
DIFFERENCES IN PULMONARY REGULATION OF THE RODENT,RABBIT, AND HUMAN CCSP/UG GENES
It is clear that TTF-1, HNF-3, and C/EBP are important transcription factors thatdetermine expression of the CCSP/UG gene in rodent lung. However, whether thesetranscription factors are also major players in other mammals seems questionable.The role of TTF-1 for expression of the CCSP/UG gene in rabbit and human lung isespecially unclear. Studies of the rabbit CCSP/UG gene in transgenic mice show that600 bp of the 5′-flanking sequence is sufficient to confer lung expression. Similar towhat has been described for the rodent genes, additional sequences between 0.6 and2.3 kb are necessary for full high-level pulmonary expression.67 Transfection studiesusing the Clara cell–like NCI-H441 cell line as a model have demonstrated six re-gions containing cis-acting elements of importance for expression within the first250 bp of the rabbit CCSP/UG promoter.68 Two of these elements are regulated bythe ubiquitous transcription factors, Sp1 and Sp3,69 and two are HNF-3/forkheadregulatory sites.41 In contrast to these HNF-3/forkhead-binding sites and the distalC/EBP-binding site, the TTF-1 sites in the rat and mouse CCSP/UG promoters arenot conserved in the corresponding rabbit and human promoter regions. The rabbitCCSP/UG promoter contains recognition sites for TTF-1 at different positions thatbind TTF-1 in vitro (unpublished data). In transfection experiments, however,neither the rabbit nor the human CCSP/UG promoters are activated by TTF-1.41
Moreover, in immunohistochemical studies of human term lung, TTF-1 was detectedprimarily in alveolar type II cells and was only rarely seen in distal conducting air-way cells.70
Both HNF-3α and HNF-3β can activate not only the rodent, but also the rabbitand human CCSP/UG promoters. However, binding of HNF-3α and β on their ownis not sufficient for activation of these promoters. The action of both transcriptionfactors is strongly dependent on the presence of the ubiquitous transcription factorsSp1 or Sp3.41 Whether a similar combinatorial action of Sp1/Sp3 transcription fac-tors and HNF-3α and HNF-3β occurs in rat and mouse is unclear. The rat gene con-tains Sp1/Sp3-binding sites in the proximal promoter region. However, in

160 ANNALS NEW YORK ACADEMY OF SCIENCES
transfection experiments, Sp1/Sp3 factors do not seem to activate these promoters(reference 33 and unpublished observations). In summary, it appears that the mech-anisms that determine Clara cell–specific expression of the CCSP/UG gene in lungare similar, but not identical in rodents, rabbits, and humans.
COOPERATIVE INTERACTIONS IN THE REGULATIONOF THE CCSP/UG PROMOTER
As discussed above, HNF-3, TTF-1, and C/EBP control the Clara cell–specific,differentiation-dependent expression of the rodent CCSP/UG genes through interac-tions with promoter proximal elements (FIG. 2). It seemed as an attractive hypothesisthat these factors synergistically act together to produce the high-level CCSP/UG ex-pression characteristic for the differentiated Clara cell. To investigate this possibility,we performed cotransfection studies in Drosophila Schneider SL-2 cells. SL-2 cellslack many mammalian transcription factors and are thus well suited for cooperativitystudies.71 Transient transfection of a 172-bp murine CCSP/UG promoter fragmentalong with expression plasmids for HNF-3α, HNF-3β, TTF-1, and C/EBPα activat-ed the CCSP/UG promoter by 1.5-, 1.9-, 3.7-, and 1.6-fold, respectively (see FIG. 3).When TTF-1 and C/EBPα expression plasmids were transfected together, a strongsynergy resulting in a 40-fold induction of the reporter gene was seen (FIG. 3). Nosynergism was observed between C/EBPα and HNF-3 nor between HNF-3 and TTF-1. Synergistic activation by C/EBPα and TTF-1 was further increased by HNF-3αor β (FIG. 3). No synergism was observed between TTF-1 and C/EBPδ (unpublishedresults).
FIGURE 3. Activation of the CCSP/UG promoter by various transcription factors.Drosophila SL-2 cells were transfected with a 172-bp murine CCSP/UG-promoter-reportergene construct (4 µg) along with expression plasmids for TTF-1 (2 µg), C/EBPα (2 µg),HNF-3α (20 ng), and HNF-3β (20 ng). The amount of DNA was kept constant by inclusionof empty pPac expression plasmid. Cells also received a β-galactosidase expression plasmid(2 µg) to allow normalization for variations in transfection efficiency. Assays were carriedout in duplicates. Cell culture methods, transfections, and reporter gene assays have beendescribed previously.71

161NORD et al.: CCSP/UG PROMOTER
Additional TTF-1 binding sites exist upstream of the 172-bp CCSP/UG promoterfragment.32 When a 2.1-kb fragment containing these upstream TTF-1 sites wasused instead, stronger activation by TTF-1 was observed. Synergistic activation byTTF-1 and C/EBPα, however, did not increase. This result indicates that the coop-erative action of TTF-1 and C/EBPα is mediated via TTF-1-binding sites within the172 most-proximal base pairs of the promoter (unpublished results). At this stage,the molecular mechanisms behind the synergistic activity of C/EBPα and TTF-1 andthe additional effects of HNF-3 are unclear. Several possibilities exist, including co-operative binding of C/EBPα and TTF-1 mediated by protein-protein interactionsand/or changes of the local DNA conformation. Another possibility would be coop-erative recruitment of coactivators. In summary, these results show that C/EBPα actsin concert with TTF-1 and HNF-3 to regulate CCSP/UG gene expression. Our datasuggest that C/EBPα expression is essential for high-level expression of CCSP/UGin differentiated Clara cells and that the onset of C/EBPα expression in the develop-ing lung is crucial for the increase in CCSP/UG expression (FIG. 1). These findingsalso support a role for C/EBP factors in the control of lung epithelial differentiation.
CONCLUSIONS
In lung, the Clara cell secretory protein/uteroglobin is specifically expressed inClara cells of the conducting airway epithelium in a differentiation-dependent man-ner. Studies during the last decade have advanced our understanding of the regula-tion of this gene and also started to uncover the molecular determinants of cellspecification and differentiation in the developing lung. Collectively, these studiessupport a hypothesis for the regulation of the rodent CCSP/UG genes in which TTF-1 and HNF-3 act in concert with C/EBPα to give cell-specific and differentiation-dependent expression, respectively.
ACKNOWLEDGMENTS
This work was supported by the Swedish Medical Research Council (Grant No.13115); the Swedish Heart-Lung Foundation (Grant No. 199941330); SwedishMatch (Grant No. 199914); the Swedish Medical Society; the Research Foundations“Tore Nilssons stiftelse för medicinsk forskning”, “Stiftelsen Lars Hiertas minne”,“Sigurd och Elsa Goljes minne”, “Magnus Bergvalls stiftelse”, and “Robert Lund-bergs stiftelse”; the Research Foundations of the Karolinska Institute (M. Nord andT. N. Cassel); and the BMBF and Deutsche Forschungsgemeinschaft (G. Suske).
REFERENCES
1. GAIL, D.B. & C.J.M. LENFANT. 1983. Cells of the lung: biology and clinical applica-tions. Am. Rev. Respir. Dis. 127: 366–387.
2. MASSARO, G.D., G. SINGH, R. MASON et al. 1994. Biology of the Clara cell: conferencereport. Am. J. Physiol. 266: L101–L106.
3. BEDETTI, C.D., J. SINGH, G. SINGH et al. 1987. Ultrastructural localization of rat Claracell 10 kD secretory protein by the immunogold technique using polyclonal andmonoclonal antibodies. J. Histochem. Cytochem. 35: 789–794.

162 ANNALS NEW YORK ACADEMY OF SCIENCES
4. LUND, J., T. DEVEREUX, H. GLAUMANN & J-Å. GUSTAFSSON. 1988. Cellular and sub-cellular localization of a binding protein for polychlorinated biphenyls in rat lung.Drug Metab. Dispos. 16: 590–599.
5. PATTON, S.E., L.B. GILMORE, A.M. JETTEN et al. 1986. Biosynthesis and release of pro-teins by isolated pulmonary Clara cells. Exp. Lung Res. 11: 277–294.
6. BERNARD, A., H. ROELS, R. LAUWERYS et al. 1992. Human urinary protein 1: evidencefor identity with the Clara cell protein and occurrence in respiratory tract and uro-genital secretions. Clin. Chim. Acta 207: 239–249.
7. ANDERSSON, O., T.N. CASSEL, R. GRONNEBERG et al. 1999. In vivo modulation ofglucocorticoid receptor mRNA by inhaled fluticasone propionate in bronchialmucosa and blood lymphocytes in subjects with mild asthma. J. Allergy Clin. Immu-nol. 103: 595–600.
8. STRUM, J.M., R.S. COMPTON, S.L. KATYAL & G. SINGH. 1992. The regulated expressionof mRNA for Clara cell protein in the developing airways of the rat, as revealed bytissue in situ hybridization. Tissue Cell 24: 461–471.
9. NORD, M., O. ANDERSSON, M. BRÖNNEGÅRD & J. LUND. 1992. Rat lung polychlorinatedbiphenyl–binding protein: effect of glucocorticoids on the expression of the Clara cell–specific protein during fetal development. Arch. Biochem. Biophys. 296: 302–307.
10. TEN HAVE–OPBROEK, A.A. 1981. The development of the lung in mammals: an analy-sis of concepts and findings. Am. J. Anat. 162: 201–219.
11. OTTO-VERBERNE, C.J. & A.A. TEN HAVE–OPBROEK. 1987. Development of the pulmo-nary acinus in fetal rat lung: a study based on an antiserum recognizing surfactant-associated proteins. Anat. Embryol. 175: 365–373.
12. SINGH, G., S.L. KATYAL & C.M. WONG. 1986. A quantitative assay for a Clara cell–specific protein and its application in the study of development of pulmonary airwaysin the rat. Pediatr. Res. 20: 802–805.
13. CARDOSO, W.V., L.G. STEWART, K.E. PINKERTON et al. 1993. Secretory product expressionduring Clara cell differentiation in the rabbit and rat. Am. J. Physiol. 264: L543–L552.
14. MASSARO, G.D., L. DAVIS & D. MASSARO. 1984. Postnatal development of the bronchi-olar Clara cell in rats. Am. J. Physiol. 247: C197–C203.
15. PLOPPER, C.G., D.M. HYDE & A.R. BUCKPITT. 1997. In The Lung: Scientific Founda-tions, pp. 517–533. Lippincott-Raven. Philadelphia.
16. ANDERSSON, O., G. NOACK, B. ROBERTSSON et al. 1994. Ontogeny of a human poly-chlorinated biphenyl binding protein: level of expression in tracheal aspirates inbroncopulmonary dysplasia. Chest 105: 17–22.
17. BERNARD, A., N. THIELEMANS, R. LAUWERYS et al. 1994. Clara cell protein in humanamniotic fluid—a potential marker of fetal lung growth. Pediatr. Res. 36: 771–775.
18. STRIPP, B.R., K. MAXSON, R. MERA & G. SINGH. 1995. Plasticity of airway cell prolif-eration and gene expression after acute naphthalene injury. Am. J. Physiol. 269:L791–L799.
19. VAN WINKLE, L.S., A.R. BUCKPITT, S.J. NISHIO et al. 1995. Cellular response innaphthalene-induced Clara cell injury and bronchiolar epithelial repair in mice. Am.J. Physiol. 269: L800–L818.
20. JENSEN, S.M., J.E. JONES, H. PASS et al. 1994. Clara cell 10 kDa protein mRNA in nor-mal and atypical regions of human respiratory epithelium. Int. J. Cancer 58: 629–637.
21. NORD, M., M. LÅG, T.N. CASSEL et al. 1998. Regulation of CCSP (PCB-BP/utero-globin) expression in primary cultures of lung cells—involvement of C/EBP. DNACell Biol. 17: 481–492.
22. MIELE, L., M.E. CORDELLA & A.B. MUKHERJEE. 1987. Uteroglobin: structure, molecu-lar biology, and new perspectives on its function as a phospholipase A2 inhibitor.Endocr. Rev. 8: 474–490.
23. MUKHERJEE, A.B., G.C. KUNDU, G. MANTILE-SELVAGGI et al. 1999. Uteroglobin: anovel cytokine? Cell. Mol. Life Sci. 55: 771–787.
24. STRIPP, B.R., P.L. SAWAYA, D.S. LUSE et al. 1992. Cis-acting elements that confer lungepithelial cell expression of the CC10 gene. J. Biol. Chem. 267: 14703–14712.
25. HACKETT, B.P. & J.D. GITLIN. 1992. Cell-specific expression of a Clara cell secretoryprotein–human growth hormone gene in the bronchiolar epithelium of transgenicmice. Proc. Natl. Acad. Sci. U.S.A. 89: 9079–9083.

163NORD et al.: CCSP/UG PROMOTER
26. RAY, M.K., S.W. MAGDALENO, M.J. FINEGOLD & F.J. DE MAYO. 1995. Cis-actingelements involved in the regulation of mouse Clara cell–specific 10-kDa proteingene—in vitro and in vivo analysis. J. Biol. Chem. 270: 2689–2694.
27. BINGLE, C.D. & J.D. GITLIN. 1993. Identification of hepatocyte nuclear factor-3 bind-ing sites in the Clara cell secretory protein gene. Biochem. J. 295: 227–232.
28. BINGLE, C.D., B.P. HACKETT, M. MOXLEY et al. 1995. Role of hepatocyte nuclearfactor-3α and hepatocyte nuclear factor-3β in Clara cell secretory protein geneexpression in the bronchiolar epithelium. Biochem. J. 308: 197–202.
29. STRIPP, B.R., J.A. HUFFMAN & R.J. BOHINSKI. 1994. Structure and regulation of themurine Clara cell secretory protein gene. Genomics 20: 27–35.
30. SAWAYA, P.L., B.R. STRIPP, J.A. WHITSETT & D.S. LUSE. 1993. The lung-specific CC10gene is regulated by transcription factors from the AP-1, octamer, and hepatocytenuclear factor-3 families. Mol. Cell. Biol. 13: 3860–3870.
31. SAWAYA, P.L. & D.S. LUSE. 1994. Two members of the HNF-3 family have oppositeeffects on a lung transcriptional element; HNF-3α stimulates and HNF-3β inhibitsactivity of region I from the Clara cell secretory protein (CCSP) promoter. J. Biol.Chem. 269: 22211–22216.
32. RAY, M.K., C.Y. CHEN, R.J. SCHWARTZ & F.J. DE MAYO. 1996. Transcriptional regula-tion of a mouse Clara cell–specific protein (mCC10) gene by the NKx transcriptionfactor family members thyroid transcription factor 1 and cardiac muscle–specifichomeobox protein (CSX). Mol. Cell. Biol. 16: 2056–2064.
33. TOONEN, R.F.G., S. GOWAN & C.D. BINGLE. 1996. The lung enriched transcription fac-tor TTF-1 and the ubiquitously expressed proteins Sp1 and Sp3 interact with ele-ments located in the minimal promoter of the rat Clara cell secretory protein gene.Biochem. J. 316: 467–473.
34. ZHANG, L.Q., J.A. WHITSETT & B.R. STRIPP. 1997. Regulation of Clara cell secretoryprotein gene transcription by thyroid transcription factor-1. Biochim. Biophys. ActaGene Struct. Expr. 1350: 359–367.
35. MONAGHAN, A.P., K.H. KAESTNER, E. GRAU & G. SCHUTZ. 1993. Postimplantationexpression patterns indicate a role for the mouse forkhead/HNF-3 alpha, beta, andgamma genes in determination of the definitive endoderm, chordamesoderm, andneuroectoderm. Development 119: 567–578.
36. ANG, S.L., A. WIERDA, D. WONG et al. 1993. The formation and maintenance of thedefinitive endoderm lineage in the mouse: involvement of HNF3/forkhead proteins.Development 119: 1301–1315.
37. CLEVIDENCE, D.E., D.G. OVERDIER, R.S. PETERSON et al. 1994. Members of the HNF-3/forkhead family of transcription factors exhibit distinct cellular expression patternsin lung and regulate the surfactant protein B promoter. Dev. Biol. 166: 195–209.
38. ANG, S.L. & J. ROSSANT. 1994. HNF-3 beta is essential for node and notochord forma-tion in mouse development. Cell 78: 561–574.
39. WEINSTEIN, D.C., A. RUIZ I ALTABA, W.S. CHEN et al. 1994. The winged-helix tran-scription factor HNF-3 beta is required for notochord development in the mouseembryo. Cell 78: 575–588.
40. ZHOU, L., L. LIM, R.H. COSTA & J.A. WHITSETT. 1996. Thyroid transcription factor-1,hepatocyte nuclear factor-3 beta, surfactant protein B, C, and Clara cell secretoryprotein in developing mouse lung. J. Histochem. Cytochem. 44: 1183–1193.
41. BRAUN, H. & G. SUSKE. 1998. Combinatorial action of HNF3 and Sp family transcrip-tion factors in the activation of the rabbit uteroglobin/CC10 promoter. J. Biol. Chem.273: 9821–9828.
42. LIM, L., H. ZHOU & R.H. COSTA. 1997. The winged helix transcription factor HFH-4 isexpressed during choroid plexus epithelial development in the mouse embryo. Proc.Natl. Acad. Sci. U.S.A. 94: 3094–3099.
43. HACKETT, B.P., S.L. BRODY, M. LIANG et al. 1995. Primary structure of hepatocyte nuclearfactor/forkhead homologue 4 and characterization of gene expression in the developingrespiratory and reproductive epithelium. Proc. Natl. Acad. Sci. U.S.A. 92: 4249–4253.
44. CHEN, J., H.J. KNOWLES, J.L. HEBERT & B.P. HACKETT. 1998. Mutation of the mousehepatocyte nuclear factor/forkhead homologue 4 gene results in an absence of ciliaand random left-right asymmetry. J. Clin. Invest. 102: 1077–1082.

164 ANNALS NEW YORK ACADEMY OF SCIENCES
45. TICHELAAR, J.W., L. LIM, R.H. COSTA & J.A. WHITSETT. 1999. HNF-3/forkhead homo-logue-4 influences lung morphogenesis and respiratory epithelial cell differentiationin vivo. Dev. Biol. 213: 405–417.
46. TICHELAAR, J.W., S.E. WERT, R.H. COSTA et al. 1999. HNF-3/forkhead homologue-4(HFH-4) is expressed in ciliated epithelial cells in the developing mouse lung. J.Histochem. Cytochem. 47: 823–832.
47. BLATT, E.N., X.H. YAN, M.K. WUERFFEL et al. 1999. Forkhead transcription factorHFH-4 expression is temporally related to ciliogenesis. Am. J. Respir. Cell Mol.Biol. 21: 168–176.
48. HELLQVIST, M., M. MAHLAPUU, L. SAMUELSSON et al. 1996. Differential activation oflung-specific genes by two forkhead proteins, FREAC-1 and FREAC-2. J. Biol.Chem. 271: 4482–4490.
49. PETERSON, R.S., L. LIM, H. YE et al. 1997. The winged helix transcriptional activatorHFH-8 is expressed in the mesoderm of the primitive streak stage of mouse embryosand its cellular derivatives. Mech. Dev. 69: 53–69.
50. LAZZARO, D., M. PRICE, M. DE FELICE & R. DI LAURO. 1991. The transcription factorTTF-1 is expressed at the onset of thyroid and lung morphogenesis and in restrictedregions of the fetal brain. Development 113: 1093–1104.
51. RAMIREZ, M.I., A.K. RISHI, Y.X. CAO & M.C. WILLIAMS. 1997. TGT3, thyroid tran-scription factor I, and Sp1 elements regulate transcriptional activity of the 1.3-kilo-base pair promoter of T1 alpha, a lung alveolar type I cell gene. J. Biol. Chem. 272:26285–26294.
52. KIMURA, S., Y. HARA, T. PINEAU et al. 1996. The t/ebp null mouse: thyroid-specificenhancer-binding protein is essential for the organogenesis of the thyroid, lung, ven-tral forebrain, and pituitary. Genes Dev. 10: 60–69.
53. MINOO, P., G. SU, H. DRUM et al. 1999. Defects in tracheoesophageal and lungmorphogenesis in Nkx2.1(−/−) mouse embryos. Dev. Biol. 209: 60–71.
54. WHITSETT, J.A. & S.W. GLASSER. 1998. Regulation of surfactant protein gene tran-scription. Biochim. Biophys. Acta 1408: 303–311.
55. LI, F., E. ROSENBERG, C.I. SMITH et al. 1995. Correlation of expression of transcriptionfactor C/EBP alpha and surfactant protein genes in lung cells. Am. J. Physiol. 269:L241–L247.
56. CASSEL, T.N., L. NORDLUND-MÖLLER, O. ANDERSSON et al. 2000. C/EBP alpha and C/EBP delta activate the Clara cell secretory protein gene through activation with twoadjacent C/EBP-binding sites. Am. J. Respir. Cell Mol. Biol. 22: 469–480.
57. LEKSTROM-HIMES, J. & K.G. XANTHOPOULOS. 1998. Biological role of the CCAAT/enhancer-binding protein family of transcription factors. J. Biol. Chem. 273: 28545–28548.
58. DIEHL, A.M. 1998. Roles of CCAAT/enhancer-binding proteins in regulation of liverregenerative growth. J. Biol. Chem. 273: 30843–30846.
59. CRONIGER, C., P. LEAHY, L. RESHEF & R.W. HANSON. 1998. C/EBP and the control ofphosphoenolpyruvate carboxykinase gene transcription in the liver. J. Biol. Chem.273: 31629–31632.
60. DARLINGTON, G.J., S.E. ROSS & O.A. MACDOUGALD. 1998. The role of C/EBP genes inadipocyte differentiation. J. Biol. Chem. 273: 30057–30060.
61. POLI, V. 1998. The role of C/EBP isoforms in the control of inflammatory and nativeimmunity functions. J. Biol. Chem. 273: 29279–29282.
62. CAO, Z., R.M. UMEK & S.L. MCKNIGHT. 1991. Regulated expression of three C/EBPisoforms during adipose conversion of 3T3-L1 cells. Genes Dev. 5: 1538–1552.
63. BREED, D.R., L.R. MARGRAF, J.L. ALCORN & C.R. MENDELSON. 1997. Transcriptionfactor C/EBP delta in fetal lung: developmental regulation and effects of cyclicadenosine 3′,5′-monophosphate and glucocorticoids. Endocrinology 138: 5527–5534.
64. CASSEL, T.N., J-Å. GUSTAFSSON & M. NORD. 2000. CYP2B1 is regulated by C/EBPalpha and C/EBP delta in lung epithelial cells. Mol. Cell. Biol. Res. Commun. 3: 42–47.
65. FLODBY, P., C. BARLOW, H. KYLEFJORD et al. 1996. Increased hepatic cell proliferationand lung abnormalities in mice deficient in CCAAT/enhancer binding protein α. J.Biol. Chem. 271: 24753–24760.

165NORD et al.: CCSP/UG PROMOTER
66. CRONIGER, C., M. TRUS, K. LYSEK-STUPP et al. 1997. Role of the isoforms of CCAAT/enhancer-binding protein in the initiation of phosphoenolpyruvate carboxykinase(GTP) gene transcription at birth. J. Biol. Chem. 272: 26306–26312.
67. DE MAYO, F.J., S. DAMAK, T.N. HANSEN & D.W. BULLOCK. 1991. Expression and regu-lation of the rabbit uteroglobin gene in transgenic mice. Mol. Endocrinol. 5: 311–318.
68. SUSKE, G., W. LORENZ, J. KLUG et al. 1992. Elements of the rabbit uteroglobin pro-moter mediating its transcription in epithelial cells from the endometrium and lung.Gene Expr. 2: 339–352.
69. DENNIG, J., G. HAGEN, M. BEATO & G. SUSKE. 1995. Members of the Sp transcriptionfactor family control transcription from the uteroglobin promoter. J. Biol. Chem.270: 12737–12744.
70. STAHLMAN, M.T., M.E. GRAY & J.A. WHITSETT. 1996. Expression of thyroid transcrip-tion factor-1 (TTF-1) in fetal and neonatal human lung. J. Histochem. Cytochem. 44:673–678.
71. SUSKE, G. 2000. Transient transfection of Schneider cells in the study of transcriptionfactors. Methods Mol. Biol. 130: 175–187.
72. ADAMSON, I.Y.R. 1997. In The Lung: Scientific Foundations, pp. 993–1001. Lippin-cott-Raven. Philadelphia.





























